Processing Nouns and Verbs in the Left Frontal
Cortex: A Transcranial Magnetic Stimulation Study
Marinella Cappelletti1,2*, Felipe Fregni1, Kevin Shapiro3*,
Alvaro Pascual-Leone1, and Alfonso Caramazza3
Abstrait
& Neuropsychological and neurophysiological studies suggest
that the production of verbs in speech depends on cortical
regions in the left frontal lobe. Cependant, the precise topography
of these regions, and their functional roles in verb production,
remains matters of debate. In an earlier study with repetitive
transcranial magnetic stimulation (SMTr), we showed that stim-
ulation to the left anterior midfrontal gyrus disrupted verb
production, but not noun production, in a task that required
subjects to perform simple morphological alternations. Ce
result raises a number of questions: Par exemple, is the effect
of stimulation focal and specific to that brain region? Is the
behavioral effect limited to rule-based, regular transformations,
or can it be generalized over the grammatical category? In the
present study, we used rTMS to suppress the excitability of
distinct parts of the left prefrontal cortex to assess their role in
producing regular and irregular verbs compared to nouns. Nous
compared rTMS to sham stimulation and to stimulation of
homologous areas in the right hemisphere. Response latencies
increased for verbs, but were unaffected for nouns, following
stimulation to the left anterior midfrontal gyrus. No significant
interference specific for verbs resulted after stimulation to two
other areas in the left frontal lobe, the posterior midfrontal
gyrus and Broca’s area. These results therefore reinforce the
pour
idea that the left anterior midfrontal cortex is critical
processing verbs. De plus, none of the regions stimulated was
preferentially engaged in the production of regular or irregular
inflection, raising questions about the role of the frontal lobes in
processing inflectional morphology. &
INTRODUCTION
Evidence from a variety of sources suggests that the
language processing system includes at least some com-
ponents that discriminate between nouns and verbs at
level. Par exemple, electrophysiological
the cortical
studies have described different spatial and/or temporal
patterns of event-related potentials (ERPs) evoked by
noun and verb stimuli (par exemple., Kellenbach, Wijers, Hovius,
Mulder, & Mulder, 2002; Dehaene, 1995; Brun, Marais, &
Forgeron, 1973; Teyler, Roemer, Harrison, & Thompson,
1973). Some functional neuroimaging studies have also
found cortical areas that respond more robustly to verbs
than to nouns (Longe, Randall, Stamatakis, & Tyler,
2007; Shapiro, Moo, & Caramazza, 2006; Shapiro et al.,
2005; Tyler, Bright, Fletcher, & Stamatakis, 2004; Perani
et coll., 1999) or vice-versa (Shapiro et al., 2005, 2006).
Arguably, the most compelling source of evidence for
the noun–verb dissociation, cependant, is the study of pa-
tients with focal ischemic lesions or neurodegenerative
diseases who show either poorer performance in re-
trieving verbs than nouns, or the opposite pattern (par exemple.,
1Beth Israel Deaconess Medical Center and Harvard Medical
École, 2University College London, Londres, ROYAUME-UNI, 3Harvard
University
*M.. C. and K. S. contributed equally to this project.
Bates, Chen, Tzeng, Li, & Opie, 1991; Zingeser &
Berndt, 1988, 1990; Miceli, Silveri, Villa, & Caramazza,
1984; see Shapiro & Caramazza, 2003c; Druks, 2002, pour
reviews).1
En général, verb deficits occur in patients with damage
to the left frontal cortex, whereas noun deficits are as-
sociated with damage to the left temporal lobe (Tranel,
Adolphs, Damasio, & Damasio, 2001; Daniele, Giustolisi,
Silveri, Colosimo, & Gainotti, 1994; Damasio & Tranel, 1993;
Miceli et al., 1984). Néanmoins, a number of patients
not consistent with this pattern have been reported
(Shapiro, Shelton, & Caramazza, 2000; Silveri & di Betta,
1997; De Renzi & di Pellegrino, 1995; Hillis & Caramazza,
1995), suggesting that several different patterns of
brain damage can contribute to selective impairments
in producing words of a given category. It may be that
these various patterns are associated with functional im-
pairments at different stages of word production, tous
of which lead to a behavioral dissociation in producing
nouns and verbs.
Careful neuropsychological studies offer some sup-
port for this postulate. Case series by Crepaldi et al.
(2006) and Berndt, Haendiges, and Burton (2002) have
shown that dissociations in noun and verb production
disappear for some patients, but not for others, quand
the imageability of target words is controlled across cat-
egories. Some single-case studies have reported patients
D 2008 Massachusetts Institute of Technology
Journal des neurosciences cognitives 20:4, pp. 707–720
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
0
je
4
e
–
7
p
0
d
7
F
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
oui
0
g
0
toi
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
e
0
2
s
3
t
/
j
.
t
F
.
/
.
o
n
1
8
M.
un
oui
2
0
2
1
whose deficits in noun and verb retrieval seem to reflect
problems in accessing word meaning, while grammatical
aspects of verb production remain intact (par exemple., Laiacona
& Caramazza, 2004; Shapiro & Caramazza, 2003un). Dans
other cases, category-selective deficits have been linked
to specific impairments in morphological processing
(Laiacona & Caramazza, 2004; Shapiro & Caramazza,
2003b; Tsapkini, Jarema, & Kehayia, 2002; Shapiro
et coll., 2000). These patients have particular difficulty
producing nouns or verbs in the context of sentences
et, in some cases, are even impaired using pseudo-
words as nouns (par exemple., ‘‘many wugs’’) or as verbs (par exemple.,
‘‘he wugs’’).
Malheureusement, the patients that have been described
have heterogeneous and sometimes diffuse patterns of
brain injury, rendering problematic the association of
particular functional deficits with precisely defined
cortical areas. Functional neuroimaging studies are ob-
viously better suited to answering questions about ce-
rebral
localisation; nevertheless, even well-designed
imaging studies present difficulties in interpretation,
the most conspicuous of which is that they cannot es-
tablish a true causal relationship between areas of ob-
served brain activation and the corresponding behavioral
task. Transcranial magnetic stimulation is a valuable
technique in this regard, complementing neuropsychol-
ogy and neuroimaging in that it allows researchers to
study the effects of modulating cortical function in a non-
invasive fashion (Walsh & Pascual-Leone, 2003).
In an earlier study, we used repetitive transcranial
stimulation magnétique (SMTr) in an attempt to determine
whether a particular brain region was critical for pro-
cessing verbs. Spécifiquement, we targeted the anterior
portion of the left middle frontal gyrus while subjects
performed a task requiring the production of verbs or
nouns in the context of short phrases (Shapiro, Pascual-
Leone, Mottaghy, Gangitano, & Caramazza, 2001). Le
results showed that stimulation to this area induced a
delay in producing verbs relative to nouns.
The task in the study by Shapiro et al. (2001) required
subjects to manipulate the inflectional form of a stimu-
lus word. Par exemple, one trial presented the word
doors, followed by a symbolic cue indicating the singular
formulaire; subjects were then required to say ‘‘door.’’ Inter-
estingly, the same dissociation between nouns and verbs
was found when the stimuli were meaningless pseudo-
mots, like wug and narfs. These data were taken as
evidence that the targeted portion of the left prefrontal
cortex is involved in processing grammatical properties
of verbs, rather than properties related to verb meaning.
Several objections to this conclusion are immediately
apparent. Perhaps the most important (and most con-
tentious) of these relates to the problem of differenti-
ating semantic from grammatical processing; we will
take up this issue in the General Discussion. Aside from
this theoretical challenge, there are a number of meth-
odological concerns that confound the interpretation of
the experiments described in Shapiro et al. (2001). Un
is that rTMS was applied only to one area within the left
frontal lobe; as a consequence, the study does not rule
out the involvement of other parts of the left frontal
cortex (or other parts of the brain generally) in verb
traitement. It is also possible that the targeted area itself
was not crucial for verb processing, and that the ob-
served effects were really attributable to the transsynap-
tic modulation of neighboring or distant cortical sites.
De plus, even if the targeted area was uniquely
crucial for performing the verb task used by Shapiro
et autres. (2001), the interpretation of this result would be
constrained by the fact that the study investigated only
the processing of regularly inflected nouns and verbs in
involving number
specific morphological alternations,
(for nouns) and agreement (for verbs). It could be
argued that the effect of rTMS on verb production in
Shapiro et coll.. resulted from the different computational
demands of these two operations—in particular, le
greater complexity of subject–verb agreement.2
A related question concerns whether the area that was
stimulated is important for the production of morpho-
logically irregular verb forms, like ran and bought, lequel
cannot be computed by adding or deleting an inflectional
affix. Some authors have proposed that regular and
irregular morphological transformations are subserved
by distinct neural circuits (Tyler et al., 2004; Miozzo, 2003;
Tyler, Russell, Fadili, & Moss, 2001; Marslen-Wilson &
Tyler, 1997; Ullman, Corkin, et coll., 1997), although this
‘‘dual-route’’ assumption has proven famously contro-
versial (voir, par exemple., McClelland & Patterson, 2002; Oiseau,
Howard, & Franklin, 2000; Joanisse & Seidenberg, 1999).
One version of the dual-route hypothesis holds that
prefrontal brain regions are particularly important for
rule-based or regular morphology (Ullman, Corkin, et coll.,
1997)—although neuroimaging studies have shown, si
anything, the opposite association (Indefrey et al., 1997;
Ullman, Bergida, & O’Craven, 1997). Consistent with the
latter results, two neuropsychological studies have de-
scribed patients with left prefrontal lesions who show an
advantage for regular over irregular verbs, albeit in the
context of a general deficit for verbs relative to nouns
(Shapiro & Caramazza, 2003b,
in an English-speaking
patient; Balaguer, Costa, Sebastian-Galles, Juncadella, &
Caramazza, 2004,
in two Spanish–Catalan bilinguals).
