Real-time Functional Architecture of Visual
Word Recognition
Caroline Whiting1,2, Yury Shtyrov2,3,4,5, and William Marslen-Wilson1,2
Abstrait
■ Despite a century of research into visual word recognition,
basic questions remain unresolved about the functional architec-
ture of the process that maps visual inputs from orthographic
analysis onto lexical form and meaning and about the units of
analysis in terms of which these processes are conducted. Ici
we use magnetoencephalography, supported by a masked prim-
ing behavioral study, to address these questions using contrasting
sets of simple (walk), complexe (swimmer), and pseudo-complex
(corner) forms. Early analyses of orthographic structure, detect-
able in bilateral posterior temporal regions within a 150–230 msec
time frame, are shown to segment the visual input into linguistic
substrings (words and morphemes) that trigger lexical access
in left middle temporal locations from 300 msec. Ceux-ci sont
primarily feedforward processes and are not initially constrained
by lexical-level variables. Lexical constraints become significant
depuis 390 msec, in both simple and complex words, with in-
creased processing of pseudowords and pseudo-complex forms.
Ces résultats, consistent with morpho-orthographic models
based on masked priming data, map out the real-time functional
architecture of visual word recognition, establishing basic feed-
forward processing relationships between orthographic form,
morphological structure, and lexical meaning. ■
INTRODUCTION
A neurocognitive account of visual word recognition—the
core process underpinning human reading—needs to
address two basic questions: What is the functional archi-
tecture of the recognition process, whereby visual inputs
are mapped via orthographic analysis onto representations
of lexical form and meaning, and what are the units of
analysis—lexical or sublexical—in terms of which these
processes are conducted? Despite an enormous research
effort over the last 100 années, involving behavioral, neuro-
psychologique, and neuroimaging techniques, there is no
agreed answer to these questions (Frost, 2012). Although
it is generally accepted that the initial analysis of visual form
and orthography engages occipitotemporal cortex, most
strongly on the left (par exemple., Vinckier et al., 2007; Cornelissen,
Tarkiainen, Helenius, & Salmelin, 2003; Cohen et al., 2000;
Bentin, Mouchetant-Rostaing, Giard, Echallier, & Pernier,
1999), and that later stages of lexical access and interpreta-
tion involve middle temporal and frontotemporal regions,
also primarily on the left (par exemple., Lau, Phillips, & Poeppel,
2008; Halgren et al., 2002; Bentin et al., 1999), the central
properties of this process remain unclear.
Here we use magnetoencephalography (MEG), dans
combination with MRI-based source reconstruction tech-
niques, to delineate the specific spatiotemporal patterns
1University of Cambridge, 2MRC Cognition and Brain Sciences
Unit, Cambridge, ROYAUME-UNI, 3Aarhus University, Denmark, 4Université de
Lund, Sweden, 5Higher School of Economics, Moscow
of neural activity elicited by a psycholinguistically rich
set of simple and complex written words and pseudo-
words. We aim to determine (1) under what description
the outputs of orthographic analysis are mapped onto
lexical-level representations and (2) what is the balance
between feedforward and feedback processes in the pro-
cessing relationship between orthographic and lexical anal-
ysis. Ce faisant, we will integrate behavioral data about
the performance characteristics of the system with direct
MEG-based evidence about its underlying neural dynamics.
Background
An important clue to the organization of visual word recog-
nition comes from masked priming research over the past
decade, demonstrating equally strong priming between
related pairs like hunter/hunt and lexically unrelated pairs
like corner/corn (par exemple., Marslen-Wilson, Bozic, & Randall,
2008; Longtin & Meunier, 2005; Rastle, Davis, & Nouveau,
2004; Longtin, Segui, & Halle, 2003; Rastle, Davis, Marslen-
Wilson, & Tyler, 2000). A masked prime like hunter is
assumed to prime hunt because it is decomposed into the
stem morpheme1 {hunt} and the grammatical morpheme
{-er}, reflecting the meaning of the whole form hunter.
The fact that significant priming is also seen for corner,
where a decompositional reading as {corn} + {-er} a
no relation to the meaning of the word, points to a process
of automatic decomposition for any word form that con-
tains potential morphological structure, regardless of the
lexical properties of the whole form. The failure of pairs like
© 2014 Massachusetts Institute of Technology Published under a
Creative Commons Attribution 3.0 Unported (CC PAR 3.0) Licence
Journal des neurosciences cognitives 27:2, pp. 246–265
est ce que je:10.1162/jocn_a_00699
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
un
/
_
j
0
o
0
c
6
n
9
9
_
un
p
_
d
0
0
b
6
oui
9
g
9
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
F
t
.
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
scandal/scan to show priming highlights the morphemic
basis for these effects. Although scan is a potential stem
morpheme, dal is not a grammatical morpheme, et
this seems to block the decomposition of scandal into
{scan} + {-dal}.
This pattern of results suggests a recognition process
that is dominated in its early stages by an analysis of the
orthographic input into sublexical morphemic units and
where a representation of the visual input in these terms
is projected onto the lexical level in a strongly bottom–up
manière, blind to lexical constraints (Marslen-Wilson et al.,
2008; Rastle & Davis, 2008). This morpho-orthographic ap-
proach is not, cependant, fully supported either behaviorally
(par exemple., Diependaele, Sandra, & Grainger, 2009; Feldman,
OʼConnor, & del Prado Martín, 2009) or in neuroimaging
studies of visual word recognition, where support can be
found for contrasting morphosemantic (or interactive)
approaches, which claim that early orthographic analysis
is modulated by top–down lexical and semantic constraints
(par exemple., Prix & Devlin, 2011).
Within the neuroimaging domain, we focus on studies
using EEG or MEG, because it is only these time-sensitive
methods that can resolve the specific temporal ordering of
different types of analysis during visual word recognition
and thus discriminate directly between different proposals
for the real-time functional architecture of the recognition
système. Recent research based on these techniques falls
broadly into two main classes. Several studies, stimulated
by the masked priming results, ask whether there is elec-
trophysiological evidence for early sensitivity to the mor-
phological content of visual word forms, independent
of lexical constraints. Working primarily with sets of mor-
phologically complex and pseudo-complex word forms,
masked priming has been combined with both EEG (par exemple.,
Morris, Grainger, & Holcomb, 2008; Lavric, Clapp, &
Rastle, 2007) and MEG (Lehtonen, Monahan, & Poeppel,
2011), whereas a further set of studies have used unprimed
lexical decision tasks (par exemple., Lavric, Elchlepp, & Rastle, 2012;
Lewis, Solomyak, & Marantz, 2011; Zwieg & Pylkkänen,
2009). Taken as a whole, these and similar studies provide
evidence for sensitivity to potential morphological struc-
ture, where complex and pseudo-complex forms like
farmer and corner initially group together relative to
orthographic controls like scandal, consistent with a
morpho-orthographic view where these processes are
not lexically driven. The spatiotemporal distribution of
these effects is quite diverse, both in terms of hemispheric
involvement (right and/or left) and posterior/anterior
location and in terms of timing, with early effects (150–
250 msec) seen in some studies (par exemple., Lavric et al., 2012;
Zwieg & Pylkkänen, 2009) and later effects (350–500 msec)
in others (par exemple., Lavric et al., 2007; Dominguez, de Vega, &
Barber, 2004).
A different set of MEG and EEG studies focus instead
on the earliness with which lexical and semantic effects
can be detected. These studies use unprimed lexical de-
cision tasks and contrast morphologically simple words
(nouns and verbs like help and gold) with matched pseudo-
words (par exemple., Hauk, Coutout, Holden, & Chen, 2012;
Hauk, Davis, Ford, Pulvermüller, & Marslen-Wilson,
2006; Assadollahi & Pulvermüller, 2003). Early lexical ef-
fects, although small relative to later N400 time frames,
have been reported in a range of posterior and middle
temporal sites. Hauk et al. (2012), Par exemple, report
word–pseudoword differences for the time period 180–
220 msec in left anterior middle and inferior temporal
lobes, whereas Shtyrov, Goryainova, Tugin, Ossadtchi,
& Shestakova, (2013) observe even earlier lexicality ef-
fects (at around 100 msec) in an EEG study using MMN
techniques.
It is hard to determine, cependant, what the implications
of these results are for the functional organization of
the word recognition process. This is partly because the
stimulus materials used rarely overlap across the two re-
search strands, with the lexically oriented work, for exam-
ple, generally not including morphologically complex
matériel. This means that there is little direct evidence,
under conditions where early lexical effects are detected,
whether these serve to modulate candidate morphological
decompositions—so that, Par exemple, the segmentation
of corner into {corn} + {-er} is inhibited.
A further issue is the use of overt response tasks in
combination with EEG and MEG, which all the studies cited
above have in common. Behavioral research into the dy-
namics of language function requires the use of these tasks
to provide information about underlying cognitive pro-
cesses. There are several concerns, cependant, that argue
against their use in the neuroimaging context. The most
salient of these is the evidence that such tasks can modu-
late the actual process under investigation, through atten-
tional tuning of neuronal computations relevant to the
task requirements, even at early stages of the cortical analy-
sis of sensory inputs (par exemple., Zanto, Rubens, Thangavel, &
Gazzaley, 2011; van Atteveldt, Formisano, Goebel, &
Blomert, 2007). This raises the possibility that early effects
seen in the EEG or MEG studies are induced by the ubiqui-
tous experimental task. Such concerns are compounded
when a priming task is used, especially in masked priming,
where prime and target overlap closely in time. Under such
conditions, it is hard to assign neural effects separately to
the properties of the prime, the target, or to interactions
between them at different levels of visual analysis.
We address these issues in the current study by (un)
ensuring that evidence about the timing of lexical and
morphological effects can be linked within the same ex-
periment to evidence about the spatiotemporal organiza-
tion of word recognition more generally; (b) presenting
the materials in a simple viewing paradigm, reducing the
likelihood that the experimental situation will induce
attentional tuning of specific aspects of the input analysis
processus; et (c) conducting a separate behavioral masked
priming study so that the functional properties of critical
stimulus materials can inform the analysis of the MEG re-
sponses evoked by a parallel set of stimuli.
Whiting, Shtyrov, and Marslen-Wilson
247
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
un
/
_
j
0
o
0
c
6
n
9
9
_
un
p
_
d
0
0
b
6
oui
9
g
9
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
/
.
F
t
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Experimental Considerations
This experiment explores the dynamic roles of morphologi-
cal, lexical, and semantic variables in the mapping between
prelexical orthographic processing and semantically sen-
sitive lexical analysis. To define the spatiotemporal co-
ordinates of these twin poles of the word recognition
process—for these stimulus sets and these participants in
this specific experimental context—we contrast morpholog-
ically simple words (par exemple., corn), pseudowords (par exemple., frum),
and length-matched consonant strings (par exemple., wvkp). These
simple forms, derived from the complex words and pseudo-
words (par exemple., corner, frumish) used elsewhere in the experi-
ment, establish the anchor points of the recognition process
using items that come from orthographic neighborhoods
matched across the main experimental conditions. Con-
trasts between words and pseudowords versus consonant
strings (par exemple., Cohen et al., 2000) should capture early ortho-
graphic effects in occipitotemporal cortex, differentiating
word-like forms from random letter strings. The same sets
of simple words and pseudowords allow us to locate the
other pole of the processing continuum, testing for lexicality
effects in a word versus pseudoword contrast. Ceux-ci sont
likely to be seen later in the access process—possibly in
the N400 time frame (par exemple., Lau et al., 2008)—with differential
responses in left-lateralized middle and anterior temporal
régions.
