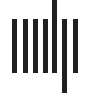RECHERCHE
Disruption of large-scale electrophysiological
networks in stroke patients with
visuospatial neglect
Tomas Ros1,2
, Abele Michela1, Anaïs Mayer3, Anne Bellmann3, Philippe Vuadens3,
Victorine Zermatten4, Arnaud Saj1,5, and Patrik Vuilleumier1
1Department of Neuroscience, University of Geneva, Genève, Suisse
2CIBM Center for Biomedical Imaging, Geneva University Hospitals, Genève, Suisse
3Romand Clinic of Readaptation, SUVA, Sion, Suisse
4Rehabilitation Clinic Valais de Coeur, Sion, Suisse
5Department of Neurology, Geneva University Hospital, Genève, Suisse
un accès ouvert
journal
Mots clés: Stroke, EEG, Hemineglect, Functional network connectivity, Alpha, sLORETA
ABSTRAIT
Stroke frequently produces attentional dysfunctions including symptoms of hemispatial neglect,
which is characterized by a breakdown of awareness for the contralesional hemispace. Recent
studies with functional MRI (IRMf) suggest that hemineglect patients display abnormal intra-
and interhemispheric functional connectivity. Cependant, since stroke is a vascular disorder
and fMRI signals remain sensitive to nonneuronal (c'est à dire., vascular) coupling, more direct
demonstrations of neural network dysfunction in hemispatial neglect are warranted. Ici, nous
utilize electroencephalogram (EEG) source imaging to uncover differences in resting-state
network organization between patients with right hemispheric stroke (N = 15) and age-
matched, healthy controls (N = 27), and determine the relationship between hemineglect
symptoms and brain network organization. We estimated intra- and interregional differences in
cortical communication by calculating the spectral power and amplitude envelope correlations
of narrow-band EEG oscillations. We first observed focal frequency-slowing within the right
posterior cortical regions, reflected in relative delta/theta power increases and alpha/beta/
gamma decreases. Secondly, nodes within the right temporal and parietal cortex consistently
displayed anomalous intra- and interhemispheric coupling, stronger in delta and gamma bands,
and weaker in theta, alpha, and beta bands. Enfin, a significant association was observed
between the severity of left-hemispace search deficits (par exemple., cancellation test omissions)
and reduced functional connectivity within the alpha and beta bands. In sum, our novel
results validate the hypothesis of large-scale cortical network disruption following stroke and
reinforce the proposal that abnormal brain oscillations may be intimately involved in the
pathophysiology of visuospatial neglect.
RÉSUMÉ DE L'AUTEUR
Stroke patients often exhibit a disabling deficit of visual awareness in the hemifield opposite to
their brain lesion, known as hemineglect. Recent studies with functional MRI (IRMf) suggérer
that hemineglect patients display abnormal functional coupling (c'est à dire., connectivité) within and
between brain hemispheres. Cependant, since stroke is a vascular disorder and fMRI measures
nonneuronal (c'est à dire., vascular) coupling, we here provide direct evidence of neural network
dysfunction in hemineglect by using electroencephalogram (EEG) source imaging, lequel
Citation: Ros, T., Michela, UN., Mayer, UN.,
Bellmann, UN., Vuadens, P., Zermatten,
V., Saj, UN., & Vuilleumier, P.. (2022).
Disruption of large-scale
electrophysiological networks in stroke
patients with visuospatial neglect.
Neurosciences en réseau, 6(1), 69–89.
https://doi.org/10.1162/netn_a_00210
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00210
Informations complémentaires:
https://doi.org/10.1162/netn_a_00210
Reçu: 12 Janvier 2021
Accepté: 17 Septembre 2021
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Tomas Ros
dr.t.ros@gmail.com
Éditeur de manipulation:
Olaf Sporns
droits d'auteur: © 2021
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
un
_
0
0
2
1
0
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
measures the electrical fluctuations of large neuronal populations. Dans l'ensemble, we observed a
breakdown of interhemispheric network connectivity within alpha/beta rhythms, lequel
specifically correlated with the degree of patients’ hemispatial errors. The high temporal
resolution and frequency content of EEG signals could lead to more sensitive markers and
targeted rehabilitation approaches of hemineglect.
INTRODUCTION
Apart from motor or sensory impairments, the sequelae of ischemic stroke may cause a signif-
icant impact on attentional function. This is most apparent when stroke damage leads to symp-
toms of hemispatial neglect (Domínguez-Borràs, Armony, Maravita, Driver, & Vuilleumier,
2013; P.. Ô. Vuilleumier & Rafal, 2000), which is characterized by an inability to attend to
and process information from the left (or more rarely right) side of space (c'est à dire., contralateral
to the lesion site). Ainsi, right-hemisphere stroke patients exhibit an impairment in detecting
visuel (or auditory) stimuli in their left hemifield (and vice versa for patients with left hemi-
sphere stroke). Despite the fact that this is a major source of disability in patients’ daily life,
current treatments for hemineglect remain minimally effective. De plus, untreated hemine-
glect leads to poorer recovery prognosis and reduced benefits from rehabilitation therapies for
other neurological deficits.
Donc, a deeper understanding of this clinical phenomenon is required for several in-
terdependent reasons: (un) to better identify the neurobiological targets for rehabilitation; (b) à
determine the underlying pathophysiological anomalies, in particular whether local and/or
distributed brain dysfunction is crucially implicated; et (c) to elucidate the neural mecha-
nisms that give rise to perceptual consciousness.
To address these issues, in line with recent work, here we investigate how brain activity dy-
namics is altered following focal hemispheric stroke, at both local and global levels, and what is
the functional impact of such changes on attentional performance in patients. To this aim, nous
use electroencephalogram (EEG) recording and adopt a network framework by reducing the
whole brain to a manageable number of regions of interest (ROI; nodes), whose functional
activity is considered to covary with each other through pairs-wise connections (edges).
Utilizing this approach, emerging work has suggested that hemineglect might be associated
with significant disruptions of brain functional connectivity. Spécifiquement, several functional
neuroimaging studies reported that the visuospatial deficits in hemineglect are accompanied
with abnormal interhemispheric and/or intrahemispheric connectivity (Baldassarre et al.,
2014; Fellrath, Mottaz, Schnider, Guggisberg, & Ptak, 2016; Guggisberg et al., 2014; Sasaki
et coll., 2013; Yordanova et al., 2016). Most recently, using functional MRI (IRMf), Ramsey
and colleagues showed that recovery from hemineglect is linked to the return of previously
depressed interhemispheric connectivity between nodes of sensorimotor and attention net-
travaux (Ramsey et al., 2016). Cependant, since stroke is a vascular disorder and fMRI signals re-
main sensitive to nonneuronal (c'est à dire., vascular) coupling, stronger validation of neural network
dysfunction in hemineglect is warranted. Physiologically speaking, fMRI measures local changes
in brain metabolism, which generally echo (but lag by a few seconds) electrophysiological
activation/deactivation patterns of neuronal populations (Bentley, Li, Snyder, Raichle, &
Snyder, 2016; Hermes, Nguyen, & Winawer, 2017; Hutchison, Hashemi, Gati, Menon, &
Everling, 2015). Fait intéressant, studies comparing noninvasive electrophysiological methods
Electroencephalogram (EEG):
A recording of brain activity where
non-invasive sensors are attached to
the scalp to detect the electrical
signals produced by neurons.
Neurosciences en réseau
70
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
un
_
0
0
2
1
0
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
such as magnetoencephalography/electroencephalography (M/EEG) with fMRI found a tight
spatiotemporal correspondence between the amplitude envelopes of neural oscillations and
neurovascular fMRI signals (Brookes et al., 2011; de Pasquale et al., 2010; Mantini, Perrucci,
Del Gratta, Romani, & Corbetta, 2007), although there are several instances of dissociations
between fMRI and neuronal signals (Schölvinck, Maier, Ye, Duyn, & Leopold, 2010).
En outre, abundant research in both human and nonhuman primates has linked specific
oscillatory activities with attentional functions in the normal brain, particularly implicating
modulations of the alpha range in relation to selective visuospatial orienting (Maréchal,
Bergmann, & Jensen, 2015b), but also the theta and beta ranges in relation to task-dependent
interactions across networks (Bauer et al., 2012; Fiebelkorn, Pinsk, & Kastner, 2018), ainsi que
the gamma band in relation to local intra-areal processing of sensory information (Szczepanski
et coll., 2014). Cependant, how such coordinated oscillatory activities are disrupted by stroke and
affected by clinical neglect deficits still remain unresolved.
Given the linkage between M/EEG and fMRI signals and their relation to attentional pro-
cesses, the goal of our current study was to investigate the neural substrates of hemineglect
using source space EEG, which reflects genuine neuroelectric activity uncontaminated by vas-
cular dynamics. Spécifiquement, we performed (un) functional whole-brain connectivity compari-
sons between stroke patients and healthy controls, et (b) regression analyses to identify
changes connectivity patterns more selectively associated with visuospatial deficits (c'est à dire., par-
ceptual omissions during search tasks) in the left and right hemifields, respectivement.
RÉSULTATS
Group Functional Connectivity in Patients Compared to Healthy Elderly Controls
We defined the EEG source space functional network connectivity (FNC) graphs for both the
stroke patients (n = 15) and the age-matched healthy controls (n = 27), and then contrasted the
graphs from each group in order to test for changes in connectivity patterns across the different
frequency bands. As illustrated in Figure 1, this revealed three main patterns in the patients
relative to controls: (un) the delta band showed a mixed pattern of both increased and
decreased FNC; (b) the theta-, alpha-, and beta- bands all displayed decreased FNC, but with
distinctive anatomical distributions; et (c) the gamma band demonstrated selective and
localized increases in FNC.
More specifically, using a network-based statistic (NBS) correction threshold of p < 0.05,
we found the delta band demonstrated significant but limited changes with mainly intrahemi-
spheric hyperconnectivity, predominating between the right paracentral lobule (BA4) and oc-
cipital areas (BA18) (d = 0.063, t = 3.28; d = −0.063, t = 3.58, for sources in right and left
cuneus, respectively), but also a more limited interhemispheric hypoconnectivity between the
right lateral occipital cortex (BA18) and left inferior prefrontal cortex/pars triangularis (BA45)
nodes (d = −0.064, t = −3.36).
For the theta band, we observed mainly long-range, intrahemispheric hypoconnectivity
changes along the anterior-posterior axis that, remarkably, were present on both sides.
These decreases were the largest between the left pericalcarine (BA17) and left inferior pre-
frontal gyrus/pars orbitalis (BA47) nodes (d = −0.079, t = −4.51), as well as between the right
pericalcarine (BA17) and right frontal pole (BA11) nodes (d = −0.081, t = −4.15).
In sharp contrast, the beta band showed an opposite pattern of hypoconnectivity, mainly
affecting interhemispheric connections, particularly those centered on posterior right-
hemisphere (RH) areas. These interhemispheric changes affected connectivity between
Functional network
connectivity (FNC):
A general term for the degree of
functional coupling between brain
regions, here measured by the
amplitude envelope correlation
(AEC).
Network-based statistic (NBS):
A statistical-correction method for
comparing network connectivity
matrices based on the largest
connected components.
Hyperconnectivity:
An increase in the amplitude
envelope correlation (AEC) between
brain regions.
Hypoconnectivity:
A decrease in the amplitude
envelope correlation (AEC) between
brain regions.
