RECHERCHE
Coupling of the spatial distributions between sMRI
and PET reveals the progression of
Alzheimer’s disease
Kun Zhao1#, Jiaji Lin2#, Martin Dyrba3, Dong Wang4, Tongtong Che1, Haoyang Wu5,
Jingyu Wang5, Yong Liu6
, and Shuyu Li7, for the Alzheimer’s Disease Neuroimaging Initiative
1Beijing Advanced Innovation Center for Biomedical Engineering, School of Biological Science and Medical Engineering,
Beihang University, Beijing, Chine
2Department of Neurology, Second Affiliated Hospital of Air Force Medical University, Xi’an, Chine
3German Center for Neurodegenerative Diseases (DZNE), Rostock, Allemagne
4School of Information Science and Engineering, Shandong Normal University, Jinan, Chine
5School of Basic Medicine, Air Force Medical University, Xi’an, Chine
6School of Artificial Intelligence, Beijing University of Posts and Telecommunications, Beijing, Chine
7State Key Laboratory of Cognitive Neuroscience and Learning, Beijing Normal University, Beijing, Chine
#These authors contributed equally to this work.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
un
_
0
0
2
7
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Mots clés: Alzheimer’s disease, Coupling, Spatial inconsistency, Multimodal, Regional radiomics
similarity network, Progression
ABSTRAIT
Amyloid-beta (Aβ) deposition and altered brain structure are the most relevant neuroimaging
biomarkers for Alzheimer’s disease (AD). Cependant, their spatial inconsistency was always
confusing and misleading. En outre, the relationship between this spatial inconsistency and
AD progression is unclear. The current study introduced a regional radiomics similarity network
(R2SN) to map structural MRI and Aβ positron emission tomography (ANIMAL DE COMPAGNIE) images to study their
cross-modal interregional coupling. A total of 790 participants (248 normal controls, 390 mild
cognitive impaired patients, et 152 AD patients) with their structural MRI and PET images were
studied. The results showed that global and regional R2SN coupling significantly decreased
according to the severity of cognitive decline, from mild cognitive impairment to AD dementia.
The global coupling patterns are discriminative between different APOE ε4, Aβ, and Tau
subgroups. R2SN coupling was probed for relationships with neuropsychiatric measures and
peripheral biomarkers. Kaplan–Meier analysis showed that lower global coupling scores could
reveal worse clinical progression of dementia. The R2SN coupling scores derived from the
coupling between Aβ and atrophy over individual brain regions could reflect the specific
pathway of AD progression, which would be a reliable biomarker for AD.
RÉSUMÉ DE L'AUTEUR
Amyloid-beta (Aβ) deposition and altered brain structure are the most relevant neuroimaging
biomarkers for Alzheimer’s disease (AD). We introduced a novel network coupling measure
based on the regional radiomics similarity network (R2SN) to explore the potential association
between the spatial distributions of brain structure and Aβ based on sMRI and Aβ positron
emission tomography (ANIMAL DE COMPAGNIE) imaging. Dans cette étude, we systematically demonstrated that the
alteration of the coupling between brain networks of brain structure and Aβ accumulation
could serve as a predictor for revealing the distinct progression of AD.
un accès ouvert
journal
Citation: Zhao, K., Lin, J., Dyrba, M.,
Wang, D., Che, T., Wu, H., Wang, J.,
Liu, Y., & Li, S. (2023). Coupling of the
spatial distributions between sMRI
and PET reveals the progression
of Alzheimer’s disease. Réseau
Neurosciences, 7(1), 86–101. https://est ce que je
.org/10.1162/netn_a_00271
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00271
Informations complémentaires:
https://doi.org/10.1162/netn_a_00271;
https://github.com/YongLiulab
Reçu: 4 May 2022
Accepté: 27 Juillet 2022
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Corresponding Authors:
Shuyu Li
shuyuli@bnu.edu.cn
Yong Liu
yongliu@bupt.edu.cn
Éditeur de manipulation:
Olaf Sporns
droits d'auteur: © 2022
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
Spatial coupling of AD based on sMRI and PET
INTRODUCTION
Both amyloid-beta (Aβ) accumulation and altered brain structure and function are the most
relevant noninvasive biomarkers for Alzheimer’s disease (AD) across the spectrum of subjec-
tive cognitive decline, mild cognitive impairment (MCI), and AD dementia (P.. Chen et al.,
2022; Lista et al., 2014; Rathore et al., 2017; H. Wang et al., 2021). In the classical amyloid
cascade hypothesis, it is suggested that Aβ aggregation and deposition in the brain paren-
chyma initiate a sequence of events that further lead to neuronal death, which eventually leads
to atrophy and dementia (Hardy & Higgins, 1992). The idea that higher Aβ causes faster neu-
rodegeneration has been very influential in research (Sepulcre et al., 2018; Zhang et al., 2020).
Cependant, the temporal sequence and causal relationship between Aβ spreading and atrophy
signatures have been questioned because of their spatial inconsistency during AD progression.
Aβ deposition starts in association cortices and spreads from the neocortex to the allocortex,
but brain structure alterations start in the hippocampus and spread from the medial temporal
lobe to the frontal lobe and then progress to the whole brain (van der Kant et al., 2020; W. Oui.
Wang et al., 2015; Yang et al., 2012; Young et al., 2018). Recent studies indicate that neuronal
injury does not necessarily occur where Aβ plaques are deposited (Terry et al., 1991).
Amyloidosis-defined “pure AD neuropathology” is observed in only 30%–50% of patients
with probable AD dementia based on sMRI (Robinson et al., 2018). En général, the relationship
between Aβ accumulation and altered brain structure is still inconclusive. Entre-temps, finding
a more accurate analysis method for their relationship has potentially valuable implications for
understanding the pathogenesis of AD.
Some efforts have been made to obtain cross-modal images of Aβ accumulation and struc-
tural architecture. A previous study applied partial correlation analysis for space-normalized
18F-florbetapir positron emission tomography (ANIMAL DE COMPAGNIE) and T1-weighted MRI scans and found a
negative relationship between global amyloid load and gray matter volume in preclinical AD
cases (X. Wang et al., 2021). A similar covariance-pattern method was applied to cognitively
normal elderly individuals to identify the collaborative relationship between Aβ deposition
and atrophy (Oh et al., 2014). All of the previous studies have demonstrated that the different
atrophy patterns driven by Aβ deposition might lead to distinct AD progression. Cependant, spa-
tial resolution varies within the imaging modalities, and inherently lower signal/noise ratios
(such as PET) make it unlikely that the features extracted from a particular imaging modality
will have the same association with the underlying characteristics as those from another imag-
ing modality (Cook et al., 2018). Network mapping of images may be an excellent solution to
this type of problem. Par exemple, by calculating the correlation coefficient in the time series
between each pair of regions of interest, functional networks were frequently applied to ana-
lyze the characteristics of brain network dynamics (Alexander-Bloch et al., 2013). Interre-
gional similarity networks, such as structural covariance networks (SCNs) or morphometric
similarity networks (MSNs), have been shown to be powerful approaches to capturing ana-
tomical indices (Seidlitz et al., 2018). This is because the brain is a complex information trans-
mission system (Bullmore & Sporns, 2012), and cross-regional mining based on a large-scale
network is better for describing the relevant properties within the brain than is isolated regional
analyse (Alexander-Bloch et al., 2013; Bullmore & Sporns, 2009; Dyrba et al., 2020; Luppi &
Stamatakis, 2021; Tijms et al., 2012).
Ici, we introduced a novel network coupling measure based on the regional radiomics
similarity network (R2SN) (Zhao, Zheng, Che, et coll., 2021; Zhao, Zheng, Dyrba, et coll., 2022) à
explore the potential association between the spatial distributions of Aβ and brain structure
based on sMRI and Aβ PET imaging. Radiomics features can provide comprehensive and sen-
sitive information about brain regions. Network mapping of both Aβ deposition and brain
87
Regional radiomics similarity
réseau (R2SN):
A novel brain network from sMRI
based on the similarity of regional
radiomics features. It has been used
successfully in investigating the
individual cognitive and defining
the subtypes of mild cognitive
impairment.
Radiomics:
A powerful, robust method to extract
more detailed information from each
brain region.
Neurosciences en réseau
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
un
_
0
0
2
7
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Spatial coupling of AD based on sMRI and PET
atrophy signatures based on radiomics features provides better inter- and inner-modal infor-
mation on spatial distribution (Z. J.. Chen et al., 2008; Z. Liu et al., 2021). It can capture alter-
ations in the AD morphological covariation network with robustness, stability, and a biological
basis and serve as a better biomarker in disease diagnosis, mechanistic studies, and progres-
sion tracking than can traditional MRI measures (Zhao, Zheng, Che, et coll., 2021; Zhao, Zheng,
Dyrba, et coll., 2022). The R2SN coupling of sMRI and PET networks was computed to evaluate
coalterations among different brain anatomical regions, which provides a new comprehensive
measure for the spatial distribution of the brain rather than isolated brain regions (Alexander-
Bloch et al., 2013; Zhao, Zheng, Che, et coll., 2021). We evaluated the basis of this R2SN cou-
pling and further systematic findings that may be a predictor for revealing the relationship
between Aβ deposition and structural alterations.