To evaluate the properties and timing of the interven-
ing processes that link orthographic analysis to lexical
representation, we present complex and pseudo-complex
stimuli that vary in morphological and lexical status. Le
morphological dimension, varying the presence or absence
of stems and affixes in potentially complex forms, asks
whether the mapping from orthographic analysis onto
lexical form and meaning is in terms of morphemic (ou
pseudo-morphemic) units (cf. Vinckier et al., 2007). Be-
cause simple words in English are always also morphemes,
this can only be tested by using complex forms that can
pull apart the lexical and morphemic properties of a given
word form—whether they are made up of potential stems
and affixes, as in farmer or brother, or whether they com-
bine an existing affix (par exemple., {-ish}) with a pseudo-stem, as in
blemish, or an existing stem (par exemple., {scan}) with a pseudo-
affix, as in scandal. From a lexical point of view, forms
like brother, blemish, and scandal are monomorphemic
and nondecompositional and should be treated differ-
ently from genuinely complex forms like farmer. From a
morpho-orthographic perspective, the form brother,
analyzable into the potential stem and affix pair {broth} +
{-er}, should behave differently from blemish and scandal
in the early stages of lexical access but similarly to farmer.
It is necessary here to treat derivational morphology
(the main focus of masked priming research) separately
from inflectional morphology, which involves word forms
like played that contain a stem and an inflectional affix
(the past tense {-éd}). Regular inflectional morphology
is systematic and transparent and does not change the
meaning of the stem. Inflected forms are argued to be
processed and represented decompositionally, relying on
a left-lateralized frontotemporal network (Marslen-Wilson
& Tyler, 2007). In contrast, derivational morphemes
change the meaning and often the grammatical category
of the stem, with a much less predictable relationship
between stem and whole form and where emerging neuro-
imaging evidence suggests that these may not be repre-
sented decompositionally (Bozic, Tyler, Su, Wingfield, &
Marslen-Wilson, 2013). Morphological structure of both
types should be parsed before lexical access based on the
presence of a stem and affix, but with potentially different
lexical outcomes.
For derivational morphology, we contrast a set of poten-
tially complex words (see Table 1) that have a stem and
affix (par exemple., farmer), a stem but no affix (par exemple., scandal ),
an affix but no stem (par exemple., blemish), and neither stem nor
affix (par exemple., biscuit). These contrasts test whether initial
morphological decomposition depends on the presence
of both a stem and an affix. Against the same backdrop
of morphologically simple forms (biscuit), we can also
evaluate the pattern of effects for inflected words like
blinked. All forms containing a potential stem and an affix
should trigger morphological segmentation, in contrast
to stem-only forms like scandal—which do not elicit a
masked priming effect—and simple forms like biscuit.
The effectiveness of an embedded affix (blemish) in trig-
gering morphological decomposition has not been tested
Tableau 1. Experimental Conditions and Example Stimuli
Condition
Exemple
Stem/
Affix
Semantic
Relatedness
Stem
Form
Derived
Transparent
farmer
+S+A
Pseudo-derived
corner
Pseudo-affix
blemish
+S+A
−S+A
+Sem
−Sem
n/a
farm
corn
blem
Inflected
Transparent
blinked
+S+A
+Sem
blink
Pseudo-inflected
ashed
+S+A
n/a
ash
Non-affixed
Pseudo-stem
No stem no affix
Pseudoword
Derived
Inflected
S = stem; A = affix.
scandal +S−A
−S−A
biscuit
−Sem
n/a
scan
bisc
frumish −S+A
−S+A
bected
n/a
n/a
frum
bect
248
Journal des neurosciences cognitives
Volume 27, Nombre 2
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
un
/
_
j
0
o
0
c
6
n
9
9
_
un
p
_
d
0
0
b
6
oui
9
g
9
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
t
.
F
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
in masked priming, although research with spoken words
shows that the presence of a potential inflectional affix does
trigger decompositional processes (Tyler, Stamatakis, Post,
Randall, & Marslen-Wilson, 2005). An inflectional pseudo-
word condition, where the affix is attached to a nonexistent
stem (par exemple., bected ), tests for this possibility in the visual
domain, matched by a further pseudoword set with deriva-
tional affixes (par exemple., frumish).
The second critical dimension of lexical status tests
competing claims for the degree of autonomy of the early
stages of visual analysis and lexical access. This dimension
contrasts semantically transparent forms like farmer with
opaque pseudo-complex pairs like corner. On a morpho-
orthographic account, no difference should be found be-
tween these forms before lexical access, but they should
begin to diverge once access to the meaning of the whole
form is in progress. To mirror the derivational contrasts,
we also include semantically transparent and opaque in-
flectional conditions. Because the inflectional equivalents
of corner words are rare in English (c'est à dire., ending in -ed with-
out being an adjective or past-tense form), we use inflected
pseudowords analogous to those tested by Longtin and
Meunier (2005) in French. Nouns that do not function as
verbs (par exemple., ash) were used as the stem, resulting in an
interpretable but nonexistent pseudoword like ashed. Both
semantically transparent inflected forms (blinked) et en-
flected pseudowords (ashed ) should generate early de-
compositional processing, as well as significant masked
priming.
En résumé, this study aims to specify the functional
architecture of visual word recognition by tracking the pat-
terns of neural activity that underlie processing of mor-
phologically simple and complex words in English. It asks
three main questions: (je) is the early output of orthographic
analysis structured into morphemic units, (ii) is there a
distinct processing phase at which potential morphological
structure is identified independent of lexical constraints,
et (iii) what is the timing with which these processes are
influenced by lexical-level constraints? To relate the MEG
results directly to the behavioral evidence for morpho-
orthographic processing, we will run a separate masked
priming study on parallel sets of complex and pseudo-
complex materials. Enfin, as noted earlier, participants
are tested in a simple word viewing situation, accompanied
by an occasional recognition task to reinforce sustained
attention to the stimuli.
MÉTHODES
MEG Participants
Sixteen participants (nine women) took part in the MEG
experiment. All were right-handed native British English
speakers between the ages of 18 et 35 (mean age of
25) with normal hearing, normal or corrected-to-normal
vision, and no history of neurological disease, who gave
written consent to take part and were paid for their time.
Stimuli and Design
In each of the nine MEG test conditions (see Table 1),
50 words were selected, which contrasted the presence
of different morphological features. Four conditions con-
tained a potential derivational affix. Three of these were
real word conditions: semantically transparent ( farmer),
pseudo-derived (corner), and pseudo-affix (blemish),
plus a pseudoword condition ( frumish), where the
stem was not a real word. Three conditions contained a
potential past-tense inflectional {-éd} affix: semantically
transparent (blinked ) and pseudo-inflected (ashed )
words, paired with a pseudoword condition (bected )
where the stem is not a real word. The pseudo-inflected
items (ashed) contained an embedded word that is only
used as a noun in English, creating a pseudoword that
could be segmented into an existing stem and an existing
affix but which was not itself an existing word. The stems
chosen for this condition appeared in the Celex English
database (Baayen, Piepenbrock, & Gulikers, 1995) only
as a noun, and no instances (or a single instance only)
of use as a verb were found in the British National Corpus
(www.natcorp.ox.ac.uk/). Two baseline conditions were
included that did not contain a potential affix: pseudo-
stem (scandal ) and no stem/no affix (biscuit).
Participants also saw the 450 embedded stems and pseudo-
stems (or first syllables for words without embedded stems)
extracted from these complex forms. These were accom-
panied by 160 strings of random consonants, matched to
the length of the target items (both stems and whole forms),
and varying in length from three to nine letters. These were
included both as a general length-matched baseline and to
allow specific contrasts with word and pseudoword stimuli
to select out regions sensitive to orthographic structure.
For use as test items in the recognition task, a set of 50 filler
items (words, pseudowords, and consonant strings) étaient
also presented. An additional 10 filler items were included
as dummy items at the beginning of each block. The total
number of stimuli in the study was 1120 items.
For all conditions where an embedded stem was present,
pairs of items were presented to native English speakers
who rated the semantic relatedness between the two words
(par exemple., corner/corn) on a scale of 1–7 (unrelated to highly
related ). Test items selected for the morphologically trans-
parent condition were rated as 6.5 or above. For the pseudo-
derived and pseudo-stem conditions, test items were rated
comme 3.5 or below (weakly related). Items for the nine condi-
tions were selected using the Celex database and the condi-
tions were matched (Tableau 2) on whole form and stem
length, percentage of orthographic overlap between stem
and whole form (where applicable), and frequency of the
whole form and embedded stem (where applicable).
Behavioral Study
To provide a bridge between masked priming research and
the current study, we ran an initial set of stimuli (40 words
Whiting, Shtyrov, and Marslen-Wilson
249
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
un
/
_
j
0
o
0
c
6
n
9
9
_
un
p
_
d
0
0
b
6
oui
9
g
9
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
.
F
t
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Tableau 2. Stimulus Properties for MEG Study
Condition
Exemple
Length
Stem Length
% Overlap (Stem/ Word)
Freq ( Wordform)
Stem Freq ( Wordform)
Derived
Transparent
farmer
Pseudo-derived
corner
Pseudo-affix
blemish
Inflected
Transparent
blinked
Pseudo-inflected
ashed
Non-affixed
Pseudo-stem
scandal
No stem no affix
biscuit
6.6
6.4
6.6
6.5
6.4
6.3
6.3
4.4
4.4
n/a
4.4
4.4
4.3
n/a
0.68
0.70
n/a
0.69
0.68
0.69
n/a
9.6
11.1
8.3
12.6
n/a
10.7
12.0
24.6
21.1
n/a
15.5
16.7
17.4
n/a
per condition) in a conventional masked priming task,
using a prime-target SOA of 40 msec with whole forms as
primes ( farmer) and stem forms as targets ( farm). Ce
study was conducted to determine what pattern of priming
effects we would see for the particular combination of
derivations, inflections, real words, and pseudowords
chosen for this research (as listed in Table 1). Previous
studies had not included all of these conditions in a single
stimulus set, and no study in English (as far as we are aware)
has used pseudowords like “ashed” and “bected” as primes.
Behavioral evidence about which combinations of real and
pseudo-stems and affixes do or do not show priming is an
essential input to the MEG study and its analysis.
We tested 29 new participants (none of whom took part
in the MEG study), all right-handed native British English
speakers between the ages of 18–34 (mean age of 24).
Each trial began with a set of hashmarks as a premask,
which appeared in the center of the screen for 500 msec.
This was followed by the prime in the same location in
lower-case letters for 40 msec and which itself was imme-
diately followed by the target in uppercase letters. Le
experiment was run in a sound-proof, dimly lit room,
using a PC-compatible microcomputer using DMDX soft-
ware (Forster & Forster, 2003). Trial order was pseudo-
randomized online using DMDX software, with two items
from each condition appearing in each scrambling block
(one related prime and one unrelated prime from each
condition). Outliers (RTs over 1500 msec) were discarded,
accounting for 0.8% of the data.
Four conditions (blemish, bected, frumish, and biscuit)
have nonwords as targets (blem, bect, etc.) where priming
was neither expected (Forster & Davis, 1984), nor found
(Tableau 3). For the remaining five conditions, there was
a main effect of Condition (F1(4, 112) = 40.71, p < .001;
F2(4, 192) = 7.80, p < .001) and Prime Type (F1(1, 28) =
17.61, p < .001; F2(1, 192) = 45.10, p < .001) and an inter-
action between the two factors (F1(4, 112) = 4.67, p <
.005; F2(4, 192) = 3.24, p < .05). Priming was found for
the farmer, corner, blinked, and ashed conditions (all of
which contain an embedded stem and affix) but not for
the pseudo-stem Condition scandal (Table 3). In a sepa-
rate analysis, no significant difference was found between
the priming effects across the four +S+A conditions. There
was a main effect of condition (F1(3, 84) = 26.80, p < .001;
F2(3, 153) = 6.52, p < .001) and Prime Type (F1(1, 28) =
28.07, p < .001; F2(1, 153) = 22.64, p < .001), but no inter-
action (F1(3, 84) = 1.65, p = .19; F2(3, 153) = 2.27, p =
.09). This is in line with previous results in English (Rastle
et al., 2000, 2004) and, in the case of ashed, with predic-
tions based on Longtin and Meunier (2005). In the MEG
study, we predict different spatiotemporal patterns of
neural activity for complex forms that do or do not elicit
masked priming, modulated over time by their lexical
status.
