RECHERCHE
Hypothalamic effective connectivity at rest is
associated with body weight and
energy homeostasis
Katharina Voigt1, Zane B. Andrews2, Ian H. Harding3, Adeel Razi1,4, and Antonio Verdejo-García1
1School of Psychological Sciences and Turner Institute for Brain and Mental Health, Monash University, Victoria, Australia
2Biomedicine Discovery Institute and Department of Physiology, Monash University, Victoria, Australia
3Department of Neuroscience, Central Clinical School, Monash University, Melbourne, Australia
4The Wellcome Centre for Human Neuroimaging, University College London, Londres, ROYAUME-UNI
un accès ouvert
journal
Mots clés: Resting-state fMRI, Spectral dynamic causal modelling, Effective connectivity, Énergie
homeostasis, Obesity
ABSTRAIT
Hunger and satiety drive eating behaviours via changes in brain function. The hypothalamus is
a central component of the brain networks that regulate food intake. Animal research parsed
the roles of the lateral hypothalamus (LH) and medial hypothalamus (MH) in hunger and
satiety, respectivement. Ici, we examined how hunger and satiety change information flow
between human LH and MH brain networks, and how these interactions are influenced by
body mass index (BMI). Forty participants (16 overweight/obese) underwent two resting-state
functional MRI scans while being fasted and sated. The excitatory/inhibitory influence of
information flow between the MH and LH was modelled using spectral dynamic causal
modelling. Our results revealed two core networks interacting across homeostatic state and
weight: subcortical bidirectional connections between the LH, MH and the substantia nigra
pars compacta (prSN), and cortical top-down inhibition from fronto-parietal and temporal
domaines. During fasting, we found higher inhibition between the LH and prSN, whereas the prSN
received greater top-down inhibition from across the cortex. Individuals with higher BMI
showed that these network dynamics occur irrespective of homeostatic state. Our findings
reveal fasting affects brain dynamics over a distributed hypothalamic-midbrain-cortical
réseau. This network is less sensitive to state-related fluctuations among people with obesity.
RÉSUMÉ DE L'AUTEUR
The hypothalamus is a central component of the brain networks regulating food intake. Animal
research subdivided the hypothalamus anatomically and functionally into lateral hypothalamus
(LH) and medial hypothalamus (MH). This is the first study showing how the LH and MH causally
interact with other neural regions and how their dynamics change with weight and homeostasis
in humans. Adopting state-of-the-art spectral dynamic causal modelling of resting-state fMRI
data, we provide new insights into how homeostasis affect hypothalamic circuit dynamics,
which involve a distributed network of midbrain and cortical areas with a key role of the
substantia nigra. We identified unique aspects of network organisation associated with obesity
involving reciprocal connections between the LH and MH, and input from the substantia nigra
to the MH.
Citation: Voigt, K., Andrés, Z. B.,
Harding, je. H., Razi, UN., & Verdejo-
García, UN. (2022). Hypothalamic
effective connectivity at rest is
associated with body weight and
energy homeostasis. Réseau
Neurosciences, 6(4), 1316–1333.
https://doi.org/10.1162/netn_a_00266
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00266
Informations complémentaires:
https://doi.org/10.1162/netn_a_00266;
https://github.com/ katharinavoigt1
/spdcm_hypo.git
Reçu: 16 Février 2022
Accepté: 27 Juin 2022
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Antonio Verdejo-García
antonio.verdejo@monash.edu
Éditeur de manipulation:
Jason N. MacLean
droits d'auteur: © 2022
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
.
t
/
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
INTRODUCTION
The hypothalamus accounts for only approximately 3% of total human brain tissue, but is one
of the most vital structures regulating a plethora of bodily functions essential for survival
(Stuber & Sage, 2016). This small subcortical region regulates our response to stress, arousal,
reward processing, body temperature, fertility and sexual behaviour, motivation, and food
intake (Gabriela Pop et al., 2018). Early preclinical lesion studies subdivided the hypothalamus
anatomically and functionally into lateral hypothalamus (LH) and medial hypothalamus (MH),
leading to the concept of a “dual centre model” (Anand & Brobeck, 1951; Brobeck et al.,
1943; Elmquist et al., 1999; Hetherington & Ranson, 1983). Lesions to the MH resulted in
increased appetite, food intake and weight gain, marking the MH as “satiety centre.” Lesions
to the LH in turn induced abnormal decreases in appetite and food intake, labelling the LH as
“hunger centre.” More recent studies show that neurons in the LH regulate food consumption
and appetitive motivation with extensive reciprocal connections to the dopaminergic midbrain
governing reward processing in support of goal-directed food seeking (Jennings et al., 2013,
2015; Nieh et al., 2016; Rossi & Stuber, 2018). The LH is a large single region with numerous
heterogeneous neuronal populations, whereas the MH can be further subdivided into many
important nuclei involved in the regulation of food intake, blood glucose, and weight control.
This includes the arcuate nucleus, the ventromedial hypothalamic nucleus, the dorsomedial
hypothalamic nucleus, and the paraventricular nucleus. Both the LH and MH nuclei function
in a metabolic state-dependent manner and can be reshaped by obesity and energy homeo-
stasis (Chen et al., 2017; Rossi et al., 2019). De plus, these hypothalamic areas are heavily
integrated into intra- and interhypothalamic neural circuits and networks, with the majority of
LH connectivity coming from outside the LH (Burdakov & Karnani, 2020).
Although animal research has greatly contributed to understanding how hypothalamic neu-
ral circuits integrate peripheral and central signals to control food intake, the connectivity in
humans to and from the LH and MH nuclei remains poorly understood. Energy homeostasis
relies on the coordinated and dynamic interactions of the hypothalamus both to (bottom-up)
et de (top-down) a broad set of cortical and subcortical brain regions (Rossi & Stuber,
2018). A precise description of how the LH and MH network functions in response to changes
in homeostatic state in humans is thus required to bridge the gap between animal and human
recherche, and to provide a critical step towards defining the neural underpinnings of maladap-
tive eating patterns leading to obesity in humans. An examination of the LH and MH networks
is further supported by the well-known psychological comorbidities associated with metabolic
diseases such as anorexia, obesity, and diabetes (Florent et al., 2020; Penninx & Lange, 2018).
Research has begun to establish the links between the hypothalamic network and obesity
using functional magnetic resonance imaging (IRMf). One study by Kullmann and colleagues
(2014) described differences in the hypothalamic network between people with excess weight
and those with healthy weight. Functional connectivity analyses revealed the LH was more
heavily connected to the dorsal striatum, anterior cingulum, and frontal operculum, et le
MH was more connected to the medial orbitofrontal cortex and nucleus accumbens (repli-
cated recently by Zhang et al., 2018). Plus loin, in participants with excess weight, the func-
tional connectivity of the MH, but not the LH, was increased with the nucleus accumbens
and medial prefrontal cortex. These results highlight the existence of two distinct circuitries
originating from the MH and LH that are modulated by obesity. Cependant, these studies do
not reveal the functional interactions (par exemple., inhibition or excitation) nor do they differentiate
between bottom-up and top-down effects within the network. Plus loin, given the metabolic
state-dependency of the LH and MH circuitries (Chen et al., 2017; Rossi et al., 2019), it is also
Neurosciences en réseau
1317
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
t
/
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
Spectral dynamic causal
modelling (spDCM):
spDCM is a method for analyzing
resting-state functional magnetic
resonance imaging (IRMf) data that
provides information about
directionality (inhibition vs.
excitation) of connectivity between
zones du cerveau.
critical to investigate this network as a function of more dynamic state-dependent changes in
energy homeostasis, such as in states of fasting versus satiety.
The current study examines the directionality (bottom-up vs. top-down) and valence (inhi-
bition vs. excitation) of connections of the LH and MH with key cortical and subcortical brain
régions. These network dynamics are examined in participants varying in weight (healthy vs.
excess weight) in a fasted or sated state. We capitalise on recent advances in modelling the
interactions within a brain network based on the low-frequency endogenous fluctuations in
resting-state functional magnetic resonance imaging (rsfMRI) data using spectral dynamic
causal modelling (spDCM; Friston et al., 2014; Razi et al., 2015; Park et al., 2018) and state
of the art anatomical labelling (Rolls et al., 2020). In contrast to conventional functional
connectivity analyses (par exemple., Kullmann et al., 2014; Zhang et al., 2018), spDCM predicts direc-
tional communications among distributed brain regions (c'est à dire., effective connectivity; Friston,
Harrison, & Penny, 2003). We hypothesise that the LH and MH would show distinct effective
connectivité. Spécifiquement, based on reviewed previous functional connectivity studies distin-
guishing between MH and LH (Kullmann et al., 2014; Zhang et al., 2018), we predicted that
le (un) LH might be more heavily interconnected than the MH with the dorsal striatum, anterior
cingulum, and frontal operculum, et (b) that these connections from and to the MH, but not
LH, are affected by BMI. Given that there are no previous fMRI studies investigating the net-
work dynamics of the LH and MH across homeostatic state, how hunger and satiety affect the
hypothalamic network dynamics was explorative.
