RECHERCHE
Parallel processing relies on a distributed,
low-dimensional cortico-cerebellar architecture
Eli J. Müller1,2, Fulvia Palesi3,4, Kevin Y. Hou1,2, Joshua Tan1,2, Thomas Close5,6,7,
Claudia A. M.. Gandini Wheeler-Kingschott3,4,8, Egidio D’Angelo3,4,
Fernando Calamante2,6,8, and James M. Shine1,2
1Complex Systems Research Group, The University of Sydney, Sydney, NSW, Australia
2Brain and Mind Centre, The University of Sydney, Sydney, NSW, Australia
3Brain Connectivity Research Center, IRCCS Mondino Foundation, Pavia, Italy
4Department of Brain and Behavioral Sciences, University of Pavia, Pavia, Italy
5National Imaging Facility, Sydney, NSW, Australia
6School of Biomedical Engineering, The University of Sydney, Sydney, NSW, Australia
7Sydney Imaging, The University of Sydney, Sydney, NSW, Australia
8NMR Research Unit, Queen Square Multiple Sclerosis Centre, Faculty of Brain Sciences, UCL Queen
Square Institute of Neurology, UCL, Londres, ROYAUME-UNI
Mots clés: IRMf, Dual-task, Cerebellum, Cerebral cortex, Diffusion, Parallel
ABSTRAIT
A characteristic feature of human cognition is our ability to ‘multi-task’—performing two or
more tasks in parallel—particularly when one task is well learned. How the brain supports
this capacity remains poorly understood. Most past studies have focussed on identifying
the areas of the brain—typically the dorsolateral prefrontal cortex—that are required to
navigate information-processing bottlenecks. En revanche, we take a systems neuroscience
approach to test the hypothesis that the capacity to conduct effective parallel processing relies
on a distributed architecture that interconnects the cerebral cortex with the cerebellum.
The latter structure contains over half of the neurons in the adult human brain and is well
suited to support the fast, effective, dynamic sequences required to perform tasks relatively
automatically. By delegating stereotyped within-task computations to the cerebellum, le
cerebral cortex can be freed up to focus on the more challenging aspects of performing the
tasks in parallel. To test this hypothesis, we analysed task-based fMRI data from 50 participants
who performed a task in which they either balanced an avatar on a screen (équilibre),
performed serial-7 subtractions (calculation) or performed both in parallel (dual task). Using a
set of approaches that include dimensionality reduction, structure-function coupling, et
time-varying functional connectivity, we provide robust evidence in support of our hypothesis.
We conclude that distributed interactions between the cerebral cortex and cerebellum are
crucially involved in parallel processing in the human brain.
RÉSUMÉ DE L'AUTEUR
How does the brain support the performance of multiple complex tasks, in parallel? Le
distributed architecture of the cerebellum is ideally placed to interact with the cerebral cortex,
creating complex channels for segregated information processing that afford the execution
of parallel tasks. Ici, we apply time-resolved functional connectivity analyses to functional
un accès ouvert
journal
Citation: Müller, E. J., Palesi, F., Hou,
K. Y., Tan, J., Close, T., Gandini
Wheeler-Kingschott, C. UN. M., D’Angelo,
E., Calamente, F., & Briller, J.. M.. (2023).
Parallel processing relies on a
distributed, low-dimensional cortico-
cerebellar architecture. Réseau
Neurosciences, 7(2), 844–863.
https://doi.org/10.1162/netn_a_00308
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00308
Informations complémentaires:
https://doi.org/10.1162/netn_a_00308
Reçu: 22 Juillet 2022
Accepté: 11 Janvier 2023
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
James M.. Briller
mac.shine@sydney.edu.au
Éditeur de manipulation:
Lucina Uddin
droits d'auteur: © 2023
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
t
/
/
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
t
.
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
Parallel processing:
The running of two or more
processes in tandem.
Cerebellum:
A physically small but neuronally
dense structure in the hindbrain
important for sensorimotor
adaptation and anticipation.
MRI data collected while individuals performed a dual task that required either balancing,
calculating, or the two in tandem. We found robust evidence for distinct patterns of cortico-
cerebellar connectivity as a function of task performance.
INTRODUCTION
How do distributed whole-brain neural activity patterns give rise to human cognitive function?
This question lies at the heart of modern psychology and neuroscience but, despite decades of
neuroimaging experiments, we still do not have a clear answer. One reason is that conven-
tional neuroimaging methods applied to data from cognitive tasks typically represent the brain
as a static snapshot of independent parts or at best, ‘functionally connected’ pairs of brain
régions (John et al., 2022). Another important issue is that neuroimaging experiments are usu-
ally designed to identify regions that are most selectively associated with a specific task, mais
are less well suited to distinguishing the presence of multiple concurrent cognitive constructs
within the same task (Poldrack, 2012). For these reasons, many leading theories in cognitive
neuroscience have relied on relatively static descriptions of the ‘key brain regions involved’ in
a particular task.
In contrast to this view, there is evidence to suggest that the neural implementation of
cognitive function in humans is far more dynamic and integrative (Eisenreich et al., 2017).
In solving real world problems, we rarely isolate a specific cognitive capacity, such as focussed
attention or resistance to distraction, but instead combine multiple cognitive constructs
together in order to solve challenges in real time (Poldrack et al., 2011). Consider an experi-
enced driver navigating heavy highway traffic in the pouring rain—the driver must remain
focussed on the road, ensure the windshield wipers are on, regularly check their blind spots
and also keep the pedals depressed at the appropriate level. This view of cognitive function in
the real world is crucially dependent on the parallel processing of multiple distinct challenges;
cependant, for the reasons outlined above, we still lack a satisfying description of how the
human brain is capable of supporting parallel processing.
To facilitate complex coordinated behavioural responses underpinned by similarly complex
spatiotemporal activity patterns, the brain may first learn to execute at least one of the com-
putations automatically (c'est à dire., without paying close, conscious attention to the completion of
the task). Pour y parvenir, the system must be capable of responding to specific contexts with a
high degree of spatial and temporal precision (Schmitz & Duncan, 2018). Deuxièmement, le
responses must be relatively error free and reliable. Enfin, the system must be able to be trig-
gered in the presence of a specific stimulus or context without the need for deliberate atten-
tion. Without making the responses to different computational burdens relatively stereotyped
in this fashion, performing two (ou plus) computations in parallel would require the prioritisa-
tion of one of the computations, likely to the detriment of the other task(s). En outre, any two
tasks learned by the same network could potentially run into structural interference (Petri et al.,
2021), particularly if the networks required to complete the overlapping tasks use similar
cortical regions.
Surtout, the architecture of the cerebellum is ideally suited to fulfil each of the features
required for automatic processing, both in the sensorimotor and cognitive domains (D’Angelo,
2019; D’Angelo & Casali, 2013; Ramnani, 2014; Shine et al., 2019). D'abord, the cerebellum is
organized in parallel modules with different cerebrocortical regions (D’Angelo & Casali,
2013). In direct contrast to the basal ganglia, the internal circuitry of the cerebellar cortex
Neurosciences en réseau
845
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
/
t
/
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
.
t
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
Cerebral cortex:
The thin, outer layer of the
telencephalon important for
deliberate, conscious processing.
Dual task:
The performance of two
simultaneous tasks, one of which is
typically assumed to be easier to
automatise than the other.
Low-dimensional:
A system whose activity can be
expressed through a smaller number
of components without a substantial
loss in explained variance.
consists of sparse, distributed connectivity patterns that likely support dimensionality expan-
sion (Cayco-Gajic & Silver, 2019), rather than reduction (as is the case for the basal ganglia;
Bar-Gad et al., 2003; Wilson, 2013). En outre, the glutamatergic outputs of the cerebellum
through the deep cerebellar nuclei innervate ‘core’ thalamic nuclei (Kuramoto et al., 2009),
which project to the granular layers of the frontal cortex (Preuss & Sage, 2022) in a much more
precise manner than the ‘matrix’ thalamic nuclei. There is also evidence that cerebellar cir-
cuits can condition on their own outputs, and hence learn to execute specific sequences of
effects based on triggering context signals (Khilkevich et al., 2018). Anatomically, the cerebel-
lum is bidirectionally interconnected with multiple cerebrocortical areas, with major tracts
connecting the dentate nucleus to the frontal and prefrontal cerebral cortex, along with other
associative areas (Palesi et al., 2015, 2017). Functionally, the cerebellum plays a critical role in
shaping complex functional network dynamics (Palesi et al., 2020), as evidenced by its role in
both resting-state (Castellazzi et al., 2014, 2018) and task-related neuroimaging studies
(Alahmadi et al., 2016; Balsters & Ramnani, 2011; Casiraghi et al., 2019; Shine et al.,
2019). Based on these architectural features and relationships with complex, dynamic neuro-
imaging patterns, we hypothesized that connections between the cerebellar cortex and cere-
bellum are crucial for the facilitation of parallel processing. Using a set of approaches that
include dimensionality reduction, structure-function coupling, and time-varying functional
connectivité, we provide robust evidence in support of our hypothesis.
RÉSULTATS
To test this hypothesis, we reanalysed an existing fMRI dataset (Papegaaij et al., 2017) consist-
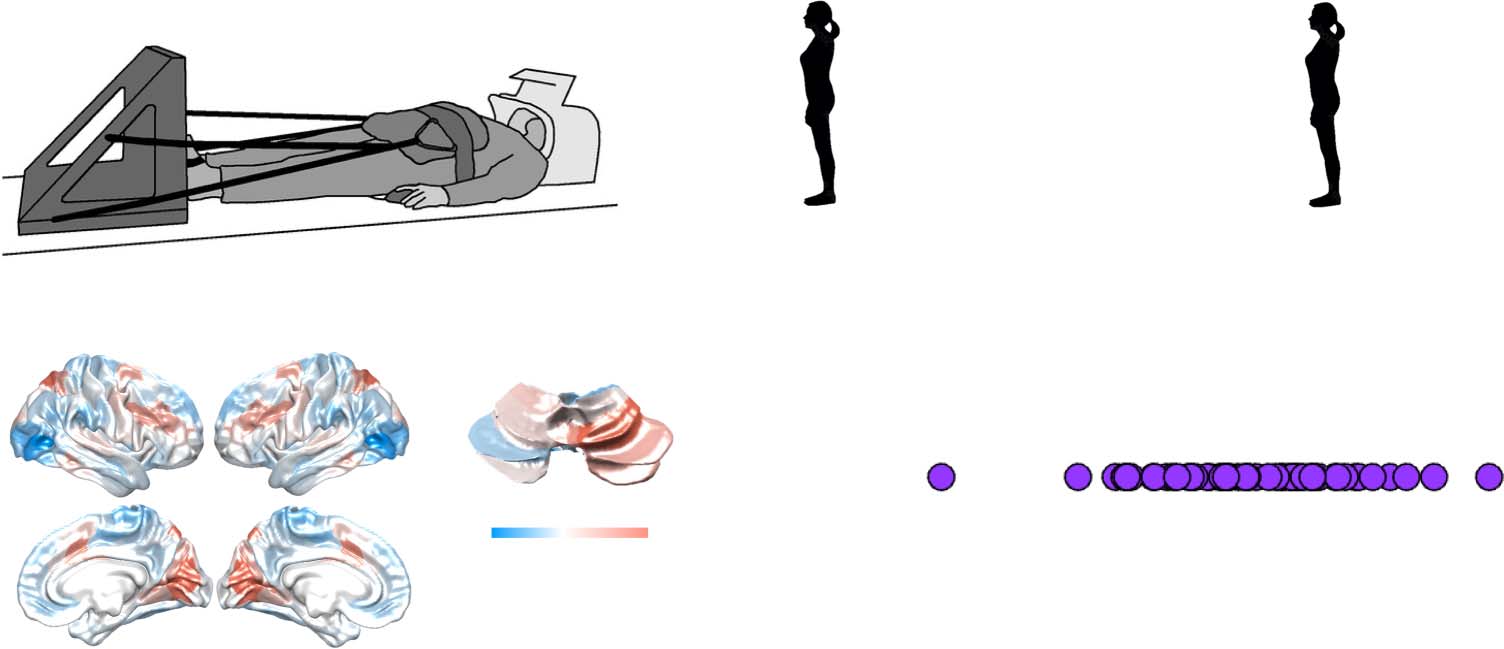
ing of 50 healthy individuals dual task in a 3T MRI scanner with their feet resting on a force
plate (Figure 1A), and their vision oriented towards a two-dimensional avatar that tilted for-
ward and backward. There were three distinct trial types: during balance blocks (Figure 1B,
bleu), participants had to stabilize the slow fluctuations of the avatar using forward and back-
ward movements on the force plate; during calculation blocks (Figure 1C, red), subjects had to
track between three and four audible beeps, and then subtract that number, multiplied by 7,
from a cue number presented prior to the trial; and during dual-task blocks (Figure 1D, purple),
subjects performed both trials simultaneously.