MEG Procedure
For the MEG study, the stimulus materials (see Tables 1
and 2) were based on those used in the masked priming
study. To enhance the signal-to-noise ratio in the MEG
environment, 10 additional stimuli were added to each
condition, increasing the number of items to 50 per con-
dition (listed in Appendix 1). The stimuli were randomly
assigned to one of two blocks, each further divided into
five subblocks with the constraint that each subblock
contained five items from each condition and 16 conso-
nant strings (for a total of 106 items per subblock). Whole
forms and their equivalent stem forms (e.g., farmer and
farm) were placed in separate blocks and always appeared
in corresponding subblocks across the experiment (i.e.,
farmer in Subblock 1 of Block 1 and farm in Subblock 1
250
Journal of Cognitive Neuroscience
Volume 27, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
of Block 2). The order of the two blocks was alternated for
each participant, so that presentation of the whole form
and equivalent stem ( farmer and farm) were alternated,
with the stem appearing first for half of the participants.
The order of the five subblocks was randomized for each
participant in a cyclical order (e.g., subblock order 1-2-3-
4-5 for Participant 1, and 2-3-4-5-1 for Participant 2). This
preserved the order of the subblocks so that the repeated
stem and whole form were always separated by the same
number of subblocks, with a mean distance of 559 trials
(range of 448–670 trials). Trial order was randomized
within each subblock using E-Prime 1.0 software (Psychology
Software Tools, Inc.).
Each trial began with a fixation cross in the middle of
the screen for 500 msec to direct the attention of the par-
ticipant to the appropriate location on the screen. This was
followed immediately by presentation of the stimulus for
100 msec, centered at the same location. The short presen-
tation prevented participants from making saccades. A
blank screen was then presented for 1.4–1.6 sec, jittered
randomly for each trial, before the next stimulus appeared.
At the end of a subblock, a screen appeared asking if
the letter string indicated had been seen in that subblock.
Participants were instructed to make a response within
3000 msec using the button boxes. Ten items were used
in each recognition task over the 10 subblocks for a total
of 100 items (50 old/50 new). Each subblock was separated
by a break at the completion of the recognition task, and
participants could control the length of each break.
Participants sat in a dimly lit magnetically shielded room
(IMEDCO AG, Switzerland), viewing items as they were
presented on a screen at eye level. All stimuli were dis-
played in bold Arial font in black letters on a light gray
background. Participants received spoken and written
instructions about the task and were given 10 practice
items. They were instructed to read the items silently but
not to articulate or make any movements. Because each
subblock contained approximately 100 items, participants
were instructed not to attempt to memorize the items
but to simply attend to them. Participants did not make
button presses during blocks of trials but used two button
boxes (one in each hand) to perform the recognition task
at the end of each subblock. The experiment was run
using E-Prime 1.0 and lasted approximately 45 min.
MEG Acquisition
MEG data were continuously acquired at a sampling rate of
1000 Hz (passband 0.01–300 Hz), with triggers placed at
the onset of each stimulus. Neuromagnetic signals were
recorded continuously with a 306-channel Vectorview
MEG system (Elekta Neuromag, Helsinki, Finland). Before
recording, four electromagnetic coils were positioned on
the head and digitized using the Polhemus Isotrak digital
tracker system (Polhemus, Colchester, VT) with respect
to three standard anatomical landmarks (nasion, left and
right preauricular points). During the recording, the posi-
tion of the magnetic coils was tracked using continuous
Table 3. Mean RTs, Error Rates, and Priming Effect in Masked Priming Behavioral Pretest
Example (Prime–Target)
Related Prime (% Errors)
Unrelated Prime (% Errors)
Priming Effect (msec)
farmer–farm
corner–corn
blemish–blem
blinked–blink
ashed–ash
scandal–scan
biscuit–bisc
frumish–frum
bected–bect
549 (2.0)
583 (3.5)
690 (9.8)
546 (1.7)
579 (4.5)
608 (9.0)
655 (4.5)
637 (2.3)
648 (2.7)
577 (3.7)
599 (4.3)
690 (9.5)
579 (3.2)
600 (5.7)
612 (5.8)
649 (2.8)
638 (1.2)
650 (3.5)
28**
16*
0
33**
21**
4
−6
1
2
Condition
Derived
Transparent
Pseudo-derived
Pseudo-affix
Inflected
Transparent
Pseudo-inflected
Non-affixed
Pseudo-stem
No stem no affix
Pseudoword
Derived
Inflected
*p < .05.
**p < .01.
Whiting, Shtyrov, and Marslen-Wilson
251
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
head position identification, providing information on
the exact head position within the MEG dewar for later
movement correction. Four EOG electrodes were placed
laterally to each eye and above and below the left eye to
monitor horizontal and vertical eye movements.
MEG Preprocessing
Continuous raw data were preprocessed offline with the
MaxFilter (Elekta Neuromag) implementation of the sig-
nal-space separation technique with a temporal extension
(Taulu & Simola, 2006). Averaging was performed using the
MNE Suite (Athinoula A. Martinos Center for Biomedical
Imaging). Epochs containing gradiometer, magnetometer,
or EOG peak-to-peak amplitudes larger than 3000 fT/cm,
6500 fT, or 200 μV, respectively, were rejected. Trials were
averaged by condition with epochs generated from −100
to 500 msec from onset of the target word. Averaged data
were baseline corrected using −100 to 0 msec interval and
low-pass filtered at 45 Hz. For sensor-level analyses, MEG
data were transformed to the head position coordinates of
the participant with the median head position within the
helmet to minimize transformation distance.
Sensor-level Analyses
These analyses were conducted on gradiometers and
magnetometers separately using the SensorSPM analysis
method implemented in SPM5 (www.fil.ion.ucl.ac.uk/
spm/). Magnetometer data were used as such, whereas
for each pair of gradiometer channels, a vector sum was
calculated that reconstructed the field gradient from its
two orthogonal components and its amplitude (computed
as a square root of the sum of squared amplitudes in the
two channels). For each participant and condition, a series
of F tests were performed on a three-dimensional topogra-
phy (2-D sensors by time image), which extended through
601 samples (1 msec each), allowing for the application of
random field theory as in fMRI analysis (Kiebel & Friston,
2004). The 3-D images were thresholded at a voxel level of
p < .005 and corrected for cluster size at p < .05. These
clusters could extend in space (distributed across the
topography) and in time. This made it possible to compare
conditions across every sensor over the entire time win-
dow while still correcting on a whole-brain basis for multi-
ple comparisons. This procedure eschews any preselection
of time windows of interest and provides a data-driven
selection process, which is not restricted to specific peaks
found through visual inspection of the data.
Source Estimation
MP-RAGE T1-weighted structural images with a 1 × 1 × 1 mm
voxel size were acquired on a 3-T Trio Siemens scanner for
each participant, which were used for reconstruction of the
cortical surface using Freesurfer (Athinoula A. Martinos
Center for Biomedical Imaging). The L2 minimum-norm
estimation (Hämäläinen & Ilmoniemi, 1994) technique
was applied for source reconstruction as implemented in
the MNE Suite. An individual MRI-based one-layer bound-
ary element model (BEM) was created for each participant
and was used to compute the forward solutions. An aver-
age cortical solution, containing 10,242 dipoles per hemi-
sphere, was created from the 16 participants, and data from
individual participants were morphed to this cortical sur-
face in 10-msec time steps. ROIs were defined from Free-
Surfer anatomical ROIs, with the exception of the large
temporal and fusiform ROIs, which were subdivided into
anterior, middle, and posterior regions. ROIs were defined
on the average cortical surface, and for each participant,
the mean value for all dipoles within each region was
extracted for statistical analysis. The source-level analyses,
using repeated-measures ANOVAs on the participant
means within a given ROI, were restricted to the time win-
dows where significant effects (after correction for multiple
comparisons) were found in the sensor analyses. The re-
sults are visualized on the inflated cortical surface of the
average participant.
RESULTS
Recognition Task Results
For the recognition task, mean accuracy was at 65% and
did not vary significantly between words, pseudowords,
and consonant strings, with accuracy at 66%, 62%, and
66%, respectively. Performance was assessed statistically
using signal detection theory to test the discriminability in-
0 = 0 would
dex (d
represent no difference between signal and noise. Dis-
0 = .98,
criminability was significantly greater than 0 (d
p < .0001), suggesting participants were reliably attending
to the items.
0) against 0 using a paired t test, where d
MEG Results
In the results, we combine sensor-based and source-space
analyses in each section. Sensor-level results are presented
separately for gradiometers and magnetometers, followed
by source-space analyses. The results are organized into
two analysis streams, relating basic stages in the visual
word recognition process to the processes that map be-
tween them. One stream focuses on the morphologically
simple word and pseudoword stems, together with
matched consonant strings, and the other on the morpho-
logically complex and pseudo-complex forms. These statis-
tically rigorous analyses, on sets of matched simple and
complex materials, provide a well-controlled backdrop
for evaluating how lexical, morphological, and semantic
variables relate to different stages of the visual word recog-
nition process. The Detecting Orthographic Structure and
Emergence of Neural Sensitivity to Morphological Structure
252
Journal of Cognitive Neuroscience
Volume 27, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
sections focus on the relationship between orthographic
analyses and the early stages of lexical access. The
Processing Lexical Identity and Lexical Effects for
Morphologically Complex Words sections address the
role of lexical constraints in the analysis of orthographic
inputs.
Detecting Orthographic Structure
The first set of analyses contrasted words and pseudo-
words with consonant strings to establish spatiotemporal
coordinates for effects associated with processing read-
able letter strings. To conduct these analyses, we used
100 pseudoword stems ( frum, bect) from the ( frumish)
and (bected) conditions, excluding the pseudoword stems
from the (blemish) and (biscuit) conditions, because
these could be interpreted as the initial portion of an ex-
isting word. One hundred word stems ( farm, corn) were
also selected, together with 100 consonant strings, all
matched in length.
