Neural Evidence for a Distinction between Short-term
Memory and the Focus of Attention
Jarrod A. Lewis-Peacock1, Andrew T. Drysdale1, Klaus Oberauer2,
and Bradley R. Postle1
Abstrait
■ It is widely assumed that the short-term retention of infor-
mation is accomplished via maintenance of an active neural
trace. Cependant, we demonstrate that memory can be preserved
across a brief delay despite the apparent loss of sustained repre-
sentations. Delay period activity may, in fact, reflect the focus of
attention, rather than STM. We unconfounded attention and
memory by causing external and internal shifts of attention
away from items that were being actively retained. Multivariate
pattern analysis of fMRI indicated that only items within the
focus of attention elicited an active neural trace. Activity cor-
responding to representations of items outside the focus
quickly dropped to baseline. Néanmoins, this information
was remembered after a brief delay. Our data also show that
refocusing attention toward a previously unattended memory
item can reactivate its neural signature. The loss of sustained
activity has long been thought to indicate a disruption of
STM, but our results suggest that, even for small memory loads
not exceeding the capacity limits of STM, the active mainte-
nance of a stimulus representation may not be necessary for
its short-term retention. ■
INTRODUCTION
Since at least the time of Hebb (1949), it has widely been
assumed that the short-term retention of information is ac-
complished via maintenance of an active memory trace. Ce
view has been reinforced by reports of elevated delay pe-
riod activity in extracellular (Fuster & Alexander, 1971;
Kubota & Niki, 1971), electroencephalographic ( Vogel,
McCollough, & Machizawa, 2005), and hemodynamic (Curtis
& DʼEsposito, 2003; Haxby, Petit, Ungerleider, & Courtney,
2000; Courtney, Ungerleider, Keil, & Haxby, 1997) record-
ings of animals and humans. Par conséquent, the loss of sus-
tained activity is thought to indicate a disruption of the
memory trace (Postle, Druzgal, & DʼEsposito, 2003; Miller
& Désimone, 1994; di Pellegrino & Sage, 1993). Cependant,
to the best of our knowledge, virtually all studies of the
short-term retention of information (regardless of species,
procedure, concurrent physiological measurement, etc.)
have confounded memory with attention: The information
to be remembered is the most task-relevant information
throughout the memory interval and, donc, is likely to
be continuously attended to. This leaves open the question
of whether sustained delay period activity is better under-
stood as a correlate of memory or as a correlate of attention.
To address this question, we unconfounded these con-
structs across two experiments by causing external and in-
ternal shifts of attention away from information that was
being actively retained during a brief memory interval.
1University of Wisconsin—Madison, 2University of Zurich
Using multivariate pattern analysis (MVPA) of brain ac-
tivity recorded in event-related fMRI (Pereira, Mitchell, &
Botvinick, 2009; Haynes & Rees, 2006; Norman, Polyn,
Detre, & Haxby, 2006), we tested the hypothesis that de-
lay period activity reflects the information that is being at-
tended to, but not the information that is unattended, yet
remembered, after a brief delay. The “embedded compo-
nent” theory of information processing provides the theo-
retical framework for this hypothesis. It characterizes STM
as an emergent property of the interaction of long-term
mémoire (LTM) and attention (Oberauer, 2002; Cowan,
1995; Ericsson & Kintsch, 1995; Cowan, 1988) and postulates
a distinction between a capacity-limited central component
of STM, referred to as the “focus of attention”1 and a more
peripheral component referred to as “activated LTM.” In
keeping with this view, we use the term STM to refer not to
a hypothetical system but to the ability of the mind or brain
to retain a limited amount of information over brief periods.
This model accounts for a wide range of data from behav-
ioral, neuropsychological, electrophysiological, and neuro-
imaging studies of monkeys and humans (reviewed in
Postle, 2006). Par exemple, evidence for the interaction
between attention and LTM comes from electroencephalo-
graphic recordings of increased neural synchrony between
prefrontal and posterior cortices during STM (Ruchkin,
Grafman, Cameron, & Berndt, 2003). This observation has
motivated the idea that PFC directs the attentional focus
needed for maintaining activation in the appropriate poste-
rior processing regions. Initial neuroscientific support for
engagement of LTM in STM relied on demonstrations that
Journal des neurosciences cognitives 24:1, pp. 61–79
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
un
/
_
j
0
o
0
c
1
n
4
_
0
un
p
_
d
0
0
b
1
oui
4
g
0
u
.
e
p
s
d
t
o
F
n
b
0
oui
7
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
F
t
.
