Processing Speech and Thoughts during Silent Reading:
Direct Reference Effects for Speech by Fictional
Characters in Voice-Selective Auditory Cortex
and a Theory-of-Mind Network
Ben Alderson-Day1, Jamie Moffatt1,2, Marco Bernini1, Kaja Mitrenga1
Bo Yao3, and Charles Fernyhough1
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
.
F
t
/
o
n
0
5
M.
un
oui
2
0
2
1
Abstrait
■ Stories transport readers into vivid imaginative worlds, mais
understanding how readers create such worlds—populating
them with characters, objets, and events—presents serious
challenges across disciplines. Auditory imagery is thought to
play a prominent role in this process, especially when repre-
senting characters’ voices. Previous research has shown that
direct reference to speech in stories (par exemple., He said, “I’m over
here”) may prompt spontaneous activation of voice-selective
auditory cortex more than indirect speech [Yao, B., Belin,
P., & Scheepers, C. Silent reading of direct versus indirect
speech activates voice-selective areas in the auditory cortex.
Journal des neurosciences cognitives, 23, 3146–3152, 2011].
Cependant, it is unclear whether this effect reflects differential
processing of speech or differences in linguistic content,
source memory, or grammar. One way to test this is to com-
pare direct reference effects for characters speaking and
thinking in a story. Ici, we present a multidisciplinary
fMRI study of 21 readers’ responses to characters’ speech
and thoughts during silent reading of short fictional stories.
Activations relating to direct and indirect references were
compared for both speaking and thinking. Eye-tracking and
independent localizer tasks (auditory cortex and theory of
esprit [ToM]) established ROIs in which responses to stories
could be tracked for individuals. Evidence of elevated auditory
cortex responses to direct speech over indirect speech was
observed, replicating previously reported effects; no reference
effect was observed for thoughts. De plus, a direct reference
effect specific to speech was also evident in regions previously
associated with inferring intentions from communication.
Implications are discussed for the spontaneous representation
of fictional characters and the potential roles of inner speech
and ToM in this process. ■
INTRODUCTION
Stories can conjure complex imaginative worlds that offer
immersion and transportation for the reader (Vert,
2004; Vert, Brock, & Kaufman, 2004; Ryan, 1999;
Gerrig, 1993). Fictional characters in particular are some-
times experienced with a vividness and complexity,
which can linger beyond the page (Alderson-Day,
Bernini, & Fernyhough, 2017; Maslej, Oatley, & Mar,
2017). Understanding how these experiences are created
by the mind—often with apparent automaticity and
spontaneity—is a challenge for a wide range of disciplines
beyond psychology, including literary theory, narratology,
philosophy of mind, and cognitive neuroscience
(Herman, 2013). Far from passively “receiving” informa-
tion from the writer, readers actively and creatively en-
gage with fictional texts in a way that draws on multiple
1Durham University, 2University of Sussex, 3Université de
Manchester
psychological resources (Polvinen, 2016; Caracciolo,
2014; Kukkonen, 2014; Oatley, 2011; Bortolussi &
Dixon, 2003).
One approach to understanding the qualitative rich-
ness of the reading experience has been to study inner
speech (sometimes also referred to as inner monologue
or articulatory imagery; Alderson-Day & Fernyhough,
2015; Perrone-Bertolotti, Rapin, Lachaux, Baciu, &
Lœvenbruck, 2014). Intuitively, reading is often associated
with the sounding out of an “inner voice,” and the self-
reports of readers involve various kinds of auditory imag-
ery when engaged in a story (Vilhauer, 2016). Although
the reliability of readers’ introspective reports has been
questioned (Caracciolo & Hurlburt, 2016), empirical evi-
dence of inner speech involvement during silent reading
is well documented (Filik & Barber, 2011; Alexander &
Nygaard, 2008). De plus, silent reading appears to elicit
activity in perisylvian regions and auditory association
cortex (Magrassi, Aromataris, Cabrini, Annovazzi-Lodi, &
Moro, 2015; Perrone-Bertolotti et al., 2012), particularly
when characters’ voices and speech are being described
© 2020 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 International (CC PAR 4.0) Licence.
Journal des neurosciences cognitives 32:9, pp. 1637–1653
https://doi.org/10.1162/jocn_a_01571
(Brück, Kreifelts, Gößling-Arnold, Wertheimer, &
Wildgruber, 2014; Yao, Belin, & Scheepers, 2011). Tel
findings have been taken as evidence of the reading
experience—and its evocation of inner speech—being
almost akin to hearing external voices (Petkov & Belin,
2013).
A good example of this is provided by texts involving
direct speech. When direct reference is made to a char-
acter overtly speaking in a text (he said, “the cat is over
there”), it is thought to evoke a more vivid experience of
the storyworld than if the same overt speech is only indi-
rectly referred to (he said that the cat is over there). Il
has been suggested that the purpose of such construc-
tions is to demonstrate (and thus depict) a situation,
rather than merely describe it (Clark & Gerrig, 1990).
Evidence that this could resemble hearing an actual voice
is provided by Yao et al. (2011), who compared fMRI re-
sponses in auditory cortex for participants silently read-
ing short stories that contained either direct or indirect
reference to speech. Although both kinds of speech acti-
vated auditory cortex, direct speech was associated with a
greater response than indirect speech in voice-selective
regions of the right superior and middle temporal lobe
(as defined by a separate auditory localizer task; Belin,
Zatorre, Lafaille, Ahad, & Pike, 2000). As the stories were
very short (three to four sentences in total) and partici-
pants were not prompted to imagine the voices, charac-
ters, or stories in any specific way, this suggests that fairly
minimal textual markers for direct speech can elicit a re-
sponse in cortical regions that are selective for voice
perception.
If direct speech in text can prompt this kind of reac-
tion, a second question is why readers appear to respond
in this way. In a separate study, Yao, Belin, and Scheepers
(2012) observed similarly enhanced responses in voice-
selective regions for direct speech quotations when they
were being read by a monotonous voice. Building on
Barsalou’s theory of embodied cognition (Barsalou,
2008), they suggested that auditory cortex activation
may have a role in constructing a perceptual simulation
of the emotional prosody and intonation of the speaker’s
voice, given that such information is either absent or di-
minished in the case of both silent reading and monoto-
nous listening. This would not rule out perceptual
simulation during other kinds of silent reading but char-
acterizes direct reference as a cue to simulate supraseg-
mental and communicative properties of speech from
text ( Yao et al., 2011, 2012).
The effects of direct speech and its potential conse-
quences for simulation can be questioned, cependant. If di-
rect speech prompts more vivid imagery or provides
more communicative information (par exemple., tone or emotional
content), this would plausibly be reflected in reader com-
prehension. Cependant, in a series of behavioral experi-
ments, Eerland, Engelen, and Zwaan (2013) reported
inconsistent evidence for either perceptual or communi-
cative information being more available to readers after
direct speech quotations. Plutôt, they suggested that
the use of direct quotations prompts better memory for
the verbatim content of characters’ utterances, alors que
indirect speech assists the building of a situation model,
c'est, an overall “representation of the referential situa-
tion” (Eerland et al., 2013, p. 7; van Dijk & Kintsch, 1983).
Supporting this, source memory for characters’ utter-
ances is actually enhanced for indirect, not direct, speech
quotations (Eerland & Zwaan, 2018)—suggesting that the
potential vividness of direct speech is not used for track-
ing information about who said what (or could even ob-
struct such tracking, when compared to indirect speech).
Enfin, the typographical and grammatical differences be-
tween direct and indirect speech make it difficult to clearly
compare their specific consequences for mental simula-
tion. Along with potentially alerting the reader to pay
attention to text, direct sentences are typically shorter
than indirect sentences, are syntactically simpler, et
may be expected to prompt changes in reader perspective
(Köder, Maier, & Hendriks, 2015; Coulmas, 2011; Clark &
Gerrig, 1990). En tant que tel, the effect of direct speech on the
lecteur, and its potential function in the imaginative re-
sponse of reading, remains unclear.
One way to explore this topic—in a way that might be-
gin to address some of the above concerns—is to com-
pare references to characters’ speech with another kind
of representation that fictional narratives can involve:
characters’ thoughts. Although theories of mental simula-
tion during reading emphasize various forms of sensory
and embodied simulation (par exemple., Kurby & Zacks, 2013;
Zwaan, Madden, Yaxley, & Aveyard, 2004), fictional narra-
tives have been proposed to place specific sociocognitive
demands on the reader (Mar & Oatley, 2008; Zunshine,
2006). Typiquement, a reader must track the mental states of
multiple characters, following their beliefs, intentions,
and desires through a narrative, to make sense of actions,
decisions, and responses to events in the storyworld
(Spreng, Mar, & Kim, 2009; Herman, 2008; Palmer,
2004; Gerrig, Brennan, & Ohaeri, 2001),1 all of which im-
ply a central role for theory of mind (ToM) in the reading
processus.
How might this shed light on direct speech? First is be-
cause it provides a contrasting example of direct refer-
ence. Both indirect and direct references to thinking
are used in narratives. Indirect thought, which is usually
considered the representational norm (Leech & Short,
2007, p. 268), is more flexible and can be used to repre-
sent verbal, preverbal, and nonverbal mental processes
from the perspective of the character (par exemple., he thought
that X; he felt that Y; he was willing to do Z ). Direct
thought (also referred to as “quoted monologue”;
Cohn, 1978) is used to represent, verbatim, the linguistic
silent articulation of verbal thoughts (He thought “this is
so complicated!»).
The verbalized nature of depicting characters’ thoughts
is almost identical in form and complexity to direct speech
(c'est à dire., when used in a basic form; indirect thoughts in more
1638
Journal des neurosciences cognitives
Volume 32, Nombre 9
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
t
F
/
.
o
n
0
5
M.
un
oui
2
0
2
1
extended narratives can be used in highly complex ways).
Contrasting these forms of speech and verbal thought can
therefore provide a test of Yao et al.’s (2011) interpreta-
tion of direct reference effects by assessing how specific
they might be to vocal information, while at the same
time controlling for typographic features. If Yao et al.’s
conjecture is correct, direct reference to speech—but
not necessarily thoughts—may be expected to elicit re-
cruitment of voice-selective regions of auditory cortex,
to specifically simulate the perceptual qualities of charac-
ters speaking out loud in the storyworld. In contrast, si
direct speech and direct thought elicit similar responses,
then a voice-specific account of direct reference would be
harder to maintain. It could be the case that both speech
and thoughts elicit some form of perceptual simulation
under direct reference, but they would be doing so de-
spite clear dissimilarities in the auditory scenario (one is
an external utterance, and the other is a form of internal
monologue). Plutôt, showing that direct reference to
speech and thoughts prompts a generally greater re-
sponse in auditory cortex could support alternative in-
terpretations: It may, Par exemple, simply reflect a
greater level of engagement that happens when quote
marks prompt the reader to pay attention to verbatim
content (Eerland et al., 2013). In that scenario, là
would be nothing special about speech for understand-
ing direct reference effects.
Deuxième, exploring sociocognitive processing and con-
trasting how this works for speaking and thinking are po-
tentially highly informative for understanding direct
speech effects. ToM has multiple components, each with
its own developmental trajectory (Fernyhough, 2008;
Tomasello, Carpenter, Call, Behne, & Moll, 2005).