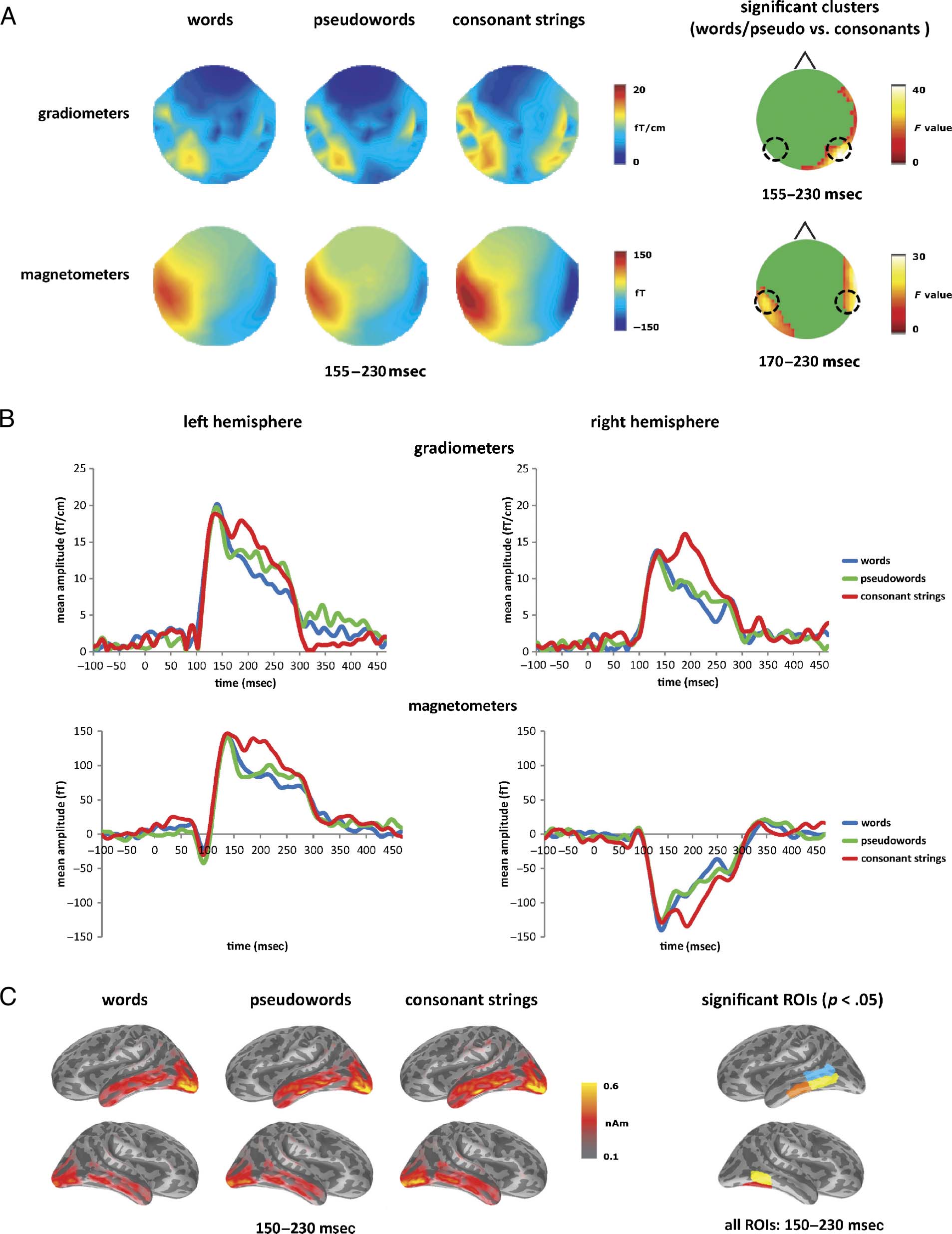
At the sensor level (see Figure 1A), the SensorSPM con-
trast of words and pseudowords against consonant strings
showed significant effects emerging between 155 and
230 msec in both gradiometers and magnetometers bilat-
erally. In the gradiometers, the cluster was significant from
155 to 230 msec within posterior right sensors with the
peak at 195 msec. In the magnetometers, significant bilat-
eral clusters appeared in left hemisphere (LH) sensors
from 170 to 230 msec, peaking at 190 msec, and in right
hemisphere (RH) sensors from 175 to 220 msec, peaking
at 200 msec. All of these clusters reflect a stronger response
to consonant strings than to words and pseudowords.
Figure 1B plots early orthographic effects for LH and
RH gradiometers and magnetometers at the peak of the
significant sensor-level cluster in each hemisphere. In both
hemispheres, there is an initial common response to all
three stimulus types, peaking at 140 msec, followed by a
second peak, at around 190 msec, which differentiates
consonant strings from words and pseudowords. This in-
dicates the presence of processes that are sensitive to
orthographic structure but not to the lexical properties of
the strings being analyzed.
At the source level (Figure 1C), focusing on the 155–
230 msec time window defined by the sensor-level results,
the strongest activity was in occipitotemporal regions bilat-
erally, with activation extending along inferior and middle
temporal gyri bilaterally. Differences between conditions
emerge at bilateral posterior and middle temporal sites,
primarily along the ventral surface in fusiform and infe-
rior temporal areas. Contrasting words and pseudowords
against consonant strings, significant differences were seen
in left posterior ITG (F(1, 15) = 8.24, p < .05), left middle
ITG (F(1, 15) = 11.84, p < .005), left posterior MTG (F(1,
15) = 8.98, p < .01), right posterior fusiform (F(1, 15) =
6.70, p < .05), and right posterior ITG (F(2, 30) = 6.85,
p < .05). All ROIs showed increased processing for conso-
nant strings over words and pseudowords.
The location and timing of these orthographically
sensitive processes is consistent with earlier research.
Previous fMRI studies have shown increased activation
for consonant strings in posterior occipital regions (e.g.,
Vinckier et al., 2007), indicating that visual word forms
are differentially processed on the basis of their ortho-
graphic structure. Processing between 150 and 200 msec
has been shown to be specific to letter strings but not yet
to word-like strings (Cornelissen et al., 2003), although
some studies have found effects associated with ortho-
graphic typicality as early as 100 msec (Hauk et al.,
2006). Here we find that the initial component peaking
at 140 msec did not differentiate between stimulus types
(although we did not explicitly test for typicality).
Emergence of Neural Sensitivity to
Morphological Structure
Here we examine the timing and distribution of analysis
processes sensitive to the presence of cues to morpho-
logical structure. If potential stems and grammatical affixes
are present in an orthographic input string, when do they
start to trigger differential neural responses? We were
guided here by the masked priming results. The four
conditions containing complex forms with a stem and an
affix (+S+A) all showed significant priming (see Table 3).
These were two derivational sets ( farmer, corner) and
two inflectional sets (blinked, ashed). We contrasted these
with two noncomplex conditions (the scandal (+S−A) and
biscuit (−S−A) sets), neither of which elicited priming.
The presence of the pseudo-derived corner forms and
the non-existing ashed forms make this a test for morpho-
logical effects that are blind to lexical-level variables. In
both cases, the significant masked priming effect is direct
behavioral evidence that stimuli of this type elicit morpho-
logically driven decomposition that is not blocked by
lexical criteria.
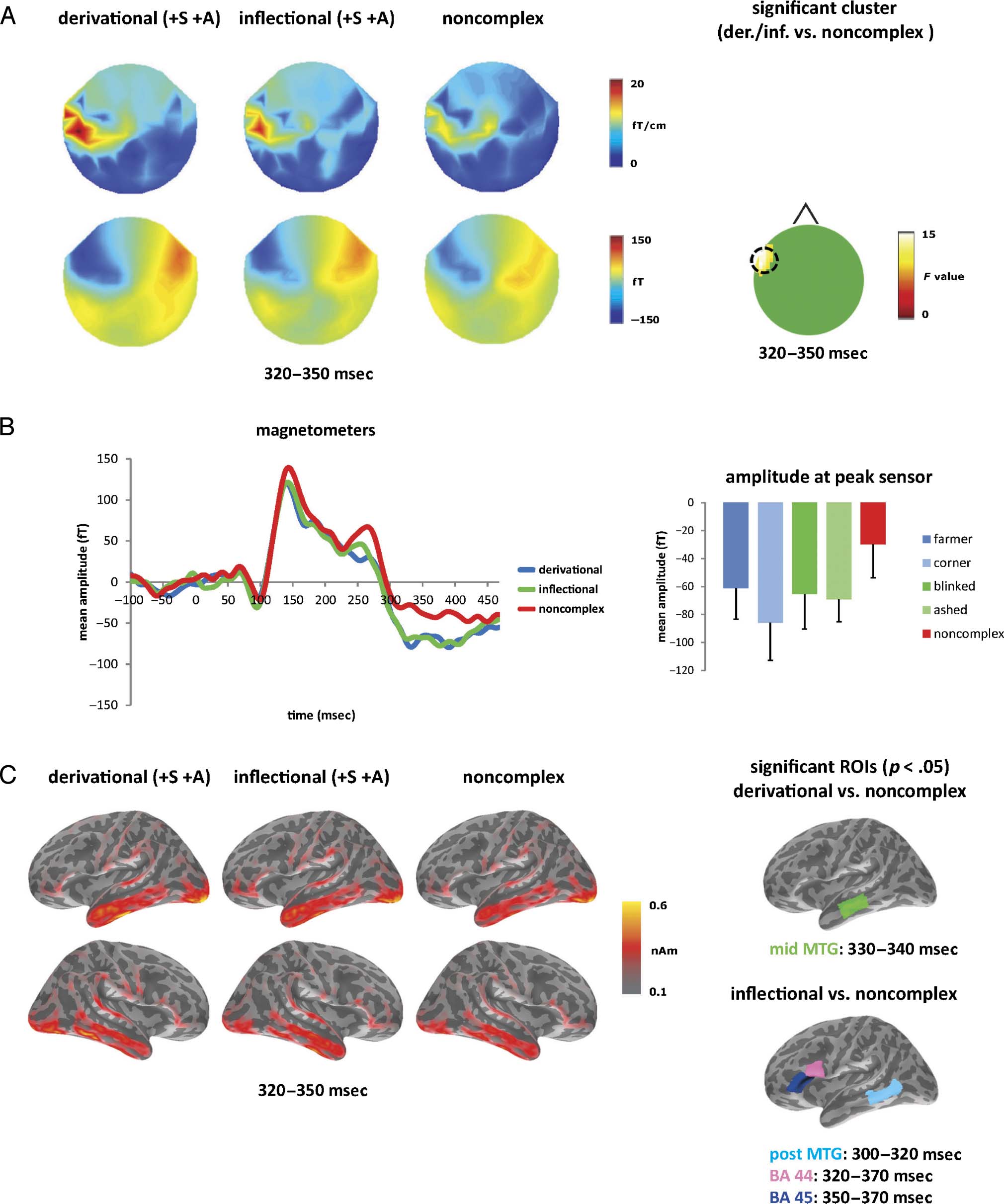
There was an increase in processing activity for the com-
bined derived and inflected forms compared with non-
complex forms in anterior left magnetometers, extending
from 325 to 350 msec with a peak at 335 msec (see Fig-
ure 2A). There was no evidence in these brain-wide (and
globally corrected) analyses for earlier or more posterior
effects of morphological structure. At the peak magnetom-
eter sensor from the SensorSPM analysis, there was no dif-
ference between the four +S+A conditions (F < 1) nor
between complex and pseudo-complex forms within com-
plexity type ( farmer vs. corner (t(15) = 1.49, p = .16);
blinked vs. ashed (t(15) < 1). Analyzing derived and in-
flected forms separately, the derived forms show the same
magnetometer cluster, from 320 to 365 msec in left ante-
rior sensors with a peak at 335 msec. The effects for the
inflected forms fall short of significance (but see source
level analyses below).
The topography of these sensor-level effects is more
anterior, left-lateralized, and later in time than the ortho-
graphic effects displayed in Figure 1. Evidence from the
Whiting, Shtyrov, and Marslen-Wilson
253
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 1. Orthographic structure effects: Word–pseudoword–consonant string contrasts. (A) Sensor level topographic maps for words, pseudowords,
and consonant strings averaged over the period 155–230 msec, corresponding to the significant clusters from the SensorSPM analyses (right-hand
column). Gradiometer results are in the upper row and magnetometer results in the lower. (B) Mean RMS gradiometer and magnetometer responses
to words (blue), pseudowords (green), and consonant strings (red) at the peak sensors of the significant clusters (locations indicated by black circles
in A). (C) Estimated source activation for words, pseudowords, and consonant strings, averaged over the period 150–230 msec, with the significant
effects in bilateral posterior ITG (yellow), left middle ITG (orange), left posterior MTG (blue), and right posterior fusiform (red) ROIs. Cortical images
are rotated to display the ventral surface.
254
Journal of Cognitive Neuroscience
Volume 27, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. Emergence of morphological structure effects. (A) Sensor-level topographic maps for derivational +S+A ( farmer, corner), inflectional
+S+A (blinked, ashed ), and noncomplex (scandal, biscuit) forms from 320 to 345 msec and the significant cluster from the magnetometer analysis.