Brain State Signatures During Dual-Task Performance
D'abord, we compared the BOLD patterns associated with the performance of the three different
tasks blocks. Spécifiquement, we created a difference map between the average group-level β
parameters estimated from 400 cortical and 28 cerebellar regions of interest for the balance
and calculation trials (Δ) (Figure 1E). By comparing this difference map to the β map from the
dual-task trials—r(Δ, βDT)—we could determine whether performing the two tasks in tandem
led to a brain map that was more or less like one or the other single tasks—a positive corre-
lation with this map (λ1) was suggestive of the dual task reflecting the more challenging
calculation task, a negative correlation with the less challenging balance task, and a null cor-
relation with the notion of optimally splitting activity between the two (or a pattern orthogonal
to the two single tasks). Consistent with the second option, we found that the low-dimensional
signature of dual-task performance was more similar to the calculation β map than the balance
β map (r = 0.192 ± 0.05, p = 6.5 × 10−5; Figure 1F), suggesting that during the dual-task trials,
the cerebral cortex and cerebellum configured their activity to ensure the effective completion
of the calculation trials.
Neurosciences en réseau
846
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
/
t
/
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
t
.
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
Chiffre 1. Low-dimensional balance between integration and segregation during dual-task performance. (UN) participants lay supine in an MRI
scanner, with their legs controlling a force plate. (B) Balance trials (bleu) involved a dynamically moving avatar that the participant had to
match. (C) calculation trials involved listening to a series of beeps, and then subtracting the multiple of 7 times the number of beeps from a cue
number (red). (D) dual-task trials required performing both tasks, simultaneously (purple). (E) The calculation trials recruited increased BOLD in
fronto-parietal and visual cortices, along with right superior cerebellum, whereas balance trials were associated with increased BOLD in lateral
visual cortex, medial motor cortex, and parietal operculum. (F) the dual-task β map across all 50 subjects was more similar to the calculation
β map (c'est à dire., positive correlation with λ1) than the balance β map (c'est à dire., inverse correlation with λ1); *** p < 0.001.
Despite the brain states during dual-task trials having more in common with the calculation
than the balance trials, close examination of the RMS error of the balance portion of the dual-
task trials suggests that subjects were performing the task as well as than when they performed
the balance trial on its own (Kolmogorov–Smirnov test: p = 0.358). So how was the brain con-
figured on these dual-task trials in order to mediate this stability? Based on previous empirical
(Balsters & Ramnani, 2011) and theoretical (D’Angelo & Casali, 2013; Shine, 2020; Shine &
Shine, 2014) work, we hypothesized that the distributed architecture integrating the cerebral
cortex and cerebellum should be important for mediating this putative parallel processing per-
formance. One straightforward prediction is that balancing multiple tasks at the same time
should recruit more regions of the cerebellum, and hence that cerebellar blood flow should
be more strongly associated with dual-task performance than either the balance or calculation
task alone. We found evidence to confirm this hypothesis—namely, greater proportion of cer-
ebellar regions were associated with a positive mean β value in dual task as compared to bal-
ance and/or calculation trials (67.3% vs. 35.7% and 39.3%, respectively; χ2 (2, N = 50) =
249.6, p < 1.0 × 10−4).
Unique Patterns of Cortico-Cerebellar Functional Connectivity During Dual-Task Performance
Given that the dual-task trials were more similar to calculation trials than balance trials
(Figure 1F), how was the brain capable of supporting multiple tasks at the same time? We
hypothesized that balance, calculation, and dual-task trials should have unique patterns of
cortico-cerebellar functional connectivity that could allow the brain to support multiple
channels of communication within the same system. To test this hypothesis, we calculated
the time-varying functional connectivity between all cortical and cerebellar parcels using
the Multiplication of Temporal Derivatives approach (window = 20 TRs; Shine et al., 2015)
and then contrasted the three trial types with one another. We observed robust differences
between the three trial types (Figure 2). For instance, calculation trials (when compared to
balance trials) were associated with widespread cortico-cerebellar connectivity between
Network Neuroscience
847
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
a
_
0
0
3
0
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
lobule V and the majority of cortical networks, as well as more targeted connections between
VIIIa/IX and primary sensorimotor networks (Figure 2A). In contrast, balance trials (when
compared to calculation trials) showed predominant increases in intermediate cerebellar
lobules (e.g., Crus I and II) with higher order cortical networks. In contrast, dual-task trials were
associated with heightened fronto-parietal connections with intermediate cerebellar lobules,
particularly Crus I and VIIIa, when compared to both balance (Figure 2B) and calculation trials
(Figure 2C).
Having confirmed a robust relationship between the cerebral cortex and cerebellum during
dual-task performance, we next asked whether cortico-cerebellar functional connectivity
patterns differentiated between correct and error dual-task trials. To test this hypothesis, we
fit a General Linear Model to each dual-task trial, independently, for each cortico-cerebellar
time-varying connectivity score. We then split dual-task trials into correct (accurate calculation
and small RMS error [<50% of population distribution]) and incorrect (inaccurate calculation,
large RMS error [>50%] or both) trials and compared (using a set of independent-samples
t tests) the task-based functional connectivity between cortical and cerebellar parcels as a
function of effective dual-task performance. We conducted a permutation test (5,000 itera-
tion) to determine the likelihood of each edge being distinct between the two groups by
chance. To summarize these results, we computed the mean significant β-value for the func-
tional connectivity between each cerebellar lobule (averaged across hemispheres, and ignor-
ing the connections of the vermis; from the cerebellar SUIT atlas (Diedrichsen, 2006)) et
each of 7 pre-identified cortical networks (the Yeo 7 parcellation from the 400-region Schaefer
atlas (Schaefer et al., 2018; Chiffre 3). We found a robust increase in task-based functional
connectivity between the ventral attention network ( VAN) and lobules Crus II, VIIb, VIIIa
and VI (Chiffre 3), as well as more distributed connections between lobule X and multiple cor-
tical subnetworks. En revanche, Crus I was relatively functionally disconnected from all cortical
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
/
/
t
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
.
t
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 2. Cortico-cerebellar task-based functional connectivity. (UN) patterns of task-based cortico-cerebellar functional connectivity during
calculation (red) when compared to balance (bleu) trials—upper: mean task-based connectivity strength for cerebral cortex and cerebellum;
lower: mean task-based connectivity strength collapsed into 7 Yeo networks (columns) et 10 average lobules (rows). (B) similar maps for dual
task (purple) versus balance. (C) similar maps for dual task versus calculation. Note: results were consistent for left and right hemispheres. VIS =
visuel; SM = somatomotor; DAN = dorsal attention network; VAN = ventral attention network; LIM = limbic network; CON = control network;
DMN = default mode network.
Neurosciences en réseau
848
Cortico-cerebellar dual tasking
L'intégration:
The formation of a unified or
coordinated whole—in the case of
brain networks, the presence of
relatively diffuse connections across
brain regions.
Segregation:
The formation of setting something
apart from others—in the case of
brain networks, the presence of
tight-knit subcommunities.
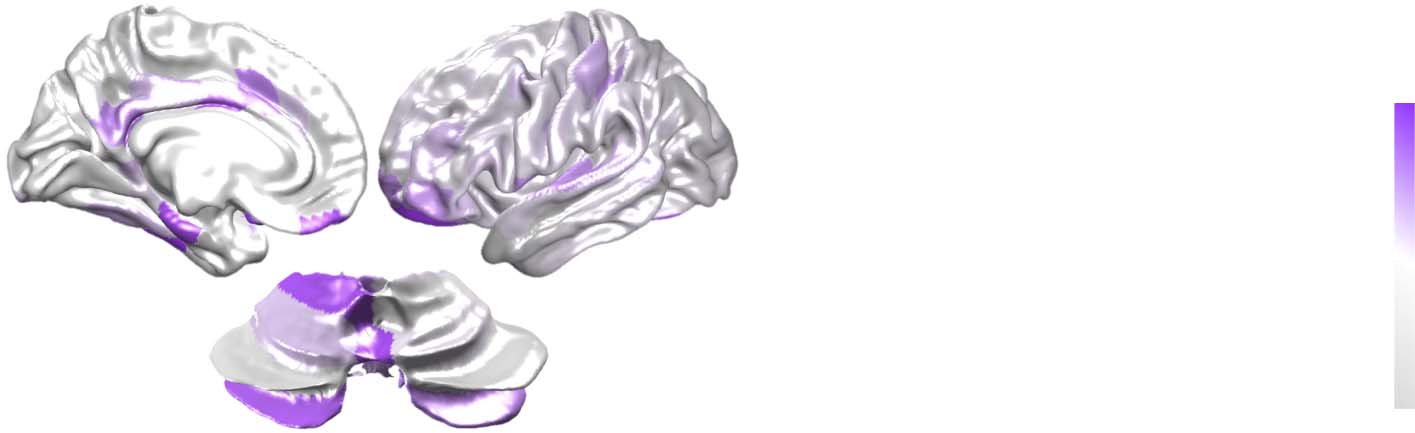
Increased cortico-cerebellar task-based functional connectivity associated with success-
Chiffre 3.
ful dual-task performance. Gauche: Key cortical and cerebellar regions with heightened task-based
functional connectivity during dual-task trials with correct versus incorrect answers. Droite: Mean
significant β-value (cortical sub-network [Yeo 7 atlas] vs. cerebellar lobule [SUIT atlas]) associated
with task-based functional connectivity values for correct versus incorrect dual-task performance
(p < 0.001; random permutation test). VIS = visual; SM = somatomotor; DAN = dorsal attention
network; VAN = ventral attention network; LIM = limbic network; CON = control network; DMN =
default mode network.
networks (except VAN) during effective dual-task performance, which is consistent with
known patterns of cerebellar lesion-related cognitive impairments (Ilg et al., 2013).