MATERIALS AND METHODS
Acquisition et prétraitement des données
This study included 790 participants (248 normal controls [NCs], 390 MCI patients, et 152
AD patients) with their T1 sMRI and Aβ PET images of AV45 tracer from the Alzheimer’s Dis-
ease Neuroimaging Initiative (ADNI, https://adni.loni.usc.edu). The clinical measures included
Mini–Mental State Examination (MMSE) scores, and Rey Auditory Verbal Learning Test (AVLT;
including AVLT1: Immediate, AVLT2: Apprentissage), Alzheimer’s Disease Assessment Scale (ADAS-
cog11 and ADAS-cog13), cerebrospinal fluid (CSF) Aβ, CSF Tau, CSF phosphorylated Tau
(p-Tau), and fluorodeoxyglucose (FDG) PET were obtained from the ADNI. Detailed informa-
tion can be found in Table 1 and Supporting Information S01. All 790 subjects have been
previously reported (Ding et al., 2021; Zhao, Zheng, Dyrba, et coll., 2022). The primary aim
of those previous studies was to verify whether radiomics features based on PET images could
serve as biomarkers for AD and whether the radiomics similarity network based on sMRI
could be applied to AD and MCI subtypes. Ici, the current study introduced a R2SN to
map structural MRI and Aβ PET imaging in order to study their cross-modal interregional
coupling.
For each participant, the T1 MRI image was aligned with Montreal Neurological Institute
(MNI) space using Advanced Normalization Tools (ANTs) after N4 bias field correction (https://
github.com/ANTsX/ANTsPy). The preliminary preprocessing of the Aβ PET image was per-
formed by the ADNI group (https://adni.loni.usc.edu/methods/pet-analysis-method/pet
-analysis/#pet-pre-processing-container). The Aβ PET image was also registered to MNI stan-
dard space using the ANTs toolkit.
R2SN Construction and Cross-Modal Coupling
The entire experimental process is shown in Figure 1A. For each image, a series of radiomics
features (N = 47) were extracted in each brain region (total of 246) defined by the Brainnetome
Atlas (Fan et al., 2016). The definitions and detailed descriptions can be found in previous
publications (Aerts et al., 2014; Ding et al., 2021; Zhao, Ding, et coll., 2020; Zhao, Zheng,
Che, et coll., 2021; Zhao, Zheng, Dyrba, et coll., 2022) and are listed in Supporting Information
S02. All features were described by Aerts and colleagues and implemented as in-house
MATLAB scripts (https://github.com/ YongLiulab/; Oui. Liu, 2022). A min-max method was first
introduced to normalize the radiomics features among different brain regions, and the redun-
dancy features were removed in further analysis, which was defined as those features that had
a high correlation with other features (R > 0.9), based on previous studies (Zhao, Zheng, Che,
Neurosciences en réseau
88
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
un
_
0
0
2
7
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Spatial coupling of AD based on sMRI and PET
Tableau 1. Detailed information on the subjects included in this study
All subjects
Subjects with an MMSE
Subjects with an FDG measurement
Subjects with an Aβ
Subjects with a Tau
Subjects with a p-Tau
Subjects with an ADAS-cog11
Groupe
Caroline du Nord (248)
MCI (390)
AD (152)
P.
Caroline du Nord (247)
MCI (388)
AD (152)
P.
Caroline du Nord (238)
MCI (383)
AD (144)
P.
Caroline du Nord (131)
MCI (278)
AD (96)
P.
Caroline du Nord (186)
MCI (324)
AD (101)
P.
Caroline du Nord (185)
MCI (324)
AD (101)
P.
Caroline du Nord (248)
MCI (389)
AD (151)
P.
Subjects with an ADAS-cog13
Caroline du Nord (246)
73.74 ± 6.05
MCI (387)
71.93 ± 7.36
AD (147)
73.72 ± 7.34
P.
0.002
Neurosciences en réseau
Age (années)
73.78 ± 6.08
71.93 ± 7.37
73.82 ± 7.37
0.002
73.72 ± 6.02
71.96 ± 7.37
73.82 ± 7.37
0.002
73.77 ± 6.12
71.85 ± 7.38
73.84 ± 7.51
0.001
73.46 ± 6.11
Sex (M/F)
125/123
217/173
89/63
0.24
125/122
217/171
89/63
0.24
120/118
214/169
83/61
0.29
65/66
Clinical measure
/
/
/
/
29.08 ± 1.19
28.03 ± 1.81
22.11 ± 3.63
<0.001
1.31 ± 0.11
1.25 ± 0.13
1.06 ± 0.15
<0.001
1,038.42 ± 386.43
71.86 ± 7.18
157/121
888.74 ± 343.83
73.79 ± 7.65
0.021
73.24 ± 6.04
55/41
0.37
93/93
646.57 ± 282.88
<0.001
243.99 ± 95.84
71.55 ± 7.28
175/149
274.52 ± 127.33
74.68 ± 7.76
0.005
73.28 ± 6.03
60/41
0.31
92/93
71.55 ± 7.28
175/149
73.66 ± 7.61
0.001
73.78 ± 6.08
71.96 ± 7.36
73.76 ± 7.36
0.001
60/41
0.29
125/123
216/173
88/63
0.26
123/123
215/172
86/61
0.21
379.12 ± 153.46
<0.001
22.59 ± 10.09
26.30 ± 14.14
37.18 ± 16.65
<0.001
5.85 ± 2.95
9.09 ± 4.30
21.61 ± 8.22
<0.001
9.22 ± 4.48
14.70 ± 6.63
32.21 ± 9.59
<0.001
89
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Spatial coupling of AD based on sMRI and PET
Subjects with an AVLT1
Subjects with an AVLT2 (N = 1,575)
Table 1.
(continued )
Group
NC (247)
MCI (389)
AD (150)
P
NC (247)
MCI (389)
AD (150)
P
Age (years)
73.75 ± 6.08
71.90 ± 7.36
73.82 ± 7.24
0.001
73.75 ± 6.08
71.90 ± 7.36
73.82 ± 7.24
0.001
Sex (M/F)
124/123
216/173
89/61
0.18
124/123
216/173
89/61
0.18
Clinical measure
45.09 ± 9.87
37.26 ± 11.15
21.37 ± 7.39
<0.001
6.01 ± 2.32
4.63 ± 2.64
1.51 ± 1.63
<0.001
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
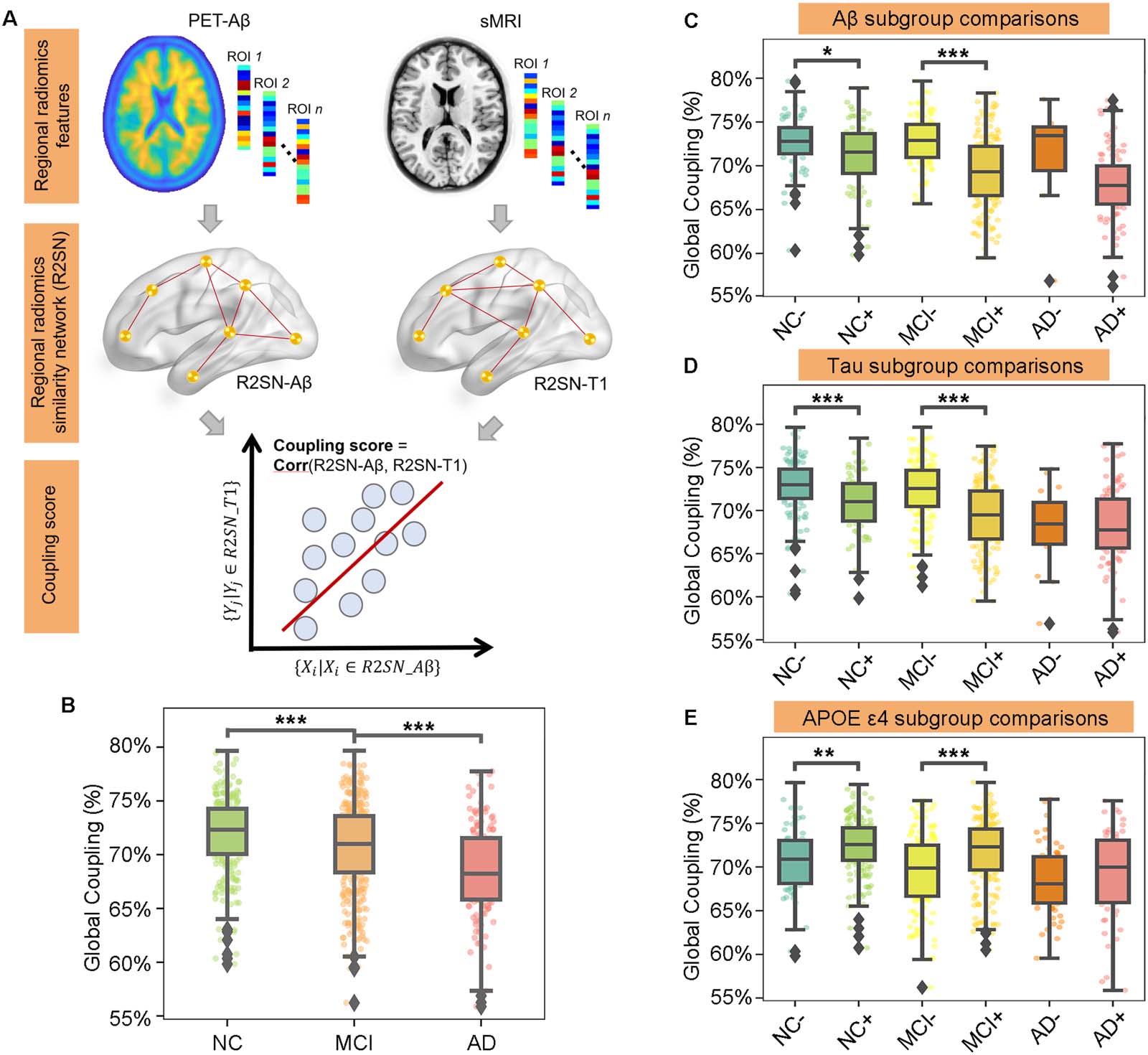
Figure 1. Global R2SN coupling patterns were discriminative for cognitively impaired states. (A) Overview of the study methodology. (B) The
global coupling scores were significantly different among the NC, MCI, and AD groups. (C–E) Further subgroup comparisons were computed
in each cognitively impaired state according to Aβ, Tau, and APOE ε4. * p < 0.05, ** p < 0.01, *** p < 0.001.