Whereas some socio-cognitive skills are evident early in
infancy—such as the ability to follow others’ attentional
cues (Behne, Liszkowski, Carpenter, & Tomasello, 2012;
Woodward, 1998)—the ability to cognitively represent
others’ mental states when incorrect is thought to emerge
later in childhood (typically around 4 ans; Wellman,
Cross, & Watson, 2001). De la même manière, understanding prag-
matic information and speaker intention from prosody
shows a competence–performance gap, in which vocal
cues to emotion are recognized very early in infancy but
are only used consistently (in the face of, par exemple., conflicting
cues) in older children (Esteve-Gibert & Guellaï, 2018).
ToM and social cognition more generally are associated
with a canonical network of regions in the medial pFC
(mPFC), precuneus, and TPJ bilaterally (Molenberghs,
Johnson, Henry, & Mattingley, 2016; Schurz, Radua,
Aichhorn, Richlan, & Perner, 2014; Saxe & Kanwisher,
2003; Fletcher et al., 1995). Of these, representing the
thoughts and intentions of others in particular has been
argued to localize to regions of the TPJ and precuneus
(Schurz, Tholen, Perner, Mars, & Sallet, 2017; Saxe &
Powell, 2006), whereas mPFC has been linked to the pro-
cessing of more constant traits associated with self and
other (van Overwalle, 2009).
If direct speech prompts a detailed simulation of su-
prasegmental vocal information (such as emotional tone
or prosody), then this may also be reflected in social–
cognitive regions—specifically for areas associated with
interpreting or reasoning about a speaker’s communica-
tive intentions. Par exemple, using a nonverbal, cartoon-
based story task, Ciaramidaro et al. (2007) observed that
bilateral TPJ regions in particular are associated with
tracking different kinds of intent associated with socio-
communicative interactions. If direct speech prompted
similar activation, this would support an extension of
Yao et al.’s (2011) original theory to suggest that direct
reference involves constructing a broader, sociopercep-
tual simulation than merely how a voice sounds.
Contrastingly, if tracking characters and their intentions
is an ultimately separate process from simulating the per-
ceptual features of characters’ voices, then no direct ef-
fect for speech would necessarily be expected in ToM
régions. Plutôt, it is possible that references to charac-
ters’ mental states—but not their speech—would be
most likely to engage such regions, irrespective of any di-
rect reference effect.
To investigate this, we adapted Yao et al.’s (2011) par-
adigm to include direct and indirect references to charac-
ters’ verbal thoughts and speech in a 2 × 2 conception. Nous
used eye tracking and an auditory localizer task to study
cortical responses specific to each individual’s reading
times and voice-selective regions. To explore the broader
effect of direct speech in regions commonly associated
with inferring communicative intentions, we also included
a version of Ciaramidaro et al.’s (2007) story task as a sec-
ond localizer. Many standard ToM tasks use written short
stories in which characters’ false beliefs must be inferred
from textual information, but using such stories could be
expected to overlap considerably with other reading
tasks (in terms of both stimuli and task demands).
Plutôt, by using a wordless, cartoon-based ToM task,
we could avoid this potential confound with the demands
of our main direct/indirect story task. On the basis of the
original findings of Yao et al. (2011, 2012), we hypothe-
sized that (je) direct reference effects would be evident for
speech but not thoughts in auditory cortex. In accor-
dance with the claim that this facilitates prosodic and
communicative processing of the utterance, we also pre-
dicted that (ii) the voice-specific effect of direct reference
would extend to ToM-related regions. In contrast, Non
direct reference effects were expected for thoughts,
in either network.
MÉTHODES
Participants
An initial sample of 30 individuals took part in the full
MRI procedure, but nine participants did not produce a
full data set because of the following exclusions (one in-
cidental finding, one insufficient accuracy [<60%] on the
Alderson-Day et al.
1639
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
9
e
1
-
6
p
3
d
7
f
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
a
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
f
.
/
o
n
0
5
M
a
y
2
0
2
1
story task, two no clear voice-selective response on the
auditory localizer task, five insufficient eye-tracking data;
three male, six female). As such, analysis proceeded with
a final sample of 21 (age: M = 23.49, SD = 6.63; three
male, 18 female). All participants were right-handed, na-
tive English speakers, with normal or corrected-to-normal
vision. All procedures were approved by a university
ethics subcommittee.
Measures
Story Task
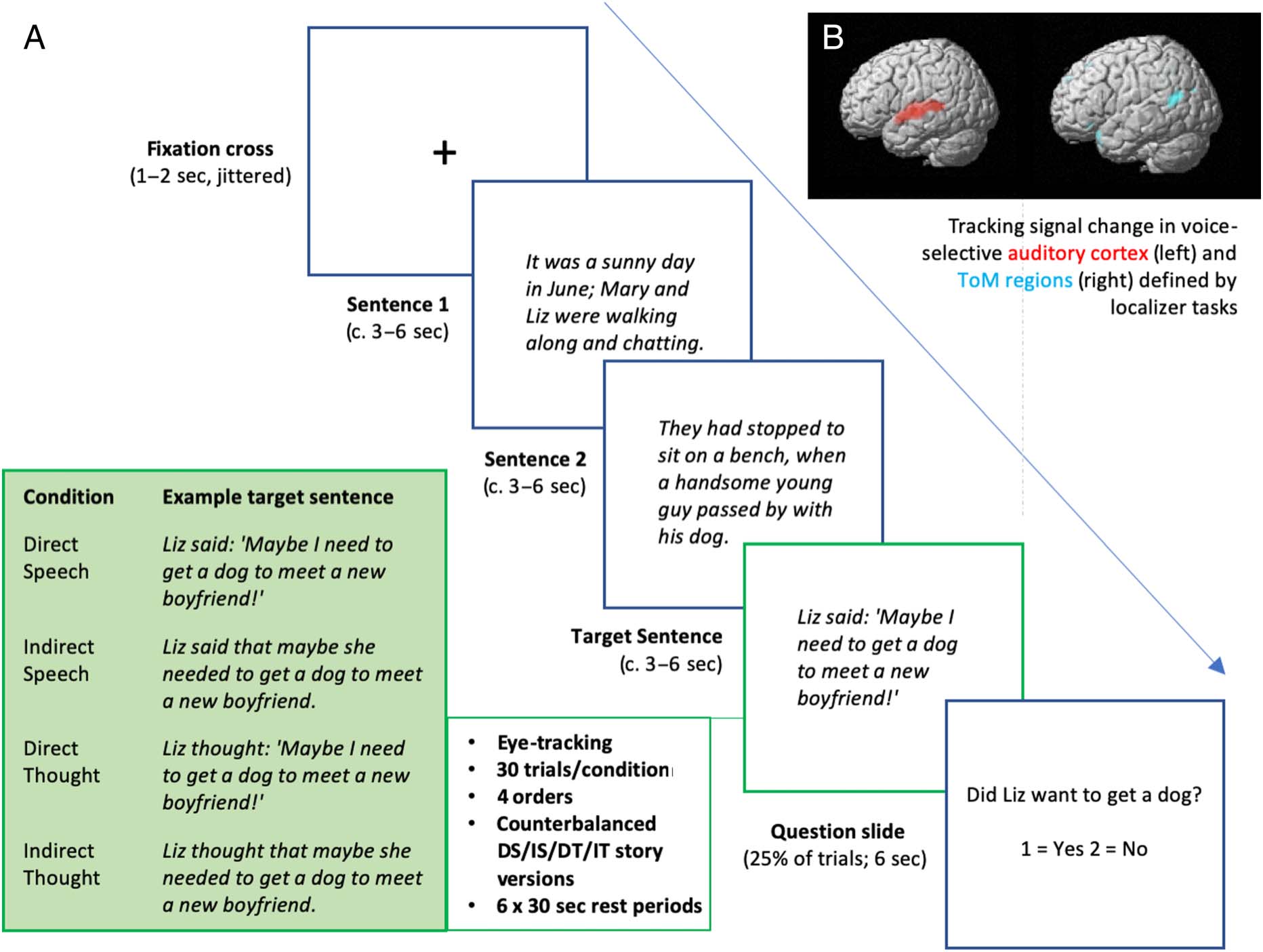
Following Yao et al. (2011), participants viewed a series of
short stories containing two preparation sentences
(Sentences 1 and 2) and a target sentence, containing a
character either (i) speaking or thinking with (ii) direct
or indirect reference. On each trial, participants viewed a
fixation cross for 1–2 sec ( jittered at random), followed by
one slide per sentence, presented sequentially (see
Figure 1). Viewing times per slide were determined using
the following formula: ( Words × 100 msec) + (Syllables ×
50 msec) + 2000 msec. Mean presentation times were
5.61 and 5.72 sec for Sentences 1 and 2, respectively,
and 5.95 and 6.22 sec for direct and indirect target sen-
tences, respectively, reflecting the slightly longer length
of indirect sentences on average (18.6 words per indirect
sentence compared to 16.8 for direct sentences). To allow
for sufficient trials in each condition, the number of stories
was increased from the 90 trials used in Yao et al. (2011) to
120, split across two 20-min runs (additional stories were
prepared by a narratologist, M. B., to follow the length,
complexity, and style of the original stimuli and ensure
balance across the four conditions). Each run also con-
tained three 30-sec break periods, occurring every 20 trials.
An attentional check (a simple comprehension question
relating to factual content from the preceding story) was
included after 25% of trials, with participants having 6 sec
to respond.2 Four random orders of trials were generated,
counterbalancing the combination of voice/thought and
direct/indirect target sentences across participants. Eye-
tracking timings were collected as an indicator of partici-
pants’ reading responses for the two preparation sentences
and the target sentence. Specifically, participants’ first
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
9
e
1
-
6
p
3
d
7
f
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
a
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
t
.
f
o
n
0
5
M
a
y
2
0
2
1
Figure 1. Adapted story task (A) with direct reference to voices and thoughts applied to auditory and ToM localizer regions (B). Figure 1B depicts
left-sided sagittal view (rendered, p < .05, FWE); note that auditory and ToM regions were observed bilaterally (see Table 1).
1640
Journal of Cognitive Neuroscience
Volume 32, Number 9
Table 1. Accuracy Rates, Reading Onsets, and Reading Times by Task Condition
Direct Speech
Indirect Speech
Direct Thought
Indirect Thought
M
SD
Accuracy (%)
79.84
18.43
Reading onset (sec)
Duration (sec)
0.56
4.02
0.32
0.54
M
79.21
0.57
4.10
SD
16.08
0.33
0.54
M
85.56
0.61
4.06
SD
15.87
0.36
0.44
M
86.35
0.67
4.11
SD
15.90
0.36
0.58
fixation (the beginning of the sentence) and last fixation
(the final line of the target sentence) within the text
area were used to define reading onsets and offsets of
characters’ speaking and thinking in the target sentence.
These were then directly included in the fMRI model
to account for individual differences in the reading
response.
Auditory Localizer Task
The auditory localizer task was identical to that used in
Yao et al. (2011). Participants listened to 20 blocks of vo-
cal stimuli and 20 blocks of nonvocal stimuli, along with
20 silent blocks that were used as a baseline. The blocks
were presented randomly. Each block was 8 sec long, and
the task lasted 10 min. The contrasting brain activity in
response to the vocal and nonvocal stimuli reliably local-
izes voice-selective areas of the auditory cortex ( Yao
et al., 2011; Belin et al., 2000).
ToM Task
The cartoon-based ToM task was adapted from a task
used by Walter et al. (2004) and Ciaramidaro et al.