Dual-Task Performance Balances Network Integration and Segregation
One way in which the distributed cortico-cerebellar architecture could facilitate effective par-
allel processing is by striking an effective balance between integration and segregation (Bassett
et al., 2015, p. 201; Mohr et al., 2016; Shine & Poldrack, 2017). In previous work, we have used
a combination of time-varying functional connectivity and a topological measure that quan-
tifies network-level integration—the participation coefficient (PC; Shine et al., 2016)—to dem-
onstrate that the systems-level network structure of functional connectivity changes during task
performance, with cognitively challenging tasks requiring higher integration than relatively
simple tasks (Shine et al., 2016). From this, we predicted that the balance task should be rela-
tively segregated (i.e., low PC), the calculation task should be relatively integrated (i.e., high
PC), and the dual-task trials should strike a balance between the two extremes (i.e., intermediate
PC). Using our standard time-varying analysis (see Methods), we observed robust evidence for
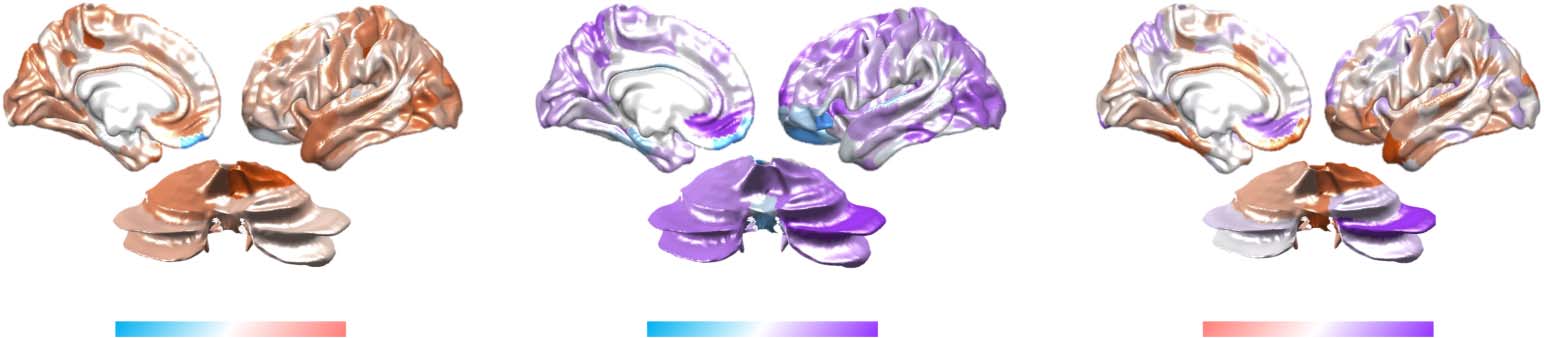
our predictions (Figure 4; F2,147 = 3.41; p = 0.036). In addition, although the dual-task topo-
logical pattern was positively correlated with the average of balance and calculation (r = 0.464;
p < 0.001), it was not a direct superposition of the two maps, suggesting topological reconfig-
uration during the different task states. Together, these results confirm that parallel processing in
the brain is supported by a topological balance between integration and segregation.
Cortico-Cerebellar Activity Flow Mapping
The input and output streams of cerebral cortex and cerebellum interact via distinct white mat-
ter pathways. Importantly, while the structural connections between these two structures are
reciprocal, they are imbalanced (Palesi et al., 2015, 2017)—different pathways exist from the
cerebral cortex to the cerebellum than from the cerebellum to the cerebral cortex. Specifically,
thick-tufted layer V pyramidal neurons in the deep layers of the cerebral cortex send projec-
tions to the mossy fibre pathway of the cerebellum (via the pontine nuclei), thus forming the
cortico-ponto-cerebellar (CPC) tract (Figure 5A). In contrast, the cerebral cortex receives feed-
back from the cerebellum via the deep cerebellar nuclei, which project via the ‘Core’ nuclei of
the thalamus—that is, the cerebello-thalamo-cortical (CTC) tract (Figure 5B). Plastic changes
Network Neuroscience
849
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
a
_
0
0
3
0
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
Figure 4. Parallel processing balances integration and segregation. Balance trials were associated
with relative segregation (low PC; blue), calculation trials with relative integration (high PC; red),
and dual-task trials with a balance between integration and segregation (intermediate PC; purple);
F2,147 = 3.41; p = 0.036. Thick lines represent the median value for each group.
between the mossy fibre pathway and the Purkinje cells of the cerebellar cortex are proposed
to act as a major site for the refinement of automatized behaviour (D’Angelo et al., 2016;
Ramnani, 2014; Shine, 2020; Shine & Shine, 2014) and hence, the capacity to perform mul-
tiple tasks simultaneously. From our observations that the time series of the cerebral cortex and
cerebellum were highly coordinated during dual-task behaviour, we hypothesized that the
specific patterns of BOLD activity in both the cortex and cerebellum should be related to
the intersection between prior BOLD activity in the cerebellum (via the CTC) and cerebral
cortex (via the CPC).
To test this hypothesis, we adapted the activity flow mapping approach (Cole et al., 2016) to
incorporate the structural connectivity between the cortex and cerebellum. Specifically, we
extracted 9 × 107 structural connectivity weights for both the contralateral CPC (Figure 6A,
orange) and CTC (Figure 6B, green) tracts (Palesi et al., 2017) from a single healthy 26–
30-year-old female (ID no. 100307) from the Human Connectome Project (a single subject
connectome was chosen so as to retain precision in the parcel-to-parcel connectivity estimates
for both CPC and CTC—note, however, that maps were highly similar to those previously
extracted from 28 healthy participants from the HCP (Palesi et al., 2017)). While both tracts
are overexpressed in the frontal cortices, there were relatively more CPC projections from the
parietal lobes and more CTC projections that innervate the frontal cortex, which is consistent
with known anatomical projection patterns (D’Angelo & Casali, 2013; Prevosto & Sommer,
2013; Ramnani, 2006; Shine, 2020). A parsimonious interpretation of these data is that
the frontal cortex benefits from the information provided to the cerebellum by posterior
cortices that process potential opportunities for action (also known as affordances; Pezzulo
& Cisek, 2016).
If cortico-cerebellar communication is required for effective dual-task performance, then
blood flow within either the cerebral cortex or cerebellum during dual-task trials should be
predictive of subsequent blood flow (assuming sufficient delay) within the cortical (or
Network Neuroscience
850
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
a
_
0
0
3
0
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
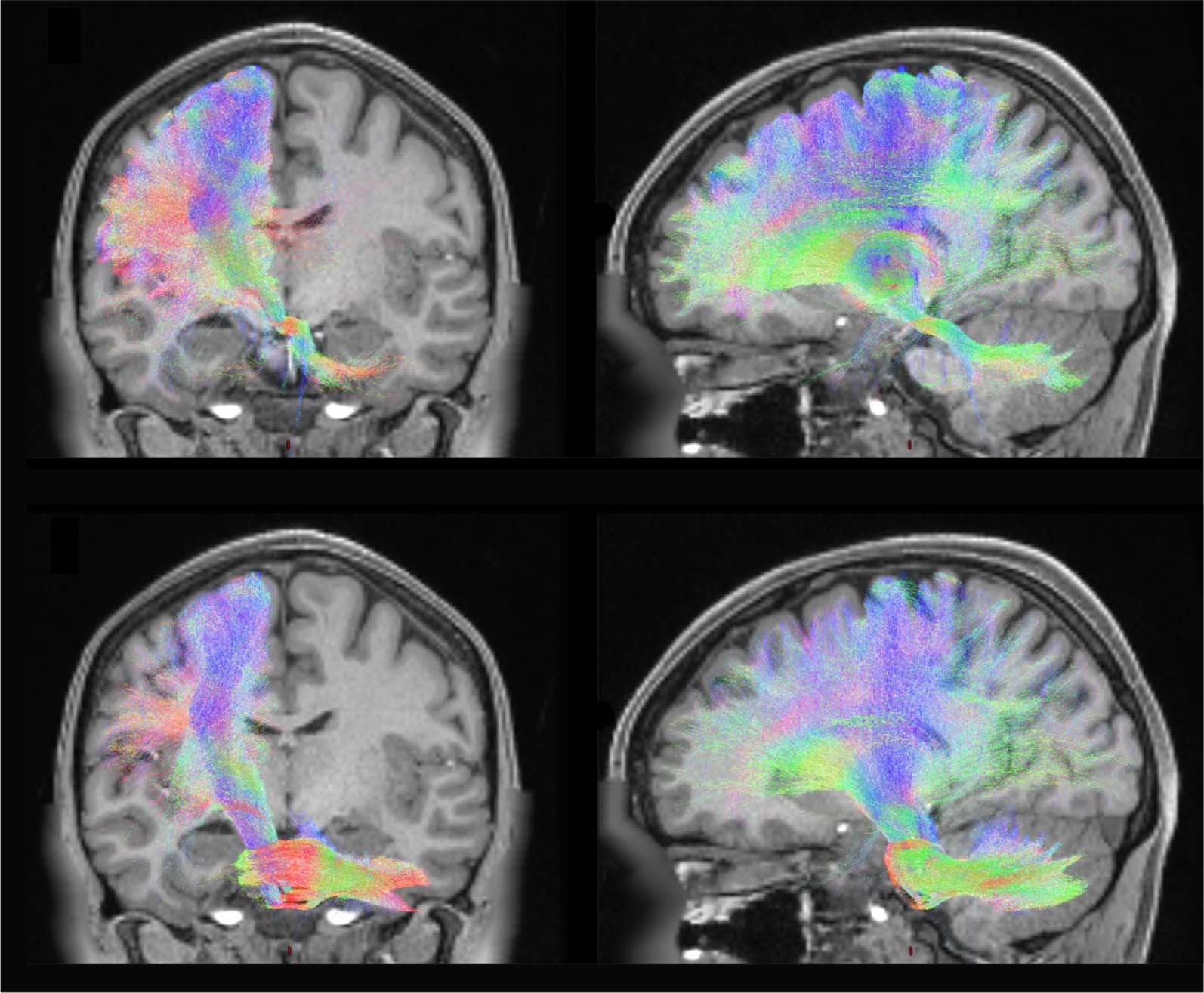
Figure 5. White matter streamlines interconnecting the cerebellum and the cerebral cortex. (A) the cortico-ponto-cerebellar (CPC) tract sends
projections from the cortex via the pontine nuclei into the mossy fibres of the cerebellum; (B) the cerebello-thalamo-cortical (CTC) tract derives
from the deep cerebellar nuclei, which project back via the core thalamic nuclei to the cerebral cortex. Tracts created using mrtrix were
projected onto a T1-weighted structural image from individual 100307 from the Human Connectome Project (de-faced to preserve autonomy).
The colours of each tract represent the primary direction of each tract: blue = inferior-superior; red = left-right; green = anterior-posterior.
f
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
a
_
0
0
3
0
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
cerebellar) regions to which they are connected by white matter projections. To create an esti-
mate of what these predicted BOLD responses should be, we created two template maps—one
for predicted cerebellar activity (estimated cerebellar activity: ACTX = WCBM . CPC) and one
for predicted cortical activity (estimated cortical activity: ACBM = WCTX . CTC)—by multiply-
ing the cortico-cerebellar structural connectivity matrices with the preprocessed BOLD pattern
observed during the three different trial types. We then correlated these prediction vectors with
the actual BOLD patterns in the respective regions. If the observed patterns of activity were
similar, we can conclude that BOLD activity patterns were intimately related to the reciprocal
structural connections between the cerebral cortex and cerebellum.
