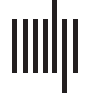RECHERCHE
Modulation of limbic resting-state networks by
subthalamic nucleus deep brain stimulation
John Eraifej1, Joana Cabral2,3,4, Henrique M. Fernandes4, Joshua Kahan5, Shenghong He6,
Laura Mancini7,8, John Thornton7,8, Mark White7,8, Tarek Yousry7,8, Ludvic Zrinzo5,
Harith Akram5, Patricia Limousin5, Tom Foltynie5, Tipu Z. Aziz1, Gustavo Deco9,10,11,
Morten Kringelbach3,4,12, and Alexander L. Green1
1Oxford Functional Neurosurgery Group, Nuffield Department of Surgical Sciences, University of Oxford,
Oxford, United Kingdom
2Life and Health Sciences Research Institute (ICVS), École de médecine, University of Minho, Braga, Portugal
3Centre for Eudaimonia and Human Flourishing, Linacre College, University of Oxford, Oxford, United Kingdom
4Center for Music in the Brain, Department of Clinical Medicine, Aarhus University, Aarhus, Denmark
5Sobell Department for Motor Neurosciences and Movement Disorders, UCL Institute of Neurology,
Londres, United Kingdom
6MRC Brain Network Dynamics Unit, Nuffield Department of Clinical Neurosciences,
University of Oxford, Oxford, United Kingdom
7Neuroradiological Academic Unit, Department of Brain Repair and Rehabilitation, UCL Institute of Neurology,
University College London, Londres, United Kingdom
8Lysholm Department of Neuroradiology, National Hospital for Neurology and Neurosurgery,
UCLH NHS Foundation Trust, Londres, United Kingdom
9Center for Brain and Cognition, Computational Neuroscience Group, Universitat Pompeu Fabra, Barcelona, Espagne
10Institució Catalana de la Recerca i Estudis Avançats, Barcelona, Espagne
11Department of Neuropsychology, Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Allemagne
12Département de psychiatrie, University of Oxford, Oxford, United Kingdom
Mots clés: Deep brain stimulation, Subthalamic nucleus, Limbic network, Functional MRI, Leading
Eigenvector Dynamics Analysis, Parkinson’s disease
ABSTRAIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Beyond the established effects of subthalamic nucleus deep brain stimulation (STN-DBS) dans
reducing motor symptoms in Parkinson’s disease, recent evidence has highlighted the effect on
non-motor symptoms. Cependant, the impact of STN-DBS on disseminated networks remains
unclear. This study aimed to perform a quantitative evaluation of network-specific modulation
induced by STN-DBS using Leading Eigenvector Dynamics Analysis (LEiDA). We calculated
the occupancy of resting-state networks (RSNs) in functional MRI data from 10 patients
with Parkinson’s disease implanted with STN-DBS and statistically compared between
ON and OFF conditions. STN-DBS was found to specifically modulate the occupancy of
networks overlapping with limbic RSNs. STN-DBS significantly increased the occupancy
of an orbitofrontal limbic subsystem with respect to both DBS OFF (p = 0.0057) et
49 age-matched healthy controls (p = 0.0033). Occupancy of a diffuse limbic RSN was
increased with STN-DBS OFF when compared with healthy controls (p = 0.021), but not when
STN-DBS was ON, which indicates rebalancing of this network. These results highlight the
modulatory effect of STN-DBS on components of the limbic system, particularly within the
orbitofrontal cortex, a structure associated with reward processing. These results reinforce
the value of quantitative biomarkers of RSN activity in evaluating the disseminated impact
of brain stimulation techniques and the personalization of therapeutic strategies.
un accès ouvert
journal
Citation: Eraifej, J., Cabral, J.,
Fernandes, H. M., Kahan, J., Il, S.,
Mancini, L., Thornton, J., Blanc, M.,
Yousry, T., Zrinzo, L., Akram, H.,
Limousin, P., Foltynie, T., Aziz, T. Z.,
Deco, G., Kringelbach, M., & Vert, UN. L.
(2023). Modulation of limbic resting-
state networks by subthalamic nucleus
deep brain stimulation. Réseau
Neurosciences, 7(2), 478–495. https://
doi.org/10.1162/netn_a_00297
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00297
Informations complémentaires:
https://doi.org/10.1162/netn_a_00297
Reçu: 8 Novembre 2021
Accepté: 29 Novembre 2022
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
John Eraifej
john.eraifej@nds.ox.ac.uk
Éditeur de manipulation:
Alex Fornito
droits d'auteur: © 2023
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
Modulation of limbic resting-state networks by STN-DBS
Deep brain stimulation (DBS):
Stimulation of deep brain nuclei
using chronically implanted
electrode and pulse generator.
Parkinson’s disease (PD):
Neurodegenerative disorder caused
by depletion of nigrostriatal
dopamine characterized by slowness
of movement, tremor, and stiffness in
addition to neuropsychiatric
symptoms.
RÉSUMÉ DE L'AUTEUR
This article addresses a burning question regarding stimulation strategies to rebalance brain
network dynamics. Using a rare fMRI dataset of Parkinson’s disease patients implanted with
deep brain stimulation, we report evidence of network-specific modulatory effects in the
dynamics of resting-state networks. En résumé, we found that Leading Eigenvector Dynamics
Analysis (LEiDA) successfully identified all seven reference resting-state networks in
participants with subthalamic deep brain stimulation (STN-DBS). En particulier, STN-DBS
increases resting-state orbitofrontal cortex activity. STN-DBS also normalizes wider resting-
state limbic network activity, and this correlated with depressive symptoms in pre- et
post-operative assessments. The work is limited by a low number of participants, et le
retrospective nature of this work, but provides evidence that STN-DBS modulates limbic
network occupancy in real time.
INTRODUCTION
Deep Brain Stimulation Motor and Non-motor Effects
Deep brain stimulation (DBS) has become an established surgical option for treating the motor
symptoms of Parkinson’s disease (PD) (Deuschl et al., 2006; Vitek et al., 2020). The procedure
involves implantation of electrodes into deep brain nuclei under stereotactic guidance and is con-
sidered for patients when pharmacological management is no longer sufficient or is associated
with intolerable side effects (Krack et al., 2003). Aujourd'hui, the two most common targets are the
globus pallidus interna (GPi) and the subthalamic nucleus (STN), the latter of which is preferred
in most cases because of the favorable reduction in dopamine replacement therapy despite the
higher risk of non-motor complications (Rughani et al., 2018; Volkmann et al., 2010).
The mechanism of action of STN-DBS remains uncertain. Initially, a functional inhibitory
effect was proposed, but more recent evidence suggests that modulation of a disseminated
cortico-striato-thalamo-cortical network contributes to the observed clinical improvement in
motor symptoms (Horn et al., 2017; Corne, Wenzel, et coll., 2019; Kahan et al., 2014; Saenger
et coll., 2017). Electrophysiological studies have demonstrated that improvement in motor
symptoms correlates with modulation of beta-range phase-amplitude coupling within the
somatomotor network (De Hemptinne et al., 2013; De Hemptinne et al., 2015; Eusebio
et coll., 2011). In vivo animal studies also corroborate this network theory of STN-DBS for the
motor symptoms of PD (Gradinaru et al., 2009; Hashimoto et al., 2003; Xu et al., 2008).
STN-DBS also demonstrates improvement in the non-motor symptoms of PD, and this is asso-
ciated with improved quality of life scores (Abbes et al., 2018; Dafsari et al., 2016; Dafsari,
Blanc, et coll., 2018). These effects correlate with active electrode location and structural
connectivity patterns (Dafsari, Petry-Schmelzer, et coll., 2018; Mosley, Paliwal, et coll., 2020;
Mosley, Robinson, et coll., 2020; Petry-Schmelzer et al., 2019). Par exemple, improvement in
apathy has been observed with ventral STN stimulation while impulsivity is associated with
ventromedial STN and orbitofrontal cortex (OFC) connectivité (Mosley, Paliwal, et coll., 2020;
Petry-Schmelzer et al., 2019). Taken together, these studies identify a key role for disseminated
brain network modulation in the therapeutic effects seen in STN-DBS (Accolla & Pollo, 2019).
Functional Neuroimaging in the Context of Deep Brain Stimulation
Brain network dynamics can be understood as the evolution of interactions between brain
areas over time (Calhoun et al., 2014; Tononi & Edelman, 1998). In recent decades, functional
Neurosciences en réseau
479
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
Resting-state networks (RSN):
Distinct modes of long-distance
interactions occurring consistently
across brain regions and between
participants at rest.
Occupancy:
Occupancy of RSNs is the probability
of occurrence of a particular RSN
across the entire scan period.
Leading Eigenvector Dynamics
Analysis (LEiDA):
Analytic framework that captures
resting-state networks as recurrent
modes of phase-locked
synchronization of fMRI signals.
magnetic resonance imaging (IRMf) studies have revealed distinct modes of long-distance
interactions occurring consistently across participants at rest, the so-called resting-state net-
travaux (RSNs; Damoiseaux et al., 2006; De Luca et al., 2006; Yeo et al., 2011). Although
the precise origin and function of RSNs remains under debate, their spatial patterns reveal
functionally relevant brain subsystems whose integrity appears disrupted in a wide range of
neurological and psychiatric disorders (Chen et al., 2020; Fornito et al., 2015; Stam, 2014;
Williams, 2016). En tant que tel, analyzing how STN-DBS affects brain activity at the level of RSNs
may identify possible mechanisms of action and permit novel therapeutic strategies targeted at
specific network patterns (Kringelbach et al., 2011).
Previous fMRI network analysis has shown that STN-DBS modulates all the major compo-
nents of the motor cortico-striato-thalamo-cortical loop with normalization of widespread
somatomotor resting-state networks (Corne, Wenzel, et coll., 2019; Kahan et al., 2014;
Tahmasian et al., 2015). Cependant, these studies focused on the analysis of correlations
between brain areas evaluated over the whole recording time and did not aim for a quantita-
tive comparison of temporally resolved RSN activity between ON and OFF conditions.
Dynamic Resting-State Network Analysis
Since RSNs represent patterns of connectivity that form transiently and recurrently over time, un
recent methodology has been developed to quantify occupancy of RSNs (Lord et al., 2019). Dans
this analytic framework, termed Leading Eigenvector Dynamics Analysis (LEiDA), RSNs are
captured as recurrent modes of phase-locked synchronization of fMRI signals, which were found
to overlap closely with RSNs from the literature (Lord et al., 2019; Vohryzek et al., 2020). Un
advantage of the method is that it allows calculation of the proportion of time points during
an fMRI session assigned to a given RSN, providing a quantitative measure that can be statisti-
cally compared between conditions (Alonso Martínez et al., 2020; Cabral, Vidaurre, et coll., 2017;
Figueroa et al., 2019; Larabi et al., 2020; Magalhães et al., 2021; Wong et al., 2021). Ce
approach allows detection of network-specific modulations; Par exemple, a previous study using
LEiDA revealed that psilocybin, a psychoactive molecule, selectively decreases the occupancy
of the frontoparietal RSN (associated with executive control), leaving the occupancy of the other
RSNs unchanged (Lord et al., 2019), while another study revealed the specific engagement of
the orbitofrontal cortex reward system during music listening (Fasano et al., 2023).