Network Neuroscience
90
Spatial coupling of AD based on sMRI and PET
et al., 2021; Zhao, Zheng, Dyrba, et al., 2022). The node was defined as the brain regions
defined by the Brainnetome Atlas, and the edge was defined as the Pearson’s correlations
between interregional radiomics features. As a result, two models of R2SN (R2SN-T1,
R2SN-Aβ) were constructed for each participant. Detailed information can be found in Sup-
porting Information S02 and S03.
To evaluate the relationship between R2SN-T1 and R2SN-Aβ, we defined the coupling of
the two networks at the global and local levels. Briefly, the global coupling score was defined
as the Pearson’s correlations between the edge of R2SN-T1 and R2SN-Aβ.
(cid:3)
Global coupling score ¼ Corr Mij jMij 2 R2SN-T 1g; Nij jNij 2 R2SN-Aβg
(cid:1)(cid:4)
(cid:1)
:
Briefly, all edges of the R2SN based on MRI can be converted into a vector with 30,315 × 1
(upper triangular matrix of 246 × 246), the same as R2SN based on PET. Here, the correlations
between two networks were converted into Pearson’s correlations between two vectors with
30,315 × 1.
The local coupling score was defined as the Pearson’s correlations between the connec-
tions of each node based on R2SN-T1 and R2SN-Aβ (Figure 1).
Local coupling scorei ¼ Corr
(cid:1)(cid:4)
(cid:3)
(cid:1)
xijjj¼1;…;246 2 R2SN-T 1g; yijjj¼1;…;246 2 R2SN-Aβg
;
where i and j represent the ith and jth brain regions, respectively.
Grouped Comparison for Cognitively Impaired States
We first tested whether the global coupling score was differentiable among the NC, MCI, and
AD groups by ANOVA with age and sex as the covariates. Then, a two-tailed, two-sample t test
was used to estimate the difference in the global coupling score in AD versus NC, MCI versus
NC, and AD versus MCI, with age and sex as the covariates. Then, further subgroup compar-
isons were computed according to apolipoprotein E (APOE) ε4, Aβ, and Tau. Aβ+ was defined
as a CSF Aβ value < 980 pg/ml, and Tau+ was defined as a CSF total Tau > 245 pg/ml, based on
https://files.alz.washington.edu/presentations/2018/spring/biomarkers/SHAW.pdf and a previ-
ous study (Zhao, Zheng, Dyrba, et coll., 2022). The local coupling scores were analyzed among
the NC, MCI, and AD groups with a two-tailed, two-sample t test with age and sex as the
covariables.
Clinical Significance of the Global Coupling Score
To investigate the potential biological basis of the global coupling score, Pearson correlations
between the global coupling score and neuropsychiatric measures (including MMSE, AVLT1,
AVLT2, ADAS-cog11, and ADAS-cog13) and peripheral biomarkers (including CSF Aβ, CSF
Tau, CSF p-Tau, and FDG) were computed with age and sex as covariates, as in previous stud-
ies (Ding et al., 2021; Zhao, Zheng, Dyrba, et coll., 2022). To further assess whether the cou-
pling level between R2SN-T1 and R2SN-Aβ can reveal the different clinical progression, le
MCI patients were subdivided into two subgroups (0%–50% with low coupling scores and
50%–100% with high coupling scores) and four subgroups (S1: 0%–25%, S2: 25%–50%,
S3: 50%–75%, and S4: 75%–100%). Survival curves for each subgroup were computed with
Kaplan–Meier analysis based on real follow-up duration information. Ici, 1 was defined as
Neurosciences en réseau
91
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
un
_
0
0
2
7
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Spatial coupling of AD based on sMRI and PET
the MCI subject developing to AD, et 0 was defined as the MCI subject not developing to
AD (Li et al., 2019).
RÉSULTATS
Demographic and Neuropsychological Characteristics
A total of 790 participants were studied, y compris 248 NCs, 390 MCI patients, et 152 AD
patients. The mean age was significantly different (P = 0.002) among the groups, and the sex
proportion was not significantly different (P = 0.24). The clinical measures (including MMSE
score, ADAS-cog11 score, ADAS-cog13 score, CSF Aβ level, CSF Tau level, CSF p-Tau level,
AVLT score, and FDG) were significantly different among the NC, MCI, and AD groups (P. <
0.001 with ANOVA; Table 1).
Global R2SN Coupling Patterns Are Discriminative for Cognitively Impaired States
The global coupling score for NCs was 0.72 ± 0.04, while the global coupling scores for MCI
and AD patients were 0.71 ± 0.04 and 0.68 ± 0.04, respectively (Figure 1B). The global cou-
pling scores were significantly different among the NC, MCI, and AD groups (P < 0.001 by
ANOVA), with scores of AD significantly lower than those of MCI (P < 0.001 by t test), scores
of AD significantly lower than those of NC (P < 0.001 by t test), and scores of MCI significantly
lower than those of NC (P < 0.001 by t test; Figure 1B).
Further subgroup comparisons were computed in each cognitively impaired state according
to Aβ, Tau, and APOE ε4. The global coupling scores were significantly decreased in the Aβ+
NC subgroup compared with the Aβ− subgroup (P = 0.015) and decreased in the Aβ+ MCI
subgroup compared with the Aβ− MCI subgroup (P < 0.001; Figure 1C). They were also sig-
nificantly decreased in the Tau+ NC subgroup compared with the Tau− NC subgroup (P <
0.001) and decreased in the Tau+ MCI subgroup compared with the Tau− MCI subgroup
(P < 0.001; Figure 1D). There was also a significant decrease in global coupling scores in
APOE ε4+ NC and MCI in contrast with APOE ε4− NC and MCI, respectively (P = 0.003
and P < 0.001; Figure 1E).
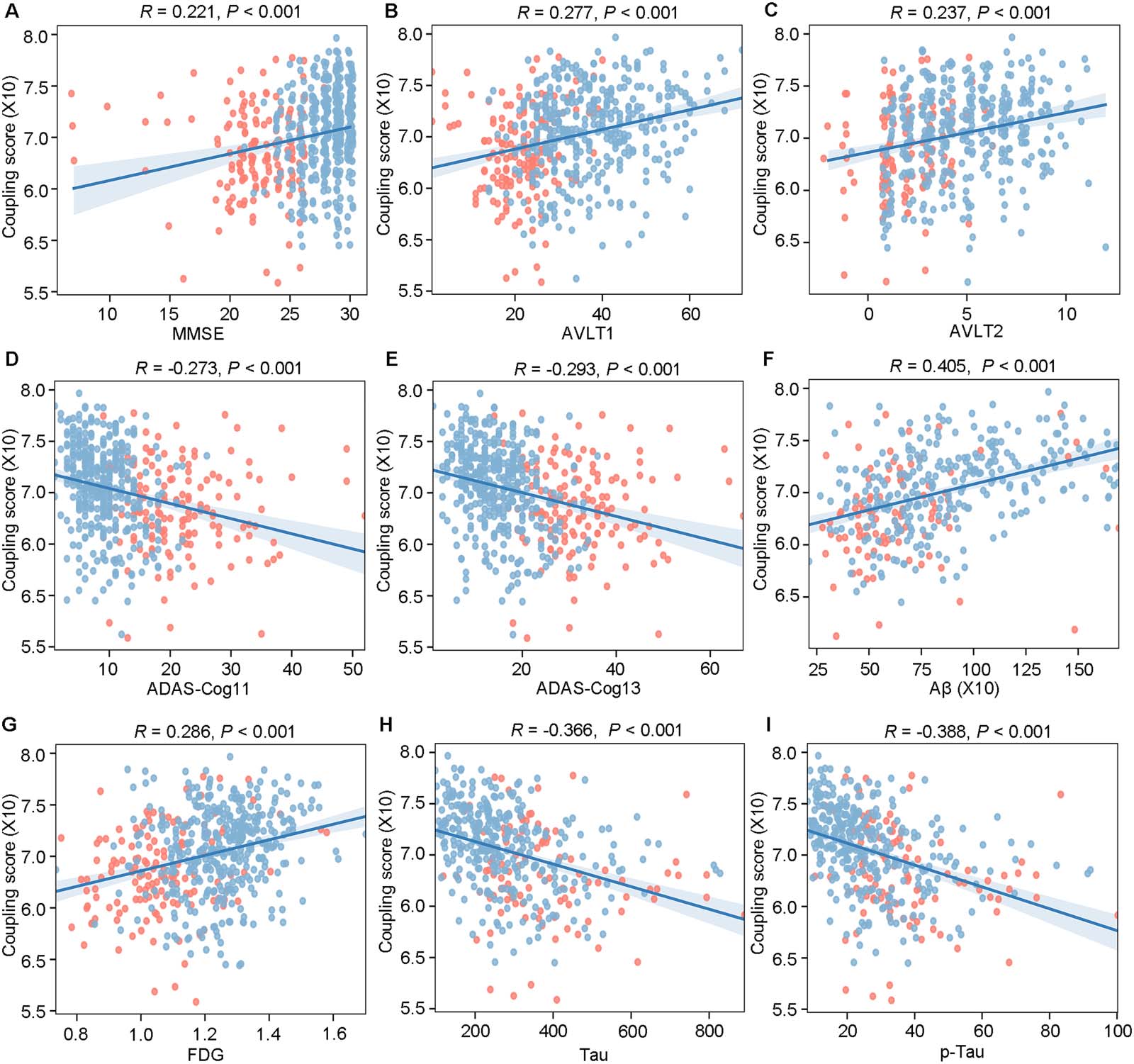
Global Coupling Scores Were Significantly Correlated With Clinical Measures
Multiple clinical measures were positively correlated with the global coupling scores,
including MMSE (R = 0.221, P < 0.001), AVLT1 (R = 0.277, P < 0.001), and AVLT2 (R =
0.237, P < 0.001; Figure 2A–C), while ADAS-cog11 (R = −0.273, P < 0.001) and ADAS-
cog13 (R = −0.293, P < 0.001) were found to be negatively correlated with the global cou-
pling scores (Figure 2D–E). Meanwhile, there were also positive correlations between the
global coupling scores and CSF Aβ (R = 0.405, P < 0.001) and FDG (R = 0.286, P <
0.001; Figure 2F–G), as well as negative correlations with Tau (R = −0.366, P < 0.001)
and p-Tau (R = −0.388, P < 0.001; Figure 2H–I), all of which were corrected by Bonferroni
correction with P < 0.05/9.