These findings do not necessarily call into question the
differential role of the left prefrontal cortex in noun and
verb processing, but they do raise the possibility that
noun or verb effects may be modulated by an orthog-
onal dimension of regularity.
Current Study
Here we report two new rTMS experiments that were
designed to investigate the functional roles of different
regions of the left frontal cortex in processing regular
and irregular verbs and nouns. Aside from replicating
708
Journal des neurosciences cognitives
Volume 20, Nombre 4
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
0
je
4
e
–
7
p
0
d
7
F
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
oui
0
g
0
toi
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
e
0
2
s
3
t
/
j
.
.
t
.
/
F
o
n
1
8
M.
un
oui
2
0
2
1
the effects reported in Shapiro et al. (2001), the experi-
ments address at least two other important questions:
d'abord, whether any of the targeted areas in the prefrontal
cortex is important in distinguishing regular from irreg-
ular morphological computations; et deuxieme, si
any role is played by other prefrontal cortical areas in
grammatical processing of nouns and verbs.
The primary goal of the first experiment was to deter-
mine whether the area targeted in Shapiro et al. (2001)
is important only for rule-based morphological trans-
formations, or alternatively, whether it is involved in any
computational process involving verbs, regardless of
morphological type. In order to compare morphologi-
cally regular and irregular word forms, we modified the
behavioral task, retaining the singular–plural alternation
for nouns but using the present–past tense alternation
for verbs. This change had the additional advantage of
making the noun and verb tasks more comparable in
terms of syntactic complexity because past tense verbs do
not require the computation of an agreement relation
with the external object of the verb phrase. As in our
earlier study, rTMS was applied to a portion of the ante-
rior midfrontal gyrus (aMFG) just anterior and superior
to Broca’s area. We expected that this would again result
in a delay in response latencies for verb trials (relative to
a baseline), but no comparable delay for noun trials.
In the second experiment, we tried to determine
whether the noun–verb difference obtained with stimu-
lation to the left aMFG is specific to that region. To this
end, we repeated the behavioral task in Experiment 1
with rTMS applied to two other regions in the frontal
lobe: Broca’s area (inferior frontal gyrus, or IFG) and a
more posterior portion of the midfrontal gyrus (pMFG).
These sites were approximately equidistant from each
other and from the aMFG with respect to the surface of
le cerveau. If rTMS applied at these sites does not result
in a verb–noun dissociation, this can be taken as evi-
dence for the focality (if not the locality) of any effect of
stimulation to the aMFG. To put it somewhat differently,
this finding would support the conclusion that the
aMFG—or some network to which the aMFG has a
privileged connection (see General Discussion), is cru-
cial for verb processing, but not for noun processing.
The IFG and the pMFG also serve as useful control
areas from a functional perspective, as both have been
implicated in verb production by some authors. A few
neuroimaging studies have shown that verb processing
induces greater activation in the left IFG than does noun
traitement (Tyler et al., 2004; Perani et al., 1999); dans
some studies this effect is specific to the processing of
inflected words (Longe et al., 2007; Tyler et al., 2004).
This dissociation may reflect the greater grammatical or
morphological complexity of inflected verbs, compared
to nouns.
D'autre part, it has been postulated that pre-
motor brain regions are crucial for representing seman-
tic features of verbs, by virtue of their role in storing
action schemata (Pulvermu¨ller, 1999, 2001, 2005). Quelques
en effet, shown activation
neuroimaging studies have,
in premotor regions during tasks involving the process-
ing of verbs referring to imageable actions (Buccino
et coll., 2005; Tettamanti et al., 2005; Hauk, Johnsrude,
& Pulvermu¨ller, 2004). Fait intéressant, an fMRI study using
a morphological alternation task identical to the one used
here also showed increased activation for verbs com-
pared to nouns in the left pMFG, regardless of whether
the verbs were concrete or abstract (Shapiro et al., 2006).
Ainsi, there are reasons to suspect that both the IFG and
the pMFG might be more involved in verb production
than in noun production, which would make the finding
of an effect restricted to the aMFG theoretically more
striking.
The current study incorporates several methodolog-
ical improvements over the design used in Shapiro et al.
(2001). In the previous study noun and verb trials were
blocked, amplifying potential effects of practice, alors que
in the current study, trials with words of each category
were interleaved. Deuxième, we replaced the iconic cues
used by Shapiro et al. with more natural phrasal cues.
Enfin, in addition to sham TMS, we used real rTMS over
homologous right hemisphere sites as a control for left-
sided stimulation. In the previous experiments, we used
only sham stimulation.
MÉTHODES
Participants
Eight right-handed native speakers of English (4 men)
âgé 19 à 33 années (mean = 22.9 années) participated in
Experiment 1. Twelve right-handed native speakers of
English (5 men) âgé 18 à 36 années (mean = 21.1 années)
participated in Experiment 2. All subjects were healthy,
with no history of neurological or psychiatric illness,
and no contraindications to TMS as determined by the
TMS safety screening questionnaire (Keel, Forgeron, &
Wassermann, 2001). They were all screened for TMS
exclusion criteria and gave their written informed con-
sent before participating. This study was performed in
close adherence to TMS safety guidelines (Wasserman,
1998) and was approved by institutional review boards at
Harvard University and at Beth Israel Deaconess Medical
Centre. In Experiment 2, the same subjects were studied
on two different days to minimize carryover effects.
Tâche comportementale
Word production was cued by a simple completion task
consisting of visually presented noun and verb phrases.
Subjects were required to produce the singular and
plural forms of regular and irregular nouns (par exemple., song/
songs or child/children), and the present or past tense
forms of regular and irregular verbs (par exemple., walk/walked
or sleep/slept). In verb trials, subjects saw stimulus
Cappelletti et al.
709
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
0
je
4
e
–
7
p
0
d
7
F
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
oui
0
g
0
toi
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
e
0
2
s
3
t
/
j
.
/
F
.
t
.
o
n
1
8
M.
un
oui
2
0
2
1
phrases of the form ‘‘today I walk,’’ followed by cue
phrases like ‘‘yesterday I. . .’’ (Dans ce cas, the subject
was instructed to say walked.). Noun trials consisted of
stimulus phrases such as ‘‘one child,’’ followed by the
cue ‘‘many. . .’’ (enfants).
The task was performed on a Dell PC using the DMDX
package (Forster & Forster, 2003). Participants’ viewing
distance from the computer monitor was about 0.5 m.
Response latencies were recorded with a custom-built
microphone connected to the computer through a pre-
amplifier. Each trial began with a fixation cross, lequel
appeared in the center of the screen for 500 msec. Ce
was followed by a written stimulus phrase (par exemple., ‘‘today I
walk’’), which appeared in the position of the fixation
cross for 500 msec, followed for another 500 msec by the
cue phrase (‘‘yesterday I. . .’’), indicating the morpho-
logical form in which the word in the stimulus phrase
was to be produced aloud. Participants were instructed
to supply appropriate single word responses immedi-
ately upon seeing the cue phrase. The next trial began
1500 msec after the offset of the cue phrase so that
subjects had a maximum of 2000 msec to respond in
each trial.
Stimulus Preparation
Eighty words were used in the experiment, y compris 20
words of each of the following types: regular verbs,
irregular verbs, regular nouns, and irregular nouns. Nous
excluded verbs that can function as auxiliaries (faire,
have), verbs with syncretic past tense forms (pleaded/
pled, dreamed/dreamt), and nouns with imported plu-
ral forms (datum, octopus). The four sets of words were
matched as closely as possible for length in phonemes
(average of inflected and unmarked forms),
lemma
frequency, and surface frequency of the relevant in-
flected forms (Kucera & Francis, 1982) (see Table 1).
Bidirectional Student’s t tests showed no significant
differences on these measures between nouns and verbs
within each morphological type, or between regular and
irregular words within each grammatical category (a =
0.05, uncorrected for multiple comparisons).
Ratings of concreteness, imageability and familiarity
on Likert scales of 1–7 were obtained from a group of
10 subjects at Harvard University, none of whom partic-
ipated in the rTMS experiment (Shapiro et al., 2006).
Confirmatory bidirectional t tests revealed no differen-
ces in imageability or familiarity within morphological
types or grammatical categories. For both regular words
[t(38) = 5.86, p < .001] and irregular words [t(38) =
5.82, p < .001], nouns were rated as significantly more
concrete than verbs (although there was no difference
between regular and irregular words of each grammat-
ical category). The dissociation between imageability
and concreteness in these stimuli is probably attribut-
able to the fact that ‘‘concreteness’’
for verbs was
defined as the extent to which they referred to actions.
Some of the verbs in the stimulus set, such as to sleep
and to bleed, may be considered highly imageable, but
poor examples of actions.
The words were subsequently used to create eight
lists of 160 trials each. In each list, every stimulus word
appeared twice: once in a trial with a marked-form target
(plural noun or past tense verb) and once in a trial with
an unmarked-form target (singular noun or present
tense verb). One of these trials required a morphological
change relative to the cue phrase, whereas the other did
not. Moreover, the lists were balanced so that each cue/
stimulus pair appeared in exactly two lists. The order of
trials in each list was pseudorandomized.