Dans cette étude, we aimed to investigate how RSN occupancy changes with STN-DBS turned
ON and OFF. To do so, we applied LEiDA to an fMRI dataset from patients with PD implanted
with STN-DBS and compared this with healthy age-matched controls (Saenger et al., 2017).
Understanding how STN-DBS modulates brain activity at the level of RSNs can be crucial to
advance in the design of more efficient and personalized therapeutic strategies targeting the
non-motor effects of DBS (Kringelbach et al., 2011).
MATERIALS AND METHODS
Scanning of all participants was performed in accordance with the Declaration of Helsinki
(59th amendment) and approved by the relevant local ethics committees (see the
Informations complémentaires). All the fMRI data from both PD patients and controls used in this
study were previously published (Saenger et al., 2017).
PD Patients
Ten patients (Tableau 1) who met the UK Brain Bank criteria for idiopathic Parkinson’s disease
and had received bilateral STN-DBS for more than 6 months were recruited. All operations
Neurosciences en réseau
480
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
Tableau 1.
Parkinson’s disease patient characteristics
UPDRS-III
Right electrode
Patient
1
Age
65
Sex
F
Dom.
main
R.
Months since
surgery
20
Off/OFF Off/ON Volts
0.5
21
53
Pulse
width (μs)
60
Freq.
(Hz)
180
2
3
4
5
6
7
8
9
10
54
65
50
64
54
43
61
56
45
F
M.
F
F
M.
M.
M.
M.
M.
R.
R.
L
R.
R.
L
R.
R.
R.
9
67
102
29
19
48
8
28
48
33
60
51
46
45
51
46
44
53
10
20
17
19
26
23
25
42
44
Mean
55.7
SD
8.1
37.8
29.3
48.2
7.2
24.7
10.6
2.4
3.7
3.8
2.5
2.4
5.4
3.2
3.7
2.4
3.0
1.3
60
60
60
60
60
60
60
60
60
130
130
185
130
130
80
130
130
130
60.0
0.0
135.5
29.3
Left electrode
Pulse
width (μs)
90
Freq.
(Hz)
180
60
90
60
60
60
60
60
60
60
130
130
185
130
130
80
130
130
130
66.0
12.6
135.5
29.3
Volts
3.30
2.40
3.45
3.60
2.50
2.30
4.10
2.90
4.10
3.15
3.20
0.70
Unified Parkinson’s Disease Rating
Scale part III (UPDRS-III):
Clinical rating scale of disease
severity in Parkinson’s disease. Part
III measures motor symptom severity.
were performed at the National Hospital for Neurology and Neurosurgery (NHNN), Queen
Square, Londres (using Model 3389, Medtronic). Stimulation parameters were set to produce
optimal clinical responses. Medication was withdrawn overnight (10–12 hr) before scanning.
Inclusion was limited to patients who tolerated lying flat with minimal head tremor while
being both OFF medication and with DBS OFF. Pour chaque patient, before scanning, both ON
and OFF stimulation, Unified Parkinson’s Disease Rating Scale part III (UPDRS-III) scores were
recorded.
Healthy Controls
Forty-nine healthy age-matched controls (30 males; mean age 57.95, standard deviation 4.05)
were selected from a larger cohort recruited in the University of Minho, Portugal (Cabral,
Vidaurre, et coll., 2017; Saenger et al., 2017). Participants with psychiatric or neurological dis-
ordres (or a history thereof ) were excluded from participation in the study.
Data Acquisition
The scanning of patients was performed at NHNN, using a safe previously published protocol
(Boertien et al., 2011; Carmichael et al., 2007; Kahan et al., 2015). Briefly, scanning was
performed in a Siemens Avanto 1.5 T MRI scanner using a transmit-receive (Tx/Rx) head coil.
The specific absorption ratio in the head was limited to 50.1 W/kg. Subjects received two
resting-state functional MRI scans during resting state with eyes closed (repetition time [TR] =
2,420 ms; echo time [TE] = 40 ms; flip angle [FA] = 90°; field of view [FoV] = 192 × 192 mm2;
matrix size = 64 × 64; 32 tranches axiales 3.5 mm thick, gap between slices of 0.7 mm; spatial
resolution = 3 × 3 × 4.2 mm3; 200 scans) within safe parameters (Carmichael et al., 2007).
The order of data collection (c'est à dire., ON stimulation then OFF stimulation, and vice versa) était
randomly assigned, such that half the patients were scanned ON then OFF, and half were
Neurosciences en réseau
481
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Lead location for included participants. (UN) Coronal view. (B) Axial view. (C) Sagittal
Chiffre 1.
voir. (D) Oblique view with STN subdivisions identified in lead-DBS: orange = motor, blue = asso-
ciative, yellow = limbic.
scanned OFF then ON. Stimulation was switched off approximately 15 min before the collec-
tion of OFF condition resting-state data. DBS lead localization was visualized using Lead-DBS
(Corne, Li, et coll., 2019; Corne & Kühn, 2015). Chiffre 1 demonstrates the electrode position for
9 out of 10 participants, as the original T1 volumetric scan for one participant could not
be obtained.
The scanning of age-matched healthy controls was conducted separately at Hospital de
Braga (Portugal). De la même manière, resting-state data were collected with eyes closed using whole-
brain EPI using a clinically approved 1.5T Siemens Magnetom Avanto (Siemens Medical
Solutions, Erlangen, Allemagne; parameters: 30 tranches axiales, TR/TE = 2,000/30 ms, FA = 90°,
slice thickness = 3.5 mm, slice gap = 0.48 mm, voxel size = 3.5 × 3.5 mm2, FoV =
1,344 mm and 180 volumes).
Data Analysis
fMRI preprocessing. Resting fMRI standard preprocessing was performed with FMRIB Software
Library tools (FSL v5.07; https://fsl.fmrib.ox.ac.uk/fsl/). Voxel-level fMRI signals were reduced
à 92 non-cerebellar brain areas (cortical areas and bilateral STN), by averaging the signals
across all voxels belonging to each brain area defined according to the Automated Anatomical
Labelling (AAL) atlas (Tzourio-Mazoyer et al., 2002). Analysis was restricted to non-cerebellar
zones du cerveau, in agreement with previous studies using LEiDA, with the intention of mapping
RSNs onto those identified by Yeo et al.
Neurosciences en réseau
482
Modulation of limbic resting-state networks by STN-DBS
Detection of RSNs. LEiDA was then used to capture recurrent phase-locking (PL) patterns in the
fMRI signal from implanted PD patients. Briefly, the phase of the fMRI signal in each brain area
was obtained via the Hilbert transform. Subsequently, the N × N phase-locking matrix was
calculated at each time point as cos(je(n, t ) − θ(p, t)), and the corresponding leading eigenvector
(a vector of size 1 × N ) was extracted. The leading eigenvectors calculated for all time points,
representing the PL patterns observed across scans, were partitioned into a repertoire of K
clusters using k-means clustering. Since the precise number of RSNs remains unclear, we varied
K between 5 et 20 and analyzed the results across the range of partitions explored (10,000
replicates of the k-means were run to ensure stability of the results; Cabral, Vidaurre, et coll.,
2017; Figueroa et al., 2019; Lord et al., 2019; Vohryzek et al., 2020). After detecting the num-
ber of RSNs that better distinguished between conditions, clustering was repeated for the
selected K using 100,000 replicates. The overlap with seven reference networks of intrinsic
functional connectivity (Yeo et al., 2011) was calculated for the cluster centroids representa-
tive of each PL state, and the same color code from the original paper was used to render the
phase-shifted subsystems revealed in each PL state, following the methodology from Vohryzek
et autres. (2020). Given the significant overlap between the cluster centroids with canonical RSNs,
in the following we refer to the PL states by the name of the RSN with most significant overlap.
Occupancy of RSNs. The occupancy of each RSN was calculated for each fMRI scan, as the
proportion of time points in a scan assigned to a given cluster by the k-means algorithm
(Cabral, Kringelbach, et coll., 2017; Cabral, Vidaurre, et coll., 2017).
Statistical comparisons. Detection of the RSNs that most significantly changed in occupancy
between ON and OFF conditions of STN-DBS was conducted using a permutation-based
paired sample t test with 100,000 permutations to ensure stability of the results (Figueroa
et coll., 2019; Lord et al., 2019). To evaluate the significance of results taking into account
the probability of false positives arising from multiple comparisons, the p values are reported
with respect to both the standard threshold α = 0.05 (red dashed line in Figure 2C), and a
corrected threshold αCorrected = 0.05/K (green dashed line in Figure 2C), taking into account
the number of independent hypothesis tested in each partition model (Figueroa et al., 2019).
Correlations between UPDRS sub-scores and occupancy of RSNs were identified by calcu-
lating Pearson’s correlation coefficients of change scores. Analyses were run in MATLAB 2020
using LEiDA scripts.
Validation using the control dataset. After selecting the number of RSNs (K ) that maximized the
difference between DBS ON and OFF (c'est à dire., returning the lowest p value), the occupancy of the
same RSNs was calculated for the control subjects (Lord et al., 2019). To do so, after obtaining
the leading eigenvectors from all control fMRI scans, a single iteration of the k-means algo-
rithm was run, inputting the K cluster centroids (for the selected K, here K = 15) detected from
the patient dataset as “start vectors.” This validation strategy, introduced in Lord et al. (2019),
allows verification if the same RSNs detected in the patient dataset are also detected with sim-
ilar probabilities in a dataset from a different research center (Lord et al., 2019). A permutation-
based unpaired sample t test was used to compare the RSN probabilities from controls with the
probabilities from patients, again with 100,000 permutations to ensure stability of the results.
