Recoding between Two Types of STM Representation
Revealed by the Dynamics of Memory Search
Marcin Leszczyński1, Nicholas E. Myers1,2, Elkan G. Akyürek3,
and Anna Schubö4
Abstrait
■ Visual STM (VSTM) is thought to be related to visual attention
in several ways. Attention controls access to VSTM during memory
encoding and plays a role in the maintenance of stored informa-
tion by strengthening memorized content. We investigated the
involvement of visual attention in recall from VSTM. In two experi-
ments, we measured electrophysiological markers of attention in
a memory search task with varying intervals between VSTM en-
coding and recall, and so we were able to track recoding of rep-
resentations in memory. Results confirmed the involvement of
attention in VSTM recall. Cependant, the amplitude of the N2pc
and N3rs components, which mark orienting of attention and
search within VSTM, decreased as a function of delay. Inversement,
the amplitude of the P3 and sustained posterior contralateral nega-
tivity components increased as a function of delay, effectively the
opposite of the N2pc and N3rs modulations. These effects were
only observed when verbal memory was not taxed. Ainsi, the re-
sults suggested that gradual recoding from visuospatial orienting
of attention into verbal recall mechanisms takes place from short
to long retention intervals. Fait intéressant, recall at longer delays
was faster than at short delays, indicating that verbal representa-
tion is coupled with faster responses. These results extend the
orienting-of-attention hypothesis by including an account of rep-
resentational recoding during short-term consolidation and its
consequences for recall from VSTM. ■
INTRODUCTION
Look at the desk in front of you. Now, close your eyes and
try recalling the color of the pencil lying next to the paper
pile on the right side of your desk. How do we recall such
an object and its features? Do we reconstruct an exact pic-
ture in the “mindʼs eye” and recall its features by attending
to the object like we do while categorizing a feature from
a visual scene? Visual STM (VSTM) has been found to be
closely related to visual attention (Akyürek, Leszczyński,
& Schubö, 2010; Soto & Humphreys, 2006, 2009; Soto,
Humphreys, & Heinke, 2006; Chelazzi, Duncan, Miller, &
Désimone, 1998; Désimone, 1998; Désimone & Duncan,
1995). Although memory encoding and maintenance have
long been shown to depend on attention mechanisms
(Vogel, McCollough, & Machizawa, 2005; Vogel & Machizawa,
2004; Awh, Anllo-Vento, & Hillyard, 2000; Logan, 1988), concernant-
cent studies have also linked orienting of attention to active
search within VSTM (DellʼAcqua, Sessa, Toffanin, Luria, &
Jolicœur, 2010; Eimer & Kiss, 2010; Kuo, Rao, Lepsien, &
Nobre, 2009) and to subliminally stored objects (Astle, Nobre,
& Scerif, 2010).
Modulations in attentional orienting to VSTM have been
assessed by measuring the amplitude of attention markers—
N2pc and N3rs. The N2pc component—a negative de-
1Ludwig Maximilian University Munich, 2Technical University
Munich, 3Université de Groningue, 4Philipps-University Marburg
flection observed over posterior contralateral electrodes
200 msec after stimulus presentation—has been found to
reflect lateralized attention (Eimer, 1996; Luck & Hillyard,
1994). The N3rs—a negative deflection observed 300 msec
after the presentation of a retro-(spective) search cue—
component has been reported to be triggered by orienting
attention toward locations within VSTM (Nobre, Griffin, &
Rao, 2008). It has been reported in paradigms that use a
so-called retro-search cue (with the cue indicating target
location in a memory template) as a fronto-centrally dis-
tributed negativity starting 300 msec after the presentation
of the retro-cue (Kuo et al., 2009; Nobre et al., 2008). Le
N3rs has been found to increase in amplitude with increas-
ing VSTM load. Fait intéressant, the N3rs was modulated by
neutral retro-cues only and not by spatial retro-cues (Nobre
et coll., 2008). This suggests that the N3rs might be suscep-
tible to the amount of spatial information during memory
recherche. Particularly, orienting attention to locations main-
tained in VSTM has been reflected in the amplitude of
the N3rs, which is reduced for increased memory load
(Nobre et al., 2008). Another component that has been
associated with memory processing and which might be
also susceptible to the amount of spatial information dur-
ing memory search is the SPCN (sustained posterior con-
tralateral negativity). The SPCN is also calculated as a
lateralization index, and therefore, it might be expected to
convey spatial information in VSTM, aussi. It is observed fol-
lowing encoding of spatial visual stimuli (Prime, DellʼAcqua,
© 2012 Massachusetts Institute of Technology
Journal des neurosciences cognitives 24:3, pp. 653–663
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
un
/
_
j
0
o
0
c
1
n
0
2
_
un
p
_
d
0
0
b
1
oui
0
g
2
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
t
.
F
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Arguin, Gosselin, & Jolicœur, 2010; Jolicœur, Brisson, &
Robitaille, 2008). Particularly, it has been found to increase
in amplitude with increasing memory load ( Jolicœur et al.,
2008), which makes it a possible marker of the amount
of visuospatial memory search. Similar to the N2pc, it is
calculated by subtracting activity measured at ipsilateral
electrode sites from activity measured at contralateral elec-
trode sites for lateralized presented targets. The SPCN has
been thought to mark maintenance of information stored
in STM ( Jolicœur et al., 2008).
Investigating mechanisms of recall from VSTM after var-
ious delays might require recurring to the concept of STM
consolidation ( Jolicœur & DellʼAcqua, 1998). Consolida-
tion has been found to operate rather in serial manner
( Jolicœur & DellʼAcqua, 1998). Ainsi, it takes time for in-
formation maintained in VSTM to be fully consolidated.
Varying retention interval (time lag between encoding
and recall) might, donc, correlate with the amount of
successfully performed consolidation. The current study
aimed at investigating ERP correlates of possible repre-
sentational changes over short-term consolidation by
manipulating the retention interval. En outre, STM
consolidation has been found to involve central processes,
which are also required for response selection, thereby
causing delays ( Jolicœur & DellʼAcqua, 1998). To inves-
tigate such behavioral effects, we also analyzed RT (cf.,
Jolicœur & DellʼAcqua, 1998).
The present study sought to address how attentional
orienting in memory develops over time. In two experi-
ments, we investigated the properties of recalled infor-
mation. We used various retention intervals in a memory
search task to cover the dynamics of the recall process.
We devised a memory search paradigm to obtain an elec-
trophysiological measure of attentional orienting strength
within VSTM. Memory search was triggered by a centrally
presented probe (voir la figure 1). The primary measure was
the amplitude of the N2pc elicited by the probe. We relate
the magnitude of the measured amplitude to the degree
of attentional orienting within VSTM. As a lateralized com-
ponent, the N2pc is usually measured in relation to a later-
ally presented object. The N2pc in the current study was
measured in response to a centrally presented probe (cf.
