Pre-output Language Monitoring in Sign Production
Stephanie K. Riès1,2, Linda Nadalet1, Soren Mickelsen1, Megan Mott1,
Katherine J. Midgley1, Phillip J. Holcomb1,2, and Karen Emmorey1,2
Abstrait
■ A domain-general monitoring mechanism is proposed to be
involved in overt speech monitoring. This mechanism is re-
flected in a medial frontal component, the error negativity
(Ne), present in both errors and correct trials (Ne-like wave)
but larger in errors than correct trials. In overt speech produc-
tion, this negativity starts to rise before speech onset and is
therefore associated with inner speech monitoring. Ici, we in-
vestigate whether the same monitoring mechanism is involved in
sign language production. Twenty deaf signers (American Sign
Language [ASL] dominant) et 16 hearing signers (English dom-
inant) participated in a picture–word interference paradigm in
ASL. As in previous studies, ASL naming latencies were measured
using the keyboard release time. EEG results revealed a medial
frontal negativity peaking within 15 msec after keyboard release
in the deaf signers. This negativity was larger in errors than cor-
rect trials, as previously observed in spoken language produc-
tion. No clear negativity was present in the hearing signers. Dans
addition, the slope of the Ne was correlated with ASL proficiency
(measured by the ASL Sentence Repetition Task) across signers.
Our results indicate that a similar medial frontal mechanism is
engaged in preoutput language monitoring in sign and spoken
language production. These results suggest that the monitoring
mechanism reflected by the Ne/Ne-like wave is independent of
output modality (c'est à dire., spoken or signed) and likely monitors pre-
articulatory representations of language. Differences between
groups may be linked to several factors including differences in
language proficiency or more variable lexical access to motor
programming latencies for hearing than deaf signers. ■
INTRODUCTION
Healthy adult speakers only err about once every 1000
words under natural speech conditions (Levelt, 1999). Le
same has been shown in sign language (Hohenberger,
Happ, & Leuninger, 2002). Such highly efficient behavior
is enabled in part by language monitoring processes,
which are responsible for controlling our linguistic pro-
duction as it is being output. Although the cognitive
and neuronal mechanisms underlying speech monitoring
have received some attention in the past few years, ces
mechanisms have been understudied in sign language
production.
Various cognitive models of language monitoring have
been proposed (Nozari, Dell, & Schwartz, 2011; Postma
& Oomen, 2005; Postma, 2000), and all of these models
make a distinction between monitoring processes in-
volved before versus after language output. These moni-
toring processes have been referred to as the inner and
outer loops of speech monitoring, respectivement. The role
of the inner loop is to monitor internal linguistic rep-
resentations, whereas the outer loop relies on auditory
feedback (in overt speech). Differences can be expected
between sign and speech monitoring concerning the im-
plementation of the outer loop as the auditory system
should not be engaged when signing, and there is
1San Diego State University, 2Université de Californie, San Diego
© 2020 Massachusetts Institute of Technology
evidence that signers do not rely on visual feedback
when monitoring sign production for errors (Emmorey,
Bosworth, & Kraljic, 2009; Emmorey, Gertsberg, Korpics, &
Wright, 2009; Emmorey, Korpics, & Petronio, 2009).
Cependant, it is unclear whether or not differences between
the inner loop monitoring mechanisms engaged in sign
versus speech production would be observed. The way in-
ternal linguistic representations are monitored has been
conceptualized in different ways. En particulier, speech mon-
itoring models differ in terms of whether the inner loop
depends on the language comprehension system (Levelt,
Roelofs, & Meyer, 1999) or on the language production
système (Nozari et al., 2011; Postma, 2000). They also dif-
fer in terms of whether or not a domain-general monitor-
ing mechanism is involved in inner speech monitoring
(Acheson & Hagoort, 2014; Riès, Janssen, Dufau, Alario,
& Burle, 2011) and whether or not this domain-general
monitoring mechanism is conflict-based (Zheng, Roelofs,
Farquhar, & Lemhöfer, 2018). Donc, clarifying whether
or not a similar brain mechanism is involved in sign lan-
guage monitoring before signs are actually produced is a
necessary step in furthering the understanding of sign
language monitoring. De plus, finding a similar inner
loop mechanism in sign and speech production would
be of interest in furthering the understanding of language
monitoring more generally, as this would suggest the
representations involved are not dependent of language
output modality.
Journal des neurosciences cognitives 32:6, pp. 1079–1091
https://doi.org/10.1162/jocn_a_01542
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
6
e
1
–
0
p
7
d
9
F
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
un
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
F
t
/
.
o
n
0
5
M.
un
oui
2
0
2
1
Similar brain regions have been shown to be engaged
in sign and in speech production at the single word level
but also at the phrase and narrative levels, including left
temporal and left inferior frontal regions (Blanco-Elorrieta,
Kastner, Emmorey, & Pylkkänen, 2018; Emmorey, Mehta, &
Grabowski, 2007; Brun, Guillemin, Hosey, & Varga, 2001).
This common neuronal substrate has been argued to un-
derlie common semantic, lexical, and syntactic properties
between sign and spoken languages. Cependant, sign and
spoken languages differ in several ways, particularly in
how they are perceived (visual vs. auditory) and in the
modality of output during production (manual and facial
movements vs. a phonatory system). Such differences
could arguably lead to differences in how sign and spoken
languages are monitored. Par exemple, somatosensory
feedback in speech monitoring is linked to the movement
of speech articulators (Tremblay, Shiller, & Ostry, 2003) mais
will include manual and facial movements in sign language
monitoring (Emmorey, McCullough, Mehta, & Grabowski,
2014; Emmorey, Bosworth, et coll., 2009).
En effet, brain regions known to be associated with
speech perception, in particular, the superior temporal
cortex, have been found to be sensitive to manipulations
affecting speech output monitoring (more specifically the
outer loop mentioned above) such as speech distortion or
delayed auditory feedback (Tourville, Reilly, & Guenther,
2008; Fu et al., 2006; Hashimoto & Sakai, 2003; McGuire,
Silbersweig, & Frith, 1996). These results have been inter-
preted as supporting the idea that overt language output
monitoring occurs through the language comprehension
système, as proposed by Levelt (1983).
In sign language, linguistic output is visual and not audi-
tory. Ainsi, this could imply that visual brain areas could
be involved in sign language monitoring, mirroring the in-
volvement of the auditory system in overt speech monitoring.
Cependant, the monitoring of sign language production has
been proposed to rely more heavily on proprioceptive feed-
back than visual feedback (Emmorey, Bosworth, et coll., 2009).
In agreement with this proposal, several studies have reported
that parietal regions and not visual regions are more active
in sign than in speech production (Emmorey et al., 2007,
2014), including the left supramarginal gyrus and the left
superior parietal lobule. Activation of the superior parietal
lobule, in particular, has been associated with proprio-
ceptive monitoring during motoric output (Emmorey, Mehta,
McCullough, & Grabowski, 2016). Enfin, activation of
the superior temporal cortex has not been associated with
language output monitoring in sign language production,
which is likely due to the different modality of output in
sign and speech production.
Speech monitoring has also been shown to rely on the
activation of medial frontal regions, such as the ACC and
the SMA (Christoffels, Formisano, & Schiller, 2007). Acti-
vation of the ACC has been shown to be associated with con-
flict monitoring in and outside language (Piai, Roelofs,
Acheson, & Takashima, 2013; Barch, Plus courageux, Sabb, & Noll,
2000; Botvinick, Nystrom, Fissell, Carter, & Cohen, 1999).
Donc, speech monitoring has been proposed to
depend not only on the perception of one’s own speech
through brain mechanisms associated with speech com-
prehension (see Indefrey, 2011, for a review) mais aussi
through the action of a domain-general action monitoring
mechanism in medial frontal cortex (Christoffels et al.,
2007).
EEG studies of speech monitoring have focused on a
component referred to as the error negativity (Ne) ou
error-related negativity (Zheng et al., 2018; Riès, Fraser,
McMahon, & de Zubicaray, 2015; Acheson & Hagoort,
2014; Riès, Xie, Haaland, Dronkers, & Knight, 2013;
Riès et al., 2011; Ganushchak & Schiller, 2008un, 2008b;
Masaki, Tanaka, Takasawa, & Yamazaki, 2001). This com-
ponent has a frontocentral distribution (maximal at elec-
trode FCz) and peaks within 100 msec following vocal
début. This component was initially only reported fol-
lowing erroneous utterances and was therefore in-
terpreted as reflecting an error detection mechanism
(Masaki et al., 2001). Cependant, this component was more
recently also found in correct trials only with a smaller am-
plitude, suggesting it reflects a monitoring mechanism op-
erating before error detection (Riès et al., 2011). À cause de
the similar topography, time course, and origin of this com-
ponent in correct trials and in errors, the component in cor-
rect trials has been referred to as the Ne-like wave (Bonini
et coll., 2014; Roger, Bénar, Vidal, Hasbroucq, & Burle, 2010;
Vidal, Burle, Bonnet, Grapperon, & Hasbroucq, 2003). Dans
speech production, this medial frontal monitoring mecha-
nism also starts to be engaged before the onset of verbal
réponses, suggesting that it reflects the monitoring of in-
ner speech (c'est à dire., the inner loop mentioned above) rather
than that of overt speech production (Riès et al., 2011,
2015; Riès, Xie, et coll., 2013). Combining a neuropsycholog-
ical and computational approach, Nozari and colleagues
have suggested that accurate speech production relies
more heavily on this domain-general monitoring mecha-
nism operating before speech onset than on the speech
comprehension-based monitor (Nozari et al., 2011), lequel
would be hosted in the superior temporal cortex. Si
or not a domain-general monitor in medial frontal cortex is
also engaged in sign monitoring before signs are produced
is unknown.