RÉSULTATS
Clinical Outcome Measures
Patient characteristics can be seen in Table 1. Patient UPDRS was significantly lower (p =
0.00012) with DBS ON (mean = 24.7, SD = 10.6) compared with DBS OFF (mean = 48.2,
Neurosciences en réseau
483
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
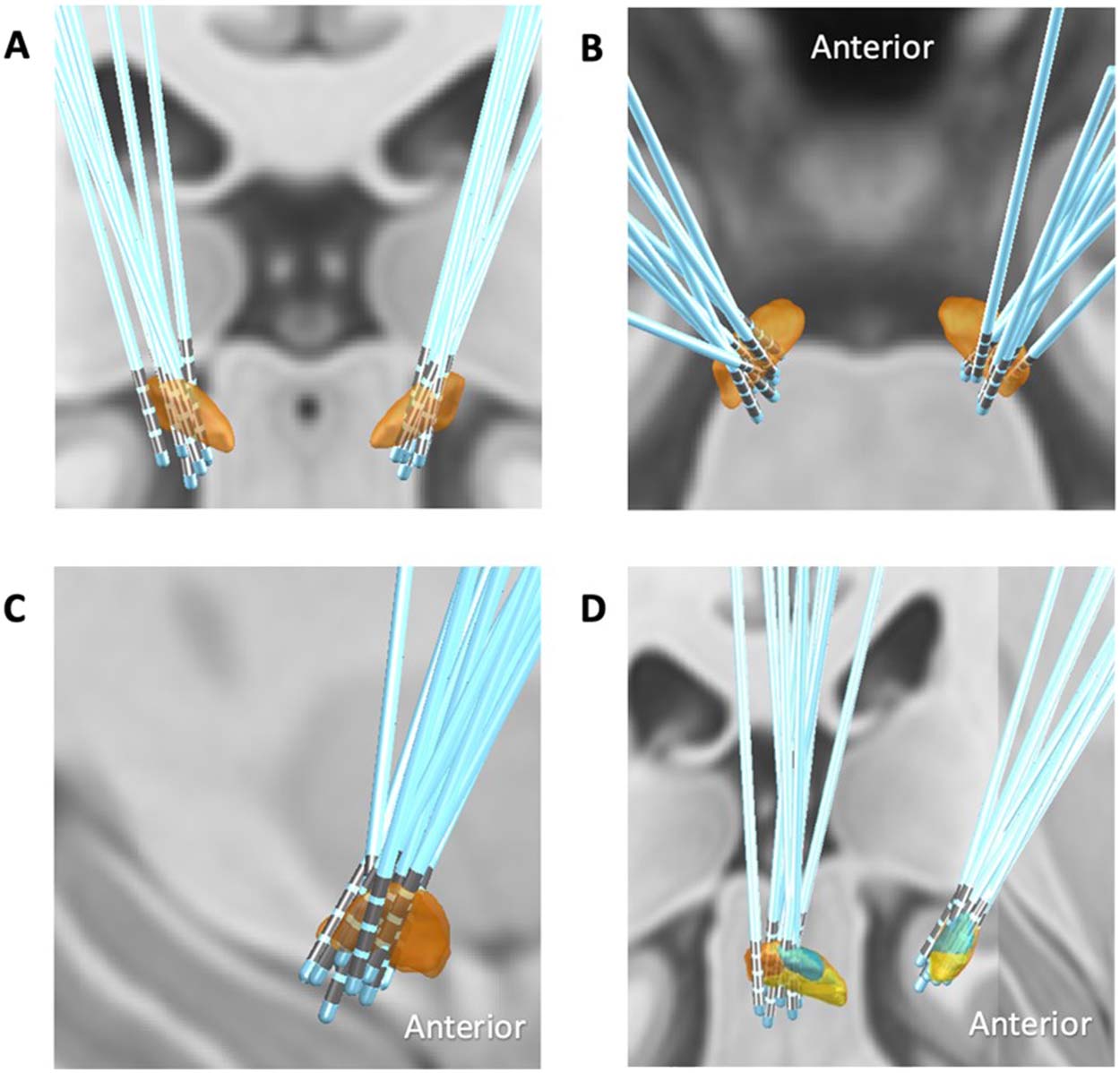
Chiffre 2. DBS modulates the occurrence of specific RSNs. (UN) Representation of the RSNs detected with LEiDA for each partition into K
clusters, with K ranging between 5 et 20. RSNs whose occupancy changes significantly when DBS is turned ON are highlighted by reporting
the corresponding p value in the title (see panel C). RSNs are represented in cortical space, rendering only the brain regions whose fMRI signal
is shifted in phase. RSNs are colored according to overlap with reference RSNs defined in Yeo et al. (2011) (shown in B, black if no significant
overlap detected with p > 0.05/K ). (B) Reference RSNs estimated from 1,000 subjects from correlation-based intrinsic functional connectivity
(Yeo et al., 2011). (C) For each RSN detected in each partition (c'est à dire., for each K ), the probabilities are compared between STN-DBS ON and
OFF, and the corresponding p values are reported with respect to the standard statistical threshold (α = 0.05, red dashed line) and the threshold
corrected by the number of independent hypotheses tested (αCorrected = 0.05/K, green dashed line). Although most RSNs do not differ in occu-
pancy between conditions (p > α, black asterisks), a number of patterns exhibit significant differences between conditions (red and green
asterisks). Notably, these changes occur exclusively in limbic and somatomotor RSNs.
Unified Parkinson’s Disease Rating
Scale part I (UPDRS-I):
Clinical rating scale of disease
severity in Parkinson’s disease. Part I
measures non-motor symptom
severity.
SD = 7.2) while off medication, with all participants demonstrating improvement. UPDRS-I
scores were collated retrospectively from pre-operative and post-operative assessments. After
STN-DBS, one participant had increased intellectual impairment, one participant had
increased thought disorder, three participants had reduced motivation/initiative, and all partic-
ipants either improved or had no change in their depression scores (see the Supporting
Information).
Neurosciences en réseau
484
Modulation of limbic resting-state networks by STN-DBS
Tableau 2. Non-motor scores. Scoring: Intellectual impairment: 0 = None. 1 = Mild. Consistent forgetfulness with partial recollection of events
and no other difficulties. 2 = Moderate memory loss, with disorientation and moderate difficulty handling complex problems. Mild but definite
impairment of function at home with need of occasional prompting. 3 = Severe memory loss with disorientation for time and often to place.
Severe impairment in handling problems. 4 = Severe memory loss with orientation preserved to person only. Unable to make judgements or
solve problems. Requires much help with personal care. Cannot be left alone at all. Thought disorder: 0 = None. 1 = Vivid dreaming. 2 =
“Benign” hallucinations with insight retained. 3 = Occasional to frequent hallucinations or delusions; without insight; could interfere with daily
activités. 4 = Persistent hallucinations, delusions, or florid psychosis. Not able to care for self. Depression: 0 = None. 1 = Periods of sadness or
guilt greater than normal, never sustained for days or weeks. 2 = Sustained depression (1 week or more). 3 = Sustained depression with
vegetative symptoms (insomnia, anorexia, weight loss, loss of interest). 4 = Sustained depression with vegetative symptoms and suicidal
thoughts or intent. Motivation/initiative: 0 = Normal. 1 = Less assertive than usual; more passive. 2 = Loss of initiative or disinterest in elective
(nonroutine) activités. 3 = Loss of initiative or disinterest in day-to-day (routine) activités. 4 = Withdrawn, complete loss of motivation.
Note that mean and standard deviation are presented, although this is a semi-quantitative scoring system.
Patient
1
Age
65
Intellectual impairment
Post-op
Pre-op
0
0
Thought disorder
Depression
Pre-op
0
Post-op
0
Pre-op
2
Post-op
0
Motivation/initiative
Post-op
Pre-op
1
1
UPDRS-I
2
3
4
5
6
7
8
9
10
Mean
SD
54
65
50
64
54
43
61
56
45
1
1
1
0
2
1
1
1
0
0
1
1
1
2
0
1
1
0
1
1
0
2
0
0
0
1
0
0
1
0
1
1
0
0
1
0
1
0
1
0
1
1
0
0
0
0
0
0
0
1
1
0
0
0
1
0
3
0
3
1
0
0
0
0
2
1
0
2
2
1
0
0
55.7
8.1
0.8
0.6
0.7
0.7
0.5
0.7
0.4
0.5
0.6
0.7
0.2
0.4
0.9
1.2
0.9
0.9
Non-motor scores are measured as part of the UPDRS part I (UPDRS-I). These are graded
depuis 1 à 4 and measure severity of symptoms based on patient recall. Retrospective data
collection was completed to collate the data obtained during routine clinical assessments
from the pre-operative clinical review and the post-operative review at 6 à 12 months after
surgery (Tableau 2).
Effects of DBS on RSN Occupancy
The network patterns detected with LEiDA in the patient fMRI dataset are reported in Figure 2A
for the whole range of partitions explored (c'est à dire., given the undefined number of RSNs, motifs
where clustered into K = 5, 6, …, 20 clusters) and sorted according to occupancy. Chaque
network pattern is represented by the corresponding cluster centroid, coloring the subset of
brain areas exhibiting phase synchronization, while shifted in phase from the rest of the brain.
Consistently, across all clustering solutions, the most dominant pattern is a global state in
which the fMRI signals of all brain areas are aligned in phase, not revealing the segregation
of any particular subsystem (hence represented as a transparent brain), occupying the first
column of Figure 2 as the state with the highest occupancy across scans.
Neurosciences en réseau
485
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
The remaining network patterns detected exhibit the segregation of a subset of brain areas,
within which the fMRI signals appear aligned in phase and shifted from the rest of the brain. Chaque
of these network patterns was compared with the seven reference RSNs defined by Yeo et al.
(2011) shown in Figure 2B, and the same color code was used when the overlap was statistically
significant (with p > 0.05/K ). Despite the distinct methodologies, most of the cluster centroids
obtained demonstrate a statistically significant overlap to reference RSNs, except for the networks
colored in black, which involve subcortical structures not considered in the Yeo et al. (2011) étude.
To evaluate the effects of STN-DBS on the modulation of RSNs, the occupancy of each RSN
shown in Figure 2A was calculated for all patient fMRI sessions and compared between ON
and OFF conditions. The p values of the statistical comparison are reported in panel C with
respect to the standard statistical threshold (α = 0.05, red dashed line) and the threshold
corrected for multiple comparisons (αCorrected = 0.05/K, green dashed line). Although most
RSNs do not differ in occupancy between conditions (p > α, black asterisks), a few patterns
were found to exhibit significant differences between conditions (red and green asterisks).
Observing the corresponding RSNs in panel A (for which the p value is reported in the title),
these changes occur exclusively in the occupancy of the global state (for K = 5, 6, et 8), le
somatomotor RSN (K = 8, 10, et 11) and the limbic RSN (for all K between 10 et 20, sur-
viving correction for the number of independent hypotheses tested only with K = 15 et 18).