Chiffre 1). As the probe in the present study was not later-
alized, lateralization effects observed in this paradigm can
be attributed to attentional orienting triggered by the probe
but performed within VSTM. We additionally varied the
duration of the retention interval between the presentation
of the memory array and the probe ranging from 300 à
1700 msec. Varying the retention interval allowed us to
measure the strength of attentional orienting within VSTM
across time. On the basis of the observation that the am-
plitude of the N2pc in visual search is increased for condi-
tions requiring more spatial attention (par exemple., Eimer, 1996),
we hypothesized that the strength of attentional orienting
within the memory array would follow a similar pattern,
c'est, show a larger amplitude when more attention is
needed for recall. We expected performance at short re-
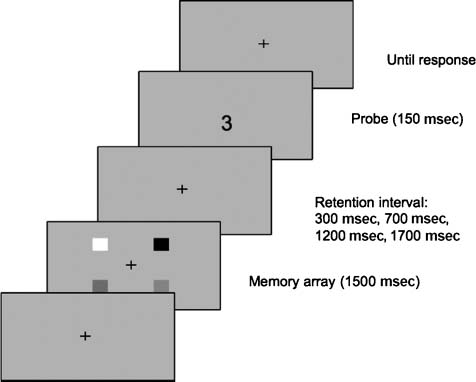
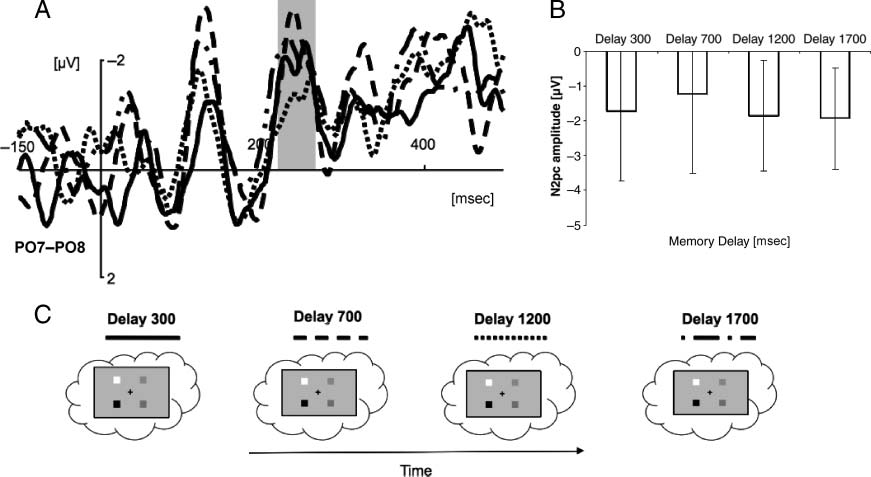
Chiffre 1. Schematic representation of the experimental trial sequence.
First a memory array consisting of four (Load 4) or two (Load 2) color
patches appeared for 1500 msec. Suivant, a blank display was presented
throughout the retention interval (300, 700, 1200, ou 1700 msec). Enfin,
a central probe was flashed for 150 msec, associatively indicating one
location. Participants were instructed to report the color that was
presented at the probed location in the memory array. Associating
probe numbers with locations was learned during a practice block
and balanced between subjects.
tention intervals to rely largely on the visuospatial represen-
tation of the stored information and to elicit larger N2pc
(and N3rs) component amplitude. The weaker the atten-
tional orienting to memorized array, the smaller compo-
nent amplitude was expected to be. Decreasing amplitude
with increasing retention interval would indicate gradual
recoding from a visuospatial form into a new format.
In Experiment 2, we used a verbal working memory task
to reveal the nature of this new representational format as
it emerged over time (cf., Luck & Vogel, 1997). The verbal
memory task should make recoding into a verbal format
more difficult by engaging the articulatory loop. If VSTM
representations are usually transformed into a verbal code
au fil du temps, this would be prevented here, and the attention
markers should be equally visible across retention inter-
vals. In that case, recall would have depended solely on
the visuospatial format.
EXPERIMENT 1
Methods
Apparatus
Four color patches placed on a plane of invisible square
(each stimulus placed 1° of visual angle away from center;
each patch was a square extending 1° of visual angle in
horizontal and vertical plane) served as stimuli in Experi-
ments 1 et 2. Colors used in the sample were green
(RGB 0, 255, 0), blue (RGB 0, 0, 255), red (255, 0, 0),
and yellow (255, 255, 0). Participants responded with the
654
Journal des neurosciences cognitives
Volume 24, Nombre 3
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
un
/
_
j
0
o
0
c
1
n
0
2
_
un
p
_
d
0
0
b
1
oui
0
g
2
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
t
.
F
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
index and middle fingers of the left and right hand using
four keys. The keys were “w,” “s,” “k,” and “o” on a
standard, international keyboard. Each of the keys was
associated with a single color. The retrieval cues were as-
signed to an invisible “Z” plane starting with Position 1 dans
the top left corner of the “Z”; Position 2 in the top right
corner, Pposition 3 in the bottom left corner, and Posi-
tion 4 in bottom right corner, for half of the participants.
For the other half the positions were placed on an in-
verted “Z” plane with Position 1 in the top right corner;
Position 2 in the top left corner; Position 3 in the bottom
right corner, and Position 4 in the bottom left corner.
Procedure
Participants (n = 12, âge moyen = 25 années, SD = 2.5 années)
viewed the memory array for 1500 msec and were asked
to memorize the colors and their locations. After various
retention intervals (300, 700, 1200, et 1700 msec), un
centrally presented probe was flashed for 150 msec, sym-
bolically indicating a location: A centrally presented digit
number indicated a particular location. Locations were
associated with numbers from a set of 1–4 during the prac-
tice block. Participants were asked to press as fast as pos-
sible (keeping accuracy above 90%) one of four buttons
indicating the color of the square presented at the probed
location. Pressing a response button triggered a feedback
screen followed after 200 msec by the next trial. The ex-
periment consisted of 14 blocks (64 trials each). In total,
a participant performed 896 experimental trials. Before
the experiment, a practice block of 64 trials had to be
completed.
EEG Recordings and Preprocessing
The EEG signal was recorded from 23 électrodes (sampling
frequency of 500 Hz, 0.1–125 Hz bandwidth, referenced
to Cz and re-referenced off-line to the average of all elec-
trodes), subsequently the signal was low-pass filtered
à 40 Hz and probe-locked. Epochs containing eye move-
ments or eye blinks were excluded from analysis (for the
procedure, see Wykowska & Schubö, 2010).