Several arguments suggest that the medial frontal cor-
tex should be similarly engaged in sign and in spoken lan-
guage monitoring. One of these arguments is that the
representations that this monitoring mechanism oper-
ates on are likely to be prearticulatory. Evidence for this
proposal comes from the finding that the amplitude of
the Ne is modulated by variables that have been tied to
stages that precede articulation such as semantic related-
ness, lexical frequency, or interference from another lan-
guage in bilinguals (Riès et al., 2015; Ganushchak &
Schiller, 2008un, 2008b, 2009). Such internal representa-
tions are likely to be commonly engaged in spoken and
sign language production. Another argument is the
domain-general nature of the monitoring mechanism
1080
Journal des neurosciences cognitives
Volume 32, Nombre 6
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
6
e
1
–
0
p
7
d
9
F
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
un
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
.
/
t
F
o
n
0
5
M.
un
oui
2
0
2
1
hosted in the medial frontal cortex. En effet, the Ne and Ne-
like waves have been shown to be present in overt speech
production, in typing (Pinet & Nozari, 2020; Kalfaoğlu,
Stafford, & Milne, 2018), but also in other actions such as
in manual button-press tasks (Roger et al., 2010; Burle,
Roger, Allain, Vidal, & Hasbroucq, 2008; Vidal et al., 2003;
Vidal, Hasbroucq, Grapperon, & Bonnet, 2000). The source
of the Ne and Ne-like waves has been localized to the medial
frontal cortex and, in particular, the ACC (Debener et al.,
2005; Dehaene, Posner, & Tucker, 1994) and/or the SMA,
as shown through intracranial investigations with depth
electrodes inserted in the medial frontal cortex (Bonini
et coll., 2014). These brain regions are associated with action
monitoring generally and are therefore also likely to be
engaged in sign language monitoring.
This Study
Dans cette étude, we hypothesized that the domain-general
monitoring mechanism hosted in the medial frontal cortex
and reflected in the Ne and Ne-like wave is similarly engaged
during signing and speaking. This study used a picture-
naming task and scalp EEG to examine the error (Ne) et
error-like (Ne-like) negativities time-locked to the initiation
of sign production (as measured through manual key re-
lease, as in Emmorey, Petrich, & Gollan, 2013). En particulier,
we used data gathered during a picture–word interference
(PWI) paradigm, which has been shown to elicit more errors
than simple picture naming. In the PWI task, used exten-
sively in the field of psycholinguistics, pictures are pre-
ceded by or presented with superimposed distractor
words (par exemple., Bürki, 2017; Roelofs & Piai, 2015, 2017; Piai,
Roelofs, Jensen, Schoffelen, & Bonnefond, 2014; Piai,
Roelofs, & Schriefers, 2014; Piai et al., 2013; Costa,
Alario, & Caramazza, 2005). In the semantic version of
the task (used here), the distractor words can be se-
mantically related to the picture (par exemple., picture of a dog,
distractor word: “cat”) or unrelated (par exemple., picture of a
dog, distractor word: “chair”). Typically in this task, nam-
ing the picture takes longer and error rates are higher in
the semantically related, compared with the unrelated
condition, although the presence of this effect appears
to depend on the language input and output modalities
(Emmorey, Mott, Meade, Holcomb, & Midgely, under re-
voir; Giezen & Emmorey, 2016). Nevertheless, error rates
are expected to be higher in this task than in simpler
picture naming, which made this paradigm of interest
for this study.
We tested both deaf and hearing signers as they named
pictures by signing the picture names in American Sign
Language (ASL). En outre, we investigated whether or
not ASL proficiency, as measured through the ASL
Sentence Repetition Task (ASL-SRT), had an effect on the
medial frontal monitoring mechanism. Language proficiency
has been found to be a potential factor affecting this mech-
anism, as suggested by several studies (Ganushchak &
Schiller, 2009; Sebastian-Gallés, Rodríguez-Fornells, de
Diego-Balaguer, & Díaz, 2006). We note, cependant, que
these studies used button-press responses and not overt
speech. Donc, more direct investigations involving
overt language production are needed. We had reasons
to believe ASL proficiency may be different between the
deaf and hearing group because, although the hearing sign-
ers participating in these experiments are selected to be
highly proficient in ASL, their use of ASL in their everyday
lives is typically less frequent than that of deaf signers (voir
Paludneviciene, Hauser, Daggett, & Kurz, 2012), and hear-
ing signers are also surrounded by spoken English in the
environment.
Finding similar components in sign language pro-
duction would provide strong evidence for the universal
nature of inner language monitoring. En effet, it would
suggest that the mechanism reflected by the Ne and
Ne-like waves is involved in inner language monitoring
irrespective of the language output modality. This would
constitute a further argument in support of the idea that
the representations monitored by this medial frontal
monitoring mechanism are prearticulatory.
MÉTHODES
The data analyzed in this study was initially collected for
another study focusing on the effect of the PWI manipu-
lation on ERPs time-locked to stimulus presentation
(Emmorey et al., under review). Dans cette étude, we focused
on the Ne and Ne-like wave time-locked to keyboard re-
lease, which reflected the point at which sign production
began. There were not enough errors per participant to
investigate the effect of the PWI manipulation on the Ne,
so we averaged across conditions to increase the number
of trials per component.
Participants
A total of 26 deaf signers (15 femmes, âge moyen = 34
années, SD = 9 années) et 21 hearing signers (17 femmes,
âge moyen = 36 années, SD = 10 années) participated in this
étude. They were recruited through the San Diego area
(California) and gave informed consent in accordance
with the San Diego State University institutional review
board. They received monetary compensation for their
temps. All had normal or corrected-to-normal vision and
no history of neurological impairment. Thirteen deaf par-
ticipants and five hearing participants were excluded
from our analyses because they had less than five error
trials remaining after artifact rejection or because they
did not follow instructions. Our analyses were therefore
conducted on the remaining 11 deaf (eight women,
âge moyen = 35 années, SD = 12 années) et 15 hearing par-
ticipants (12 femmes, âge moyen = 37 années, SD = 12
années). Of these remaining participants, 7 of the 11 deaf
participants acquired ASL from birth from their deaf
signing families, et 4 acquired ASL in early childhood
(before age of 6 années). Of the included 15 hearing
Riès et al.
1081
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
6
e
1
–
0
p
7
d
9
F
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
un
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
.
F
/
t
o
n
0
5
M.
un
oui
2
0
2
1
participants, 4 acquired ASL from birth from their deaf
signing families and 11 acquired ASL later, at a mean age
de 15 années (SD = 7 années), 7 were interpreters, and all had
been signing for at least 7 years before the experiment
(mean = 24 années, SD = 10 années). All included participants
were right-handed. English proficiency was objectively mea-
sured using the Peabody Individual Achievement Test
(PIAT) reading comprehension subtest (Markwardt, 1998)
and a spelling test from Andrews and Hersch (2010). ASL
proficiency was objectively measured using the extended
(35 sentence) version of the ASL-SRT (Supalla, Hauser, &
Bavelier, 2014). In this task, participants view an ASL sen-
tence and then sign back what was just viewed. Sentence
complexity and length increased after each trial. The ASL-
SRT task has been shown to differentiate deaf from hearing
users of sign language, as well as native from nonnative
users (Supalla et al., 2014).
Materials and Design
The stimuli consisted of 200 words representing com-
mon nouns and 100 pictures (c'est à dire., line drawings) selected
from various sources (Snodgrass & Vanderwart, 1980),
presented on a white background. Name agreement in
English for the pictures was high (average = 90%, SD =
14.4).1 The average length in letters for the words was
5.05 letters (SD = 1.87). The words were presented in cap-
ital letters in Arial font (size 60 in height by 30 in width).
Fifty of the pictures were presented in an identity condi-
tion (par exemple., the word “house” followed by a picture of a
maison), and the other 50 were presented in the semanti-
cally related condition (par exemple., the word “paper” followed by
a picture of scissors). All of the pictures were also pre-
sented in an unrelated condition (par exemple., the word “ring”
followed by a picture of scissors). Donc, each picture
appeared twice (once in a related condition and once in
an unrelated condition). Lists were constructed so that
they only contained each target item once, half (50 pic-
photos) in the unrelated condition, one fourth or 25 pic-
tures in the semantically related condition, et un
fourth or 25 pictures in the identity condition. Lists were
counterbalanced across participants so that any target item
was presented first in the related condition to half of the
participants and first in the unrelated condition to the rest
of the participants.
the naming onset. They were asked to name the pictures
as quickly and as accurately as possible and ignore the
words. They were each given one practice round that
consisted of six trials (these stimuli were not used in
the experiment). During the practice, they were in-
structed to blink during the breaks between stimuli and
to minimize facial movements while signing to avoid pro-
ducing artifacts in the EEG recordings.