Network Neuroscience
71
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Group differences in EEG source functional connectivity. Significant differences of the
magnitude of amplitude envelope correlations between stroke patients and controls. Line thickness
represents larger absolute connectivity difference (Pearson’s correlation coefficient r). Red color in-
dicates greater connectivity for patients versus controls, while blue color indicates greater connectiv-
ity for controls versus patients (only significant connectivity components are displayed, p < 0.05
network-based statistic corrected).
homologous areas of the superior parietal and temporal cortices. The greatest reductions in
connectivity were found for the right hemisphere, between the right superior parietal lobule
(BA7) and left posterior cingulate gyrus (d = −0.051, t = −5.59), as well as between the right
middle temporal (BA20) and right precuneus (BA3) nodes (d = –0.041, t = −4.93).
Alpha-band hypoconnectivity was characterized by a more focal cluster of reduced con-
nections emerging from the posterior right hemisphere. The most salient interhemispheric dis-
connectivities were observed between right posterior temporal (BA20) and left superior
parietal (BA7) nodes (d = −0.130, t = −4.65), as well as between right inferior parietal
(BA39) and left superior parietal (BA7) nodes (d = −0.126, t = −5.32).
Finally, the gamma band displayed a local hyperconnectivity pattern predominating mainly
within the anterior right hemisphere. These difference were maximal between the right pre-
central (BA6) and right superior frontal (BA32) gyri (d = 0.065, t = 2.50), between the right
precentral (BA6) and right inferior temporal (BA20) gyri (d = 0.064, t = 2.30), as well as be-
tween the right precentral (BA6) and right frontal pole (BA10) (d = 0.062, t = 2.35).
Relationships Between Network Connectivity and Visuospatial Bias in Hemineglect Patients
Although group-level analyses indicated anomalous FNC between stroke patients and control
subjects, they do not disambiguate which network connections, if any, may be related to the
Network Neuroscience
72
EEG network correlates of visuospatial neglect
emergence of visuospatial deficits. Many of our patients presented signs of hemispatial ne-
glect, as commonly observed in right brain–damaged patients (Verdon et al., 2010), but to
a varying degree. The FNC changes described above could more generally reflect the effect
of posterior stroke damage. Hence, in order to disentangle neurobehaviorally specific from
nonspecific connections in stroke-associated hemineglect, we performed regression analyses
directly testing for any relation between individual patients’ FNC matrices and the severity of
neglect symptoms, as indexed by the number of left/right omission errors in the visual cancel-
lation test. These errors are considered as one of the most reliable markers of neglect (Klinke,
Hjaltason, & Tryggvadóttir, 2018).
Using a NBS correction threshold of p < 0.05, we found significant brain–behavior relation-
ships that distinctively affected the alpha, beta, and gamma bands. Changes in the delta and
theta bands did not predict the severity of visuospatial neglect.
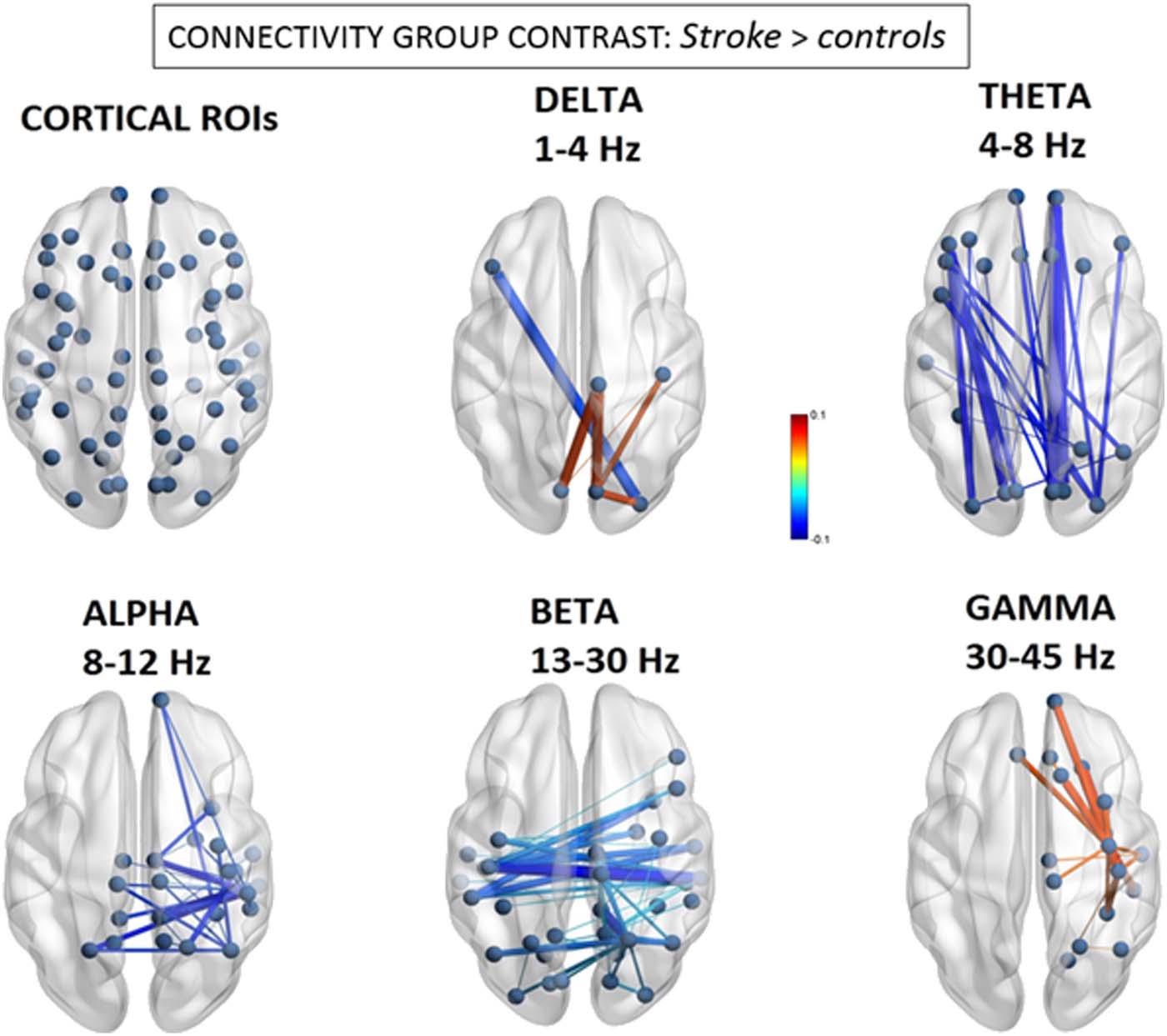
As seen in Figure 2, the alpha band demonstrated a remarkable contrast between omissions
in left and right space, implicating near mirror image interhemispheric connections. Consistent
with the group-level hypoconnectivity described above, the number of leftward misses was
associated with selective decreases in connectivity between extrastriate visual regions in the
right inferior temporal gyrus (BA20) with the left precuneus (BA3) (beta = −0.99) and left peri-
calcarine region (BA17) (beta = −0.98). Conversely, the number of misses within the right
hemifield (typically reflecting more extensive spatial neglect) was significantly predicted by
increases in alpha-band connectivity within a homologous set of visual and parietal nodes,
which included the left inferior temporal cortex (BA20) and the left precuneus (BA3).
Visuospatial neglect:
Cognitive deficit characterized by
reduction or loss of spatial awareness
for the contralesional space, most
commonly after unilateral stroke or
brain injury.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Alpha (8–12 Hz) connectivity as a function of neglect symptoms. Depicted connections
correspond to changes in individual FNC components correlating with the number of omissions in
the left and right hemifields, respectively, during the cancellation task. Red/blue values indicate
statistically significant beta coefficients ( p < 0.05 network-based statistic corrected). Further con-
junction analysis (see text) revealed that reduced connections associated with left omissions (in left
panel) were those showing the most significant overlap with general stroke-related decreases at the
group level.
Network Neuroscience
73
EEG network correlates of visuospatial neglect
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
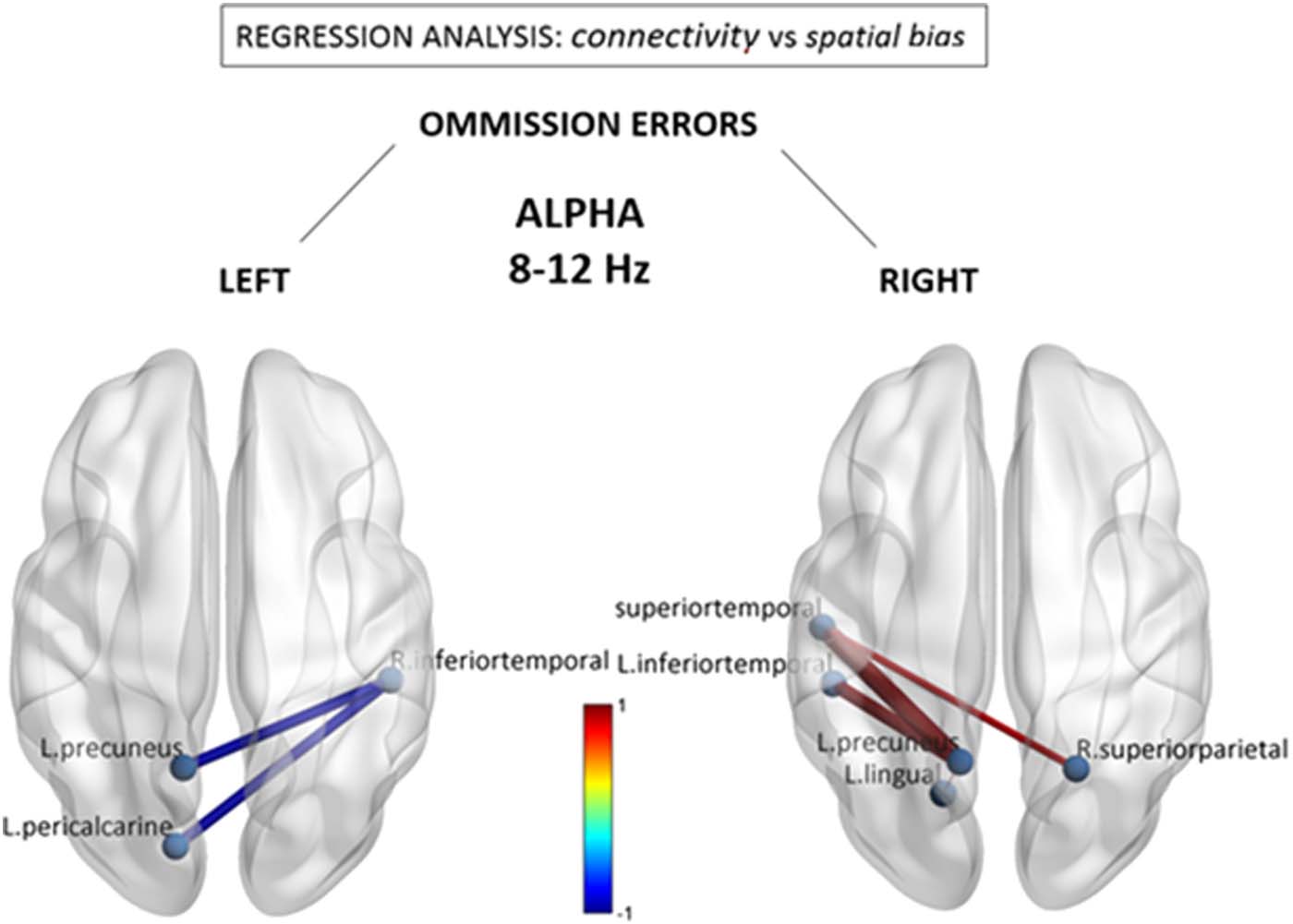
Figure 3. Beta (13–30 Hz) connectivity as a function of neglect symptoms. Depicted connections
correspond to changes in individual FNC components correlating with the number of omissions in
the left and right hemifields, respectively, during the cancellation task. Red/blue values indicate
statistically significant beta coefficients ( p < 0.05 network-based statistic corrected). R. bankssts =
right bank of the superior temporal sulcus.