(2007). Participants viewed a sequence of three cartoon
story vignettes (“story” phase) and were required to indi-
cate a logical end of each story based on the three pre-
sented images (“choice” phase). The story phase
included either reasoning about characters’ intentions
when communicating with others (e.g., a man indicating
whether a seat is free on a train) or physical reasoning
(e.g., a water pipe bursting). The images were displayed
sequentially for 3 sec in the story phase and for 7 sec in
the choice phase. The intertrial intervals lasted between 7
and 11 sec. In total, 10 ToM stories and 10 physical
reasoning stories were presented in a random order.
Participants answered (A, B, or C) by a button press.
The task took 9 min to complete. The contrasting
brain activity in response to the ToM reasoning stories
compared to physical reasoning stories has been ob-
served to prompt activity in brain regions often associated
with ToM, including the right TPJ, precuneus, and anterior
paracingulate cortex (Alderson-Day et al., 2016; Ciaramidaro
et al., 2007; Walter et al., 2004).
Data Acquisition
fMRI data were acquired at Durham University
Neuroimaging Centre using a 3-T Magnetom Trio MRI
system (Siemens Medical Systems) with standard gradi-
ents and a 32-channel head coil. T2*-weighted axial EPI
scans were acquired with the following parameters:
field of view = 212 mm, flip angle = 90°, repetition
time = 2000 msec, echo time = 30 msec, number of
slices = 32, slice thickness = 3.0 mm, interslice gap =
0.3 mm, and matrix size = 64 × 64. Story task data
were collected across 2 × 20-min runs consisting of
600 volumes each; auditory and ToM tasks took roughly
10 min each and consisted of 300 and 281 volumes, re-
spectively. The first three volumes of each EPI run were
discarded to allow for equilibrium of the T2 response.
For each participant, an anatomical scan was acquired
using a high-resolution T1-weighted 3-D sequence
(number of slices = 192, slice thickness = 1 mm, matrix
size = 512 × 512, field of view = 256 mm, echo time =
2.52 msec, repetition time = 2250 msec, flip angle =
9°). Eye-tracking data were collected using a LiveTrack
system (Cambridge Research Systems) with MATLAB
2016b (The Mathworks, Inc.).
Data Analysis
All MRI analyses were conducted using SPM Version 12
( Wellcome Department of Cognitive Neurology) imple-
mented in MATLAB. Images were slice-time corrected
before being realigned to the first image to correct for
head movement. Volumes were then normalized into
standard stereotaxic anatomical Montreal Neurological
Institute space using the transformation matrix calculated
from the first EPI scan of each participant and the EPI
template. The default settings for normalization in
SPM12 and the standard EPI template supplied with
SPM12 were used. The normalized data with a resliced
voxel size of 2 × 2 × 2 mm were smoothed with an
8-mm FHWM isotropic Gaussian kernel to accommodate
intersubject anatomical variation. The time-series data
were high-pass filtered with a high-pass cutoff of 1/128 Hz,
and first-order autocorrelations of the data were estimated
and corrected for. Movement parameters from the re-
alignment phase were visually inspected for outliers and
included as regressors for single-participant (first-level)
Alderson-Day et al.
1641
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
9
e
1
-
6
p
3
d
7
f
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
a
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
.
f
/
o
n
0
5
M
a
y
2
0
2
1
Table 2. Whole-Brain Coordinates for (A) Auditory and (B) ToM Localizer Tasks
k
t
z
pFWE
Location
x
y
(A) Auditory (vocal > nonvocal)
L middle temporal gyrus
L superior temporal gyrus
L middle temporal gyrus
R superior temporal gyrus
R temporal pole
R superior temporal gyrus
−60
−58
−60
56
48
64
−14
−2
−36
−18
12
−4
(B) ToM (communicative inference > physical reasoning)
R middle cingulate cortex
L precuneus
WM
R middle temporal gyrus
R superior temporal gyrus
R middle temporal gyrus
WM
L temporal pole
L middle temporal gyrus
L superior medial frontal gyrus
R superior medial frontal gyrus
L posterior-medial frontal gyrus
L gyrus rectus
4
−2
−14
48
58
56
−36
−38
−62
−8
4
−4
0
−56
−50
−50
−50
−46
2
−56
22
−56
32
54
14
52
z
−2
−4
6
0
−18
0
40
46
34
24
20
−22
16
−22
18
52
36
60
3512
5051
1674
5835
5472
4021
17.08
11.67
11.42
12.76
11.85
10.58
12.43
10.25
9.51
12.24
12.03
10.44
11.59
11.45
10.35
10.68
9.21
7.92
7.76
7.34
6.34
6.29
6.58
6.39
6.08
5.95
5.10
5.33
5.92
5.88
5.55
5.79
5.76
5.53
5.60
5.26
4.90
4.85
<.001
<.001
<.001
<.001
<.001
<.001
.015
−14
279
All results p < .05, FWE, at peak/cluster level. Minimum cluster size: k = 50. L = left; R = right; ToM = theory of mind; WM = white matter.
analyses. ROI analyses were conducted using the
Marsbar toolbox (Brett, Anton, Valabregue, & Poline,
2002). Individual ROIs were defined using p < .05 cor-
rected for FWE at cluster level, in temporal cortical re-
gions for the auditory localizer task and clusters in
mPFC, precuneus, and TPJ regions for the ToM task.
Where significant clusters were not evident for individ-
ual participants at this level, a more liberal threshold of
p < .001 (uncorrected) was used to maximize sensitiv-
ity to individual differences; participants who showed
no clusters in these regions even at the more liberal
threshold were excluded from analyses (two auditory,
six ToM). All whole-brain analyses are presented at
p < .05, FWE, cluster-level corrected. All statistical anal-
yses of mean beta values were conducted using R and
jamovi; figures were generated using ggplot2 and
MicroGL. Effect sizes are reported as Cohen’s d for
pairwise comparisons and ηp
2 values for ANOVA main
and interaction effects. ηp
2 values can be considered
as small, moderate, and large effects with values of
.099, .0588, and .1379, respectively (Richardson, 2011;
Cohen, 1969).
RESULTS
Accuracy on the task was generally high (M = 82.5%, SD =
7.4%) indicating that participants maintained attention
despite the 40-min duration of the task. Repeated-
measures ANOVA with a 2 × 2 (Form × Reference) de-
sign was used to compare behavioral responses for the
four conditions (see Table 1). No main effects, interac-
tion effects, or pairwise comparisons were significant
for condition accuracy, although we observed a nonsig-
nificant trend for participants to be slightly less accurate
on speech trials compared with thought trials, F(1, 20) =
3.34, p = .082, ηp
2 = .14 ( p > .14 for all other effects
and comparisons). For the duration of reading times,
the only effect close to significance was for direct com-
pared with indirect reference, F(1, 20) = 3.57, p = .073,
1642
Journal des neurosciences cognitives
Volume 32, Nombre 9
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
t
/
F
.
o
n
0
5
M.
un
oui
2
0
2
1
Tableau 3. Whole-Brain Coordinates for Speech and Thought Sentences vs. Baseline
Location
Direct speech
L middle temporal gyrus
L middle temporal gyrus
L middle temporal gyrus
R temporal pole
R temporal pole
L temporal pole
L inferior frontal gyrus
L inferior frontal gyrus
L superior medial frontal gyrus
L precentral gyrus
Indirect speech
L middle temporal gyrus
L middle temporal gyrus
L middle temporal gyrus
L temporal pole
L inferior frontal gyrus
L temporal pole
R medial temporal pole
R temporal pole
L middle temporal gyrus
L inferior frontal gyrus
L middle temporal gyrus
L middle temporal gyrus
L middle temporal gyrus
Direct thought
L middle temporal gyrus
L middle temporal gyrus
L middle temporal gyrus
R medial temporal pole
L superior medial frontal gyrus
L superior frontal gyrus
R middle temporal gyrus
L inferior frontal gyrus
X
oui
z
k
t
z
pFWE
−50
−62
−56
48
44
−48
−50
−46
−10
−44
−54
−50
−60
−50
−48
−46
50
44
−52
−54
−54
−50
−60
−54
−50
−62
50
−8
−10
52
−54
−26
−18
−46
14
22
20
22
16
54
−2
−34
−26
−20
12
28
22
16
22
−52
20
−34
−26
−20
−34
−26
−18
10
56
58
−36
20
−6
−8
4
−24
−28
−14
20
24
28
54
−2
−4
−6
−22
−8
−14
−28
−26
18
22
−2
−4
−6
0
−4
−8
−24
28
38
−2
24
1198
409
895
124
112
550
292
308
144
76
550
11.25
9.66
8.55
10.92
9.21
8.83
8.64
8.31
8.63
8.43
11.84
10.42
7.73
9.35
7.57
7.52
9.31
8.46
7.87
7.79
11.84
10.42
7.73
689
10.79
8.6
8.31
9.6
8.75
6.69
8.16
7.18
344
65
50
53
6.25
5.83
5.49
6.17
5.69
5.58
5.51
5.41
5.51
5.45
6.38
6.04
5.2
5.74
5.14
5.12
5.73
5.45
5.25
5.22
6.38
6.04
5.2
6.13
5.5
5.41
5.81
5.55
4.79
5.35
4.99
<.001
<.001
<.001
<.001
<.001
<.001
<.001
<.001
<.001
<.001
<.001
<.001
<.001
<.001
<.001
<.001
Alderson-Day et al.
1643
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
9
e
1
-
6
p
3
d
7
f
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
a
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
t
.
o
n
0
5
M
a
y
2
0
2
1
Table 3. (continued )
Location
Indirect thought
L middle temporal gyrus
L middle temporal gyrus
L middle temporal gyrus
R medial temporal pole
R temporal pole
R middle temporal gyrus
L superior medial gyrus
L precentral gyrus
L temporal pole
L temporal pole
L temporal pole
x
y
z
k
t
z
pFWE
−54
−50
−54
50
46
50
−10
−42
−52
−44
−46
−34
−26
−54
12
20
−38
54
−2
12
18
16
−2
−6
16
−24
−26
−2
28
56
−20
−16
−32
1048
343
56
76
70
206
11.2
11.07
9.74
10.78
10.4
8.91
8.89
8.52
8.42
8.42
6.69
6.23
6.2
5.85
6.13
6.03
5.6
5.59
5.48
5.44
5.44
4.79
<.001
<.001
<.001
<.001
<.001
<.001
All results p < .05, FWE, at cluster and peak levels. Minimum cluster size: k = 50. L = left; R = right; ToM = theory of mind; WM = white matter.
ηp
2 = .15, which likely reflected the slightly longer lengths
of indirect sentences. All other effects and comparisons
for duration were also nonsignificant (all ps > .15).
Reading onsets, in contrast, showed a main effect of
Form, F(1, 20) = 7.10, p = .015, ηp
2 = .26, such that readers
were quicker to start reading speech trials; follow-up
pairwise comparisons indicated that this was only signifi-
cantly quicker for direct speech compared with indirect
thought, t = 2.20, df = 36.24, p = .035 (uncorrected), d =
0.4, all other ps > .10.
Whole-brain analyses—included here for descriptive
purposes—indicated that the vocal > nonvocal contrast
from the auditory localizer task was associated with sig-
nificantly greater activation in bilateral auditory cortices,
across the middle temporal gyrus (MTG) and superior
temporal gyrus (see Figure 1B and Table 2). Compared
with baseline, each of the four reading task conditions
was associated with temporal activation bilaterally, avec
the largest clusters being observed along the dorsal bank
of the left MTG (Tableau 3).