Each trial of the experiment began with a fixation cross
that was presented in the center of the screen. The cross
remained on the screen until the participant placed their
hands on the spacebar. The word was then presented for
200 msec and was replaced by the picture that was pre-
sented for 2000 msec. Participants were asked to produce
the sign corresponding to the picture name as quickly
and as accurately as possible, without hesitating. After
signing the picture name, the participants were asked
to place their hands back on the spacebar. The fixation
cross replaced the picture after 2000 msec, and the next
trial would start only after the participant placed their
hands back on the spacebar. Participants were video-
recorded during the experiment so that their sign accu-
racy could be analyzed off-line. The task was self-paced
by use of the space bar. Participants were instructed to
rest during the fixation periods before placing their
hands back on the keyboard. The whole experiment
lasted around 20 min with some variability in time de-
pending on how many breaks each participant took.
EEG Recordings
EEG was continually recorded from a 32-channel tin elec-
trode cap (Electro-Cap International, Inc.; using a 10–20
electrode placement). The EEG signal was amplified by a
SynAmpsRT amplifier (Neuroscan-Compumedics), et
data were collected by Curry Data Acquisition software
at a sampling rate of 500 Hz with a band-pass filter of
DC to 100 Hz. To monitor for eye blinks and movements,
electrodes were placed under the left eye and on the
outer canthus of to the right eye. The reference electrode
was placed on the left mastoid, and an electrode was
placed on the right mastoid for monitoring differential
mastoid activity. Impedances were measured before the
experiment started and kept below 2.5 kΩ.
Procedure
The stimuli were presented within a 2° × 3° visual angle
at the center of an LCD computer screen at a viewing
distance of approximately 150 cm from the participant’s
eyes. This ensured that participants did not have to make
large eye movements to fully perceive the stimuli. The par-
ticipants were seated in a dimly lit, sound-attenuated room
and were asked to hold down the spacebar of a keyboard
and only lift their hands when they were ready to pro-
duce the sign corresponding to the picture, marking
Data Processing
Behavioral Data Processing
RTs were defined as the time separating the picture onset
from the release of the spacebar to initiate sign produc-
tion. The accuracy of sign production was determined
off-line by visual inspection of the video recordings from
the experiment, and all hesitations were discarded from
the analysis. Accuracy and hesitation coding was done by
two raters, a deaf native signer and a hearing highly pro-
ficient ASL signer. Correct trials were those in which an
1082
Journal des neurosciences cognitives
Volume 32, Nombre 6
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
6
e
1
–
0
p
7
d
9
F
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
un
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
/
F
t
.
o
n
0
5
M.
un
oui
2
0
2
1
accurate sign was produced at the time of keyboard release
with no hesitations. Error trials were trials in which the par-
ticipant produced an off-target sign (par exemple., LION instead of
TIGER). Trials in which the participant produced an UM
sign or where there was a perceptible pause between the
keyboard liftoff and the initiation of the sign were excluded
from analysis (see Emmorey, Petrich, & Gollan, 2012). Trials
in which the participant did not respond were also ex-
cluded from the behavioral and EEG analyses.
EEG Processing
After acquisition, vertical eye movements (c'est à dire., eye blinks)
were removed using independent component analysis as
implemented in EEGLAB (Delorme & Makeig, 2004).
Additional artifacts caused by EMG activity associated with
facial movements were reduced using a blind source sepa-
ration algorithm based on canonical correlation analysis
(De Clercq, Vergult, Vanrumste, Van Paesschen, & Van Huffel,
2006), previously adapted to speech production (De Vos
et coll., 2010), and as used successfully in previous studies in-
vestigating speech monitoring processes (Riès et al., 2011,
2015; Riès, Xie, et coll., 2013). Enfin, any remaining artifacts
were removed through manual inspection in Brain Vision
Analyzer (Brain Products). Laplacian transformation (c'est à dire.,
current source density, estimation), as implemented in
Brain Vision Analyzer, was applied to each participant’s aver-
ages and on the grand averages (as in Riès et al., 2011, 2015;
Riès, Janssen, Burle, & Alario, 2013; Riès, Xie, et coll., 2013;
degree of spline = 3, Legendre polynomial = 15° maxi-
mum). We assumed a radius of 10 cm for the sphere repre-
senting the head. The resulting unit was μV/cm2. Grand
averages were created for correct and incorrect trials in both
the deaf and hearing groups for the participants with more
than five error trials remaining after artifact rejection.
Data Analysis
Proficiency scores for English and ASL as measured by
the above-listed tests were compared between groups
using two-tailed Student t tests.
Behavioral data were analyzed using linear (for RTs) et
generalized mixed-effects models (for accuracy rates). Nous
tested for main effects of Accuracy and Group (deaf vs. hear-
ing) and the interaction between Accuracy × Group on RTs
and tested for a main effect of Group on accuracy rates and
controlled for random effects of subjects and items. p Values
were obtained using Type II analyses of deviance tables pro-
viding the Wald χ2 tests for the fixed effects in the mixed-
effects models. For all models, we report the Wald χ2 values
and p values from the analyses of deviance tables as well as
raw β estimates (βraw), 95% confidence intervals around the-
se β estimates (CI), standard errors (SE), t values for RTs, et
Wald Z and associated p values for accuracy rates.
EEG data were analyzed using two types of EEG mea-
sures following methods described in previous studies
(Riès, Janssen, et coll., 2013; Riès, Xie, et coll., 2013; Riès
et coll., 2011). The first measure was the slope of the wave-
forms on a 150-msec time window preceding the key
release (the onset of sign production). To find the slope,
a linear regression was fitted to the data, and then non-
parametric exact Wilcoxon signed-rank tests were used to
compare the slopes to 0 for both the errors and correct
trials in the deaf and hearing group as the number of
error trials was low and the data could not be assumed to
be normally distributed. The second measure was peak-to-
peak amplitude (c'est à dire., the difference between the amplitude
of two consecutive peaks of activity). Peak-to-peak ampli-
tude was calculated by first determining the peak latencies
of the Ne and Ne-like wave as well as the preceding positive
peak (also referred to as the start of the rise of the nega-
tivité) on the by-participant averages. Latencies were
measured on smoothed data to minimize the impact of
background noise (the length of the smoothing window
était 40 msec) and within 100-msec time windows centered
around the latency of the peak on grand averages. Alors, pour
each participant, the surface area was calculated between
the waveform and the baseline on a 50-msec time window
centered around each peak latency as measured in each
participant’s average waveform. Enfin, the difference be-
tween the surface areas measured around the Ne or Ne-like
wave and around the preceding positivity was considered
the peak-to-peak amplitude and is hence independent from
the baseline. Encore, nonparametric exact Wilcoxon signed-
rank one-sided tests (Wilcoxon t tests) were used to com-
pare peak-to-peak amplitudes in errors versus correct trials
because the measures were based on few error trials and
the normality of the data could not be assumed (as in
Riès, Xie, et coll., 2013; Riès et al., 2011). The use of one-
sided tests was justified as the direction of the difference
was expected based on several preceding studies in lan-
guage (Riès et al., 2011, 2015; Riès, Xie, et coll., 2013) et
outside language (Vidal et al., 2000, 2003). For each test,
we report the W statistic for Wilcoxon signed-rank tests,
general Z statistic, associated p value, and effect size r value.
En outre, the effect of Group (deaf vs. hearing) on these
differences between correct and error trials were tested
using an ANOVA.
Enfin, we tested for a correlation between the slope
of the Ne and Ne-like wave, and the ASL proficiency score
as measured with the ASL-SRT, using Spearman’s rank
correlation coefficient ρ. We report the rho correlation
coefficients, S, and associated p values.
All statistical analyses were conducted using R (R Core
Team, 2014).
RÉSULTATS
Behavioral Results
Language Proficiency
English. Raw PIAT scores for the deaf group ranged
depuis 42 à 99 (M = 79, SD = 13), and spelling scores
ranged from 62 à 80 (M = 73, SD = 6). PIAT scores from
Riès et al.
1083
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
6
e
1
–
0
p
7
d
9
F
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
un
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
t
.
F
/
o
n
0
5
M.
un
oui
2
0
2
1
a significant main effect of Accuracy on RTs ( Wald χ2 =
21.08, p < .001), but no effect of Group ( Wald χ2 = 0.71,
p = .398), and no interaction between Group × Accuracy
( Wald χ2 = 0.40, p = .530). RTs were shorter in correct
than in incorrect trials (βraw = −120.20, CI [−191.81,
−48.59], SE = 36.54, t = −3.29). The median error
rate was 5.6% (IQR = 4.6–12.4%), and there was no
effect of Group on accuracy rates ( Wald χ2 = 0.91, p =
.341; βraw = −0.29, CI [−0.88, 0.30], SE = 0.30, Z = 0.34,
p = .341). Mean RT, median error rates, and number of
errors are reported in Table 1 (see Tables S1 and S2 for
full fixed-effect results from the mixed-effects models2).
On average, 76% (σ = 17%) of correct trials and 74%
(σ = 20%) of errors were left after artifact rejection.
Figure 1. ASL proficiency scores as measured through the ASL-SRT in
deaf (blue) and hearing signers (orange).
EEG Results
Deaf Signers
the hearing group ranged from 67 to 99 (M = 92, SD = 7),
and spelling test scores ranged from 62 to 85 (M = 78,
SD = 5). There was a marginal difference between groups
on the spelling test, t(21.06) = −2.0352, p = .055, and a
significant difference between groups on the PIAT,
t(14.56) = −3.016, p = .0089: The hearing participants
showed higher performance on these tests of English
proficiency than the deaf participants.