Statistical Analysis
Separate repeated measures ANOVA were computed on
the mean voltages time-locked to the centrally presented
probe for the N2pc, SPCN, N3rs, and P3. The N2pc was
measured from electrodes ipsilateral and contralateral to
the probed item location in the memory template within
a 220–280 msec time window at parieto-occipital electrode
pairs (PO7–PO8 and PO3–PO4). The ANOVA included
the factors electrode laterality (ipsilateral, contralateral to
the position where the target had been presented in mem-
ory array), load (2 items, 4 items), retention interval (clus-
tered short: 300 et 700 averaged together, clustered
long: 1200 et 1700 averaged together), and electrode pair
(PO7–PO8, PO3–PO4). A repeated measure ANOVA on
the SPCN amplitude was measured within 300–400 msec
poststimulus and also included the factor delay (300, 700,
1200, 1700 msec), laterality (contralateral, ipsilateral) et
load (2 items, 4 items). The ANOVA for the N3rs mea-
sured 300-400 msec poststimulus over four (AFz, Fz, FCz,
CPz) midline electrodes (cf. Nobre et al., 2008) and con-
tained the factors electrode, load (2 items, 4 items), et
retention interval (300, 700, 1200, 1700). A repeated mea-
sures ANOVA on P3 amplitude was measured within 280–
380 msec poststimulus and included the factors delay (300,
700, 1200, 1700 msec) and electrode (Pz, POz; cf. Akyürek,
Leszczyński, et coll., 2010; Vogel, Luck, & Shapiro, 1998).
Results
Attentional Orienting across Loads Suggests Nonspatial
Load Costs
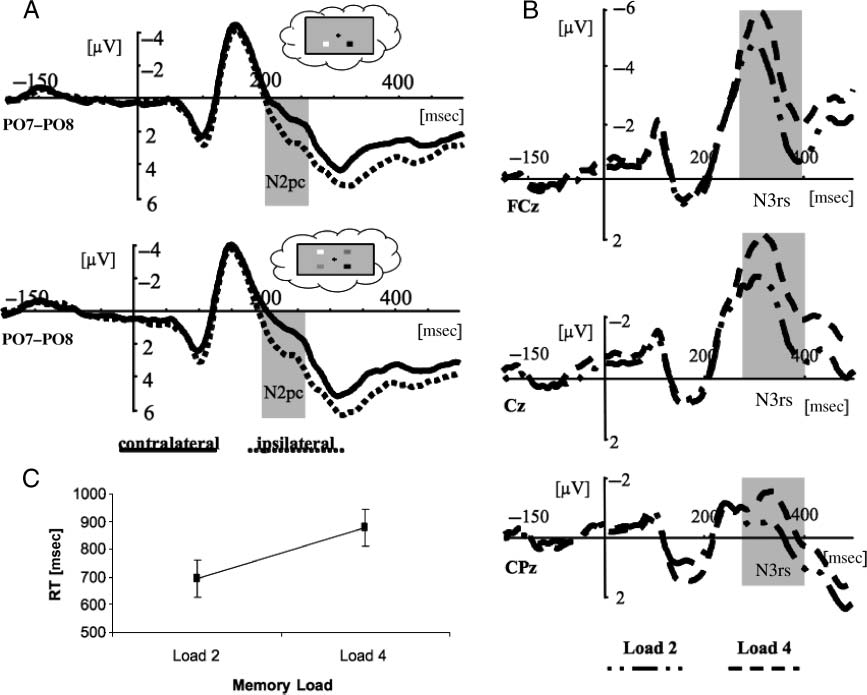
Figure 2A presents contralateral and ipsilateral activity
measured at parieto-occipital electrodes. The main effect
of Electrode Laterality [F(1, 11) = 6.58; p < .05] confirmed
the presence of a pronounced N2pc as expected from pre-
vious research in visual search (Eimer, 1996) and memory
search (DellʼAcqua et al., 2010; Eimer & Kiss, 2010; Kuo
et al., 2009). Subtracting ipsilateral (3.21 μV) from contra-
lateral (1.90 μV) activity resulted in a negativity (−1.31 μV),
marking attentional orienting to the location where the
target had been presented in the memory array. Notice
that the N2pc was triggered by a centrally presented visual
probe, which ensures that the negative deflection is only
the result of orienting to a representation in memory and
not orienting to any actual visual stimulation. This sug-
gests that the centrally presented probe triggered a shift
of spatial attention within VSTM. None of the further main
effects nor the interaction of factors was reliable. Although
memory orienting (as marked by the N2pc amplitude) was
not affected by load, the N3rs component, which has been
found to reflect retroactive target selection mechanisms
(Kuo et al., 2009; Nobre et al., 2008), showed a greater
negativity for Load 4 than for Load 2 [F(1, 11) = 20.00,
p < .001]. Figure 2B plots mean amplitude of the N3rs
component over representative midline electrodes. The
main effect of load revealed that Load 4 was accompanied
by a greater negativity than load 2 (−2.92 vs. −1.94 μV).
This replicates a previously observed pattern (Kuo et al.,
2009) with a load-dependent N3rs over midline elec-
trodes (Figure 2B) accompanied by a load-independent
N2pc (Figure 2A).
Gradual Recoding of the Visuospatial Format into a
Different Type of Representation
Figure 3A shows difference waves (contralateral minus
ipsilateral) plotted separately for the four delays. The mean
amplitudes and a potential interpretation are presented
Leszczyński et al.
655
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
a
/
_
j
0
o
0
c
1
n
0
2
_
a
p
_
d
0
0
b
1
y
0
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
a
/
_
j
0
o
0
c
1
n
0
2
_
a
p
_
d
0
0
b
1
y
0
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. The memory load effect. (A) Contralateral (solid line) and ipsilateral (dotted line) waveforms as a function of time (msec) plotted
separately for memory Loads 2 and 4. Analyses were time-locked to the onset of the centrally presented probe. The N2pc contralateral minus
ipsilateral time window that entered statistical analysis is depicted with a gray bar (for all plots and panels). (B) N3rs waveforms for Load 2
(dashed-dotted line) and Load 4 (dashed line) plotted separately for three representative midline electrodes. (C) Behavioral results of the
experimental task showing RTs plotted separately for two memory loads. The error bars depict SEM.