Experimental Sessions
Experiment 1 was completed in a single session and
included four blocks of trials, corresponding to four
stimulation conditions (right hemisphere rTMS,
left
hemisphere rTMS, right hemisphere sham, and left
hemisphere sham), as well as an initial practice block.
In Experiment 2, subjects returned for two sessions,
separated by at least 48 hr. In each session of the
experiment, subjects completed a practice block and
three blocks of trials (left hemisphere rTMS, right hemi-
sphere rTMS, and either right or left hemisphere sham).
Each block corresponded to one of the trial lists. Lists
Table 1. Psycholinguistic Characteristics of Words Included in the Experiment
Morphology
Category
Fl
FSZ
FSI
Length Concreteness
Imageability Familiarity
Regular (n = 40) Noun (n = 20)
93.1 (105.5) 53.1 (62.4)
30.1 (37.1) 3.6 (0.8)
6.1 (1.0)*
6.0 (1.3)
6.2 (0.6)
Verb (n = 20)
115.2 (91.3)
35.4 (34.6)
34.5 (33.5) 3.5 (0.6)
4.5 (0.7)
5.3 (0.9)
6.3 (0.4)
Irregular (n = 40) Noun (n = 20) 122.1 (218.8) 68.9 (157.0) 44.5 (95.8) 3.7 (0.8)
6.1 (0.8)*
5.9 (1.4)
6.1 (0.7)
Verb (n = 20)
120.1 (86.9)
32.0 (28.4)
32.2 (34.2) 3.9 (0.6)
4.8 (0.6)
5.6 (0.8)
6.3 (0.3)
Fl = cumulative (lemma) frequency; FSZ = surface frequency of unmarked form; FSI = surface frequency of inflected form. Standard deviations are
given in parentheses. Morphology refers to the marked form (plural for nouns, past tense for verbs). Frequency values are from Kucera and Francis
(1982). Length refers to the mean length in phonemes of the inflected and unmarked forms, with diphthongs counted as two phonemes. Values for
concreteness, imageability, and familiarity are mean ratings on Likert scales of 1–7 (see text).
*p < .001.
710
Journal of Cognitive Neuroscience
Volume 20, Number 4
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
f
.
.
t
/
o
n
1
8
M
a
y
2
0
2
1
were assigned to blocks by subject in a Latin square fash-
ion. In total, there were 640 trials in Experiment 1 and
960 trials in Experiment 2, excluding the practice trials.
Data collected during the practice blocks were discarded.
The experimental blocks were completed immediately
after each of the stimulation conditions in each session
(right hemisphere rTMS,
left hemisphere rTMS, and
sham). A 30-min rest period followed the first non-
practice block to allow any effect of rTMS to wash out.
In both experiments, the order of the areas stimulated
with rTMS was counterbalanced across subjects to avoid
carryover effects. Similarly, the order of real rTMS and
sham stimulation was counterbalanced across subjects.
In Experiment 2, the two areas stimulated with rTMS in
each session were never located in the same hemisphere
and were never homologous.
Areas Targeted with rTMS
Prior to the TMS experiments, each subject underwent a
high-resolution structural MRI scan. Three-dimensional
cortical surface maps were reconstructed from the MR
images using the Brainsight neuronavigation system
(Rogue Research, Montreal, Canada), and target sites
for stimulation were marked on these maps.
In Experiment 1 we targeted the inferior portion of
the left aMFG and its right hemisphere homologue. To
localize this area in each hemisphere, we initially iden-
tified the middle frontal gyrus in the 3-D anatomical
brain reconstruction, and then marked the anterior part
of the gyrus near the intersection of the inferior and
superior frontal sulci and above the frontal pole. Sham
stimulation was applied at the same sites as real rTMS.
In Experiment 2, three different stimulation sites were
identified in each hemisphere. The first site of interest
was the superior posterior third of the midfrontal gyrus
(pMFG), near the precentral and superior frontal sulci
and above the opercular part of the IFG. In functional
terms, this corresponds to the premotor cortex. The
second site was within the triangular part of the IFG,
corresponding to the anterior portion of Broca’s area
in the left hemisphere. We targeted an area near the
anterior ascending rami, roughly at the midpoint be-
tween the lateral sulcus (Sylvian fissure) and the inferior
frontal sulcus. A third, intermediate site was marked in
each hemisphere for sham stimulation.
Stereotactic Guidance and Application of rTMS
The application of rTMS during the experiments was
guided with frameless stereotaxy using the Brainsight
system and a Polaris infrared camera. Subjects wore
tightly fitting elastic headbands with infrared-reflective
trackers. At the beginning of each experimental session,
surface anatomical landmarks were registered to the re-
constructed MR images using a pointing device. A reflec-
tive tracker was also affixed to the TMS coil so that the
position and orientation of the coil with respect to the
marked TMS target sites could be monitored on-line.
rTMS was applied using a Magstim Rapid Rate stimula-
tor (Magstim, UK) and a focal eight-shaped coil with
wings each measuring 70 mm in outer diameter. Stimu-
lation was applied at a frequency of 1 Hz over 12 min for a
total of 720 pulses. Figure 1 illustrates where TMS was
delivered in each of the four target brain areas in a
representative subject. Although many rTMS studies (in-
cluding those reported in Shapiro et al., 2001) tailor the
intensity of stimulation to the motor threshold in indi-
vidual subjects, this approach is problematic inasmuch as
cortical excitability may vary across different brain areas
(Robertson, Theoret, & Pascual-Leone, 2003). We there-
fore opted to use a fixed intensity of stimulation across
subjects, corresponding to 65% of the maximum stimu-
lator output. For sham stimulation, we used a specially
designed coil that induces no magnetic field, but pro-
duces an acoustic artifact and a tapping sensation on the
scalp similar to real TMS (www.magstim.com). Using this
sham coil, TMS-naive subjects are truly blinded to the
sham nature of the stimulation (unpublished data).
Data Analysis
Individual trials were excluded from the dataset when sub-
jects responded incorrectly, self-corrected, or hesitated,
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
f
.
/
.
.
t
o
n
1
8
M
a
y
2
0
2
1
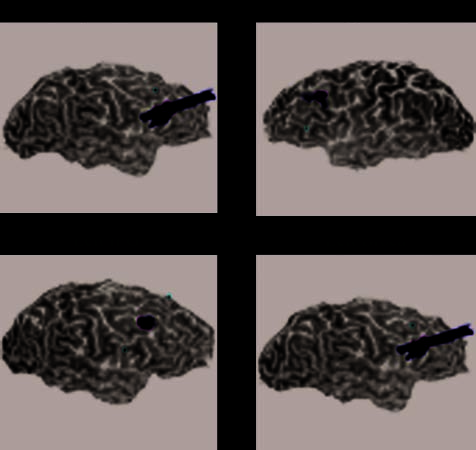
Figure 1. Sites and targeting of rTMS and sham stimulation. In
Experiments 1 and 2, the order of stimulation conditions was fully
counterbalanced across subjects. Yellow lines illustrate an example of
where stimulation was applied during an entire 10-min rTMS train in a
representative subject for each of the four stimulation positions; the
green dots correspond to the point on the scalp where each TMS pulse
was delivered. The stimulation coil rested tangentially on the subject’s
scalp and the handle pointing posteriorly parallel to the subject’s
midsagittal plane as calculated by the frameless stereotactic.
Cappelletti et al.
711
when no response was recorded, and when recorded
reaction times (RTs) were less than 200 msec. Using
these criteria, 3.8% and 3.5% of the total number of
responses were counted as errors in Experiments 1 and
2, respectively. Visual inspection of histograms of the
remaining data confirmed that RTs were approximately
normally distributed. Experimental conditions in which
real stimulation was applied were compared to condi-
tions in which sham stimulation was applied. RTs fol-
lowing left- and right-sided sham stimulation were
averaged. We used Fisher’s LSD correction for multiple
comparisons, and statistical significance refers to a two-
tailed p value < .05.
EXPERIMENTS
Experiment 1: Stimulation to Left and Right
Anterior Midfrontal Regions (aMFG)
Our first experiment was designed to replicate and extend
the study by Shapiro et al. (2001), with two specific aims:
(1) to determine whether the left aMFG is critical for
processing both regular and irregular verbs—or, con-
versely, whether the effect of rTMS on this region is
modulated by the morphological regularity of the words
to be produced; and (2) to investigate the role of the
homologous midfrontal area in the right hemisphere. We
reasoned that if the left aMFG is critical for processing
verbs irrespective of their morphological features, rTMS
should equally affect regular and irregular verbs. More-
over, if verb processing critically depends on the left
aMFG, we predicted that no specific impairment in per-
formance should follow stimulation to the right aMFG.
Results
We initially tested for effects of stimulation using a 4 (cid:1) 3
repeated measures analysis of variance (ANOVA) on
mean RTs by subject, with trial types (regular and irreg-
ular verbs and nouns) and conditions of stimulation (left
aMFG, right aMFG, and sham) as factors. When appro-
priate, post hoc t tests were carried out to compare spe-
cific conditions to their respective controls.
Following real stimulation, mean response latencies dif-
fered significantly between the sham condition (643 msec),
stimulation to the right aMFG (645 msec), and stimula-
tion to the left aMFG [697 msec; main effect of stimula-
tion condition, F(2, 14) = 6.23 p < .02]. However, the
effect of stimulation differed across trial types [F(3, 21) =
6.03, p < .005], and a significant two-factor interaction
[F(6, 42) = 2.56, p < .05] emerged.3
Post hoc comparisons showed that RTs increased
following rTMS to the left aMFG, compared to sham
stimulation, only for regular and irregular verb produc-
tion [t(7) = (cid:2)2.61, p < .02 and t(7) = (cid:2)2.14 p < .005,
respectively], but not for the production of regular or
irregular nouns [t(7) = (cid:2)0.358 and t(7) = (cid:2)1.28, ns,
respectively; see Figure 2].