Responses to Characters’ Speech and Thoughts in
Voice-Selective Auditory Cortex
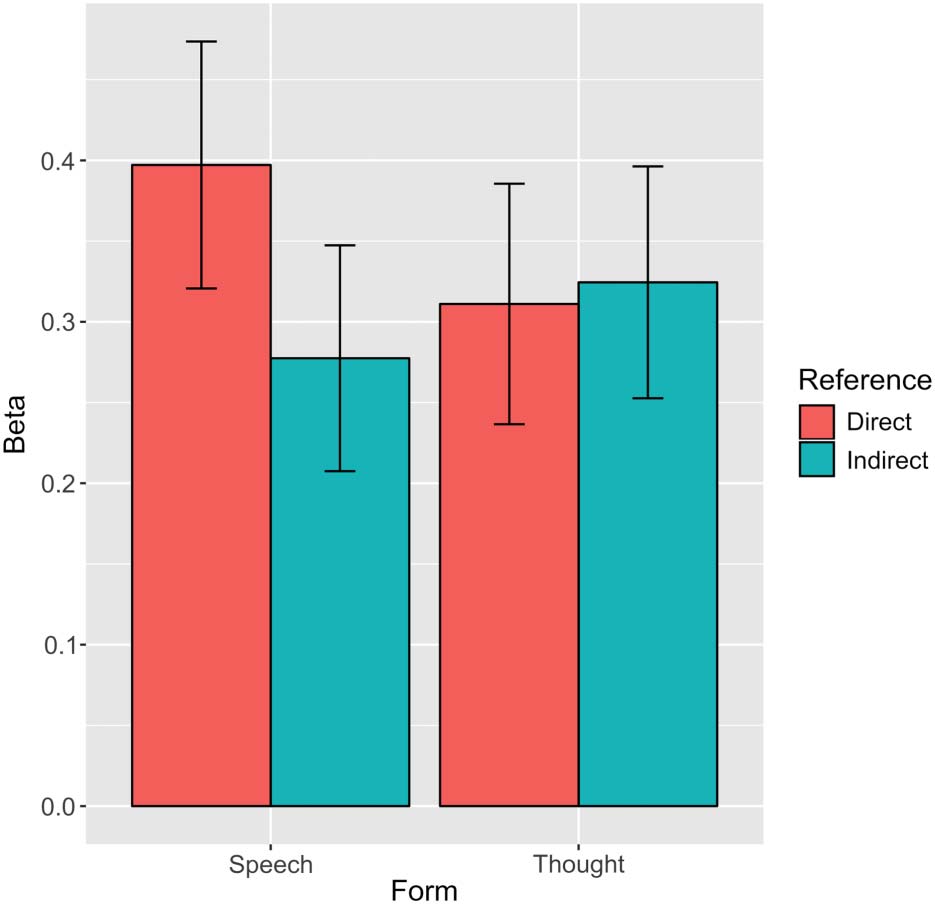
A repeated-measures ANOVA was used to compare mean
beta values in auditory ROIs for story passages containing
characters’ speech or thoughts (c'est à dire., Form) in direct or
indirect reference, in a 2 × 2 conception. No significant main
effect of Form was evident, F(1, 20) = 0.31, p = .584,
ηp
2 = .02, although a trend was observed for reference
in favor of direct quotation, F(1, 20) = 4.00, p = .059,
ηp
2 = .17. The interaction of Form and Reference was sig-
nificant, F(1, 20) = 7.08, p = .015, ηp
2 = .26. As displayed
in Figure 2, this was largely driven by a specific direct
reference effect for character’s speech, but not thoughts.
Pairwise comparisons indicated that mean beta values for
direct speech were significantly higher than those for in-
direct speech ( p = .006, d = 0.84, Bonferroni corrected),
but no other pairwise contrasts were significant (all ps > .25).
Responses to Speech and Thoughts in a
ToM Network
We then applied the same analyses to responses in a ToM
network identified via the cartoons task. As shown in
o
n
0
5
M.
un
oui
2
0
2
1
Chiffre 2. Mean beta values for direct and indirect references to
characters’ speech and thoughts in voice-selective auditory cortex.
Error bars represent SEM.
1644
Journal des neurosciences cognitives
Volume 32, Nombre 9
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
/
.
F
t
in voice-selective regions of the auditory cortex, no signif-
icant increase in signal was seen for this region when cor-
recting across the whole brain for direct versus indirect
speech (see Table 3). No regions were more active in
the reverse contrast (indirect > direct speech).
Other exploratory whole-brain comparisons indicated
few differences between conditions. Two exceptions
were direct speech versus direct thought and direct ref-
erence versus indirect reference (c'est à dire., with speech and
thought sentences combined). Direct speech compared
to direct thought was associated with greater activation
in the right insula and anterior and middle cingulate, dans-
cluding regions bordering on the pre-SMA (voir
Table 4B). Direct reference was observed to predomi-
nantly activate occipital and parietal regions more than
indirect reference (Table 4C). Their reverse contrasts (di-
rect thought > direct speech; indirect > direct) pro-
duced no significant clusters, even at an uncorrected
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
t
.
/
F
o
n
0
5
M.
un
oui
2
0
2
1
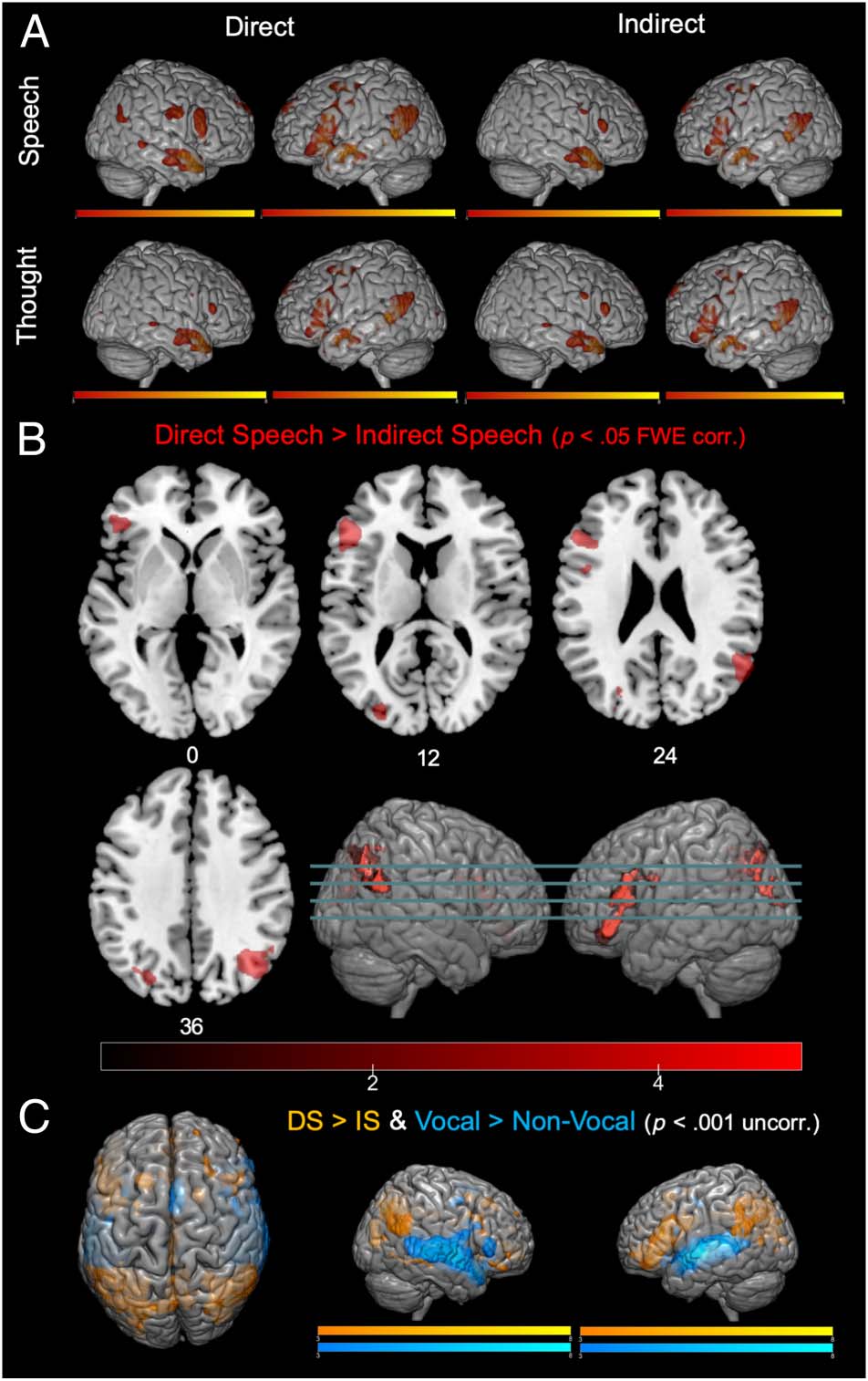
Chiffre 4. Whole-brain activation for (UN) direct and indirect speech and
thoughts versus baseline, (B) direct > indirect speech (DS > IS), et
(C) DS > IS with group-level vocal > nonvocal contrast from auditory
localizer data. All activations significant at cluster level, p < .05, FWE,
apart from C, which reflects p < .001, uncorrected (uncorr.). corr. =
corrected.
Alderson-Day et al.
1645
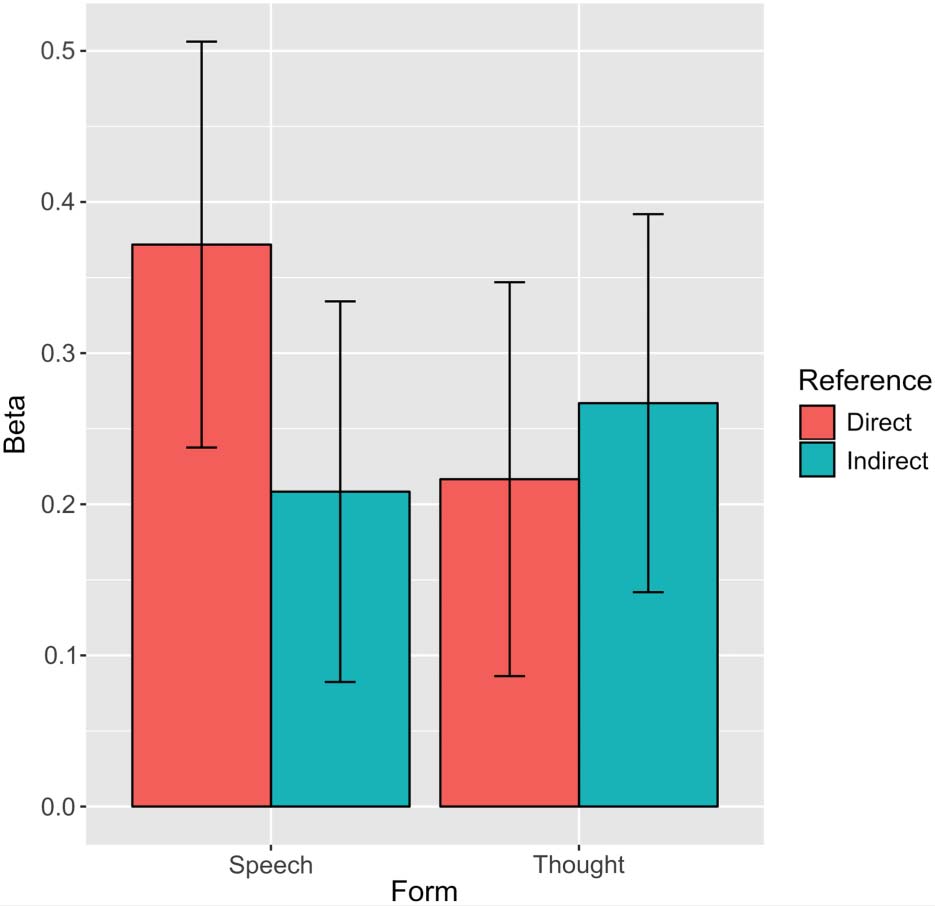
Figure 3. Mean beta values for direct and indirect references to
characters’ speech and thoughts in ToM regions. Error bars represent
SEM.