RSN Occupancy in Patients and Controls
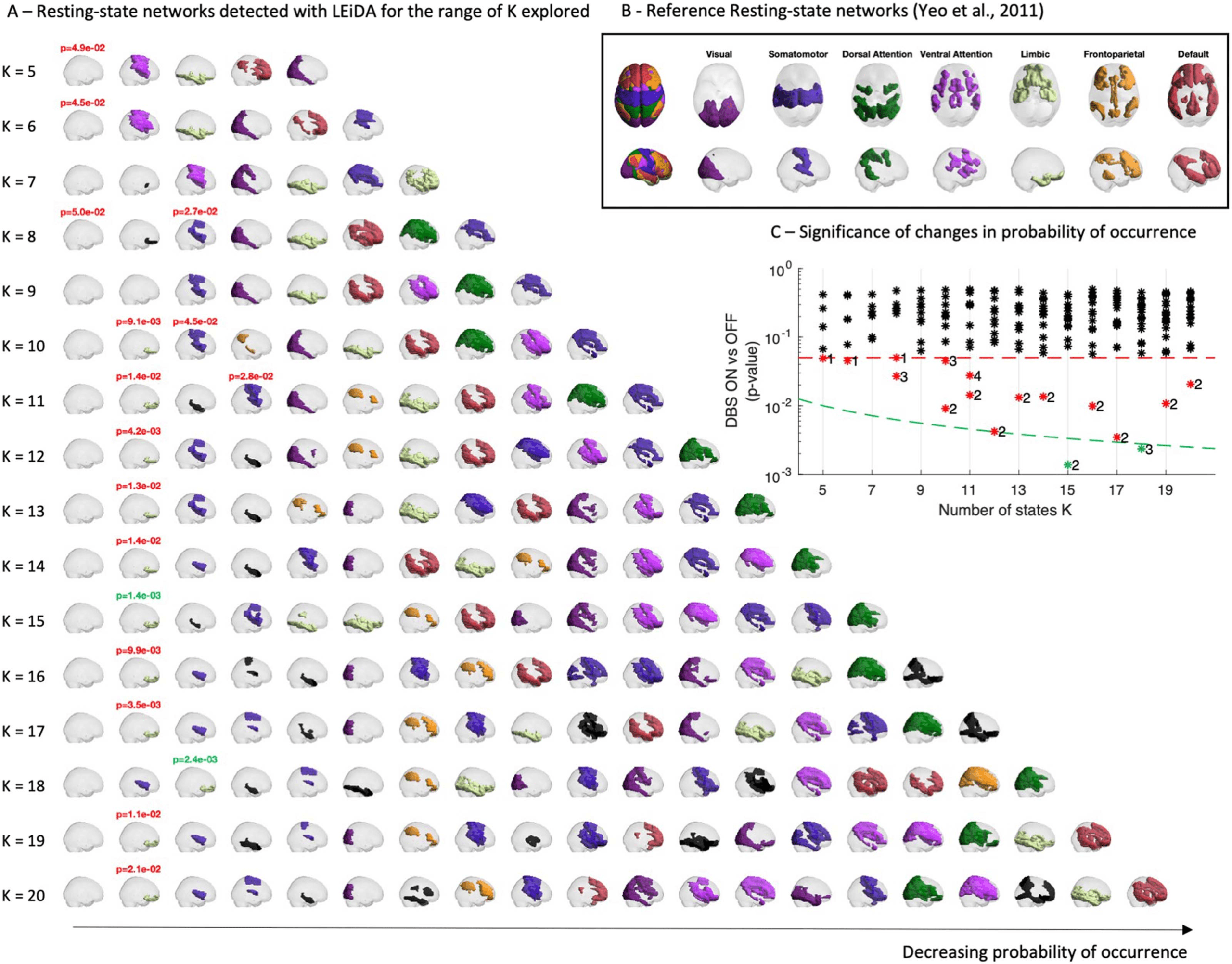
For the subsequent analysis, the partition into 15 network patterns was selected for revealing the
RSN that most significantly differed between the DBS OFF and ON conditions. In Figure 3, the full
repertoire of network patterns detected with K = 15 is reported, together with the corresponding
probabilities in patients (in both OFF and ON conditions) and in healthy controls. Notably,
despite the low number of scans and the artefacts due to the electrode lead and motion, all seven
RSNs used as reference (shown in Figure 2B) are represented in this repertoire, with some being
represented by more than one network pattern. Par exemple, motifs 3, 4, 9, 12, et 13 overlap
significantly with the somatomotor network, although each reveals the engagement of different
substructures. De plus, one RSN involving only subcortical structures was detected (5).
Most RSNs detected did not change in occupancy across scans, neither between DBS OFF
versus ON in patients, nor between patients and healthy age-matched controls (all p values >
Chiffre 3. Le 15 RSNs identified in the patients’ fMRI data. (UN) The repertoire of BOLD PL states obtained for K = 15, represented by coloring
only the brain areas whose BOLD signal phase is shifted with respect to the dominant BOLD phase orientation (see Figure 2A). The patches are
colored according to the reference functional network with most significant overlap, and black otherwise. (B) Probabilities of occurrence
(mean ± standard error of the mean) of the different states during the fMRI recordings in the two DBS conditions and in healthy controls.
Neurosciences en réseau
486
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
0.1 for patterns 3, 4, 5, 6, 7, 9, 10, 13, 14, et 15). The fact that the prevalence of most RSNs
(detected from the patient dataset only) remains stable in the control dataset (from a different
étude, recorded at a different institution) reinforces the validity of the method.
Regarding the network patterns that did exhibit differences in occupancy between condi-
tion, these involve the globally synchronized pattern (1), two RSNs overlapping with the
limbic system (one involving only structures in the orbitofrontal cortex [2, OFC] and another
involving more diffuse limbic structures [8]), and finally one overlapping with the somatomo-
tor system (12). Although STN-DBS only slightly increased the occurrence of the globally syn-
chronized mode (1), this tendency (0.05 > p > 0.1) was observed both with respect to DBS
OFF and with respect to healthy controls. This pattern, in which no particular RSN is detected,
may be due to diffuse effects of DBS stimulation on the whole brain. Since we are mostly
interested in network-specific modulation induced by STN-DBS, the effects on network pat-
terns 2, 8, et 12 are discussed in more detail below.
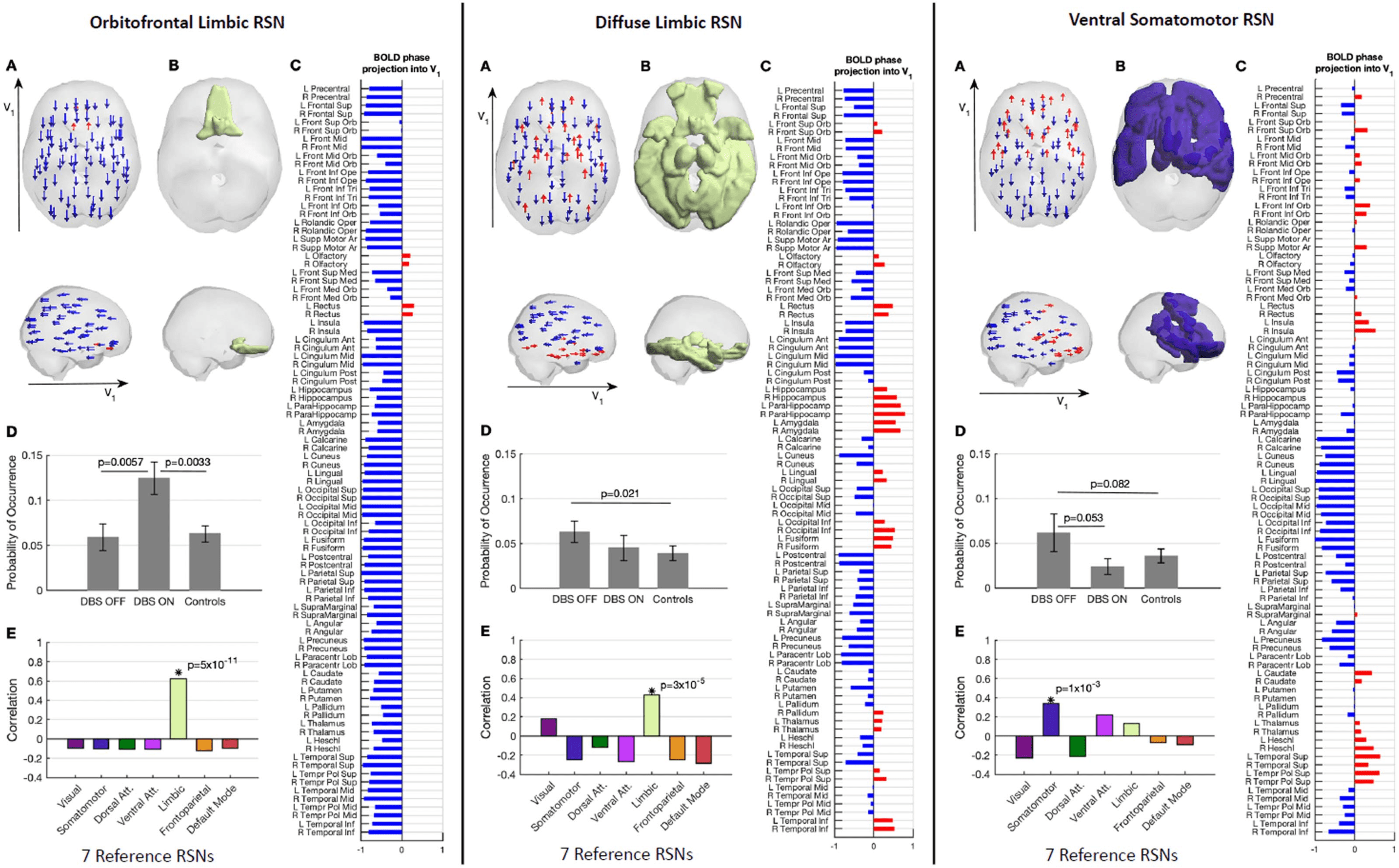
Effects of STN-DBS on the orbitofrontal RSN. The RSN that showed the most significant change
in occupancy during STN-DBS involves regions within the OFC, labeled in the AAL atlas as
the bilateral olfactory and rectus (voir la figure 4, left panel). In more detail, the fMRI signal in
these brain areas is shifted in phase with respect to the rest of the brain (see red and blue
arrows in panel A). With STN-DBS ON, this orbitofrontal RSN had a mean occupancy (± stan-
dard deviation) de 0.124 ± 0.0571 compared with 0.059 ± 0.047 (p = 0.0057) for STN-DBS
OFF and 0.063 ± 0.064 (p = 0.0033) in age-matched healthy controls (panel D). Notably, le
occupancy of this state when STN-DBS was OFF did not show statistically significant differ-
ences compared with age-matched healthy controls. This RSN was found to significantly
overlap with the network identified as the limbic network in Yeo et al. (2011) (panel E). In a
leave-one-out sensitivity analysis, this change in orbitofrontal RSN occupancy remains the
most significant change observed with STN-DBS (see the Supporting Information).
Effects of STN-DBS on a diffuse limbic RSN. Occupancy of another RSN mapping onto the limbic
network was found to normalize with STN-DBS (Chiffre 4, middle panel). This diffuse RSN
includes the same AAL regions found in state 2, together with subcortical structures such as
the bilateral hippocampus, parahippocampus, amygdala, thalamus, left pallidum and cortical
areas within the temporal and occipital lobes (the list of brain areas is reported in panel C).