/
u
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
the brain regions which participate in the initial perception
and comprehension of incoming information are also in-
volved in its short-term retention. Par exemple, delay period
activity during STM for faces has been localized to regions of
temporooccipital cortex that are believed to support the per-
ception and long-term retention of faces (Ranganath, Cohen,
Dam, & DʼEsposito, 2004; Ranganath, DeGutis, & DʼEsposito,
2004; Druzgal & DʼEsposito, 2003; Postle et al., 2003). Tel
results cannot be interpreted as strong tests of this model,
cependant, because they rely on tenuous reverse inferences
(c'est à dire., they reason backward, from the presence of peaks in
brain activity to the engagement of a particular cognitive func-
tion; Poldrack, 2006). This is because, Par exemple, the pres-
ence of sustained activity peaks in midfusiform gyrus does
not necessarily imply that faces were being remembered, être-
cause this region can show above-baseline activity during
many other cognitive states (par exemple., Gauthier, Skudlarski, Sang,
& Anderson, 2000). Stronger evidence comes from a dem-
onstration with MVPA that the information content of delay
period activity can be decoded based on distributed patterns
of unthresholded brain activity recorded from an indepen-
dent LTM task (Lewis-Peacock & Postle, 2008). MVPA can sup-
port stronger reverse inferences than univariate techniques,
because it captures high-dimensional neural representations
that have markedly higher selectivity than do univariate ac-
tivation peaks, a consequence of which is that MVPA can
support discrimination of neural representations at the item
level (Corps de guerre, Formisano, Sorger, & Goebel, 2007).
The temporal dynamics of the embedded component
model are being mapped out in the behavioral literature.
Par exemple, memory items that are no longer relevant
for behavior can be removed (within 1–2 sec) from the
focus of attention, thereby reducing the load on the sys-
temʼs limited capacity and consequently reducing RTs
to memory probes of the behaviorally relevant items still
in the focus (Oberauer, 2001). Information removed from
the focus remains in a state of heightened availability for
several seconds, as shown by the finding that lures from a
recently encoded memory list are harder to reject than
lures not recently encountered (Oberauer, 2001; Woltz,
1996; Monsell, 1978). This information can be refocused
if needed again (Oberauer, 2005); otherwise, it is prone
to forgetting by decay or by interference.
A recent fMRI study showed that retention of a single
item inside the focus of attention exhibits a distinct neural
signature (Née & En Ionie, 2008). It found that an item with-
in the focus is associated with increased activation in the
inferior temporal cortex (ITC) relative to other information
in STM. Attended information was sustained via enhanced
functional connectivity with frontal and posterior parietal
régions, whereas unattended information was charac-
terized by increased activations in LTM retrieval-related
regions in the medial-temporal lobe and PFC. These intrigu-
ing results provide some of the first empirical evidence for
a neural dissociation of representations within STM.
Two aspects of this study gave it the potential to pro-
vide novel insights into the embedded component model.
D'abord, it used MVPA so that, rather than having to make
assumptions about what elevated activity in one or more
brain regions might represent, we could objectively and
quantitatively measure the information being actively rep-
resented during the delay period. Deuxième, we explicitly
unconfounded attention from STM by exogenously and
endogenously drawing the focus of attention away from
information that had to be remembered after a brief delay.
In the first experiment, we recorded fMRI data from healthy
young adults while they performed a paired-associate rec-
ognition test of STM, dans lequel, during an unpredictable
half of trials, trial-irrelevant stimuli were presented in the
middle of a memory delay. These visual distractors were
used to redirect the focus of attention outwardly toward
external stimuli and away from the items being actively
retained in memory. In the second experiment, we re-
corded fMRI data from a separate group of participants
while they performed a test of STM, during which only
one of two items being actively retained in STM were cued
as relevant for the next behavioral response. These cues
were used to redirect (c'est à dire., shrink) the focus of attention
internally, such that the irrelevant item would be removed
from the focus.