ASL. ASL-SRT scores ranged from 12 to 27 (M = 22, SD =
5) for deaf signers and ranged from 6 to 24 (M = 14, SD =
5) for hearing signers. An ASL-SRT score was not collected
from one of the hearing participants because of her famil-
iarity with the test. There was a significant difference in
ASL-SRT scores between the deaf and the hearing signers,
t(22.82) = 3.60, p = .0015: The deaf signers had higher
ASL-SRT scores than the hearing signers. Figure 1 provides
an illustration of the distribution of the ASL-SRT scores.
RTs and Error Rates
The average RT was 742 msec (σ = 270 msec) for correct
trials and 848 msec (σ = 376 msec) for errors. There was
We observed negativities for correct and incorrect trials.
The negativity began to rise on average 209 msec (SD =
64 msec) before keyboard release in correct trials and
−212 msec (SD = 60 msec) in errors. There was no statis-
tical difference between the latency of the positive peak in
errors and correct trials, t(10) < 1; W = 30.5, Z = −.22,
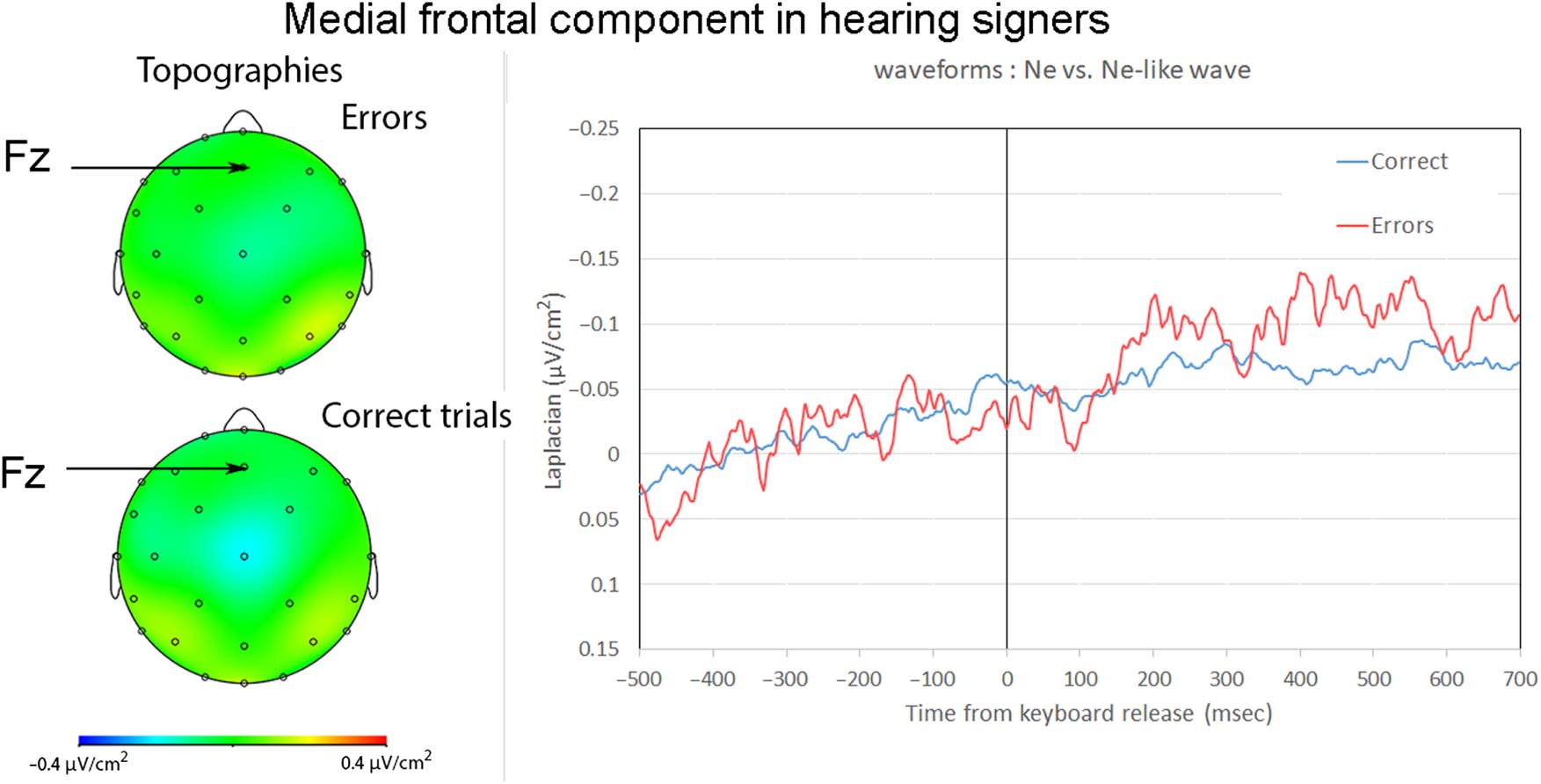
p = .852, r = .04. The negativity reached its maximum on
average 13 msec (SD = 31 msec) before keyboard release
in correct trials and 32 msec (SD = 59 msec) after key-
board release in errors. The negativity peaked significantly
later in errors than correct trials, t(10) = 3.10, p = .011;
W = 63, Z = −2.67, p = .005, r = .56. The negativities
reached their maximum at frontocentral electrodes, just
posterior to Fz and anterior to Cz (see Figure 2; the used
montage did not include electrode FCz, which is typically
used to study the Ne and Ne-like waves). Slopes measured
from −150 to 0 msec were significantly different from 0 in
correct trials, t(10) = −2.31, p = .022; W = 7, Z = −2.60,
p = .009, r = .55, and incorrect trials, t(10) = −2.52, p =
.015; W = 7, Z = −2.60, p = .009, r = .55. The amplitude
of the negativity for incorrect trials was significantly larger
than for correct trials, t(10) = 4.03, p = .001; W = 66,
Z = −3.49, p < .001, r = .74.
Table 1. Mean RTs per Group (Deaf vs. Hearing) for Errors and Correct Trials with Standard Deviations around the Mean (σ),
Median Error Rates, Range, and Median (M ) Number of Errors per Group with Interquartile Ranges (IQR)
Deaf
Hearing
Correct
Errors
Correct
Errors
Mean RTs
688 msec
(σ = 242 msec)
796 msec
(σ = 394 msec)
781 msec
(σ = 290 msec)
886 msec
(σ = 372 msec)
Median error rates
5.08% (IQR = 4.12–11.81%)
6.70% (IQR = 4.94–11.52%)
Range and median number of errors
Range = 5–26, M = 8, IQR = 7–10
Range = 5–33, M = 11, IQR = 9–13
1084
Journal of Cognitive Neuroscience
Volume 32, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
6
e
1
-
0
p
7
d
9
f
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
a
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
f
/
.
o
n
0
5
M
a
y
2
0
2
1
Figure 2. Ne and Ne-like wave in deaf participants. On the left, topography of Ne at the peak latency. On the right, waveforms for the Ne
(in red, errors) and Ne-like waves (in blue, correct trials) time-locked to keyboard release (in msec). The Ne and Ne-like wave reach their
maxima around keyboard release time and the Ne is marginally larger than the Ne-like wave.