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Importantly, a separate regression with the total number of left and right misses (see Supporting
Information Figure S2) did not reveal any shared connections with either left or right omissions,
but limited reductions in connectivity between left medial parietal and right inferior frontal
areas, reinforcing the idea that the changes in alpha band may be specifically related to spa-
tially directed attentional processing.
Figure 3 shows results from similar regression analyses for the beta band, demonstrating an
association of leftward omissions with a different pattern of reduced functional connections
between the right posterior-superior temporal region (BA22) and anterior brain regions in
the left middle frontal gyrus (BA6) (beta = −0.67) and left inferior frontal gyrus/pars orbitalis
(BA47) (beta = −0.70). Reductions were also seen within the anterior left hemisphere. No sig-
nificant associations were observed for the number of rightward errors, nor were there any
systematic functional connectivity changes that regressed with the total number of left and
right omissions (see Supporting Information Figure S3).
Lastly, as can be seen in Figure 4, the relationship between leftward omissions and FNC
within the gamma band revealed a node also implicated within the beta band, namely, the
right posterior-superior temporal region (BA22) that acted as a hub for reduced connections
with mirror areas in left temporal regions. Also similar to the beta band, additional reductions
were seen within the anterior left hemisphere. However, surprisingly, these negative changes
contrasted with the overall increases in connectivity observed for the gamma band at the
Network Neuroscience
74
EEG network correlates of visuospatial neglect
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
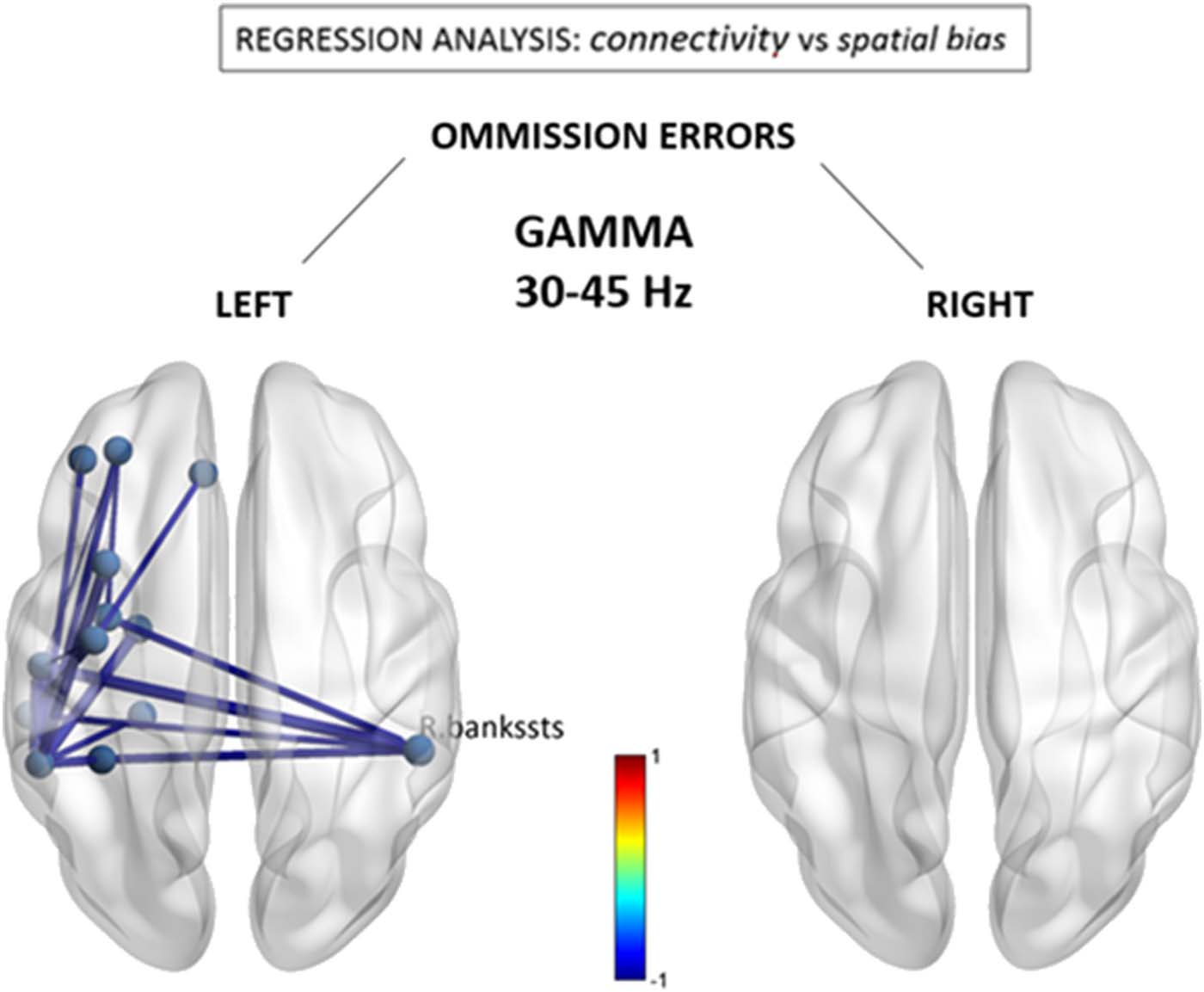
Figure 4. Gamma (30–40 Hz) connectivity as a function of neglect symptoms. Depicted connec-
tions correspond to changes in individual FNC components correlating with the number of omis-
sions in the left and right hemifields, respectively, during the cancellation task. Red/blue values
indicate statistically significant beta coefficients ( p < 0.05 network-based statistic corrected). R.
bankssts = right bank of the superior temporal sulcus.
group level (see Figure 1). There was no correlation between connectivity patterns in gamma
band and omissions in right space.
Conjunction Analysis Between Group Functional Connectivity Abnormalities and Correlates of
Visuospatial Errors
Finally, we asked whether the abnormal functional components seen as a direct marker of
stroke at the group level are consistent with those that interindividually predict stronger left-
ward visuospatial deficits. In other words, is there evidence for a distinctive disruption of FNC
components associated with clinical symptoms of hemineglect? This question was addressed
by using a standard conjunction analysis, with the aim to identify the overlapping or “in com-
mon” components that were significant in the group-wise difference test and in the behavioral
regression test. Accordingly, a correct test for a logical AND requires that all the comparisons
in the conjunction are individually significant at p < 0.05 (Nichols et al., 2005). Hence we
performed a conjunction of tests (i.e., global null hypothesis) to identify the overlap—if any—
between only the statistically significant ( p < 0.05 corrected) group-level FNC difference maps
(i.e., Figure 1) and the counterpart visuospatial regression maps (i.e., Figures 2 and 3), across
the statistically significant alpha, beta, and gamma bands.
Interestingly, among all abnormal connections, we identified a single overlap with a net-
work component that includes a node in the right inferior temporal gyrus (BA20, MNI coor-
dinates X = 49, Y = −31, Z = −23) within the alpha band. This network component was
Network Neuroscience
75
EEG network correlates of visuospatial neglect
interhemispherically disconnected with the left precuneus (BA3) and left pericalcarine region
(BA17), identical to the connections illustrated in the left panel of Figure 2. This suggests that
among connections globally disrupted by right hemispheric stroke, these may be the most re-
liable marker of left hemineglect symptoms.
Group Spectral Power in Patients Compared to Healthy Elderly Controls
In addition to interregional FNC measures, regional source spectral power
Absolute power.
(i.e., current source density) differences were estimated between stroke patients and controls
using independent t tests across the five EEG bands (false discovery rate [FDR] corrected).
Absolute power was consistently more elevated in the right (damaged) hemisphere of patients
relative to controls for all EEG bands (delta, theta, beta, gamma) except for alpha, where no
significant differences were detected ( p < 0.05 corrected). As seen in Supporting Information
Figure S4, absolute delta power was stronger within posterior parietal areas (t = 3.1, p < 0.05
corrected), theta power within widespread regions with a maximum over supramarginal re-
gions (t = 3.8, p < 0.05 corrected), while beta was higher mostly over frontal (t = 2.7, p <
0.05 corrected), and gamma mostly over precentral motor regions (t = 3.8, p < 0.05 corrected).
Supporting Information Figure S6 illustrates the absolute power spectral density of hemineglect
and control groups at the sensor level (using a current source density montage at electrode P4,
over the right parietal cortex). Interestingly, no significant associations were detected between
absolute EEG power changes and visuospatial performance ( p < 0.05 corrected).
Relative power. We also tested for differences in relative (%) power, often used to normalize
spectra under a constant value of broadband (1–40 Hz) power, and reflecting the degree of
spectral slowing (i.e., greater relative power in lower frequencies) or spectral acceleration
(i.e., greater relative power in higher frequencies). As evidenced by Supporting Information
Figure S5, neglect was associated with relative spectral slowing of right posterior cortical
regions, with relative delta power being more elevated in patients mainly within the inferior
parietal/posterior temporal junction (t = 3.6, p < 0.05 corrected) while relative theta power was
higher within the superior parietal cortex (t = 3.4, p < 0.05 corrected). Conversely, relative
alpha power was maximally reduced within the inferior parietal/posterior temporal junction
(t = −3.6, p < 0.05 corrected), in conjunction with relative beta (t = −4.4, p < 0.05 corrected)
and relative gamma (t = −3.2, p < 0.05 FDR corrected) reductions within the superior parietal
lobe. Supporting Information Figure S7 illustrates the relative power spectral density of
hemineglect and control groups at the sensor level (using a current source density montage
at electrode P4, over the right parietal cortex). No significant associations were detected
between relative EEG power changes and visuospatial performance ( p < 0.05 corrected).
Relating Connectivity and Oscillatory Power Changes With Lesion Topology
Given that changes in functional connectivity (FC) and source power were topographically
specific, a reasonable question to ask is whether connectivity differences could be explained
firstly by changes in signal power? According to this hypothesis, reduced connectivity would
indirectly reflect a degraded signal-to-noise ratio caused by a loss of source EEG power. The
latter could result in particular from neuronal loss at the site of the stroke lesion. However,
additional analyses allowed us to rule out this hypothesis, given the rather limited overlap be-
tween anatomical regions with maximal lesion extent and those with strongest disruptions of
connectivity. As shown in Supporting Information Figure S8, the location of largest lesion over-
lap occurred in fronto-insular cortex (which was common to no more than 10 patients), en-
compassing regions such as BA44, BA45, and BA6. This co-localized with 5 RH nodes/parcels
Network Neuroscience
76
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
of the Desikan–Killiani atlas used for FNC calculations: entorhinal, insula, temporal pole, su-
perior temporal, and precentral regions (see Supporting Information Figure S4 for their exact
locations in MNI space). Importantly, removal of these particular nodes (or their connections)
from our analysis did not alter our principal findings, which highlight alpha/beta disconnec-
tivity with more posterior temporo-parietal foci (see Supporting Information Figure S4 for an-
atomical verification).
On the other hand, although regions with significant relative power differences (indicative
of relative frequency shifts in EEG power) did partly overlap with temporo-parietal nodes pre-
senting abnormal FC power in alpha/beta bands (see Supporting Information Figure S1), this
was clearly not the case for absolute source power (see Supporting Information Figure S2),
which provides a more direct measure of electrical signal strength. Notably, each narrow-band
EEG frequency range (from delta to gamma) exhibited a unique topographical fingerprint (e.g.,
see Figure 2), arguing against a simple generative mechanism of reduced EEG signal-to-noise
ratio due to neuronal loss that would presumably lead to a broadband attenuation of electrical
activity across the full EEG frequency spectrum.