Across all three trial types, both cortico-cerebellar (via CPC; Figure 6C, circles) and
cerebello-cortical (via CTC; Figure 6D, squares) activity flow patterns were significantly greater
for actual versus randomly shuffled data (all p < 0.05), suggesting that functional activity was
coordinated by connections both from the cerebral cortex to the cerebellum (i.e., CPC) and
vice versa (i.e., CTC) across all tasks. Interestingly, despite the consistent positive relationships,
cerebello-cortical connections (i.e., CTC) were more robustly able to predict subsequent
cortical patterns than cortico-cerebellar connections (i.e., CPC), suggesting that the feedback
from the cerebellum to the cerebral cortex was more crucial for task performance. Finally, we
Network Neuroscience
851
Cortico-cerebellar dual tasking
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
a
_
0
0
3
0
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
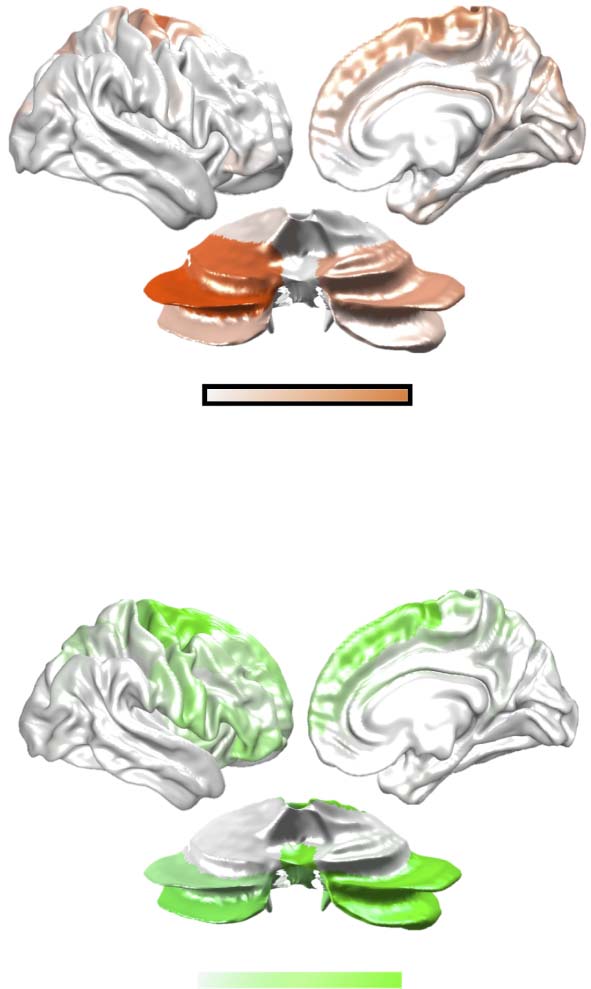
Figure 6. Cortico-cerebellar structure-function mapping across trial types. (A) normalized (in log10 of white matter connectivity) map of
projections from the cerebral cortex to cerebellum via CPC (orange). (B) normalized (in log10 of white matter connectivity) map of projections
from cerebellum to the cerebellar cortex via CTC (green). (C) activity flow mapping (Cole et al., 2016) between cerebellar BOLD patterns
predicted from CPC tract in balance (Bal, blue), calculation (Calc, orange), and dual-task (DT, purple) trials (circles); see Methods for details.
(D) the same for cortical BOLD patterns predicted from the CTC tract (squares). All activity flow map correlations were greater than permuted
null levels.
found that the match between ACTX/ACBM and the raw data was greater in correct versus
incorrect dual-task trials for both cerebral cortex (T = 2.397, p = 0.017) and cerebellum
(T = 2.049, p = 0.041), further confirming the importance of cortico-cerebellar interaction
for parallel processing.
DISCUSSION
In this study, we used systems-level neuroimaging analysis to demonstrate that robust interac-
tions between the cerebral cortex and cerebellum are associated with effective dual-task per-
formance. We hypothesized that, through distributed white matter pathways that interconnect
these major cortical systems, the brain can differentiate different task contexts so as to effec-
tively maintain the performance of two computational tasks in parallel. To test this hypothesis,
we analysed BOLD data from the cerebral cortex and cerebellum, and in doing so demon-
strated that dual-Task performance recruited heightened cerebellar activity (Figure 1) and func-
tional connectivity between the cerebral cortex and cerebellum (Figures 2 and 3) that was
linked to the balance between integration and segregation (Figure 4) and related to the struc-
tural connections between the cerebellum and cerebral cortex (Figures 5 and 6). Together,
these results highlight the importance of systems-level interactions in the manifestation of
complex cognitive capacities.
Our results clearly demonstrate that models that incorporate the cerebellum and its massive,
high-dimensional architecture provide a more parsimonious account for how the brain can
Network Neuroscience
852
Cortico-cerebellar dual tasking
Automaticity:
The ability to perform a behaviour
without deliberate, focussed
attention.
balance the challenges inherent with parallel processing (Balsters & Ramnani, 2011; D’Angelo &
Casali, 2013; Shine, 2020; Wu et al., 2013). The distributed circuits that interconnect the cerebral
cortex and cerebellum are optimally set up to fulfil this capacity. Specifically, the major output of
the cerebral cortex—layer V PT-type pyramidal neurons—provides the primary afferent input to
the cerebellar cortex (i.e., granule cells), by way of the pontine nuclei (D’Angelo & Casali, 2013;
Kratochwil et al., 2017; Shine, 2020). Following a massive dimensionality expansion that has
been argued to facilitate pattern separation (Cayco-Gajic & Silver, 2019), the outputs of the
cerebellum (the deep cerebellar nuclei) send large glutamatergic projections to the ventral tier
of the thalamus (Prevosto & Sommer, 2013), wherein they innervate the cerebral cortex. The
thalamic targets of the cerebellum then go on to drive activity in the cerebral cortex, typically
in a high-frequency, precise fashion (Nashef et al., 2022) that we have argued form the basis
of relatively automatic modes of behaviour (Shine, 2020; Shine & Shine, 2014). Here, we
extend these functional neuroanatomical principles to incorporate the completion of
challenging dual tasks, thus augmenting and reinforcing conclusions from previous functional
neuroimaging work on dual-task performance (Balsters & Ramnani, 2011; Shine & Poldrack,
2017; Wu et al., 2013). We anticipate that similar patterns will be observed in future
experiments that interrogate different types of dual tasks, particularly those in which one (or
both) of the tasks is capable of relative automaticity. Whether such automaticity benefits
extend to purely perceptual tasks, such as the attentional blink (Sergent & Dehaene, 2004), is
an interesting open question for future work.
The topological signature of functional networks estimated from BOLD data have previ-
ously been linked to effective performance on cognitive tasks. For instance, an integrated brain
has been linked to the completion of a range of complex tasks, such as those that probe work-
ing memory (Cruzat et al., 2018; Fransson et al., 2018; Shine et al., 2016), logical reasoning
(Hearne et al., 2017), and attentional tracking (Mäki-Marttunen, 2021; Wainstein et al., 2021).
In contrast, a relatively segregated functional network has been linked to relative sensorimotor
automaticity (Bassett et al., 2015; Mohr et al., 2016), as well as to attentional vigilance
(Sadaghiani et al., 2015). Our results are consistent with the spectrum implied by these pre-
vious results—the balance task, which presumably tapped into relatively well-learned behav-
iours, was associated with a segregated functional network; and the calculation task, which
likely required more focussed, flexible attention, was associated with a relatively integrated
network. Interestingly, although the dual-task trials were arguably more challenging than the
calculation trials on their own, the topology of the network actually demonstrated a balance
between integration and segregation, suggesting that performing tasks in parallel requires an
ability to avoid topological extremes, perhaps so as to maximise information-processing capa-
bilities (Sporns, 2013). In addition, there are theoretical reasons to believe that the finite nature
of biological networks may imbue specific limits on the number of possible tasks that can be
run in parallel, although we expect that the high-dimensional architecture of the cerebellum
(Cayco-Gajic & Silver, 2019) will likely boost this capacity, particularly as a function of expe-
rience (Shine, 2020; Shine & Shine, 2014). Precisely which systems in the brain help to control
this balance remains an open question; however, there are intriguing results that suggest that
the neuromodulatory system may play a crucial role in this process (Breton-Provencher et al.,
2022; Shine, 2020; Shine et al., 2021).
Systems-level neuroimaging analysis provides an integrated perspective of cognitive capac-
ities; however, BOLD dynamics are necessarily indirect, that is, they don’t measure neural
activity directly, but rather filtered through the low-dimensional lens of perfusion (Aquino
et al., 2014; Pang et al., 2016). While the BOLD signal remains a robust measurement for
neural signalling (Attwell & Iadecola, 2002; Moore & Cao, 2008), it only reveals a part of
Network Neuroscience
853
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
a
_
0
0
3
0
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
how the brain functions. This is particularly true for the cerebellar cortex, whose complex,
convoluted anatomy (Caligiore et al., 2016; D’Angelo & Casali, 2013) and idiosyncratic firing
properties (Khilkevich et al., 2018; Kostadinov et al., 2019; Person & Raman, 2011) render
simple, linear readouts of neural activity from BOLD problematic. Specifically, there is evi-
dence to suggest that BOLD measurements in the cerebellar cortex predominantly track activ-
ity in the mossy fibre pathway (via the CPC; Caesar et al., 2003; Mathiesen et al., 2000),
whereas outputs from the Purkinje cells (via the CTC) are more difficult to characterize with
BOLD signalling (Diedrichsen et al., 2019; Thomsen et al., 2009). While this does suggest
caution with respect to the interpretation of our results, it makes the presence of robust
cerebello-cortical activity flow mapping via the CTC (Figure 6D) all the more fascinating of
a result, as it suggests that the fate of the Purkinje cells is relatively sealed by the specific pat-
tern of mossy fibre inputs that they received, although we anticipate that this mapping is likely
augmented by the process of learning, that is, it should be less profound when facing highly
novel task contexts. Irrespectively, we hope that by consolidating analysis from multiple neu-
roimaging techniques, we have provided a robust illustration of changes to cortico-cerebellar
circuits during a parallel processing task.
The capacity to perform tasks in parallel clearly scales positively with experience. In the
future, it will be fascinating to examine the interactions between the cerebral cortex and cer-
ebellum as individuals learn to perform individual tasks to relative automaticity. There is robust
empirical previous work linking cerebellar output with highly overtrained behaviours in
rodents (Callu et al., 2013). Similar arguments have been made when analysing automaticity
in the performance of challenging cognitive tasks (Balsters & Ramnani, 2011). Interestingly,
there is also evidence suggesting that, over the course of learning a simple sensorimotor task,
the brain shifts from a relatively integrated to a segregated architecture (Bassett et al., 2015;
Mohr et al., 2016). This suggests a novel prediction: the extent to which a particular task has
been well learned will lead to relative segregation of the topological network signature of the
brain, which in turn will make the task easier to automatise, and hence to combine success-
fully with other, more challenging dual tasks.
One factor that was not well controlled in this study was cognitive load, which is known to
play an important role in our capacity to perform multiple tasks in parallel (Just et al., 2001;
Michael et al., 2001; Whelan, 2007). Simply put, it is much easier to perform two tasks simul-
taneously if (at least) one of the tasks is either highly automatic or is sufficiently easy that its
performance requires little to no focussed attention (Fischer & Plessow, 2015). In these cases,
the simpler or more automatic task can be performed with minimal awareness, freeing up
higher cognitive systems to aid in the completion of the second, harder/more challenging task.