Similar findings were obtained when the effect of
stimulation to the left aMFG was contrasted with the
effect of stimulation to the right aMFG (rather than to
sham). A significant increase in RTs was observed for
verbs [regular: t(7) = 3.56, p < .01; irregular: t(7) =
4.29, p < .005], but not for nouns [regular: t(7) = 0.094,
ns; irregular: t(7) = 1.91, ns]. No significant effects or
trends (.05 < p < .10) were found when we compared
RTs after stimulation to the right aMFG with RTs after
sham stimulation.
Summary and Interim Discussion
These findings indicate that the left aMFG appears to be
more strongly involved in the morphological processing
of verbs than nouns. The data are consistent with the
results obtained by Shapiro et al. (2001), and extend the
conclusions of that study in two ways.
First, the results show that the effect of stimulation on
response latency applies to both regular and irregular
morphological transformations with verbs. Just as im-
portantly, the effect was replicated with a different set of
inflectional features—in this case, tense rather than
agreement. This suggests that the computation carried
out by the left aMFG (or the network to which it is
immediately connected) operates at a level more general
than those at which either morphophonological features
Figure 2. Effects of rTMS
in Experiment 1. Difference
in average response times
(RTs) between left aMFG
and sham stimulation (left
columns) and left aMFG and
right MFG (right columns).
712
Journal of Cognitive Neuroscience
Volume 20, Number 4
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
f
.
t
/
.
o
n
1
8
M
a
y
2
0
2
1
(such as regularity) or particular inflectional features
(such as tense or agreement) are represented. We pro-
pose that this level can be referred to as verbal, as
opposed to nominal, morphosyntax; or, in other words,
as grammatical computations pertaining to verbs. Alter-
natives to this formulation will be considered in the
General Discussion.
Second, these results demonstrate that rTMS is effec-
tive in delaying verb production only when applied to
the left hemisphere, and therefore, that the results
obtained by Shapiro et al. (2001) cannot be attributed
to a general effect of TMS on cortical function. However,
this is obviously a very coarse-grained statement about
the organization of the cortex; even if it is correct, it
does not rule out the possibility that the effect at the left
aMFG site results from a spread of inhibition to neigh-
boring regions, or that other areas in the left (or right)
frontal lobe are also important for verb processing in
this task. Experiment 2 addressed this issue.
Experiment 2: Stimulation to Other Frontal
Regions (pMFG and IFG)
This experiment was designed to clarify whether the
effects of stimulation to the left aMFG obtained in our
Experiment 1 and by Shapiro et al. (2001) are focal and
specific to this region. To this end, we targeted two
areas within the frontal lobe adjacent to, but distinct
from, the region targeted in Experiment 1. Specifically,
we used the same experimental approach to target the
pMFG and the triangular part of the IFG (corresponding
to Broca’s area in the left hemisphere).
Results
A 4 (cid:1) 5 repeated measures ANOVA on the RTs of correct
answers tested for stimulation effects with stimulus type
(regular and irregular verbs and nouns) and condition
(left and right IFG, left and right pMFG, and sham) as
factors. Post hoc comparisons were performed as in
Experiment 1.
Response latencies differed significantly between real
and sham stimulation conditions [left IFG: 591 msec;
right IFG: 450 msec; left pMFG: 518 msec; right pMFG:
597 msec; sham: 536 msec; main effect of condition,
F(4, 44) = 10.85, p < .0001 and stimulus, F(3, 33) =
26.73, p < .0001; Condition (cid:1) Stimulus interaction,
F(12, 132) = 1.78, p = .056]. Specifically, real stimula-
tion to the left IFG produced a significant delay relative
to sham stimulation for regular nouns [t(11) = 2.69,
p < .05] but not for irregular nouns [t(11) = 1.8, ns],
and for both regular [t(11) = 2.96, p < .02] and irregular
verbs [t(11) = 2.50, p < .05]. However, a direct com-
parison showed no significant difference between RTs to
regular and irregular nouns following stimulation to the
left IFG area [t(11) = 0.01, ns].
When stimulation to the left IFG was compared to
stimulation to the right IFG, significant delays in re-
sponse latency were observed for both regular nouns
[t(11) = 5.75, p < .0001] and irregular nouns [t(11) =
6.11, p < .0001], and for both regular verbs [t(11) =
7.99, p < .0001] and irregular verbs [t(11) = 7.23,
p < .0001]. Stimulation to the left pMFG did not
produce effects that differed from sham stimulation
for either regular or irregular nouns [t(11) = 1.31 and
t(11) = 0.093, ns, respectively], or for regular or irreg-
ular verbs [t(11) = 1.87, ns, p = .087 and t(11) = 1.59,
ns, respectively]. On the other hand, stimulation to
the right pMFG resulted in RTs that were significantly
lower than stimulation to the left pMFG for all trial
types [regular nouns: t(11) = 3.63, p < .005; irregular
nouns: t(11) = 3.55, p < .005; regular verbs: t(11) =
4.19, p < .001; irregular verbs t(11) = 2.44, p < .05; see
Figure 3].4
Comparison of Experiments 1 and 2
An ANOVA with the factor area (3 levels) nested within
experiment (2 levels) compared real rTMS effects at the
three different sites within the left hemisphere (aMFG,
pMFG, and IFG). This analysis showed that the effects of
stimulation to the left aMFG differed reliably from
stimulation to the left pMFG and left IFG when verbs
and nouns were considered together [main effect of
experiment: F(1, 5) = 4.97, p < .001]. Post hoc compar-
isons showed that stimulation to the left aMFG resulted
in significantly greater interference with verb production
that stimulation to either the pMFG [t(18) = 2.557,
p < .02] or the IFG [t(11) = (cid:2)3.51, p < .05]. No
significant difference emerged between the aMFG and
IFG conditions for noun production [t(11) = 1.36, ns],
although there was a trend toward increased interfer-
ence for nouns following stimulation to the left aMFG
compared to the left pMFG [t(18) = 2.07, p < .07].
Summary
Experiment 2 demonstrated that neither the IFG nor the
pMFG, in either hemisphere, differentiates between verb
and noun processing. Although rTMS to the left IFG
produced a delay in RTs (compared to sham stimulation
and to stimulation to the homologous region in the right
hemisphere), this effect was not specific to verbs. Stim-
ulation to the left pMFG did not produce any delay in
RTs compared to sham stimulation.
Likewise, no category-specific effects were observed
following stimulation to the right hemisphere. However,
response latencies for all trial types following stimulation
to the right pMFG were significantly shorter than they
were following stimulation to the left pMFG. These re-
sults suggest that category-general cross-hemisphere in-
teractions may emerge when we compare the effects of
rTMS to areas in the left and right frontal lobes (see
Cappelletti et al.
713
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
.
f
.
t
/
o
n
1
8
M
a
y
2
0
2
1
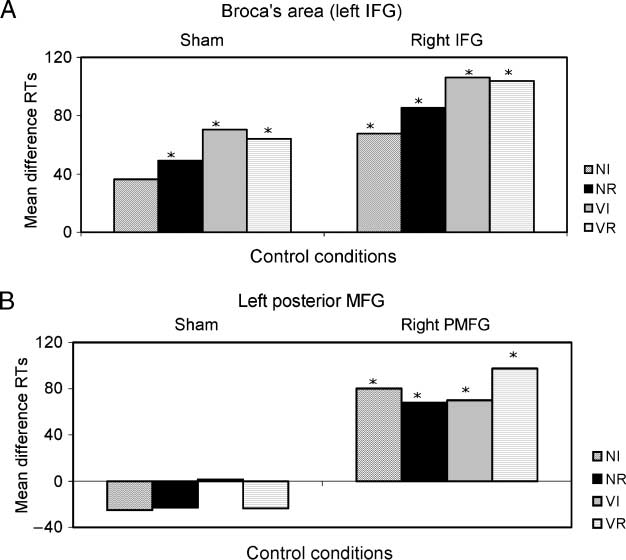
Figure 3. Effects of rTMS in
Experiment 2. (A) Difference
in average response times
(RTs) between left Broca’s
areas and sham stimulation
(left columns) and left and
right Broca’s areas (right
columns). (B) Difference in
average RTs between the
left posterior IMG and sham
stimulation (left columns) and
the left and right posterior
IMG (right columns).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
t
/
.
.
f
.
o
n
1
8
M
a
y
2
0
2
1
General Discussion, below). Moreover, in the case of
verb processing, the effects of stimulation were signifi-
cantly different in the aMFG compared to the other two
regions in the left hemisphere.
There was no change in error rate following stimula-
tion to any area in the left or right hemisphere. This is
consistent with findings from other TMS studies, which
have shown that TMS produces an increase in mean RTs,
but not an increase in error rates, for tasks performed at
a high level of accuracy (e.g., Cappelletti, Barth, Fregni,
Pascual-Leone, & Spelke, 2007; Alexander et al., 2005;
Ashbridge et al., 1997).
GENERAL DISCUSSION
In this study we used rTMS to explore the role of frontal
cortical regions in processing regular and irregular verbs
and nouns. We found greater interference for verb
production than noun production when stimulation
was applied to the left aMFG, but not to the right aMFG
(Experiment 1) or to two other areas in the frontal lobes
that are hypothesized to play important roles in verb
processing (Experiment 2).