Instead, and given the partial FC overlap with nodal reductions in relative alpha/beta power,
we suggest that this pair of EEG rhythms may act as carrier waves for attentional information
(Wróbel, 2000) compared to other frequency channels (e.g., delta, theta, gamma), and that it is
their relative degradation compared to other frequencies that may be an index of impaired
network communication (Akam & Kullmann, 2012).
DISCUSSION
Overall, our EEG results converge with previous M/EEG studies reporting electrocortical abnor-
malities in stroke patients with hemineglect (Colson, Demeurisse, Hublet, & Slachmuylder,
2001; Demeurisse, Hublet, & Paternot, 1998; Ros et al., 2017; Yordanova et al., 2016) and
add novel neurophysiological evidence to support recent findings of neurometabolic (i.e.,
fMRI) alterations in functional brain networks associated with specific cognitive deficits after
stroke (Siegel et al., 2016). Remarkably, we found widespread changes of FNC extending far
beyond focal areas of structural brain lesions in RH, including connections with the
opposite/intact left hemisphere (LH), with very distinctive patterns across different frequency
bands. Moreover, a conjunction analysis that sought to overlap (a) group-level FC differences
between patients and controls, together with (b) connections predicting the magnitude of left-
ward visuospatial inattention in a regression analysis, revealed a circumscribed reduction of in-
terhemispheric FC between the posterior right temporal lobe and left precuneal/pericalcarine
regions. This is consistent with and supportive of anatomical lesion studies that have long im-
plicated right posterior temporal and inferior parietal areas as a critical site associated with vi-
suospatial attention deficits (Chelazzi, Duncan, Miller, & Desimone, 1998; Heilman & Van Den
Abell, 1980; Hillis et al., 2005; Karnath, Ferber, & Himmelbach, 2001; Mort et al., 2003; Ptak &
Valenza, 2005; Vallar & Perani, 1986), and with the notion of an impaired interhemispheric
balance in the pathological spatial biases associated with left hemineglect (Corbetta &
Shulman, 2011; Kinsbourne, 1977; Mesulam, 1981). Importantly, the posterior temporal cortex
has previously been associated with mechanisms of visuospatial awareness in both human and
animal studies and found to be among the most frequent sites of brain damage in hemineglect
(Karnath, Ferber, & Himmelbach, 2001). Finally, a separate regression with the total number of
(left and right) errors did not reveal any overlap with connections specifically regressing with
either left or right omissions, reinforcing the idea that the changes in alpha-band connectivity
may be specifically related to spatially directed attentional processing.
Network Neuroscience
77
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
Taken together, our results therefore reinforce the theory that network disturbances charac-
terized by a weakened functional communication between key cortical regions is a fundamen-
tal mechanism underlying spatial neglect (Doricchi, Thiebaut de Schotten, Tomaiuolo, &
Bartolomeo, 2008; Driver & Vuilleumier, 2001; P. Vuilleumier et al., 2001, 2008), including
both interhemispheric and intrahemispheric disconnectivity.
The Multiplex FNC Signature of Hemineglect
Compared to age-matched healthy subjects, our source space analyses of amplitude envelope
correlations indicated that patients with RH stroke displayed a multiplex reorganization of neu-
ral connectivity that spans the full frequency range captured by EEG, involving delta, theta, al-
pha, beta, and gamma rhythms, and that covers both hemispheres in a frequency-specific
manner. Specifically, delta FC was mainly elevated within posterior RH regions, while it was
reduced between the posterior RH and anterior LH. These changes are consistent with nonspe-
cific effects of brain lesions on focal delta activity (van Dellen et al., 2013) and they did not cor-
relate with neglect symptoms. On the other hand, theta connectivity mainly exhibited a bilateral
reduction of long-range intrahemispheric connections, partially consistent with recent work
reporting reductions in fronto-parietal theta and beta coherence in other EEG studies of stroke
patients with hemineglect (Fellrath et al., 2016; Yordanova et al., 2017). However, we found
that the theta connectivity difference did not correlate directly with neglect severity, suggesting
that previously reported changes in this band may reflect task-specific abnormalities during goal
directed attention or nonspecific effects of stroke on executive aspects of attention unrelated
to spatial neglect (see Fellrath et al., 2016).
In contrast, we found that alpha and beta rhythms both demonstrated important reductions in
intra- and interhemispheric connectivity of posterior temporal and parietal cortices, that is, re-
gions traditionally found to be implicated in hemineglect (Thimm, Fink, & Sturm, 2008;
Umarova et al. 2011; Verdon et al., 2010). Furthermore, a subset of these connections was sig-
nificantly associated with neglect symptoms, in particular those linking right temporo-parietal
areas with left posterior parietal regions in the alpha band and with left prefrontal regions in the
beta band (see Figures 2 and 3). Remarkably, changes in alpha-band connections significantly
correlated with the severity of visuospatial biases and implicated symmetrical regions over the
extrastriate visual cortex (pericalcarine and lingual), where hypoconnectivity with the LH pre-
dicted more left visual field omissions, while the converse (hypoconnectivity with the right hemi-
sphere) was observed for right visual field omissions. This topographically specific
disconnection pattern is compatible with the role of alpha rhythms in contralateral visuospatial
attention (Ikkai, Dandekar, & Curtis, 2016; Lobier, Palva, & Palva, 2017; Marshall, Bergmann, &
Jensen, 2015a; Okazaki et al., 2015; van Schouwenburg, Zanto, & Gazzaley, 2017), as well as
previous studies on hemineglect utilizing functional (Ramsey et al., 2016; Sasaki et al., 2013)
and structural (Karnath, Rennig, Johannsen, & Rorden, 2011; Vaessen et al., 2016) neuroimaging
to delineate the functional neuroanatomy of visuospatial deficits. In particular, it is thought that
lateralized modulation of alpha activity across hemispheres might gate sensory information flow
from early visual areas to the ventral temporo-occipital stream, under top-down influences from
the dorsal attention network (Geng & Mangun, 2009; Liu et al., 2014; van Diepen, Miller,
Mazaheri, & Geng, 2016). Moreover, it is interesting to note that while the alpha band was im-
plicated in the disconnectivity pattern of the posterior right temporal lobe with sensory visual
areas, the beta band was distinctively involved in disconnectivity with left prefrontal areas,
which presumably subserve more executive processes of attention (Antzoulatos & Miller, 2016).
Importantly, a selective depression of interhemispheric connectivity has in itself been
shown to be a pathological hallmark of hemineglect in both fMRI and EEG studies (Ramsey
Network Neuroscience
78
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
et al., 2016; Sasaki et al., 2013), including for the alpha (Sasaki et al., 2013) and beta bands
(Guggisberg et al., 2014). Furthermore, increased alpha and/or beta FC has been reported to be
a predictive marker of clinical status after stroke or traumatic brain injury in patients with
motor (Dubovik et al., 2012; Guggisberg et al., 2014; Kawano et al., 2017) as well as language
(Castellanos et al., 2010; Guggisberg et al., 2014; Nicolo et al., 2015) impairments.
Finally, we also found connectivity changes in the gamma range, including increases within
right frontal areas at the whole group level, as well as decreases between right temporo-parietal
and LH regions that correlated with neglect symptoms and partly overlapped with beta changes.
These divergent effects make it difficult to associate them with a clear functional role in
spatial deficits after stroke. Moreover, gamma-range neuronal activity is usually associated with
local interactions between nearby cortical populations rather than with long-distance interac-
tions at network level (Vinck, Womelsdorf, Buffalo, Desimone, & Fries, 2013). Given theoret-
ical proposals linking gamma activity to conscious perceptual processes (Melloni et al., 2007),
one tentative hypothesis might be that a reduction of synchronous activity in gamma band be-
tween right posterior brain areas and left fronto-temporal networks would reflect the loss of
access of spatial representations held in the right hemisphere (Karnath et al., 2001) to LH pro-
cesses mediating conscious awareness and verbal report, in line with work in split-brain pa-
tients demonstrating an intimate link between conscious behavior and language abilities of
the LH (Volz & Gazzaniga, 2017). Furthermore, through phase–amplitude coupling, gamma-
band activity has recently been demonstrated to be modulated by both alpha and theta rhythms
(observed to be disrupted above) in the service of spatial attention (Fiebelkorn et al., 2018;
Jensen, Gips, Bergmann, & Bonnefond, 2014). Hence, it is possible that hemineglect constitutes
a clinical syndrome resulting from a concerted disruption of a family of nested rhythms
(Bonnefond, Kastner, & Jensen, 2017), rather than a single one. Within this theoretical frame-
work, communication between two regions can be established by phase synchronization of
oscillations at lower frequencies (i.e., theta, alpha, beta, <25 Hz), which serve as temporal ref-
erence frame for information carried by high-frequency activity (i.e., gamma,>40 Hz)
(Szczepanski et al., 2014). Cependant, this interpretation still remains speculative and the role
of phase–amplitude coupling in neglect, if any, remains unsettled and undoubtedly needs fur-
ther investigation to be clarified. Enfin, given the lack of significant overlap between rightward
omissions (which are specific markers of hemineglect) and group-level connectivity differences
in the beta and gamma bands, it is equally possible that such reorganization may be the result of
naturalistic recovery of cortical function that has occurred since the time of the stroke.
Limitations
The first important limitation of our study is that it was not preregistered and that it lacked a
patient control group with stroke but without presentation of hemineglect symptoms. Ce
would have had the benefit of additionally controlling for a general and deleterious effect
of stroke itself on brain activity. We acknowledge this as an important shortcoming in the strict
context of our primary analysis of group-wise differences. Cependant, beyond this comparison
in the first step of our analysis, the current study was able to leverage hemineglect-specific
behavior with the second, most important step of analysis (c'est à dire., regression), as well as with
its functional intersection with the group-wise stroke-related difference (c'est à dire., conjunction).
This double-step approach allowed us to exploit the full sample of patients and circumvent
difficulties associated with a dichotomous diagnosis of neglect (Checketts et al., 2020).
Ainsi, despite the lack of separate patient subgroups, we are confident that the network com-
ponent(s) that ultimately survived this sequential statistical selection are not only robust but
also clinically relevant. Cependant, in light of the limited number of participants in our study,
Neurosciences en réseau
79
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
un
_
0
0
2
1
0
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
this does not exclude a potential region-of-interest bias due to the pooling of different anatom-
ical lesion locations within this sample of stroke patients.
Secondly, as already noted above, the connectivity values may have been contaminated by
degraded signal-to-noise ratio and global losses in source EEG power due to neuronal tissue
dommage. Cependant, as shown by our supplementary analyses, we consider this interpretation
unlikely for several reasons, most notably because connectivity changes included both de-
creases and increases, they did not correspond to the local peaks of absolute source power
that constitutes a more direct measure of electrical signal strength, and distinct patterns were
observed for each EEG frequency sub-band. Taken together, this argues against a single, mondial
mechanism of reduced EEG signal-to-noise ratio due to neuronal loss. Plutôt, and given the
partial FC overlap with nodal reductions in relative alpha/beta power, we suggest that this pair
of EEG rhythms may act as major carrier waves for attentional information (Wróbel, 2000),
distinct from other frequency channels (par exemple., delta, theta, gamma), and hence it is their relative
degradation compared to other frequencies that may constitute a functional index of impaired
network communication (Akam & Kullmann, 2012).
Other potential limitations are reflective of the imaging modality we used, c'est, EEG.