Hearing Signers
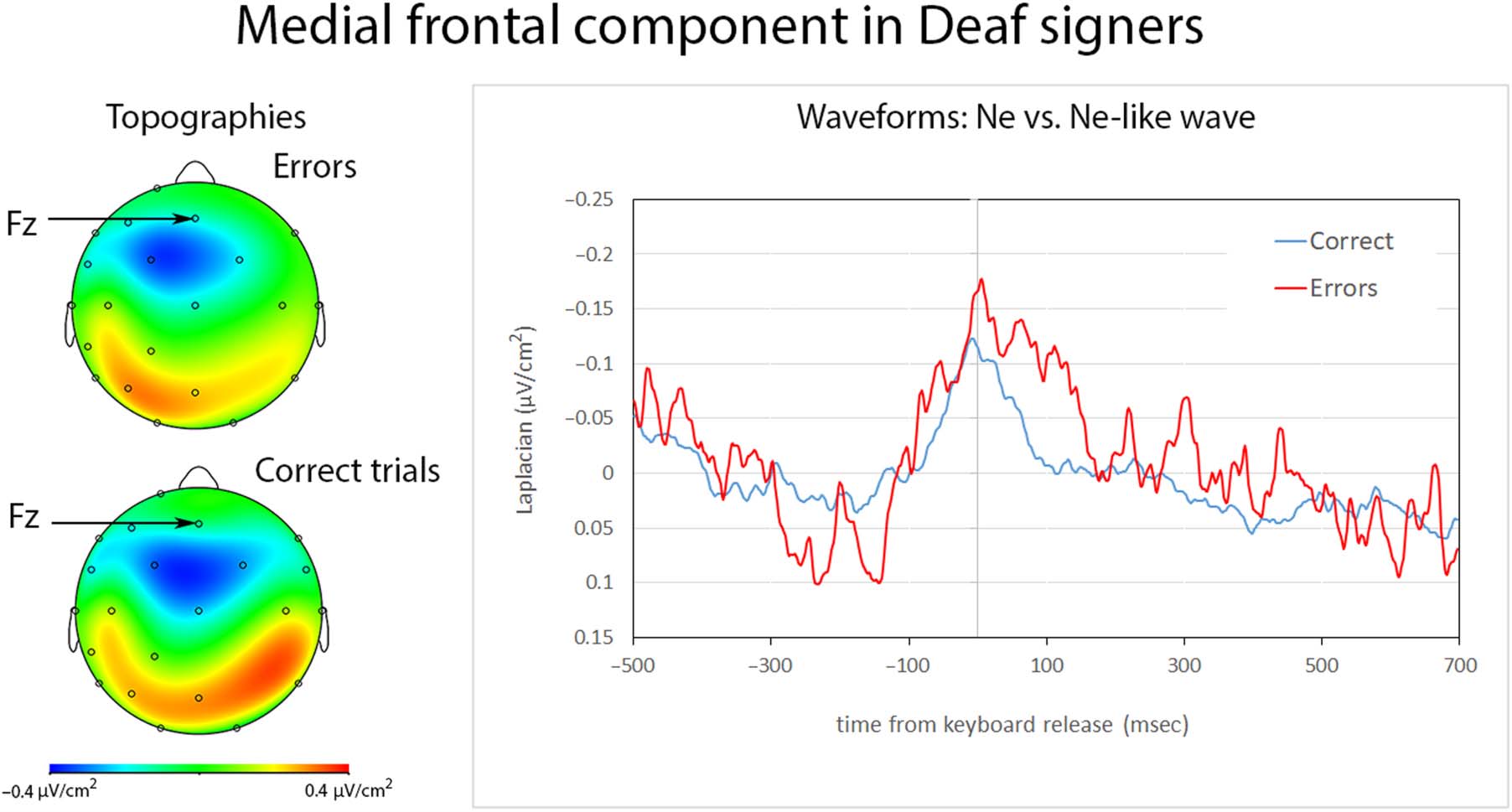
Slopes measured from −150 to 0 msec were significantly
different from zero in correct trials, t(14) = −2.14, p =
.025 (W = 24, Z = −2.06, p = .039, r = .38). Slopes were
not significantly different from zero on the same time
window for errors, t(14) < 1 (W = 56, Z = −.53, p =
.596, r = .10). This indicated there was no reliable Ne
in errors for the hearing signers at the same recording
site as for deaf signers (i.e., Fz). We note, however, that
a negativity seemed present at Cz (see Figure 3), al-
though this activity was much smaller than the one re-
ported in deaf participants (see Figure 2; the same
scale was used in Figures 2 and 3).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
6
e
1
-
0
p
7
d
9
f
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
a
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
.
f
t
o
n
0
5
M
a
y
2
0
2
1
Figure 3. Ne and Ne-like wave in hearing signers. Left: Topography around the peak of the negativity in errors and correct trials. Right: Waveforms
for the Ne (in red, errors) and Ne-like waves (in blue, correct trials) time-locked to the keyboard release (in msec). For ease of comparison, the
same scales were used as for deaf signers (Figure 2).
Riès et al.
1085
frontal negativity in errors versus correct trials was larger
in the deaf than in the hearing signers (Figure 4). When
tested independently, we however did not find a signifi-
cant difference between the slope in errors versus cor-
rect trials in deaf signers (W = 19, Z = −1.55, p =
.120, r = .33), although we note that the amplitudes
were significantly different as reported in the previous
section. In the hearing group, there was no indication
of a difference between errors and correct trials (W =
76, Z = −.08, p = .932, r = .02).
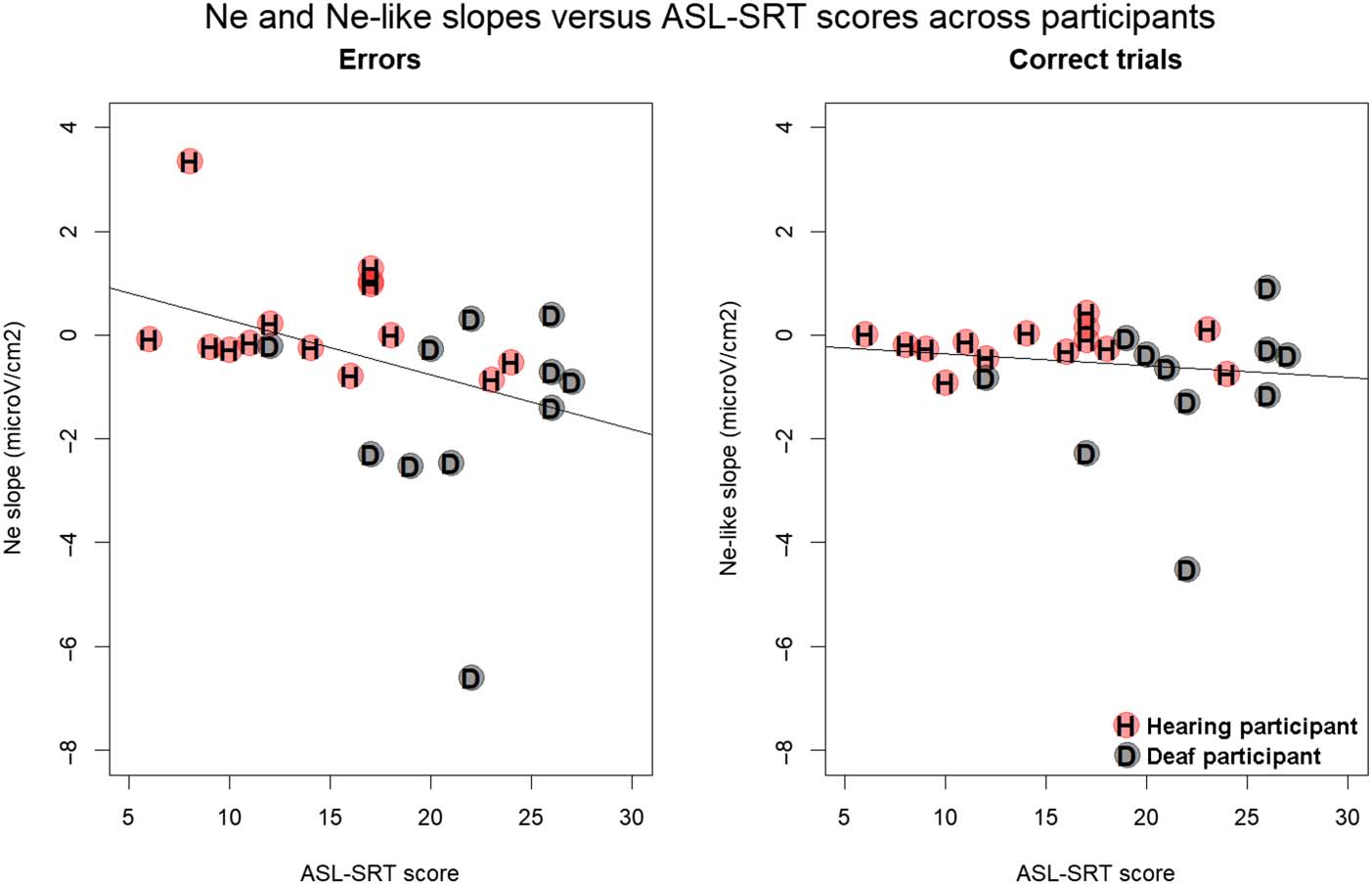
Because the effect of hearing status may have been
confounded with ASL proficiency, we tested for a corre-
lation between the slope of the Ne and Ne-like wave (as
calculated between −150 msec and keyboard release
time) and ASL proficiency scores on the ASL-SRT. The
slope of the Ne (in errors) was negatively correlated with
ASL proficiency scores across deaf and hearing participants
(rho = −0.41, S = 3677.3, p = .039), meaning that steeper
slopes for the Ne (the steeper the slope, the more nega-
tive) were associated with higher ASL proficiency (see
Figure 5). No significant correlation was observed between
ASL proficiency and the slope of the Ne-like wave (in cor-
rect trials, rho = −0.16, S = 3019.3, p = .441).
DISCUSSION
Our results showed that a medial frontal component is
present in sign production for both correct responses
and errors when deaf signers name pictures in ASL
(time-locked to keyboard release). This component is
larger in errors than in correct trials. In addition, the
slope of the Ne was correlated with proficiency levels
in ASL across hearing and deaf signers. The slope was
steeper with increasing proficiency, as measured through
the ASL-SRT. In hearing signers, this medial frontal
Figure 4. Histogram plotting the mean slope of the Ne and Ne-like
wave (with standard deviations from the mean) in deaf and hearing
signers.
Deaf versus Hearing Signers and Effect of
ASL Proficiency
We tested for effects of Accuracy, Group, and ASL-SRT
score on the slope of the medial frontal component
(the data of the participant for whom we did not have
an ASL-SRT score was excluded). We found a significant
effect of Group, F(1, 21) = 7.14, p = .014, and an inter-
action between Group × Accuracy, F(1, 21) = 4.35, p =
.050. There was no significant main effect of Accuracy,
F(1, 21) < 1; ASL-SRT score, F(1, 21) < 1; nor any inter-
action between Group × ASL-SRT score, F(1, 21) < 1, and
Accuracy × ASL-SRT score, F(1, 21) = 1.21, p = .284; and
no three-way interaction, F(1, 21) < 1.
The significant interaction between Group × Accuracy
suggests the difference between the slope of the medial
Figure 5. Slopes of the Ne
(left) and Ne-like wave (right)
versus ASL-SRT scores across
participants. Hearing
participants are indicated in red
and with the letter “H,” and deaf
participants are indicated in
gray with the letter “D.”