Table 1, a range of typical regions were identified in the
contrast between communicative inference reasoning
and physical reasoning on the task, including the
mPFC, precuneus, and TPJ bilaterally. Sixteen of the 21
individuals produced ToM networks with significant clus-
ters in at least one of these regions, and their beta values
were taken forward for ROI analysis (15/16, right TPJ;
12/16, left TPJ; 7/16, precuneus; 6/16, mPFC). When the
mean beta values were compared in these areas in a
repeated-measures ANOVA, no main effects of Form,
F(1, 15) = 0.49, p = .493, ηp
2 = .03, or Reference, F(1,
15) = 1.74, p = .207, ηp
2 = .10, were observed, but a
significant interaction was again evident, F(1, 15) =
9.39, p = .008, ηp
2 = .38.3 As Figure 3 shows, this too
was driven by responses for direct speech (compared
with indirect speech), and this was the only significant
difference between the conditions ( p = .016, d = 0.90,
Bonferroni corrected).
We then conducted an exploratory whole-brain analysis
to investigate any further potential differences for direct
versus indirect speech. Significant increases in signal for
direct over indirect speech were evident in three regions:
right TPJ (encompassing right angular gyrus [AG] and
MTG), left inferior frontal gyrus (IFG), and left superior
parietal lobule (see Figure 4). Using the online meta-
analytic tool Neurosynth ( Yarkoni, Poldrack, Nichols,
Van Essen, & Wager, 2011), the most common functional
terms associated with these regions were “network
DMN” for the right AG (posterior probability = 0.73),
“theory mind” for the right MTG ( p = .88), “semantic”
for the left IFG ( p = .88), and “imagery” for the left
SPL ( p = .78). Despite the apparent direct speech effect
Table 4. Whole-Brain Activation Differences for (A) Direct vs. Indirect Speech, (B) Direct Speech vs. Direct Thought, and (C) Direct
vs. Indirect Reference
Location
(A) Direct speech > indirect speech
R angular gyrus
R angular gyrus
R middle temporal gyrus
L inferior frontal gyrus
L inferior frontal gyrus
L inferior frontal gyrus
L superior parietal lobule
L middle occipital gyrus
L middle occipital gyrus
(B) Direct speech > direct thought
R middle cingulate cortex
R ACC
L ACC
Right insula
Right insula
(C) Direct > indirect
L superior parietal lobule
WM
L superior parietal lobule
L middle occipital gyrus
WM
R superior occipital gyrus
WM
R superior occipital gyrus
R middle occipital gyrus
R middle occipital gyrus
WM
R middle temporal gyrus
R middle temporal gyrus
R inferior temporal gyrus
L inferior occipital gyrus
L middle occipital gyrus
L inferior occipital gyrus
X
42
40
56
−50
−52
−44
−24
−28
−26
14
6
−10
34
28
−22
−22
−28
−26
−24
26
20
24
32
34
30
62
70
48
−42
−44
−36
oui
z
−66
−72
−56
30
26
32
−78
−90
−80
26
36
28
18
24
−60
−52
−60
−94
−78
−74
−58
−60
−88
−78
−84
−46
−36
−54
−66
−78
−72
36
42
24
14
22
0
48
12
18
32
28
28
−6
−10
54
44
46
14
16
40
42
54
16
12
6
−8
−8
−10
−8
−2
−4
k
920
865
413
438
231
1303
239
543
323
240
233
t
z
pFWE
5.64
5.54
5.30
5.30
5.23
4.66
4.69
4.67
4.19
5.36
4.92
4.55
5.26
5.20
6.61
6.17
5.85
6.38
4.15
5.95
4.78
4.64
5.56
4.63
4.34
4.96
4.89
4.45
4.73
4.45
4.18
4.31
4.26
4.14
4.14
4.11
3.79
3.81
3.80
3.51
4.17
3.93
3.73
4.12
4.09
4.76
4.57
4.42
4.66
3.48
4.46
3.86
3.78
4.27
3.77
3.60
3.96
3.92
3.67
3.83
3.67
3.50
<.001
<.001
.007
.002
.036
.00
.01
.00
.00
.01
.01
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
9
e
1
-
6
p
3
d
7
f
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
a
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
f
.
/
o
n
0
5
M
a
y
2
0
2
1
All results p < .05, FWE, at cluster level, and p < .001, uncorrected, at peak level. Minimum cluster size: k = 50. WM = white matter.
1646
Journal of Cognitive Neuroscience
Volume 32, Number 9
significance level ( p < .001, uncorrected, k > 50).
De la même manière, no whole-brain differences were observed be-
tween voices and thoughts overall or between indirect
forms of speech and thought, either at corrected or un-
corrected levels.
DISCUSSION
The aim of this study was to explore further the effect of
direct speech in the brains of readers. The main finding
of our results was to replicate the original effect reported
by Yao et al. (2011), namely, that direct speech in short
stories is accompanied by elevated responses in voice-
selective auditory regions of the brain, when compared
with indirect speech. Our findings go further than those
of Yao et al. in two key ways. D'abord, by comparing direct
and indirect references for speech and thoughts, our ROI
results demonstrate a specific effect of reference for char-
acters who are represented as speaking, but not when
they are represented as thinking. Deuxième, this direct
speech effect appears to extend beyond voice-selective
auditory cortex to also include regions that are used
when making inferences about communicative inten-
tion, based on a ToM localizer task (Ciaramidaro et al.,
2007). This pattern of results, donc, supports the ear-
lier observation that readers spontaneously engage sen-
sory cortices when faced with direct speech, but it also
implicates higher-order processes associated with gaug-
ing character intention and meaning.
Evidence of a direct speech effect in auditory cortex is
consistent with previous findings that such regions are
recruited during silent reading of characters’ speech
(Yao et al., 2011, 2012), which is in turn suggestive of au-
ditory verbal imagery being used during this process.
This aligns with behavioral evidence of phonologically
detailed imagery being involved in silent reading of vari-
ous kinds (Kurby & Zacks, 2013; Filik & Barber, 2011).
There is debate around how specific any such voice rep-
resentation would be: Kurby, Magliano, and Rapp (2009)
have argued that such effects are specific to familiar
voices only, whereas Petkov and Belin (2013) propose
that any kind of voice simulation is likely to reflect a ge-
neric speaking voice. Their argument for this is based on
phonological information specific to voice identity usually
being associated with anterior temporal cortex, alors que
those associated with direct speech in Yao et al. (2011),
Par exemple, are more focused on posterior temporal re-
gions (Petkov & Belin, 2013). Our findings cannot easily
arbitrate between these two possibilities (general vs. spe-
cific voices), as voice-selective auditory regions were
identified along the length of the superior temporal gyri
bilaterally. Cependant, we would speculate that any simula-
tion of a generic or specific voice is likely to vary consid-
erably across individuals. When asked, readers describe
drawing upon a wide range of active and creative strate-
gies to imagine the voices of characters, including other
familiar voices and their own voice (Alderson-Day et al.,
2017).
Perhaps more notable is the suggestion of direct
speech effects also being present in cortical regions often
associated with ToM in general and understanding
others’ intentions in particular.4 We chose a localizer
task that aimed to minimize superficial overlaps with
the primary task—using cartoons instead of a written
story format—and focused specifically on assessing un-
derstanding of communicative intentions over other
types of ToM reasoning, such as inferring false beliefs
(Ciaramidaro et al., 2007; Walter et al., 2004). This pro-
duced a network that, in our sample, primarily centered
around bilateral TPJ regions but also included precuneus
and mPFC in subsets of participants. Evidence of a direct
speech effect in these regions provides at least prima
facie support for the idea that text presented in this
way prompts engagement with what a character intends
to say (Yao et al., 2011, 2012), despite the mixed behavioral
evidence that direct reference primes any further commu-
nicative information about characters (Eerland et al., 2013).
De plus, our analysis suggests involvement of these re-
gions at a comparable level to responses in auditory net-
travaux, as indicated by the lack of any interaction effect
across the two ROIs.
Drawing strong conclusions about the role of these
regions in processing direct speech is fraught with dif-
ficulty. The areas highlighted by our ToM task are often
implicated in a range of attentional and cognitive pro-
cesses (Spreng et al., 2009; Mitchell, 2008), and making
broader claims based on the prior literature raises the risk
of reverse inference (Poldrack, 2006). Using Neurosynth
( Yarkoni et al., 2011), which provides at least a systematic
approach to informal reverse inference (Poldrack, 2011),
the strongest responses in the localizer task were in two
regions where the most common associations in the lit-
erature are with “mind tom” and “theory mind” (avec
posterior probabilities of .87–.90). De la même manière, in the ex-
ploratory whole-brain analysis, the right MTG peak in par-
ticular showed high z scores for tests of association (z =
12.00) and uniformity (z = 14.39) in a ToM meta-analytic
map of 181 études ( Yarkoni et al., 2011).
These regions have also been observed in similar work
examining sociocognitive responses to fiction reading by
Tamir, Bricker, Dodell-Feder, and Mitchell (2016), al-
though in their study, they observed preferential engage-
ment of the mPFC for social content in stories (describing
a person’s mental content), with medial temporal cortex
more closely indexing story vividness. In contrast, most
of our participants (15/16) activated the right TPJ on
our localizer task (compared with only six for mPFC),
and this was the only ToM region to be identified in
our whole-brain analysis comparing direct and indirect
speech. The right TPJ cluster that we observed in this
analysis included peaks in the right AG, extending dorsally
and caudally from areas that are often linked to represent-
ing others’ mental states (Bzdok et al., 2013). Both left
Alderson-Day et al.
1647
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
/
t
.
F
o
n
0
5
M.
un
oui
2
0
2
1
and right AG have been associated with support for the
default mode network, via the generation and processing
of transmodal information in the absence of stimulus in-
put (Murphy et al., 2018) and modality-independent con-
tributions to imagery (Daselaar, Porat, Huijbers, &
Pennartz, 2010). The right AG has also recently been im-
plicated in making valence judgments from nonverbal
cues: In a paradigm where participants were asked to
judge the intentions of musical alien “signals,” variations
in the consonance and dissonance of the stimuli (roughly
corresponding to positive and negative emotions) modu-
lated this region specifically (Bravo et al., 2017). Le
broader extension of this cluster, donc, may reflect
the generation and maintenance of intention-related im-
agery, rather than representing characters’ mental states,
or social content more generally. This being associated
with posterior ToM regions over mPFC would also be con-
sistent with van Overwalle’s (2009) distinction between a
posterior ToM subsystem supporting representation of
temporary and perceptually based intentions and goals,
versus an anterior pFC system that tracks and integrates
enduring social information over time.
When taken together, these findings broadly support
the interpretation of direct speech made by Yao et al.
(2011). Recall that, for Yao and colleagues, direct speech
prompts auditory imagery as a means of modeling
speaker prosody (et, finalement, communicative intent).