MÉTHODES
Participants
Forty participants were recruited via flyers and social media advertisements. Participants were
required to be 18–55 years old, right-handed, and have a body mass index (BMI) entre 18
et 30 kg/m2. Screening criteria excluded people with a history of hypertension or diabetes,
neurological or psychiatric illness, or who had recently taken psychoactive medications. Addi-
tionally, participants could not be subject to MRI contradiction, such as metal implants or
pregnancy. The number of participants was chosen based on a sample size estimation study
révélant que 20 participants provided for reliable DCM predictions (Goulden et al., 2012). Dans
agreement, recent research showed robust model predictions using similar sample sizes when
applying spDCM to rsfMRI data (Park et al., 2018; Preller et al., 2019; Voigt et al., 2020). Out
of the 40 participants, two were excluded from analysis as they did not complete both fasted
and sated rsfMRI scans. In total, data from 38 participants were included into the analyses
(Tableau 1 for participants’ demographics). From these participants, 22 had healthy weight
(18.56–24.27 kg/m2), 4 were overweight (25.12–28.82 kg/m2), et 12 were obese (30.84–
55.55 kg/m2). All participants gave written consent before participating and were reimbursed
avec $100 gift card vouchers. The Monash University Human Research Ethics Committee
approved the study (2019-5979-30222) following the Declaration of Helsinki.
Experimental Procedure
Participants completed two resting-state fMRI scans, one after an overnight fast (fasted condi-
tion) and one after a standard breakfast (sated condition). In both conditions, participants were
instructed to have a standard meal (700–1,000 kJ) entre 7.30 pm and 8.30 pm on the night
prior to their scan and subsequently to refrain from eating or drinking (except for water) jusqu'à
their morning scan. Fasting blood glucose levels were measured via a standard finger prick
test. For the sated condition, participants received a breakfast (293 kcal) 1 hour prior to their
Neurosciences en réseau
1318
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
.
t
/
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
Tableau 1.
Participants’ demographics
Characteristic
Age
Female/male
Obese and overweight/healthy weighta
BMIb
Hip-waist ratioc
Blood glucosed
Hunger fastede
Hunger satede
M.
27.40
25/13
16/22
26.44
0.92
4.94
4.41
3.22
SD
8.10
7.16
0.10
0.79
1.44
1.48
Range
18–48
18.56–55.56
0.79–1.25
3.8–8.4
1.18–7
1.45–6.21
Note. M = mean; SD = standard deviation; BMI = body mass index.
a Healthy weight = BMI between 18–25 kg/m2; Obese = BMI from 30 kg/m2.
b BMI = weight (kg)/height (m2).
c Waist circumference divided by hip circumference.
d Blood glucose levels were assessed via finger prick test conducted at the start of the fasted condition.
e Hunger levels were based on self-report via a 1 (not at all) à 7 (very much) Likert scale.
scan. Subjective self-reports of hunger (1 = not hungry at all; 7 = very hungry) revealed a sig-
nificant difference in the perception of hunger during the fasted (M = 4.42; SD = 1.44) et
sated (M = 3.22; SD = 1.48) condition (t(36) = 4.72, p < 0.001). There was no interaction
between subjective reported hunger and BMI, but there was between BMI and fasting blood
glucose level (see Supporting Information Figure S1 and Figure S2). All scans were scheduled
in the morning between 9 am and 10 am. On average there were 5.82 days (SD = 3.73 days)
between the two scanning sessions. The order of fasted and sated scans was counterbalanced
across participants.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
a
_
0
0
2
6
6
p
d
/
.
t
Resting-State fMRI Data Acquisition
Resting-state fMRI data were acquired using a 3-Tesla Siemens Skyra MRI scanner equipped
with a 32-channel head coil at the Monash Biomedical Imaging Research Centre (Melbourne,
Victoria, Australia). During a total acquisition time of 7.8 minutes, 600 volumes were acquired
for each participant and homeostatic condition using a multiband gradient echo pulse
sequence (45 axial slices; time of repetition, TR = 780 ms; echo time, TE = 21 ms, resolution
3 × 3 × 3 mm). In order to obtain structural brain information for each participant, a high-
resolution T1-weighted magnetisation-prepared rapid gradient echo covering the whole brain
was measured (repetition time = 2,300 ms; echo time = 2.07 ms; flip angle = 9°; 192 slices;
field of view = 256 × 256 mm, voxel resolution = 1 mm isotropic). Participants were instructed
to rest while fixating on a central black crosshair (i.e., eyes-open resting-state protocol).
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Resting-State fMRI Data Analyses
Functional images were preprocessed using SPM12 (revision 12.2, www.fil.ion.ucl.ac.uk). The
preprocessing steps consisted of spatial realignment, tissue segmentation, and spatial normal-
isation to the standard EPI template of the Montreal Neurological Institute (MNI), and spatial
smoothing using a Gaussian kernel of 6-mm FWHM. None of the participants exceeded exces-
sive head motion of larger than 3 mm. For the seed-based functional connectivity analyses, we
Network Neuroscience
1319
Hypothalamic effective connectivity
applied an additional temporal band-pass filter (0.01–0.08 Hz) to remove low-frequency drifts
and high-frequency physiological noise as well as linear detrending the data. Nuisance covar-
iate regression was performed to remove signal variance of nonneuronal origin using time
series extracted from the white matter, and independently from the cerebrospinal fluid, in addi-
tion to the six parameters to define the magnitude of frame-by-frame head motion (3 × trans-
lation; 3 × rotation).
Statistical fMRI Analyses
We first conducted an initial seed-based functional connectivity analysis (using the bilateral
MH and LH as seeds; Figure 1). This analysis was used to obtain the brain areas that are asso-
ciated with the MH and LH at rest (i.e., the hypothalamic functional resting-state network).
Next, we conducted a spDCM analyses to investigate the causal interactions between these
areas and how they differ as a function of homeostatic state (fasted vs. sated), BMI, and
the interaction between homeostatic state and BMI. The details of these two analyses are
outlined next.
Seed-Based Functional Connectivity Analysis
Functional connectivity maps of the hypothalamic functional resting-state network were
obtained using an initial seed-based functional connectivity analyses across all subjects based
on the data obtained during the fasted condition (Kullmann et al., 2014). We defined two ROIs
according to Baroncini et al. (2012): the bilateral LH; MNI coordinates x = ±6, y = −9, z =
−10) and bilateral MH; MNI coordinates x = ±4, y = −2, z = −12) using 2-mm-radius spheres
(Figure 1). To minimise overlap between the two ROIs, we chose the peak voxel of the LH to
be in the posterior part of the LH according to Baroncini et al. (2012). The seeds were, as
such, spatially separated by >6 mm (c'est à dire., >1 mm after smoothing).
In order to define the general hypothalamus network that was associated with either of the
LH or MH across subjects, we extracted the average time series from LH and MH combined.
This time series was then correlated with the time series of activity within each of voxel across
the rest of the brain. The resulting functional connectivity maps were transferred to z-scores
using Fisher’s transformation and analysed using a one-sample’s t test in SPM12 ( Wellcome
Department of Cognitive Neurology, Londres, ROYAUME-UNI). Brain voxels with a threshold of p < 0.05,
family-wise error (FWE) corrected for multiple comparisons on the voxel-level were consid-
ered significant. Anatomical regions were labelled using the recent update of the automatic
anatomical labelling atlas AAL3 (Rolls et al., 2020). This updated atlas includes brain areas
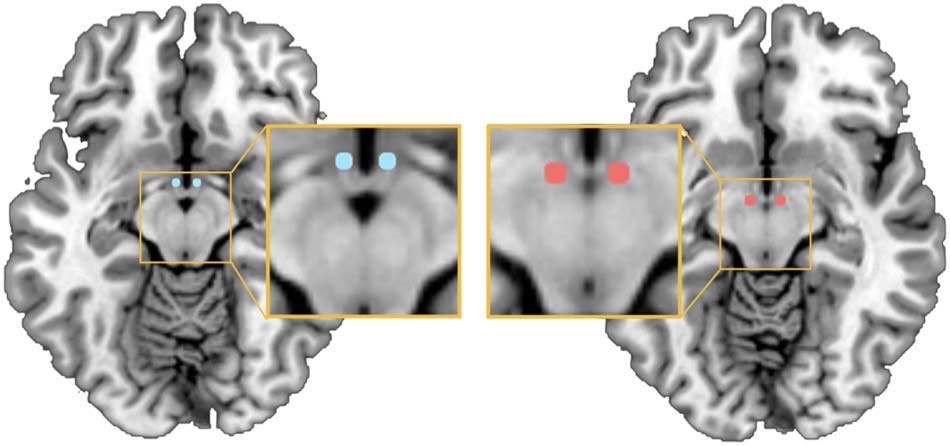
Seed regions of the medial hypothalamus (MH; MNI coordinates x = ±4, y = −2, z = −12;
Figure 1.
left depicted in blue), and lateral hypothalamus (LH; MNI coordinates x = ±6, y = −9, −10; right
depicted in red) (based on Baroncini et al., 2012) used to obtain the hypothalamic network for
subsequent spDCM analyses.
Network Neuroscience
1320
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
a
_
0
0
2
6
6
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
that have not generally been defined in other atlases, such as subdivisions of the thalamus or
the substantia nigra. Previous studies investigating the functional connectivity of the MH and
LH (Kullmann et al., 2014; Zhang et al., 2018) have not used such precise anatomical
labelling.
Spectral Dynamic Causal Modelling
The spDCM analyses were performed using the functions of DCM12 (revision 7196) imple-
mented in SPM12 (version 7487) in MATLAB 2018b. In order to address our main hypotheses,
we focused on spDCM analyses that assessed four questions: (1) effective connectivity of the
hypothalamic network in the fasted and sated states independently; (2) changes in hypotha-
lamic effective connectivity between the fasted versus sated state, independent of BMI (main
effect of fasting); (3) changes in hypothalamic effective connectivity modulated by BMI, inde-
pendent of energy state (main effect of BMI); and (4) changes in fasting-related effective con-
nectivity of the hypothalamus modulated by BMI (fasting-by-BMI interaction). Question (1) will
be assessed via the first-level spDCM analysis and Question (2)–(4) will be assessed via the
second-level spDCM analyses. The first level of the spDCM is an intercept model, which esti-
mates the direction and valence (i.e., inhibition/excitation) regardless of any behavioural var-
iable (e.g., BMI, homeostasis). As such, it estimates the neuronal network dynamics of our
chosen network (as established via an initial functional connectivity analyses) during the
resting-state fMRI whilst individuals where fasted and sated. On the next level, we estimated
additional spDCM to assess the effects of BMI and homeostasis and their interaction on the
neuronal network dynamics.