1086
Journal of Cognitive Neuroscience
Volume 32, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
6
e
1
-
0
p
7
d
9
f
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
a
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
t
/
o
n
0
5
M
a
y
2
0
2
1
component was not larger in errors than correct trials
and was absent in errors at the same recording site as
for deaf signers, but was present in correct trials.
Frontomedial Monitoring Mechanism in
Sign Language Production
The first important result is that a medial frontal compo-
nent is present in sign language production in both cor-
rect (Ne-like wave) and error trials (Ne). This component
had a frontocentral distribution, started to rise before
naming onset (as indexed by keyboard release), and
peaked just after keyboard release in errors and slightly
earlier in correct trials. In addition, it had a larger ampli-
tude in errors than in correct trials. This result suggests
that a similar medial frontal mechanism is engaged in pre-
output language monitoring in both signed and spoken
language production. Indeed, a similar medial frontal neg-
ativity was previously reported in overt speech production
(Zheng et al., 2018; Riès et al., 2011, 2015; Acheson &
Hagoort, 2014; Riès, Xie, et al., 2013). Because this activity
starts to rise before vocal onset, it was associated with
inner speech monitoring. The topography and time course
of this component in sign language production is very sim-
ilar to that observed in overt speech. In overt speech, the
Ne was reported to peak between 30 and 40 msec after vo-
cal onset (Riès, Xie, et al., 2013; Riès et al., 2011), which
corresponds to what we observed in this study (mean la-
tency of negative peak: 32 msec after keyboard release).
In addition, the preceding positive peak was found to pre-
cede the onset of language output (speech and sign) across
studies even though there were some differences in the
latency of this preceding positive peak: 166 msec (SD =
80 msec) prevocal onset in Riès, Xie, et al. (2013), 46 msec
(SD = 52 msec) prevocal onset in Riès et al. (2011), and
212 msec (SD = 60 msec) before keyboard release in this
study. One possible explanation for these differences in
latencies for the positive peak could be that the task used
here was more complex as it required participants to
ignore the distractor word and to release the spacebar
to start signing. The relatively small difference in am-
plitude between the Ne and Ne-like wave in this study
would be in agreement with this interpretation. Indeed,
the difference in amplitude between the Ne and Ne-like
wave has been shown to be smaller with increasing task
difficulty (leading to higher error rates; e.g., due to de-
creased response certainty or in time pressure situations;
Ganushchak & Schiller, 2006, 2009; Sebastian-Gallés
et al., 2006). We note that the RTs were on average lon-
ger (742 msec, SD = 270 msec, for correct trials) and that
the error rates were higher (median = 5.6%, IQR = 4.6–
12.4%) in this study compared with Experiment 2 of Riès
et al. (2011), which used simple overt picture naming
(mean RT for correct trials = 651 msec, SD = 72 msec;
mean error rate = 1.31%, SD = 0.96%). In addition, in
this study, the requirement to release the spacebar be-
fore signing constitutes an additional step in response
programming that may have caused the increased delay
between the latency of the peak of the Ne and that of the
preceding negativity. However, the similarity in the la-
tency of the negative peak and the fact that it starts to
rise before articulation onset, as well as the similar topog-
raphies associated with this component in speech and in
sign language production, suggest that the medial frontal
component we report here is similar to the medial frontal
component reported in overt speech (e.g., Riès, Xie,
et al., 2013; Riès et al., 2011). This suggests that this me-
dial frontal component is involved in the inner loop of
language output monitoring irrespective of the output
modality, which would be in line with the idea that the
representations that are monitored by this mechanism
are prearticulatory.
In addition to specifying the role of the medial frontal mon-
itoring mechanism in language production, our results also
shed light on sign language monitoring more specifi-
cally. Indeed, based on the perceptual loop theory of self-
monitoring (Levelt, 1983, 1989), previous studies had
investigated the role of visual feedback in sign language
monitoring. Emmorey, Gertsberg, et al. (2009) found that
preventing visual feedback with a blindfold had little
impact on sign production (i.e., there is no Lombard effect
for sign language). Emmorey, Bosworth, et al. (2009) showed
that blurring or completely masking visual feedback did
not alter how well novel signs were learned, suggesting
that signers do not rely on visual feedback to fine-tune
articulation during learning. In fact, production perfor-
mance of hearing nonsigners was slightly worse with than
without visual feedback. This led the authors to suggest
that sign language monitoring may rely more heavily on
proprioceptive feedback than on visual feedback (see also
Emmorey, Korpics, et al., 2009). What the present results
suggest is that a medial frontal monitoring mechanism
may also be involved in sign language monitoring and that
this monitoring mechanism is engaged before propriocep-
tive feedback is available (i.e., before beginning to move the
hand(s) to sign). Evidence for this claim comes from the
time course of the Ne and Ne-like waves, which start to rise
before sign production onset (i.e., before key release). In
addition, Allain, Hasbroucq, Burle, Grapperon, and Vidal
(2004) reported Ne and Ne-like waves in a completely deaf-
ferented patient. This rare clinical case was tested using a
two-response RT task and a go/no-go task and showed the
expected Ne and Ne-like wave patterns in both tasks. Our
results therefore indicate yet another similarity in the pro-
cessing of sign and speech production and imply that cur-
rent models of speech monitoring should be adapted to
sign language production (Nozari et al., 2011; Postma &
Oomen, 2005; Levelt, 1983).
Difference between Deaf and Hearing Signers
At the group level, no clear Ne was visible in the hearing
signers at the same recording site as for deaf signers.
Although the hearing signers were highly proficient in
Riès et al.
1087
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
6
e
1
-
0
p
7
d
9
f
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
a
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
f
.
o
n
0
5
M
a
y
2
0
2
1
ASL (many worked as interpreters), their scores on the
ASL-SRT were significantly lower than for the deaf signers
we tested. Therefore, we tested for an effect of profi-
ciency on the slope of the Ne and found that the slope
of the Ne was negatively correlated with ASL proficiency
scores across deaf and hearing signers, meaning that the
higher the ASL-SRT score, the more negative-going the
slope of the Ne was. Nevertheless, we note that there
was no significant effect of ASL proficiency when tested
along with the effect of group. It therefore appeared that
group and ASL proficiency were confounded in our
study. Consequently, more detailed examinations of the
possible effect of ASL proficiency on the medial frontal
monitoring mechanism are needed in future studies.
Previous studies investigating the Ne and/or Ne-like wave
in overt speech monitoring have been performed in bilin-
guals (Acheson, Ganushchak, Christoffels, & Hagoort, 2012;
Ganushchak & Schiller, 2009; Sebastian-Gallés et al., 2006).
In particular, Ganushchak and Schiller (2009) compared
German–Dutch bilinguals to Dutch monolingual partici-
pants as they performed a phoneme monitoring “go/no-
go” task (i.e., they were asked to press a button if the
Dutch name of the presented picture contained a specific
phoneme) under time pressure versus not. In the time
pressure condition, the stimuli were presented for a shorter
duration than in the control condition, and this duration
was adapted on an individual basis. The results showed
differential effects of time pressure on the amplitude of
the Ne (referred to as the error-related negativity in their
study) as a function of group. German–Dutch bilinguals,
who were performing the task in their nonnative language,
showed a larger Ne in the time pressure condition than in
the control condition, and Dutch monolingual speakers
showed the reverse effect. Importantly, the bilingual indi-
viduals tested in this study were not balanced bilinguals
and had lower proficiency in the language in which they
were tested (i.e., Dutch) than in their native language
(i.e., German). Although the task we used was very differ-
ent from the task used in Ganushchak and Schiller (2009),
their results suggest that the Ne may be sensitive to lan-
guage proficiency. Interestingly and similarly to our results,
the mean amplitude of the Ne in the control condition,
which is more comparable to the setup of our study, ap-
peared to be lower in the German–Dutch bilinguals than
in the Dutch monolinguals, although this difference was
not directly tested.
Relatedly, Sebastian-Gallés et al. (2006) compared
Spanish–Catalan bilinguals who were Spanish versus
Catalan dominant in a lexical decision task in Catalan.
For Catalan-dominant bilinguals, they observed the ex-
pected pattern of a larger negativity in errors than correct
trials. However, for Spanish-dominant bilinguals, the
amplitude of the negativity was not larger in errors than
in correct trials. These results suggest language domi-
nance is an important variable influencing inner speech
monitoring abilities. However, we did not test for an effect
of language dominance independently from language
proficiency. Typically, English is considered the dominant
language for hearing signers because English is the
language of schooling and the surrounding community,
whereas ASL is considered the dominant language for deaf
signers (for discussion, see Emmorey, Giezen, & Gollan,
2016). Interestingly, Sebastian-Gallés et al. (2006) reported
a negativity in correct trials (our Ne-like wave) in both
groups of bilinguals, which was larger when lexical deci-
sion was more difficult (i.e., for nonword trials vs. word
trials). This finding is in line with our results as we also
found a significant Ne-like wave in the hearing signers,
even though the Ne was not statistically reliable at the
group level.