A counterhypothesis, provided by Eerland, Zwaan, et
colleagues, is that direct reference acts primarily as a
cue to simulate verbatim linguistic content—in other
words, emphasizing the words but, arguably, not the
conférencier (Eerland & Zwaan, 2018; Eerland et al., 2013).
Our data suggest that direct reference has a specific effect
for speech, and this extends to regions that would be
consistent with inferring communicative intentions.
De plus, this can be distinguished from the overall ef-
fect of direct reference, which primarily shows greater
engagement in visual areas of occipital and parietal cortex
(see Table 4C).
A curious characteristic of our data is the apparently
contradictory results for a direct speech effect in auditory
régions, which was evident in the ROI analysis, but not
for the whole-brain contrast. This likely reflects (je) indi-
vidual variability in the temporal voice area (Belin et al.,
2000), (ii) the effect of the more conservative statistical
correction required across the whole brain, et (iii) le
fact that both direct and indirect speech activate a range
of overlapping temporal regions, with any subsequent
difference in beta values being likely to be subtle.
Nevertheless, it should be noted that prominent differ-
ences across the cortex were observed in the right TPJ
(as discussed), left SPL, and left IFG, much more obviously
than for regions of the auditory cortex. The involvement of
the latter in particular is consistent with greater demand
being placed on inner speech production to support the
representation of direct speech, given the common asso-
ciation of Broca’s area with silent articulation (Alderson-
Day & Fernyhough, 2015; Kühn, Fernyhough, Alderson-
Day, & Hurlburt, 2014; Simons et al., 2010; Shergill et al.,
2001). Evidence from psycholinguistics research suggests
that greater involvement of articulatory processes in si-
lent speech results in more detailed acoustic properties
being represented in auditory imagery (Oppenheim &
Dell, 2010), and both external and internal speech have
been shown to consistently modulate auditory cortical re-
sponses (Okada, Matchin, & Hickok, 2018; Ylinen et al.,
2014; Shergill et al., 2002). En outre, two recent studies
of inner speech have highlighted how right-hemisphere
homologs of left-hemisphere language regions are re-
cruited when speech of others must be imagined
(Grandchamp et al., 2019; Alderson-Day et al., 2016). UN
potential model, alors, would be that a reader coming
across direct speech in a text is prompted to generate a
communicatively plausible perceptual simulation, via in-
ner speech, which involves the left IFG and right TPJ
working in concert to modulate voice-selective regions
of the auditory cortex. This is not to suggest that inner
speech (and other auditory imagery processes) would
not be evidenced in each of the task conditions (given
the widespread activation vs. baseline seen for all condi-
tion; see Table 2 and Figure 4A) but rather that direct
speech could place a specific demand on internal articu-
latory processes. In this scenario, direct reference effects
in auditory cortex would plausibly not be the primary
component of the reader’s response but a secondary
consequence of inner speech (and ToM) processes,
which may explain their relative prominence in our
whole-brain results.
Although the present results appear to have much to
say about how speech is treated by readers, they perhaps
say less about what is happening for characters’ thoughts.
Despite having received early theoretical attention in sty-
listics (Sharvit, 2008; Sotirova, 2004), until now, qualita-
tive differences between direct and indirect modes of
speech and thought representation have scarcely been
empirically investigated (for some exceptions using free
indirect discourse, see Fletcher & Monterosso, 2015;
Bray, 2007). A plausible assumption would be that
thought presentation (in direct or indirect reference)
would be more likely to engage ToM resources—that
est, a main effect of thoughts—compared with speech.
Why, alors, was this not seen? Insights from contempo-
rary cognitive narratology may be useful here, particularly
in relation to the problem of “accessibility” of others’
thoughts. Ordinarily, stories that are used to assess
ToM require the reader to make inferences about the
mental states of others; their actual beliefs are not made
explicit and may even conflict with the literal and imme-
diate content of what they say and how they act (par exemple., Saxe
& Kanwisher, 2003). Fictional narratives may sometimes
exploit this “accessibility gap” (par exemple., a suspect in a mystery
could have hidden motives), but they are also notable be-
cause they can give us apparent access to other minds via
direct and indirect references (Bernini, 2016; Cohn,
1648
Journal des neurosciences cognitives
Volume 32, Nombre 9
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
/
.
F
t
o
n
0
5
M.
un
oui
2
0
2
1
1978). Although the stimuli used in our experiment con-
tained mental content, they did not necessarily make de-
mands in terms of mental state inference—in the thought
trials used in our experiment, the inner life of the charac-
ter is laid bare (par exemple., “He thought that he should go to the
shop”). Direct speech, in contrast, does not signal the in-
tonation, emotion, or intention of a character—they
must be simulated or otherwise inferred by the reader,
in a way that the ToM system is often considered to do
(Saxe & Kanwisher, 2003). En tant que tel, although counterin-
tuitive, our findings are in line with common views about
mental state inference (Spreng et al., 2009). It may also
be the case that direct speech in general is more vivid
and salient than direct reference to thinking, given the
whole-brain differences between speaking and thinking
seen in anterior insula and dorsal ACC (Uddin, 2016),
and the quicker orienting times we observed for speech
trials. Engagement with fictional storyworlds and charac-
ters is often argued to depend on the “experiential
traces” the reader brings from his or her own life
(Zwaan, 2008): The more we have access to an experi-
ence in the real world, the more it will be used to gener-
ate vivid and imaginative responses during reading. Quand
one considers the diminished, quasi-perceptual phenom-
enology that verbal thoughts are often claimed to possess
(Prinz, 2011; Jones & Fernyhough, 2007), it is perhaps no
surprise that characters’ thinking in a text did not provide
distinct patterns of activation that were as distinct as for
direct speech.
Another perspective—also provided by cognitive liter-
ary studies—is to consider how fictional minds may be
differently represented from the outside and the inside.
Kuzmičová (2013), Par exemple, has suggested that we
experience characters’ speech in literary texts as either
“outer reverberations” (when we read, as vicarious lis-
teners, about a character overtly speaking) and “inner re-
verberations” (when we voice a character’s words within
his or her perspective). In parallel, Caracciolo (2014) a
highlighted the contrast between attributing intentions
to characters and the direct, inner enactment of a charac-
ter’s thoughts and fictional consciousnesses more broadly.
These distinctions parallel the extensive literature on
perspective-taking and how this is instantiated in the
brain (par exemple., Ruby & Decety, 2001). It could be the case
that our different conditions prompted readers to adopt
d'abord- or third-person perspectives in response to speech
compared with thoughts or direct compared with indirect
reference. Cependant, the direction of these shifts is not
straightforward: Although it is sometimes assumed that
direct speech necessarily prompts adopting a first-person
perspective (speaking as the character), it is also under-
stood as focusing the reader on what it would be like to
hear the character speak to them (Clark & Gerrig, 1990).
De la même manière, thoughts could be seen to prime a first-person
perspective (thinking “from the inside”), but this will likely
depend on the position of the narrator, the reader’s iden-
tification with the character, and the wider context of the
narrative (Kuiken, Miall, & Sikora, 2004). En tant que tel, a key
area for further exploration is to systematically examine
how perspective shifts potentially interact with direct ref-
erence effects and speech/thought distinctions.
This study has a number of limitations. D'abord, it was
necessary to exclude some participants because of partial
data from eye-tracking or either of the independent loca-
lizer tasks, limiting the overall sample size. This also fur-
ther skewed our sex ratio, such that male participants are
underrepresented in our eventual sample (as can often
be the case for psychology studies recruited from univer-
sity populations, par exemple., Dickinson, Adelson, & Owen,
2012). Given the wide variability in individual differences
for reading responses, we chose to deploy these mea-
sures to be as specific as possible about both participants’
onset and offset times of reading target sentences and to
allow for the use of individually specific cortical networks.
This did not prohibit the recruitment of a larger sample
than the original study we sought to replicate ( Yao et al.,
2011), but for a topic (imagery) with typically small ef-
fects and potentially large variation, replication in larger
samples will be required for the exploration of individual
differences in imagery production across different kinds
of readers. Inner speech and imagery are highly suscep-
tible to individual differences in day-to-day use (Alderson-
Day et al., 2016), and effects of expertise (Borst, Niven, &
Logie, 2011) and variation across readers seem highly
likely.
Deuxième, our use of direct reference for thoughts (tel
as he thought “I should have finished this paper by
now”) could be questioned in terms of its relative famil-
iarity for readers. One of our aims for the study was to
use a stimulus that could act as a typographical and gram-
matical control comparison for direct and indirect
speech. Although use of quotation marks for thoughts
does feature in narratives, indirect references might be
thought of as many authors’ default option when refer-
ring to characters’ mental states (Leech & Short, 2007).
An alternative form of reference—such as using italics
to mark characters’ thoughts—may have been more fa-
miliar to readers but would also have added further typo-
graphical differences to the original contrast of interest:
direct versus indirect speech. The lack of any behavioral
differences (in terms of accuracy or reading time) être-
tween the thought conditions, and the lack of any pair-
wise or whole-brain differences, would suggest that this
had little effect on our participants. Cependant, further
careful behavioral (et, arguably, interdisciplinary)
work—incorporating the valuable insights of cognitive lit-
erary studies—is clearly required to elucidate how
readers interpret these kinds of text constructions when
depicting characters’ mental states.
Enfin, a related point about generalizability concerns
the fictional stories used in the experiment. For exper-
imental use, we used very minimal stories that were un-
likely to prompt extensive use of many of the processes
thought to be relevant to a reader’s experience of a
Alderson-Day et al.
1649
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
F
/
.
t
o
n
0
5
M.
un
oui
2
0
2
1
text, whether that involves identification with characters,
use of prior knowledge, management of expectations,
or feelings of transportation (Miall, 2011; Vert, 2004;
Kuiken et al., 2004). En tant que tel, this is still a very artificial
reading scenario for many participants. We cannot rule
out the possibility that there was something about this
situation in particular that may have posed unusual de-
mands or biased readers’ responses, such as encourag-
ing them to pay attention to or engage more with
specific aspects of the text (such as voices in particular).
Our attentional checks would adjudicate against this
interpretation—no significant differences in accuracy
were observed across the various task conditions—but,
in considering ecological validity, the experiential gap be-
tween full stories and these experimental sketches must
be borne in mind.
Notwithstanding these limitations, our findings have
important implications for future research on fiction,
reading, and imagination more generally. Our data broadly
support social cognitive approaches to fiction (Oatley,
2016; Tamir et al., 2016), but in a complex and unexpected
chemin. D'une part, the potential involvement of ToM
in simulating episodes of characters’ speech opens a new
avenue for research on fiction and mentalizing; on the
other hand, our findings for representing characters’
thoughts challenge the idea that engaging with the mental
states of others via fiction necessarily involves (or could
even enhance) ToM processes. Our findings also highlight
how readers likely draw on multiple perceptuomotor re-
sources to support a socially informed simulation of
speech, where prompted by the text. This is, arguably,
a creative and constructive process on the part of the
lecteur, which will be contingent on their own imaginative
skills and experience. Along with comparing individual
differences in this process, contrasting forms of reference
for speech and verbal thoughts offer a comparative meth-
odology for exploring how readers track speaking and
thinking through more complicated narratives. Free indi-
rect discourse, as seen in many modernist texts, demands
that the reader follow closely, or even make their own
inference, about exactly who is speaking or thinking in
a story (see Waugh, 2011, for a discussion of this topic).