In DCM, each connection has a prior distribution, which assumes them to have a normal
(Gaussian) distribution (the so-called Laplace approximation). The priors on connectivity
parameters are given in Table 1 of Friston et al. (2014). Each extrinsic (between-region) con-
nection has a small but positive prior mean value (i.e., excitatory connectivity) but with a var-
iance that allows it to take on a posterior connectivity which can either be inhibitory (i.e.,
becoming negative) or become more excitatory after model fitting is performed. The intrinsic
or self-connections are modelled as inhibitory only as they represent a recurrent activity within
a region. A more detailed description of spDCM is provided in the Supporting Information.
Since the development of spDCM, a number of studies have used this method in order to
establish the directed excitation/inhibition between brain areas in the various contexts, such
as, recently in relation to emotional intelligence (Bajaj & Killgore, 2021), internet gaming dis-
order (Dong et al., 2021), or dementia (Benhamou et al., 2020).
First-Level spDCM Analysis
In order to assess the effective connectivity of the hypothalamus network, regions revealed by
the initial functional connectivity analyses of both the MH and LH in conjunction with a min-
imum voxel size of 20 were used as ROIs for the subsequent spDCM analyses (Figure 2,
Table 2). As such, our ROIs were not defined based on previous literature (e.g., Kullmann
et al., 2014), but rather in a data-driven fashion. This ROI selection approach is statistically
valid (Poldrack, 2007) and has been used previously (e.g., Esménio et al., 2019). At the first-
level, a fully connected model was created for each participant and each session. Next, we
inverted (i.e., estimated) the DCMs using spectral DCM, which fits the complex cross-spectral
density using a parameterised power-law model of endogenous neural fluctuations (Razi et al.,
2015). This analysis provides measures of causal interactions between regions, as well as the
amplitude and exponent of endogenous neural fluctuations within each region (Razi et al.,
2015). Model inversion was based on standard variational Laplace procedures (Friston
Network Neuroscience
1321
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
a
_
0
0
2
6
6
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
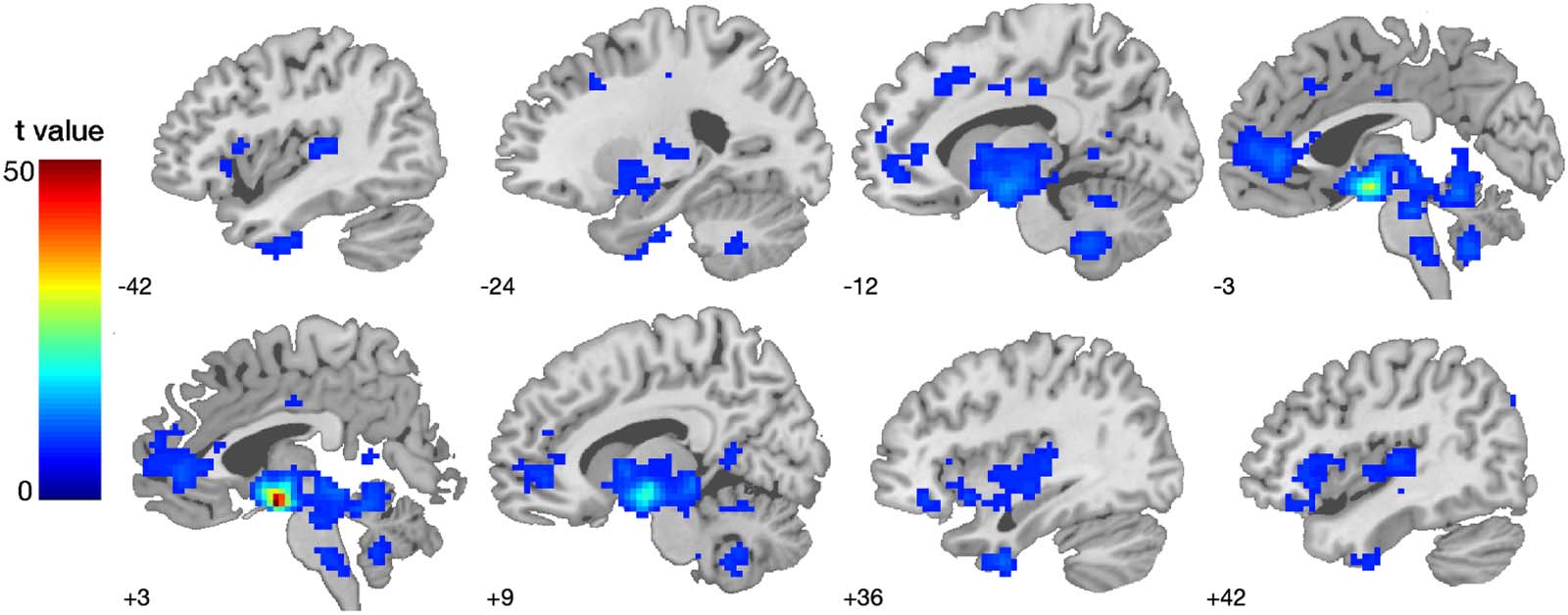
Figure 2. Functional connectivity network of the lateral and medial hypothalamus. Seed-based functional connectivity results by using the
bilateral seed mask LH and MH (Figure 1). Results reflect whole-brain one-sample t tests at p < 0.001 FWE-corrected. For differences in MH
and LH functional connectivity, refer to Table 2. The colour bar represents voxel T values.
et al., 2007). This Bayesian inference method uses Free Energy as a proxy for (log) model evi-
dence, while optimising the posterior density under Laplace approximation.
Second-Level spDCM Analysis
To characterise how group differences in neural circuitry were modulated by BMI and energy state,
hierarchical models over the parameters were specified within a parametric empirical Bayes (PEB)
framework for DCM (Friston et al., 2016). The five models we used were based on our hypotheses
as follows. Firstly, to investigate the effective connectivity of the MH and LH, two PEB models were
estimated for the fasted and sated condition separately. These (intercept) models provide the base-
line effective connectivity independent of any behavioural measures. Secondly, we were inter-
ested in the group difference between fasted versus sated conditions, and in this PEB analysis,
we contrasted the DCMs for the fasted against the sated condition whilst controlling for BMI.
Thirdly, we were interested in associating effective connectivity with BMI and used BMI as a main
regressor of interest whilst controlling for homeostatic condition. Lastly, we were interested in
interaction between group factor (fasted vs. sated) and BMI, and in this PEB analysis, we used
the interaction between BMI and the group factor (fasted vs. sated) as main variables of interest.
For each of the presented models, all behavioural regressors were mean centred so that the
intercept of each model was interpretable as the mean connectivity. We tested the relation-
ships between all covariates (i.e., age, gender, BMI, subjective hunger reports, blood glucose
levels; see Supporting Information) and included blood glucose as a mean-centred covariate,
as it correlated with BMI in interaction with the experimental condition. We further controlled
for age and gender in every model. Hunger condition was a grouping variable, whereas BMI
was a continuous variable. BMI was treated as continuous, as opposed to a grouping variable,
as our sample contained overweight (BMI = 25–30 kg/m2) and obese (BMI > 30 kg/m2)
individuals.
Bayesian model reduction was used to test all combinations of parameters (c'est à dire., reduced
models) within each parent PEB model (assuming that a different combination of connections
could exist; Friston et al., 2016) and ‘pruning’ redundant model parameters. Parameters of the
set of best-fit pruned models (in the last Occam’s window) were averaged and weighted by
their evidence (c'est à dire., Bayesian model averaging) to generate final estimates of connection
parameters. To identify important effects (c'est à dire., changes in directed connectivity), we compared
models, using log Bayesian model evidence to ensure the optimal balance between model
Neurosciences en réseau
1322
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
.
t
/
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
Tableau 2.
Peak coordinates of hypothalamus intrinsic functional connectivity networks
Brodmann’s
area (BA)
Hemisphere
Peak location
(X, oui, z)b
Cluster
sizec
T score
Local maxima labellinga
MH and LH
Substantia nigra, pars compacta (prSN)
Cortex cingulaire antérieur, pregenual (pACC)
Inferior temporal gyrus (ITG)
Middle cingulate & paracingulate gyri (MCC)
Inferior temporal gyrus (ITG)
Inferior frontal gyrus, opercular part
Angular gyrus
MH > LH
Substantia nigra, pars compacta (prSN)
Lobule IX of cerebellar hemisphere
LH > MH
Red nucleus
Middle frontal gyrus
Inferior frontal gyrus, opercular part
Putamen
Cortex cingulaire antérieur, supracallosal
Middle frontal gyrus
Inferior frontal gyrus, opercular part
Inferior parietal gyrus, excluding supramarginal and angular gyri
Inferior parietal gyrus, excluding supramarginal and angular gyri
Lobule VIII of cerebellar hemisphere
Cuneus
Lobule X of cerebellar hemisphere
Superior temporal gyrus
Middle cingulate gyrus
Gyrus frontal supérieur, dorsolateral
Putamen
Superior occipital gyrus
BA32
BA32
BA6
BA44
BA10
BA38
R.
L
R.
L
L
L
R.
R.
L
R.
L
R.
R.
R.
R.
L
R.
L
L
L
L
R.
R.
R.
R.