Gradiometer and magnetometer results are in the upper and lower rows, respectively. (B) Mean response to derivational (blue), inflectional (green),
and noncomplex (red) forms at the peak of the significant magnetometer cluster (indicated by black circle in A). At right, the mean amplitude during
the significant cluster for +S+A and noncomplex forms. (C) Estimated source activation for derivational, inflectional, and noncomplex forms, averaged
over the period 300–360 msec, with the significant effects in left MTG (green) for derivational forms and in left posterior MTG (light blue), left pars
triangularis (dark blue), and left pars opercularis (pink) for inflectional forms.
Whiting, Shtyrov, and Marslen-Wilson
255
magnetometers (Figure 2B) showed a stronger response to
the derivational and inflectional forms at the peak left tem-
poral sensors, sustained over the period 300–450 msec,
with peak effects at around 320–330 msec.
Turning to the contribution of stems and affixes to these
morphologically driven processes, the results confirm that
both these elements need to be present, whether to elicit
masked priming or to generate different distributions of
neural activity. In a further analysis, the (−S, + A) blemish
set patterned with the noncomplex scandal and biscuit con-
ditions, consistent with the view that both a potential stem
and a potential affix are needed to trigger early segmentation.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
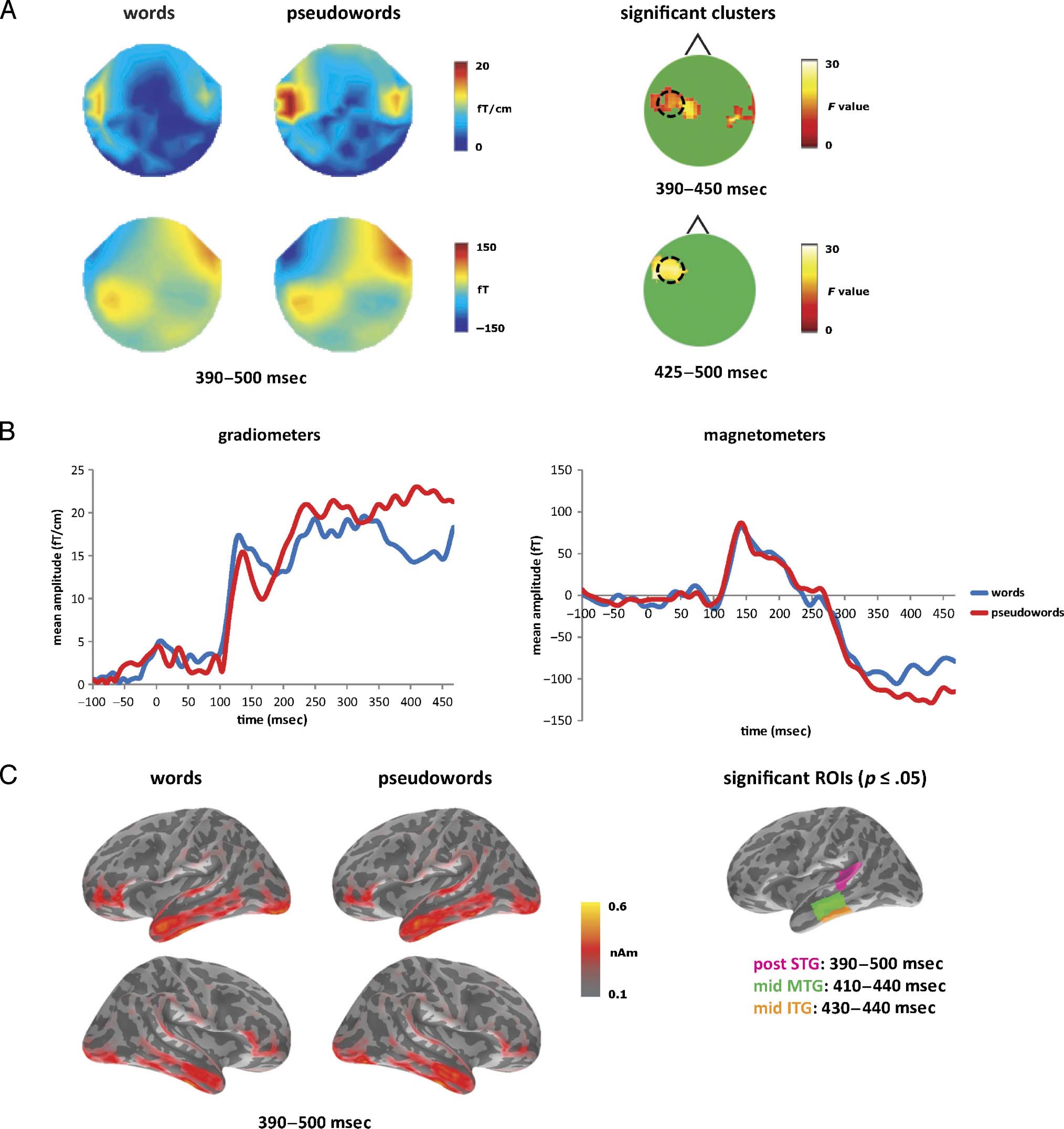
Figure 3. Lexicality effects: Word–pseudoword contrast. (A) Sensor-level topographic maps for words and pseudowords from 390 to 500 msec,
with significant clusters from the sensor analysis. Gradiometer and magnetometer results are in the upper and lower rows, respectively. (B) Mean
RMS gradiometer and magnetometer responses to words (blue) and pseudowords (red) at the peak of the significant cluster (indicated by black
circles in A). (C) Estimated source activation for words and pseudowords from 390 to 500 msec, with the significant effects in left posterior STG
(pink), left middle MTG (green), and left middle ITG (orange).
256
Journal of Cognitive Neuroscience
Volume 27, Number 2
At the source level (Figure 2C), focusing on the time
window during which significant sensor-level effects were
found, activation has shifted more anteriorly and now
includes inferior frontal areas, most strongly on the left.
Specific contrasts between conditions within ROIs showed
that the derivational/noncomplex contrast was significant
in a 330–340 msec time window in left middle MTG (F(1,
15) = 4.69, p < .05), at the peak of the effect found at
the sensor level. For the inflectional/noncomplex contrast,
we see a more complex pattern, with effects in left poste-
rior MTG from 300 to 320 msec (F(1, 15) = 4.47, p = .05)
together with slightly later effects in the left inferior
frontal gyrus (LIFG), in left BA 44 from 320 to 370 msec
(F(1, 15) = 5.21, p < .05) and in left BA 45 from 350 to
370 msec (F(1, 15) = 4.81, p < .05).
The left frontotemporal patterning of these inflectional
effects, which is identical for both the (+S+A) inflectional
conditions (blinked, ashed), is consistent with extensive
research using spoken words (e.g., Marslen-Wilson &
Tyler, 2007) locating morphosyntactic effects in exactly
these left peri-sylvian locations. The pseudoword bected,
in contrast, which contains a potential inflectional affix
but no stem, did not elicit significant effects in these ROIs
compared with the noncomplex forms. This suggests that
the requirement for both a stem and an affix to be present
extends to inflectional as well as derivational morphology
in visual word recognition.
The timing of these morphological effects is generally
in line with several earlier EEG and MEG studies showing
sensitivity to morphological complexity between 300 and
500 msec in primed and unprimed studies (e.g., Leinonen
et al., 2009; Vartiainen et al., 2009; Lavric et al., 2007;
Lehtonen et al., 2007; Dominguez et al., 2004). The hypoth-
esized M350 component, which peaks at approximately
350 msec in left temporal cortex, has been linked to
initial activation of lexical representations (Pylkkänen,
Stringfellow, & Marantz, 2002), as well as multimorphemic
forms such as compounds (Fiorentino & Poeppel, 2007).
Earlier morphological effects have also been reported—
for example, by Lavric et al. (2012)—with effects potentially
emerging as early as 200 msec postonset. We discuss these
below, in the context of potential task influences on the
timing of such reports. In terms of spatial location, left
MTG has previously been implicated in morphological
decomposition in several studies using fMRI (e.g., Mar-
slen-Wilson & Tyler, 2007; Lehtonen, Vorobyev, Hugdahl,
Tuokkola, & Laine, 2006; Tyler et al., 2005) as well as being
linked to lexical retrieval and semantic processing (cf.
Zhuang, Tyler, Randall, Stamatakis, & Marslen-Wilson,
2012; Turken & Dronkers, 2011; Hickok & Poeppel, 2007).
Processing Lexical Identity
A complementary set of analyses focused on the word and
pseudoword stems to test for effects linked to successful
lexical access. The same two sets of 100 morphologically
simple words and pseudowords were used as before. At
the sensor level (Figure 3A), the gradiometers revealed
one cluster at 390–450 msec in left temporal sensors and
a smaller cluster from 410 to 440 msec in right temporal
sensors, peaking at 430 msec in both hemispheres. In the
magnetometers, one cluster emerged at 425–500 msec
within anterior left sensors with a peak at 470 msec. All
clusters showed increased processing of pseudowords
over words. Figure 3B plots the gradiometer and magne-
tometer response amplitudes for words and pseudowords
at the peak of the significant LH cluster, with the two con-
ditions starting to separate at 350 msec and peaking at
430 msec.
Source-level analyses (Figure 3C) focused on the 390–
500 msec time window where significant lexicality ef-
fects were found in the sensor-level analyses. The overall
distribution of activation has shifted anteriorly and fron-
tally, especially on the left, where there is strong activity
in temporal and inferior frontal regions for both words
and pseudowords. Differences between these conditions
emerge more posteriorly, with stronger responses to
pseudowords in left posterior STG from 390 to 500 msec
(F(1, 15) = 5.12, p < .05), left middle MTG from 410 to
440 msec (F(1, 15) = 5.48, p < .05), and left middle ITG
from 430 to 440 msec (F(1, 15) = 4.50, p = .05). These
lexicality effects overlap spatially with the morphological
effects (Figure 2) but emerge around 100 msec later.
These spatiotemporal and functional patterns are consis-
tent with the standard N400-like effects seen in MEG in
terms of timing as well as location (Pylkkänen & Marantz,
2003; Halgren et al., 2002) and with evidence from fMRI
showing the involvement of left posterior temporal regions
in semantic processing (Hickok & Poeppel, 2007). Lexicality
effects on the N400 have been interpreted as reflecting
access to lexical representations (e.g., Lau et al., 2008; Kutas
& Federmeier, 2000), whereas Dominguez et al. (2004)
report prolonged N400 effects at the level of meaning selec-
tion because of incorrect morphological decomposition.
Lexical Effects for Morphologically Complex Words
The four +S+A ( farmer, corner, blinked, ashed) condi-
tions were used to examine potential interactions of lexical-
level effects with the processing of morphologically complex
letter strings. For the derivational pair, both farmer and
corner are existing words, but corner has a potential inter-
pretation as a pseudo-stem plus a pseudo-affix. For the
inflectional pair, ashed is not an existing word, although it
is potentially interpretable as a real stem plus a real affix.
In each case, if an early segmentation process identifies
these forms as potentially morphologically complex real
words, a later process sensitive to lexical-level informa-
tion will need to rescue the perceptual system from these
potential garden paths.
A significant cluster emerged from 400 to 500 msec
within left anterior temporal magnetometers, showing
increased activation of both corner and ashed forms
relative to their corresponding semantically transparent
Whiting, Shtyrov, and Marslen-Wilson
257
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
258
Journal of Cognitive Neuroscience
Volume 27, Number 2
conditions, peaking at 450 msec (Figure 4A). The ampli-
tude plots indicate comparable effects for the two contrasts.