With STN-DBS OFF, the mean occupancy for this RSN was 0.063 ± 0.038 compared with
0.045 ± 0.044 (p = 0.176) for STN-DBS ON and 0.039 ± 0.058 (p = 0.021) in age-matched
healthy controls. Although the occupancy did not change significantly between STN-DBS OFF
versus ON, the fact that the difference with respect to age-matched healthy controls is not
significant when DBS is ON indicates a normalization towards healthy values.
Effects of STN-DBS on a ventral somatomotor RSN. A network pattern involving brain areas of the
somatomotor network with components of the ventral attention and limbic networks, tel que
the insula, the caudate nucleus, and the thalamus, showed a tendency for normalization
towards healthy control values under STN-DBS (Chiffre 4, right panel). Although not surviving
the standard threshold of 0.05 for the selected partition into K = 15 clusters, the occupancy
was found to reduce from 0.063 ± 0.067 with STN-DBS OFF to 0.024 ± 0.028 (p = 0.053) avec
DBS turned ON as compared with 0.036 ± 0.054 (p = 0.082) in healthy controls. Plus loin, comme
can be seen in Figure 2, on the first analysis of the results across the range of K explored, quand
choosing K = 8, 10, et 11, a network overlapping with the somatomotor RSN was found to
differ between ON and OFF at higher significance levels, avec p = 0.027 for K = 8, indicating
that the effects of STN-DBS on the somatomotor system are not negligible.
Neurosciences en réseau
487
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
un
_
0
0
2
9
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 4. RSNs modulated by STN-DBS. The three RSNs exhibiting the most sensitivity to STN-DBS modulation, namely the orbitofrontal
limbic RSN (gauche), the diffuse limbic RSN (middle), and ventral somatomotor RSN (droite). (UN) In the LEiDA framework, the occurrence of an RSN
is characterized by a phase shift of the fMRI signals in a subset of brain areas with respect to the leading eigenvector V1, as illustrated by the
arrows placed at the center of gravity of each brain area, colored in blue when projecting in the negative direction of V1, or in red when
projecting in the opposite direction of V1. (B) The phase-shifted areas (represented in red in panel A) are colored according to the reference
network with significant overlap (see panel E). (C) List of AAL brain areas and the corresponding projection into V1. (D) Probabilities of occur-
rence (mean ± standard error of the mean) of each RSN during the fMRI recordings in the two DBS conditions and in healthy controls. (E)
Overlap with seven reference RSNs from Yeo et al. (2011).
Correlation with disease scores. When comparing motor and non-motor scores with resting-
state network occupancy, we found that change in UPDRS-III (moteur) scores did not correlate
with change in somatomotor RSN occupancy. UPDRS-I scores were obtained as a proxy mea-
sure of non-motor symptoms (Tableau 2). Change in depressive symptoms correlated with
change in diffuse limbic RSN occupancy (correlation coefficient = 0.698, p = 0.025), alors que
change in intellectual impairment correlated with change in somatomotor RSN occupancy
(correlation coefficient = 0.795, p = 0.006). There was no correlation between these RSNs
and symptoms of thought disorder or motivation and initiative.
DISCUSSION
Many questions remain open regarding the origin of RSNs and their role in brain function.
Cependant, irrespective of their number or physiological origin, there is growing evidence
reporting alterations in RSN integrity in different types of neuropsychiatric disorders, indicating
that RSNs are, at least in part, related to the formation of coordinated thought and behavior. Comme
Neurosciences en réseau
488
Modulation of limbic resting-state networks by STN-DBS
tel, understanding the external factors—be they pharmacological, electromagnetic, ou
behavioral—that modulate RSN activity is crucial to advance in the development of novel
therapeutic strategies.
This study revealed interesting findings that contribute to our understanding of the effects of
STN-DBS on the occupancy of RSNs. Our analysis shows that orbitofrontal RSN occupancy
increases significantly when STN-DBS is turned ON, compared with when it is turned OFF and
compared with healthy controls. STN-DBS showed a tendency to normalize the occupancy of
a somatomotor RSN, although not sufficiently significant (p = 0.053 for K = 15; p = 0.027 pour
K = 8, uncorrected). En plus, the occupancy of a diffuse limbic RSN was increased
compared with controls only with STN-DBS OFF, which demonstrates a trend towards healthy
controls with STN-DBS ON. We found that the occupancy of these RSNs was only weakly
correlated with UPDRS-III, the motor component of UPDRS (0.05 < p < 0.1).
STN, the OFC, and the Limbic Network
The OFC is associated with reward processing, decision-making and prediction (Kringelbach,
2005). This coincides with the finding that ventromedial STN-DBS modulates mood in PD
patients and that orbitofrontal-STN structural connectivity correlates with impulsivity and
behavior during a gambling task (Mosley, Paliwal, et al., 2020; Petry-Schmelzer et al.,
2019). More broadly, intrinsic STN connectivity to a wider limbic network has been identified
both in health and in STN-DBS, particularly in the context of post-operative neuropsychiatric
side effects (Morris et al., 2016; Petry-Schmelzer et al., 2019). Furthermore, electrophysiolog-
ical studies have identified frontosubthalamic network coherence modulation during
STN-DBS (Aron et al., 2016; Frank et al., 2007). Anatomically, Haynes and Haber (2013) also
previously demonstrated that axons project directly from the OFC to the medial tip of the STN,
providing a possible direct pathway for orbitofrontal-STN interaction. The results of our study
correlate well with previously published work, and this hyperdirect pathway may represent the
mechanism by which OFC modulation occurred.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
a
_
0
0
2
9
7
p
d
t
.
STN and Somatomotor Network
STN-DBS is known to modulate BOLD signal within the somatomotor network (Kahan et al.,
2014, 2019; Shen et al., 2020). The degree to which this somatomotor network modulation
occurs correlates with DBS electrode placement within the motor STN; indeed, improvement
in motor outcome is predicted by STN-DBS connectivity to primary motor cortex and nega-
tively associated with other regions, which interestingly included the OFC (Horn et al., 2017;
Horn, Wenzel, et al., 2019). In this study, it was expected that STN-DBS would normalize the
occupancy of somatomotor network connectivity towards that of healthy controls. However,
while our study data demonstrate a trend towards normalization of somatomotor PL state
occupancy, it does not reach statistical significance. This may be due, on one side, to the small
patient numbers leading to an underpowered study, and on the other, to the methodological
constraints of LEiDA, which considers only the “dominant” RSN at each instant of time. This
means that there may be modulation of RSNs occurring at a secondary level that is not
detected using this method.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Global Signal Detection
The most dominant pattern is a global state in which the fMRI signals of all brain areas are aligned
in phase, as observed in previous resting-state fMRI analyses using LEiDA (Cabral, Vidaurre, et al.,
2017; Figueroa et al., 2019; Lord et al., 2019). This is related to the global signal present in fMRI
Network Neuroscience
489
Modulation of limbic resting-state networks by STN-DBS
studies, the significance of which remains controversial. This global signal has been treated as a
nuisance signal and regressed out of fMRI analysis, but growing evidence suggests that it contains
neurophysiological information (Hahamy et al., 2014; Liu et al., 2017). In our dataset there is an
indication that the occupancy of this global state may be increased with STN-DBS ON, but this
finding is not statistically significant (p > 0.05). The role of this global BOLD phase coherence
state remains unclear and needs further investigation (Cabral, Vidaurre, et coll., 2017).
Clinical Relevance
The non-motor side effects of STN-DBS are not fully understood, but they may be associated with
the interaction of STN-DBS with disseminated neural networks. This study indicates that STN-DBS
modulates limbic networks that may contribute to the neuropsychiatric side effects seen in
STN-DBS (Volkmann et al., 2010). Frontosubthalamic network coherence has previously been
identified as a key component of cognitive processes such as high-conflict decision-making, lequel
may be disrupted by STN-DBS (Antoniades et al., 2014; Aron et al., 2016; Frank et al., 2007).
In PD patients, impulse-control disorders are associated with altered dynamic functional
connectivity within the limbic network as well as increased BOLD activation in OFC (Tessitore
et coll., 2017). With respect to STN-DBS for PD, Mosley et al. demonstrated the direct structural
and functional relevance of STN-OFC connectivity in the development of non-motor symptom
modulation (Mosley et al., 2019; Mosley, Paliwal, et coll., 2020; Mosley, Robinson, et coll., 2020).
The results of this study add further evidence of STN-OFC functional connectivity and limbic
network modulation during STN-DBS.
This study also demonstrates a significant difference in diffuse limbic RSN occupancy with
DBS OFF compared with healthy controls, which is no longer present with DBS ON. Previous
studies have demonstrated a correlation between diffuse limbic connectivity and
apathy/depression in PD, and recent work has highlighted the potential for improvement in these
symptoms according to DBS electrode placement within the STN (Dafsari, Petry-Schmelzer,
et coll., 2018; Dan et al., 2017; Petry-Schmelzer et al., 2019). Although post-operative UPDRS-
I was used here as an estimate of non-motor symptoms, we also identified a correlation between
depressive symptoms and diffuse limbic RSN occupancy. In the context of previous work, ce
diffuse limbic modulation may contribute to the post-operative clinical presentation.
Strengths and Limitations
Strengths. Because of historic concerns about the MRI compatibility of DBS hardware, only a
few post-operative fMRI datasets exist (Corne, Wenzel, et coll., 2019; Shen et al., 2020). Func-
tional neuroimaging both ON and OFF STN-DBS allowed us to investigate real-time effects of
STN-DBS on the brain’s functional connectivity, giving more granular insights into the wide-
spread effects of STN-DBS.
Another key strength is the use of LEiDA, which demonstrates high sensitivity to RSN activity
in the dynamical analysis of fMRI data, crucial for the estimation of RSN. This approach reveals
network patterns that may occur only briefly and sporadically (c'est à dire., <5% of the time) but that are
recurrent across scans and subjects. It should be taken into consideration that the results of
LEiDA analysis differ from conventional connectivity methods of fMRI data analysis, relying
on coactivation or correlation. Importantly, the validity of the results obtained herein is rein-
forced by the correspondence with structural MRI, electrophysiological, and anatomical studies.
The partition of phase-locking patterns using the k-means clustering algorithm allowed the
identification of the most significant change in RSN occupancy between STN-DBS ON and
OFF (Figueroa et al., 2019). Although it is possible to critique this method as being circular,
Network Neuroscience
490
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
a
_
0
0
2
9
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
this is rather an informed selection that is subsequently validated by the fact that the PL state
revealing the most significant difference in occupancy between STN-DBS ON and OFF
remained robust and stable for partitions between 10 to 20 (see Figure 2C) when compared
a posteriori with the data from healthy controls, and our results were found to remain
consistent and meaningful. It is important to note that the objective of this study was not
to identify all RSNs that occur but to identify those that are most significantly modulated by
STN-DBS.