R.
a Labelling based on AAL3 atlas (Rolls et al., 2020).
b Peak voxel location in MNI space.
c The report of the functional network for the hypothalamus network (MH and LH) was limited to regions sized > 20 voxels.
Neurosciences en réseau
3, −3, −15
3,407
50.42
−3, 42, 3
516
12.03
36, −3, −45
144
11.51
−9, 24, 33
−42, −6, −42
−42, 15, 9
42, −78, 39
123
137
47
27
8.85
8.66
8.37
7.90
3, −3, −12
−6, −36, −60
92
57
32.66
7.48
6, −12, −9
978
23.38
−39, 39, 24
334
11.14
48, 9, 27
30, 15, 0
9, 24, 21
36, 42, 27
−54, 9, 24
51, −36, 45
−45, −45, 42
−33, −57, −60
−18, −78, 36
−9, −30, −39
54, 15, −9
12, −30, 39
30, 3, 57
36, −12, −9
21, −72, 36
61
85
301
265
119
322
88
17
64
23
42
42
82
22
22
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
9.98
9.82
9.82
9.71
9.64
9.63
9.47
9.45
9.15
8.80
8.63
8.44
8.41
7.95
7.71
1323
Hypothalamic effective connectivity
complexity and accuracy, with and without each effect, and calculated the posterior probability
for each model as a softmax function of the log Bayes factor. We treat effects (c'est à dire., connection
strengths and their changes) with a strong posterior probability > 0.99 (equivalent of very strong
evidence in classical inference) as significant for reporting purposes. This posterior probability
indicates very strong evidence for effects in Bayesian statistics (Kass & Raftery, 1995).
Enfin, in order to determine the predictive validity (par exemple., whether BMI can be predicted
from the final, reduced spDCM’s individual connections), leave-one-out cross-validation
was performed within the PEB framework (Zeidman et al., 2019). This procedure fits the
PEB model in all but one participant and predicts the covariate of interest (par exemple., homeostatic
state) for the left-out participant. This is repeated with each participant to assess the averaged
prediction accuracy for each model.
RÉSULTATS
We first provide the overview of the seed-based functional connectivity analysis, ce qui était
conducted for the derivation of the hypothalamic network. Secondly, we describe the causal
dynamics within this derived hypothalamic network.
Seed-Based Functional Connectivity Analyses
The seed-based functional connectivity analyses (combined LH and MH seed mask, Chiffre 1)
revealed a hypothalamic network comprising seven regions in the substantia nigra, anterior
and middle cingulate, inferior temporal and frontal gyrus, and the angular gyrus (Tableau 2).
These seed-based functional connectivity results subtending the MH and LH functional
connectivity were used to define the network for each participant for the subsequent effec-
tive connectivity analysis. The differential contrast (MH > LH) revealed a stronger functional
connectivity between the MH and substantia nigra, cerebellum, and precuneus. The lateral
hypothalamus (LH > MH contrast) showed stronger functional connectivity to the red
nucleus, inferior and middle frontal gyrus, putamen, anterior and middle cingulate cortex,
putamen and regions of the parietal, occipital lobe, as well as cerebellum, ( p < 0.001, FWE-
corrected; Table 2, Figure 2).
Spectral Dynamic Causal Modelling Results
The average variance explained across subject-level DCM inversion was very high (Hunger:
M = 86.78, SD = 3.23, range = 80.20–94.90; Satiety: M = 86.66, SD = 2.58, range = 80.02–
91.68), indicating very good model convergence.
Effective Connectivity of the MH and LH During the Fasted and Sated Condition
Figure 3 illustrates the effective connectivity results of the LH and MH during the fasted and
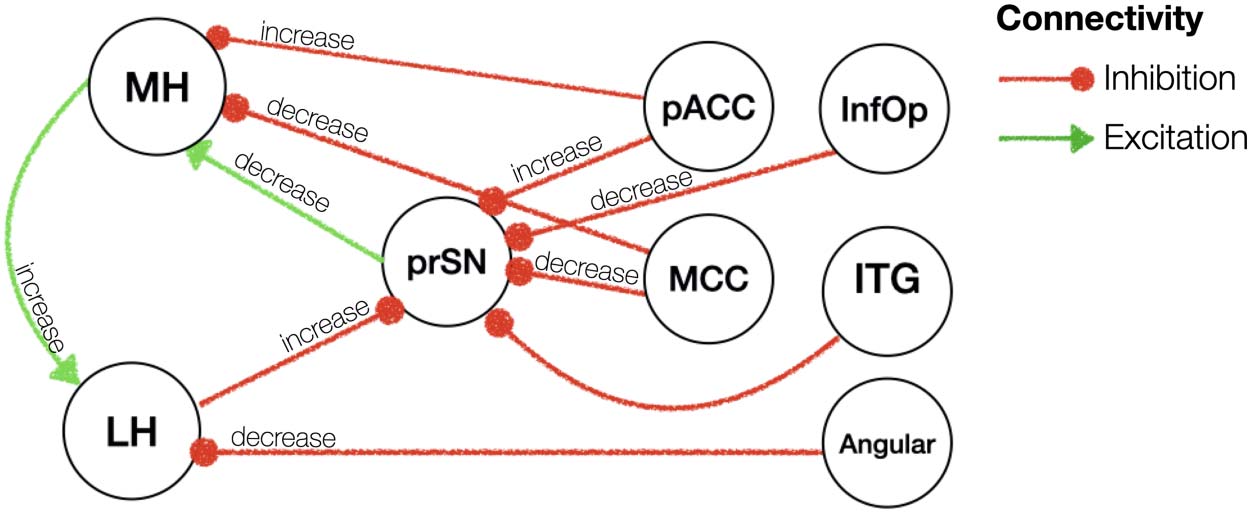
sated condition separately. Across the fasted and sated condition, there was inhibition from
anterior and mid-cingulate, frontal, temporal and parietal cortex to the LH and MH. There
was no evidence for connections going from the LH or MH to these cortical regions in the
fasted condition. During the sated condition, there was excitation from the right MH to the
angular gyrus, and from the right LH to the pACC.
Effective Connectivity of the MH and LH in Fasted Versus Sated States
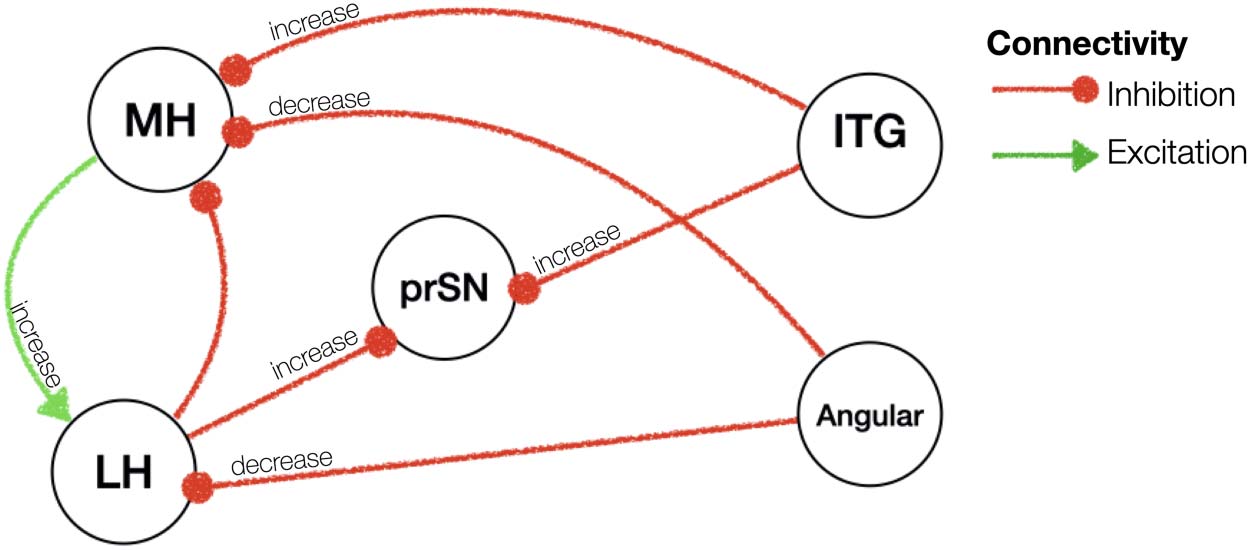
Fasting, compared to satiety, was associated with a decreased excitatory influence from the
substantia nigra to the left MH (0.01 Hz, 95% CI [−0.03, 0.004]). The left MH in turn showed
Network Neuroscience
1324
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
a
_
0
0
2
6
6
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
Figure 3. Effective resting-state connectivity of the hypothalamic network across during the fasted versus sated condition. Colour bar indi-
cates effect sizes in hertz. Posterior probability of >.99 (very strong evidence). prSN = substantia nigra pars compacta; MH = medial hypo-
thalamus; LH = lateral hypothalamus; pACC = anterior cingulate cortex pregenual; MCC = middle cingulate and paracingulate gyri; ITG =
inferior temporal gyrus; InfOp = inferior frontal gyrus, opercular part.
an increased excitation to the left LH (0.06 Hz, 95% CI [−0.002, 0.11]) and to the right MH
(0.06 Hz, 95% CI [0.007, 0.12]). We further found an increased inhibition from the pregenual
anterior cingulate cortex onto the bilateral MH (right hemisphere: 0.14 Hz, 95% CI [0.08,
0.20]; left hemisphere: 0.07 Hz, 95% CI [0.004, 0.14]) and a decreased inhibition from the
middle cingulate onto the right MH (−0.01 Hz, 95% CI [−0.03, −0.01]). The left LH in turn
received less inhibition from the angular gyrus (−0.01 Hz, 95% CI [−0,02, 0.01]) and exerted
stronger inhibition on the substantia nigra (0.01 Hz, 95% CI [−0.007, 0.02]).