Three important conclusions can be drawn from these
data. First, they strengthen the evidence that rTMS ap-
plied to the anterior portion of the left midfrontal gy-
rus interferes with the ability to produce verbs, but
not nouns, in agreement with the results obtained by
Shapiro et al. (2001). Second, they extend the previous
results by showing that rTMS affects the production of
both regular and irregular verb forms, suggesting that
this area is involved in processing at a level of compu-
tation that does not discriminate between rule-based
and irregular morphophonological transformations. In-
deed, we found no such distinction, for either nouns or
verbs, at any of the cortical sites targeted with rTMS.
Third, the current results demonstrate that the effects
of rTMS on verb processing are specific to the left aMFG;
no category-specific interference effects were observed
when rTMS was applied to the left pMFG and to Broca’s
area. Instead, stimulation to these areas seemed to
result in generic interference with both noun and verb
trials, suggesting that they may be engaged in produc-
tion processes that do not discriminate between words
of different grammatical categories. This is in contradis-
tinction to results from functional neuroimaging studies,
which have revealed noun–verb differences in both the
IFG (Longe et al., 2007; Tyler et al., 2004; Perani et al.,
1999) and the pMFG (Shapiro & Caramazza, 2006).
Verb Processing in the Left Anterior Midfrontal
Gyrus: Functional Considerations
Observations of language deficits in brain-damaged pa-
tients suggest that the production of nouns and verbs in
speech can be differentially impaired (e.g., Daniele et al.,
1994; Miozzo, Soardi, & Cappa, 1994; Damasio & Tranel,
1993; Caramazza & Hillis, 1991; Miceli et al., 1984; see
Shapiro & Caramazza, 2003c; Druks, 2002, for reviews).
These behavioral dissociations have been taken as evi-
dence that distinct cortical areas are involved in noun
and verb production. The most common anatomical
714
Journal of Cognitive Neuroscience
Volume 20, Number 4
correlate of verb impairment involves damage to the left
frontal lobe. The association between verb production
and frontal regions has been replicated, albeit inconsis-
tently, by some neuroimaging studies, which have
shown that verb processing is associated with increased
activation in prefrontal regions (Shapiro et al., 2005,
2006; Tyler et al., 2001, 2004; Perani et al., 1999; Herholz
et al., 1996; Warburton et al., 1996; Crivello et al., 1995).
However, limited evidence has so far been provided as
to whether frontal brain regions are really critical for the
production of verbs as defined by their grammatical
role—as opposed, for example, to the processing of
words specifying concrete actions (Pulvermu¨ller, 1999,
2001, 2005; Cappa, Sandrini, Rossini, Sosta, & Miniussi,
2002; Neininger & Pulvermu¨ller, 1997), or of words that
happen to be poorly imageable (Bird et al., 2000). Some
patient studies have attempted to address this problem
by using real word and pseudoword stimuli in tasks that
explicitly emphasize morphosyntactic transformation
(Laiacona & Caramazza, 2004; Shapiro & Caramazza,
2003c; Shapiro et al., 2000), on the assumption that
category-selective deficits in this context are difficult to
impair-
interpret as anything other than grammatical
ments. The same logic was used in our earlier rTMS
study, which showed that suppression of the left aMFG
can be used to dissociate the processing of real words
and pseudowords as nouns and verbs in unimpaired
subjects.
The results of the current experiments show that
stimulation to the aMFG, and not to other frontal regions,
results in temporary interference with verb production,
but not noun production, thus reinforcing the idea that
the left aMFG, in particular, plays a critical role in verb
processing. Using the same logic that was applied in our
earlier studies, we have argued that this effect arises
because the left aMFG is critical for processing certain
kinds of grammatical operations—specifically, morpho-
syntactic operations that call for the application of in-
flectional morphemes restricted to the category of verbs.
This, of course, is not the only possible interpretation of
our results. We have not definitively ruled out an expla-
nation on the basis of the different kinds of semantic
features of nouns and verbs.
In the present study, groups of noun and verb stimuli
were not matched for concreteness, although they did
not differ significantly in imageability, a correlated se-
mantic variable that has more often been invoked to
account for specific verb deficits in aphasia (Crepaldi
et al., 2006; Luzzatti et al., 2002; Bird et al., 2000). It is
therefore possible that what we have called a noun–verb
dissociation in the left aMFG could be attributable to this
factor. We believe, however, that this is improbable
for several reasons. To begin with, an effect of concrete-
ness would more naturally be expected in premotor
cortical regions, where no noun–verb effects were found
(Pulvermu¨ller, 1999, 2001, 2005). An explanation in
terms of concreteness is also unparsimonious on the
basis of past results, including the pseudoword data in
Shapiro et al. (2001).
One could argue that the effects with pseudowords
were also ‘‘semantic,’’ but this would require one to
qualify what is meant by ‘‘semantics.’’ In this context, it
might be supposed that the postulated verbal semantic
features are so abstract as to apply even to words with
no specific meaning, which would divorce verb seman-
tics from a notion of concreteness that reflects the
activation of action schemata. This was essentially the
argument we adopted in Shapiro et al. (2006), in which
we proposed that a different cortical region activated in
fMRI may have a role in representing the ‘‘core’’ or
generic semantics of eventhood (but see below for a
novel interpretation of that study). Alternatively, it may
be that subjects adopted a strategy of associating pseu-
dowords (like wug) with particular actions (like wag). It
is unclear, however, why this additional, meta-cognitive
assumption should be preferred to a simple explanation
in terms of grammatical processing.
Verb Processing in the Left Frontal Cortex:
Anatomical Considerations
The area that we stimulated in the left aMFG is included
wholly or partially in the lesions observed in several
patients with selective verb deficits, including one pa-
tient who had analogous difficulties in producing verbs
and pseudowords in the context of a morphological
alternation task (Shapiro & Caramazza, 2003b). It may
also correspond to the brain region implicated in so-
called dynamic aphasia, which was characterized in part
as involving a selective deficit in predication—that is, in
using verbs in their natural syntactic context (Luria &
Tsvetkova, 1967).
It must be said, however, that it is difficult to make
more precise claims about the degree of anatomical
similarity between the area targeted in this study and
cortical regions damaged in patients or activated in
functional neuroimaging studies. Even if we are mindful
of concerns about the reliability of anatomical localiza-
tion in functional neuroimaging (Brett, Johnsrude, &
Owen, 2002), it is notable that patterns of frontal acti-
vation for verb production have varied widely, including
almost every major division of the frontal lobes (in both
hemispheres) in one study or another.
Role of the Posterior Middle Frontal Gyrus
In a recent fMRI study we conducted, using a behavioral
paradigm identical
in important respects to the task
used here, the contrast between verb production and
noun production isolated three areas that seemed to be
more engaged for verb trials, including one area in the
left frontal cortex (Shapiro et al., 2006). However, this
area is anatomically more comparable to the pMFG re-
gion in this study—where TMS produced no differential
Cappelletti et al.
715
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
f
.
.
/
.
t
o
n
1
8
M
a
y
2
0
2
1
effects across categories—than to the aMFG area where
category-specific suppression was found. Other func-
tional neuroimaging studies have also shown that verb
production activates parts of the left posterior prefrontal
cortex (Herholz et al., 1996; Crivello et al., 1995). The
implications of this apparent discrepancy between TMS
and fMRI/PET results are unclear.
One possibility is that the stages of cognitive process-
ing illuminated by functional neuroimaging and sup-
pressed by rTMS are not entirely isomorphic. In the
case at hand, it may be that the various neuroimaging
studies revealed an area which is activated by the ‘‘core’’
semantic properties of verb stimuli (O’Grady, 1997), but
which is not necessarily critical for the production of
verbal morphology. If we grant that the fMRI study
conducted by Shapiro et al. (2006) did not reveal areas
engaged in morphological processing, it may be that this
insensitivity resulted from the massive repetition of the
specific morphological alternations, with adaptation in
the blood oxygenation level-dependent (BOLD) re-
sponse to the aspects of the task relevant to grammatical
processing. The residual significant BOLD response may
have been more sensitive to semantic features of the
noun and verb stimuli, which were obviously repeated
less often than the morphological cues. By contrast,
rTMS interferes directly with cortical activity so the
effects induced by rTMS may be less affected by repeti-
tion in a task.
Role of the Inferior Frontal Gyrus
As we mentioned earlier, some functional imaging stud-
ies have also reported increased activation in the left IFG
for verbs compared to nouns (Longe et al., 2007; Tyler
et al., 2004; Perani et al., 1999). This effect has been
attributed to the processing of inflectional morphology
for verbs, which is assumed to be computationally more
demanding than processing nominal morphology (Longe
et al., 2007; Tyler et al., 2004). A full discussion of
the problems raised by this account is beyond the scope
of the present article. One question that is pertinent,
however, is how much of this postulated computational
disparity can be credited to task-specific or even stimulus-
specific differences in various experiments. In the most
recent and probably the best controlled of the above-
mentioned studies (Longe et al., 2007), subjects per-
formed a lexical decision task on unambiguous plural
nouns (bullets) and third-person singular verbs (sings).
It is probably reasonable to argue that the syntactic
functions signaled by these inflections are mismatched
in complexity—indeed, this is, in part, what motivated
us to use the past/unmarked alternation for verbs in the
present study (see Introduction).
Moreover, the presentation of English nouns and
verbs as single words, outside of a phrasal context, is
much less natural for verbs, which do not occur in many
inflectional forms without an accompanying subject
(consider What are these? Bullets. vs. What does he do?