Although EEG provides a direct measure of neural activity, it is most sensitive to sources within
the cortical mantle. Ainsi, our analyses were restricted to cortical network dynamics and did
not allow for reliable assessment of subcortical structures that may play an important role in the
control of attention (Gitelman et al., 1999; P.. Vuilleumier, 2013). Another weakness involves the
relatively low number of EEG channels that constitute the 10–20 montage, raising potential con-
cerns of excessive source localization error and/or spread, which would lead to spurious activity
mixing between cortical ROIs. Ici, given the clinical nature of our study, we found that the 10–
20 montage met an important trade-off between EEG setup time and patient fatigue during daily
rehabilitation sessions. Secondly, our methodology allows for replication and application in
standard clinical settings where high-density EEG is not routinely available. In this context, un
number of studies have shown that low-density montages may be used to reconstruct a consid-
erably greater number of dipolar sources than there are sensors (c'est à dire., électrodes) (for a review see
Grech et al., 2008). En particulier, a source localization simulation study with low-resolution
electromagnetic tomography analysis (sLORETA) en utilisant 1,000 cortical sources (c'est à dire., patches)
of ~1 cm2 (Song et al., 2015) and an electrode montage based on the 10–20 system indicated
a mean localization accuracy of ~0.5 cm and source spread of ~0.7 cm, which remains within
the lowest inter-ROI distances of the Desikan–Killiany atlas (~1.5 cm) used in the present study.
Surtout, as the Desikan–Killiany atlas contains ROIs of variable patch sizes, empirical work
has not revealed a systematic bias between patch size and sLORETA reconstruction accuracy
(Cosandier-Rimélé, Ramantani, Zentner, Schulze-Bonhage, & Dümpelmann, 2017). Dernièrement, dans
line with published work leveraging a similar low-density EEG montage in clinical conditions
(Tokariev et al., 2018), our analysis pipeline included EEG signal orthogonalization prior to FC
estimation, which has been shown to minimize any instantaneous (c'est à dire., at zero-phase lag) activ-
ity that is spuriously shared between ROIs and that may have arisen from (low-resolution) source
blurring (Brookes et al., 2011). Nevertheless, as source localization errors cannot entirely be
excluded, caution must be exercised regarding the precise anatomical locations of cortical net-
work nodes, given the use of a generic head template and low-density montage in this study.
Conclusion
In a nutshell, our results show that the hemineglect syndrome following RH stroke is associ-
ated with a widely distributed but anatomically specific disruption of cortical network connec-
tivity that involves temporo-occipital regions of the right hemisphere and their functional
Low-resolution electromagnetic
tomography analysis (sLORETA):
A method of mathematically
analyzing multiple EEG signals from
across the scalp to determine their
anatomical source within the brain.
Neurosciences en réseau
80
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
un
_
0
0
2
1
0
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
interactions both within and between the two hemispheres. En particulier, losses in the connec-
tivity of the posterior RH with left parieto-occipital cortex in alpha channels and with left pre-
frontal cortex in beta channels appear to be critically involved in impaired control of
visuospatial attention toward the contralesional/left hemispace. Better mapping and under-
standing of these neurophysiological markers of hemineglect is an important step toward de-
signing novel tools to assess poststroke deficits in patients and more effective rehabilitation
approaches. Par exemple, by directly training alpha oscillations by using neurofeedback
(Mottaz et al., 2018; Ros et al., 2017) or transcranial alternating stimulation (also known as
tACS) (van Schouwenburg, Sörensen, de Klerk, Reteig, & Slagter, 2018).
MATERIALS AND METHODS
Study Participants
Stroke patients participated after giving their written informed consent. The study was ap-
proved by Geneva State Ethics Committee and accorded with the Helsinki Declaration.
Patients were admitted after a first RH stroke and consecutively recruited from a primary clin-
ical center. We excluded patients with bilateral lesions, previous neurological or psychiatric
disorders, impairment in primary visual perception (except partial visual field defect), psychi-
atric disorders, severe motor difficulties in the right upper limb, pusher syndrome (c'est à dire., contra-
lateral trunk deviation with active resistance to any attempt of external correction), or current
psychotropic treatment. In total, we recruited 15 right-hemisphere-lesioned patients (mean
âge: 59.4; SD: 11.3; 2 femmes, 13 men) who fulfilled these criteria and were prospectively
admitted to two primary clinical centers at the Clinique de Réadaptation of SUVA in Sion
(https://www.crr-suva.ch) and the Foundation Valais de Coeur of Sion and Sierre (https://
www.valaisdecoeur.ch).
Spatial neglect was assessed using a standard clinical battery similar to other research in our
group (Saj, Cojan, Vocat, Luauté, & Vuilleumier, 2013; Vaessen, Saj, Lovblad, Gschwind, &
Vuilleumier, 2016). Details on clinical tests are provided below. Time since stroke onset was
7.5 months on average (SD 5.3 mois). Hemianopia or quadranopia was present in 4 out of
15 patients (26%).
All patients underwent structural MRI scans to delineate the location and extent of brain
dommage. Individual stroke lesions were manually delineated and the group average lesion
map was created using the MRICron toolbox (https://people.cas.sc.edu/rorden/mricron). Le
maximal lesion overlap affected posterior parts of the lateral prefrontal cortex, the anterior
and middle temporal lobe, as we all as the deep paraventricular white matter in the right hemi-
sphère (see Supporting Information Figure S1).
As we sought to explore electrophysiological differences between stroke patients and the
healthy population, we also collected data from a control group of 27 matched healthy adults
(mean age: 56; SD: 7; males: 23, females: 4), randomly sampled from the Human Brain
Institut (HBI) normative database (https://www.hbimed.com/) (Grin-Yatsenko, Baas,
Ponomarev, & Kropotov, 2009). Surtout, the healthy subject in the HBI data were collected
using the same EEG amplifier (Mitsar-201) and EEG montage (10–20 international montage) comme
the recordings in stroke patients.
Clinical Battery for Visuospatial Neglect
A series of standard paper-and-pencil tasks were presented to each patient at their first visit, dans
order to determine the presence and severity of baseline visuospatial biases in attention.
Neglect severity was measured with the Schenkenberg line bisection task (Schenkenberg,
Neurosciences en réseau
81
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
un
_
0
0
2
1
0
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
Bradford, & Ajax, 1980un) (18 horizontal lines, 10–20 cm) and a variant of the bell cancellation
test (Gauthier, Dehaut, & Joanette, 1989) (35 animal targets among distractor objects)
(Gassama, Deplancke, Saj, Honoré, & Rousseaux, 2011). In the latter animal cancellation test,
the search sheet was divided into seven virtual columns, each containing five targets. In total,
there were 15 targets in the left side, 15 in the right side, et 5 in the central column. Clinique
neglect was defined as abnormal performance in the line bisection test (cutoff: rightward de-
viation > 11%) (Schenkenberg, Bradford, & Ajax, 1980b) and target cancellation test (cutoff:
left-right omissions ≥ 4 out of 15 omissions) (Gauthier et al., 1989). Cependant, some patients
showed deficits in one of these tests only, and all those selected for our sample (n = 15) dem-
onstrated some signs of hemineglect according to at least one of these tests (86% des malades
had cancellation deficits, 79% had line bisection deficits), as commonly observed after RH
dommage (Halligan et al., 1991; Verdon et al., 2010). Cependant, neglect severity varied substan-
tially between patients (leftward omission error range: 13%–100%; mean: 77%; SD: 29%). UN
group summary of the clinical and behavioral characteristics of the patients is reported in
Supporting Information Table S1.
EEG Recording and Processing
A multichannel EEG cap was used to measure whole-scalp activity in each baseline recording.
This consisted of resting-state measurement of 3 min under eyes open conditions, pendant
which participants gazed at an empty black computer screen with their head comfortably po-
sitioned against at headrest. All EEG recordings were performed in standardized conditions
during the patient’s clinical rehabilitation visits. Using a similar clinical EEG setup as in pre-
ceding work (Tokariev et al., 2018), scalp voltages were recorded with a 19 Ag/AgCl electrode
cap (Electro-cap International, Inc., https://www.electro-cap.com) according to the 10–20 in-
ternational system. The ground electrode was placed on the scalp, at a site equidistant be-
tween Fpz and Fz. Electrical signals were amplified with the Mitsar 21-channel EEG system
(Mitsar-201, CE0537, Mitsar, Ltd. https://www.mitsar-medical.com), and all electrode imped-
ances were kept under 5 kΩ. For online recording, electrodes were referenced to linked ear-
lobes, and then the common average reference was calculated off-line before further analysis.
EEG data was recorded at 250 Hz and then filtered with a 0.5–40 Hz band-pass filter off-line.
All EEG data were imported into the MATLAB toolbox EEGLAB v12 (https://sccn.ucsd.edu
/eeglab/) for off-line processing. We used Infomax ICA decomposition to remove usual eye
movement such as saccades or blinking (Jung et al., 2000). Recordings were further cleaned
with an automated z-score-based method, using the FASTER plugin (Nolan, Whelan, & Reilly,
2010), rejecting 1-s epochs that deviated from the mean by more than 1.5 standard deviations,
in light of frequent occurrences of involuntary head movements in the stroke patients. Ce
strict selection procedure avoided an inflation of spurious increases in connectivity while pre-
serving a large dataset in each individual. Enfin, EEG discontinuities were minimized by uti-
lizing Hann windowing (MATLAB hann() window function) on all clean epochs before
concatenating them into continuous EEG.
Source Space Measures of EEG Activity
Artifact-free EEG data were processed in MATLAB with the Brainstorm Toolbox (https://
neuroimage.usc.edu/brainstorm/). In line with previous approaches using a similar EEG setup
in clinical populations (Tokariev et al., 2018), we first computed a head model of the cortex
surface for each EEG recording using the (symmetric) boundary element method from
OpenMEEG (Gramfort, Papadopoulo, Olivi, & Clerc, 2010) and then estimated unconstrained
cortical sources by using the minimum-norm sLORETA algorithm implemented in Brainstorm.
Neurosciences en réseau
82
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
un
_
0
0
2
1
0
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
Amplitude envelope
correlation (AEC):
The correlation coefficient between
the instantaneous amplitude
fluctuations of two brain regions.
To normalize sources across participants, we projected (warped) the sources from each par-
ticipant onto the MNI/Colin27 template brain surface (Holmes et al., 1998). Le 15,000 voxel
source space was then divided into 68 cortical ROIs according to the Desikan–Killiany neu-
roanatomical atlas (Desikan et al., 2006). Temporal source activities across all the voxels in
each ROI were then averaged and band-pass filtered in the following six frequency bands:
delta 1–4 Hz, theta 4–8 Hz, alpha 8–12 Hz, beta 13–30 Hz, and gamma 30–45 Hz. For every
sujet, each frequency band was quantified in Brainstorm to examine differences in spectral
power and FC between the patient and control groups.
Spectral Power
Band-limited EEG power was estimated with a standard fast Fourier transform approach using
Welch’s method (MATLAB pwelch() fonction) and a Hanning windowing function (4-s epoch,
50% overlap). Relative spectral power (c'est à dire., % pouvoir) was calculated as the ratio of the mean
power in a specific EEG band and the broadband power (1–45 Hz). Multiple comparison cor-
rection was performed using the FDR option in the Brainstorm Toolbox.