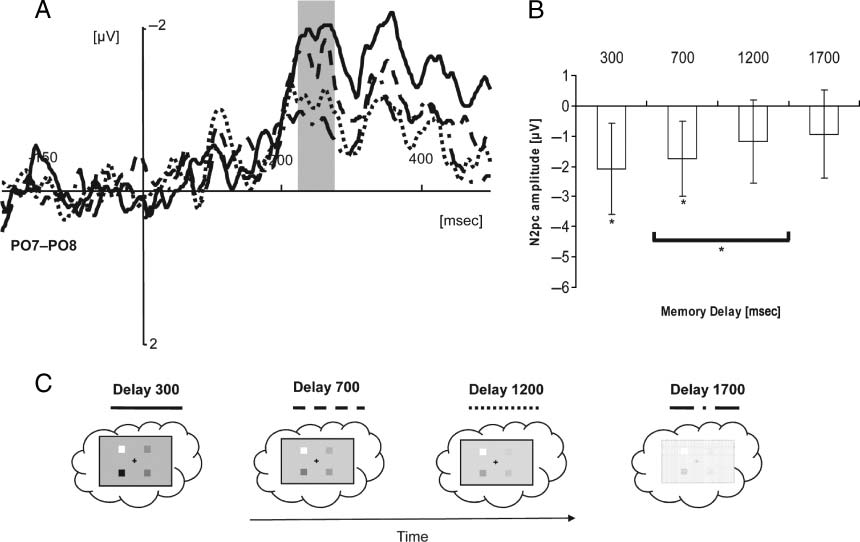
in Figure 3B and C, respectively. The negative deflection in
the N2pc time window confirmed that observers were re-
calling by means of attentional orienting within VSTM. An
interaction of electrode laterality and retention interval
[F(1, 11) = 4.98, p < .05] showed that the amplitude of
the N2pc was more pronounced at short delays than at
long delays (−1.65 μV vs. −0.972 μV, respectively; see Fig-
ure 3A and B). The decrease of N2pc amplitude as a func-
tion of retention interval reflects a decrease in the strength
of attentional orienting over time. It is worth to notice that
there was no significant N2pc at longest intervals. The
longer the interval between memory template presenta-
tion and the probe, the less spatial attention seemed re-
quired for recall. This leads us to conclude that the initial
spatial format of the VSTM representation is gradually re-
coded into another form. We further computed the SPCN
component, which revealed that laterality did not interact
significantly with any other factor. However, post hoc com-
parison reflected there was a reliable SPCN component,
but only at the shortest [t(11) = 2.41, p < .05] delay
(300 msec: −1.48 μV; 700 msec: −0.80 μV; 1200 msec:
−0.65 μV; 1700 msec: −0.83 μV). This could again re-
flect preserved spatial layout of memory representation
at shortest retention interval and gradual recoding with
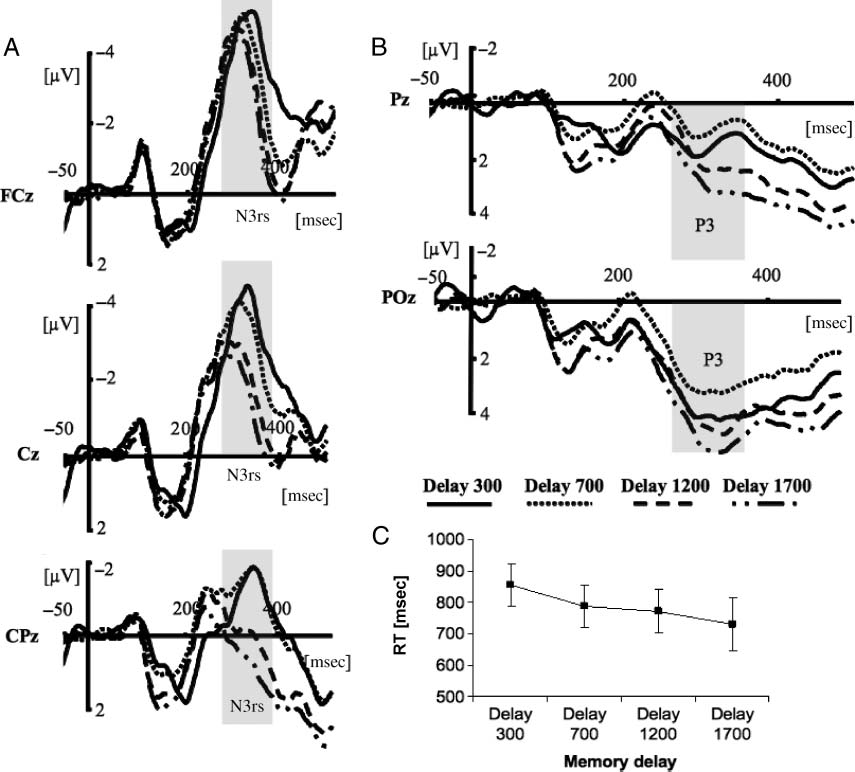
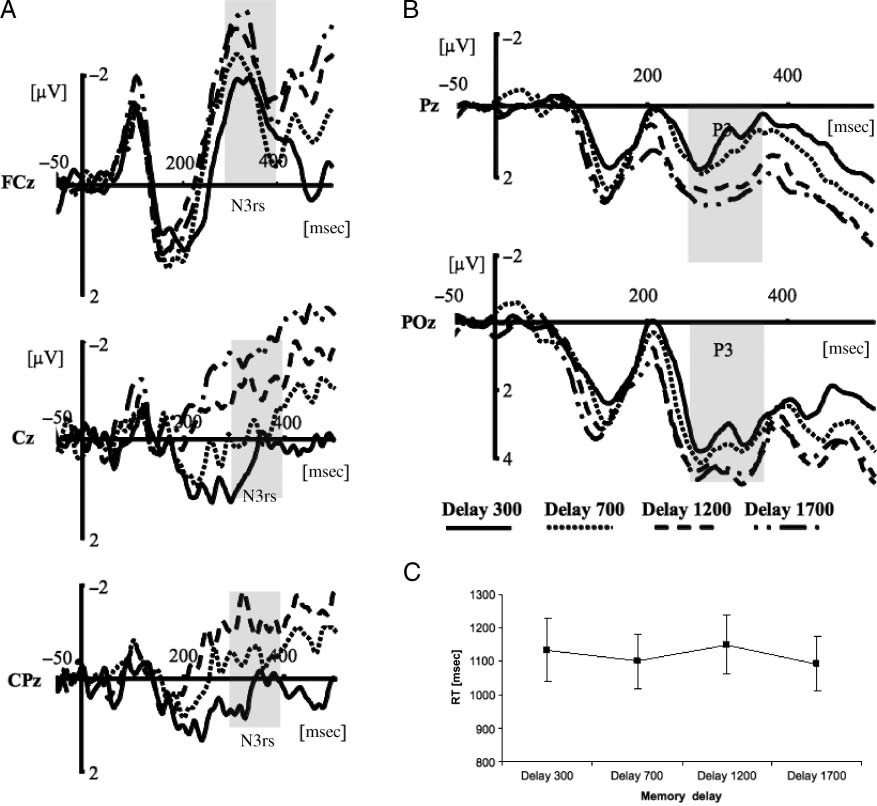
increasing delay. Interestingly, the amplitude of the N3rs
component (Figures 4A and 5A), thought to reflect retro-
active spatial target selection (Kuo et al., 2009; Nobre et al.,
2008), showed a similar pattern. It also decreased as a
function of delay [F(3, 33) = 10.07, p < .001]. Post hoc
contrasts revealed that there was a decrease in negativ-
ity of the N3rs component from delays of 300–1700 msec
[F(1, 11) = 12.53, p < .005], 300–1200 msec [F(1, 11) =
14.03, p < .005], and 300–700 msec [F(1, 11) = 17.81,
p < .001]. Mean amplitudes were −3.95 μV (300 msec),
−2.31 μV (700 msec), −1.91 μV (1200 msec), and −1.56 μV
(1700 msec). The decreasing N3rs negativity suggested
that it becomes more and more difficult to select a spatial
target from the memory array with increasing retention in-
terval. These variations in difficulty could mirror a decrease
in the degree of retroactive attentional orienting and retro-
active spatial target selection (cf. Kuo et al., 2009; Nobre
et al., 2008). Such an interpretation fits well with earlier
findings of Nobre et al. (2008), where the N3rs was sus-
ceptible to the amount of spatial information conveyed
656
Journal of Cognitive Neuroscience
Volume 24, Number 3
by retro-cues. In the current experiment, the amount of
spatial information seemed to be changing over retention
intervals. Thus, the longer the retention interval between
memory array and probe, the more difficult retroactive
spatial target selection became. Taken together, results
reflected by the N2pc and N3rs component indicate that
the spatial memory representation is gradually being re-
coded into a new format, thus making attentional orient-
ing more difficult as a recall strategy.