Ici, reference or its absence could be considered as an
experimental tool to challenge the readers and place
them in situations of uncertainty about the speech and
thoughts in a narrative (as in Fletcher & Monterosso,
2015). A cet égard, more challenging texts offer an op-
portunity to push at the limits of readers’ creative and
imaginative capacities.
Conclusions
In conclusion, references to direct speech in fictional
stories are associated with the recruitment of not only
voice-selective auditory cortex but also regions that may
implicate gauging of characters’ communicative inten-
tion. De plus, this is a process that is apparently
specific to speech. We cannot conclude on the basis of
these findings that the function of this process is commu-
nicative inference per se, but we speculate that it goes
beyond a purely perceptual simulation of voice and re-
quires coordination between inner speech and ToM re-
sources. To experience a character’s voice in a story, dans
this sense, may be about not only what they say but also
how they say it and what they intend.
Remerciements
This work was supported by the Wellcome Trust ( WT098455
and WT108720). John Foxwell, Lucy May, and Anthony
Atkinson are thanked for their assistance with piloting and
eye-tracking; and David Smailes and the Hearing the Voice team
are thanked for their contributions to the early development of
the research question. For more information on this process,
please see Fernyhough (2015).
Reprint requests should be sent to Ben Alderson-Day,
Département de psychologie, Durham University, Science
Laboratories, South Road, Durham DH1 3LE, ROYAUME-UNI, ou par e-mail:
benjamin.alderson-day@durham.ac.uk.
Remarques
1. Exposure to literary fiction in particular has also been pro-
posed to enhance readers’ ToM skills (Oatley, 2016; Kidd &
Castano, 2013), although such claims have not always been sup-
ported in replication attempts (Kidd & Castano, 2018; Klein
et coll., 2018). For recent meta-analyses on this topic, voir
Dodell-Feder and Tamir (2018) and Mumper and Gerrig (2017).
2. A full list of the stories and questions used is available at
community.dur.ac.uk/benjamin.alderson-day/RVT_full_stim_
aldersonday.pdf.
3. This analysis was also run with ROIs that explicitly excluded
areas identified in the auditory localizer task, leading to almost
identical results: no main effects and a significant interaction,
F(1, 15) = 9.29, p = .008, ηp
2 = .38. On an individual level,
auditory and ToM ROIs overlapped in only 2 de 16 participants,
and this was to a minimal degree.
4. Although Yao et al.’s (2011) main analysis was ROI driven,
they also conducted an exploratory whole-brain analysis that
primarily identified regions of posterior temporal cortex and
occipital-fusiform regions associated with word reading.
RÉFÉRENCES
Alderson-Day, B., Bernini, M., & Fernyhough, C. (2017).
Uncharted features and dynamics of reading: Voices,
characters, and crossing of experiences. Consciousness and
Cognition, 49, 98–109.
Alderson-Day, B., & Fernyhough, C. (2015). Inner speech:
Développement, cognitive functions, phenomenology, et
neurobiology. Psychological Bulletin, 141, 931–965.
Alderson-Day, B., Weis, S., McCarthy-Jones, S., Moseley, P.,
Smailes, D., & Fernyhough, C. (2016). The brain’s
conversation with itself: Neural substrates of dialogic inner
speech. Social Cognitive and Affective Neuroscience, 11,
110–120.
Alexander, J.. D., & Nygaard, L. C. (2008). Reading voices and
hearing text: Talker-specific auditory imagery in reading.
Journal de psychologie expérimentale: Human Perception
et performances, 34, 446–459.
1650
Journal des neurosciences cognitives
Volume 32, Nombre 9
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
F
/
t
.
o
n
0
5
M.
un
oui
2
0
2
1
Barsalou, L. W. (2008). Grounded cognition. Annual Review of
Psychologie, 59, 617–645.
Behne, T., Liszkowski, U., Carpenter, M., & Tomasello, M..
(2012). Twelve-month-olds’ comprehension and production
of pointing. British Journal of Developmental Psychology,
30, 359–375.
Belin, P., Zatorre, R.. J., Lafaille, P., Ahad, P., & Pike, B. (2000).
Voice-selective areas in human auditory cortex. Nature, 403,
309–312.
Bernini, M.. (2016). The opacity of fictional minds:
Transparency, interpretive cognition and the exceptionality
thesis. In P. Garratt (Ed.), The cognitive humanities:
Embodied mind in literature and culture (pp. 35–54).
Londres: Palgrave Macmillan.
Borst, G., Niven, E., & Logie, R.. H. (2011). Visual mental image
generation does not overlap with visual short-term memory:
A dual-task interference study. Mémoire & Cognition, 40,
360–372.
Bortolussi, M., & Dixon, P.. (2003). Psychonarratology:
Foundations for the empirical study of literary response.
Cambridge: la presse de l'Universite de Cambridge.
Bravo, F., Cross, JE., Hawkins, S., González, N., Docampo, J.,
Bruno, C., et autres. (2017). Neural mechanisms underlying
valence inferences to sound: The role of the right angular
gyrus. Neuropsychologie, 102, 144–162.
Bray, J.. (2007). The ‘dual voice’ of free indirect discourse: UN
reading experiment. Language and Literature:
International Journal of Stylistics, 16, 37–52.
Eerland, UN., Engelen, J.. UN., & Zwaan, R.. UN. (2013). The influence
of direct and indirect speech on mental representations. PLoS
Un, 8, e65480.
Eerland, UN., & Zwaan, R.. UN. (2018). The influence of direct and
indirect speech on source memory. Collabra: Psychologie,
4, 5.
Esteve-Gibert, N., & Guellaï, B. (2018). Prosody in the auditory
and visual domains: A developmental perspective. Frontiers
in Psychology, 9, 338.
Fernyhough, C. (2008). Getting Vygotskian about theory of
esprit: Mediation, dialogue, and the development of social
understanding. Developmental Review, 28, 225–262.
Fernyhough, C. (2015). The experimental design hackathon. Dans
C. Fernyhough, UN. Woods, & V. Patton (Éd.), Working
connaissance: Transferable methodology for interdisciplinary
recherche. Hearing the voice. Durham: Durham University.
Filik, R., & Barber, E. (2011). Inner speech during silent reading
reflects the reader’s regional accent. PLoS One, 6, e25782.
Fletcher, P.. C., Happé, F., Frith, U., Boulanger, S. C., Dolan, R.. J.,
Frackowiak, R.. S., et autres. (1995). Other minds in the brain:
A functional imaging study of “theory of mind” in story
comprehension. Cognition, 57, 109–128.
Fletcher, UN., & Monterosso, J.. (2015). The science of free-
indirect discourse: An alternate cognitive effect. Narrative,
24, 82–103.
Gerrig, R.. J.. (1993). Experiencing narrative worlds: On the
psychological activities of reading. New Haven, CT: Yale
Presse universitaire.
Brett, M., Anton, J.-L., Valabregue, R., & Poline, J.-B. (2002).
Gerrig, R.. J., Brennan, S. E., & Ohaeri, J.. Ô. (2001). What
Region of interest analysis using the MarsBar toolbox for SPM
99. Neuroimage, 16, S497.
Brück, C., Kreifelts, B., Gößling-Arnold, C., Wertheimer, J., &
Wildgruber, D. (2014). ‘Inner voices’: The cerebral
representation of emotional voice cues described in literary
texts. Social Cognitive and Affective Neuroscience, 9,
1819–1827.
Bzdok, D., Langner, R., Schilbach, L., Jakobs, O., Roski, C.,
Caspers, S., et autres. (2013). Characterization of the temporo-
parietal junction by combining data-driven parcellation,
complementary connectivity analyses, and functional
decoding. Neuroimage, 81, 381–392.
Caracciolo, M.. (2014). The experientiality of narrative: Un
enactivist approach. Boston: Walter de Gruyter.
Caracciolo, M., & Hurlburt, R.. T. (2016). A passion for
specificity: Confronting inner experience in literature and
science (Forthcoming ed.). Columbus: The Ohio State
Presse universitaire.
characters know: Projected knowledge and projected co-
presence. Journal of Memory and Language, 44, 81–95.
Grandchamp, R., Rapin, L., Perrone-Bertolotti, M., Pichat, C.,
Haldin, C., Cousin, E., et autres. (2019). The ConDialInt model:
Condensation, dialogality, and intentionality dimensions of
inner speech within a hierarchical predictive control
framework. Frontiers in Psychology, 10, 2019.
Vert, M.. C. (2004). Transportation into narrative worlds: Le
role of prior knowledge and perceived realism. Discourse
Processes, 38, 247–266.
Vert, M.. C., Brock, T. C., & Kaufman, G. F. (2004).
Understanding media enjoyment: The role of transportation
into narrative worlds. Communication Theory, 14, 311–327.
Herman, D. (2008). Narrative theory and the intentional stance.
Partial Answers: Journal of Literature and the History of
Ideas, 6, 233–260.
Herman, D. (2013). Storytelling and the sciences of mind.
Cambridge, MA: AVEC Presse.
Ciaramidaro, UN., Adenzato, M., Enrici, JE., Erk, S., Pia, L., Bara, B. G.,
et autres. (2007). The intentional network: How the brain reads
varieties of intentions. Neuropsychologie, 45, 3105–3113.
Jones, S. R., & Fernyhough, C. (2007). Thought as action: Inner
speech, self-monitoring, and auditory verbal hallucinations.
Consciousness and Cognition, 16, 391–399.
Clark, H. H., & Gerrig, R.. J.. (1990). Quotations as
demonstrations. Language, 66, 764–805.
Kidd, D. C., & Castano, E. (2013). Reading literary fiction
improves theory of mind. Science, 342, 377–380.
Cohen, J.. (1969). Statistical power analysis for the behavioural
Kidd, D. C., & Castano, E. (2018). Reading literary fiction and
sciences (2nd ed.). New York: Academic Press.
Cohn, D. (1978). Transparent minds. Princeton, New Jersey: Princeton
Presse universitaire.
Coulmas, F. (2011). Direct and indirect speech. New York:
Walter de Gruyter.
Daselaar, S. M., Porat, Y., Huijbers, W., & Pennartz, C. M.. (2010).
Modality-specific and modality-independent components of
the human imagery system. Neuroimage, 52, 677–685.
Dickinson, E. R., Adelson, J.. L., & Owen, J.. (2012). Gender
équilibre, representativeness, and statistical power in sexuality
research using undergraduate student samples. Archives of
Sexual Behavior, 41, 325–327.
Dodell-Feder, D., & Tamir, D. je. (2018). Fiction reading has a
small positive impact on social cognition: A meta-analysis.
Journal de psychologie expérimentale: General, 147, 1713–1727.
theory of mind: Three preregistered replications and
extensions of Kidd and Castano (2013). Social Psychological
and Personality Science, 10, 522–531.
Klein, R.. UN., Vianello, M., Hasselman, F., Adams, B. G., Adams, R.. B.,
Alper, S., et autres. (2018). Many labs 2: Investigating variation in
replicability across sample and setting. Advances in Methods
and Practices in Psychological Science, 1, 443–490.
Köder, F., Maier, E., & Hendriks, P.. (2015). Perspective shift
increases processing effort of pronouns: A comparison
between direct and indirect speech. Language, Cognition
and Neuroscience, 30, 940–946.
Kühn, S., Fernyhough, C., Alderson-Day, B., & Hurlburt, R.. T.
(2014). Inner experience in the scanner: Can high fidelity
apprehensions of inner experience be integrated with fMRI?