Functional Connectivity
A single time course was constructed for each ROI, which was then defined as a node in a
network graph. Connectivity between nodes was subsequently estimated using the amplitude
envelope correlation, yielding a 68 × 68 node adjacency matrix. The first step in estimating the
amplitude envelope correlation is to compensate for spatial leakage confounds by using a bi-
directional orthogonalization procedure (Hipp, Hawellek, Corbetta, Siegel, & Ange, 2012) à
remove all shared signal at zero lag between filtered EEG signals. After this, we computed the
instantaneous amplitude (c'est à dire., envelope) across time for each frequency band by using the ab-
solute value of the Hilbert transform. Enfin, the linear correlation between the amplitude time
series of each node pair was calculated using the Pearson correlation coefficient (Brookes
et coll., 2011). For visualization of group differences (c'est à dire., Chiffre 1), we reported the absolute
difference in Pearson correlation (r) which may be regarded as a standardized measure of ef-
fect size (Nakagawa & Cuthill, 2007).
Statistical Analyses
Source space (voxel-wise) statistical comparisons of band-limited spectral power were per-
formed using the Brainstorm Toolbox via independent two-tailed t tests with a p < 0.05 thresh-
old. Separately, we used the GraphVar Toolbox (Kruschwitz, List, Waller, Rubinov, & Walter,
2015) for statistical comparisons of network connectivity using intergroup t tests with a p <
0.05 threshold. Individual neglect severity was measured by performance on the cancellation
test and subsequently used in a between-subject regression analysis (with a p < 0.05
threshold).
Our statistical analyses followed a stepwise approach. First, we performed a direct compar-
ison of whole-brain connectivity changes in stroke patients versus healthy controls, regardless
of neglect severity. This allowed us to optimize our statistical power by considering the whole
sample data and avoid a dichotomous categorization of neglect presence versus absence
based on arbitrary criteria, given that neglect diagnosis may vary according to the test used
(Checketts et al., 2020; Verdon et al., 2010) and that many of our patients showed some ne-
glect deficits in one task but not in others. Next, we performed a regression analysis to identify
those connections whose strength varied as a function of the severity of neglect deficits. The
specificity of this relationship was further ensured by a final conjunction test (Nichols, Brett,
Network Neuroscience
83
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
Andersson, Wager, & Poline, 2005) to isolate neglect-related changes among the stroke-
related anomalies in connectivity patterns. Impaired visuospatial attention was quantified by
the absolute number of omitted targets (i.e., errors) within the left and right hemifields during
the cancellation test in each individual patient. Please note we preferred the absolute rather
than the relative number of omissions (e.g., difference between left and right) as the latter did
not reliably reflect the stroke severity and extent of network damage in EEG. This is because
stroke-related lateralized damage in brain network should induce a proportional degree of la-
teralized behavioral deficit, while a numerically similar difference in relative number (left mi-
nus right) could occur with different absolute values and different extent of brain damage. A
similar quantitative analysis of brain–behavior relationships using deviation severity on the
line bisection test did not yield any significant results and was not further reported.
For all tests, and to statistically correct for multiple comparisons in FC measures, we used
the NBS (Zalesky, Fornito, & Bullmore, 2010) based on MATLAB code from the Brain
Connectivity Toolbox (brain-connectivity-toolbox.net). We performed n = 5,000 permutations
to generate a null network model based on a random shuffle of all connections. Basically, the
NBS is used to control the family-wise error rate when the null hypothesis is tested indepen-
dently at each of the N(N-1)/2 edges comprising the connectivity matrix. The NBS may provide
greater statistical power than conventional procedures such as FDR, when the set of edges at
which the null hypothesis is rejected constitutes large component(s). The theoretical basis of
the NBS is to consider the pairwise similarity matrix of functional connections by using the
framework of graph theory, insofar as conventional cluster statistics are applied to a graph
structure, with the main difference that graph components (i.e., significantly interconnected
nodes) play the role of voxel clusters. This proceeds as follows. First, the test statistic computed
for each link is thresholded to construct a set of suprathreshold links. Any connected struc-
tures, or components in graph terminology, that may be present in the set of suprathreshold
links are then identified. Lastly, using permutation tests, a p value is assigned to each identified
component by indexing its size with the null distribution of maximal component size. As a
result, in the NBS framework, it is not possible to declare individual links (pairwise connec-
tions) as being significant, but only the network component to which they belong can be de-
clared significant. Hence, the individual (i.e., link level) connection t values were reported
mainly indicatively, together with d as the absolute difference in Pearson correlation (r), which
may be regarded as a standardized measure of effect size.
Data Availability
Anonymized EEG data and MATLAB analysis scripts that were used in this study are available
from the corresponding author upon request.
ACKNOWLEDGMENTS
The authors would like to thank all the participants and their families, the neuropsychologists,
all members of the neuropsychology unit, and the general rehabilitation team of the Clinique
Romande de Réadaptation and Vallée de Coeur Institute in Sion for their help and useful
advice.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00210.
Network Neuroscience
84
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
AUTHOR CONTRIBUTIONS
Tomas Ros: Conceptualization; Formal analysis; Methodology; Visualization; Writing – origi-
nal draft; Writing – review & editing. Abele Michela: Data curation; Project administration.
Anaïs Mayer: Data curation; Methodology. Anne Bellmann: Conceptualization;
Investigation; Project administration. Philippe Vuadens: Conceptualization; Data curation;
Investigation; Project administration. Victorine Zermatten: Investigation; Project administration.
Arnaud Saj: Conceptualization; Funding acquisition; Investigation; Project administration;
Resources. Patrik Vuilleumier: Conceptualization; Funding acquisition; Supervision; Writing –
review & editing.
FUNDING INFORMATION
This study was generously supported by grants from the Leenaards Foundation, as well as the EU
Marie-Curie COFUND program BRIDGE (Grant no. 267,171), Geneva University Hospitals
(HUG), Société Académique de Genève, and the Swiss National Science Foundation (SNF;
Grant no. 320030-166704).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
REFERENCES
Akam, T. E., & Kullmann, D. M. (2012). Efficient “communication
through coherence” requires oscillations structured to minimize
interference between signals. PLoS Computational Biology, 8(11),
e1002760. https://doi.org/10.1371/journal.pcbi.1002760,
PubMed: 23144603
Antzoulatos, E. G., & Miller, E. K. (2016). Synchronous beta rhythms
of frontoparietal networks support only behaviorally relevant
representations. ELife, 5. https://doi.org/10.7554/eLife.17822,
PubMed: 27841747
Baldassarre, A., Ramsey, L., Hacker, C. L., Callejas, A., Astafiev,
S. V., Metcalf, N. V., … Corbetta, M. (2014). Large-scale changes
in network interactions as a physiological signature of spatial
neglect. Brain, 137, 3267–3283. https://doi.org/10.1093/ brain
/awu297, PubMed: 25367028
Bauer, M., Kluge, C., Bach, D., Bradbury, D., Heinze, H. J., Dolan,
R. J., & Driver, J. (2012). Cholinergic enhancement of visual
attention and neural oscillations in the human brain. Current
Biology, 22(5), 397–402. https://doi.org/10.1016/j.cub.2012.01
.022, PubMed: 22305751
Bentley, W. J., Li, J. M., Snyder, A. Z., Raichle, M. E., & Snyder,
L. H. (2016). Oxygen level and LFP in task-positive and task-
negative areas: Bridging BOLD fMRI and electrophysiology.
Cerebral Cortex, 26(1), 346–357. https://doi.org/10.1093/cercor
/bhu260, PubMed: 25385710
Bonnefond, M., Kastner, S., & Jensen, O. (2017). Communication
between brain areas based on nested oscillations. ENeuro, 4(2).
https://doi.org/10.1523/ ENEURO.0153-16.2017, PubMed:
28374013
Brookes, M. J., Hale, J. R., Zumer, J. M., Stevenson, C. M., Francis,
S. T., Barnes, G. R., … Nagarajan, S. S. (2011). Measuring func-
tional connectivity using MEG: Methodology and comparison
with fcMRI. NeuroImage, 56(3), 1082–1104. https://doi.org/10
.1016/j.neuroimage.2011.02.054, PubMed: 21352925
Castellanos, N. P., Paúl, N., Ordóñez, V. E., Demuynck, O., Bajo,
R., Campo, P., … Maestú, F. (2010). Reorganization of functional
connectivity as a correlate of cognitive recovery in acquired
brain injury. Brain, 133(8), 2365–2381. https://doi.org/10.1093
/brain/awq174, PubMed: 20826433
Checketts, M., Mancuso, M., Fordell, H., Chen, P., Eskes, G. A.,
Vuilleumier, P., … Bowen, A. (2020). Current clinical practice in
the screening and diagnosis of spatial neglect post-stroke: Findings
from a multidisciplinary international survey. Neuropsychological
Rehabilitation, 31(9), 1495–1526. https://doi.org/10.1080
/09602011.2020.1782946, PubMed: 32691688
Chelazzi, L., Duncan, J., Miller, E. K., & Desimone, R. (1998).
Responses of neurons in inferior temporal cortex during
memory-guided visual search. Journal of Neurophysiology, 80(6),
2918–2940. https://doi.org/10.1152/jn.1998.80.6.2918, PubMed:
9862896
Colson, C., Demeurisse, G., Hublet, C., & Slachmuylder, J.-L.
(2001). Subcortical neglect as a consequence of a remote
parieto-temporal dysfunction. A quantitative EEG study. Cortex,
37(5), 619–625. https://doi.org/10.1016/S0010-9452(08)70609-9,
PubMed: 11804212
Corbetta, M., & Shulman, G. L. (2011). Spatial neglect and atten-
tion networks. Annual Review of Neuroscience, 34, 569–599.
https://doi.org/10.1146/annurev-neuro-061010-113731,
PubMed: 21692662
Cosandier-Rimélé, D., Ramantani, G., Zentner, J., Schulze-
Bonhage, A., & Dümpelmann, M. (2017). A realistic multimodal
modeling approach for the evaluation of distributed source
analysis: Application to sLORETA. Journal of Neural Engineering,
14(5), 056008. https://doi.org/10.1088/1741-2552/aa7db1,
PubMed: 28677591
de Pasquale, F., Della Penna, S., Snyder, A. Z., Lewis, C., Mantini,
D., Marzetti, L., … Corbetta, M. (2010). Temporal dynamics of
Network Neuroscience
85
EEG network correlates of visuospatial neglect
spontaneous MEG activity in brain networks. Proceedings of the
National Academy of Sciences of the United States of America,
107(13), 6040–6045. https://doi.org/10.1073/pnas.0913863107,
PubMed: 20304792
Demeurisse, G., Hublet, C., & Paternot, J. (1998). Quantitative EEG
in subcortical neglect. Clinical Neurophysiology, 28(3), 259–265.
https://doi.org/10.1016/S0987-7053(98)80116-0, PubMed:
9686401
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson, B. C.,
Blacker, D., Buckner, R. L., Dale, A. M., Maguire, R. P., Hyman,
B. T., Albert, M. S., Killiany, R. J. (2006). An automated labeling
system for subdividing the human cerebral cortex on MRI
scans into gyral based regions of interest. NeuroImage, 31(3),
968–980. https://doi.org/10.1016/j.neuroimage.2006.01.021,
PubMed: 16530430
Domínguez-Borràs, J., Armony, J. L., Maravita, A., Driver, J., &
Vuilleumier, P. (2013). Partial recovery of visual extinction by
Pavlovian conditioning in a patient with hemispatial neglect.
Cortex, 49(3), 891–898. https://doi.org/10.1016/j.cortex.2012
.11.005, PubMed: 23337458
Doricchi, F., Thiebaut de Schotten, M., Tomaiuolo, F., &
Bartolomeo, P. (2008). White matter (dis)connections and gray
matter (dys)functions in visual neglect: Gaining insights into the
brain networks of spatial awareness. Cortex, 44(8), 983–995.
https://doi.org/10.1016/j.cortex.2008.03.006, PubMed:
18603235
Driver, J., & Vuilleumier, P. (2001). Perceptual awareness and its loss
in unilateral neglect and extinction. Cognition, 79(1–2), 39–88.
https://doi.org/10.1016/S0010-0277(00)00124-4, PubMed:
11164023
Dubovik, S., Pignat, J.-M., Ptak, R., Aboulafia, T., Allet, L.,
Gillabert, N., … Guggisberg, A. G. (2012). The behavioral signif-
icance of coherent resting-state oscillations after stroke.