*Sings.) It is not even obvious how one should construe
the English /s/ suffix without contextual cues. If gram-
matical category information is not encoded in the
representation of lexical roots, as some theorists have
argued (Marantz, 1997; Halle & Marantz, 1993), then
part of the lexical decision process might involve reject-
ing the construals that are unlikely or prohibited on
semantic or thematic grounds. This decision, in turn,
could interact with factors like the relative frequency
of the distinct /s/ morphemes. Because of these numer-
ous uncertainties,
it may be difficult to interpret the
presence or absence of grammatical category effects in
paradigms that do not explicitly involve grammatical
processing (Shapiro & Caramazza, 2003c).
In the present study, we did not observe any categor-
ical dissociation following stimulation to the left IFG.
Instead, stimulation of Broca’s area resulted in an in-
crease in response latencies for both noun and verb
trials, suggesting that this part of the brain subserves
category-general aspects of morphological production.
Such a pattern of performance is in agreement with a
large corpus of neuropsychological studies showing that
damage to Broca’s area results in impaired processing of
inf lectional morphology for both nouns and verbs
(Menn & Obler, 1990; Miceli et al., 1984). Further
evidence along these lines comes from a TMS study by
Sakai, Noguchi, Takeuchi, and Watanabe (2002), who
showed that stimulation of Broca’s area results in selec-
tive impairment in a task that involves the detection of
anomalous morphemes (Japanese case markers, which
are postnominal clitics).
The result could also be compatible with the idea that
Broca’s area is engaged in a way that is proportional to
the difficulty of the computation required, or the num-
ber of alternative responses that exist (if not in the
context of the task, then in the language as a whole).
On the other hand, a number of prior studies using both
functional neuroimaging and TMS have suggested that
Broca’s area may in fact be parcellated into functionally
distinct subregions (see Devlin & Watkins, 2007, for a
review). Although this work has focused primarily on the
difference between semantic and phonological process-
ing, finer-grained distinctions may exist, suggesting that
the region targeted here is not representative of the IFG
as a whole. We ourselves have predicted elsewhere that
part of Broca’s area might be important for processing
of nouns (Shapiro & Caramazza, 2003b), based on the
observation of a patient who had a left suprasylvian
lesion that was accompanied by noun production diffi-
culties (Shapiro et al., 2000). The present results offer no
support for this prediction, although of course it may be
that the processing of nouns depends on some other
region within Broca’s area or elsewhere in the left
frontal cortex that was not targeted in the present study.
Finally, we should reiterate the possibility raised
elsewhere in this article that the effects of stimulation
716
Journal of Cognitive Neuroscience
Volume 20, Number 4
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
f
.
.
t
.
/
o
n
1
8
M
a
y
2
0
2
1
to the left aMFG are not attributable to the disruption of
the left aMFG per se, but are rather attributable to the
transsynaptic suppression of some other cortical region
or regions. If this is true, the distant site is presumably
not one of those stimulated in Experiment 2, nor one to
which either of those areas is connected very strongly.
Therefore, a conservative conclusion would be that the
effects arise either at the aMFG itself, or in a region or
network to which the left aMFG has what we have called
a ‘‘privileged’’ connection.
Processing of Other Grammatical Categories
The experiments reported here were aimed, in part, at
clarifying the role of a part of the left prefrontal cortex
that is known from prior studies to be engaged in verb
processing. We did not explicitly aim to target areas that
might be involved in the processing of other grammat-
ical categories, although as we have noted, we found
no support for a prior prediction that the IFG might
be engaged selectively in noun processing (Shapiro &
Caramazza, 2003b). At present, there are no data that
point to any particular brain region that might be
engaged specifically in the grammatical processing of
nouns or words of other categories (e.g., adjectives).
The available data from neuropsychology (Daniele et al.,
1994; Damasio & Tranel, 1993) and neuroimaging
(Shapiro et al., 2006) suggest that lexical–semantic pro-
cessing of nouns relies on portions of the left temporal
lobe that, unfortunately, are largely inaccessible to TMS.
Therefore, any attempt to demonstrate a double disso-
ciation in processing between verbs and another cate-
gory using TMS, however desirable this might be from
a theoretical perspective, should await further advances
in empirical knowledge about the anatomy of lexical
processing.
Processing Regular and Irregular
Morphological Inflection
The contrast between regular and irregular nouns and
verbs in Experiments 1 and 2 allowed us to assess the
level at which category-specific effects arise in the left
aMFG. In other words, we tested whether repeated
stimulation interferes with the production of words of
a specific category (verbs), words undergoing a specific
morphological process (regular morphological
inflec-
tion), or some interaction between these two dimen-
sions. We showed that rTMS applied to the left aMFG
interferes equally with the production of regularly and
irregularly inflected verbs, and does not interfere with
the production of either regular or irregular nouns.
Moreover, no regularity effect was observed following
stimulation to the pMFG or Broca’s area.
These results do not seem to support the view that
regular and irregular forms are processed by two distinct
and neuroanatomically segregated mechanisms, with
regularly inflected forms relying on the left inferior
frontal cortex (e.g., Tyler et al., 2001, 2004; Ullman,
Corkin, et al., 1997). At none of the three sites of
stimulation in the left frontal cortex (nor at any of their
right hemisphere homologues) did we observe a specific
effect on the production of regular words.
Two explanations may be suggested to account for
our results. First, failure to detect any difference be-
tween regular and irregular form processing may sup-
port the view that these forms are implemented by a
single cognitive mechanism (McClelland & Patterson,
2002; Joanisse & Seidenberg, 1999). This view holds that
inflectional processes arise from a single integrated
system, namely, that both regular and irregular verb
inflection emerge from one distinct mechanism. How-
ever, some recent neuropsychological cases of selective
impairment of irregular verb forms (e.g., Miozzo, 2003)
seem problematic for the single-mechanism account and
require further explanations.
An alternative account is that regular and irregular
forms are processed, at some level, by distinct neural
circuits, but that this distinction is not reflected in the
organization of the cortical regions targeted in our ex-
periments. We have proposed that the area targeted in
the left aMFG is crucial for morphosyntactic processes
that apply to the grammatical category of verbs (cf. also
Shapiro et al., 2001). Such processes could, in principle,
operate over rather abstract kinds of representations; for
example, a morphosyntactic rule might mark a lexical
representation to be produced with the tense feature
[+PAST], without specifying the phonological conse-
quences of that feature setting. This would then require
some other system to cash out abstract morphosyntactic
features in phonological terms (e.g., the suffix /ed/ for
regular verbs, or an internal vowel change like ablaut for
some irregular verbs). Our results do not speak to the
question of whether some brain regions might be more
crucial for the morphophonological computation of
regular or irregular forms for nouns or verbs, except to
say that the areas we stimulated in the frontal lobes do
not seem to encode any such distinction. It may be
the case that portions of Broca’s area caudal to the re-
gion stimulated here, which are thought to be engaged
in phonological processing (see Devlin & Watkins, 2007),
are important for this kind of morphophonological
process.
Right Hemisphere Effects
For each condition in which an area in the left frontal
cortex was targeted with rTMS, there were two controls:
sham stimulation and rTMS to the homologous area in
the right hemisphere. We reasoned that if any area in
the left hemisphere was found to be critical for verb or
noun processing, it would be unlikely that a similarly
specific impairment in performance should be observed
after stimulation to right hemisphere areas. Indeed,
Cappelletti et al.
717
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
.
t
/
f
.
o
n
1
8
M
a
y
2
0
2
1
Experiment 1 confirmed this prediction: Verb processing
was delayed following left aMFG stimulation, but not
after sham stimulation or stimulation to the right aMFG.
Interestingly, however, the overall effects of stimulat-
ing the right aMFG were not the same as the effects of
sham stimulation. The difference in RTs for every trial
type was greater when left hemisphere rTMS was com-
pared to right hemisphere rTMS than when left hemi-
sphere rTMS was compared to sham stimulation. In
Experiment 2, this trend was more pronounced: The
difference between left rTMS and right rTMS was greater
than the difference between left rTMS and sham for all
trial types in both areas that were stimulated (pMFG and
IFG). In other words, stimulation to the right hemi-
sphere seemed to result in a relative facilitation of
responses across categories.
We should reiterate that this facilitation effect did not
seem to be stimulus-specific; the only specific effect
observed in either experiment was the delay in verb
processing following stimulation to the left aMFG. Having
said this, two interesting aspects of the left–right pattern
deserve comment. One is that the facilitatory effect of
stimulation to the right hemisphere (compared to sham)
appeared to increase in an anterior-to-posterior direc-
tion. The second is that the absolute difference between
stimulation to the left and right appeared to be relatively
constant, especially for the IFG and the pMFG. Although
the importance of these results is not obvious, they
suggest that right hemisphere cortical areas may modu-
late the activity of homologous areas in the left hemi-
sphere that are engaged in language production. This
kind of interhemispheric interaction is believed to be
mediated by callosal fibers, and has been described in
areas such as the motor cortex and parietal lobes in
both neurologically healthy subjects and in patients
with stroke (Mansur et al., 2005; Naeser et al., 2005a,
2005b; Kobayashi, Hutchinson, Theoret, Schlaug, & Pascual-
Leone, 2004; Theoret, Kobayashi, Valero-Cabre, & Pascual-
Leone, 2003; Hilgetag, Theoret, & Pascual-Leone, 2001).
The present results suggest that transcallosal inhibition
may also be extended to lexical processing.