In our study, the balance task was presumed to be easier than the calculation task, as partic-
ipants were expected to have been unlikely to have practiced the subtraction of the digit “7”
from random large numbers, whereas balance is something many of us perform so much as to
take it for granted. In future studies, it will be important to attempt to stack together multiple
tasks that are difficult to perform in the same manner, such as comprehending an auditory
stream while performing a calculation on concurrent visual input. Although we anticipate that
both the cerebellum and cerebral cortex would be engaged in such a task, it is less likely that
effective performance would be as crucially dependent on their interaction, as the mechanism
we propose invokes the cerebellar-mediated anticipation of expected consequences as a
means for freeing up higher cognitive resources (Ramnani, 2014; Shine, 2020; Shine & Shine,
2014). It is not currently clear whether these anticipatory processes are as important in the
more deliberate, flexible stages of cognitive processing that would be required to complete
two more deliberate cognitive tasks simultaneously.
Network Neuroscience
854
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
a
_
0
0
3
0
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
Here, we have demonstrated that dynamic interactions between the cerebral cortex and
cerebellum are critically related to the performance of a challenging dual task. Future research
is required to determine whether similar principals are related to parallel processing of other
simultaneous cognitive and perceptual challenges, as well as across distinct spatiotemporal
scales.
METHODS
Experimental Setup
The functional data from this study arose from a re-analysis of a previously published dataset
(Papegaaij et al., 2017); here, we will include the minimal information required to interpret the
results, and point the interested reader to the original study for full details. 50 healthy female par-
ticipants (mean age = 49 ± 20 years; Papegaaij et al., 2017) lay supine in the MRI scanner with their
feet against a custom-made force platform attached to the MRI bed (Figure 1A; sample frequency
of 100 Hz), with the position of the force platform was adjusted to subject height. To minimize
excessive head movement, participants were pulled towards the force platform using thick elastic
ropes attached to a hip belt (Papegaaij et al., 2017). A four-button device was placed underneath
the right hand for the calculation task. The tasks were projected onto a white screen placed at the
head of the scanner. Participants could see the screen via a mirror attached to the head coil.
During the balance task, an avatar in the shape of a woman was displayed on the screen. The
avatar swayed forward and backward. Participants were instructed to try to keep the avatar in
the upright position by increasing or decreasing the level of plantar flexion force measured by
the load cell. As in normal standing, increasing the plantar flexion force led to a backward sway
(and v.v.). At the start of every balance condition, participants were given 2 seconds to bring the
avatar in the upright position. After these 2 seconds, a disturbance signal was added, causing
the avatar to sway forward and backward. To keep the avatar upright, participants had to
counteract these disturbances. The disturbance signal was made by combining 15 sinusoidal
signals with random phases and with frequency characteristics based on an average frequency
spectrum of centre of pressure movement during upright standing (0.025–1 Hz), measured in
10 young and 10 old adults. The maximum amplitude of the disturbance was ±30°. The error
for each balance trial was created by calculating the sum of the root-mean-squared error
between the optimal balanced avatar (i.e., 900) and the position of the actual avatar. Trials were
subsequently median split to identify ‘good’ and ‘bad’ balance trials.
The calculation task consisted of serial subtractions with increments of seven—at the begin-
ning of each trial, a number between 50 and 100 was projected on the screen for 2 seconds,
after which a plus sign was displayed on the screen and a beep was generated every 3 to
4 seconds through an MRI compatible headphone (MR confon Optime 1, Magdeburg,
Germany), with a total of four beeps per trial. Participants were instructed to subtract the num-
ber 7 with every beep. At the end of each trial, four answer possibilities were displayed on the
screen: one indicating the correct answer, two erroneous answers, and the option that none of
the other answers is correct. Participants indicated which answer they thought was correct by
pressing the corresponding button of the four-button device.
During the dual-task condition, subjects performed the balance and calculation tasks
simultaneously. The distribution of RMS errors in the balance trials and dual-task trials were
compared using a Kolmogorov–Smirnov test.
An fMRI block design was used to alternate between the three conditions: balance, calcu-
lation, and dual task. Every participant performed 12 blocks, each block including one trial of
Network Neuroscience
855
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
a
_
0
0
3
0
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
each condition (thus three trials), with the order of the conditions randomized, both across
blocks and between subjects. At the end of every block a 15-second rest period was given
in which the participants fixated their gaze on a plus sign.
MRI Acquisition and Preprocessing
Brain imaging was performed on a 3-T SIEMENS Magnetom Skyra System (Siemens, Erlangen,
Germany) with a 20-channel head/neck coil. For functional scans, a T2*-weighted multiband
gradient echo-planar imaging sequence was used (TR = 700 msec, TE = 30 msec, flip angle =
55°, 48 axial slices, slice thickness = 3 mm, no gap, in-plane resolution 3 × 3 mm) (Feinberg
et al., 2010). After the functional scanning session, a high-resolution magnetization-prepared
rapid acquisition gradient echo (MPRAGE) T1-weighted sequence (TR = 2,100 msec, TE =
4.6 msec, TI = 900 msec, flip angle = 8°, 192 contiguous slices, voxel resolution 1 mm3,
FOV = 256 × 256 × 192 mm, iPAT factor of 2) was obtained in sagittal orientation. These ana-
tomical scans were used to coregister the functional runs using SPM 12. The anatomical scan
was segmented using the SPM tissue probability maps. The functional data were preprocessed
as part of a different study (Papegaaij et al., 2017). For each subject, interscan movement was
corrected by realigning and unwarping the data, with the first scan as a reference. All func-
tional scans were then coregistered to the anatomical scan and normalized to the Montreal
Neurological Institute (MNI) template brain via the forward deformations revealed by the
structural segmentation. Movement in the scanner was assessed by calculating framewise dis-
placement (FD) from the derivatives of the six rigid body realignment parameters estimated
during standard volume realignment, as well as the root-mean-square change in BOLD signal
from volume to volume (aka DVARS). Across the cohort, head motion was found to be minimal
(group mean FD = 0.183 ± 0.08 mm; group mean DVARS = 0.811 ± 0.13).
Temporal artifacts were identified in each dataset by calculating FD from the derivatives of
the six rigid body realignment parameters estimated during standard volume realignment
(Power et al., 2014), as well as the root-mean-square change in BOLD signal from volume
to volume (DVARS). Frames associated with FD > 0.25 mm or DVARS > 2.5% were identified;
cependant, as no participants were identified with greater than 10% of the resting time points
exceeding these values, no trials were excluded from further analysis.
Brain Parcellation
Following preprocessing, the mean time series was extracted from 400 predefined cortical par-
cels using the Schaefer atlas (Schaefer et al., 2018) et 28 predefined cerebellar parcels from
the SUIT atlas (Diedrichsen, 2006) (cerebellar nuclei were not included). The mean BOLD
signal intensity from each region was extracted and then used for subsequent analyses.
General Linear Model and Principal Component Analysis
A general linear model was fit to preprocessed, parcellated BOLD data with separate terms
modelling each trial type (c'est à dire., équilibre, calculation, and dual task). The event time series used
to analyse the task included a convolution with a canonical haemodynamic response function.
The proportion of cerebellar regions associated with positive cerebellar β-values was com-
pared across balance, calculation and dual-task trials using a χ2 test with degrees of freedom =
(rows – 1) × (columns – 1) = (3 – 1) × (2 – 1) = 2.
The average β-value for the balance and calculation trials were demeaned and analysed
with a principal component analysis. The coefficient of the leading principal component
was correlated with the mean β map from the balance and calculation trials to demonstrate
Neurosciences en réseau
856
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
t
/
/
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
t
.
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
its utility as a linear decoder between balance and calculation. The dot product between the
dual-task β map for each subject and the leading principal component was calculated, et
then subjected to a one-sample t test to determine whether the loading was more similar to
calculation (positive loadings) or balance (negative loadings).
Time-Varying Functional Connectivity
We used the multiplication of temporal derivatives (MTD) approche (Shine et al., 2015) à
calculate time-resolved dynamic functional connectivity between the selected ROIs; code is
freely available at https://github.com/macshine/coupling/ with a window size of 20 TRs (résultats
were stable for window sizes of 10–50 TR). For each node, n, with time points, t, a vector of
t − 1 temporal derivatives was calculated and normalized (temporal derivatives divided by the
standard deviation of temporal derivatives, p). Alors, we created a matrix of functional cou-
pling between the ith and jth nodes for each time point, by multiplying the temporal deriva-
tives of each pair of nodes across each time point.
(cid:2)
(cid:2)
(cid:3)
(cid:3)
X
tþw
1
(1)
MTDijt ¼
w
t
dtit (cid:2) dtjt
(cid:2) σdtj
σdti
where dt is the first temporal derivative of the i th and j th time series, and σ standard deviation
of the temporal derivative, w is the window length of the simple moving average (Shine et al.,
2015). The MTD values for the cortico-cerebellar system (c'est à dire., 400 × 28 = 11,200 edges) étaient
entered into a similar general linear model to the cortico-cerebellar BOLD values, with a per-
mutation test (5,000 iterations) used to test for statistical significance.
Modularity Maximization
The Louvain modularity algorithm from the Brain Connectivity Toolbox (BCT; Rubinov &
De la Sporn, 2010; https://www.brain-connectivity-toolbox.net) was used on the neural network
edge weights to estimate community structure. The Louvain algorithm iteratively maximizes
the modularity statistic, Q, for different community assignments until the maximum possible
score of Q has been obtained (see Equation 2). The modularity of a given network is therefore
a quantification of the extent to which the network may be subdivided into communities with
stronger within-module than between-module connections.
(cid:4)
X
w þ
ij
(cid:5)
δMi Mj
(cid:5)
δMi Mj
QT ¼
− eþ
ij
− e−
ij
w −
ij
X
(2)
−
(cid:4)
1
υþ þ υ−
1
υþ
ij
ij
where v is the total weight of the network (sum of all negative and positive connections), wij is
the weighted and signed connection between regions i and j, eij is the strength of a connection
divided by the total weight of the network, and δMiMj
is set to 1 when regions are in the same
community and 0 otherwise; + and − superscripts denote all positive and negative connec-
tion, respectivement.
For each epoch, we assessed the community assignment for each region 500 times and a
consensus partition was identified using a fine-tuning algorithm (BCT). We calculated all graph
theoretical measures on unthresholded, weighted, and signed connectivity matrices (Rubinov
& De la Sporn, 2010). The stability of the γ parameter was estimated by iteratively calculating the
modularity across a range of γ values (0.5–2.5; mean Pearson’s r = 0.859 ± 0.01) on the
time-averaged connectivity matrix for each subject—across iterations and subjects, a γ value
de 1.0 was found to be the least variable, and hence was used for the resultant topological
analyses.
Neurosciences en réseau
857
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
t
/
/
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
t
.
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
Participation Coefficient
The participation coefficient, PC, quantifies the extent to which a region connects across all
modules (c'est à dire., between-module strength) and has previously been used to successfully charac-
terize hubs within brain networks (Shine et al., 2016, 2019). The PC for each region was
calculated within each temporal window using Equation 3, where kisT is the strength of the
positive connections of region i to regions in module s at time T, and kiT is the sum of strengths
of all positive connections of region i at time T. Negative connections were discarded prior to
calculation. The PC of a region is therefore close to 1 if its connections are uniformly distrib-
uted among all the modules and 0 if all of its links are within its own module.
PC ¼ 1 −
X
nM
s¼1
(cid:6) (cid:7)2
κisT
κiT
(3)
The PC for each parcel was compared across balance, calculation and dual-task trials using
paired t tests (FDR q = 0.05).
Diffusion MRI Analysis
Data were selected from a single 26–30-year-old female subject from the HCP (code: 100307).