The effect of increasing difficulty for both attentional
orienting in VSTM and retroactive spatial target selection
were accompanied by a surprising pattern of RTs (see Fig-
ure 4C). RT decreased as a function of delay [F(3, 33) =
30.000, p < .005]. Post hoc contrasts revealed that partici-
pants were slower in responding with a delay of 300 msec
than with any other delay. Significant decreases in RT were
observed between the delays of 300 versus 700 msec [F(1,
11) = 21.228, p < .001] and between the delays of 1200
versus 1700 msec [F(1, 11) = 18.257, p < .001] but not
between the delays of 700 and 1200 msec [p > .18]. Mean
RTs were 855 msec (delay of 300 msec), 786 msec (delay
de 700 msec), 771 msec (delay of 1200 msec), et 729 msec
(delay of 1700 msec). Overall error rate was 8%. Là
was main effect of load [F(1, 11) = 17.45, p < .05] with
Load 2 resulting in less errors than Load 4 [7% vs. 11%].
The main effect of retention interval was also significant
[F(3, 33) = 4.91, p < .05]. Planned comparison revealed
that the difference between delays of 1200 msec (11%)
and 1700 msec (8%) was significant whereas the other dif-
ferences were not (delay of 300 msec: 9% and delay of
700 msec: 7%). A significant interaction between both
factors [F(3, 33) = 10.35, p < .001] indicated higher er-
ror rates for Load 4 at longer delays (14% for delay of
1200 msec and 9% for delay of 1700 msec) than at short
delays (12% for delay of 300 msec and 7% for delay of
700 msec) and no variation in error rate for Load 2 (delay
of 300, 700, 1200, and 1700 msec = 7%).
From the improvement in recall performance, one may
conclude that the memory search task became easier
with longer retention intervals. Given that the initial for-
mat of representation becomes gradually less susceptible
to attentional orienting as recall strategy, we hypothesized
that this initial memory representation was transformed
into some other, less spatial form. This new representa-
tion facilitated memory recall as indicated by the decrease
in RT. This claim was corroborated by the analysis of the
P3 component (cf. Figure 4B). P3 amplitude increased as
a function of delay [F(3, 33) = 9.447, p < .005]. Post hoc
contrasts revealed a significant increase in positivity from
700 to 1200 msec [F(1, 11) = 13.520, p < .005] and from
1200 to 1700 msec [F(1, 11) = 8.766, p < .05], but not
from 300 to 700 msec [p > .14]. The mean amplitude of
the P3 was 2.715 μV (300 msec), 1.973 μV (700 msec),
3.220 μV (1200 msec), et 3.933 μV (1700 msec). Précédent
research has related the P3 component to updating or
maintenance of information (Donchin & Coles, 1988).
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
un
/
_
j
0
o
0
c
1
n
0
2
_
un
p
_
d
0
0
b
1
oui
0
g
2
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
/
t
F
.
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Chiffre 3. Fading of spatial representation during retention. (UN) The N2pc difference waveforms (calculated as subtraction of the signal obtained
from electrodes ipsilateral and contralateral to the probed item location in memory) plotted separately for four levels of the memory delay (300 msec,
solid line; 700 msec, dashed line; 1200 msec, dotted line; 1700 msec, dash-dotted line). (B) Mean amplitude differences at PO7–PO8. (C) Un
interpretation of the results showing that spatial memory representation fades away with increasing memory delay.
Leszczyński et al.
657
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
un
/
_
j
0
o
0
c
1
n
0
2
_
un
p
_
d
0
0
b
1
oui
0
g
2
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
t
/
.
F
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Chiffre 4. Exchange of representations during the retention interval. (UN) The N3rs component plotted as a function of time (msec) separately
for four retention time intervals (delay of 300 msec, solid line; 700 msec, dotted line; 1200 msec, dashed line; 1700 msec, dash-dotted line).
Three electrodes over midline are presented. (B) P3 amplitude as a function of time (msec) plotted separately for four retention intervals observed
at two representative electrodes. (C) Behavioral results showing RTs plotted separately for four memory delays. The error bars depict SEM.
Attentional blink studies have associated the amplitude of
the P3 to the amount of information that reaches memory
(Vogel et al., 1998). Taken together, this could explain the
current pattern. The maintenance or updating phase in-
creased with increasing retention interval, in other words,
the longer the retention interval, the more time partici-
pants had for maintenance. This would explain the ob-
served increase in the P3 amplitude with longer delays.
En outre, increasing P3 amplitude has also been related
to the formation of a durable VSTM representation, a pro-
cess also known as consolidation (Akyürek, Leszczyński,
et coll., 2010; Vogel et al., 1998). Given that the retention in-
tervals varied from 300 à 1700 msec, the current P3 ampli-
tude increase could be explained by the gradual formation
of a durable VSTM representation, thereby marking the
process of consolidation. The current pattern reveals how
the initial spatial representation in VSTM may be replaced
by another, nonspatial representation. This nonspatial rep-
resentation seems to be developing during memory con-
solidation. Ainsi, the transition between the nature of the
N2pc, SPCN, and N3rs effects to those of the P3/RT effects
might be viewed as a correlate of gradual representational
change from a spatial to a non-spatial memory representa-
tion. To examine the nature of the representation that is
gradually built up during short-term consolidation, we con-
ducted Experiment 2.