Consistent with this, the corresponding magnetometer
and gradiometer responses (Figure 4B) show similar trajec-
tories over time, although differences between conditions
emerge earlier for the inflectional pairs. Notably, the timing
and distribution of these effects are very similar to those
seen for the lexicality effects reported in Processing Lexical
Identity section for morphologically simple words and
pseudowords.
At the source level (Figure 4C), we see substantial bilat-
eral activation in temporal and inferior frontal regions for
both complex and noncomplex conditions, but significant
differences between them only emerge in more posterior
and inferior temporal sites. In all cases these reflect in-
creased processing for pseudo-complex over complex
forms and are likely be the result of top–down feedback
processes. The combined contrast of complex versus
pseudo-complex is significant from 400 to 470 msec in
left posterior fusiform (F(1, 15) = 7.92, p < .05) and ap-
proaches significance from 400 to 410 msec in left middle
ITG (F(1, 15) = 4.20, p = .058) and left middle fusiform
(F(1, 15) = 3.85, p = .068). Broken down by morpholog-
ical type, the 400–470 msec effect in left posterior fusiform
gyrus is significant only for the derivational farmer/corner
contrast (F(1, 15) = 8.66, p < .01), whereas the brief effect
in left middle ITG from 400 to 410 msec is found only for
the inflectional blinked/ashed contrast (F(1, 15) = 4.94,
p < .05).
DISCUSSION
This research shows that we can unify the functional char-
acteristics of real-time neural analysis with the functional
properties of visual word recognition as revealed in the
masked priming data. We can use this linkage across
methodological domains to determine the functional
architecture of the neurobiological system that generates
these properties. In doing so, we benefit in particular from
the spatiotemporally specific constraints provided by MEG.
Unlike other imaging methodologies, MEG data mapped
into neuroanatomically constrained source space allow us
to specify not only when processes of different types begin
and end, but also (within the limits of MEG source recon-
struction) where these processes take place.
The proposed architecture based on these results con-
ceptualizes visual word recognition as a two-phase process,
where primarily feedforward orthographically driven
analyses segment the visual input into potentially mean-
ingful linguistic substrings (words and morphemes) and
where these substrings initiate lexical access in middle
and frontotemporal locations from about 300 msec after
stimulus onset. The initial stages of this access process
are dominated by morpho-orthographic factors, with lexi-
cal constraints becoming detectable around 100 msec
later, as reflected in effects of lexical identity and in in-
creased processing for pseudo-complex strings like corner.
We review below the evidence for these claims and their
implications.
Morphemically Driven Lexical Access
The joint behavioral and neuroimaging false segmentation
effects for ashed- and corner-type stimuli are compelling
evidence that the output of orthographic analysis is not
in terms of lexical words per se, but in terms of mor-
pheme-like linguistically relevant substrings. The critical
MEG contrast is between materials that show decomposi-
tional effects in masked priming—the derivationally and
inflectionally complex farmer, corner, blinked, and ashed
conditions—and materials (scandal, biscuit) that do not
(Figure 2). Consistent with the masked priming results,
this contrast reveals a time period—between 300 and
370 msec from stimulus onset—where both complex sets
diverge from the noncomplex sets, but where there are
no significant differences within each set as a function of
their lexical properties. At the source level, the derivational
set differs from the noncomplex set in left middle MTG
at 330–340 msec, but the farmer/corner subsets do not
differ. Similarly, the inflectional set differs from the non-
complex set between 300 and 370 msec in left posterior
MTG and LIFG, but the blinked/ashed subsets do not dif-
fer. This pattern of results closely parallels the morpho-
orthographic process hypothesized on behavioral grounds,
where the input is analyzed in terms of its morphological
properties, but is blind to the lexical properties of the
words involved. This is particularly clear for the inflected
forms, where both blinked and ashed deviate from the
noncomplex forms at around 300 msec and where the
presence of the inflectional morpheme activates classic
LIFG regions (BA 44 and BA 45) irrespective of the lexical
status of the whole form.
The finding that these early processes do not discrimi-
nate between genuinely complex and pseudo-complex
strings demonstrates that the processes generating can-
didates for lexical access and recognition are blind to
the lexical properties of the strings they are generating.
The results for ashed and corner also demonstrate that the
manner in which these output processes interface with
lexical representations is morphologically compositional.
Figure 4. Lexicality effects: Morphologically complex words. (A) Topographic maps for complex ( farmer, blinked ) and pseudo-complex (corner,
ashed ) forms from 400 to 470 msec and the significant cluster from 400 to 470 msec from the sensor analysis. At far right, the mean amplitude in
the significant left anterior cluster for farmer, corner, blinked, and ashed. (B) Mean response to derivational ( farmer, corner), left, and inflectional
forms (blinked, ashed ), right, at the peak of the significant magnetometer cluster (indicated by black circle in A). (C) Estimated source activation
for complex and pseudo-complex forms and separate derivational ( farmer, corner) and inflectional (blinked, ashed ) forms from 400 to 470 msec,
with the significant effects in left middle ITG (orange) and left posterior fusiform (red).
Whiting, Shtyrov, and Marslen-Wilson
259
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
For the inflectional morphology, strings like ashed cannot
be accessed as stored forms, because they are not existing
words. Instead, they must be compositionally constructed,
combining the potential stem (ash) and affix (−ed). More
striking still, a monomorphemic form like corner—
no different at the lexical level from a simple form like
biscuit—seems to be temporarily reconstructed as the
nonexistent complex form {corn} + {-er}. This patterns
in the relevant neural time window—as well as in masked
priming—with genuinely complex forms like farmer and
blinked and not with noncomplex forms like scandal
and biscuit. These effects require the output of ortho-
graphical analysis to be morphemically decomposed.
More evidence for morphemic constraints on early
orthographically driven string segmentation and identifi-
cation comes from the sensitivity of this process to the
morphemic status of both elements of a potential com-
plex form. The masked priming data—here and in earlier
studies—together with the MEG analyses involving the
partially complex scandal, blemish, frumish, and bected
sets, suggest that pseudo-complex forms are not treated
as complex forms in the 300–370 msec time window
unless they contain both a potential stem and a potential
affix—as in the corner and ashed conditions. This points
to a segmentation process that is not only sensitive to
the presence of linguistically relevant subunits but also
to the contexts in which they co-occur.
The view that orthographic analysis results in a mor-
phemic output is consistent with the proposals of Dehaene
and colleagues (Vinckier et al., 2007; Dehaene, Cohen,
Sigman, & Vinckier, 2005), where the endpoint of ortho-
graphic analysis in posterior temporal cortex is seen as
the identification of “small words and recurring substrings
(e.g., morphemes).” It is also consistent with earlier MEG
evidence that orthographic processing in inferior and poste-
rior temporal regions, over early 150–250 time windows, is
sensitive to the presence of potential stems or affixes (e.g.,
Lehtonen et al., 2011). More generally, these can be seen
as aspects of a ventral stream object recognition process
tuned to orthographic analysis over decades of intensive
experience with written text.
Visual Word Recognition as a Two-phase Process
The evidence for the salience of morphemic factors in the
visual word recognition process, together with the demon-
stration of a short period during which structural morpho-
logical factors seem to dominate, raises the question of
whether this indicates a separate, specifically morphologi-
cal processing stage, intervening between orthographic
analysis and access to lexical representations. The exis-
tence of such a stage is both a frequent postulate in cog-
nitive theories of visual word recognition and a major
source of disagreement between competing theories.
The evidence here is that there is no such separable pro-
cessing stage and that what we see instead are two inter-
secting phases of neurocognitive activity. The first, as
described above, is located in posterior and inferior tem-
poral and occipitotemporal regions and is concerned with
the analysis of the visual input into higher-order linguisti-
cally relevant orthographic units. These processes are in
themselves neither lexical nor semantic in nature and form
a spatiotemporally distinct phase in the recognition pro-
cess. This can be viewed as a modality-specific input sys-
tem that projects onto a more distributed morpholexical
system, more anterior and frontotemporal, that is sensitive
to the morphological structure of complex forms and to
lexically represented variables more generally—and which
is likely to be largely in common with the target systems
accessed from auditory inputs.
The separation into two phases is reflected in the spatio-
temporal distribution of processes sensitive to orthograph-
ic variables but not to morphological structure or lexical
identity. For orthographic structure, there is increased
activation for consonant strings (relative to words and
pseudowords) in the time period 150–230 msec, seen bi-
laterally in posterior brain regions (Figure 1). Analyses
sensitive to morphological structure emerge at around
300 msec, peaking 100–150 msec later than the ortho-
graphic effects, whereas the spatial center of gravity shifts
anteriorly to more dorsal left frontotemporal sites (Fig-
ure 2). None of the inferior temporal and fusiform ROIs
that were significant in the orthographic analyses are active
in the contrasts sensitive to morphological structure.
Although there continues to be RH activation for both
complex and pseudo-complex conditions, the only effects
that differentiate between conditions are seen in LH mid-
dle temporal and inferior frontal sites. A similar though
spatiotemporally more clear cut separation from early or-
thographic processing is seen for the lexically sensitive
effects, with increased processing for pseudoword stems
from 390 to 500 msec, chiefly in left middle temporal re-
gions (Figure 3). In complementary analyses contrasting
complex pseudowords ( frumish, bected ) with complex
real words ( farmer, blinked ), we found comparable
effects, with increased processing for pseudowords emerg-
ing in left temporal sensors from 425 to 465 msec.
Taken together, these data show that there is a clear
separation in neural space and time between orthographi-
cally centered analyses and those sensitive to morphologi-
cal structure and to lexical variables. There is little evidence
for a similar separation between the latter types of process,
on the basis of the contrasts plotted across Figures 2–4.
Although different phases of analysis peak at different
points in time, with the earliest morphological structure
effects emerging around 100 msec earlier than the effects
of lexicality, there is no evidence that these activities are
spatially distinct—especially where core middle temporal
locations are concerned. Given these timing and location
constraints—which are consistent, as noted earlier, with
earlier EEG and MEG studies—the most plausible account
is that, although the engagement and interpretation of
lexical constraints have a sequential time course, these pro-
cesses involve the same set of frontotemporal brain regions
260
Journal of Cognitive Neuroscience
Volume 27, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
as those implicated in the analysis of morphological
structure.
On this account, morphological effects in visual word
recognition will emerge as an interaction between the
outputs of orthographic analysis and the properties of
morpholexical representation and analysis. These in turn
will depend on the properties of simple and complex
words in the language and how they are lexically repre-
sented. Research in the auditory domain suggests that
inflectionally complex words in English are decomposi-
tionally represented and analyzed in the neural language
system (e.g., Marslen-Wilson & Tyler, 2007), whereas de-
rivationally complex words are accessed as whole forms
(Bozic et al., 2013), although with some preservation
of internal morphological structure (cf. Marslen-Wilson,
2007). There are signs of this differentiation here, with
the analysis of inflected and pseudo-inflected forms
(blinked, ashed) closely paralleling the decompositional
neural patterns seen in the auditory domain (Bozic, Tyler,
Ives, Randall, & Marslen-Wilson, 2010; Marslen-Wilson &
Tyler, 2007). Derivationally complex forms, in contrast,
primarily activate middle temporal sites, although further
research is needed here.
Feed-forward Processing and Recurrence
The third defining feature of the proposed functional
architecture concerns the processing relationship be-
tween the orthographic analysis of the input and the
broader lexical and contextual context in which this analy-
sis occurs: Does this context modulate early orthographic
analyses, as interactionist accounts would require, or
do these analyses operate in a primarily feedforward (or
bottom–up) manner? There are two aspects to this—
whether lexical constraints are directly coded into the
orthographic analysis process and whether this process is
dynamically modulated by top–down predictive or recur-
rent processes.