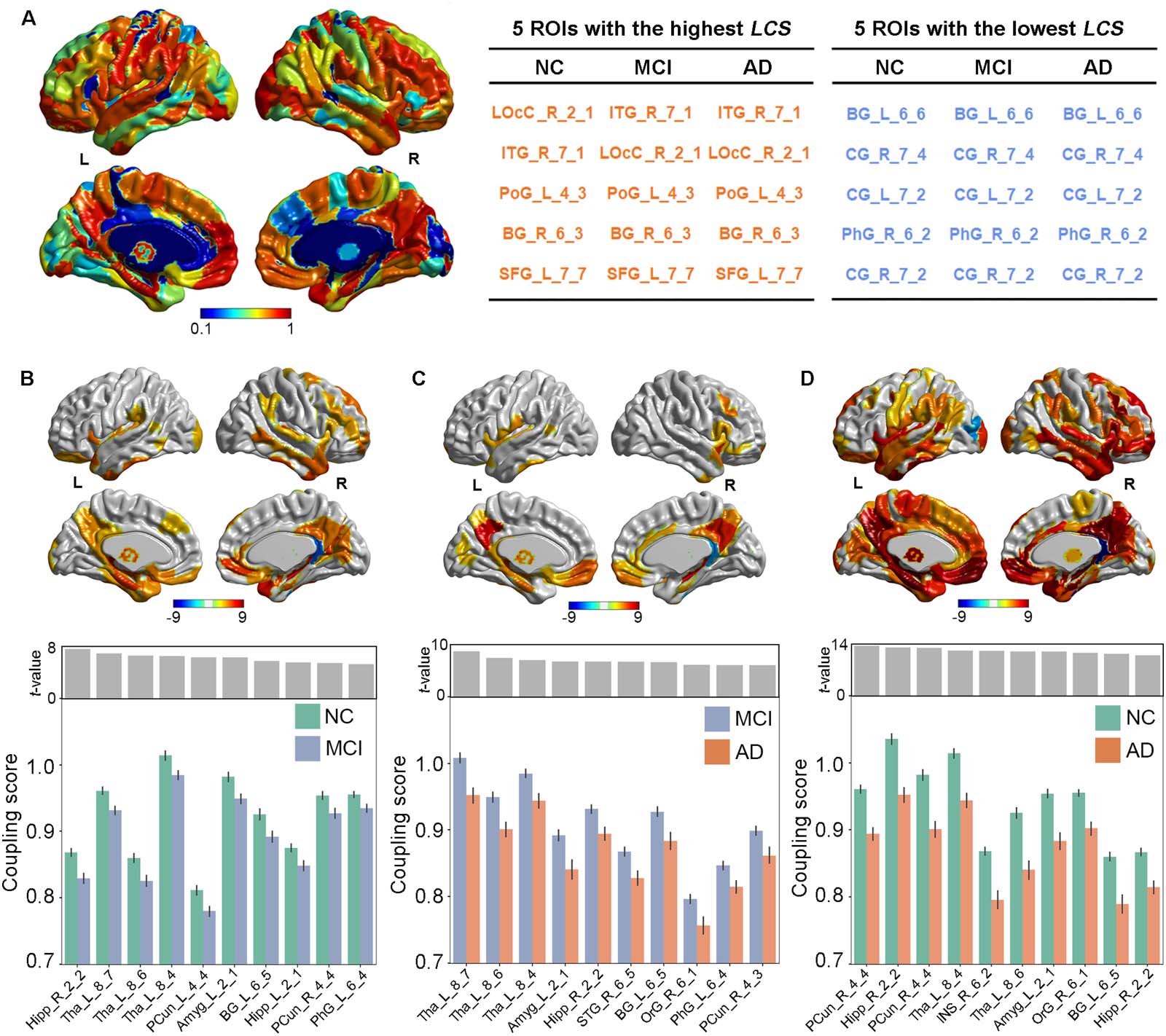
Local Coupling Scores Suggested Regional Heterogeneity Links to Aβ and Atrophy
After clarifying the global R2SN coupling patterns, we used the image datasets to analyze
regional heterogeneity. Overall, in the AD, MCI, and NC groups, the distribution of regional
R2SN coupling was basically the same, with LOcC_R_2_1, ITG_R_7_1, PoG_L_4_3,
BG_R_6_3, and SFG_L_7_7 having the highest scores and BG_L_6_6, CG_R_7_4,
Network Neuroscience
92
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Spatial coupling of AD based on sMRI and PET
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Global coupling scores were significantly correlated with clinical measures. The correlation between global coupling score and (A)
MMSE, (B) AVLT1, (C) AVLT2, (D) ADAS-Cog11, (E) ADAS-Cog13, (F) Aβ, (G) FDG, (H) Tau, and (I) p-Tau. The blue dots indicate the MCI
subjects, and the red dots indicate the AD subjects.
CG_L_7_2, PhG_R_6_2, and CG_R_7_2 having the lowest scores (Figure 3A). The detailed
names of the brain regions are shown in Table S5 in the Supporting Information.
We further compared regional R2SN coupling across cognitively impaired state groups
with age and sex as covariates, and there was a gradual decrease from NC to MCI/AD.
The local coupling scores were significantly decreased in the MCI group compared with
the NC group, including PCun_L_4_4 (t = 7.54, P < 0.001), Hipp_R_2_2 (t = 6.90, P <
0.001), PCun_R_4_4 (t = 6.58, P < 0.001), Tha_L_8_4 (t = 6.51, P < 0.001), and INS_R_6_2
(t = 6.32, P < 0.001; Figure 3B). The local coupling scores of AD were also decreased com-
pared with MCI, including Tha_L_8_7 (t = 8.66, P < 0.001), Tha_L_8_6 (t = 7.44, P <
0.001), Tha_L_8_4 (t = 7.03, P < 0.001), Amg_L_2_1 (t = 6.77, P < 0.001), and Hipp_R_2_2
(t = 6.76, P < 0.001; Figure 3C). Finally, there was a dramatic decrease in local coupling
scores in AD compared with NC, such as Hipp_R_2_2 (t = 13.39, P < 0.001), Tha_L_8_7
(t = 12.98, P < 0.001), Tha_L_8_6 (t = 12.87, P < 0.001), Tha_L_8_4 (t = 12.13, P < 0.001),
and PCun_L_4_4 (t = 12.09, P < 0.001; Figure 3D). It should be noted that all of the brain
regions shown in Figure 3 have been corrected by Bonferroni correction with P < 0.05/
Network Neuroscience
93
Spatial coupling of AD based on sMRI and PET
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Local coupling scores suggested regional heterogeneity in the link between Aβ and atrophy. (A) Overview of the general local
coupling scores across the NC, MCI, and AD groups as well as their representative brain regions. The color bar represents the strength of
local coupling scores. The discriminative regions of local R2SN coupling patterns in (B) MCI vs. NC, (C) AD vs. MCI and (D) AD vs. NC.
The color bar represents the T scores between each pair of groups. The bottom bar figures indicate the top 10 regions between each pair
of groups. The error bar indicates the standard deviation of the coupling score. LCS = local coupling scores.
246. Detailed information on these regions can be found at https://atlas.brainnetome.org
/index.html.