NeuroImage, 61(1), 249–257. https://doi.org/10.1016/j
.neuroimage.2012.03.024, PubMed: 22440653
Fellrath, J., Mottaz, A., Schnider, A., Guggisberg, A. G., & Ptak, R.
(2016). Theta-band functional connectivity in the dorsal
fronto-parietal network predicts goal-directed attention.
Neuropsychologia, 92, 20–30. https://doi.org/10.1016/j
.neuropsychologia.2016.07.012, PubMed: 27422540
Fiebelkorn, I. C., Pinsk, M. A., & Kastner, S. (2018). A dynamic
interplay within the frontoparietal network underlies rhythmic
spatial attention. Neuron, 99(4), 842–853. https://doi.org/10
.1016/j.neuron.2018.07.038, PubMed: 30138590
Gassama, S., Deplancke, A., Saj, A., Honoré, J., & Rousseaux, M.
(2011). Do supine position and deprivation of visual environ-
ment influence spatial neglect? Journal of Neurology, 258(7),
1288–1294. https://doi.org/10.1007/s00415-011-5926-z,
PubMed: 21287184
Gauthier, L., Dehaut, F., & Joanette, Y. (1989). The Bells Test: A
quantitative and qualitative test for visual neglect. International
Journal of Clinical Neuropsychology, 11(2), 49–54.
Geng, J. J., & Mangun, G. R. (2009). Anterior intraparietal sulcus is
sensitive to bottom-up attention driven by stimulus salience.
Journal of Cognitive Neuroscience, 21(8), 1584–1601. https://
doi.org/10.1162/jocn.2009.21103, PubMed: 18752405
Gitelman, D. R., Nobre, A. C., Parrish, T. B., LaBar, K. S., Kim, Y. H.,
Meyer, J. R., & Mesulam, M. (1999). A large-scale distributed
network for covert spatial attention: Further anatomical delinea-
tion based on stringent behavioural and cognitive controls.
Brain, 122(Pt 6), 1093–1106. https://doi.org/10.1093/brain/122
.6.1093, PubMed: 10356062
Gramfort, A., Papadopoulo, T., Olivi, E., & Clerc, M. (2010).
OpenMEEG: Opensource software for quasistatic bioelectromag-
netics. BioMedical Engineering OnLine, 9(1), 45. https://doi.org/10
.1186/1475-925X-9-45, PubMed: 20819204
Grech, R., Cassar, T., Muscat, J., Camilleri, K. P., Fabri, S. G.,
Zervakis, M., … Vanrumste, B. (2008). Review on solving the
J o u r n a l o f
i n v e r s e p r o b l e m i n E E G s o u r c e a n a l y s i s .
NeuroEngineering and Rehabilitation, 5(1), 25. https://doi.org
/10.1186/1743-0003-5-25, PubMed: 18990257
Grin-Yatsenko, V. A., Baas, I., Ponomarev, V. A., & Kropotov,
J. D. (2009). EEG power spectra at early stages of depressive
disorders. Journal of Clinical Neurophysiology, 26(6), 401–406.
https://doi.org/10.1097/ WNP.0b013e3181c298fe, PubMed:
19952564
Guggisberg, A. G., Rizk, S., Ptak, R., Di Pietro, M., Saj, A.,
Lazeyras, F., … Pignat, J.-M. (2014). Two intrinsic coupling types
for resting-state integration in the human brain. Brain
Topography, 28(2), 318–329. https://doi.org/10.1007/s10548
-014-0394-2, PubMed: 25182143
Halligan, P., Robertson, I., Pizzamiglio, L., Homberg, V., Weber, E.,
& Bergego, C. (1991). The laterality of visual neglect after right
hemisphere damage. Neuropsychological Rehabilitation, 1(4),
281–301. https://doi.org/10.1080/09602019108402259
Heilman, K. M., & Van Den Abell, T. (1980). Right hemisphere
dominance for attention: The mechanism underlying hemi-
spheric asymmetries of inattention (neglect). Neurology, 30(3),
327–330. https://doi.org/10.1212/ WNL.30.3.327, PubMed:
7189037
Hermes, D., Nguyen, M., & Winawer, J. (2017). Neuronal synchro-
ny and the relation between the blood-oxygen-level dependent
response and the local field potential. PLoS Biology, 15(7),
e2001461. https://doi.org/10.1371/journal.pbio.2001461,
PubMed: 28742093
Hillis, A. E., Newhart, M., Heidler, J., Barker, P. B., Herskovits, E. H.,
& Degaonkar, M. (2005). Anatomy of spatial attention: Insights
from perfusion imaging and hemispatial neglect in acute stroke.
Journal of Neuroscience, 25(12), 3161–3167. https://doi.org/10
.1523/JNEUROSCI.4468-04.2005, PubMed: 15788773
Hipp, J. F., Hawellek, D. J., Corbetta, M., Siegel, M., & Engel, A. K.
(2012). Large-scale cortical correlation structure of spontaneous
oscillatory activity. Nature Neuroscience, 15(6), 884–890.
https://doi.org/10.1038/nn.3101, PubMed: 22561454
Holmes, C. J., Hoge, R., Collins, L., Woods, R., Toga, A. W., &
Evans, A. C. (1998). Enhancement of MR images using registra-
tion for signal averaging. Journal of Computer Assisted
Tomography, 22(2), 324–333. https://doi.org/10.1097
/00004728-199803000-00032, PubMed: 9530404
Hutchison, R. M., Hashemi, N., Gati, J. S., Menon, R. S., & Everling,
S. (2015). Electrophysiological signatures of spontaneous BOLD
fluctuations in macaque prefrontal cortex. NeuroImage, 113,
257–267. https://doi.org/10.1016/ J.NEUROIMAGE.2015.03
.062, PubMed: 25837599
Ikkai, A., Dandekar, S., & Curtis, C. E. (2016). Lateralization in
alpha-band oscillations predicts the locus and spatial distribution
Network Neuroscience
86
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
of attention. PLoS One, 11(5), e0154796. https://doi.org/10.1371
/journal.pone.0154796, PubMed: 27144717
Jensen, O., Gips, B., Bergmann, T. O., & Bonnefond, M. (2014).
Temporal coding organized by coupled alpha and gamma
oscillations prioritize visual processing. Trends in Neurosciences,
37(7), 357–369. https://doi.org/10.1016/ J.TINS.2014.04.001,
PubMed: 24836381
Jung, T. P., Makeig, S., Humphries, C., Lee, T. W., McKeown, M. J.,
Iragui, V., & Sejnowski, T. J. (2000). Removing electroencepha-
lographic artifacts by blind source separation. Psychophysiology,
37(2), 163–178. https://doi.org/10.1111/1469-8986.3720163,
PubMed: 10731767
Karnath, H.-O., Ferber, S., & Himmelbach, M. (2001). Spatial
awareness is a function of the temporal not the posterior parietal
lobe. Nature, 411(6840), 950–953. https://doi.org/10.1038
/35082075, PubMed: 11418859
Karnath, H.-O., Rennig, J., Johannsen, L., & Rorden, C. (2011). The
anatomy underlying acute versus chronic spatial neglect: A lon-
gitudinal study. Brain, 134(Pt 3), 903–912. https://doi.org/10
.1093/brain/awq355, PubMed: 21156661
Kawano, T., Hattori, N., Uno, Y., Kitajo, K., Hatakenaka, M.,
Yagura, H., … Miyai, I. (2017). Large-scale phase synchrony reflects
clinical status after stroke: An EEG study. Neurorehabilitation and
Neural Repair, 31(6), 561–570. https://doi.org/10.1177
/1545968317697031, PubMed: 28506148
Kinsbourne, M. (1977). Hemi-neglect and hemisphere rivalry.
Advances in Neurology, 18, 41–49. PubMed: 920524
Klinke, M. E., Hjaltason, H., & Tryggvadóttir, G. B. (2018). Topics
in stroke rehabilitation hemispatial neglect following right hemi-
sphere stroke: Clinical course and sensitivity of diagnostic tasks.
Topics in Stroke Rehabilitation, 9357, 1–11. https://doi.org/10
.1080/10749357.2017.1394632, PubMed: 29105618
Kruschwitz, J. D., List, D., Waller, L., Rubinov, M., & Walter, H.
(2015). GraphVar: A user-friendly toolbox for comprehensive
graph analyses of functional brain connectivity. Journal of
Neuroscience Methods, 245, 107–115. https://doi.org/10.1016
/J.JNEUMETH.2015.02.021, PubMed: 25725332
Liu, Y., Bengson, J., Huang, H., Mangun, G. R., & Ding, M. (2014).
Top-down modulation of neural activity in anticipatory visual at-
tention: Control mechanisms revealed by simultaneous EEG-
fMRI. Cerebral Cortex, 26(2), 517–529. https://doi.org/10.1093
/cercor/bhu204, PubMed: 25205663
Lobier, M., Palva, J. M., & Palva, S. (2017). High-alpha band syn-
chronization across frontal, parietal and visual cortex mediates
behavioral and neuronal effects of visuospatial attention.
NeuroImage, 165, 222–237. https://doi.org/10.1016/j.neuroimage
.2017.10.044, PubMed: 29074278
Mantini, D., Perrucci, M. G., Del Gratta, C., Romani, G. L., &
Corbetta, M. (2007). Electrophysiological signatures of resting
state networks in the human brain. Proceedings of the National
Academy of Sciences of the United States of America, 104(32),
13170–13175. https://doi.org/10.1073/pnas.0700668104,
PubMed: 17670949
Marshall, T. R., Bergmann, T. O., & Jensen, O. (2015a).
Frontoparietal structural connectivity mediates the top-down
control of neuronal synchronization associated with selective at-
tention. PLoS Biology, 13(10), e1002272. https://doi.org/10.1371
/journal.pbio.1002272, PubMed: 26441286
Marshall, T. R., Bergmann, T. O., & Jensen, O. (2015b).
Frontoparietal structural connectivity mediates the top-down
control of neuronal synchronization associated with selective
attention. PLoS Biology, 13(10), e1002272. https://doi.org/10
.1371/journal.pbio.1002272, PubMed: 26441286
Melloni, L., Molina, C., Pena, M., Torres, D., Singer, W., &
Rodriguez, E. (2007). Synchronization of neural activity across
cortical areas correlates with conscious perception. The Journal
of Neuroscience, 27(11), 2858–2865. https://doi.org/10.1523
/JNEUROSCI.4623-06.2007, PubMed: 17360907
Mesulam, M. M. (1981). A cortical network for directed attention
and unilateral neglect. Annals of Neurology, 10(4), 309–325.
https://doi.org/10.1002/ana.410100402, PubMed: 7032417
Mort, D. J., Malhotra, P., Mannan, S. K., Rorden, C., Pambakian, A.,
Kennard, C., & Husain, M. (2003). The anatomy of visual ne-
glect. Brain, 126(9), 1986–1997. https://doi.org/10.1093/brain
/awg200, PubMed: 12821519
Mottaz, A., Corbet, T., Doganci, N., Magnin, C., Nicolo, P.,
Schnider, A., & Guggisberg, A. G. (2018). Modulating functional
connectivity after stroke with neurofeedback: Effect on motor
deficits in a controlled cross-over study. NeuroImage: Clinical,
20, 336–346. https://doi.org/10.1016/j.nicl.2018.07.029,
PubMed: 30112275
Nakagawa, S., & Cuthill, I. C. (2007). Effect size, confidence inter-
val and statistical significance: A practical guide for biologists.