Outside the MH and LH, the substantia nigra pars compacta received a large number of
inhibitory inputs from the areas of the cingulate, frontal and temporal cortices. Spécifiquement,
during the fasted as opposed to sated state, the substantia nigra pars compacta received a
greater inhibition from the bilateral inferior temporal gyrus (gauche: 0.01 Hz, 95% CI [−0.007,
0.02]; droite: 0.01 Hz, 95% CI [−0.007, 0.024]) and the pregenual anterior cingulate cortex
(0.01 Hz, 95% CI [−0,02, 0.01]) as well as lower inhibition from the middle cingulate cortex
(−0.02 Hz, 95% CI [−0,02, 0.006]) and inferior opercular frontal gyrus (−0.01 Hz, 95% CI
[−0,02, 0.01]) (results are summarised in Supporting Information Table S1 and illustrated in
Chiffre 4).
Effective Connectivity Changes of the MH and LH as a Function of BMI
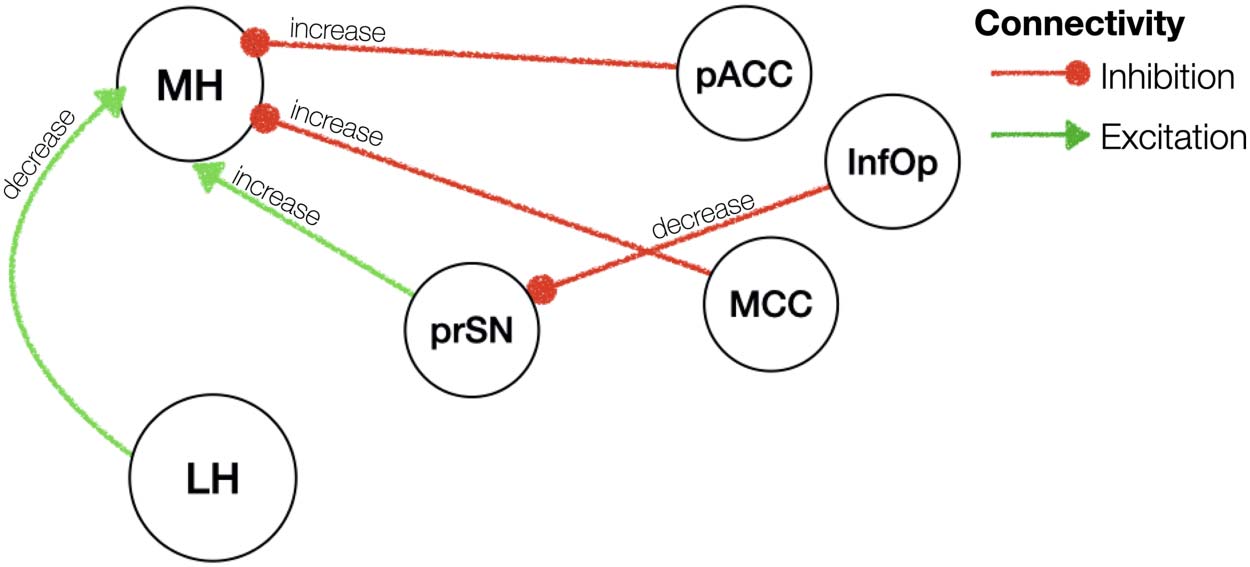
BMI, independent of homeostatic state, was associated with a greater excitatory influence from
the left MH to the left LH (0.017 Hz, 95% CI [0.01, 0.03]) and greater inhibition from the right
LH to the right MH (0.02 Hz, 95% CI [0.01, 0.02]). We further found a greater inhibition from
the inferior temporal gyrus and angular gyrus to the MH (see Table 2 for lateralities and effect
sizes). A greater inhibitory influence of the right LH on the substantia nigra (0.007 Hz, 95% CI
[0.003, 0.012]) and the right MH (0.02 Hz, 95% CI [0.02, 0.024]) was also evident in individ-
uals with a higher BMI. The substantia nigra pars compacta in turn received a greater inhibi-
tion from the bilateral inferior temporal gyrus (gauche: 0.014 Hz, 95% CI [0.01, 0.02]; droite:
Neurosciences en réseau
1325
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
t
/
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
Chiffre 4. Effective connectivity of fasted versus sated state. Red/green arrows indicate inhibitory/
excitatory connectivity. All these connections had posterior probability of >0.99 (very strong evi-
dence). Note that hemispheric information is not shown in this figure for illustration purposes.
Please refer to text for details on the hemispheric results. prSN = substantia nigra pars compacta;
MH = medial hypothalamus; LH = lateral hypothalamus; pACC = anterior cingulate cortex pregen-
ual; MCC = middle cingulate and paracingulate gyri; ITG = inferior temporal gyrus; InfOp = inferior
frontal gyrus, opercular part; Angular = Angular gyrus.
0.008 Hz, 95% CI [0.004, 0.01]) (results are summarised in Supporting Information Table S2
and illustrated in Figure 5).
Effective Connectivity Changes in Fasted Versus Sated States in Interaction With BMI
In the final analysis, we investigated how fasted-related connectivity changes were modulated
by differences in BMI (Supporting Information Table S3 and Figure 6). During fasting relative to
satiety, higher BMI was associated with a higher excitatory influence from the substantia nigra
to the left MH (0.006 Hz, 95% CI [0.002, 0.009]). The left MH received an increased excita-
tion from the right MH (0.013 Hz, 95% CI [0.004, 0.02]) as well as a decreased excitation from
the left LH (−0.014 Hz, 95% CI [−0,02, −0.005]). The left MH also received higher inhibition
from the anterior (0.001 Hz, 95% CI [0, 0.001]) and middle cingulate cortex (0.001 Hz, 95%
CI [0, 0.002]). The substantia nigra pars compacta received a greater inhibition from the infe-
rior opercular frontal gyrus (0.006 Hz, 95% CI [−0.01, −0.002]) (results are summarised in
Supporting Information Table S3 and illustrated in Figure 6).
Effective connectivity of BMI. Red/green arrows indicate inhibitory/excitatory connec-
Chiffre 5.
tivité. Note that hemispheric information is not shown in this figure for illustration purposes.
Please refer to text for details on the hemispheric results. prSN = substantia nigra pars compacta;
MH = medial hypothalamus; LH = lateral hypothalamus; ITG = inferior temporal gyrus; Angular =
angular gyrus.
Neurosciences en réseau
1326
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
t
.
/
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
Effective Connectivity of BMI × Homeostatic state interaction effect. Red/green arrows
Chiffre 6.
indicate inhibitory/excitatory connectivity. Note that hemispheric information is not shown in this
figure for illustration purposes. Please refer to text for details on the hemispheric results. prSN =
substantia nigra pars compacta; MH = medial hypothalamus; LH = lateral hypothalamus; pACC =
anterior cingulate cortex pregenual; MCC = middle cingulate and paracingulate gyri; ITG = inferior
temporal gyrus; InfOp = inferior frontal gyrus, opercular part.
DISCUSSION
This is the first study to reveal how the LH and MH causally interact with other neural regions
and how their dynamics change with weight and energy state in humans. Adopting state of the
art spectral dynamic causal modelling of resting-state fMRI data (Friston et al., 2014; Parc
et coll., 2018; Razi et al., 2015), our results show two core networks interacting: (un) subcortical
bidirectional connections between the LH, MH, and the prSN, et (b) cortical top-down inhi-
bition from frontal, cingulate, and temporal onto the subcortical network. The prSN seems to
represent a central hub interconnecting the subcortical and cortical neural systems. During the
fasted compared to the sated state, regardless of weight status, we found increased inhibition
between the right LH and prSN as well as decreased excitation between the prSN and left MH,
whereas the prSN received top-down inhibition from across the cortex, which may represent
an adaptive motivational drive to seek food while hungry, in fitting with animal studies
(Cassidy & Tong, 2017; Nieh et al., 2016; Rossi et al., 2019; Rossi & Stuber, 2018). Cependant,
individuals with excess weight revealed a similar hypothalamic network communication irre-
spective of being in a fasted or sated state. Plus loin, when taking into consideration excess
weight, they showed a reverse communication pattern of decreased substantia nigra-MH inhi-
bition during the fasted state. The neural network communications involved in the regular pro-
cesses of food seeking after fasting may therefore be disrupted in individuals with excess
weight, providing a compelling hypothesis for food overconsumption beyond metabolic
needs. We previously used spDCM to investigate how the food choice network, lequel
includes the hypothalamus relates to BMI and homeostasis. Ici, we found that BMI and
homeostatic state modulated the hypothalamus connections to subcortical and cortical areas
(Voigt et al., 2021). Cependant, this work did not distinguish between the LH and MH, which are
known to have significant different functionalities (Anand & Brobeck, 1951; Brobeck et al.,
1943; Elmquist et al., 1999; Hetherington & Ranson, 1983), and did not specifically focus
on the hypothalamic network as established in this work, but rather on a food choice network
(Voigt et al., 2021).
Our results from whole-brain functional connectivity analyses largely reflect those reported
in earlier neuroimaging studies (Kullmann et al., 2014; Martín-Pérez et al., 2019; Zhang et al.,
2018). Both resting-state activity of LH and the MH was correlated with resting-state activity of
Neurosciences en réseau
1327
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
prSN, middle and anterior cingulate cortex, inferior frontal and temporal gyrus as well as
angular gyrus. Consistent with animal and human research, the LH was more strongly
connected than the MH across the entire brain, ranging from subcortical areas (par exemple., red
nucleus, putamen) to all neocortical areas. This finding strengthens the established role of
the LH as a central interface integrating diverse central and peripheral signals through a
complex large-scale neural network that may coordinate adaptive behavioural responses
related to motivation and controlled feeding behaviour (Bonnavion et al., 2016; Petrovich,
2018). The MH in turn was more strongly connected to the prSN, cerebellum, and precuneus.