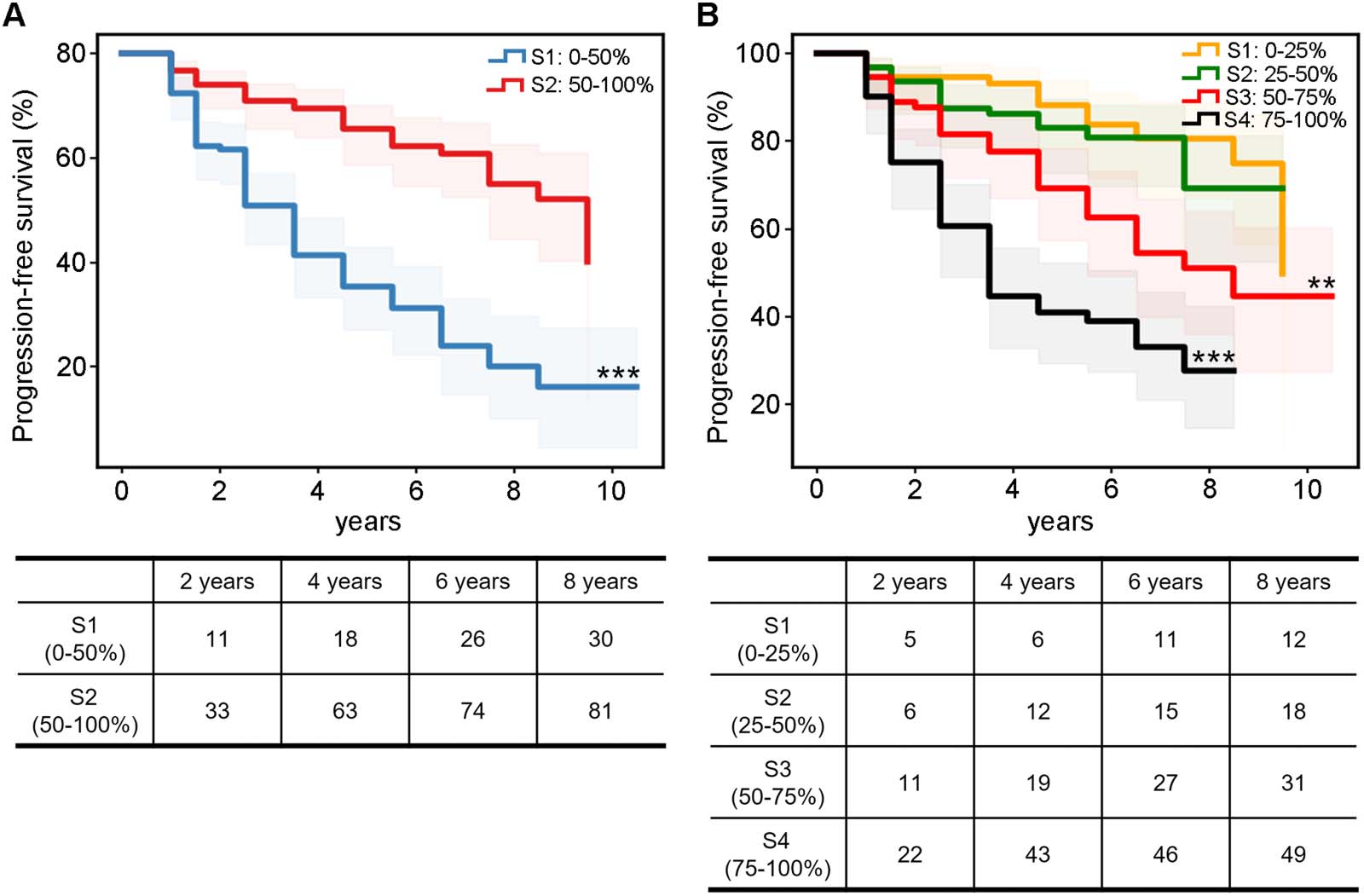
Global Coupling Scores Indicate the Distinct Progression of MCI Patients
To further assess whether the R2SN coupling level could reveal the clinical progression of
dementia, we subdivided the MCI patients into two subgroups according to the average
global coupling scores. Kaplan–Meier analysis demonstrated that the progression pattern of
the low-coupling subgroup was significantly worse than that of the high-coupling subgroup
(P < 0.001; Figure 4A). To further confirm this result, we subdivided the MCI patients into
four subgroups according to the quartiles of global coupling scores (S1: top 0%–25%, S2:
25%–50%, S3: 50%–75%, and S4: 75%–100%). Kaplan–Meier analysis showed that lower
global coupling scores could reveal worse clinical progression of dementia (PS1 vs. S2 =
0.466, PS1 vs. S3 = 0.002, PS1 vs. S4 < 0.001, PS2 vs. S3 = 0.014, PS2 vs. S4 < 0.001, PS3 vs. S4 <
0.001; Figure 4B).
Network Neuroscience
94
Spatial coupling of AD based on sMRI and PET
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Global coupling scores indicated the distinct progression of the MCI patients. The survival curve of the different coupling levels: (A)
two equal subgroupings and (B) four quartile subgroupings. ** p < 0.01, *** p < 0.001.
DISCUSSION
The initial motivation for establishing these network mappings is to use analytical tools from
graph theory, system theory, control theory, and the like to mine the underlying peculiarities
behind the original data. There are also many times when networking data can lead to unique
mechanistic annotations; for example, human MSNs recapitulate cortical cytoarchitectonic
divisions and better structural connectomes (Seidlitz et al., 2018). In the present study, we
applied R2SNs to construct a morphological covariation network by radiomics, providing a
network perspective to analyze the relationship between Aβ accumulation and altered brain
structure.
Network relationships can be investigated using methods ranging from simple approaches,
such as statistical models (Messé et al., 2014; Mišić et al., 2016), to more complex ones, such
as communication models (Goñi et al., 2014; Mišić et al., 2015) or biophysical models (Breakspear,
2017; Sanz-Leon et al., 2015). Typically, network coupling by correlational analyses is simple
but useful, and it could offer new insights into individual fingerprinting, that is, how functional
brain networks align with the underlying structural network as measured with diffusion MRI
(Honey et al., 2009) or how microstructural covariance network is correlated with region-to-
region connectivity (Huntenburg et al., 2017; Paquola et al., 2019). This statistical model
offers a data-driven way to associate structural and functional connectivity without assuming
a specific mode of interaction among neuronal populations, which has proven to be helpful in
quantifying the effects of manipulations and perturbations, such as development and aging
(Baum et al., 2020), neurological and psychiatric diseases (Jirsa et al., 2017), and lesions
(Rosenthal et al., 2018). Extended studies further explain this coupling as systematic hierarchi-
cal variation in laminar differentiation (Paquola et al., 2019) and cytoarchitecture (Vazquez-
Rodriguez et al., 2019). Similar to the causal interpretation between brain structure and
Network Neuroscience
95
Spatial coupling of AD based on sMRI and PET
function, brain atrophy was always taken as an inevitable event associated with Aβ deposition
in dementia progression (Sepulcre et al., 2018; Zhang et al., 2020). Several AD studies have
shown a temporal sequence between the spreading of Aβ aggregation species and other
neurodegeneration-based biological signatures underlying atrophy, including tauopathy, neu-
roinflammation, and neurochemical systems (Hampel et al., 2021). Although no causal effect
was established between the Aβ pathway and AD-related pathophysiological changes, it has
been suggested that the Aβ pathway exerts a permissive/facilitating effect on other pathophys-
iological pathways and/or unfolds synergistically at different temporal scales (Hampel et al.,
2021). The relationship between the R2SN-T1 and R2SN-Aβ networks could better reflect the
complex process secondary to the Aβ pathway and its direct link to atrophy.
In the analysis of local coupling scores, it was found that the spatial coupling distribution of
brain regions of both the NC and clinical groups was very similar, with the highest local cou-
pling scores in LOcC_R_2_1, ITG_R_7_1, PoG_L_4_3, BG_R_6_3, and SFG_L_7_6 and the
lowest local coupling scores in BG_L_6_6, CG_R_7_4, CG_L_7_2, PhG_R_6_2, and
CG_r_7_2. The consistent distribution of local coupling scores across different groups once
again suggested that the contribution of Aβ to the disease varied; that is, the contribution to
the morphological changes of brain tissue was higher in brain regions such as the temporal
lobe. Future biological studies of highly coupled areas may provide a more in-depth mecha-
nistic explanation.
In further analysis of global and local coupling scores, we noticed that the coupling pat-
tern constantly declines with the severity of cognitive impairment. A previous study high-
lighted that the coupling of altered gray matter volume and Aβ was correlated with subjective
cognitive decline–related worries (X. Wang et al., 2021). Even in cognitively normal elderly
individuals, there was a covarying alteration in Aβ deposition and atrophy (Oh et al., 2014).
However, the contribution of Aβ to brain atrophy gradually decreased. In the early course of
cognitively impaired states rather than normal aging, Aβ depositions acted as a more pro-
nounced accelerator to advance the onset of brain degeneration (Donohue et al., 2017; Fandos
et al., 2017; Lim et al., 2014; Sheline et al., 2010). Aβ deposition of upstream events drives
downstream events (neocortical Tau spread, impaired glucose metabolism, and widespread
neurodegeneration; Hansson, 2021). These inconsistent biomarkers also result from the differ-
ent AD pathways with distinct progression (Reimand et al., 2020). This significant stage effect
leads to the differential sensitivity of classical ATN (A [CSF Ab42, or Aβ42/Ab40 ratio and amy-
loid PET], T [CSF p-Tau, Tau PET], and N [atomic MRI, FDG PET, and CSF total Tau]) in the
diagnosis sensitivity over AD progression (Hammond et al., 2020; Sperling et al., 2011). We
supposed that the lower the coupling, the more abnormal pathophysiological mechanisms
were involved, which was also an important reason for the poor prognosis of AD. This hypoth-
esis is also reflected in the progressive disease outcome of MCI patients. The lower the global
coupling scores are, the faster the rate at which MCI patients transition to AD. Brain regions
with lower local coupling scores tended to be brain regions with more obvious atrophy
changes in previous reports (Grothe et al., 2018). All these results indicate that the progression
of AD is the result of the simultaneous effect of multiple biological mechanisms, and the assess-
ment of the contribution of different biomarkers may have clinical value in predicting disease
prognosis.