This work reinforces the value of quantitative measures to evaluate the impact of perturba-
tive strategies on resting-state activity. Such methods offer sensitivity to network-specific
modulation and can be applied to any existing dataset contrasting two or more resting-state
conditions (Caetano et al., 2022; Magalhães et al., 2021).
Limitations. The clinical generalizability of this particular work is limited. Non-motor symp-
tom scores were not collected for these patients during the experimental visit, so it was not
possible to directly correlate the results of this study with the clinical symptoms associated
with OFC and limbic network modulation. Given the historical nature of the data, and as
an estimate of the non-motor effects of STN-DBS, the pre- and post-operative UPDRS-I scores
were retrospectively collected and used in the analysis here. The correlations presented here
should be interpreted with caution, but the inclusion of this analysis does give an indication of
possible correlated phenotypes and generates hypotheses for future studies.
There are also two key areas of variability both within and between groups that should also
be highlighted. First, within the patient group, the variability of treatment duration (from 8 to
102 months) may affect the generalizability of this analysis. With a larger sample size, the
effect of treatment duration on RSN occupancy could be probed, but this was not possible
here. Second, the fMRI acquisition parameters were slightly different between healthy controls
and the patient group. This was due to safety considerations but should be noted when
replicating this work.
The participant population excluded those with significant head tremor, so only 10 PD
patients were included in this study, which increases the risk of false negative results (type
II error), limiting the detection of differences in RSN occupancy between STN-DBS ON and
OFF and controls. The small sample size may also account for the fact that the correlation
between somatomotor RSN occupancy and UPDRS-III did not meet statistical significance.
Future Work
This work highlighted the modulation of orbitofrontal and limbic network activity during
STN-DBS. The correlation between these networks and the non-motor symptoms of PD in
larger patient cohorts with non-motor clinical outcome measurement certainly deserves
further investigation. Further, understanding how the modulation of different RSNs relates to
electrode location relative to white fiber tracts (“sweet spot” analysis), volume of tissue
activated, and stimulation frequency/intensity is likely to provide a deeper understanding of
the therapeutic potential of brain stimulation strategies (Dembek et al., 2019; Hollunder
et al., 2021). Electrophysiology methods including EEG and MEG should also be considered
to address the temporal limitations of fMRI data analysis.
CONCLUSION
This study provides further insights into the neuromodulatory effect of STN-DBS on the reper-
toire of disseminated brain networks at rest. In particular, the most significant network
Network Neuroscience
491
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
a
_
0
0
2
9
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
modulation occurred within the OFC, which was more likely to occur with STN-DBS ON.
Future work should optimize patient selection, sample size, data acquisition, and outcome
measurement in order to interrogate the correlation of these findings with clinical outcomes.
ACKNOWLEDGMENTS
This study was funded by the Brain Research Trust (https://www.brt.org.uk). J.E. received fund-
ing from the National Institute of Health Research (NIHR). J.C. was funded by the Portuguese
Foundation for Science and Technology, Portugal (UIDB/50026/2020, UIDP/50026/2020, and
CEECIND/03325/2017) and by “La Caixa” Foundation project BRAINSTIM (LCF/BQ/PR22/
11920014). J.K. received funding from the Astor Foundation, the Rosetrees Trust, and the
MHMS General Charitable Trust. T.F. The work was undertaken by UCL/UCLH, who receives
a proportion of funding from the UK Department of Health’s NIHR Biomedical Research
Centres funding scheme. The funders had no role in study design, data collection and analysis,
decision to publish, or preparation of the manuscript.
AUTHOR CONTRIBUTIONS
John Eraifej: Conceptualization; Formal analysis; Methodology; Writing – original draft;
Writing – review & editing. Joana Cabral: Conceptualization; Investigation; Methodology;
Visualization; Writing – original draft. Henrique M. Fernandes: Formal analysis; Methodology.
Joshua Kahan: Investigation. Shenghong He: Visualization; Writing – review & editing. Laura
Mancini: Data acquisition. John Thornton: Data acquisition; Writing – review & editing. Mark
White: Data acquisition. Tarek Yousry: Data acquisition. Ludvic Zrinzo: Patient recruitment
and data acquisition; Supervision. Harith Akram: Data acquisition; Resources; Writing –
review & editing. Patricia Limousin: Supervision. Tom Foltynie: Supervision. Tipu Z. Aziz:
Supervision. Gustavo Deco: Conceptualization; Methodology. Morten Kringelbach: Concep-
tualization; Investigation; Methodology; Supervision; Writing – original draft. Alexander L.
Green: Conceptualization; Investigation; Methodology; Supervision; Writing – original draft.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
a
_
0
0
2
9
7
p
d
.
t
REFERENCES
Abbes, M., Lhommée, E., Thobois, S., Klinger, H., Schmitt, E.,
Bichon, A., Castrioto, A., Xie, J., Fraix, V., Kistner, A., Pélissier,
P., Seigneuret, É., Chabardès, S., Mertens, P., Broussolle, E.,
Moro, E., & Krack, P. (2018). Subthalamic stimulation and neu-
ropsychiatric symptoms in Parkinson’s disease: Results from a
long-term follow-up cohort study. Journal of Neurology, Neuro-
surgery, and Psychiatry, 89(8), 836–843. https://doi.org/10.1136
/jnnp-2017-316373, PubMed: 29436490
Accolla, E. A., & Pollo, C. (2019). Mood effects after deep brain
stimulation for Parkinson’s disease: An update. Frontiers in Neu-
rology, 10, 617. https://doi.org/10.3389/fneur.2019.00617,
PubMed: 31258509
Alonso Martínez, S., Deco, G., Ter Horst, G. J., & Cabral, J. (2020).
The dynamics of functional brain networks associated with
depressive symptoms in a nonclinical sample. Frontiers in Neural
Circuits, 14, 570583. https://doi.org/10.3389/fncir.2020.570583,
PubMed: 33071760
Antoniades, C. A., Bogacz, R., Kennard, C., FitzGerald, J. J., Aziz,
T., & Green, A. L. (2014). Deep brain stimulation abolishes
slowing of reactions to unlikely stimuli. Journal of Neuroscience,
34(33), 10844–10852. https://doi.org/10.1523/JNEUROSCI.1065
-14.2014, PubMed: 25122887
Aron, A. R., Herz, D. M., Brown, P., Forstmann, B. U., & Zaghloul,
K. (2016). Frontosubthalamic circuits for control of action and
cognition. Journal of Neuroscience, 36(45), 11489–11495.
https://doi.org/10.1523/ JNEUROSCI.2348-16.2016, PubMed:
27911752
Boertien, T., Zrinzo, L., Kahan, J., Jahanshahi, M., Hariz, M.,
Mancini, L., Limousin, P., & Foltynie, T. (2011). Functional
imaging of subthalamic nucleus deep brain stimulation in
Parkinson’s disease. Movement Disorders, 26(10), 1835–1843.
https://doi.org/10.1002/mds.23788, PubMed: 21674623
Cabral, J., Kringelbach, M. L., & Deco, G. (2017). Functional
connectivity dynamically evolves on multiple time-scales over
a static structural connectome: Models and mechanisms. Neuro-
Image, 160, 84–96. https://doi.org/10.1016/j.neuroimage.2017
.03.045, PubMed: 28343985
Cabral, J., Vidaurre, D., Marques, P., Magalhães, R., Silva Moreira,
P., Miguel Soares, J., Deco, G., Sousa, N., & Kringelbach, M. L.
(2017). Cognitive performance in healthy older adults relates to
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
492
Modulation of limbic resting-state networks by STN-DBS
spontaneous switching between states of functional connectivity
during rest. Scientific Reports, 7(1), 5135. https://doi.org/10.1038
/s41598-017-05425-7, PubMed: 28698644
Caetano, I., Ferreira, S., Coelho, A., Amorim, L., Castanho, T. C.,
Portugal-Nunes, C., Soares, J. M., Gonçalves, N., Sousa, R., Reis,
J., Lima, C., Marques, P., Moreira, P. S., Rodrigues, A. J., Santos,
N. C., Morgado, P., Magalhães, R., Picó-Pérez, M., Cabral, J., &
Sousa, N. (2022). Perceived stress modulates the activity
between the amygdala and the cortex. Molecular Psychiatry,
27(12), 4939–4947. https://doi.org/10.1038/s41380-022-01780-8,
PubMed: 36117211
Calhoun, V. D., Miller, R., Pearlson, G., & Adali, T. (2014). The
chronnectome: Time-varying connectivity networks as the next
frontier in fMRI data discovery. Neuron, 84(2), 262–274.
https://doi.org/10.1016/j.neuron.2014.10.015, PubMed:
25374354
Carmichael, D. W., Pinto, S., Limousin-Dowsey, P., Thobois, S.,
Allen, P. J., Lemieux, L., Yousry, T., & Thornton, J. S. (2007).
Functional MRI with active, fully implanted, deep brain stimula-
tion systems: Safety and experimental confounds. NeuroImage,
37(2), 508–517. https://doi.org/10.1016/j.neuroimage.2007.04
.058, PubMed: 17590355
Chen, J. E., Lewis, L. D., Chang, C., Tian, Q., Fultz, N. E., Ohringer,
N. A., Rosen, B. R., & Polimeni, J. R. (2020). Resting-state “phys-
iological networks.” NeuroImage, 213, 116707. https://doi.org
/10.1016/j.neuroimage.2020.116707, PubMed: 32145437
Dafsari, H. S., Petry-Schmelzer, J. N., Ray-Chaudhuri, K., Ashkan,
K., Weis, L., Dembek, T. A., Samuel, M., Rizos, A., Silverdale, M.,
Barbe, M. T., Fink, G. R., Evans, J., Martinez-Martin, P., Antonini,
A., Visser-Vandewalle, V., & Timmermann, L. (2018). Non-motor
outcomes of subthalamic stimulation in Parkinson’s disease
depend on location of active contacts. Brain Stimulation, 11(4),
904–912. https://doi.org/10.1016/j.brs.2018.03.009, PubMed:
29655586
Dafsari, H. S., Reddy, P., Herchenbach, C., Wawro, S., Petry-
Schmelzer, J. N., Visser-Vandewalle, V., Rizos, A., Silverdale,
M., Ashkan, K., Samuel, M., Evans, J., Huber, C. A., Fink,
G. R., Antonini, A., Chaudhuri, K. R., Martinez-Martin, P., &
Timmermann, L. (2016). Beneficial effects of bilateral subtha-
lamic stimulation on non-motor symptoms in Parkinson’s
disease. Brain Stimulation, 9(1), 78–85. https://doi.org/10.1016/j
.brs.2015.08.005, PubMed: 26385442
Dafsari, H. S., Weiss, L., Silverdale, M., Rizos, A., Reddy, P.,
Ashkan, K., Evans, J., Reker, P., Petry-Schmelzer, J. N., Samuel,
M., Visser-Vandewalle, V., Antonini, A., Martinez-Martin, P.,
Chaudhuri, K. R., & Timmermann, L. (2018). Short-term quality
of life after subthalamic stimulation depends on non-motor
symptoms in Parkinson’s disease. Brain Stimulation, 11(4),
867–874. https://doi.org/10.1016/j.brs.2018.02.015, PubMed:
29655587
Damoiseaux, J. S., Rombouts, S. A. R. B., Barkhof, F., Scheltens, P.,
Stam, C. J., Smith, S. M., & Beckmann, C. F. (2006). Consistent
resting-state networks across healthy subjects. Proceedings of the
National Academy of Sciences, 103(37), 13848–13853. https://
doi.org/10.1073/pnas.0601417103, PubMed: 16945915
Dan, R., Růžička, F., Bezdicek, O., Růžička, E., Roth, J., Vymazal, J.,
Goelman, G., & Jech, R. (2017). Separate neural representations
of depression, anxiety and apathy in Parkinson’s disease.