These and previous findings (Kullmann et al., 2014; Martín-Pérez et al., 2019; Zhang et al., 2018)
highlight the potential of a dual hypothalamic functionality resulting from distinct LH and MH
neural networks. Cependant, these characterisations have been limited as the directionality and
valence of the interactions within these networks have remained unknown. Ici, we have
extended the characterisation of these networks by means of spDCM, investigating the
directed communication of the LH and MH networks and their changes as a function of both
BMI and energy state.
Irrespective of BMI and energy state, the prSN emerges as a key area connecting the hypo-
thalamus with neocortical regions. The prSN processes autonomic, gut-induced rewards
regulating motivational and emotional states (par exemple., Gutierrez et al., 2020; Han et al., 2018).
Hormones implicated in regulating the homeostatic system also impinge directly on dopamine
neurons in the prSN (Palmiter, 2007). Anatomically and functionally the prSN is highly inter-
connected with the ventral tegmental area ( VTA), and both regions contribute to motivation
and reward processing (Ilango et al., 2014; Kwon & Jang, 2014). In hungry mice, inhibitory
inputs from the LH to the VTA inhibited dopamine release, resulting in increased motivation to
seek and approach food (Nieh et al., 2016). In this study we found an increased inhibitory
influence from the LH to the prSN when participants were hungry. Given the strong prSN-
VTA interconnectivity and interchangeable functionality (Ilango et al., 2014; Kwon & Jang,
2014), it is reasonable to assume that the increased inhibition from the LH to the prSN in
humans might mirror a necessary survival mechanism to increase appetitive motivation to pre-
vent starvation. Notably, in individuals with higher BMI, regardless of their energy state, ce
inhibition from LH to prSN persisted. This failure to ‘shut off’ the inhibitory signalling might
reflect an underlying neural trigger for increased motivation for food regardless of homeostatic
state (Berthoud, 2004, 2012; Berthoud et al., 2017; Cassidy & Tong, 2017).
During the fasted compared to the sated state, we also found a decreased excitation from
the prSN to the MH. The MH contains a diverse array of nuclei and circuits, including the
anorectic melanocortin system that reduces food consumption, as well as increasing energy
expenditure (Kühnen et al., 2019). Ainsi, the reduced excitation during fasting compared to
satiety may reflect reduced activation of this anorectic pathway. Cependant, it is also important
to note that the MH contains strong drivers of appetite and motivation. Agouti-related peptide
neurons in the hypothalamic arcuate nucleus drive food intake and motivation (Andermann &
Lowell, 2017). In individuals with excess weight, this excitatory connectivity between prSN
and MH increased during the fasted compared to the sated state. This might contribute to
increased food seeking and consumption in response to energy deprivation among individuals
with overweight/obesity (Kühnen et al., 2019). Clairement, future research in humans is required to
examine the activity of specific hypothalamic nuclei within the MH region in relation to food
seeking and obesity; cependant, this is currently beyond the technical capability of MRI.
In addition to the communication between MH and LH with the prSN, we also found intra-
hypothalamic connectivity between the LH and MH across weight and homeostatic state. Le
dynamic from the ventral MH to the LH has been previously observed in animals (Canteras
Neurosciences en réseau
1328
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
t
.
/
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
et coll., 1994; Horst & Luiten, 1986; Luiten et al., 1980); cependant, its functional significance
remains untested and further studies in humans are needed to elucidate the functionality of
intrahypothalamic connectivity patterns.
During the fasted as opposed to the sated state, we found an increased top-down inhibition
from the pregenual anterior cingulate cortex to the MH, yet a decreased top-down inhibition
from the middle cingulate gyrus to the MH and prSN. While it is not possible to definitively
disentangle the role of these network dynamics in the current context, previous neuroimaging
studies that have dissociated the functions of the pregenual anterior cingulate cortex and the
middle cingulate gyrus provide for some conjecture (Stevens et al., 2011). En particulier, spon-
taneous activity in the pregenual anterior cingulate cortex is associated with affective process-
ing and anticorrelated with activity in sensorimotor areas. In contrast, activity in the middle
cingulate gyrus is temporally coupled with activity in sensorimotor areas, and functionally
connected with areas involved in cognitive control. The causal dynamics we report herein
between the middle cingulate gyrus, pregenual anterior cingulate cortex, and MH might
therefore suggest interactions between homeostatic inputs and affective, sensorimotor, et
cognitive networks dynamics. Note that the increased inhibition from the pregenual anterior
cingulate cortex to the MH was not related to BMI. Cependant, increased inhibition from the
middle cingulate gyrus to the MH was exacerbated in participants with higher BMIs. Future
studies are needed to clarify if this alteration is associated with core symptoms of obesity.
In addition to the cingulate cortex, the angular gyrus had a decreased impact on the LH,
when individuals were fasted or in individuals with excess weight. En outre, all cortical
areas inhibited either the prSN or hypothalamus irrespective of weight and homeostatic state.
Whereas the cingulate cortex is a hub for sensory, motivational and cognitive information, le
prefrontal and parietal cortex are more predominantly associated with executive control (Seeley
et coll., 2007). In participants with excess weight, a differential pattern within the executive con-
trol network has been observed in fMRI activation studies using food stimuli (Franssen et al.,
2020). Recently, it has also been shown that obesity is related to prominent functional connec-
tivity alterations mainly in prefrontal regions during resting-state as well as in response to food
stimuli (García-García et al., 2013; Kullmann et al., 2012). Ainsi, our resting-state findings might
further add to the possibility of disrupted communication between the executive control net-
work and regions regulating metabolic needs in individuals with excess weight.
In the light of the proposed mechanisms here, we note, cependant, that the relationship
between resting-state effective connectivity and its cognitive correlates remains elusive. Le
interindividual variations in effective connectivity do not necessarily overlap with the interin-
dividual variations in effective connectivity during task performance (Fox & Raichle, 2007;
Jung et al., 2018). At this stage, only one study has revealed that resting-state effective connec-
tivity might facilitate task performance but may not reflect task-based network dynamics (Jung
et coll., 2018). Future studies are needed to address whether the resting-state dynamics revealed
in our study are also engaged during task performance and how potential deviations might
translate to differences in behaviour or clinical phenotypes.
Food intake is orchestrated by a large-scale network of subcortical and cortical areas on
distinct higher and lower order cognitive functions (de Araujo et al., 2020). Both hemispheres
need cross talk to support functions such as food seeking and balanced food intake (Voigt
et coll., 2021). Our results support this view by revealing that there is no specific hemisphere
favoured and regions occur equally distributed in both, in the right and left hemisphere of the
brain. Previous studies included the bilateral MH/LH, but did not reveal whether lateralisation
might be functionally significant (Kullmann et al., 2014; Zhang et al., 2018), and reviews or
Neurosciences en réseau
1329
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
opinion papers on that matter did not discuss laterality (Petrovich, 2018). Although there has
been some animal research discussed that the hypothalamus might have functional lateralisa-
tion (Kiss et al., 2020), more research is needed to systematically test this claim in humans and
in relation to BMI and homeostasis. En tant que tel, in this discussion, we do not distinguish between
the left and right hemisphere and discussed each region functionality and bilaterally.
In conclusion, our study provides new insights into how hunger and satiety states affect
hypothalamic circuit dynamics, which involve a distributed network of midbrain and cortical
areas with a key role of the substantia nigra pars compacta. We also identified unique aspects
of network organisation associated with obesity, which involve the reciprocal connections
between the lateral and MH, and the input from the substantia nigra to the MH.
REMERCIEMENTS
The authors thank Richard McIntyre, Naomi Kakoschke, Amelia Romei, Tori Gaunson, et
Tiffany Falcone for help with MRI data acquisition.
INFORMATIONS À L'APPUI
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00266.
Data for main spectral dynamic causal modelling are available at GitHub: https://github
.com/KatharinaVoigt1/spDCM_Hypo.git (Voigt, 2021).
CONTRIBUTIONS DES AUTEURS
Katharina Voigt: Conceptualisation; Analyse formelle; Gestion de projet; Visualisation;
Rédaction – ébauche originale; Rédaction – révision & édition. Zane B. Andrés: Conceptualisation;
Acquisition de financement; Rédaction – révision & édition. Ian H. Harding: Conceptualisation; Formal
analyse; Acquisition de financement; Rédaction – révision & édition. Adeel Razi: Analyse formelle; Meth-
odology; Rédaction – révision & édition. Antonio Verdejo-Garcia: Conceptualisation; Funding
acquisition; Rédaction – révision & édition.
INFORMATIONS SUR LE FINANCEMENT
Antonio Verdejo-García, Zane Andrews, and Ian Harding, NHMRC Grant, Award ID:
GNT1140197.