This study has some limitations. First, the sMRI and PET scans for some subjects were not
obtained at the same time (e.g., 3–6 months). Second, we explored the coupling score of the
R2SN between sMRI and PET in only the ADNI dataset. Another dataset to verify the robust-
ness should be analyzed in future studies. Third, the altered coupling pattern among different
imaging modalities would benefit our understanding of AD pathophysiology in the future.
Network Neuroscience
96
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Spatial coupling of AD based on sMRI and PET
Conclusion
In this study, we systematically demonstrated that the alteration of the coupling between brain
networks of brain structure and Aβ accumulation was related to the solid biological/clinical
basis, which could serve as a predictor for revealing the distinct progression of AD. This study
provided a new measure for exploring the coupling between Aβ and atrophy.
ACKNOWLEDGMENTS
Data collection and sharing for this project were funded by the Alzheimer’s Disease Neuro-
imaging Initiative (ADNI) (National Institutes of Health Grant No. U01 AG024904) and DOD
ADNI (Department of Defense Award No. W81XWH-12-2-0012). The ADNI was funded by
the National Institute on Aging, the National Institute of Biomedical Imaging and Bioengineer-
ing, and generous contributions from AbbVie; Alzheimer’s Association; Alzheimer’s Drug Dis-
covery Foundation; Araclon Biotech; BioClinica, Inc.; Biogen; Bristol-Myers Squibb Company;
CereSpir, Inc.; Cogstate; Eisai Inc.; Elan Pharmaceuticals, Inc.; Eli Lilly and Company; Euro-
Immun; F. Hoffmann-La Roche Ltd., and its affiliated company Genentech, Inc.; Fujirebio;
GE Healthcare; IXICO, Ltd.; Janssen Alzheimer Immunotherapy Research & Development,
LLC.; Johnson & Johnson Pharmaceutical Research & Development, LLC; Lumosity; Lundbeck;
Merck & Co., Inc.; Meso Scale Diagnostics, LLC.; NeuroRx Research; Neurotrack Technolo-
gies; Novartis Pharmaceuticals Corporation; Pfizer, Inc.; Piramal Imaging; Servier; Takeda
Pharmaceutical Company; and Transition Therapeutics. The Canadian Institutes of Health
Research provided funds to support ADNI clinical sites in Canada. Private sector contributions
were facilitated by the Foundation for the National Institutes of Health (https://www.fnih.org).
The grantee organization was the Northern California Institute for Research and Education,
and the study was coordinated by the Alzheimer’s Therapeutic Research Institute at the
University of Southern California. ADNI data were disseminated by the Laboratory of Neuro
Imaging at the University of Southern California. As data used in preparation of this article
were obtained from the ADNI database (https://adni.loni.usc.edu), the investigators within
the ADNI contributed to the design and implementation of the ADNI and/or provided data
but did not participate in the analysis or writing of this report. A complete listing of ADNI
investigators can be found at https://adni.loni.usc.edu/wp-content/uploads/ how_to_apply
/ADNI_Acknowledgement_List.pdf.
DATA AVAILABILITY
All subjects of this study were downloaded from the ADNI (https://adni.loni.usc.edu), and the
scripts of the radiomics features (Zhao, Ding, et al., 2020) are available at https://github.com
/ YongLiulab (Y. Liu, 2022).
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00271.
AUTHOR CONTRIBUTIONS
Kun Zhao: Conceptualization; Data curation; Formal analysis; Investigation; Methodology;
Writing – original draft. Jiaji Lin: Conceptualization; Investigation; Writing – original draft.
Martin Dyrba: Writing – review & editing. Dong Wang: Data curation. Tongtong Che: Writing –
review & editing. Haoyang Wu: Software. Jingyu Wang: Software. Yong Liu: Conceptualization;
Network Neuroscience
97
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Spatial coupling of AD based on sMRI and PET
Supervision; Validation; Visualization; Writing – review & editing. Shuyu Li: Conceptualization;
Supervision; Validation; Writing – review & editing.
FUNDING INFORMATION
Shuyu Li, Startup Funds for Leading Talents at Beijing Normal University. Shuyu Li, National
Natural Science Foundation of China, Award ID: 81972160. Yong Liu, Fundamental Research
Funds for the Central Universities, Award ID: 2021XD-A03-1. Yong Liu, Beijing Natural Sci-
ence Funds for Distinguished Young Scholars, Award ID: JQ20036. Yong Liu, National Natural
Science Foundation of China, Award ID: 81871438. Yong Liu, National Natural Science Foun-
dation of China, Award ID: 82172018.
REFERENCES
Aerts, H. J., Velazquez, E. R., Leijenaar, R. T., Parmar, C.,
Grossmann, P., Carvalho, S., Bussink, J., Monshouwer, R.,
Haibe-Kains, B., Rietveld, D., Hoebers, F., Rietbergen, M. M.,
Leemans, C. R., Dekker, A., Quackenbush, J., Gillies, R. J., &
Lambin, P. (2014). Decoding tumour phenotype by noninvasive
imaging using a quantitative radiomics approach. Nature
C o m m u n i c a t i o n s , 5 , 4 0 0 6 . h t t p s : / / d o i . o r g / 1 0 . 1 0 3 8
/ncomms5006, PubMed: 24892406
Alexander-Bloch, A., Giedd, J. N., & Bullmore, E. (2013). Imaging
structural co-variance between human brain regions. Nature
Reviews Neuroscience, 14(5), 322–336. https://doi.org/10.1038
/nrn3465, PubMed: 23531697
Baum, G. L., Cui, Z., Roalf, D. R., Ciric, R., Betzel, R. F., Larsen, B.,
Cieslak, M., Cook, P. A., Xia, C. H., Moore, T. M., Ruparel, K.,
Oathes, D. J., Alexander-Bloch, A. F., Shinohara, R. T.,
Raznahan, A., Gur, R. E., Gur, R. C., Bassett, D. S., &
Satterthwaite, T. D. (2020). Development of structure-function
coupling in human brain networks during youth. Proceedings
of the National Academy of Sciences, 117(1), 771–778. https://
doi.org/10.1073/pnas.1912034117, PubMed: 31874926
Breakspear, M. (2017). Dynamic models of large-scale brain activ-
ity. Nature Neuroscience, 20(3), 340–352. https://doi.org/10
.1038/nn.4497, PubMed: 28230845
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph
theoretical analysis of structural and functional systems. Nature
Reviews Neuroscience, 10(3), 186–198. https://doi.org/10.1038
/nrn2575, PubMed: 19190637
Bullmore, E., & Sporns, O. (2012). The economy of brain network
organization. Nature Reviews Neuroscience, 13(5), 336–349.
https://doi.org/10.1038/nrn3214, PubMed: 22498897
Chen, P., Yao, H., Tijms, B. M., Wang, P., Wang, D., Song, C., Yang,
H., Zhang, Z., Zhao, K., Qu, Y., Kang, X., Du, K., Fan, L., Han, T.,
Yu, C., Zhang, X., Jiang, T., Zhou, Y., Lu, J., … Liu, Y. (2022). Four
distinct subtypes of Alzheimer’s disease based on resting-state
connectivity biomarkers. Biological Psychiatry. https://doi.org
/10.1016/j.biopsych.2022.06.019, PubMed: 36137824
Chen, Z. J., He, Y., Rosa-Neto, P., Germann, J., & Evans, A. C.
(2008). Revealing modular architecture of human brain structural
networks by using cortical thickness from MRI. Cerebral Cortex,
18(10), 2374–2381. https://doi.org/10.1093/cercor/ bhn003,
PubMed: 18267952
Cook, G. J. R., Azad, G., Owczarczyk, K., Siddique, M., & Goh, V.
(2018). Challenges and promises of PET radiomics. International
Journal of Radiation Oncology, Biology, Physics, 102(4),
1083–1089. https://doi.org/10.1016/j.ijrobp.2017.12.268,
PubMed: 29395627
Ding, Y., Zhao, K., Che, T., Du, K., Sun, H., Liu, S., Zheng, Y., Li, S.,
Liu, B., Liu, Y., & Alzheimer’s Disease Neuroimaging Initiative.
(2021). Quantitative radiomic features as new biomarkers for
Alzheimer’s disease: An amyloid PET study. Cerebral Cortex,
31(8), 3950–3961. https://doi.org/10.1093/cercor/ bhab061,
PubMed: 33884402
Donohue, M. C., Sperling, R. A., Petersen, R., Sun, C. K., Weiner,
M. W., Aisen, P. S., & Alzheimer’s Disease Neuroimaging Initia-
tive. (2017). Association between elevated brain amyloid and
subsequent cognitive decline among cognitively normal persons.
JAMA, 317(22), 2305–2316. https://doi.org/10.1001/jama.2017
.6669, PubMed: 28609533
Dyrba, M., Mohammadi, R., Grothe, M. J., Kirste, T., & Teipel, S. J.