Scientific Reports, 7(1), 12164. https://doi.org/10.1038/s41598
-017-12457-6, PubMed: 28939804
De Hemptinne, C., Ryapolova-Webb, E. S., Air, E. L., Garcia, P. A.,
Miller, K. J., Ojemann, J. G., Ostrem, J. L., Galifianakis, N. B., &
Starr, P. A. (2013). Exaggerated phase-amplitude coupling in the
primary motor cortex in Parkinson disease. Proceedings of the
National Academy of Sciences, 110(12), 4780–4785. https://doi
.org/10.1073/pnas.1214546110, PubMed: 23471992
De Hemptinne, C., Swann, N. C., Ostrem, J. L., Ryapolova-Webb, E. S.,
San Luciano, M., Galifianakis, N. B., & Starr, P. A. (2015). Thera-
peutic deep brain stimulation reduces cortical phase-amplitude
coupling in Parkinson’s disease. Nature Neuroscience, 18(5),
779–786. https://doi.org/10.1038/nn.3997, PubMed: 25867121
De Luca, M., Beckmann, C. F., De Stefano, N., Matthews, P. M., &
Smith, S. M. (2006). fMRI resting state networks define distinct
modes of long-distance interactions in the human brain. Neuro-
Image, 29(4), 1359–1367. https://doi.org/10.1016/j.neuroimage
.2005.08.035, PubMed: 16260155
Dembek, T. A., Roediger, J., Horn, A., Reker, P., Oehrn, C., Dafsari,
H. S., Li, N., Kühn, A. A., Fink, G. R., Visser-Vandewalle, V.,
Barbe, M. T., & Timmermann, L. (2019). Probabilistic sweet spots
predict motor outcome for deep brain stimulation in Parkinson
disease. Annals of Neurology, 86(4), 527–538. https://doi.org
/10.1002/ana.25567, PubMed: 31376171
Deuschl, G., Schade-Brittinger, C., Krack, P., Volkmann, J., Schäfer,
H., Bötzel, K., Daniels, C., Deutschländer, A., Dillmann, U.,
Eisner, W., Grube, D., Hamel, W., Herzog, J., Hilker, R., Klebe,
S., Kloss, M., Koy, J., Krause, M., Kupsch, A., … German
Parkinson Study Group, Neurostimulation Section. (2006). A
randomized trial of deep-brain simulation for Parkinson. New
England Journal of Medicine, 355(9), 896–908. https://doi.org
/10.1056/NEJMoa060281, PubMed: 16943402
Eusebio, A., Thevathasan, W., Doyle Gaynor, L., Pogosyan, A., Bye,
E., Foltynie, T., Zrinzo, L., Ashkan, K., Aziz, T., & Brown, P.
(2011). Deep brain stimulation can suppress pathological
synchronisation in parkinsonian patients. Journal of Neurology,
Neurosurgery, and Psychiatry, 82(5), 569–573. https://doi.org/10
.1136/jnnp.2010.217489, PubMed: 20935326
Fasano, M. C., Cabral, J., Stevner, A., Vuust, P., Cantou, P., Brattico,
E., & Kringelbach, M. L. (2023). The early adolescent brain on
music: Analysis of functional dynamics reveals engagement of
orbitofrontal cortex reward system. Human Brain Mapping, 44(2),
429–446. https://doi.org/10.1002/hbm.26060, PubMed: 36069619
Figueroa, C. A., Cabral, J., Mocking, R. J. T., Rapuano, K. M., van
Hartevelt, T. J., Deco, G., Expert, P., Schene, A. H., Kringelbach,
M. L., & Ruhé, H. G. (2019). Altered ability to access a clinically
relevant control network in patients remitted from major depres-
sive disorder. Human Brain Mapping, 40(9), 2771–2786. https://
doi.org/10.1002/hbm.24559, PubMed: 30864248
Fornito, A., Zalesky, A., & Breakspear, M. (2015). The connec-
tomics of brain disorders. Nature Reviews Neuroscience, 16(3),
159–172. https://doi.org/10.1038/nrn3901, PubMed: 25697159
Frank, M. J., Samanta, J., Moustafa, A. A., & Sherman, S. J. (2007).
Hold your horses: Impulsivity, deep brain stimulation, and med-
ication in parkinsonism. Science, 318(5854), 1309–1312. https://
doi.org/10.1126/science.1146157, PubMed: 17962524
Gradinaru, V., Mogri, M., Thompson, K. R., Henderson, J. M., &
Deisseroth, K. (2009). Optical deconstruction of parkinsonian
Network Neuroscience
493
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
a
_
0
0
2
9
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
neural circuitry. Science, 324(5925), 354–359. https://doi.org/10
.1126/science.1167093, PubMed: 19299587
Hahamy, A., Calhoun, V., Pearlson, G., Harel, M., Stern, N., Attar,
F., Malach, R., & Salomon, R. (2014). Save the global: Global
signal connectivity as a tool for studying clinical populations
with functional magnetic resonance imaging. Brain Connectivity,
4(6), 395–403. https://doi.org/10.1089/ brain.2014.0244,
PubMed: 24923194
Hashimoto, T., Elder, C. M., Okun, M. S., Patrick, S. K., & Vitek, J. L.
(2003). Stimulation of the subthalamic nucleus changes the firing
pattern of pallidal neurons. Journal of Neuroscience, 23(5),
1916–1923. https://doi.org/10.1523/JNEUROSCI.23-05-01916
.2003, PubMed: 12629196
Haynes, W. I. A., & Haber, S. N. (2013). The organization of
prefrontal-subthalamic inputs in primates provides an anatomical
substrate for both functional specificity and integration: Implica-
tions for basal ganglia models and deep brain stimulation.
Journal of Neuroscience, 33(11), 4804–4814. https://doi.org/10
.1523/JNEUROSCI.4674-12.2013, PubMed: 23486951
Hollunder, B., Ganos, C., & Horn, A. (2021). Deep brain stimula-
tion: From sweet spots to sweet networks? Biological Psychiatry:
Cognitive Neuroscience and Neuroimaging, 6(10), 939–941.
https://doi.org/10.1016/j.bpsc.2021.06.002, PubMed: 34625219
Horn, A., & Kühn, A. A. (2015). Lead-DBS: A toolbox for deep brain
stimulation electrode localizations and visualizations. Neuro-
Image, 107, 127–135. https://doi.org/10.1016/j.neuroimage
.2014.12.002, PubMed: 25498389
Horn, A., Li, N., Dembek, T. A., Kappel, A., Boulay, C., Ewert, S.,
Tietze, A., Husch, A., Perera, T., Neumann, W. J., Reisert, M., Si,
H., Oostenveld, R., Rorden, C., Yeh, F. C., Fang, Q., Herrington,
T. M., Vorwerk, J., & Kühn, A. A. (2019). Lead-DBS v2: Towards a
comprehensive pipeline for deep brain stimulation imaging.
NeuroImage, 184, 293–316. https://doi.org/10.1016/j
.neuroimage.2018.08.068, PubMed: 30179717
Horn, A., Reich, M., Vorwerk, J., Li, N., Wenzel, G., Fang, Q.,
Schmitz-Hübsch, T., Nickl, R., Kupsch, A., Volkmann, J., Kühn,
A. A., & Fox, M. D. (2017). Connectivity predicts deep brain
stimulation outcome in Parkinson disease. Annals of Neurology,
82(1), 67–78. https://doi.org/10.1002/ana.24974, PubMed:
28586141
Horn, A., Wenzel, G., Irmen, F., Huebl, J., Li, N., Neumann, W. J.,
Krause, P., Bohner, G., Scheel, M., & Kühn, A. A. (2019). Deep
brain stimulation induced normalization of the human functional
connectome in Parkinson’s disease. Brain, 142(10), 3129–3143.
https://doi.org/10.1093/brain/awz239, PubMed: 31412106.
Kahan, J., Mancini, L., Flandin, G., White, M., Papadaki, A.,
Thornton, J., Yousry, T., Zrinzo, L., Hariz, M., Limousin, P.,
Friston, K., & Foltynie, T. (2019). Deep brain stimulation has
state-dependent effects on motor connectivity in Parkinson’s
disease. Brain, 142(8), 2417–2431. https://doi.org/10.1093
/brain/awz164, PubMed: 31219504
Kahan, J., Papadaki, A., White, M., Mancini, L., Yousry, T., Zrinzo, L.,
Limousin, P., Hariz, M., Foltynie, T., & Thornton, J. (2015). The
safety of using body-transmit MRI in patients with implanted deep
brain stimulation devices. PLoS ONE, 10(6), e0129077. https://doi
.org/10.1371/journal.pone.0129077, PubMed: 26061738
Kahan, J., Urner, M., Moran, R., Flandin, G., Marreiros, A.,
Mancini, L., White, M., Thornton, J., Yousry, T., Zrinzo, L., Hariz,
M., Limousin, P., Friston, K., & Foltynie, T. (2014). Resting state
functional MRI in Parkinson’s disease: The impact of deep brain
stimulation on “effective” connectivity. Brain, 137(4), 1130–1144.
https://doi.org/10.1093/brain/awu027, PubMed: 24566670
Krack, P., Batir, A., Van Blercom, N., Chabardes, S., Fraix, V., Ardouin,
C., Koudsie, A., Limousin, P. D., Benazzouz, A., LeBas, J. F.,
Benabid, A.-L., & Pollak, P. (2003). Five-year follow-up of bilateral
stimulation of the subthalamic nucleus in advanced Parkinson’s
disease. New England Journal of Medicine, 349(20), 1925–1934.
https://doi.org/10.1056/NEJMoa035275, PubMed: 14614167
Kringelbach, M. L. (2005). The human orbitofrontal cortex: Linking
reward to hedonic experience. Nature Reviews Neuroscience,
6(9), 691–702. https://doi.org/10.1038/nrn1747, PubMed:
16136173
Kringelbach, M. L., Green, A. L., & Aziz, T. Z. (2011). Balancing
the brain: Resting state networks and deep brain stimulation.