RÉFÉRENCES
Anand, B. K., & Brobeck, J.. R.. (1951). Hypothalamic control of food
intake in rats and cats. The Yale Journal of Biology and Medicine,
24(2), 123–140. PubMed: 14901884
Andermann, M.. L., & Lowell, B. B. (2017). Toward a wiring diagram
understanding of appetite control. Neurone, 95(4), 757–778.
https://doi.org/10.1016/j.neuron.2017.06.014, PubMed:
28817798
Bajaj, S., Killgore, W. D. S. (2021). Association between emotional
intelligence and effective brain connectome: A large-scale spec-
tral DCM study. NeuroImage, 229, 117750. https://est ce que je.org/10
.1016/j.neuroimage.2021.117750, PubMed: 33454407
Baroncini, M., Jissendi, P., Balland, E., Besson, P., Pruvo, J.-P.,
Francke, J.-P., Dewailly, D., Blond, S., & Prevot, V. (2012). IRM
atlas of the human hypothalamus. NeuroImage, 59(1), 168–180.
https://doi.org/10.1016/j.neuroimage.2011.07.013, PubMed:
21777680
Benhamou, E., Maréchal, C. R., Russell, L. L., Hardy, C. J.. D., Bond,
R.. L., Sivasathiaseelan, H., … Razi, UN. (2020). The neurophysio-
logical architecture of semantic dementia: spectral dynamic
causal modelling of a neurodegenerative proteinopathy. Scien-
tific Reports, 10, 16321. https://doi.org/10.1038/s41598-020
-72847-1, PubMed: 33004840
Berthoud, H.-R. (2004). Neural control of appetite: Cross-talk
between homeostatic and non-homeostatic systems. Appetite,
43(3), 315–317. https://doi.org/10.1016/j.appet.2004.04.009,
PubMed: 15527935
Berthoud, H.-R. (2012). The neurobiology of food intake in an obe-
sogenic environment. Proceedings of the Nutrition Society, 71(4),
Neurosciences en réseau
1330
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
t
.
/
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
478–487. https://doi.org/10.1017/S0029665112000602,
PubMed: 22800810
Berthoud, H.-R., Münzberg, H., & Morrison, C. D. (2017). Blaming
the brain for obesity: Integration of hedonic and homeostatic
mechanisms. Gastroenterology, 152(7), 1728–1738. https://est ce que je
.org/10.1053/j.gastro.2016.12.050, PubMed: 28192106
Bonnavion, P., Mickelsen, L. E., Fujita, UN., de Lecea, L., & Jackson,
UN. C. (2016). Hubs and spokes of the lateral hypothalamus: Cell
les types, circuits and behaviour: LHA cell types and circuits. Le
Journal of Physiology, 594(22), 6443–6462. https://est ce que je.org/10
.1113/JP271946, PubMed: 27302606
Brobeck, J.. R., Tepperman, J., & Long, C. N. (1943). Experimental
hypothalamic hyperphagia in the albino rat. The Yale Journal of
Biology and Medicine, 15(6), 831–853. PubMed: 21434115
Burdakov, D., & Karnani, M.. M.. (2020). Ultra-sparse connectivity
within the lateral hypothalamus. Biologie actuelle, 30(20),
4063–4070. https://doi.org/10.1016/j.cub.2020.07.061,
PubMed: 32822604
Canteras, N. S., Simerly, R.. B., & Swanson, L. W. (1994). Organiza-
tion of projections from the ventromedial nucleus of the hypo-
thalamus: A Phaseolus vulgaris-Leucoagglutinin study in the
rat. Journal of Comparative Neurology, 348(1), 41–79. https://
doi.org/10.1002/cne.903480103, PubMed: 7814684
Cassidy, R.. M., & Tong, Q. (2017). Hunger and satiety gauge
reward sensitivity. Frontiers in Endocrinology, 8, 104. https://est ce que je
.org/10.3389/fendo.2017.00104, PubMed: 28572791
Chen, R., Wu, X., Jiang, L., & Zhang, Oui. (2017). Single-cell
RNA-Seq reveals hypothalamic cell diversity. Cell Reports, 18(13),
3227–3241. https://doi.org/10.1016/j.celrep.2017.03.004,
PubMed: 28355573
de Araujo, je. E., Schatzker, M., & Petit, D. M.. (2020). Rethinking food
reward. Revue annuelle de psychologie, 71, 139–164. https://doi.org
/10.1146/annurev-psych-122216-011643, PubMed: 31561741
Dong, G.-H., Dong, H., Wang, M., Zhang, J., Zhou, W., Du, X., &
Potenza, M.. N. (2021). Dorsal and ventral striatal functional con-
nectivity shifts play a potential role in internet gaming disorder.
Communications Biology, 4, 866. https://doi.org/10.1038/s42003
-021-02395-5, PubMed: 34262129
Elmquist, J.. K., Elias, C. F., & Saper, C. B. (1999). From lesions to
leptin: Hypothalamic control of food intake and body weight.
Neurone, 22(2), 221–232. https://doi.org/10.1016/S0896-6273
(00)81084-3, PubMed: 10069329
Esménio, S., Soares, J.. M., Oliveira-Silva, P., Zeidman, P., Razi, UN.,
Gonçalves, Ô. F., … Coutinho, J.. (2019). Using resting-state DMN
effective connectivity to characterize the neurofunctional archi-
tecture of empathy. Rapports scientifiques, 9, 2603. https://est ce que je.org/10
.1038/s41598-019-38801-6, PubMed: 30796260
Florent, V., Baroncini, M., Jissendi-Tchofo, P., Lopes, R., Vanhoutte,
M., Rasika, S., … Prevot, V. (2020). Hypothalamic structural and
functional imbalances in anorexia nervosa. Neuroendocrinology,
110(6), 552–562. https://doi.org/10.1159/000503147, PubMed:
31484186
Fox, M.. D., & Raichle, M.. E. (2007). Spontaneous fluctuations in
brain activity observed with functional magnetic resonance
imaging. Nature Revues Neurosciences, 8(9), 700–711. https://
doi.org/10.1038/nrn2201, PubMed: 17704812
Franssen, S., Jansen, UN., Schyns, G., van den Akker, K., & Roefs, UN.
(2020). Neural correlates of food cue exposure intervention for
obesity: A case-series approach. Frontiers in Behavioral Neuro-
science, 14, 46. https://doi.org/10.3389/fnbeh.2020.00046,
PubMed: 32372924
Friston, K. J., Harrison, L., & Penny, W. (2003). Dynamic causal
modelling. NeuroImage, 19(4), 1273–1302. https://est ce que je.org/10
.1016/S1053-8119(03)00202-7, PubMed: 12948688
Friston, K. J., Kahan, J., Biswal, B., & Razi, UN. (2014). A DCM for
resting state fMRI. NeuroImage, 94, 396–407. https://est ce que je.org/10
.1016/j.neuroimage.2013.12.009, PubMed: 24345387
Friston, K. J., Litvak, V., Oswal, UN., Razi, UN., Stephan, K. E., van
Wijk, B. C. M., … Zeidman, P.. (2016). Bayesian model reduction
and empirical Bayes for group (DCM) études. NeuroImage, 128,
413–431. https://doi.org/10.1016/j.neuroimage.2015.11.015,
PubMed: 26569570
Friston, K. J., Mattout, J., Trujillo-Barreto, N., Ashburner, J., &
Penny, W. (2007). Variational free energy and the Laplace
approximation. NeuroImage, 34(1), 220–234. https://est ce que je.org/10
.1016/j.neuroimage.2006.08.035, PubMed: 17055746
Gabriela Pop, M., Crivii, C., & Opincariu, je. (2018). Anatomy and
function of the hypothalamus. In S. J.. Baloyannis & J.. Oxholm
Gordeladze (Éd.), Hypothalamus health in and diseases.
Londres, ROYAUME-UNI: IntechOpen. https://doi.org/10.5772/intechopen
.80728
García-García, JE., Jurado, M.. Á., Garolera, M., Segura, B., Sala-
Llonch, R., Marqués-Iturria, JE., … Junqué, C. (2013). Alterations
of the salience network in obesity: A resting-state fMRI study:
Salience network and obesity. Human Brain Mapping, 34(11),
2786–2797. https://doi.org/10.1002/ hbm.22104, PubMed:
22522963
Goulden, N., Elliott, R., Suckling, J., Williams, S. R., Deakin,
J.. F. W., & McKie, S. (2012). Sample size estimation for compar-
ing parameters using dynamic causal modeling. Brain Connectiv-
ville, 2(2), 80–90. https://doi.org/10.1089/ brain.2011.0057,
PubMed: 22559836
Gutiérrez, R., Fonseca, E., & Simon, S. UN. (2020). The neuroscience
of sugars in taste, gut-reward, feeding circuits, and obesity. Cel-
lular and Molecular Life Sciences, 77(18), 3469–3502. https://est ce que je
.org/10.1007/s00018-020-03458-2, PubMed: 32006052
Han, W., Tellez, L. UN., Perkins, M.. H., Perez, je. O., Qu, T., Ferreira,
J., … de Araujo, je. E. (2018). A neural circuit for gut-induced
reward. Cell, 175(3), 665–678. https://doi.org/10.1016/j.cell
.2018.08.049, PubMed: 30245012
Hetherington, UN. W., & Ranson, S. W. (1983). Hypothalamic lesions
and adiposity in the rat. Nutrition Reviews, 41(4), 124–127.