Previous reports have also shown modulations of the Ne-
like wave outside language as a function of response uncer-
tainty (Pailing & Segalowitz, 2004) and the accuracy of the
following trial (Allain, Carbonnell, Falkenstein, Burle, &
Vidal, 2004). In particular, the amplitude of the Ne-like
wave has been shown to increase with response uncertainty,
whereas the amplitude of the Ne has been shown to de-
crease with response uncertainty (Pailing & Segalowitz,
2004). Hence, one possible interpretation of our results
could be that the hearing signers experienced greater
response uncertainty compared with deaf signers. This
hypothesis would also be in line with the proposal that
hearing signers are less aware of their sign errors com-
pared with deaf signers, as suggested by Nicodemus and
Emmorey (2015). Another (possibly related) reason for
the lack of an Ne in hearing signers could be linked to a
time alignment issue with the event used to mark sign pro-
duction onset, namely the keyboard release. Even though
we carefully rejected all trials containing a perceptible pause
between the keyboard release time and the onset of the sign
(e.g., when the dominant hand reached the target location
of the sign; see Caselli, Sehyr, Cohen-Goldberg, and
Emmorey (2017) for a detailed description of how sign on-
sets are determined), naming onset may not have been as
closely aligned to keyboard release time in hearing signers
as compared with deaf signers. That is, hearing signers may
have been more likely to prematurely release the spacebar
before they had completely encoded the sign for articula-
tion. This could explain why a later, though not strongly re-
liable, negativity was observed in the subgroup of proficient
hearing signers. Indeed, for these signers, the sign onset it-
self, occurring after keyboard release, might be a better
marker to use for the observation of an Ne. Future studies
are needed to clarify this issue.
Conclusion
In summary, our study reports for the first time the pres-
ence of a medial frontal negativity associated with inner
language output monitoring in sign language production.
The presence of this negativity in sign language produc-
tion strongly suggests that a similar medial frontal mecha-
nism is involved in language monitoring before response
initiation irrespective of language output modality and
1088
Journal of Cognitive Neuroscience
Volume 32, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
6
e
1
-
0
p
7
d
9
f
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
a
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
suggests the representations that are monitored by this
mechanism are prearticulatory. In addition, in line with
previous studies using phonological monitoring and lexical
decision tasks, our results showed that this mechanism
was modulated by language proficiency in sign language
production, suggesting similar factors affect medial frontal
language output monitoring across modalities.
Acknowledgments
This research was supported by an award to S. K. R. and K. E.
from the Collaborative Pilot Grant Program from the Center for
Cognitive and Clinical Neuroscience at San Diego State
University and by NIH grant DC010997 (K. E.). We are very
thankful to the participants who took part in this study.
Reprint requests should be sent to Stephanie Riès, School of
Speech, Language, and Hearing Sciences, Center for Clinical
and Cognitive Neuroscience, Joint Doctoral Program in
Language and Communicative Disorders, San Diego State
University, 5500 Campanile Drive, San Diego, CA 92182, or
via e-mail: sries@sdsu.edu.
Notes
1. ASL name agreement was available for 61 of the stimuli
(from an ongoing ASL picture naming study in the Emmorey
Lab), and agreement was also high for these stimuli (average =
83.0%, SD = 21.6%).
2. Supplementary material for this paper can be retrieved from
https://lbdl.sdsu.edu/wp-content/uploads/2020/02/Supplementary_
tables.pdf.
REFERENCES
Acheson, D. J., Ganushchak, L. Y., Christoffels, I. K., & Hagoort,
P. (2012). Conflict monitoring in speech production:
Physiological evidence from bilingual picture naming. Brain
and Language, 123, 131–136.
Acheson, D. J., & Hagoort, P. (2014). Twisting tongues to test
for conflict-monitoring in speech production. Frontiers in
Human Neuroscience, 8, 206.
Allain, S., Carbonnell, L., Falkenstein, M., Burle, B., & Vidal, F.
(2004). The modulation of the Ne-like wave on correct
responses foreshadows errors. Neuroscience Letters, 372,
161–166.
Allain, S., Hasbroucq, T., Burle, B., Grapperon, J., & Vidal, F.
(2004). Response monitoring without sensory feedback.
Clinical Neurophysiology, 115, 2014–2020.
Andrews, S., & Hersch, J. (2010). Lexical precision in skilled
readers: Individual differences in masked neighbor priming.
Journal of Experimental Psychology: General, 139,
299–318.
Barch, D. M., Braver, T. S., Sabb, F. W., & Noll, D. C. (2000).
Anterior cingulate and the monitoriing of response conflict:
Evidence from an fMRI study of overt verb generation.
Journal of Cognitive Neuroscience, 12, 298–309.
Blanco-Elorrieta, E., Kastner, I., Emmorey, K., & Pylkkänen, L.
(2018). Shared neural correlates for building phrases in
signed and spoken language. Scientific Reports, 8, 5492.
Bonini, F., Burle, B., Liégeois-Chauvel, C., Régis, J., Chauvel, P.,
& Vidal, F. (2014). Action monitoring and medial frontal
cortex: Leading role of supplementary motor area. Science,
343, 888–891.
Botvinick, M., Nystrom, L. E., Fissell, K., Carter, C. S., & Cohen,
J. D. (1999). Conflict monitoring versus selection-for-action
in anterior cingulate cortex. Nature, 402, 179–181.
Braun, A. R., Guillemin, A., Hosey, L., & Varga, M. (2001). The
neural organization of discourse: An H2
narrative production in English and American sign language.
Brain, 124, 2028–2044.
15O-PET study of
Bürki, A. (2017). Electrophysiological characterization of
facilitation and interference in the picture–word interference
paradigm. Psychophysiology, 54, 1370–1392.
Burle, B., Roger, C., Allain, S., Vidal, F., & Hasbroucq, T.
(2008). Error negativity does not reflect conflict: A
reappraisal of conflict monitoring and anterior cingulate
cortex activity. Journal of Cognitive Neuroscience, 20,
1637–1655.
Caselli, N. K., Sehyr, Z. S., Cohen-Goldberg, A. M., & Emmorey,
K. (2017). ASL-LEX: A lexical database of American sign
language. Behavior Research Methods, 49, 784–801.
Christoffels, I. K., Formisano, E., & Schiller, N. O. (2007). Neural
correlates of verbal feedback processing: An fMRI study
employing overt speech. Human Brain Mapping, 28,
868–879.
Costa, A., Alario, F.-X., & Caramazza, A. (2005). On the
categorical nature of the semantic interference effect in the
picture–word interference paradigm. Psychonomic Bulletin
& Review, 12, 125–131.
Debener, S., Ullsperger, M., Siegel, M., Fiehler, K., von
Cramon, D. Y., & Engel, A. K. (2005). Trial-by-trial coupling
of concurrent electroencephalogram and functional
magnetic resonance imaging identifies the dynamics of
performance monitoring. Journal of Neuroscience, 25,
11730–11737.
De Clercq, W., Vergult, A., Vanrumste, B., Van Paesschen, W., &
Van Huffel, S. (2006). Canonical correlation analysis applied
to remove muscle artifacts from the electroencephalogram.
IEEE Transactions on Biomedical Engineering, 53,
2583–2587.
Dehaene, S., Posner, M. I., & Tucker, D. M. (1994). Localization
of a neural system for error detection and compensation.
Psychological Science, 5, 303–305.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
De Vos, M., Riès, S., Vanderperren, K., Vanrumste, B., Alario,
F.-X., Van Huffel, S., et al. (2010). Removal of muscle artifacts
from EEG recordings of spoken language production.
Neuroinformatics, 8, 135–150.
Emmorey, K., Bosworth, R., & Kraljic, T. (2009). Visual feedback
and self-monitoring of sign language. Journal of Memory
and Language, 61, 398–411.
Emmorey, K., Gertsberg, N., Korpics, F., & Wright, C. E. (2009).
The influence of visual feedback and register changes on sign
language production: A kinematic study with deaf signers.
Applied Psycholinguistics, 30, 187–203.
Emmorey, K., Giezen, M. R., & Gollan, T. H. (2016).
Psycholinguistic, cognitive, and neural implications of
bimodal bilingualism. Bilingualism: Language and
Cognition, 19, 223–242.
Emmorey, K., Korpics, F., & Petronio, K. (2009). The use of
visual feedback during signing: Evidence from signers with
impaired vision. Journal of Deaf Studies and Deaf
Education, 14, 99–104.
Emmorey, K., McCullough, S., Mehta, S., & Grabowski, T. J.
(2014). How sensory-motor systems impact the neural
organization for language: Direct contrasts between
spoken and signed language. Frontiers in Psychology, 5,
484.
Riès et al.
1089
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
6
e
1
-
0
p
7
d
9
f
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
a
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
t
.
f
o
n
0
5
M
a
y
2
0
2
1
Emmorey, K., Mehta, S., & Grabowski, T. J. (2007). The neural
correlates of sign versus word production. Neuroimage, 36,
202–208.
Emmorey, K., Mehta, S., McCullough, S., & Grabowski, T. J.
(2016). The neural circuits recruited for the production of
signs and fingerspelled words. Brain and Language, 160,
30–41.
Emmorey, K., Mott, M., Meade, G., Holcomb, P. J., & Midgely,
K. J. (under review). Bilingual lexical selection without lexical
competition: ERP evidence from bimodal bilinguals and
picture–word interference. Language, Cognition and
Neuroscience.
Emmorey, K., Petrich, J. A. F., & Gollan, T. H. (2012). Bilingual
processing of ASL–English code-blends: The consequences
of accessing two lexical representations simultaneously.