EXPERIMENT 2
In Experiment 1, we found that the visuospatial format of
VSTM was recoded into some other, less spatially-specific
code. Experiment 2 was performed to reveal the nature
of the recoded VSTM representation. Given that the stim-
uli in Experiment 1 were easy to verbalize, one could
expect that the visuospatial format was recoded into a
verbal representation. To test whether a verbal code is
a possible format,1 we used a verbal working memory
658
Journal des neurosciences cognitives
Volume 24, Nombre 3
task, which was adapted from Luck and Vogel (1997), à
make it suitable for the EEG technique. This was done to
increase the difficulty of recoding spatial VSTM contents
into a verbal code. In case recoding of the visuospatial
code into a verbal code was actually part of the partici-
pantsʼ strategy to perform the task, then increasing re-
coding difficulty because of verbal working memory task
should lead to a different pattern of visuospatial ERP mark-
ers as found in Experiment 1. If the N2pc–SPCN–N3rs
pattern in Experiment 1 indicates gradual recoding into
a verbal format then increasing the recoding difficulty
should also affect the markers of memory search. Tel
modulations would indicate that loading verbal working
memory changed the recall strategy used by participants.
En plus, if recoding into a verbal code is associated
with efficient memory retrieval, the RT benefit for long
retention intervals observed in Experiment 1 should also
disappear in Experiment 2.
Methods
Procedure
Twelve participants (âge moyen = 24.87 années, SD =
3.57 années; eight women) participated in Experiment 2.
Unless stated differently, the method was the same as in
Experiment 1. The VSTM task was embedded within a
verbal working memory task. Participants were presented,
via stereo loudspeakers, a verbal sequence of three letters
(par exemple., UN, B, C sampled randomly without replacement from
the set of four letters A, B, C, and D) at the beginning of
each trial. Après 1000 msec, a VSTM search task like in Ex-
periment 1 was performed. Once participants responded
to the VSTM task, they were asked to recall the verbal
memory content by repeating the letters in correct order.
The load condition was dropped in Experiment 2 (only
load 4 was used) for efficiency reasons. Furthermore and
to make verbal coding even more difficult, the memory
array presentation time was set to 300 msec.
Separate repeated measures ANOVAs were computed
on mean amplitudes time-locked to the centrally presented
probe for the N2pc, SPCN, N3rs, and P3. The same time
windows and electrodes were selected as in Experiment 1.
Results
Recoding VSTM Content from a Visuospatial into a
Verbal Representation
Figure 5A presents mean amplitude of the lateral differ-
ence waves (contralateral minus ipsilateral) plotted as
a function of retention interval. Similar to Experiment 1,
the main effect of laterality was significant [F(1, 11) =
6.561, p < .05]. This replicates the result from Experi-
ment 1, demonstrating a significant N2pc component to
a centrally presented probe. Interestingly, unlike Experi-
ment 1, laterality and retention interval did not interact
significantly (F < 1). The linear decrease in amplitude of
the N2pc was not observed in Experiment 2 (see Figure 2
for Experiment 1 and Figure 5 for the pattern in Experi-
ment 2). The sustained N2pc amplitude across retention
intervals suggests that the visuospatial representation was
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
a
/
_
j
0
o
0
c
1
n
0
2
_
a
p
_
d
0
0
b
1
y
0
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 5. Sustained spatial representation during retention with suppressed verbal recoding (Experiment 2). (A) The N2pc difference waveforms
(contralateral minus ipsilateral to the probed item location in memory template) plotted for four delays (300 msec, solid line; 700 msec, dashed
line; 1200 msec, dotted line; 1700 msec, dash-dotted line). (B) Mean amplitude differences at PO7–PO8. (C) An interpretation of the data
showing that spatial memory format does not fade away with increasing memory delay.
Leszczyński et al.
659
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
a
/
_
j
0
o
0
c
1
n
0
2
_
a
p
_
d
0
0
b
1
y
0
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 6. Sustained visuospatial representation during retention interval. (A) The N3rs component plotted separately for four (delays of 300 msec,
solid line; 700 msec, dotted line; 1200 msec, dashed line; 1700 msec, dash-dotted line) retention intervals (Experiment 2). (B) P3 amplitude
observed for four retention intervals (Experiment 2). (C) Behavioral results showing RTs plotted for four memory delays (Experiment 2). The
error bars depict SEM.
not recoded into a new format while the verbal working
memory task was used. A repeated measures ANOVA com-
puted on the SPCN data revealed a main effect of Lateral-
ity [F(1, 11) = 8.34, p < .01] with ipsilateral (4.12 μV)
subtracted from contralateral (3.52 μV) potentials indicat-
ing a significant SPCN (−1.40 μV). Similar to Experiment 1,
laterality did not interact with retention interval indicating
no significant changes in amplitude of the SPCN as a func-
tion of retention interval.
Figure 6A plots mean amplitudes of the N3rs component
as a function of retention interval separately for three mid-
line electrodes. A repeated measures ANOVA performed
on the N3rs revealed a significant main effect of Retention
Interval [F(3, 33) = 4.232, p < .01], indicating an increas-
ing negativity as a function of retention interval. The N3rs
negativity rose from delay of 300 msec (−0.62 μV), across
delay of 700 msec (−0.81 μV) and delay of 1200 msec
(−1.75 μV) up to delay of 1700 msec (−1.971 μV). This
effectively contrasted with the results from Experiment 1.
Given that the N3rs is susceptible to the amount of spa-
tial information represented in memory, it might reflect
the persistent spatial coding of VSTM contents in Experi-
ment 2. Having suppressed the possibility of recoding con-
tents into a verbal code, spatial coding seems to have
been maintained to serve as recall mechanism even at the
longest intervals.
Figure 6C shows behavioral performance in Experi-
ment 2. A repeated measure ANOVA with Greenhouse–
Geisser correction for the RT data revealed no significant
effect of Retention Interval, although response times dif-
fered numerically between conditions. Contrary to the
findings in Experiment 1, no linear trend was observed
(cf. Figures 4C and 6C). For error rates, a repeated mea-
sures ANOVA with Greenhouse–Geisser correction de-
termined a significant main effect of Retention Interval
[F(1.841, 23.934) = 8.703, p < .05]. A contrast analysis
660
Journal of Cognitive Neuroscience
Volume 24, Number 3
revealed that delay of 1200 msec resulted in the highest
error rate (15%) as compared with 8% (delay of 300 msec),
7% (delay of 700 msec), and 9% (delay of 1700 msec). No
sign of linear improvement of performance was observed
either in RT or in error rates.
A repeated measures ANOVA on the P3 data revealed
a significant main effect of Retention Interval [F(3, 33) =
4.039, p < .01]. Similar to Experiment 1, the positivity
increased linearly from delay of 300 msec (1.880 μV )
to 700 msec (2.202 μV ), 1200 msec (2.939 μV ), and
1700 msec (3.137 μV). The main effect of Electrode was
also significant [F(1, 11) = 33.651, p < .001], with greater
positivity over electrode POz (3.59 μV) than Pz (1.49 μV).