https://doi.org/10.1111/j.1753-4887.1983.tb07169.x, PubMed:
6346152
Horst, G. J.. T., & Luiten, P.. G. M.. (1986). The projections of the
dorsomedial hypothalamic nucleus in the rat. Brain Research
Bulletin, 16(2), 231–248. https://doi.org/10.1016/0361-9230(86)
90038-9
Ilango, UN., Kesner, UN. J., Keller, K. L., Stuber, G. D., Bonci, UN., &
Ikemoto, S. (2014). Similar roles of substantia nigra and ventral
tegmental dopamine neurons in reward and aversion. Journal de
Neurosciences, 34(3), 817–822. https://est ce que je.org/10.1523
/JNEUROSCI.1703-13.2014, PubMed: 24431440
Jennings, J.. H., Rizzi, G., Stamatakis, UN. M., Ung, R.. L., & Stuber,
G. D. (2013). The inhibitory circuit architecture of the lateral
hypothalamus orchestrates feeding. Science, 341(6153),
Neurosciences en réseau
1331
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
1517–1521. https://doi.org/10.1126/science.1241812, PubMed:
24072922
Jennings, J.. H., Ung, R.. L., Resendez, S. L., Stamatakis, UN. M.,
Taylor, J.. G., Huang, J., … Stuber, G. D. (2015). Visualizing hypo-
thalamic network dynamics for appetitive and consummatory
behaviors. Cell, 160(3), 516–527. https://doi.org/10.1016/j.cell
.2014.12.026, PubMed: 25635459
Jung, K., Friston, K. J., Pae, C., Choi, H. H., Tak, S., Choi, Oui. K., …
Parc, H.-J. (2018). Effective connectivity during working memory and
resting states: A DCM study. NeuroImage, 169, 485–495. https://est ce que je
.org/10.1016/j.neuroimage.2017.12.067, PubMed: 29284140
Kass, R.. E., & Raftery, UN. E. (1995). Bayes factors. Journal of the
American Statistical Association, 90(430), 773–795. https://est ce que je
.org/10.1080/01621459.1995.10476572
Kiss, D. S., Toth, JE., Jocsak, G., Barany, Z., Bartha, T., Frenyo, L. V.,
… Zsarnovsyky, UN. (2020). Functional aspects of hypothalamic
asymmetry. Sciences du cerveau, 10(6), 389. https://doi.org/10.3390
/brainsci10060389, PubMed: 32575391
Kühnen, P., Krude, H., & Biebermann, H. (2019). Melanocortin-4
receptor signalling: Importance for weight regulation and obe-
sity treatment. Trends in Molecular Medicine, 25(2), 136–148.
https://doi.org/10.1016/j.molmed.2018.12.002, PubMed:
30642682
Kullmann, S., Heni, M., Linder, K., Zipfel, S., Häring, H.-U., Veit, R.,
Fritsche, UN., & Preissl, H. (2014). Resting-state functional connec-
tivity of the human hypothalamus: Hypothalamus functional
connectivity networks. Human Brain Mapping, 35(12),
6088–6096. https://doi.org/10.1002/ hbm.22607, PubMed:
25131690
Kullmann, S., Heni, M., Veit, R., Ketterer, C., Schick, F., Häring,
H.-U., Fritsche, UN., & Preissl, H. (2012). The obese brain: Asso-
ciation of body mass index and insulin sensitivity with resting
state network functional connectivity. Human Brain Mapping,
33(5), 1052–1061. https://doi.org/10.1002/ hbm.21268,
PubMed: 21520345
Kwon, H. G., & Jang, S. H. (2014). Differences in neural connectiv-
ity between the substantia nigra and ventral tegmental area in the
cerveau humain. Frontiers in Human Neuroscience, 8, 41. https://est ce que je
.org/10.3389/fnhum.2014.00041, PubMed: 24567711
Luiten, P.. G., Room, P., & Lohman, UN. H. (1980). Ependymal tany-
cytes projecting to the ventromedial hypothalamic nucleus as
demonstrated by retrograde and anterograde transport of HRP.
Brain Research, 193(2), 539–542. https://doi.org/10.1016/0006
-8993(80)90184-5, PubMed: 7388607
Martín-Pérez, C., Contreras-Rodríguez, O., Vilar-López, R., &
Verdejo-García, UN. (2019). Hypothalamic networks in adoles-
cents with excess weight: Stress-related connectivity and associ-
ations with emotional eating. Journal of the American Academy
of Child & Adolescent Psychiatry, 58(2), 211–220. https://doi.org
/10.1016/j.jaac.2018.06.039, PubMed: 30738548
Nieh, E. H., Vander Weele, C. M., Matthieu, G. UN., Presbrey, K. N.,
Wichmann, R., Leppla, C. UN., … Tye, K. M.. (2016). Inhibitory
input from the lateral hypothalamus to the ventral tegmental area
disinhibits dopamine neurons and promotes behavioral activa-
tion. Neurone, 90(6), 1286–1298. https://est ce que je.org/10.1016/j
.neuron.2016.04.035, PubMed: 27238864
Palmiter, R.. D. (2007). Is dopamine a physiologically relevant
mediator of feeding behavior? Trends in Neurosciences, 30(8),
375–381. https://doi.org/10.1016/j.tins.2007.06.004, PubMed:
17604133
Parc, H.-J., Friston, K. J., Pae, C., Parc, B., & Razi, UN. (2018).
Dynamic effective connectivity in resting state fMRI. Neuro-
Image, 180, 594–608. https://doi.org/10.1016/j.neuroimage
.2017.11.033, PubMed: 29158202
Penninx, B. W. J.. H., & Lange, S. M.. M.. (2018). Metabolic syndrome
in psychiatric patients: Overview, mechanisms, and implications.
Dialogues in Clinical Neuroscience, 20(1), 63–73. https://doi.org
/10.31887/DCNS.2018.20.1/bpenninx, PubMed: 29946213
Petrovich, G. D. (2018). Lateral hypothalamus as a motivation-
cognition interface in the control of feeding behavior. Frontiers
in Systems Neuroscience, 12, 14. https://doi.org/10.3389/fnsys
.2018.00014, PubMed: 29713268
Poldrack, R.. UN. (2007). Region of interest analysis for fMRI. Social
Cognitive and Affective Neuroscience, 2(1), 67–70. https://est ce que je
.org/10.1093/scan/nsm006, PubMed: 18985121
Preller, K. H., Razi, UN., Zeidman, P., Stämpfli, P., Friston, K. J., &
Vollenweider, F. X. (2019). Effective connectivity changes in
LSD-induced altered states of consciousness in humans. Proceed-
ings of the National Academy of Sciences, 116(7), 2743–2748.
https://doi.org/10.1073/pnas.1815129116, PubMed: 30692255
Razi, UN., Kahan, J., Rees, G., & Friston, K. J.. (2015). Construct val-
idation of a DCM for resting state fMRI. NeuroImage, 106, 1–14.
https://doi.org/10.1016/j.neuroimage.2014.11.027, PubMed:
25463471
Rolls, E. T., Huang, C.-C., Lin, C.-P., Feng, J., & Joliot, M.. (2020).
Automated anatomical labelling atlas 3. NeuroImage, 206,
116189. https://doi.org/10.1016/j.neuroimage.2019.116189,
PubMed: 31521825
Rossi, M.. UN., Basiri, M.. L., McHenry, J.. UN., Kosyk, O., Otis, J.. M.,
van den Munkhof, H. E., … Stuber, G. D. (2019). Obesity
remodels activity and transcriptional state of a lateral hypotha-
lamic brake on feeding. Science, 364(6447), 1271–1274.
https://doi.org/10.1126/science.aax1184, PubMed: 31249056
Rossi, M.. UN., & Stuber, G. D. (2018). Overlapping brain circuits for
homeostatic and hedonic feeding. Cell Metabolism, 27(1), 42–56.
https://doi.org/10.1016/j.cmet.2017.09.021, PubMed: 29107504
Seeley, W. W., Menon, V., Schatzberg, UN. F., Keller, J., Glover,
G. H., Kenna, H., … Greicius, M.. D. (2007). Dissociable intrinsic
connectivity networks for salience processing and executive
control. Journal des neurosciences, 27(9), 2349–2356. https://est ce que je
.org/10.1523/JNEUROSCI.5587-06.2007, PubMed: 17329432
Stevens, F. L., Hurley, R.. UN., & Taber, K. H. (2011). Anterior cingu-
late cortex: Unique role in cognition and emotion. Journal de
Neuropsychiatry and Clinical Neurosciences, 23(2), 121–125.
https://doi.org/10.1176/jnp.23.2.jnp121, PubMed: 21677237
Stuber, G. D., & Sage, R.. UN. (2016). Lateral hypothalamic circuits
for feeding and reward. Neurosciences naturelles, 19(2), 198–205.
https://doi.org/10.1038/nn.4220, PubMed: 26814589
Voigt, K. (2021). spDCM_Hypo, GitHub, https://github.com
/KatharinaVoigt1/spDCM_Hypo.git
Voigt, K., Murawski, C., Speer, S., & Bode, S. (2020). Effective brain
connectivity at rest is associated with choice-induced preference
formation. Human Brain Mapping, 41(11), 3077–3088. https://
doi.org/10.1002/hbm.24999, PubMed: 32243689
Voigt, K., Razi, UN., Harding, je. H., Andrés, Z. B., & Verdejo-
Garcia, UN. (2021). Neural network modelling reveals changes
Neurosciences en réseau
1332
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
t
/
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Hypothalamic effective connectivity
in directional connectivity between cortical and hypothalamic
regions with increased BMI. International Journal of Obesity,
45, 2447–2454. https://doi.org/10.1038/s41366-021-00918-y,
PubMed: 34341471
Zeidman, P., Jafarian, UN., Seghier, M.. L., Litvak, V., Cagnan, H.,
Prix, C. J., & Friston, K. (2019). A guide to group effective con-
nectivity analysis, part 2: Second level analysis with PEB.
NeuroImage, 200, 12–25. https://doi.org/10.1016/j.neuroimage
.2019.06.032, PubMed: 31226492
Zhang, S., Wang, W., Zhornitsky, S., & Li, C. R.. (2018). Resting state
functional connectivity of the lateral and medial hypothalamus
in cocaine dependence: An exploratory study. Frontiers in Psy-
chiatry, 9, 344. https://doi.org/10.3389/fpsyt.2018.00344,
PubMed: 30100886
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
3
1
6
2
0
5
9
7
5
6
n
e
n
_
un
_
0
0
2
6
6
p
d
.
t
/
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurosciences en réseau
1333