(2020). Gaussian graphical models reveal inter-modal and
inter-regional conditional dependencies of brain alterations in
Alzheimer’s disease. Frontiers in Aging Neuroscience, 12, 99.
https://doi.org/10.3389/fnagi.2020.00099, PubMed: 32372944
Fan, L., Li, H., Zhuo, J., Zhang, Y., Wang, J., Chen, L., Yang, Z., Chu,
C., Xie, S., Laird, A. R., Fox, P. T., Eickhoff, S. B., Yu, C., & Jiang,
T. (2016). The Human Brainnetome Atlas: A new brain atlas
based on connectional architecture. Cerebral Cortex, 26(8),
3508–3526. https://doi.org/10.1093/cercor/ bhw157, PubMed:
27230218
Fandos, N., Perez-Grijalba, V., Pesini, P., Olmos, S., Bossa, M.,
Villemagne, V. L., Doecke, J., Fowler, C., Masters, C. L., Sarasa,
M., & AIBL Research Group. (2017). Plasma amyloid beta 42/40
ratios as biomarkers for amyloid beta cerebral deposition in
cognitively normal individuals. Alzheimer’s and Dementia:
Diagnosis, Assessment and Disease Monitoring, 8, 179–187.
https://doi.org/10.1016/j.dadm.2017.07.004, PubMed:
28948206
Goñi, J., van den Heuvel, M. P., Avena-Koenigsberger, A., Velez de
Mendizabal, N., Betzel, R. F., Griffa, A., Hagmann, P., Corominas-
Murtra, B., Thiran, J. P., & Sporns, O. (2014). Resting-brain func-
tional connectivity predicted by analytic measures of network
communication. Proceedings of the National Academy of
Network Neuroscience
98
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Spatial coupling of AD based on sMRI and PET
Sciences, 111(2), 833–838. https://doi.org/10.1073/pnas
.1315529111, PubMed: 24379387
Grothe, M. J., Sepulcre, J., Gonzalez-Escamilla, G., Jelistratova, I.,
Schöll, M., Hansson, O., & Teipel, S. J. (2018). Molecular prop-
erties underlying regional vulnerability to Alzheimer’s disease
pathology. Brain, 141(9), 2755–2771. https://doi.org/10.1093
/brain/awy189, PubMed: 30016411
Hammond, T. C., Xing, X., Wang, C., Ma, D., Nho, K., Crane,
P. K., Elahi, F., Ziegler, D. A., Liang, G., Cheng, Q., Yanckello,
L. M., Jacobs, N., & Lin, A. L. (2020). Beta-amyloid and tau
drive early Alzheimer’s disease decline while glucose hypome-
tabolism drives late decline. Communications Biology, 3(1),
352. https://doi.org/10.1038/s42003-020-1079-x, PubMed:
32632135
Hampel, H., Hardy, J., Blennow, K., Chen, C., Perry, G., Kim, S. H.,
Villemagne, V. L., Aisen, P., Vendruscolo, M., Iwatsubo, T.,
Masters, C. L., Cho, M., Lannfelt, L., Cummings, J. L., & Vergallo,
A. (2021). The amyloid-beta pathway in Alzheimer’s disease.
Molecular Psychiatry, 26(10), 5481–5503. https://doi.org/10
.1038/s41380-021-01249-0, PubMed: 34456336
Hansson, O. (2021). Biomarkers for neurodegenerative diseases.
Nature Medicine, 27(6), 954–963. https://doi.org/10.1038
/s41591-021-01382-x, PubMed: 34083813
Hardy, J. A., & Higgins, G. A. (1992). Alzheimer’s disease: The
amyloid cascade hypothesis. Science, 256(5054), 184–185.
https://doi.org/10.1126/science.1566067, PubMed: 1566067
Honey, C. J., Sporns, O., Cammoun, L., Gigandet, X., Thiran, J. P.,
Meuli, R., & Hagmann, P. (2009). Predicting human resting-state
functional connectivity from structural connectivity. Proceedings
of the National Academy of Sciences, 106(6), 2035–2040. https://
doi.org/10.1073/pnas.0811168106, PubMed: 19188601
Huntenburg, J. M., Bazin, P. L., Goulas, A., Tardif, C. L., Villringer,
A., & Margulies, D. S. (2017). A systematic relationship between
functional connectivity and intracortical myelin in the human
cerebral cortex. Cerebral Cortex, 27(2), 981–997. https://doi.org
/10.1093/cercor/bhx030, PubMed: 28184415
Jirsa, V. K., Proix, T., Perdikis, D., Woodman, M. M., Wang, H.,
Gonzalez-Martinez, J., Bernard, C., Bénar, C., Guye, M.,
Chauvel, P., & Bartolomei, F. (2017). The virtual epileptic patient:
Individualized whole-brain models of epilepsy spread. Neuro-
Image, 145(Pt. B), 377–388. https://doi.org/10.1016/j
.neuroimage.2016.04.049, PubMed: 27477535
Li, H., Habes, M., Wolk, D. A., & Fan, Y., for the Alzheimer’s Dis-
ease Neuroimaging Initiative and the Australian Imaging Bio-
markers and Lifestyle Study of Aging. (2019). A deep learning
model for early prediction of Alzheimer’s disease dementia
based on hippocampal magnetic resonance imaging data.
Alzheimer’s and Dementia, 15(8), 1059–1070. https://doi.org
/10.1016/j.jalz.2019.02.007, PubMed: 31201098
Lim, H. K., Nebes, R., Snitz, B., Cohen, A., Mathis, C., Price, J.,
Weissfeld, L., Klunk, W., & Aizenstein, H. J. (2014). Regional
amyloid burden and intrinsic connectivity networks in cogni-
tively normal elderly subjects. Brain, 137(Pt. 12), 3327–3338.
https://doi.org/10.1093/brain/awu271, PubMed: 25266592
Lista, S., Garaci, F. G., Ewers, M., Teipel, S., Zetterberg, H.,
Blennow, K., & Hampel, H. (2014). CSF Abeta1-42 combined
with neuroimaging biomarkers in the early detection, diagnosis
and prediction of Alzheimer’s disease. Alzheimer’s and Demen-
tia, 10(3), 381–392. https://doi.org/10.1016/j.jalz.2013.04.506,
PubMed: 23850330
Liu, Y. (2022). Yong Liu Lab, GitHub. https://github.com
/ YongLiulab
Liu, Z., Palaniyappan, L., Wu, X., Zhang, K., Du, J., Zhao, Q., Xie,
C., Tang, Y., Su, W., Wei, Y., Xue, K., Han, S., Tsai, S. J., Lin, C. P.,
Cheng, J., Li, C., Wang, J., Sahakian, B. J., Robbins, T. W., …
Feng, J. (2021). Resolving heterogeneity in schizophrenia
through a novel systems approach to brain structure: Individual-
ized structural covariance network analysis. Molecular Psychia-
try, 26, 7719–7731. https://doi.org/10.1038/s41380-021-01229
-4, PubMed: 34316005
Luppi, A. I., & Stamatakis, E. A. (2021). Combining network topol-
ogy and information theory to construct representative brain net-
works. Network Neuroscience, 5(1), 96–124. https://doi.org/10
.1162/netn_a_00170, PubMed: 33688608
Messé, A., Rudrauf, D., Benali, H., & Marrelec, G. (2014). Relating
structure and function in the human brain: Relative contributions
of anatomy, stationary dynamics, and non-stationarities. PLoS
Computational Biology, 10(3), e1003530. https://doi.org/10
.1371/journal.pcbi.1003530, PubMed: 24651524
Mišić, B., Betzel, R. F., de Reus, M. A., van den Heuvel, M. P.,
Berman, M. G., McIntosh, A. R., & Sporns, O. (2016). Net-
work-level structure-function relationships in human neocortex.
Cerebral Cortex, 26(7), 3285–3296. https://doi.org/10.1093
/cercor/bhw089, PubMed: 27102654
Mišić, B., Betzel, R. F., Nematzadeh, A., Goñi, J., Griffa, A.,
Hagmann, P., Flammini, A., Ahn, Y. Y., & Sporns, O. (2015).
Cooperative and competitive spreading dynamics on the human
connectome. Neuron, 86(6), 1518–1529. https://doi.org/10.1016
/j.neuron.2015.05.035, PubMed: 26087168
Oh, H., Habeck, C., Madison, C., & Jagust, W. (2014). Covarying
alterations in Aβ deposition, glucose metabolism, and gray mat-
ter volume in cognitively normal elderly. Human Brain Mapping,
35(1), 297–308. https://doi.org/10.1002/hbm.22173, PubMed:
22965806
Paquola, C., Vos De Wael, R., Wagstyl, K., Bethlehem, R. A. I.,
Hong, S. J., Seidlitz, J., Bullmore, E. T., Evans, A. C., Mišić, B.,
Margulies, D. S., Smallwood, J., & Bernhardt, B. C. (2019).