On an encoding account, the orthographic mapping
process is tuned to the specific lexical context of the lan-
guage, so that it would not generate (or would disprefer)
outputs that were not lexically valid. The results here
argue against this, with the first-pass analysis of letter strings
into potential stems and affixes being conducted with-
out reference to the lexical identities of these strings (cf.
Marslen-Wilson et al., 2008). Otherwise the misanalysis of
corner would be blocked, along with the rejection of non-
words like ashed. The results are also inconsistent with a
weaker encoding account, where lexical variables modulate
but do not determine early segmentational hypotheses.
Significant lexical effects on the analysis of potential com-
plex forms—indexed by increased processing for corner
over farmer and for ashed over blinked—are not seen until
the 400–470 msec time period, substantially later than the
initial emergence of morphological structure effects. On an
encoding account, these effects should be seen at earlier
time points as well.
This leaves open the possibility that early analyses can
be modulated by externally generated constraints—for
example, by predictive constraints generated in a sentence
context or by recurrent constraints generated top–down
as a letter string is being processed. In the current study,
we only see candidate effects of this type in late time
windows (400 msec from word onset), with increased
processing for pseudo-complex forms like corner at pos-
terior temporal sites (Figure 4). If this is a top–down feed-
back effect, then it occurs too late to be evidence for early
morphosemantic interactions. Note that in the context of
an fMRI study, where the BOLD response sums over a
multisecond time window of neural activity, this critical
temporal separation is lost, making it possible to mis-
interpret such an effect as evidence for early interactions
between form and meaning. Similar caveats apply to con-
ventional RT tasks, where the temporal ordering of the mul-
tiple processes contributing to overall RT is also opaque.
However, although the current experiment allows us to
evaluate the role of system-internal constraint, it does not
allow us to evaluate the effects of contextual variables
more generally. The letter strings were presented in isola-
tion, and we took care to minimize task-based effects. The
natural habitat of the visual word recognition system is of
course reading in context, with potential constraints gen-
erated at syntactic, semantic, and pragmatic levels as
words are being read. Without properly time-resolved pro-
cessing measures, however, it is not possible to determine
how these contexts operate—whether they operate
primarily in the second phase domain of morpholexical
interpretation, or whether they directly modulate the
operations of the orthographic input system.
Finally, we note that some findings from EEG and MEG
suggest that the initial sweep through the visual word rec-
ognition system to the level of lexical access occurs within
200 msec of word onset (e.g., Shtyrov et al., 2013; Lavric
et al., 2012; Hauk et al., 2006; Pammer et al., 2004). These
timings contrast with this study, where morphological
effects emerge at 300 msec and lexicality effects appear
later still. This divergence could be attributed to a number
of sources.
One of these, as we proposed earlier, may be differences
in task demands. Task-driven top–down effects can modu-
late sequential feed-forward processes in the visual system
at early stages of sensory analysis (e.g., Twomey, Kawabata
Duncan, Price, & Devlin, 2011), and similar effects may
also modulate early processes relevant to the performance
of tasks such as lexical decision. Direct evidence for such
effects in visual word recognition comes from a further
study by Whiting (2011), parallel to the research reported
here, which ran the same set of stimuli but under differ-
ent task conditions. This study replaced the end-of-block
recognition test with an occasional lexical decision task,
occurring on 10% of trials. The results show both common-
alities and divergences relative to the current study. The
timing and pattern of early orthographic effects (compar-
ing words and pseudowords with consonant strings) were
Whiting, Shtyrov, and Marslen-Wilson
261
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
essentially unchanged. Morphological decompositional
effects are detected earlier, but with a similar spatio-
temporal distribution—for example, activity is seen in BA 44
for inflectional morphemes from 260 msec, rather than the
320 msec onset seen in the current study. Lexical effects
(comparing words and pseudowords) emerged around
100 msec earlier, starting at 310 msec and showing a similar
spatial distribution to this study. These selective effects on
the timing of different processes leave their relative order-
ing intact (and consistent with a morpho-orthographic
account) but suggest that task demands can indeed shift
the timing with which neural processes can be detected.
An additional source of divergences between studies
may be differences in statistical methods. In the majority
of published EEG and MEG studies of visual word recog-
nition, the dominant analysis strategy is to identify poten-
tial temporal or spatiotemporal ROIs on the basis of visual
inspection of the global energy profile and to focus sub-
sequent analyses around the visible peaks in this profile.
In the current study, we avoided any preselection of
areas of interest in favor of a brain-wide analysis process
(SensorSPM), conducted in sensor space, where the sig-
nificance of any contrast is corrected on a brain-wide basis
for multiple comparisons. We then used the outcome of
these analyses to select the time windows of interest with-
in which we conducted the source space analyses. This
approach, which is a more conservative—because globally
corrected—procedure for selecting time windows of in-
terest, will be less likely to pick up effects that are weak
and transient, and this may disfavor some very early ef-
fects. This is a possibility, however, that will need careful
evaluation in further research.
We suggest, in conclusion, that our findings are a robust
reflection of the basic underlying structure of the process-
ing system supporting visual word recognition, as revealed
in the context of reading words in the absence of a lexical
decision task. Top–down effects may well be at work in more
predictive natural perceptual contexts—such as the reading
of continuous text—to maximize the speed and efficiency
of the reading process. Nonetheless, such effects would
serve to modulate the performance of the basic feedforward
process we have described, not to replace it.
APPENDIX 1. MEG Study “Complex” Experimental Stimuli2
Derived
Transparent
Pseudo-
derived
Inflected
Transparent
Pseudo-
inflected
Pseudo-
stem
Simple
Pseudoword
Derived
Pseudoword
Inflected
aimed
altered
angered
arked
ashed
acorned
accordion
biscuit
ballot
bandit
billow
bishop
bleach
annoyed
barned
blossom
clibly
blamed
beasted
brothel
bourbon
bremful
brifter
chothly
arbed
bected
blumped
cloister
blended
birded
brown
cactus
boothed
burrow
cinnamon
blinded
blinked
bluffed
blurred
blushed
chanted
cheated
chewed
crashed
dared
drifted
earned
exited
glowed
command
cranium
chessed
chapel
clamp
clayed
colted
creeded
cordial
culted
cramp
doughed
curtain
dusked
dialect
earled
disco
faithed
dragon
filthed
electron
clench
collide
cream
cringe
dream
fiasco
flinch
fossil
fringe
giraffe
hinge
drowned
germed
fleet
glanded
fortune
griefed
harpoon
hollow
halled
hatchet
horizon
clongly
coiger
cralish
douper
droder
drooty
dunchly
farfer
flarly
flenly
foaper
fonkish
fowenly
fraiter
frinkful
frumish
fulker
bruffed
calked
caphed
crusked
dolded
donked
drelled
drissed
dunted
durped
fasked
fouted
fumped
furked
geeted
goined
grawed
hained
accuser
baker
baldness
bluntly
boldly
bomber
briskly
calmly
charmer
cheerful
chilly
climber
coyly
creepy
curable
curly
dancer
densely
dimly
diver
brandish
dizzy
broker
feather
bully
bunker
burnish
filter
finger
flutter
butcher
folly
archer
badger
balmy
beaker
belly
blazer
caper
coaster
corner
county
cracker
cranky
dumpy
fairy
Pseudo-
affix
bladder
blemish
blister
clatter
clever
garnish
gender
glitter
jolly
lavish
lorry
lumber
nourish
pamper
drummer
flipper
262
Journal of Cognitive Neuroscience
Volume 27, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
APPENDIX 1. (continued )
Derived
Transparent
Pseudo-
derived
Pseudo-
affix
Inflected
Transparent
Pseudo-
inflected
Pseudo-
stem
Simple
Pseudoword
Derived
Pseudoword
Inflected
farmer
feverish
fiercely
foolish
fussy
golfer
graceful
grimly
guilty
harmless
healer
hunter
joyful
lonely
loosely
lucky
mildly
mixer
painter
politely
quietly
quitter
sadness
singer
smoker
stiffly
thriller
frisky
plaster
groaned
heighted
limbo
jaguar
fruitless
plunder
guessed
heroed
manicure
lemon
grateful
probable
gulped
gullible
prosper
halter
quiver
hurled
jeered
hulked
jeeped
pasta
pillow
lamped
plaintiff
lounge
maggot
mellow
hammer
rally
laughed
lioned
plump
mundane
reckless
rowdy
saunter
sever
shelter
leaked
loaned
melted
mayored
pulpit
mealed
raven
nephew
orphan
menued
scandal
paradise
mended
oathed
scarf
phantom
obeyed
oceaned
shovel
plunge
skirmish
plucked
panged
shrewd
peckish
slander
phoney
slender
ponder
slither
poured
prayed
pushed
pathed
sight
pewed
pinted
slumber
spinach
salmon
prison
queen
rabbit
portable
smother
roared
poemed
spurt
sermon
squander
rolled
sheafed
stagnant
shadow
irony
larder
ledger
listless
master
mister
rafter
saucer
spanner
syllable
shouted
snailed
stark
stigma
stretcher
tally
shunned
stooled
sternum
stomach
study
tarnish
sweater
tether
sniffed
spilled
thunder
sprayed
tacky
treaty
vanish
varnish
weather
whisker
whisper
wicker
wistful
stirred
thawed
warned
yawned
yielded
sworded
stunt
symptom
traited
verbed
tartan
trample
trauma
tribune
wanded
trapeze
trivia
wasped
twinge
wombed
vowel
wooled
wrench
turban
turmoil
wreath
yeasted
yellow
zebra
funcher
gackish
gesher
goaper
jusher
marfy
nendy
pawker
prinker
prokish
purtly
sengly
shalker
shander
sligness
sooder
speepy
sterpful
storter
sustly
swener
tecter
telsely
theetless
thocker
tiefer
tridable
tullable
wrelful
heabed
heened
jucted
jurfed
learted
meefed
mirphed
narted
nooped
nouded
paimed
peffed
phized
plerked
porped
remped
sested
slasked
slimped
sorked
soubed
spoated
stawed
telked
terted
vacked
vissed
yomped
yooted
painless
sneaker
sully
stammer
ruined
rushed
sheeped
stallion
shuffle
skulled
stampede
staunch
traceable
witness
yonder
Acknowledgments
This research was supported by a grant to W. M. W. from the
European Research Council (NEUROLEX 230570) and by MRC Cog-
nition and Brain Sciences Unit funding (U.1055.04.002.00001.01).
C. W. was supported by funding from the Cambridge Trusts
and a Howard Research Studentship from Sidney Sussex College,
Cambridge. Y. S. was supported by MRC core program funding
(MC-A060-5PQ90). We thank Daniel Wakeman for his assistance
with MNE and FreeSurfer; Kathy Rastle, Lorraine Tyler, Tim
Shallice, and Ram Frost for their comments on the manuscript;
and the MEG operators at the MRC CBU for their help.
Reprint requests should be sent to Caroline Whiting or William
Marslen-Wilson, Department of Psychology, University of Cam-
bridge, Downing Street, Cambridge CB2 3EB, UK, or via e-mail:
cmw59@cam.ac.uk, wdm10@cam.ac.uk.
Notes
1. We follow here, and throughout, the standard linguistic defi-
nition of the morpheme as the minimal meaning-bearing linguis-
tic unit (e.g., Matthews, 1991), distinguishing between “content”
Whiting, Shtyrov, and Marslen-Wilson
263
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
morphemes like the stems {corn} and {hunt} and grammatical
morphemes like the derivational affix {-er}.
2. Participants also saw a further 450 “stem” stimuli constructed
from the complex stimuli and 160 consonant strings (for details,
see text).
REFERENCES
Assadollahi, R., & Pulvermüller, F. (2003). Early influences of
word length and frequency: A group study using MEG.
NeuroReport, 14, 1183.