The minimally processed HCP diffusion dataset (which included correction for motion,
susceptibility distortions, gradient nonlinearity and eddy currents) were subject to additional
image processing, which multishell multitissue constrained spherical deconvolution to gener-
ate the fibre orientation distribution (FOD) in each voxel (Jeurissen et al., 2014; Tournier et al.,
2004, 2007). These steps were implemented in accordance with previous work (Civier et al.,
2019) and were performed using the MRtrix software package (https://www.mrtrix.org;
Tournier et al., 2012, 2019).
The T1-weighted images were used to generate a so-called ‘five-tissue-type’ (5TT) image
(R.. E. Smith et al., 2012) using FSL (S. M.. Smith et al., 2004); the 5TT image classifies the voxel
into one of five tissue types: cortical grey matter, subcortical grey matter, white matter, cere-
brospinal fluid, and ‘5th type’ (par exemple., pathology). The FOD data and the 5TT image were used to
generate 120 million streamlines using the anatomically constrained tractography framework
(R.. E. Smith et al., 2012), using dynamic and the second-order integration over fibre orientation
distributions (iFOD2; Tournier et al., 2012) probabilistic fibre-tracking algorithm, using default
MRtrix parameters, with the exception of FOD cutoff 0.06, maximum length 250 mm, step size
1 mm, and backtrack specified. This set of streamlines is referred to as the whole-brain-
tractogram thereafter.
Spécifiquement, we calculated from a highly curated tractography rendering of the cerebro-
cerebellar loop, after thresholding the streamlines to eliminate possible spurious tracts. Un
average tract obtained from 5 à 10 à 28 sujets, thresholded to represent the group, may
lose the finer details of the connectome that are key when using a ∼400 region grey matter
parcellation atlas as in this work. D'autre part, the connectome from the union of the
tracts, if not thresholded, would inflate this finer connectivity (if thresholded, these connec-
tions would be downweighted). The impact of averaging individual subject streamlines on
the actual connectome has to be demonstrated in a separate study, as the high variability of
the streamlines is likely to correspond to a fairly stable connectome.
The cerebello-thalamo-cortical (CTC) and cortico-ponto-cerebellar (CPC) tracts were
extracted from the whole-brain tractogram by using contralateral cerebral and cerebellar
cortices, cerebellar peduncles, contralateral red nuclei, and thalami as regions of interest
Neurosciences en réseau
858
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
/
/
t
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
.
t
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
(for more details, see Palesi et al., 2015, 2017). To define the strength of the cerebellar con-
nectivity with each of brain parcel, the log10 of the number of streamlines was used to weight
the CTC and CPC tracts (Abos et al., 2019; Palesi et al., 2021). To ensure that the single-subject
connectome was representative of the group-level parcellation, we calculated the DICE
coefficient between the mean map of both the CTC and CPC tracts (L and R) of a further 27
subjects in MNI space (including only those voxels that were common to at least 70% of sub-
projets, c'est, less than the 90,000,000 streamlines used for the individual connectome); le
DICE was 0.7, suggesting strong correspondence between our single subject (who preserved
the fine-scale nature of the connectome) and the group template.
Cortico-Cerebellar Activity Flow Mapping
To determine whether cortico-cerebellar interactions could transform cortical or cerebellar
task-evoked activity into respective cerebello-cortical task activity, we modified the activity
flow mapping procedure (Cole et al., 2016) to incorporate estimates of cortico-cerebellar
(CPC) and cerebello-cortical (CTC) structural connectivity. Spécifiquement, for each trial type,
block and subject, we calculated:
ACTX ¼ W t
CBM
⋅ CPC
ACBM ¼ W t
CTX
⋅ CTC
(4)
(5)
where Wt is the evoked response estimate for every cortical (WCTX) or cerebellar (WCBM)
parcel, CPC and CTC are the structural connectivity matrices described above, and ACTX
and ACBM are the predicted activity pattern for each subgroup. For each trial type, block
and subject, the predicted cortical and cerebellar activity patterns were then empirically com-
pared to the observed activity patterns using Pearson correlations. A series of t tests were used
to compare the Pearson’s correlation loadings, with the nonmatching predictions (par exemple., en utilisant
the cortical BOLD for balance trials to predict cerebellar BOLD for calculation trials) used a
simple null model that contained all the same spectral features but spatiotemporal sequences
that did not match the data. Enfin, we created separate null distributions following a random
permutation (Nichols & Holmes, 2002) of both CPC and CTC, separately (each with 5,000
iterations).
REMERCIEMENTS
Data were provided by the Human Connectome Project, WU-Minn Consortium (PI: David Van
Essen and Kamil Ugurbil; 1U54MH091657) funded by the 16 NIH Institutes and Centers that
support the NIH Blueprint for Neuroscience Research; and by the McDonnell Center for
Systems Neuroscience, Washington University.
INFORMATIONS À L'APPUI
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00308.
CONTRIBUTIONS DES AUTEURS
Eli J. Müller: Conceptualisation; Enquête; Méthodologie; Rédaction – révision & édition.
Fulvia Palesi: Analyse formelle; Méthodologie; Rédaction – révision & édition. Kevin Y. Hou: Formal
analyse; Méthodologie. Joshua Tan: Rédaction – révision & édition. Thomas Close: En écrivant -
revoir & édition. Claudia A. M.. Gandini Wheeler-Kingschott: Enquête; Rédaction – révision
Neurosciences en réseau
859
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
t
/
/
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
t
.
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
& édition. Egidio D’Angelo: Enquête; Rédaction – révision & édition. Fernando Calamante:
Analyse formelle; Enquête; Méthodologie; Rédaction – révision & édition. James Shine: Con-
ceptualization; Conservation des données; Analyse formelle; Enquête; Méthodologie; Project adminis-
tration; Ressources; Logiciel; Surveillance; Validation; Visualisation; Rédaction – ébauche originale;
Rédaction – révision & édition.
INFORMATIONS SUR LE FINANCEMENT
James Shine, National Health and Medical Research Council (https://dx.doi.org/10.13039
/501100000925), Pièce d'identité: 1193857.
RÉFÉRENCES
Abos, UN., Baggio, H. C., Segura, B., Campabadal, UN., Uribe, C.,
Giraldo, D. M., Perez-Soriano, UN., Muñoz, E., Compta, Y.,
Junque, C., & Marti, M.. J.. (2019). Differentiation of multiple sys-
tem atrophy from Parkinson’s disease by structural connectivity
derived from probabilistic tractography. Rapports scientifiques, 9(1),
16488. https://doi.org/10.1038/s41598-019-52829-8, PubMed:
31712681
Alahmadi, UN. UN. S., Samson, R.. S., Gasston, D., Pardini, M., Friston,
K. J., D’Angelo, E., Toosy, UN. T., & Wheeler-Kingshott, C. UN. M..
(2016). Complex motor task associated with non-linear BOLD
responses in cerebro-cortical areas and cerebellum. Brain Struc-
ture and Function, 221(5), 2443–2458. https://doi.org/10.1007
/s00429-015-1048-1, PubMed: 25921976
Aquino, K. M., Robinson, P.. UN., & Drysdale, P.. M.. (2014). Spatio-
temporal hemodynamic response functions derived from physi-
œnologie. Journal of Theoretical Biology, 347, 118–136. https://est ce que je
.org/10.1016/j.jtbi.2013.12.027, PubMed: 24398024
Attwell, D., & Iadecola, C. (2002). The neural basis of functional
brain imaging signals. Tendances des neurosciences, 25(12), 621–625.
https://doi.org/10.1016/S0166-2236(02)02264-6, PubMed:
12446129
Balsters, J.. H., & Ramnani, N. (2011). Cerebellar plasticity and the
automation of first-order rules. Journal des neurosciences, 31(6),
2305–2312. https://doi.org/10.1523/JNEUROSCI.4358-10.2011,
PubMed: 21307266
Bar-Gad, JE., Morris, G., & Bergman, H. (2003). Information pro-
cessation, dimensionality reduction and reinforcement learning in
the basal ganglia. Progrès en neurobiologie, 71(6), 439–473.
https://doi.org/10.1016/j.pneurobio.2003.12.001, PubMed:
15013228
Bassett, D. S., Lequel, M., Wymbs, N. F., & Grafton, S. T. (2015).
Learning-induced autonomy of sensorimotor systems. Nature
Neurosciences, 18(5), 744–751. https://doi.org/10.1038/nn.3993,
PubMed: 25849989
Breton-Provencher, V., Drummond, G., Feng, J., Li, Y., & Sur, M..
(2022). Spatiotemporal dynamics of norepinephrine during
learned behavior. Nature, 606(7915), 732–738. https://doi.org
/10.1038/s41586-022-04782-2, PubMed: 35650441
Caesar, K., Gold, L., & Lauritzen, M.. (2003). Context sensitivity of
activity-dependent increases in cerebral blood flow. Procédure
of the National Academy of Sciences, 100(7), 4239–4244. https://
doi.org/10.1073/pnas.0635075100, PubMed: 12655065
Caligiore, D., Pezzulo, G., Baldassarre, G., Bostan, UN. C.,
Strick, P.. L., Doya, K., Helmich, R.. C., Dirkx, M., Houk, J.,
Jörntell, H., Lago-Rodriguez, UN., Galea, J.. M., Miall, R.. C.,
Popa, T., Kishore, UN., Verschure, P.. F. M..
J., Zucca, R., &
(2016). Consensus paper: Towards a systems-
Herreros,
level view of cerebellar
fonction: The interplay between
cerebellum, basal ganglia, and cortex. The Cerebellum, 16(1),
203–229. https://doi.org/10.1007/s12311-016-0763-3, PubMed:
26873754
je.
Callu, D., Lopez, J., & El Massioui, N. (2013). Cerebellar deep
nuclei involvement in cognitive adaptation and automaticity.
Apprentissage & Mémoire, 20(7), 344–347. https://doi.org/10.1101/lm
.030536.113, PubMed: 23772087
Casiraghi, L., Alahmadi, UN. UN. S., Monteverdi, UN., Palesi, F., Castellazzi,
G., Savini, G., Friston, K., Gandini Wheeler-Kingshott, C. UN. M.,
& D’Angelo, E. (2019). I see your effort: Force-related BOLD
effects in an extended action execution–observation network
involving the cerebellum. Cortex cérébral, 29(3), 1351–1368.
https://doi.org/10.1093/cercor/bhy322, PubMed: 30615116
Castellazzi, G., Bruno, S. D., Toosy, UN. T., Casiraghi, L., Palesi, F.,
Savini, G., D’Angelo, E., & Wheeler-Kingshott, C. UN. M.. G.
(2018). Prominent changes in cerebro-cerebellar functional
connectivity during continuous cognitive processing. Frontiers
in Cellular Neuroscience, 12, 331. https://doi.org/10.3389/fncel
.2018.00331, PubMed: 30327590
Castellazzi, G., Palesi, F., Casali, S., Vitali, P., Sinforiani, E.,
Wheeler-Kingshott, C. UN. M., & D’Angelo, E. (2014). A compre-
hensive assessment of resting state networks: Bidirectional
modification of functional integrity in cerebro-cerebellar net-
works in dementia. Frontiers in Neuroscience, 8, 223. https://
doi.org/10.3389/fnins.2014.00223, PubMed: 25126054
Cayco-Gajic, N. UN., & Silver, R.. UN. (2019). Re-evaluating circuit
mechanisms underlying pattern separation. Neurone, 101(4),
584–602. https://doi.org/10.1016/j.neuron.2019.01.044,
PubMed: 30790539
Civier, O., Forgeron, R.. E., Yeh, C.-H., Connelly, UN., & Calamante, F.