The interaction between factors was not significant (see
Figure 6B).
DISCUSSION
In the present study we were able to show a dynamic
representational change in memory. Initially objects were
stored in visuospatial form. However, as the retention
interval increased the spatial form was gradually replaced
by a verbal representation. Using electrophysiological mea-
sures together with a memory search task and varying the
retention interval between memory array presentation and
the recall probe stimulus, we were able to study the neural
basis for representational recoding in VSTM. We observed
an initial representation that is suited to attentional orient-
ing at the shortest retention intervals. However, with an
increasing interval, this early representation was gradually
recoded into a verbal form, indicating that both attentional
orienting to VSTM (N2pc) and spatial retroactive target
selection (N3rs) got suppressed with increasing delay. The
small (but nonsignificant) N2pc or N3rs at the longest in-
terval in Experiment 1 do not contradict this claim, as we
are suggesting a gradual representational change that may
not follow the same time course on every trial. Interestingly,
behavioral performance as well as the amplitude of the P3
in Experiment 1 showed an inverse pattern (see Figure 4C
and B). The P3 has been related to updating of information
in working memory (Nobre et al., 2008; Vogel et al., 1998).
This updating process has been interpreted as leading to
the formation of a more durable, consolidated short-term
representation ( Vogel et al., 1998). The current increase
in P3 with an increased delay suggests that with a longer
retention interval updating was also prolonged, which was
reflected by an enlarged P3. This prolonged updating pro-
cess may have allowed for a more consolidated, durable
representation at long delays. Interestingly, the increase in
P3 amplitude was also observed in Experiment 2, without
the improvement in behavioral performance at longer
intervals. Given that recoding into verbal code was sup-
pressed in that experiment, the P3 seems to reflect the
formation of a more abstract representation, irrespective
of the code in use. Together the results from the two ex-
periments led us to conclude that the initial visuospatial
memory representation was replaced by a verbal memory
representation during the consolidation process.
Alternatively one may explain the increasing amplitude
of the P3 component and the decreasing RT observed with
longer retention intervals as a consequence of response
probability (Brookhuis, Mulder, Mulder, & Gloerich, 1983;
Bertelson & Boons, 1960) as the likelihood of having to give
a response gets larger as a function of retention interval. As
in Experiment 2, the same probabilities were used as in
Experiment 1 and no linear decrease in RT was observed,
it seems unlikely that probability differences could explain
the current result pattern. Modulations of the P3 component,
however, may also be attributed to changes in response
probabilities over delay.
The attentional orienting hypothesis (Kuo et al., 2009;
Lepsien & Nobre, 2007; Griffin & Nobre, 2003) proposes
that recall is performed by shifting attention toward spa-
tially organized memorized items. Our results extend the
current attentional orienting hypothesis, which does not
predict additional VSTM recall mechanisms for spatial in-
formation. By providing an electrophysiological measure
of the dynamics of representational change in memory
recall, the current study speaks in favor of attentional
orienting as an initial strategy used only before recoding
of information into a verbal code. Using a verbal working
memory task, we demonstrated that, when possible, the
initial visuospatial representational format was gradually
recoded into verbal information as the retention interval
increased. Additionally, as revealed by RT data, the bene-
fit from recoding at long retention intervals was not ob-
served when the articulatory suppression task was used.
Together this led us to conclude that verbal recoding is
associated with a representation that is more efficient in
guiding the recall process.
The fact that visuospatial information is getting grad-
ually recoded into verbal form with an increasing retention
interval may be linked to concepts of temporal integra-
tion. Studies using the temporal integration task (Akyürek,
Schubö, & Hommel, 2010; Jiang, 2004) and the attentional
blink task (Raymond, Shapiro, & Arnell, 1992) have shown
evidence for the existence of a temporal window in which
consolidation is expected to happen. Here we provide
evidence for consolidation by revealing correlates of the
representational change of memory content throughout
the consolidation process. We show that the early, precon-
solidated representation is spatiotopic and that the spatial
layout is still preserved at that time. This spatiality of early
representation is gradually recoded into a verbal form over
time. This could serve as a mechanism that closes the win-
dow of opportunity for temporal integration. Paradigms
that are used to study temporal integration often require
combining spatially distinct information across successive
presentations to form a coherent percept (Akyürek, Schubö,
et al., 2010; Jiang, 2004). Here, we show that with increas-
ing retention interval the spatiality of the memorized repre-
sentation is gradually recoded into verbal form, making
spatial information less obvious. This could be one reason
Leszczyński et al.
661
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
a
/
_
j
0
o
0
c
1
n
0
2
_
a
p
_
d
0
0
b
1
y
0
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
why integration is mostly observed at short time delays,
when the spatial format is still available ( Jiang, 2004).
Further consolidation brings up a verbal representation
that prevents orienting as a recall strategy. Lacking spatial
information, the late form of VSTM representation might
also prevent successful temporal integration at long delays
(Akyürek, Schubö, et al., 2010; Jiang, 2004). As the spatial
information about the initial stimulus array in a sequence
is not available anymore, the following array, although
being represented in spatial form itself, cannot be com-
bined with the representation of the initial array. Thus,
the observed representational change might also provide
a limit for temporal integration in VSTM.
The decreasing amplitude of the N2pc and the N3rs
could also reflect changes in the way attention operates
in VSTM over retention intervals. For example, Zhang and
Luck (2009) reported that a representation of feature–
location binding fades away earlier than the feature (i.e.,
color or shape) itself. Participants might still represent the
red color of stimuli which they are asked to recall, but the
missing feature–location binding prevents them from ac-
cessing this information. Thus, participants still might have
a representation of a feature after long delays, yet the fea-
ture–location bindings are missing, and therefore, they do
not know which item they are expected to report. Given the
results from Experiment 2 with sustained N2pc and increas-
ing N3rs amplitudes, we claim that locations are still repre-
sented in VSTM at long retention intervals. Using the verbal
working memory task to increase the difficulty of recoding
into a verbal form, we showed that it was possible to use
visuospatial attention to orient toward memorized locations
even at the longest intervals. Along the same line, Treisman
and Zhang (2006) have reported that locations play an im-
portant role for maintenance and recall at the initial encod-
ing and that locations and object information are separated
in VSTM. The current results extend this view providing
evidence that locations could be used to recall information
even at longer intervals. This, however, holds only when a
more efficient (i.e., verbal) way of representing information
in VSTM is prevented.
Acknowledgments
This research was supported by the German Research Founda-
tion (DFG), by a grant to A. S. as part of the Excellence Cluster
“Cognition for Technical Systems” (CoTeSys; Project #433), and
by a scholarship to M. L. and N. M. by the Graduate School of
Systemic Neurosciences GSN-LMU.