Baayen, R. H., Piepenbrock, R., & Gulikers, L. (1995). The
CELEX lexical database (CD-ROM). Philadelphia: Linguistic
Data Consortium, University of Pennsylvania.
Bentin, S., Mouchetant-Rostaing, Y., Giard, M. H., Echallier, J. F.,
& Pernier, J. (1999). ERP manifestations of processing printed
words at different psycholinguistic levels: Time course and
scalp distribution. Journal of Cognitive Neuroscience, 11,
235–260.
Bozic, M., Tyler, L. K., Ives, D. T., Randall, B., & Marslen-Wilson,
W. D. (2010). Bihemispheric foundations for human speech
comprehension. Proceedings of the National Academy of
Sciences, U.S.A., 107, 17439–17444.
Bozic, M., Tyler, L. K., Su, L., Wingfield, C., & Marslen-Wilson,
W. D. (2013). Neurobiological systems for lexical
representation and analysis in English. Journal of Cognitive
Neuroscience, 25, 1678–1691.
Cohen, L., Dehaene, S., Naccache, L., Lehéricy, S., Dehaene-
Lambertz, G., Hénaff, M. A., et al. (2000). The visual word
form area—Spatial and temporal characterization of an initial
stage of reading in normal subjects and split-brain patients.
Brain, 123, 291–307.
Cornelissen, P., Tarkiainen, A., Helenius, P., & Salmelin, R.
(2003). Cortical effects of shifting letter position in letter
strings of varying length. Journal of Cognitive Neuroscience,
15, 731–746.
Dehaene, S., Cohen, L., Sigman, M., & Vinckier, F. (2005).
The neural code for written words: A proposal. Trends in
Cognitive Sciences, 9, 335–341.
Diependaele, K., Sandra, D., & Grainger, J. (2009). Semantic
transparency and masked morphological priming: The case
of prefixed words. Memory & Cognition, 37, 895–908.
Dominguez, A., de Vega, M., & Barber, H. (2004). Event-related
brain potentials elicited by morphological, homographic,
orthographic, and semantic priming. Journal of Cognitive
Neuroscience, 16, 598–608.
Feldman, L. B., OʼConnor, P. A., & del Prado Martín, F. M.
(2009). Early morphological processing is morphosemantic
and not simply morpho-orthographic: A violation of
form-then-meaning accounts of word recognition.
Psychonomic Bulletin & Review, 16, 684–691.
Hämäläinen, M. S., & Ilmoniemi, R. J. (1994). Interpreting
magnetic fields of the brain: Minimum norm estimates.
Medical and Biological Engineering and Computing, 32,
35–42.
Hauk, O., Coutout, C., Holden, A., & Chen, Y. (2012).
The time-course of single-word reading: Evidence from
fast behavioral and brain responses. Neuroimage, 60,
1462–1477.
Hauk, O., Davis, M. H., Ford, M., Pulvermüller, F., &
Marslen-Wilson, W. D. (2006). The time course of visual
word recognition as revealed by linear regression analysis
of ERP data. Neuroimage, 30, 1383–1400.
Hickok, G., & Poeppel, D. (2007). The cortical organization
of speech processing. Nature Reviews Neuroscience, 8,
393–402.
Kiebel, S. J., & Friston, K. J. (2004). Statistical parametric
mapping for event-related potentials (II): A hierarchical
temporal model. Neuroimage, 22, 503–520.
Kutas, M., & Federmeier, K. D. (2000). Electrophysiology
reveals semantic memory use in language comprehension.
Trends in Cognitive Sciences, 4, 463–470.
Lau, E. F., Phillips, C., & Poeppel, D. (2008). A cortical network
for semantics: [De]constructing the N400. Nature Reviews
Neuroscience, 9, 920–933.
Lavric, A., Clapp, A., & Rastle, K. (2007). ERP evidence of
morphological analysis from orthography: A masked priming
study. Journal of Cognitive Neuroscience, 19, 1–12.
Lavric, A., Elchlepp, H., & Rastle, K. (2012). Tracking
hierarchical processing in morphological decomposition
with brain potentials. Journal of Experimental Psychology:
Human Perception and Performance, 38, 811–816.
Lehtonen, M., Cunillera, T., Rodríguez-Fornells, A., Hultén,
A., Tuomainen, J., & Laine, M. (2007). Recognition of
morphologically complex words in Finnish: Evidence from
event-related potentials. Brain Research, 1148, 123–137.
Lehtonen, M., Monahan, P. J., & Poeppel, D. (2011). Evidence
for early morphological decomposition: Combining masked
priming with magnetoencephalography. Journal of Cognitive
Neuroscience, 23, 3366–3379.
Lehtonen, M., Vorobyev, V. A., Hugdahl, K., Tuokkola, T.,
& Laine, M. (2006). Neural correlates of morphological
decomposition in a morphologically rich language:
An fMRI study. Brain and Language, 98, 182–193.
Leinonen, A., Grönholm-Nyman, P., Järvenpää, M.,
Söderholm, C., Lappi, O., Laine, M., et al. (2009).
Neurocognitive processing of auditorily and visually
presented inflected words and pseudowords: Evidence
from a morphologically rich language. Brain Research,
1275, 54–66.
Lewis, G., Solomyak, O., & Marantz, A. (2011). The neural
basis of obligatory decomposition of suffixed words.
Brain and Language, 118, 118–127.
Fiorentino, R., & Poeppel, D. (2007). Compound words and
Longtin, C. M., & Meunier, F. (2005). Morphological
structure in the lexicon. Language and Cognitive Processes,
22, 1–48.
Forster, K. I., & Davis, C. (1984). Repetition priming and frequency
attenuation in lexical access. Journal of Experimental
Psychology: Learning, Memory and Cognition, 10, 680–698.
Forster, K. I., & Forster, J. C. (2003). DMDX: A Windows display
program with millisecond accuracy. Behavior Research
Methods, Instruments, & Computers, 35, 116–124.
Frost, R. (2012). Towards a universal model of reading.
Brain & Behavioral Sciences, 35, 1–67.
Halgren, E., Dhond, R. P., Christensen, N., Van Petten, C.,
Marinkovic, K., Lewine, J. D., et al. (2002). N400-like
magnetoencephalography responses modulated by
semantic context, word frequency, and lexical class in
sentences. Neuroimage, 17, 1101–1116.
decomposition in early visual word processing. Journal
of Memory and Language, 53, 26–41.
Longtin, C. M., Segui, J., & Halle, P. A. (2003). Morphological
priming without morphological relationship. Language
and Cognitive Processes, 18, 313–334.
Marslen-Wilson, W. D. (2007). Morphological processes in
language comprehension. Oxford handbook of psycholinguistics
(pp. 175–193). Oxford: Oxford University Press.
Marslen-Wilson, W. D., Bozic, M., & Randall, B. (2008). Early
decomposition in visual word recognition: Morphology,
form, and meaning. Language and Cognitive Processes,
23, 394–421.
Marslen-Wilson, W. D., & Tyler, L. K. (2007). Morphology,
language and the brain: The decompositional substrate
for language comprehension. Philosophical Transactions
264
Journal of Cognitive Neuroscience
Volume 27, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
of the Royal Society, Series B, Biological Sciences, 362,
823–836.
Matthews, P. H. (1991). Morphology. Cambridge: Cambridge
University Press.
Morris, J., Grainger, J., & Holcomb, P. J. (2008). An
electrophysiological investigation of early effects of masked
morphological priming. Language and Cognitive Processes,
23, 1021–1056.
Pammer, K., Hansen, P. C., Kringelbach, M. L., Holliday, I.,
Barnes, G., Hillebrand, A., et al. (2004). Visual word
recognition: The first half second. Neuroimage, 22,
1819–1825.
Price, C. J., & Devlin, J. T. (2011). The interactive account of
ventral occipitotemporal contributions to reading. Trends
in Cognitive Sciences, 15, 246–253.
Pylkkänen, L., & Marantz, A. (2003). Tracking the time course of
word recognition with MEG. Trends in Cognitive Sciences,
7, 187–189.
Pylkkänen, L., Stringfellow, A., & Marantz, A. (2002). Neuromagnetic
evidence for the timing of lexical activation: An MEG
component sensitive to phonotactic probability but
not to neighborhood density. Brain and Language, 81,
666–678.
Rastle, K., & Davis, M. H. (2008). Morphological decomposition
based on the analysis of orthography. Language and
Cognitive Processes, 23, 942–971.
Rastle, K., Davis, M. H., Marslen-Wilson, W. D., & Tyler, L. K.
(2000). Morphological and semantic effects in visual word
recognition. Language and Cognitive Processes, 15, 507–537.
Rastle, K., Davis, M. H., & New, B. (2004). The broth in my
brotherʼs brothel: Morpho-orthographic segmentation
in visual word recognition. Psychonomic Bulletin and
Review, 11, 1090–1098.
Shtyrov, Y., Goryainova, G., Tugin, S., Ossadtchi, A., &
Shestakova, A. (2013). Automatic processing of unattended
lexical information in visual oddball presentation:
Neurophysiological evidence. Frontiers in Human
Neuroscience, 7, 1–10.
Taulu, S., & Simola, J. (2006). Spatiotemporal signal space
separation method for rejecting nearby interference in
MEG measurements. Physics in Medicine and Biology,
51, 1759–1768.
Turken, U., & Dronkers, N. F. (2011). The neural architecture of
the language comprehension network: Converging evidence
from lesion and connectivity analyses. Frontiers in Systems
Neuroscience, 5, 1–20.
Twomey, T., Kawabata Duncan, K. J., Price, C. J., & Devlin,
J. T. (2011). Top–down modulation of occipitotemporal
responses during visual word recognition. Neuroimage,
55, 1242–1251.
Tyler, L. K., Stamatakis, E. A., Post, B. E., Randall, B., &
Marslen-Wilson, W. D. (2005). Temporal and frontal systems
in speech comprehension: An fMRI study of past tense
processing. Neuropsychologia, 43, 1963–1974.
van Atteveldt, N. M., Formisano, E., Goebel, R., & Blomert, L.
(2007). Top–down task effects overrule automatic
multisensory responses to letter–sound pairs in auditory
association cortex. Neuroimage, 36, 1345–1360.
Vartiainen, J., Aggujaro, S., Lehtonen, M., Hulten, A.,
Laine, M., & Salmelin, R. (2009). Neural dynamics of
reading morphologically complex words. Neuroimage,
47, 2064–2072.
Vinckier, F., Dehaene, S., Jobert, A., Dubus, J. P., Sigman, M.,
& Cohen, L. (2007). Hierarchical coding of letter strings in
the ventral stream: Dissecting the inner organization of
the visual word-form system. Neuron, 55, 143–156.
Whiting, C. M. (2011). Spatiotemporal dynamics of
morphological and lexical processing in the brain.
Ph.D. thesis, University of Cambridge, Cambridge, UK.
Zanto, T. P., Rubens, M. T., Thangavel, A., & Gazzaley, A.
(2011). Causal role of the prefrontal cortex in top–down
modulation of visual processing and working memory.
Nature Neuroscience, 14, 656–661.
Zhuang, J., Tyler, L. K., Randall, B., Stamatakis, E. A., & Marslen-
Wilson, W. D. (2012). Optimally efficient neural systems for
processing spoken language. Cerebral Cortex. doi:10.1093/
cercor/bhs366.
Zwieg, E., & Pylkkänen, L. (2009). A visual M170 effect of
morphological complexity. Language and Cognitive
Processes, 24, 412–439.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
2
2
7
2
/
4
2
6
/
2
2
0
4
0
6
6
/
2
1
7
8
6
2
o
3
c
4
n
9
_
7
a
/
_
j
0
o
0
c
6
n
9
9
_
a
p
_
d
0
0
b
6
y
9
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Whiting, Shtyrov, and Marslen-Wilson
265