(2019). Is removal of weak connections necessary for graph-
theoretical analysis of dense weighted structural connectomes
from diffusion MRI? NeuroImage, 194, 68–81. https://est ce que je.org/10
.1016/j.neuroimage.2019.02.039, PubMed: 30844506
Cole, M.. W., Ito, T., Bassett, D. S., & Schultz, D. H. (2016). Activity
flow over resting-state networks shapes cognitive task
Neurosciences en réseau
860
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
/
t
/
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
.
t
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
activations. Neurosciences naturelles, 19(12), 1718–1726. https://est ce que je
.org/10.1038/nn.4406, PubMed: 27723746
Cruzat, J., Déco, G., Tauste-Campo, UN., Principe, UN., Costa, UN.,
Kringelbach, M.. L., & Rocamora, R.. (2018). The dynamics of
human cognition: Increasing global integration coupled with
decreasing segregation found using iEEG. NeuroImage, 172,
492–505. https://doi.org/10.1016/j.neuroimage.2018.01.064,
PubMed: 29425897
D’Angelo, E. (2019). The cerebellum gets social. Science, 363(6424),
229–229. https://doi.org/10.1126/science.aaw2571, PubMed:
30655429
D’Angelo, E., & Casali, S. (2013). Seeking a unified framework for
cerebellar function and dysfunction: From circuit operations to
cognition. Frontiers in Neural Circuits, 6, 116. https://est ce que je.org/10
.3389/fncir.2012.00116, PubMed: 23335884
D’Angelo, E., Mapelli, L., Casellato, C., Garrido, J.. UN., Luque, N.,
Monaco, J., Prestori, F., Pedrocchi, UN., & Ros, E. (2016). Distrib-
uted circuit plasticity: New clues for the cerebellar mechanisms
of learning. The Cerebellum, 15(2), 139–151. https://est ce que je.org/10
.1007/s12311-015-0711-7, PubMed: 26304953
Diedrichsen, J.. (2006). A spatially unbiased atlas template of the
human cerebellum. NeuroImage, 33(1), 127–138. https://est ce que je
.org/10.1016/j.neuroimage.2006.05.056, PubMed: 16904911
Diedrichsen, J., King, M., Hernandez-Castillo, C., Séréno, M., &
Ivry, R.. B. (2019). Universal transform or multiple functionality?
Understanding the contribution of the human cerebellum across
task domains. Neurone, 102(5), 918–928. https://est ce que je.org/10.1016
/j.neuron.2019.04.021, PubMed: 31170400
Eisenreich, B. R., Akaishi, R., & Hayden, B. Oui. (2017). Control with-
out controllers: Toward a distributed neuroscience of executive
control. Journal des neurosciences cognitives, 29(10), 1684–1698.
https://doi.org/10.1162/jocn_a_01139, PubMed: 28430042
Feinberg, D. UN., Moeller, S., Forgeron, S. M., Auerbach, E., Ramanna,
S., Glasser, M.. F., Miller, K. L., Voiture d'Ugur, K., & Yacoub, E. (2010).
Multiplexed echo planar imaging for sub-second whole brain
FMRI and fast diffusion imaging. Plos un, 5(12), e15710.
https://doi.org/10.1371/journal.pone.0015710, PubMed:
21187930
Fischer, R., & Plessow, F. (2015). Efficient multitasking: Parallel
versus serial processing of multiple tasks. Frontières en psychologie,
6, 1366. https://doi.org/10.3389/fpsyg.2015.01366, PubMed:
26441742
Fransson, P., Schiffler, B. C., & Thompson, W. H. (2018). Brain net-
work segregation and integration during an epoch-related working
memory fMRI experiment. NeuroImage, 178, 147–161. https://est ce que je
.org/10.1016/j.neuroimage.2018.05.040, PubMed: 29777824
Hearne, L. J., Cocchi, L., Zalesky, UN., & Mattingley, J.. B. (2017).
Reconfiguration of brain network architectures between
resting-state and complexity-dependent cognitive reasoning.
Journal des neurosciences, 37(35), 8399–8411. https://est ce que je.org/10
.1523/JNEUROSCI.0485-17.2017, PubMed: 28760864
Ilg, W., Christensen, UN., Mueller, Ô. M., Goericke, S. L., Giese,
M.. UN., & Timmann, D. (2013). Effects of cerebellar lesions on
working memory interacting with motor tasks of different com-
plexities. Journal de neurophysiologie, 110(10), 2337–2349.
https://est ce que je.org/10.1152/jn.00062.2013, PubMed: 23966680
Jeurissen, B., Tournier, J.-D., Dhollander, T., Connelly, UN., & Sijbers,
J.. (2014). Multi-tissue constrained spherical deconvolution for
improved analysis of multi-shell diffusion MRI data. NeuroImage,
103, 411–426. https://doi.org/10.1016/j.neuroimage.2014.07
.061, PubMed: 25109526
John, Oui. J., Sawyer, K. S., Srinivasan, K., Müller, E. J., Munn, B. R., &
Briller, J.. M.. (2022). It’s about time: Linking dynamical systems
with human neuroimaging to understand the brain. Réseau
Neurosciences, 6(4), 960–979. https://doi.org/10.1162/netn_a
_00230, PubMed: 36875012
Just, M.. UN., Carpenter, P.. UN., Keller, T. UN., Emery, L., Zajac, H., &
Thulborn, K. R.. (2001). Interdependence of nonoverlapping cortical
systems in dual cognitive tasks. NeuroImage, 14(2), 417–426.
https://doi.org/10.1006/nimg.2001.0826, PubMed: 11467915
Khilkevich, UN., Zambrano, J., Richards, M.-M., & Mauk, M.. D.
(2018). Cerebellar implementation of movement sequences
through feedback. eLife, 7, e06262. https://doi.org/10.7554
/eLife.37443, PubMed: 30063004
Kostadinov, D., Beau, M., Blanco-Pozo, M., & Häusser, M.. (2019).
Predictive and reactive reward signals conveyed by climbing fiber
inputs to cerebellar Purkinje cells. Neurosciences naturelles, 22(6),
950–962. https://doi.org/10.1038/s41593-019-0381-8, PubMed:
31036947
Kratochwil, C. F., Maheshwari, U., & Rijli, F. M.. (2017). The long
journey of pontine nuclei neurons: From rhombic lip to cortico-
ponto-cerebellar circuitry. Frontiers in Neural Circuits, 11, 33.
https://doi.org/10.3389/fncir.2017.00033, PubMed: 28567005
Kuramoto, E., Furuta, T., Nakamura, K. C., Unzai, T., Hioki, H., &
Kaneko, T. (2009). Two types of thalamocortical projections from
the motor thalamic nuclei of the rat: A single neuron-tracing
study using viral vectors. Cortex cérébral, 19(9), 2065–2077.
https://doi.org/10.1093/cercor/bhn231, PubMed: 19174446
Mäki-Marttunen, V. (2021). Pupil-based states of brain integration
across cognitive states. Neurosciences, 471, 61–71. https://est ce que je
.org/10.1016/j.neuroscience.2021.07.016, PubMed: 34303781
Mathiesen, C., Caesar, K., & Lauritzen, M.. (2000). Temporal cou-
pling between neuronal activity and blood flow in rat cerebellar
cortex as indicated by field potential analysis. Journal of Phys-
iology, 523(1), 235–246. https://doi.org/10.1111/j.1469-7793
.2000.t01-1-00235.x, PubMed: 10673558
Michael, E. B., Keller, T. UN., Carpenter, P.. UN., & Just, M.. UN. (2001).
fMRI investigation of sentence comprehension by eye and by
ear: Modality fingerprints on cognitive processes. Cerveau humain
Cartographie, 13(4), 239–252. https://doi.org/10.1002/ hbm.1036,
PubMed: 11410952
Mohr, H., Wolfensteller, U., Bétzel, R.. F., Mišić, B., De la Sporn, O.,
Richiardi, J., & Ruge, H. (2016). Integration and segregation of
large-scale brain networks during short-term task automatization.
Communications naturelles, 7, 13217. https://est ce que je.org/10.1038
/ncomms13217, PubMed: 27808095
Moore, C. JE., & Cao, R.. (2008). The hemo-neural hypothesis: Sur
the role of blood flow in information processing. Journal of Neu-
rophysiology, 99(5), 2035–2047. https://est ce que je.org/10.1152/jn
.01366.2006, PubMed: 17913979
Nashef, UN., Cohen, O., Perlmutter, S. JE., & Prut, Oui. (2022). A cere-
bellar origin of feedforward inhibition to the motor cortex in
non-human primates. Cell Reports, 39(6), 110803. https://est ce que je
.org/10.1016/j.celrep.2022.110803, PubMed: 35545040
Nichols, T. E., & Holmes, UN. P.. (2002). Nonparametric permutation
tests for functional neuroimaging: A primer with examples.
Neurosciences en réseau
861
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
/
/
t
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
.
t
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
Human Brain Mapping, 15(1), 1–25. https://doi.org/10.1002
/hbm.1058, PubMed: 11747097
Palesi, F., De Rinaldis, UN., Castellazzi, G., Calamante, F., Muhlert,
N., Chard, D., Tournier, J.. D., Magenes, G., D’Angelo, E., &
Gandini Wheeler-Kingshott, C. UN. M.. (2017). Contralateral
cortico-ponto-cerebellar pathways reconstruction in humans in
vivo: Implications for reciprocal cerebro-cerebellar structural
connectivity in motor and non-motor areas. Rapports scientifiques,
7(1), 12841. https://doi.org/10.1038/s41598-017-13079-8,
PubMed: 28993670
Palesi, F., Ferrante, M., Gaviraghi, M., Misiti, UN., Savini, G., Lascialfari,
UN., D’Angelo, E., & Gandini Wheeler-Kingshott, C. UN. M.. (2021).
Motor and higher-order functions topography of the human
dentate nuclei identified with tractography and clustering
méthodes. Human Brain Mapping, 42(13), 4348–4361. https://
doi.org/10.1002/hbm.25551, PubMed: 34087040
Palesi, F., Lorenzi, R.. M., Casellato, C., Ritter, P., Jirsa, V., Gandini
Wheeler-Kingshott, C. UN. M., & D’Angelo, E. (2020). The impor-
tance of cerebellar connectivity on simulated brain dynamics.
Frontiers in Cellular Neuroscience, 14, 240. https://est ce que je.org/10
.3389/fncel.2020.00240, PubMed: 32848628
Palesi, F., Tournier, J.-D., Calamante, F., Muhlert, N., Castellazzi,
G., Chard, D., D’Angelo, E., & Wheeler-Kingshott, C. UN. M..
(2015). Contralateral cerebello-thalamo-cortical pathways with
prominent involvement of associative areas in humans in vivo.