CONCLUSION
We have previously suggested that verbs are processed
by distinct cortical circuits that can be identified by tar-
geted suppression of the left prefrontal cortex (Shapiro
et al., 2001). In the present investigation, with the use of
more sophisticated methods and an improved experi-
mental design, we confirmed this hypothesis, at least
insofar as it applies to the contrast between verbs and
nouns. In addition, we have drawn three other impor-
tant conclusions: (1) the left aMFG is equally involved in
processing regular and irregular verbs, as no difference
in response latencies emerged between them following
rTMS; (2) the effect of rTMS on the left aMFG is rel-
atively specific and focal, as no interference resulted
after stimulating the left pMFG; (3) rTMS applied to
Broca’s area did not specifically interfere with verb pro-
duction, but seemed to result in category-general inter-
ference with word production.
Acknowledgments
This work was supported by British Academy and Wellcome
Trust Grants to M. C., a grant from the Harvard Medical School
Scholars in Clinical Science Program (NIH K30 HL04095-03) to
F. F., grants from the Sackler Scholars Programme in Psycho-
biology and the National Institute of Neurological Disorders
and Stroke (F30 NS50906) to K. S., a grant from the National
Institute of Deafness and Communication Disorders to A. C.,
and by NIH grants (K24 RR018875; RO1-DC05672; RO1-NS
20068) to A. P. L. We thank Matthew Finkbeiner and Elliot
Freeman for technical support and the Harvard-Thorndike
General Clinical Research Center (NCRR MO1 RR01032) for
help in implementing the study.
Reprint requests should be sent to Marinella Cappelletti, Ins-
titute of Cognitive Neuroscience, University College London,
17 Queen Square, London WC1N 3AR, UK, or via e-mail:
m.cappelletti@ucl.ac.uk.
Notes
1. Other word classes, like adjectives, may also be associated
with distinct neural circuits, but the status of categories other
than nouns and verbs has not been well studied. There are,
however, a number of more principled reasons for focusing on
nouns and verbs, including the consensus among linguists that
nouns and verbs (but no other categories) are distinguished in
every human language (Evans, 2000; Jacobsen, 1979; Robins,
In this article, the phrases ‘‘category-
1952; Sapir, 1921).
specific,’’ ‘‘selective,’’ and the like refer only to the categories
of nouns and verbs.
2.
In Shapiro et al., we attempted to address this concern by
showing that the effects of rTMS were not greater for trials with
longer reaction times at baseline; indeed, less interference was
observed for more difficult trials within a category.
3. A similar ANOVA on error rates showed no significant main
effect of stimulation condition [F(2, 14) = 0.173, ns] or of trial
types [F(3, 21) = 1.745, ns], and no significant interaction [F(6,
42) = 0.901, ns].
4. A similar ANOVA on error rates showed no significant main
effect of stimulation condition [F(4, 44) = 0.167, p < .95, ns]
and of stimulus [F(3, 33) = 1.507, p < .23, ns] and no signif-
icant Condition (cid:1) Stimulus interaction [F(12, 132) = 0.950,
p = .5, ns].
REFERENCES
Alexander, I., Cowey, A., & Walsh, V. (2005). The right parietal
cortex and time perception: Back to Critchley and the
Zeitraffer phenomenon. Cognitive Neuropsychology, 22,
306–315.
Ashbridge, E., Walsh, V., & Cowey, A. (1997). Temporal aspects
of visual search studies by transcranial magnetic stimulation.
Neuropsychologia, 35, 1121–1131.
Balaguer, R. D. D., Costa, A., Sebastian-Galles, N., Juncadella,
M., & Caramazza, A. (2004). Regular and irregular
morphology and its relation with agrammatism: Evidence
from two Spanish–Catalan bilinguals. Brain and Language,
91, 212–222.
718
Journal of Cognitive Neuroscience
Volume 20, Number 4
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
/
.
.
.
f
t
o
n
1
8
M
a
y
2
0
2
1
Bates, E., Chen, S., Tzeng, O., Li, P., & Opie, M. (1991). The
noun–verb problem in Chinese. Brain and Language, 41,
203–233.
Berndt, R. S., Haendiges, A. N., & Burton, M. W. (2002).
Grammatical class and imageability in aphasic word
production: Their effects are independent. Journal of
Neurolinguistics, 15, 353–371.
Bird, H., Howard, D., & Franklin, S. (2000). Why is a verb
like an inanimate object? Grammatical category and
semantic category deficits. Brain and Language, 72,
246–309.
Brett, M., Johnsrude, I. S., & Owen, A. M. (2002). The problem
of functional localization in the human brain. Nature
Reviews Neuroscience, 3, 243–249.
Brown, W. S., Marsh, J. T., & Smith, J. C. (1973). Contextual
meaning effects on speech evoked potentials. Behavioral
Biology, 9, 755–761.
Buccino, G., Riggio, L., Melli, G., Binkofski, F., Gallese, V., &
Rizzolatti, G. (2005). Listening to action-related sentences
modulates the activity of the motor system: A combined TMS
and behavioural study. Brain Research Cognitive Brain
Research, 24, 355–363.
Cappa, S. F., Sandrini, M., Rossini, P. M., Sosta, K., & Miniussi,
C. (2002). The role of the left frontal lobe in action naming:
rTMS evidence. Neurology, 59, 720–723.
Cappelletti, M., Barth, H., Fregni, F., Pascual-Leone, A., &
Spelke, E. (2007). rTMS over the left and the right
intraparietal sulcus disrupts discrete and continuous
quantity processing. Experimental Brain Research, 179,
631–642.
Caramazza, A., & Hillis, A. (1991). Lexical organization of nouns
and verbs in the brain. Nature, 349, 788–790.
Crepaldi, D., Aggujaro, S., Arduino, L. S., Zonca, G., Ghirardi,
G., Inzaghi, M. G., et al. (2006). Noun–verb dissociation in
aphasia: The role of imageability and functional locus of the
lesion. Neuropsychologia, 44, 73–89.
Crivello, F., Tzourio, N., Poline, J. B., Woods, R. P., Mazziotta,
J. C., & Mazoyer, B. (1995). Intersubject variability in
functional neuroanatomy of silent verb generation:
Assessment by a new activation detection algorithm based
on amplitude and size information. Neuroimage, 2, 253–263.
Damasio, A., & Tranel, D. (1993). Nouns and verbs are
retrieved with differentially distributed neural systems.
Proceedings of the National Academy of Sciences, U.S.A., 90,
4957–4960.
Daniele, A., Giustolisi, L., Silveri, M., Colosimo, C., & Gainotti,
G. (1994). Evidence for a possible neuroanatomical basis for
lexical processing of nouns and verbs. Neuropsychologia,
32, 1325–1341.
Dehaene, S. (1995). Electrophysiological evidence for category
specific word processing in the normal human brain.
NeuroReport, 6, 2153–2157.
De Renzi, E., & di Pellegrino, G. (1995). Sparing of verbs and
preserved but ineffectual reading in patient with impaired
word production. Cortex, 31, 619–636.
Devlin, J. T., & Watkins, K. E. (2007). Stimulating language:
Insights from TMS. Brain, 130, 610–622.
The view from building 20 (pp. 111–176). Cambridge:
MIT Press.
Hauk, O., Johnsrude, I., & Pulvermu¨ller, F. (2004).
Somatotopic representation of action words in human
motor and premotor cortex. Neuron, 41, 301–307.
Herholz, K., Thiel, A., Wienhard, K., Pietrzyk, U., von
Stockhausen, H. M., Karbe, H., et al. (1996). Individual
functional anatomy of verb generation. Neuroimage, 3,
185–194.
Hilgetag, C. C., Theoret, H., & Pascual-Leone, A. (2001).
Enhanced visual spatial attention ipsilateral to rTMS-induced
‘‘virtual lesions’’ of human parietal cortex. Nature
Neuroscience, 4, 953–957.
Hillis, A., & Caramazza, A. (1995). Representation of
grammatical categories of words in the brain. Journal of
Cognitive Neuroscience, 7, 396–407.
Indefrey, P., Brown, C., Hagoort, P., Herzog, H., Sach, M., &
Seitz, R. J. (1997). A PET study of cerebral activation patterns
induced by verb inflection. Neuroimage, 5, S548.
Jacobsen, W. (1979). Noun and verb in Nootkan. Heritage
Record, 4, 83–155.
Joanisse, M. F., & Seidenberg, M. S. (1999). Impairments in
verb morphology after brain injury: A connectionist model.
Proceedings of the National Academy of Sciences, U.S.A., 96,
7592–7597.
Keel, J. C., Smith, M. J., & Wassermann, E. M. (2001). A
safety screening questionnaire for transcranial magnetic
stimulation. Clinical Neurophysiology, 112, 720.
Kellenbach, M. L., Wijers, A. A., Hovius, M., Mulder, J., &
Mulder, G. (2002). Neural differentiation of lexico-syntactic
categories or semantic features? Event-related potential
evidence for both. Journal of Cognitive Neuroscience, 14,
561–577.
Kobayashi, M., Hutchinson, S., Theoret, H., Schlaug, G., &
Pascual-Leone, A. (2004). A repetitive TMS of the motor
cortex improves ipsilateral sequential simple finger
movements. Neurology, 62, 91–98.
Kucera, H., & Francis, W. N. (1982). Computational analysis
of present-day American English. Providence: Brown
University Press.
Laiacona, M., & Caramazza, A. (2004). The noun/verb
dissociation in language production: Varieties of causes.
Cognitive Neuropsychology, 21, 103–123.
Longe, O., Randall, B., Stamatakis, E. A., & Tyler, L. K.
(2007). Grammatical categories in the brain: The role
of morphological structure. Cerebral Cortex, 17, 1812–1820.
Luria, A. R., & Tsvetkova, L. S. (1967). Les troubles de la
resolution de problemes. Analyse neuropsychologique.