Journal of Memory and Language, 67, 199–210.
Emmorey, K., Petrich, J. A. F., & Gollan, T. H. (2013). Bimodal
bilingualism and the frequency-lag hypothesis. Journal of
Deaf Studies and Deaf Education, 18, 1–11.
Fu, C. H. Y., Vythelingum, G. N., Brammer, M. J., Williams, S. C. R.,
Amaro, E., Jr., Andrew, C. M., et al. (2006). An fMRI study of
verbal self-monitoring: Neural correlates of auditory verbal
feedback. Cerebral Cortex, 16, 969–977.
Ganushchak, L. Y., & Schiller, N. O. (2006). Effects of time
pressure on verbal self-monitoring: An ERP study. Brain
Research, 1125, 104–115.
Ganushchak, L. Y., & Schiller, N. O. (2008a). Motivation and
semantic context affect brain error-monitoring activity: An
event-related brain potentials study. Neuroimage, 39,
395–405.
Ganushchak, L. Y., & Schiller, N. O. (2008b). Brain error-
monitoring activity is affected by semantic relatedness: An
event-related brain potentials study. Journal of Cognitive
Neuroscience, 20, 927–940.
Ganushchak, L. Y., & Schiller, N. O. (2009). Speaking one’s
second language under time pressure: An ERP study on
verbal self-monitoring in German–Dutch bilinguals.
Psychophysiology, 46, 410–419.
Giezen, M. R., & Emmorey, K. (2016). Language co-activation
and lexical selection in bimodal bilinguals: Evidence from
picture–word interference. Bilingualism: Language and
Cognition, 19, 264–276.
Hashimoto, Y., & Sakai, K. L. (2003). Brain activations during
conscious self-monitoring of speech production with delayed
auditory feedback: An fMRI study. Human Brain Mapping,
20, 22–28.
Hohenberger, A., Happ, D., & Leuninger, H. (2002). Modality-
dependent aspects of sign language production: Evidence from
slips of the hands and their repairs in German Sign Language. In
R. P. Meier, K. Cormier, & D. Quinto-Pozos (Eds.), Modality
and structure in signed and spoken languages (pp. 112–142).
Cambridge, UK: Cambridge University Press.
Indefrey, P. (2011). The spatial and temporal signatures of word
production components: A critical update. Frontiers in
Psychology, 2, 255.
Kalfaoğlu, Ç., Stafford, T., & Milne, E. (2018). Frontal theta band
oscillations predict error correction and posterror slowing in
typing. Journal of Experimental Psychology: Human
Perception and Performance, 44, 69–88.
Levelt, W. J. M. (1983). Monitoring and self-repair in speech.
Cognition, 14, 41–104.
Levelt, W. J. M. (1989). Speaking: From intention to
articulation. Cambridge, MA: MIT Press.
Levelt, W. J. M. (1999). Models of word production. Trends in
Cognitive Science, 3, 223–232.
Levelt, W. J. M., Roelofs, A., & Meyer, A. S. (1999). A theory of
lexical access in speech production. Behavioral and Brain
Sciences, 22, 1–38.
Markwardt, F. C. (1998). Peabody Individual Achievement
Test–Revised (Normative update). Circle Pines, MN: American
Guidance Service.
Masaki, H., Tanaka, H., Takasawa, N., & Yamazaki, K. (2001).
Error-related brain potentials elicited by vocal errors.
NeuroReport, 12, 1851–1855.
McGuire, P. K., Silbersweig, D. A., & Frith, C. D. (1996). Functional
neuroanatomy of verbal self-monitoring. Brain, 119, 907–917.
Nicodemus, B., & Emmorey, K. (2015). Directionality in ASL–
English interpreting: Accuracy and articulation quality in L1
and L2. Interpreting, 17, 145–166.
Nozari, N., Dell, G. S., & Schwartz, M. F. (2011). Is
comprehension necessary for error detection? A conflict-
based account of monitoring in speech production. Cognitive
Psychology, 63, 1–33.
Pailing, P. E., & Segalowitz, S. J. (2004). The effects of
uncertainty in error monitoring on associated ERPs. Brain
and Cognition, 56, 215–233.
Paludneviciene, R., Hauser, P. C., Daggett, D., & Kurz, K. B.
(2012). Issues and trends in sign language assessment. In
D. A. Morere & T. E. Allen (Eds.), Assessing literacy in deaf
individuals: Neurocognitive measurement and predictors
(pp. 191–207). New York: Springer.
Piai, V., Roelofs, A., Acheson, D. J., & Takashima, A. (2013).
Attention for speaking: Domain-general control from the
anterior cingulate cortex in spoken word production.
Frontiers in Human Neuroscience, 7, 832.
Piai, V., Roelofs, A., Jensen, O., Schoffelen, J.-M., & Bonnefond,
M. (2014). Distinct patterns of brain activity characterise
lexical activation and competition in spoken word
production. PLoS One, 9, e88674.
Piai, V., Roelofs, A., & Schriefers, H. (2014). Locus of semantic
interference in picture naming: Evidence from dual-task
performance. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 40, 147–165.
Pinet, S., & Nozari, N. (2020). Electrophysiological correlates of
monitoring in typing with and without visual feedback.
Journal of Cognitive Neuroscience, 32, 603–620.
Postma, A. (2000). Detection of errors during speech production:
A review of speech monitoring models. Cognition, 77, 97–132.
Postma, A., & Oomen, C. E. (2005). Critical issues in speech
monitoring. In R. Hartsuiker, Y. Bastiaanse, A. Postma, &
F. Wijnen (Eds.), Phonological encoding and monitoring in
normal and pathological speech (pp. 157–166). Hove, UK:
Psychology Press.
R Core Team. (2014). R: A language and environment for
statistical computing. Vienna: R Foundation for Statistical
Computing. Retrieved from www.R-project.org/.
Riès, S. K., Fraser, D., McMahon, K. L., & de Zubicaray, G. I.
(2015). Early and late electrophysiological effects of
distractor frequency in picture naming: Reconciling input
and outpu accounts. Journal of Cognitive Neuroscience, 27,
1936–1947.
Riès, S. K., Janssen, N., Burle, B., & Alario, F.-X. (2013).
Response-locked brain dynamics of word production. PLoS
One, 8, e58197.
Riès, S. K., Janssen, N., Dufau, S., Alario, F.-X., & Burle, B. (2011).
General-purpose monitoring during speech production.
Journal of Cognitive Neuroscience, 23, 1419–1436.
Riès, S. K., Xie, K., Haaland, K. Y., Dronkers, N. F., & Knight,
R. T. (2013). Role of the lateral prefrontal cortex in speech
monitoring. Frontiers in Human Neuroscience, 7, 703.
Roelofs, A., & Piai, V. (2015). Aspects of competition in word
production: Reply to Mahon and Navarrete. Cortex, 64,
420–424.
Roelofs, A., & Piai, V. (2017). Distributional analysis of semantic
interference in picture naming. Quarterly Journal of
Experimental Psychology, 70, 782–792.
1090
Journal of Cognitive Neuroscience
Volume 32, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
6
e
1
-
0
p
7
d
9
f
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
a
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
f
/
t
o
n
0
5
M
a
y
2
0
2
1
Roger, C., Bénar, C. G., Vidal, F., Hasbroucq, T., & Burle, B. (2010).
Rostral cingulate zone and correct response monitoring: ICA
and source localization evidences for the unicity of correct-
and error-negativities. Neuroimage, 51, 391–403.
Sebastian-Gallés, N., Rodríguez-Fornells, A., de Diego-Balaguer,
R., & Díaz, B. (2006). First- and second-language phonological
representations in the mental lexicon. Journal of Cognitive
Neuroscience, 18, 1277–1291.
Snodgrass, J. G., & Vanderwart, M. (1980). A standardized set of
260 pictures: Norms for name agreement, image agreement,
familiarity, and visual complexity. Journal of Experimental
Psychology: Human Learning and Memory, 6, 174–215.
Supalla, T., Hauser, P. C., & Bavelier, D. (2014). Reproducing
American sign language sentences: Cognitive scaffolding in
working memory. Frontiers in Psychology, 5, 859.
Tourville, J. A., Reilly, K. J., & Guenther, F. H. (2008). Neural
mechanisms underlying auditory feedback control of speech.
Neuroimage, 39, 1429–1443.
Tremblay, S., Shiller, D. M., & Ostry, D. J. (2003). Somatosensory
basis of speech production. Nature, 423, 866–869.
Vidal, F., Burle, B., Bonnet, M., Grapperon, J., & Hasbroucq,
T. (2003). Error negativity on correct trials: A
reexamination of available data. Biological Psychology,
64, 265–282.
Vidal, F., Hasbroucq, T., Grapperon, J., & Bonnet, M. (2000).
Is the “error negativity” specific to errors? Biological
Psychology, 51, 109–128.
Zheng, X., Roelofs, A., Farquhar, J., & Lemhöfer, K. (2018).
Monitoring of language selection errors in switching: Not all
about conflict. PLoS One, 13, e0200397.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
6
e
1
-
0
p
7
d
9
f
2
/
0
3
1
2
3
/
4
6
7
/
9
1
0
o
7
c
9
n
_
/
a
1
_
8
0
6
1
1
5
5
4
8
2
0
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
4
0
2
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
f
.
t
o
n
0
5
M
a
y
2
0
2
1
Riès et al.
1091