Biological Reviews, 82(4), 591–605. https://doi.org/10.1111/j
.1469-185X.2007.00027.x, PubMed: 17944619
Nichols, T., Brett, M., Andersson, J., Wager, T., & Poline, J.-B.
(2005). Valid conjunction inference with the minimum statistic.
NeuroImage, 25(3), 653–660. https://doi.org/10.1016/j
.neuroimage.2004.12.005, PubMed: 15808966
Nicolo, P., Rizk, S., Magnin, C., Di Pietro, M., Schnider, A., &
Guggisberg, A. G. (2015). Coherent neural oscillations predict
future motor and language improvement after stroke. Brain,
138(10), 3048–3060. https://doi.org/10.1093/ brain/awv200,
PubMed: 26163304
Nolan, H., Whelan, R., & Reilly, R. B. (2010). FASTER: Fully auto-
mated statistical thresholding for EEG artifact rejection. Journal of
Neuroscience Methods, 192(1), 152–162. https://doi.org/10
.1016/j.jneumeth.2010.07.015, PubMed: 20654646
Okazaki, Y. O., Horschig, J. M., Luther, L., Oostenveld, R.,
Murakami, I., & Jensen, O. (2015). Real-time MEG neurofeed-
back training of posterior alpha activity modulates subsequent
visual detection performance. NeuroImage, 107, 323–332.
https://doi.org/10.1016/j.neuroimage.2014.12.014, PubMed:
25514519
Ptak, R., & Valenza, N. (2005). The inferior temporal lobe mediates
distracter-resistant visual search of patients with spatial neglect.
Journal of Cognitive Neuroscience, 17(5), 788–799. https://doi
.org/10.1162/0898929053747676, PubMed: 15904545
Ramsey, L. E., Siegel, J. S., Baldassarre, A., Metcalf, N. V., Zinn, K.,
Shulman, G. L., & Corbetta, M. (2016). Normalization of network
connectivity in hemispatial neglect recovery. Annals of
Neurology, 80(1), 127–141. https://doi.org/10.1002/ana.24690,
PubMed: 27277836
Ros, T., Michela, A., Bellman, A., Vuadens, P., Saj, A., &
Vuilleumier, P. (2017). Increased alpha-rhythm dynamic range
promotes recovery from visuospatial neglect: A neurofeedback
Network Neuroscience
87
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
study. Neural Plasticity, 2017, 1–9. https://doi.org/10.1155/2017
/7407241, PubMed: 28529806
Saj, A., Cojan, Y., Vocat, R., Luauté, J., & Vuilleumier, P. (2013).
Prism adaptation enhances activity of intact fronto-parietal areas
in both hemispheres in neglect patients. Cortex, 49(1), 107–119.
https://doi.org/10.1016/j.cortex.2011.10.009, PubMed: 22154751
Sasaki, T., Abe, M., Okumura, E., Okada, T., Kondo, K., Sekihara,
K., … Kamada, H. (2013). Disturbed resting functional inter-
hemispherical connectivity of the ventral attentional network in
alpha band is associated with unilateral spatial neglect. PLoS
One, 8(9), e73416. https://doi.org/10.1371/journal.pone
.0073416, PubMed: 24023873
Schenkenberg, T., Bradford, D. C., & Ajax, E. T. (1980a). Line
bisection and unilateral visual neglect in patients with neurologic
impairment. Neurology, 30(5), 509–517. https://doi.org/10.1212
/ WNL.30.5.509, PubMed: 7189256
Schenkenberg, T., Bradford, D. C., & Ajax, E. T. (1980b). Line
bisection and unilateral visual neglect in patients with neurologic
impairment. Neurology, 30(5), 509–517. https://doi.org/10.1212
/ WNL.30.5.509, PubMed: 7189256
Schölvinck, M. L., Maier, A., Ye, F. Q., Duyn, J. H., & Leopold,
D. A. (2010). Neural basis of global resting-state fMRI activity.
Proceedings of the National Academy of Sciences of the
United States of America, 107(22), 10238–10243. https://doi
.org/10.1073/pnas.0913110107, PubMed: 20439733
Siegel, J. S., Ramsey, L. E., Snyder, A. Z., Metcalf, N. V., Chacko,
R. V., Weinberger, K., … Corbetta, M. (2016). Disruptions of
network connectivity predict impairment in multiple behavioral
domains after stroke. Proceedings of the National Academy of
Sciences, 113(30), E4367–E4376. https://doi.org/10.1073/pnas
.1521083113, PubMed: 27402738
Song, J., Davey, C., Poulsen, C., Luu, P., Turovets, S., Anderson, E., …
Tucker, D. (2015). EEG source localization: Sensor density and
head surface coverage. Journal of Neuroscience Methods, 256,
9–21. https://doi.org/10.1016/ J.JNEUMETH.2015.08.015,
PubMed: 26300183
Szczepanski, S. M., Crone, N. E., Kuperman, R. A., Auguste, K. I.,
Parvizi, J., & Knight, R. (2014). Dynamic changes in phase-
amplitude coupling facilitate spatial attention control in fronto-
parietal cortex. PLoS Biology, 12(8), e1001936. https://doi.org
/10.1371/journal.pbio.1001936, PubMed: 25157678
Thimm, M., Fink, G. R., & Sturm, W. (2008). Neural correlates of
recovery from acute hemispatial neglect. Restorative Neurology
and Neuroscience, 26(6), 481–492. PubMed: 19096136
Tokariev, A., Stjerna, S., Lano, A., Metsäranta, M., Palva, J. M., &
Vanhatalo, S. (2018). Preterm birth changes networks of newborn
cortical activity. Cerebral Cortex, 29(2), 814–826. https://doi.org
/10.1093/cercor/bhy012, PubMed: 30321291
Umarova, R. M., Saur, D., Kaller, C. P., Vry, M.-S., Glauche, V.,
Mader, I., … Weiller, C. (2011). Acute visual neglect and extinc-
tion: distinct functional state of the visuospatial attention system.
Brain, 134(11), 3310–3325. https://doi.org/10.1093/ brain
/awr220, PubMed: 21948940
Vaessen, M. J., Saj, A., Lovblad, K.-O., Gschwind, M., &
Vuilleumier, P. (2016). Structural white-matter connections me-
diating distinct behavioral components of spatial neglect in right
brain-damaged patients. Cortex, 77, 54–68. https://doi.org/10
.1016/j.cortex.2015.12.008, PubMed: 26922504
Vallar, G., & Perani, D.
(1986). The anatomy of unilateral
neglect after right-hemisphere stroke lesions. A clinical/CT-scan
correlation study in man. Neuropsychologia, 24(5), 609–622.
https://doi.org/10.1016/0028-3932(86)90001-1, PubMed:
3785649
van Dellen, E., Hillebrand, A., Douw, L., Heimans, J. J., Reijneveld,
J. C., & Stam, C. J. (2013). Local polymorphic delta activity in
cortical lesions causes global decreases in functional connectivity.
NeuroImage, 83, 524–532. https://doi.org/10.1016/j.neuroimage
.2013.06.009, PubMed: 23769919
van Diepen, R. M., Miller, L. M., Mazaheri, A., & Geng, J. J. (2016).
The role of alpha activity in spatial and feature-based attention.
ENeuro, 3(5). https://doi.org/10.1523/ENEURO.0204-16.2016,
PubMed: 27822505
van Schouwenburg, M. R., Sörensen, L. K. A., de Klerk, R., Reteig,
L. C., & Slagter, H. A. (2018). No differential effects of two differ-
ent alpha-band electrical stimulation protocols over fronto-
parietal regions on spatial attention. Frontiers in Neuroscience,
12, 433. https://doi.org/10.3389/fnins.2018.00433, PubMed:
30018530
van Schouwenburg, M. R., Zanto, T. P., & Gazzaley, A. (2017).
Spatial attention and the effects of
frontoparietal alpha
band stimulation. Frontiers in Human Neuroscience, 10, 658.
https://doi.org/10.3389/fnhum.2016.00658, PubMed:
28174529
Verdon, V., Schwartz, S., Lovblad, K.-O., Hauert, C.-A., &
Vuilleumier, P. (2010). Neuroanatomy of hemispatial neglect
and its functional components: A study using voxel-based
lesion-symptom mapping. Brain, 133(Pt 3), 880–894. https://doi
.org/10.1093/brain/awp305, PubMed: 20028714
Vinck, M., Womelsdorf, T., Buffalo, E. A., Desimone, R., & Fries, P.
(2013). Attentional modulation of cell-class-specific gamma-band
synchronization in awake monkey area V4. Neuron, 80(4),
1077–1089. https://doi.org/10.1016/ J.NEURON.2013.08.019,
PubMed: 24267656
Volz, L. J., & Gazzaniga, M. S. (2017). Interaction in isolation:
50 years of insights from split-brain research. Brain, 140(7),
2051–2060. https://doi.org/10.1093/ brain/awx139, PubMed:
29177496
Vuilleumier, P. (2013). Mapping the functional neuroanatomy of
spatial neglect and human parietal lobe functions: Progress
and challenges. Annals of the New York Academy of Sciences,
1296, 50–74. https://doi.org/10.1111/nyas.12161, PubMed:
23751037
Vuilleumier, P. O., & Rafal, R. D. (2000). A systematic study
of visual extinction. Between- and within-field deficits of
attention in hemispatial neglect. Brain, 123(Pt 6), 1263–1279.
https://doi.org/10.1093/brain/123.6.1263, PubMed: 10825363
Vuilleumier, P., Sagiv, N., Hazeltine, E., Poldrack, R. A., Swick, D.,
Rafal, R. D., & Gabrieli, J. D. (2001). Neural fate of seen and
unseen faces in visuospatial neglect: A combined event-related
functional MRI and event-related potential study. Proceedings
of the National Academy of Sciences of the United States of
America, 98(6), 3495–3500. https://doi.org/10.1073/pnas
.051436898, PubMed: 11248106
Vuilleumier, P., Schwartz, S., Verdon, V., Maravita, A., Hutton, C.,
Husain, M., & Driver, J. (2008). Abnormal attentional modulation
of retinotopic cortex in parietal patients with spatial neglect.
Network Neuroscience
88
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG network correlates of visuospatial neglect
Current Biology, 18(19), 1525–1529. https://doi.org/10.1016/j
.cub.2008.08.072, PubMed: 18848444
Wróbel, A. (2000). Beta activity: A carrier for visual attention. Acta
Neurobiologiae Experimentalis, 60(2), 247–260. PubMed: 10909182
Yordanova, J., Kolev, V., Verleger, R., Heide, W., Grumbt, M., &
Schürmann, M. (2016). Synchronization of fronto-parietal beta
and theta networks as a signature of visual awareness in neglect.
NeuroImage, 146, 341–354. https://doi.org/10.1016/j.neuroimage
.2016.11.013, PubMed: 27840240
Yordanova, J., Kolev, V., Verleger, R., Heide, W., Grumbt, M., &
Schürmann, M. (2017). Synchronization of fronto-parietal beta
and theta networks as a signature of visual awareness in neglect.
NeuroImage, 146(July), 341–354. https://doi.org/10.1016/j
.neuroimage.2016.11.013, PubMed: 27840240
Zalesky, A., Fornito, A., & Bullmore, E. T. (2010). Network-based
statistic: Identifying differences in brain networks. NeuroImage,
53(4), 1197–1207. https://doi.org/10.1016/j.neuroimage.2010
.06.041, PubMed: 20600983
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
1
6
9
1
9
8
4
2
4
5
n
e
n
_
a
_
0
0
2
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
89