Microstructural and functional gradients are increasingly dissoci-
ated in transmodal cortices. PLoS Biology, 17(5), e3000284.
https://doi.org/10.1371/journal.pbio.3000284, PubMed:
31107870
Rathore, S., Habes, M., Iftikhar, M. A., Shacklett, A., & Davatzikos,
C. (2017). A review on neuroimaging-based classification studies
and associated feature extraction methods for Alzheimer’s dis-
ease and its prodromal stages. NeuroImage, 155, 530–548.
https://doi.org/10.1016/j.neuroimage.2017.03.057, PubMed:
28414186
Reimand, J., Collij, L., Scheltens, P., Bouwman, F., Ossenkoppele,
R., & Alzheimer’s Disease Neuroimaging Initiative. (2020). Asso-
ciation of amyloid-beta CSF/ PET discordance and tau load 5
years later. Neurology, 95(19), e2648–e2657. https://doi.org/10
.1212/ WNL.0000000000010739, PubMed: 32913020
Robinson, J. L., Lee, E. B., Xie, S. X., Rennert, L., Suh, E.,
Bredenberg, C., Caswell, C., Van Deerlin, V. M., Yan, N., Yousef,
Network Neuroscience
99
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Spatial coupling of AD based on sMRI and PET
A., Hurtig, H. I., Siderowf, A., Grossman, M., McMillan, C. T.,
Miller, B., Duda, J. E., Irwin, D. J., Wolk, D., Elman, L., …
Trojanowski, J. Q. (2018). Neurodegenerative disease concomi-
tant proteinopathies are prevalent, age-related and APOE4-
associated. Brain, 141(7), 2181–2193. https://doi.org/10.1093
/brain/awy146, PubMed: 29878075
Rosenthal, G., Vasa, F., Griffa, A., Hagmann, P., Amico, E., Goñi, J.,
Avidan, G., & Sporns, O. (2018). Mapping higher-order relations
between brain structure and function with embedded vector rep-
resentations of connectomes. Nature Communications, 9(1),
2178. https://doi.org/10.1038/s41467-018-04614-w, PubMed:
29872218
Sanz-Leon, P., Knock, S. A., Spiegler, A., & Jirsa, V. K. (2015). Math-
ematical framework for large-scale brain network modeling in
The Virtual Brain. NeuroImage, 111, 385–430. https://doi.org/10
.1016/j.neuroimage.2015.01.002, PubMed: 25592995
Seidlitz, J., Váša, F., Shinn, M., Romero-Garcia, R., Whitaker, K. J.,
Vértes, P. E., Wagstyl, K., Kirkpatrick Reardon, P., Clasen, L., Liu,
S., Messinger, A., Leopold, D. A., Fonagy, P., Dolan, R. J., Jones,
P. B., Goodyer, I. M., Raznahan, A., & Bullmore, E. T. (2018).
Morphometric similarity networks detect microscale cortical
organization and predict inter-individual cognitive variation.
Neuron, 97(1), 231–247. https://doi.org/10.1016/j.neuron.2017
.11.039, PubMed: 29276055
Sepulcre, J., Grothe, M. J., d’Oleire Uquillas, F., Ortiz-Teran, L.,
Diez, I., Yang, H. S., Jacobs, H. I. L., Hanseeuw, B. J., Li, Q.,
El-Fakhri, G., Sperling, R. A., & Johnson, K. A. (2018). Neuroge-
netic contributions to amyloid beta and tau spreading in the
human cortex. Nature Medicine, 24(12), 1910–1918. https://doi
.org/10.1038/s41591-018-0206-4, PubMed: 30374196
Sheline, Y. I., Raichle, M. E., Snyder, A. Z., Morris, J. C., Head, D.,
Wang, S., & Mintun, M. A. (2010). Amyloid plaques disrupt rest-
ing state default mode network connectivity in cognitively nor-
mal elderly. Biological Psychiatry, 67(6), 584–587. https://doi
.org/10.1016/j.biopsych.2009.08.024, PubMed: 19833321
Sperling, R. A., Aisen, P. S., Beckett, L. A., Bennett, D. A., Craft, S.,
Fagan, A. M., Iwatsubo, T., Jack, C. R., Jr., Kaye, J., Montine, T. J.,
Park, D. C., Reiman, E. M., Rowe, C. C., Siemers, E., Stern, Y.,
Yaffe, K., Carrillo, M. C., Thies, B., Morrison-Bogorad, M., …
Phelps, C. H. (2011). Toward defining the preclinical stages of
Alzheimer’s disease: Recommendations from the National Insti-
tute on Aging–Alzheimer’s Association workgroups on diagnostic
guidelines for Alzheimer’s disease. Alzheimer’s and Dementia,
7(3), 280–292. https://doi.org/10.1016/j.jalz.2011.03.003,
PubMed: 21514248
Terry, R. D., Masliah, E., Salmon, D. P., Butters, N., DeTeresa, R.,
Hill, R., Hansen, L. A., & Katzman, R. (1991). Physical basis of
cognitive alterations in Alzheimer’s disease: Synapse loss is the
major correlate of cognitive impairment. Annals of Neurology,
30(4), 572–580. https://doi.org/10.1002/ana.410300410,
PubMed: 1789684
Tijms, B. M., Series, P., Willshaw, D. J., & Lawrie, S. M. (2012).
Similarity-based extraction of individual networks from gray mat-
ter MRI scans. Cerebral Cortex, 22(7), 1530–1541. https://doi.org
/10.1093/cercor/bhr221, PubMed: 21878484
van der Kant, R., Goldstein, L. S. B., & Ossenkoppele, R. (2020).
Amyloid-beta-independent regulators of tau pathology in
Alzheimer disease. Nature Reviews Neuroscience, 21(1),
21–35. https://doi.org/10.1038/s41583-019-0240-3, PubMed:
31780819
Vazquez-Rodriguez, B., Suarez, L. E., Markello, R. D., Shafiei, G.,
Paquola, C., Hagmann, P., van den Heuvel, M. P., Bernhardt,
B. C., Spreng, R. N., & Mišić, B. (2019). Gradients of
structure-function tethering across neocortex. Proceedings of
the National Academy of Sciences, 116(42), 21219–21227.
https://doi.org/10.1073/pnas.1903403116, PubMed: 31570622
Wang, H., Kulas, J. A., Wang, C., Holtzman, D. M., Ferris, H. A., &
Hansen, S. B. (2021). Regulation of beta-amyloid production in
neurons by astrocyte-derived cholesterol. Proceedings of the
National Academy of Sciences, 118(33), e2102191118. https://
doi.org/10.1073/pnas.2102191118, PubMed: 34385305
Wang, W. Y., Yu, J. T., Liu, Y., Yin, R. H., Wang, H. F., Wang, J., Tan,
L., Radua, J., & Tan, L. (2015). Voxel-based meta-analysis of grey
matter changes in Alzheimer’s disease. Translational Neurode-
generation, 4, 6. https://doi.org/10.1186/s40035-015-0027-z,
PubMed: 25834730
Wang, X., Wang, M., Wang, X., Zhou, F., Jiang, J., Liu, H., & Han, Y.
(2021). Subjective cognitive decline–related worries modulate
the relationship between global amyloid load and gray matter
volume in preclinical Alzheimer’s disease. Brain Imaging and
Behavior, 16, 1088–1097. https://doi.org/10.1007/s11682-021
-00558-w, PubMed: 34743296
Yang, J., Pan, P., Song, W., Huang, R., Li, J., Chen, K., Gong, Q.,
Zhong, J., Shi, H., & Shang, H. (2012). Voxelwise meta-analysis
of gray matter anomalies in Alzheimer’s disease and mild cogni-
tive impairment using anatomic likelihood estimation. Journal of
the Neurological Sciences, 316(1–2), 21–29. https://doi.org/10
.1016/j.jns.2012.02.010, PubMed: 22385679
Young, A. L., Marinescu, R. V., Oxtoby, N. P., Bocchetta, M., Yong,
K., Firth, N. C., Cash, D. M., Thomas, D. L., Dick, K. M., Cardoso,
J., van Swieten, J., Borroni, B., Galimberti, D., Masellis, M.,
Tartaglia, M. C., Rowe, J. B., Graff, C., Tagliavini, F., Frisoni,
G. B., … Alzheimer’s Disease Neuroimaging Initiative. (2018).
Uncovering the heterogeneity and temporal complexity of
neurodegenerative diseases with Subtype and Stage Inference.
Nature Communications, 9(1), 4273. https://doi.org/10.1038
/s41467-018-05892-0, PubMed: 30323170
Zhang, L., Mak, E., Reilhac, A., Shim, H. Y., Ng, K. K., Ong,
M. Q. W., Ji, F., Chong, E. J. Y., Xu, X., Wong, Z. X., Stephenson,
M. C., Venketasubramanian, N., Tan, B. Y., O’Brien, J. T., Zhou,
J. H., Chen, C. L. H., & Alzheimer’s Disease Neuroimaging Ini-
tiative. (2020). Longitudinal trajectory of Amyloid-related hippo-
campal subfield atrophy in nondemented elderly. Human Brain
Mapping, 41(8), 2037–2047. https://doi.org/10.1002/ hbm
.24928, PubMed: 31944479
Zhao, K., Ding, Y., Han, Y., Fan, Y., Alexander-Bloch, A. F., Han, T.,
Jin, D., Liu, B., Lu, J., Song, C., Wang, P., Wang, D., Wang, Q.,
Xu, K., Yang, H., Yao, H., Zheng, Y., Yu, C., Zhou, B., … Liu, Y.
(2020). Independent and reproducible hippocampal radiomic
biomarkers for multisite Alzheimer’s disease: Diagnosis, longitu-
dinal progress and biological basis. Science Bulletin, 65(13),
1103–1113. https://doi.org/10.1016/j.scib.2020.04.003
Zhao, K., Zheng, Q., Che, T., Dyrba, M., Li, Q., Ding, Y., Zheng, Y., Liu,
Y., & Li, S. (2021). Regional radiomics similarity networks (R2SNs)
Network Neuroscience
100
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Spatial coupling of AD based on sMRI and PET
in the human brain: Reproducibility, small-world properties and a
biological basis. Network Neuroscience, 5(3), 783–797. https://doi
.org/10.1162/netn_a_00200, PubMed: 34746627
Zhao, K., Zheng, Q., Dyrba, M., Rittman, T., Li, A., Che, T., Chen,
P., Sun, Y., Kang, X., Li, Q., Liu, B., Liu, Y., Li, S., & Alzheimer’s
Disease Neuroimaging Initiative. (2022). Regional radiomics
similarity networks reveal distinct subtypes and abnormality pat-
terns in mild cognitive impairment. Advanced Science, 9(12),
e2104538. https://doi.org/10.1002/advs.202104538, PubMed:
35098696
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
1
8
6
2
0
7
4
5
1
7
n
e
n
_
a
_
0
0
2
7
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
101