Structure et fonction du cerveau, 220(6), 3369–3384. https://est ce que je
.org/10.1007/s00429-014-0861-2, PubMed: 25134682
Pang, J.. C., Robinson, P.. UN., & Aquino, K. M.. (2016). Response-
mode decomposition of spatio-temporal haemodynamics. Jour-
nal of The Royal Society Interface, 13(118), 20160253. https://
doi.org/10.1098/rsif.2016.0253, PubMed: 27170653
Papegaaij, S., Hortobágyi, T., Godde, B., Kaan, W. UN., Erhard, P., &
Voelcker-Rehage, C. (2017). Neural correlates of motor-cognitive
dual-tasking in young and old adults. Plos un, 12(12),
e0189025. https://doi.org/10.1371/journal.pone.0189025,
PubMed: 29220349
Person, UN. L., & Raman, je. M.. (2011). Purkinje neuron synchrony
elicits time-locked spiking in the cerebellar nuclei. Nature,
481(7382), 502–505. https://doi.org/10.1038/nature10732,
PubMed: 22198670
Petri, G., Musslick, S., Dey, B., Özcimder, K., Tourneur, D., Ahmed,
N. K., Willke, T. L., & Cohen, J.. D. (2021). Topological limits to
the parallel processing capability of network architectures. Nature
Physics, 17, 646–651. https://doi.org/10.1038/s41567-021-01170-x
Pezzulo, G., & Cisek, P.. (2016). Navigating the affordance land-
scape: Feedback control as a process model of behavior and cog-
nition. Tendances des sciences cognitives, 20(6), 414–424. https://est ce que je
.org/10.1016/j.tics.2016.03.013, PubMed: 27118642
Poldrack, R.. UN. (2012). The future of fMRI in cognitive neurosci-
ence. NeuroImage, 62(2), 1216–1220. https://est ce que je.org/10.1016/j
.neuroimage.2011.08.007, PubMed: 21856431
Poldrack, R.. UN., Kittur, UN., Kalar, D., Miller, E., Seppa, C., Gil, Y.,
Parker, D. S., Sabb, F. W., & Bilder, R.. M.. (2011). The Cognitive
Atlas: Toward a knowledge foundation for cognitive neurosci-
ence. Frontiers in Neuroinformatics, 5, 17. https://est ce que je.org/10
.3389/fninf.2011.00017, PubMed: 21922006
Power, J.. D., Mitra, UN., Laumann, T. O., Snyder, UN. Z., Schlaggar, B. L.,
& Petersen, S. E. (2014). Methods to detect, characterize, et
remove motion artifact in resting state fMRI. NeuroImage, 84,
320–341. https://doi.org/10.1016/j.neuroimage.2013.08.048,
PubMed: 23994314
Preuss, T. M., & Sage, S. P.. (2022). Evolution of prefrontal cortex.
Neuropsychopharmacology, 47(1), 3–19. https://est ce que je.org/10.1038
/s41386-021-01076-5, PubMed: 34363014
Prevosto, V., & Sommer, M.. UN. (2013). Cognitive control of move-
ment via the cerebellar-recipient thalamus. Frontiers in Systems
Neurosciences, 7, 56. https://doi.org/10.3389/fnsys.2013.00056,
PubMed: 24101896
Ramnani, N. (2006). The primate cortico-cerebellar system:
Anatomy and function. Nature Revues Neurosciences, 7(7),
511–522. https://doi.org/10.1038/nrn1953, PubMed: 16791141
Ramnani, N. (2014). Automatic and controlled processing in the
corticocerebellar system. In Cerebellar learning (pp. 255–285).
Amsterdam, Pays-Bas: Elsevier. https://est ce que je.org/10.1016
/B978-0-444-63356-9.00010-8, PubMed: 24916296
Rubinov, M., & De la Sporn, Ô. (2010). Complex network measures of
brain connectivity: Utilisations et interprétations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003,
PubMed: 19819337
Sadaghiani, S., Poline, J.-B., Kleinschmidt, UN., & D'Esposito, M..
(2015). Ongoing dynamics in large-scale functional connectivity
predict perception. Proceedings of the National Academy of Sci-
ences of the United States of America, 112(27), 8463–8468.
https://doi.org/10.1073/pnas.1420687112, PubMed: 26106164
Schaefer, UN., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N.,
Holmes, UN. J., Eickhoff, S. B., & Yeo, B. T. T. (2018). Local-global
parcellation of the human cerebral cortex from intrinsic func-
tional connectivity MRI. Cortex cérébral, 28(9), 3095–3114.
https://doi.org/10.1093/cercor/bhx179, PubMed: 28981612
Schmitz, T. W., & Duncan, J.. (2018). Normalization and the cholin-
ergic microcircuit: A unified basis for attention. Trends in Cogni-
tive Sciences, 22(5), 422–437. https://doi.org/10.1016/j.tics.2018
.02.011, PubMed: 29576464
Sergent, C., & Dehaene, S. (2004). Is consciousness a gradual phe-
nomenon? Evidence for an all-or-none bifurcation during the
attentional blink. Sciences psychologiques, 15(11), 720–728. https://
doi.org/10.1111/j.0956-7976.2004.00748.X, PubMed: 15482443
Briller, J.. M.. (2020). The thalamus integrates the macrosystems of
the brain to facilitate complex, adaptive brain network dynamics.
Progrès en neurobiologie, 199, 101951. https://est ce que je.org/10.1016/j
.pneurobio.2020.101951, PubMed: 33189781
Briller, J.. M., Bissett, P.. G., Cloche, P.. T., Kayevo, O., Balsters, J.. H.,
Gorgolewski, K. J., Moodie, C. UN., & Poldrack, R.. UN. (2016).
The dynamics of functional brain networks: Integrated network
states during cognitive task performance. Neurone, 92(2),
544–554. https://doi.org/10.1016/j.neuron.2016.09.018,
PubMed: 27693256
Briller, J.. M., Épreuve de rupture, M., Cloche, P.. T., Ehgoetz Martens, K. UN.,
Briller, R., Kayevo, O., De la Sporn, O., & Poldrack, R.. UN. (2019).
Human cognition involves the dynamic integration of neural
activity and neuromodulatory systems. Neurosciences naturelles,
22(2), 289–296. https://doi.org/10.1038/s41593-018-0312-0,
PubMed: 30664771
Briller, J.. M., Kayevo, O., Cloche, P.. T., Gorgolewski, K. J., Gilat, M., &
Poldrack, R.. UN. (2015). Estimation of dynamic functional connec-
tivity using multiplication of temporal derivatives. NeuroImage,
Neurosciences en réseau
862
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
t
/
/
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
.
t
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cortico-cerebellar dual tasking
122, 399–407. https://doi.org/10.1016/j.neuroimage.2015.07
.064, PubMed: 26231247
Briller, J.. M., Müller, E. J., Munn, B., Cabral, J., Moran, R.. J., &
Épreuve de rupture, M.. (2021). Computational models link cellular
mechanisms of neuromodulation to large-scale neural dynamics.
Neurosciences naturelles, 24(6), 765–776. https://est ce que je.org/10.1038
/s41593-021-00824-6, PubMed: 33958801
Briller, J.. M., & Poldrack, R.. UN. (2017). Principles of dynamic net-
work reconfiguration across diverse brain states. NeuroImage,
180, 396–405. https://doi.org/10.1016/j.neuroimage.2017.08
.010, PubMed: 28782684
Briller, J.. M., & Briller, R.. (2014). Delegation to automaticity: Le
driving force for cognitive evolution? Frontiers in Neuroscience, 8,
90. https://doi.org/10.3389/fnins.2014.00090, PubMed: 24808820
Forgeron, R.. E., Tournier, J.-D., Calamante, F., & Connelly, UN. (2012).
Anatomically-constrained tractography: Improved diffusion MRI
streamlines tractography through effective use of anatomical
information. NeuroImage, 62(3), 1924–1938. https://est ce que je.org/10
.1016/j.neuroimage.2012.06.005, PubMed: 22705374
Forgeron, S. M., Jenkinson, M., Woolrich, M.. W., Beckmann, C. F.,
Behrens, T. E. J., Johansen-Berg, H., Bannister, P.. R., De Luca,
M., Drobnjak, JE., Flitney, D. E., Niazy, R.. K., Saunders, J., Vickers,
J., Zhang, Y., De Stefano, N., Brady, J.. M., & Matthieu, P.. M..
(2004). Advances in functional and structural MR image analysis
and implementation as FSL. NeuroImage, 23(Supplément 1), S208–S219.
https://doi.org/10.1016/j.neuroimage.2004.07.051, PubMed:
15501092
De la Sporn, Ô. (2013). Network attributes for segregation and integra-
tion in the human brain. Opinion actuelle en neurobiologie, 23(2),
162–171. https://doi.org/10.1016/j.conb.2012.11.015, PubMed:
23294553
Thomsen, K., Piilgaard, H., Gjedde, UN., Bonvento, G., & Lauritzen,
M.. (2009). Principal cell spiking, postsynaptic excitation, et
oxygen consumption in the rat cerebellar cortex. Journal de
Neurophysiologie, 102(3), 1503–1512. https://est ce que je.org/10.1152/jn
.00289.2009, PubMed: 19571198
Tournier, J.-D., Calamante, F., & Connelly, UN. (2007). Robust deter-
mination of the fibre orientation distribution in diffusion MRI:
Non-negativity constrained super-resolved spherical deconvolu-
tion. NeuroImage, 35(4), 1459–1472. https://est ce que je.org/10.1016/j
.neuroimage.2007.02.016, PubMed: 17379540
Tournier, J.-D., Calamante, F., & Connelly, UN. (2012). MRtrix: Diffu-
sion tractography in crossing fiber regions. International Journal
of Imaging Systems and Technology, 22(1), 53–66. https://doi.org
/10.1002/ima.22005
Tournier, J.-D., Calamante, F., Gadian, D. G., & Connelly, UN.
(2004). Direct estimation of the fiber orientation density function
from diffusion-weighted MRI data using spherical deconvolution.
NeuroImage, 23(3), 1176–1185. https://est ce que je.org/10.1016/j
.neuroimage.2004.07.037, PubMed: 15528117
Tournier, J.-D., Forgeron, R., Raffelt, D., Tabbara, R., Dhollander, T.,
Pietsch, M., Christiaens, D., Jeurissen, B., Yeh, C.-H., & Connelly,
UN. (2019). MRtrix3: A fast, flexible and open software framework
for medical image processing and visualisation. NeuroImage,
202, 116137. https://doi.org/10.1016/j.neuroimage.2019
.116137, PubMed: 31473352
Wainstein, G., Rojas-Líbano, D., Medel, V., Alnæs, D., Kolskår,
K. K., Endestad, T., Laeng, B., Ossandon, T., Crossley, N., Matar,
E., & Briller, J.. M.. (2021). The ascending arousal system promotes
optimal performance through mesoscale network integration in a
visuospatial attentional task. Neurosciences en réseau, 5(4), 890–910.
https://doi.org/10.1162/netn_a_00205, PubMed: 35024535
Whelan, R.. R.. (2007). Neuroimaging of cognitive load in instruc-
tional multimedia. Educational Research Review, 2(1), 1–12.
https://doi.org/10.1016/j.edurev.2006.11.001
Wilson, C. J.. (2013). Active decorrelation in the basal ganglia. Neu-
roscience, 250, 467–482. https://doi.org/10.1016/j.neuroscience
.2013.07.032, PubMed: 23892007
Wu, T., Liu, J., Hallett, M., Zheng, Z., & Chan, P.. (2013). Cerebel-
lum and integration of neural networks in dual-task processing.
NeuroImage, 65, 466–475. https://doi.org/10.1016/j.neuroimage
.2012.10.004, PubMed: 23063842
Neurosciences en réseau
863
l
D
o
w
n
o
À
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
J’ai
r
e
c
t
.
m
J’ai
t
.
/
t
/
e
d
u
n
e
n
À
r
t
J’ai
c
e
–
p
d
l
F
/
/
/
/
/
7
2
8
4
4
2
1
1
8
5
2
7
n
e
n
_
À
_
0
0
3
0
8
p
d
t
.
F
b
oui
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3