Frontiers in Integrative Neuroscience, 5, 8. https://doi.org/10
.3389/fnint.2011.00008, PubMed: 21577250
Larabi, D. I., Renken, R. J., Cabral, J., Marsman, J.-B. C., Aleman,
A., & Ćurčić-Blake, B. (2020). Trait self-reflectiveness relates to
time-varying dynamics of resting state functional connectivity
and underlying structural connectomes: Role of the default mode
network. NeuroImage, 219, 116896. https://doi.org/10.1016/j
.neuroimage.2020.116896, PubMed: 32470573
Liu, T. T., Nalci, A., & Falahpour, M. (2017). The global signal in
fMRI: Nuisance or information? NeuroImage, 150, 213–229.
https://doi.org/10.1016/j.neuroimage.2017.02.036, PubMed:
28213118
Lord, L. D., Expert, P., Atasoy, S., Roseman, L., Rapuano, K.,
Lambiotte, R., Nutt, D. J., Deco, G., Carhart-Harris, R. L.,
Kringelbach, M. L., & Cabral, J. (2019). Dynamical exploration
of the repertoire of brain networks at rest is modulated by
psilocybin. NeuroImage, 199, 127–142. https://doi.org/10.1016
/j.neuroimage.2019.05.060, PubMed: 31132450
Magalhães, R., Picó-Pérez, M., Esteves, M., Vieira, R., Castanho,
T. C., Amorim, L., Sousa, M., Coelho, A., Fernandes, H. M.,
Cabral, J., Moreira, P. S., & Sousa, N. (2021). Habitual coffee
drinkers display a distinct pattern of brain functional connectiv-
ity. Molecular Psychiatry, 26, 6589–6598. https://doi.org/10
.1038/s41380-021-01075-4, PubMed: 33875801
Morris, L. S., Kundu, P., Baek, K., Irvine, M. A., Mechelmans, D. J.,
Wood, J., Harrison, N. A., Robbins, T. W., Bullmore, E. T., &
Voon, V. (2016). Jumping the gun: Mapping neural correlates of
waiting impulsivity and relevance across alcohol misuse. Biolog-
ical Psychiatry, 79(6), 499–507. https://doi.org/10.1016/j
.biopsych.2015.06.009, PubMed: 26185010
Mosley, P. E., Paliwal, S., Robinson, K., Coyne, T., Silburn, P.,
Tittgemeyer, M., Stephan, K. E., Breakspear, M., & Perry, A.
(2019). The structural connectivity of discrete networks underlies
impulsivity and gambling in Parkinson’s disease. Brain, 142(12),
3917–3935. https://doi.org/10.1093/ brain/awz327, PubMed:
31665241
Mosley, P. E., Paliwal, S., Robinson, K., Coyne, T., Silburn, P.,
Tittgemeyer, M., Stephan, K. E., Perry, A., & Breakspear, M.
(2020). The structural connectivity of subthalamic deep brain
stimulation correlates with impulsivity in Parkinson’s disease.
Brain, 143(7), 2235–2254. https://doi.org/10.1093/ brain
/awaa148, PubMed: 32568370
Network Neuroscience
494
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
a
_
0
0
2
9
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Modulation of limbic resting-state networks by STN-DBS
Mosley, P. E., Robinson, K., Coyne, T., Silburn, P., Barker, M. S.,
Breakspear, M., Robinson, G. A., & Perry, A. (2020). Subthalamic
deep brain stimulation identifies frontal networks supporting
initiation, inhibition and strategy use in Parkinson’s disease.
N e u ro I m a g e , 2 2 3 , 11 7 3 5 2 . h t t p s : / / d o i . o rg / 1 0 . 1 0 1 6 / j
.neuroimage.2020.117352, PubMed: 32916288
Petry-Schmelzer, J. N., Krause, M., Dembek, T. A., Horn, A., Evans,
J., Ashkan, K., Rizos, A., Silverdale, M., Schumacher, W., Sack,
C., Loehrer, P. A., Fink, G. R., Fonoff, E. T., Martinez-Martin, P.,
Antonini, A., Barbe, M. T., Visser-Vandewalle, V., Ray-
Chaudhuri, K., Timmermann, L., & Dafsari, H. S. (2019). Non-
motor outcomes depend on location of neurostimulation in
Parkinson’s disease. Brain, 142(11), 3592–3604. https://doi.org
/10.1093/brain/awz285, PubMed: 31553039
Rughani, A., Schwalb, J. M., Sidiropoulos, C., Pilitsis, J., Ramirez-
Zamora, A., Sweet, J. A., Mittal, S., Espay, A. J., Martinez, J. G.,
Abosch, A., Eskandar, E., Gross, R., Alterman, R., & Hamani, C.
(2018). Congress of neurological surgeons systematic review and
evidence-based guideline on subthalamic nucleus and globus
pallidus internus deep brain stimulation for the treatment of patients
with Parkinson’s disease: Executive summary. Neurosurgery, 82(6),
753–756. https://doi.org/10.1093/neuros/nyy037, PubMed: 29538685
Saenger, V. M., Kahan, J., Foltynie, T., Friston, K., Aziz, T. Z., Green,
A. L., van Hartevelt, T. J., Cabral, J., Stevner, A. B. A., Fernandes,
H. M., Mancini, L., Thornton, J., Yousry, T., Limousin, P., Zrinzo,
L., Hariz, M., Marques, P., Sousa, N., Kringelbach, M. L., &
Deco, G. (2017). Uncovering the underlying mechanisms and
whole-brain dynamics of deep brain stimulation for Parkinson’s
disease. Scientific Reports, 7(1), 9882. https://doi.org/10.1038
/s41598-017-10003-y, PubMed: 28851996
Shen, L., Jiang, C., Hubbard, C. S., Ren, J., He, C., Wang, D., Dahmani,
L., Guo, Y., Liu, Y., Xu, S., Meng, F., Zhang, J., Liu, H., & Li, L. (2020).
Subthalamic nucleus deep brain stimulation modulates 2 distinct
neurocircuits. Annals of Neurology, 88(6), 1178–1193. https://doi
.org/10.1002/ana.25906, PubMed: 32951262
Stam, C. J. (2014). Modern network science of neurological disor-
ders. Nature Reviews Neuroscience, 15(10), 683–695. https://doi
.org/10.1038/nrn3801, PubMed: 25186238
Tahmasian, M., Bettray, L. M., van Eimeren, T., Drzezga, A.,
Timmermann, L., Eickhoff, C. R., Eickhoff, S. B., & Eggers, C.
(2015). A systematic review on the applications of resting-state
fMRI in Parkinson’s disease: Does dopamine replacement
therapy play a role? Cortex, 73, 80–105. https://doi.org/10
.1016/j.cortex.2015.08.005, PubMed: 26386442
Tessitore, A., De Micco, R., Giordano, A., di Nardo, F., Caiazzo, G.,
Siciliano, M., De Stefano, M., Russo, A., Esposito, F., & Tedeschi,
G. (2017). Intrinsic brain connectivity predicts impulse control
disorders in patients with Parkinson’s disease. Movement Disor-
ders, 32(12), 1710–1719. https://doi.org/10.1002/mds.27139,
PubMed: 28949049
Tononi, G., & Edelman, G. M. (1998). Consciousness and complex-
ity. Science, 282(5395), 1846–1851. https://doi.org/10.1126
/science.282.5395.1846, PubMed: 9836628
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F.,
Etard, O., Delcroix, N., Mazoyer, B., & Joliot, M. (2002). Auto-
mated anatomical labeling of activations in SPM using a macro-
scopic anatomical parcellation of the MNI MRI single-subject
brain. NeuroImage, 15(1), 273–289. https://doi.org/10.1006
/nimg.2001.0978, PubMed: 11771995
Vitek, J. L., Jain, R., Chen, L., Tröster, A. I., Schrock, L. E., House,
P. A., Giroux, M. L., Hebb, A. O., Farris, S. M., Whiting, D. M.,
Leichliter, T. A., Ostrem, J. L., San Luciano, M., Galifianakis, N.,
Verhagen Metman, L., Sani, S., Karl, J. A., Siddiqui, M. S., Tatter,
S. B., … Starr, P. A. (2020). Subthalamic nucleus deep brain stim-
ulation with a multiple independent constant current-controlled
device in Parkinson’s disease (INTREPID): A multicentre,
double-blind, randomised, sham-controlled study. The Lancet
Neurology, 19(6), 491–501. https://doi.org/10.1016/S1474-4422
(20)30108-3, PubMed: 32470421
Vohryzek, J., Deco, G., Cessac, B., Kringelbach, M. L., & Cabral, J.
(2020). Ghost attractors in spontaneous brain activity: Recurrent
excursions into functionally relevant BOLD phase-locking states.
Frontiers in Systems Neuroscience, 14, 20. https://doi.org/10
.3389/fnsys.2020.00020, PubMed: 32362815
Volkmann, J., Daniels, C., & Witt, K. (2010). Neuropsychiatric
effects of subthalamic neurostimulation in Parkinson disease.
Nature Reviews Neurology, 6(9), 487–498. https://doi.org/10
.1038/nrneurol.2010.111, PubMed: 20680036
Williams, L. M. (2016). Precision psychiatry: A neural circuit
taxonomy for depression and anxiety. The Lancet Psychiatry,
3(5), 472–480. https://doi.org/10.1016/S2215-0366(15)00579-9,
PubMed: 27150382
Wong, W., Cabral, J., Rane, R., Ly, R., Kringelbach, M. L., &
Feusner, J. D. (2021). Effects of visual attention modulation on
dynamic functional connectivity during own-face viewing in
body dysmorphic disorder. Neuropsychopharmacology, 46(11),
2030–2038. https://doi.org/10.1038/s41386-021-01039-w,
PubMed: 34050267
Xu, W., Russo, G. S., Hashimoto, T., Zhang, J., & Vitek, J. L. (2008).
Subthalamic nucleus stimulation modulates thalamic neuronal
activity. Journal of Neuroscience, 28(46), 11916–11924. https://
doi.org/10.1523/ JNEUROSCI.2027-08.2008, PubMed:
19005057
Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari,
D., Hollinshead, M., Roffman, J. L., Smoller, J. W., Zöllei, L.,
Polimeni, J. R., Fisch, B., Liu, H., & Buckner, R. L. (2011). The
organization of the human cerebral cortex estimated by intrinsic
functional connectivity. Journal of Neurophysiology, 106(3),
1125–1165. https://doi.org/10.1152/jn.00338.2011, PubMed:
21653723
Network Neuroscience
495
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
2
4
7
8
2
1
1
8
3
9
3
n
e
n
_
a
_
0
0
2
9
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3