Frontiers in Psychology, 5, 1393.
Alderson-Day et al.
1651
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
F
.
t
/
o
n
0
5
M.
un
oui
2
0
2
1
Kuiken, D., Miall, D. S., & Sikora, S. (2004). Forms of self-
implication in literary reading. Poetics Today, 25, 171–203.
Kukkonen, K. (2014). Presence and prediction: The embodied
reader’s cascades of cognition. Style, 48, 367–384.
Kurby, C. UN., Magliano, J.. P., & Rapp, D. N. (2009). Those voices
in your head: Activation of auditory images during reading.
Cognition, 112, 457–461.
Kurby, C. UN., & Zacks, J.. M.. (2013). The activation of modality-
specific representations during discourse processing. Cerveau
and Language, 126, 338–349.
Kuzmičová, UN. (2013). Outer vs. inner reverberations: Verbal
auditory imagery and meaning-making in literary narrative.
Journal of Literary Theory, 7, 111–134.
Leech, G., & Short, M.. (2007). Style in fiction: A linguistic
introduction to english fictional prose (2nd ed.). New York:
Routledge.
Magrassi, L., Aromataris, G., Cabrini, UN., Annovazzi-Lodi, V., &
Moro, UN. (2015). Sound representation in higher language
areas during language generation. Actes du
Académie nationale des sciences, USA., 112, 1868–1873.
Mar, R.. UN., & Oatley, K. (2008). The function of fiction is the
abstraction and simulation of social experience. Perspectives
on Psychological Science, 3, 173–192.
Maslej, M.. M., Oatley, K., & Mar, R.. UN. (2017). Creating fictional
characters: The role of experience, personality, and social
processes. Psychology of Aesthetics, Creativity, and the Arts,
11, 487–499.
Miall, D. S. (2011). Emotions and the structuring of narrative
réponses. Poetics Today, 32, 323–348.
Mitchell, J.. P.. (2008). Activity in right temporo-parietal junction
is not selective for theory-of-mind. Cortex cérébral, 18,
262–271.
Molenberghs, P., Johnson, H., Henry, J.. D., & Mattingley, J.. B.
(2016). Understanding the minds of others: A neuroimaging
meta-analysis. Neurosciences & Biobehavioral Reviews, 65,
276–291.
Mumper, M.. L., & Gerrig, R.. J.. (2017). Leisure reading and social
cognition: A meta-analysis. Psychology of Aesthetics,
Creativity, and the Arts, 11, 109–120.
Murphy, C., Jefferies, E., Rueschemeyer, S.-A., Sormaz, M.,
Wang, H., Margulies, D. S., et autres. (2018). Distant from input:
Evidence of regions within the default mode network
supporting perceptually-decoupled and conceptually-guided
cognition. Neuroimage, 171, 393–401.
Oatley, K. (2011). Such stuff as dreams. Malden, MA:
Wiley-Blackwell.
Oatley, K. (2016). Fiction: Simulation of social worlds. Trends in
Cognitive Sciences, 20, 618–628.
Poldrack, R.. UN. (2006). Can cognitive processes be inferred from
neuroimaging data? Tendances des sciences cognitives, 10, 59–63.
Poldrack, R.. UN. (2011). Inferring mental states from
neuroimaging data: From reverse inference to large-scale
decoding. Neurone, 72, 692–697.
Polvinen, M.. (2016). Enactive perception and fictional worlds. Dans
P.. Garratt (Ed.), The cognitive humanities (pp. 19–34). Londres:
Palgrave Macmillan.
Prinz, J.. (2011). The sensory basis of cognitive phenomenology.
In T. Bayne & M.. Montague (Éd.), Cognitive phenomenology
(pp. 174–196). Oxford: Presse universitaire d'Oxford.
Richardson, J.. T. (2011). Eta squared and partial eta squared as
measures of effect size in educational research. Educational
Research Review, 6, 135–147.
Ruby, P., & Decety, J.. (2001). Effect of subjective perspective
taking during simulation of action: A PET investigation of
agency. Neurosciences naturelles, 4, 546–550.
Ryan, M.-L. (1999). Immersion vs. interactivity: Virtual reality
and literary theory. SubStance, 28, 110–137.
Saxe, R., & Kanwisher, N. (2003). People thinking about
thinking people. The role of the temporo-parietal junction in
“theory of mind”. Neuroimage, 19, 1835–1842.
Saxe, R., & Powell, L. J.. (2006). It’s the thought that counts:
Specific brain regions for one component of theory of mind.
Sciences psychologiques, 17, 692–699.
Schurz, M., Radua, J., Aichhorn, M., Richlan, F., & Perner, J..
(2014). Fractionating theory of mind: A meta-analysis of
functional brain imaging studies. Neurosciences &
Biobehavioral Reviews, 42, 9–34.
Schurz, M., Tholen, M.. G., Perner, J., Mars, R.. B., & Sallet, J..
(2017). Specifying the brain anatomy underlying temporo-
parietal junction activations for theory of mind: A review
using probabilistic atlases from different imaging modalities.
Human Brain Mapping, 38, 4788–4805.
Sharvit, Oui. (2008). The puzzle of free indirect discourse.
Linguistics and Philosophy, 31, 353–395.
Shergill, S. S., Brammer, M.. J., Fukuda, R., Bullmore, E., Amaro,
E., Jr., Murray, R.. M., et autres. (2002). Modulation of activity in
temporal cortex during generation of inner speech. Human
Brain Mapping, 16, 219–227.
Shergill, S. S., Bullmore, E. T., Brammer, M.. J., Williams, S. C.,
Murray, R.. M., & McGuire, P.. K. (2001). A functional study
of auditory verbal imagery. Psychological Medicine, 31,
241–253.
Simons, C. J., Tracy, D. K., Sanghera, K. K., O’Daly, O., Gilleen,
J., Dominguez, M.. G., et autres. (2010). Functional magnetic
resonance imaging of inner speech in schizophrenia.
Biological Psychiatry, 67, 232–237.
Okada, K., Matchin, W., & Hickok, G. (2018). Neural evidence
Sotirova, V. (2004). Connectives in free indirect style:
for predictive coding in auditory cortex during speech
production. Psychonomic Bulletin & Review, 25, 423–430.
Oppenheim, G. M., & Dell, G. S. (2010). Motor movement
matters: The flexible abstractness of inner speech. Mémoire &
Cognition, 38, 1147–1160.
Palmer, UN. (2004). Fictional minds. Lincoln, NE: Université de
Nebraska Press.
Perrone-Bertolotti, M., Kujala, J., Vidal, J.. R., Hamame, C. M.,
Ossandon, T., Bertrand, O., et autres. (2012). How silent is silent
reading? Intracerebral evidence for top–down activation of
temporal voice areas during reading. Journal de
Neurosciences, 32, 17554–17562.
Perrone-Bertolotti, M., Rapin, L., Lachaux, J.. P., Baciu, M., &
Lœvenbruck, H. (2014). What is that little voice inside my
head? Inner speech phenomenology, its role in cognitive
performance, and its relation to self-monitoring. Behavioural
Brain Research, 261, 220–239.
Petkov, C. JE., & Belin, P.. (2013). Silent reading: Does the brain
‘hear’ both speech and voices? Biologie actuelle, 23, R155–R156.
Continuity or shift? Language and Literature: International
Journal of Stylistics, 13, 216–234.
Spreng, R.. N., Mar, R.. UN., & Kim, UN. S. (2009). The common
neural basis of autobiographical memory, prospection,
navigation, theory of mind, and the default mode: UN
quantitative meta-analysis. Journal of Cognitive
Neurosciences, 21, 489–510.
Tamir, D. JE., Bricker, UN. B., Dodell-Feder, D., & Mitchell, J.. P..
(2016). Reading fiction and reading minds: The role of
simulation in the default network. Social Cognitive and
Affective Neuroscience, 11, 215–224.
Tomasello, M., Carpenter, M., Call, J., Behne, T., & Moll, H.
(2005). Understanding and sharing intentions: The origins
of cultural cognition. Behavioral and Brain Sciences, 28,
675–691.
Uddin, L. Q. (2016). Salience network of the human brain. San
Diego, Californie: Academic Press.
van Dijk, T. UN., & Kintsch, W. (1983). Strategies of discourse
comprehension. San Diego, Californie: Academic Press.
1652
Journal des neurosciences cognitives
Volume 32, Nombre 9
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
t
F
/
.
o
n
0
5
M.
un
oui
2
0
2
1
van Overwalle, F. (2009). Social cognition and the brain: A meta-
analyse. Human Brain Mapping, 30, 829–858.
Vilhauer, R.. P.. (2016). Inner reading voices: An overlooked form
of inner speech. Psychosis: Psychological, Social and
Integrative Approaches, 8, 37–47.
Walter, H., Adenzato, M., Ciaramidaro, UN., Enrici, JE., Pia, L., &
Bara, B. (2004). Understanding intentions in social
interaction: The role of the anterior paracingulate cortex.
Journal des neurosciences cognitives, 16, 1854–1863.
Waugh, P.. (2011). Thinking in Literature: Modernism and
contemporary neuroscience. In D. James (Ed.), The legacies
of modernism: Historicising postwar and contemporary
fiction (pp. 75–95). Cambridge: la presse de l'Universite de Cambridge.
Wellman, H. M., Cross, D., & Watson, J.. (2001). Meta-analysis of
theory-of-mind development: The truth about false belief.
Child Development, 72, 655–684.
Woodward, UN. L. (1998). Infants selectively encode the goal
object of an actor’s reach. Cognition, 69, 1–34.
Yao, B., Belin, P., & Scheepers, C. (2011). Silent reading of
direct versus indirect speech activates voice-selective areas in
the auditory cortex. Journal des neurosciences cognitives, 23,
3146–3152.
Yao, B., Belin, P., & Scheepers, C. (2012). Brain “talks over”
boring quotes: Top–down activation of voice-selective areas
while listening to monotonous direct speech quotations.
Neuroimage, 60, 1832–1842.
Yarkoni, T., Poldrack, R.. UN., Nichols, T. E., Van Essen, D. C., &
Wager, T. D. (2011). Large-scale automated synthesis of human
functional neuroimaging data. Nature Methods, 8, 665–670.
Ylinen, S., Nora, UN., Leminen, UN., Hakala, T., Huotilainen, M.,
Shtyrov, Y., et autres. (2014). Two distinct auditory-motor circuits
for monitoring speech production as revealed by content-
specific suppression of auditory cortex. Cortex cérébral, 25,
1576–1586.
Zunshine, L. (2006). Why we read fiction: Theory of mind and
the novel. Columbus, OH: Ohio State University Press.
Zwaan, R.. UN. (2008). Experiential traces and mental simulations
in language comprehension. En M. de Vega, UN. Glenberg, & UN.
Graesser (Éd.), Symbols and embodiment: Debates on
meaning and cognition (pp. 165–180). Oxford: Oxford
Presse universitaire.
Zwaan, R.. UN., Madden, C. J., Yaxley, R.. H., & Aveyard, M.. E.
(2004). Moving words: Dynamic representations in language
comprehension. Sciences cognitives, 28, 611–619.
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
9
e
1
–
6
p
3
d
7
F
2
/
0
3
1
2
3
/
8
9
2
/
9
1
6
o
3
c
7
n
_
/
un
1
_
8
0
6
1
2
5
0
7
1
1
7
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
7
0
1
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
/
F
.
t
o
n
0
5
M.
un
oui
2
0
2
1
Alderson-Day et al.
1653