Reprint requests should be sent to Marcin Leszczyński, Depart-
ment of Experimental Psychology, Ludwig Maximilian University,
Leopoldstr. 13, 80802 Munich, Germany, or via e-mail: leszczynski.
marcin@gmail.com.
Note
1. The authors would like to thank Werner Schneider for sug-
gesting verbal coding as a possible recoding strategy and, there-
fore, inspiring Experiment 2.
REFERENCES
Akyürek, E. G., Leszczyński, M., & Schubö, A. (2010). The
temporal locus of the interaction between working
memory and the attentional blink. Psychophysiology, 47,
1134–1141.
Akyürek, E. G., Schubö, A., & Hommel, B. (2010). Fast temporal
event integration in the visual domain demonstrated by
event-related potentials. Psychophysiology, 47, 512–522.
Astle, D. E., Nobre, A. C., & Scerif, G. (2010). Subliminally
presented and stored objects capture spatial attention.
Journal of Neuroscience, 30, 3567–3571.
Awh, E., Anllo-Vento, L., & Hillyard, S. A. (2000). The role of
spatial selective attention in working memory for locations:
Evidence from event-related potentials. Journal of Cognitive
Neuroscience, 12, 840–847.
Bertelson, P., & Boons, J. P. (1960). Time uncertainty and
choice reaction time. Nature, 187, 531–532.
Brookhuis, K. A., Mulder, G., Mulder, L. J., & Gloerich, A. B.
(1983). The P3 complex as an index of information processing:
The effects of response probability. Biological Psychology,
17, 277–296.
Chelazzi, L., Duncan, J., Miller, E. K., & Desimone, R. (1998).
Responses of neurons in inferior temporal cortex during
memory-guided visual search. Journal of Neurophysiology,
80, 2918–2940.
DellʼAcqua, R., Sessa, P., Toffanin, P., Luria, R., & Jolicœur, P.
(2010). Orienting attention to objects in visual short-term
memory. Neuropsychologia, 48, 419–428.
Desimone, R. (1998). Visual attention mediated by biased
competition in extrastriate visual cortex. Philosophical
Transactions of the Royal Society, Series B, Biological
Sciences, 353, 1245–1255.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of
selective visual attention. Annual Review of Neuroscience,
18, 193–222.
Donchin, E., & Coles, M. G. H. (1988). Is the P300 component
a manifestation of context updating? Behavioral and
Brain Sciences, 11, 357–374.
Eimer, M. (1996). The N2pc component as an indicator of
attentional selectivity. Electroencephalography and
Clinical Neurophysiology, 99, 225–234.
Eimer, M., & Kiss, M. (2010). An electrophysiological measure
of access to representations in visual working memory.
Psychophysiology, 47, 197–200.
Griffin, I. C., & Nobre, A. C. (2003). Orienting attention to
locations in internal representations. Journal of Cognitive
Neuroscience, 15, 1176–1194.
Jiang, Y. (2004). Time window from visual images to visual
short-term memory: Consolidation or integration?
Experimental Psychology, 51, 45–51.
Jolicœur, P., Brisson, B., & Robitaille, N. (2008). Dissociation
of the N2pc and sustained posterior contralateral negativity
in a choice response task. Brain Research, 1215, 160–172.
Jolicœur, P., & DellʼAcqua, R. (1998). The demonstration of
short-term consolidation. Cognitive Psychology, 36, 138–202.
Kuo, B.-C., Rao, A., Lepsien, J., & Nobre, A. C. (2009). Searching
for targets within the spatial layout of visual short-term
memory. Journal of Neuroscience, 29, 8032–8038.
Lepsien, J., & Nobre, A. C. (2007). Attentional modulation of
object representations in working memory. Cerebral
Cortex, 17, 2072–2083.
Logan, G. D. (1988). Toward an instance theory of
automatization. Psychological Review, 95, 492–527.
Luck, S. J., & Hillyard, S. A. (1994). Spatial filtering during
visual search: Evidence from human electrophysiology.
Journal of Experimental Psychology: Human Perception
and Performance, 20, 1000–1014.
662
Journal of Cognitive Neuroscience
Volume 24, Number 3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
a
/
_
j
0
o
0
c
1
n
0
2
_
a
p
_
d
0
0
b
1
y
0
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Luck, S. J., & Vogel, E. K. (1997). The capacity of visual
working memory for features and conjunctions. Nature,
390, 279–281.
Nobre, A. C., Griffin, I. C., & Rao, A. (2008). Spatial attention
can bias search in visual short-term memory. Frontiers
in Human Neuroscience, 1, 1–9.
Prime, D., DellʼAcqua, R., Arguin, M., Gosselin, F., & Jolicœur, P.
(2010). Spatial layout of letters in nonwords affects
visual short-term memory load: Evidence from human
electrophysiology. Psychophysiology, 48, 430–436.
Raymond, J. E., Shapiro, K. L., & Arnell, K. M. (1992).
Temporary suppression of visual processing in an RSVP task:
An attentional blink? Journal of Experimental Psychology:
Human Perception and Performance, 18, 849–860.
Soto, D., & Humphreys, G. W. (2006). Seeing the content
of the mind: Enhanced awareness through working
memory in patients with visual extinction. Proceedings
of the National Academy of Sciences, U.S.A., 103,
4789–4792.
Soto, D., & Humphreys, G. W. (2009). Automatic selection
of irrelevant object features through working memory:
Evidence for top–down attentional capture. Experimental
Psychology, 56, 165–172.
Soto, D., Humphreys, G. W., & Heinke, D. (2006). Working memory
can guide pop-out search. Vision Research, 46, 1010–1018.
Treisman, A., & Zhang, W. (2006). Location and binding
in visual working memory. Memory & Cognition, 34,
1704–1719.
Vogel, E. K., Luck, S. J., & Shapiro, K. L. (1998).
Electrophysiological evidence for a post-perceptual locus
of suppression during the attentional blink. Journal of
Experimental Psychology: Human Perception and
Performance, 24, 1656–1674.
Vogel, E. K., & Machizawa, M. G. (2004). Neural activity
predicts individual differences in visual working memory
capacity. Nature, 428, 748–751.
Vogel, E. K., McCollough, A. W., & Machizawa, M. G. (2005).
Neural measures reveal individual differences in controlling
access to working memory. Nature, 438, 500–503.
Wykowska, A., & Schubö, A. (2010). On the temporal relation
of top–down and bottom–up mechanisms during guidance
of attention. Journal of Cognitive Neuroscience, 22,
640–654.
Zhang, W., & Luck, S. J. (2009). Sudden death and gradual
decay in visual working memory. Psychological Science,
20, 423–428.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
3
2
4
6
/
5
3
3
/
1
6
9
5
4
3
4
/
0
1
5
7
8
7
o
9
c
9
n
8
_
4
a
/
_
j
0
o
0
c
1
n
0
2
_
a
p
_
d
0
0
b
1
y
0
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Leszczyński et al.
663