Paris: Gauthier-Villars.
Luzzatti, C., Raggi, R., Zonca, G., Pistarini, C., Contardi, A.,
& Pinna, G. D. (2002). Verb–noun double dissociation in
aphasic lexical impairments: The role of word frequency and
imageability. Brain and Language, 81, 432–444.
Mansur, C. G., Fregni, F., Boggio, P. S., Riberto, M.,
Gallucci-Neto, J., Santos, C. M., et al. (2005). A sham
stimulation-controlled trial of rTMS of the unaffected
hemisphere in stroke patients. Neurology, 64, 1802–1804.
Druks, J. (2002). Verbs and nouns: A review of the literature.
Marantz, A. (1997). No escape from syntax: Don’t try
Journal of Neurolingusics, 15, 289–315.
Evans, N. (2000). Word classes in the world’s languages. In
G. Booij, C. Lehmann, & J. Mugdan (Eds.), Morphology: A
handbook on the inflection and word formation. Berlin:
Mouton de Gruyter.
Forster, K. I., & Forster, J. C. (2003). DMDX: A windows
display program with millisecond accuracy. Behavioural
Research Methods, Instruments & Computers, 35, 116–124.
Halle, M., & Marantz, A. (1993). Distributed morphology
and the pieces of inflection. In K. Hale & S. J. Keyser (Eds.),
morphological analysis in the privacy of your own Lexicon.
Penn Working Papers in Linguistics, 4, 201–225.
Marslen-Wilson, W., & Tyler, L. (1997). Dissociating types of
mental computation. Nature, 5, 592–594.
McClelland, J. L., & Patterson, K. (2002). Rules or connections
in past-tense inflections: What does the evidence rule out?
Trends in Cognitive Sciences, 6, 465–472.
Menn, L., & Obler, L. K. (Eds.) (1990). Agrammatic aphasia:
A cross-language narrative sourcebook. Philadelphia:
J. Benjamins.
Cappelletti et al.
719
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
f
/
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Miceli, G., Silveri, M. C., Villa, G., & Caramazza, A. (1984). On
the basis for the agrammatic’s difficulty in producing main
verbs. Cortex, 20, 207–220.
Miozzo, A., Soardi, M., & Cappa, S. F. (1994). Pure anomia with
spared action naming due to a left temporal lesion.
Neuropsychologia, 32, 1101–1109.
Miozzo, M. (2003). On the processing of regular and irregular
forms of verbs and nouns: Evidence from neuropsychology.
Cognition, 87, 101–127.
Naeser, M. A., Martin, P. I., Nicholas, M., Baker, E. H., Seekins,
H., Helm-Estabrooks, N., et al. (2005a). Improved picture
naming in chronic aphasia after TMS to part of right Broca’s
area: An open-protocol study. Brain and Language, 93,
95–105.
Naeser, M. A., Martin, P. I., Nicholas, M., Baker, E. H.,
Seekins, H., Helm-Estabrooks, N., et al. (2005b). Improved
naming after TMS treatments in a chronic, global aphasia
patient—Case report. Neurocase, 11, 182–193.
Neininger, B., & Pulvermu¨ller, F. (1997). The right
hemisphere’s role in action word processing: A double
case study. Neurocase, 7, 303–317.
O’Grady, W. (1997). Syntactic development. Chicago:
University of Chicago Press.
Perani, D., Cappa, S., Schnur, T., Tettamanti, M., Collina, S.,
Rosa, M., et al. (1999). The neural correlates of verb and
noun processing: A PET study. Brain, 122, 2337–2344.
Pulvermu¨ller, F. (1999). Words in the brain’s language.
Behavioral and Brain Sciences, 22, 253–279.
the left frontal cortex. Journal of Cognitive Neuroscience,
13, 713–720.
Shapiro, K. A., Shelton, J., & Caramazza, A. (2000). Grammatical
class in lexical production and morphological processing:
Evidence from a case of fluent aphasia. Cognitive
Neuropsychology, 17, 665–682.
Silveri, M., & di Betta, A. (1997). Noun–verb dissociations in
brain-damaged patients: Further evidence. Neurocase, 3,
477–488.
Tettamanti, M., Buccino, G., Saccuman, M. C., Gallese, V.,
Dann, M., Scifo, M. P., et al. (2005). Listening to action
related sentences activates fronto-parietal motor circuits.
Journal of Cognitive Neuroscience, 17, 273–281.
Teyler, T., Roemer, R. R., Harrison, T., & Thompson, R. (1973).
Human scalp recorded evoked potential correlates of
linguistic stimuli. Bulletin of the Psychonomic Society, 1,
333–334.
Theoret, H., Kobayashi, M., Valero-Cabre, A., & Pascual-Leone,
A. (2003). Exploring paradoxical functional facilitation
with TMS. Supplement to Clinical Neurophysiology, 56,
211–219.
Tranel, D., Adolphs, R., Damasio, H., & Damasio, A. R. (2001). A
neural basis for the retrieval of words for actions. Cognitive
Neuropsychology, 18, 655–670.
Tsapkini, K., Jarema, G., & Kehayia, E. (2002). A morphological
processing deficit in verbs but not in nouns: A case study in a
highly inflected language. Journal of Neurolinguistics, 15,
265–288.
Pulvermu¨ller, F. (2001). Brain reflections of words and their
Tyler, L., Bright, P., Fletcher, P., & Stamatakis, E. A. (2004).
meaning. Trends in Cognitive Sciences, 5, 517–524.
Pulvermu¨ller, F. (2005). Brain mechanisms linking language
and action. Nature Reviews Neuroscience, 6, 576–582.
Robertson, E. M., Theoret, H., & Pascual-Leone, A. (2003).
The problems solved and created by transcranial magnetic
stimulation. Journal of Cognitive Neuroscience, 15,
948–960.
Robins, R. (1952). Noun and verb in universal grammar.
Language, 28, 289–298.
Sakai, K. L., Noguchi, Y., Takeuchi, T., & Watanabe, E. (2002).
Selective priming of syntactic processing by event-related
transcranial magnetic stimulation of Broca’s area. Neuron,
35, 1177–1182.
Sapir, E. (1921). Language. New York: Hartcourt, Brace
and World.
Shapiro, K. A., & Caramazza, A. (2003a). Looming a loom:
Evidence for independent access to grammatical and
phonological properties in verb retrieval. Journal of
Neurolinguistics, 16, 85–111.
Shapiro, K., & Caramazza, A. (2003b). Grammatical processing
of nouns and verbs in left frontal cortex? Neuropsychologia,
41, 1189–1198.
Shapiro, K., & Caramazza, A. (2003c). The representation of
grammatical categories in the brain. Trends in Cognitive
Sciences, 7, 201–206.
Shapiro, K. A., Moo, L. R., & Caramazza, A. (2006). Cortical
signatures of noun and verb production. Proceedings of the
National Academy of Sciences, U.S.A., 103, 1644–1649.
Shapiro, K. A., Mottaghy, F. M., Schiller, N. O., Poeppel, T. D.,
Fluss, M. O., Muller, H. W., et al. (2005). Dissociating
neural correlates for nouns and verbs. Neuroimage, 24,
1058–1067.
Shapiro, K. A., Pascual-Leone, A., Mottaghy, F. M., Gangitano,
M., & Caramazza, A. (2001). Grammatical distinctions in
Neural processing of nouns and verbs: The role of
inflectional morphology. Neuropsychologia, 42,
512–523.
Tyler, L. K., Russell, R., Fadili, J., & Moss, H. E. (2001). The
neural representation of nouns and verbs: PET studies.
Brain, 124, 1619–1634.
Ullman, M., Corkin, S., Coppola, M., Hickok, G., Growdon, J.,
Koroshetz, W., et al. (1997). A neural dissociation
within language: Evidence that the mental dictionary
is part of declarative memory, and that grammatical
rules are processed by the procedural system. Journal
of Cognitive Neuroscience, 9, 266–276.
Ullman, M. T., Bergida, R., & O’Craven, K. (1997). Distinct fMRI
activation patterns for regular and irregular past tense.
Neuroimage, 5, S549.
Walsh, V., & Pascual-Leone, A. (2003). Transcranial magnetic
stimulation. A neurochronometrics of mind. Cambridge:
MIT Press.
Warburton, E., Wise, R., Price, C., Weiller, C., Hadar, U.,
Ramsay, S., et al. (1996). Noun and verb retrieval by normal
subjects: Studies with PET. Brain, 119, 159–179.
Wasserman, E. M. (1998). Risk and safety of repetitive
transcranial magnetic stimulation: Report and
suggested guidelines from the International workshop
on the safety of Repetitive Transcranial Magnetic
Stimulation, June 5–7, 1996. Electroencelophagraphy
and Clinical Neurophysiology, 108, 1–16.
Zingeser, L., & Berndt, R. (1988). Grammatical class and
context effects in a case of pure anomia: Implications for
models of language production. Cognitive Neuropsychology,
5, 473–516.
Zingeser, L., & Berndt, R. (1990). Retrieval of nouns and
verbs in agrammatism and anomia. Brain and Language,
29, 14–32.
720
Journal of Cognitive Neuroscience
Volume 20, Number 4
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
4
e
-
7
p
0
d
7
f
1
/
9
2
3
0
6
/
8
4
3
/
7
7
o
0
c
7
n
/
1
2
0
7
0
5
8
9
4
2
2
0
6
0
4
/
5
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
0
4
0
5
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
/
.
f
t
.
.
o
n
1
8
M
a
y
2
0
2
1