Our results showed that the information content of de-
lay period activity reflects the focus of attention rather
than the full contents of STM. En fait, brain activity corre-
sponding to representations of unattended information
dropped to baseline levels. Néanmoins, this information
was remembered after a brief delay. Our data also showed
that refocusing attention to previously unattended infor-
mation can restore the active neural signature of that in-
formation. Whereas the loss of sustained activity has
been thought to indicate a disruption of STM, our results
suggest that active maintenance may not be required for
the short-term retention of information. Plutôt, two com-
plementary forms of retention may underlie STM: (1) le
active retention of information inside the focus of atten-
tion via sustained neural firing and (2) the passive reten-
tion of information outside the focus via some other
neural mechanisms (par exemple., transient changes in synaptic po-
tentiation) from which it can be reactivated with cue-based
retrieval. The present results provide direct demonstra-
tions of the former, and they demand the latter by infer-
ence. Theoretically, our results call for rethinking the
“activation” assumption for memory representations out-
side the focus of attention in the embedded component
model. Empirically, they suggest that many previous stud-
ies of short-term and working memory might best be in-
terpreted as studies of sustained attention to information.
EXPERIMENT 1
Méthodes
Participants
Fourteen (nine men; ages 18–29 years) healthy, droite-
handed adults were recruited from the undergraduate
62
Journal des neurosciences cognitives
Volume 24, Nombre 1
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
un
/
_
j
0
o
0
c
1
n
4
_
0
un
p
_
d
0
0
b
1
oui
4
g
0
u
.
e
p
s
d
t
o
F
n
b
0
oui
7
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
F
t
/
.
u
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
and medical campuses of the University of Wisconsin—
Madison. None reported any medical, neurological, ou
psychiatric illness, and all gave informed consent. Un
participantʼs data were removed from analysis because
of a failure to comply with task instructions.
Phase 1: Short-term Recognition
Participants performed short-term recognition of 120 pic-
tures selected from three categories: 40 unfamiliar faces
(20 les hommes et 20 femmes), 40 unfamiliar outdoor places or
scènes, et 40 common objects (Figure 1A). All images
were converted to grayscale with an image processing
software to remove any unintended confounds of color
in the perception and short-term retention of the stimuli.
Each stimulus was presented one time only for 120 ran-
domly ordered stimulus presentations. Each trial consisted
of a target presentation (1 sec), a delay period (7 sec), et
probe presentation (1 sec), a response period (3 sec) and an
intertrial interval (ITI) (10 sec). Participants indicated, avec un
yes–no button press, whether the probe stimulus matched
the target stimulus. Trials were configured, such that there
was a probability of .5 that the probe stimulus was the same
as the target, with foils (invalid probes) drawn from the same
category as the target. The ITI lasted 10 sec and consisted
of an arithmetic task (4 sec), requiring evaluation of the
sum of three numbers, a task intended to reduce inter-
ference between trials and encourage alertness throughout
the experiment (Lewis-Peacock & Postle, 2008; Polyn, Natu,
Chiffre 1. Task diagrams for Experiment 1. (UN) In the first session,
participants performed short-term recognition of faces, lieux, et
objects inside the scanner. (B) At the beginning of the second session,
outside the scanner, participants learned arbitrary cross-category pairs
of stimuli. (C) Participants then returned to the scanner to perform
short-term paired-associate recognition of the stimulus pairs they
learned. Half of these trials included trial-irrelevant distraction during
the delay period.
Cohen, & Norman, 2005) and a final rest period (6 sec)
before the next trial began.
Phase 2: Stimulus Pairing
Ranging from 0 to 42 days following their initial scan,
participants returned to complete Phases 2 et 3 of the
experiment. For Phase 2, which occurred outside the
scanner, 18 stimuli (six faces, six places, and six objects)
were selected at random (a different subset for each par-
ticipant) from the initial set and paired arbitrarily so that
nine stimulus pairs were created (Figure 1B). Each pair
consisted of two stimuli from different categories (face–
place, face–object, and place–object pairs). Participants
learned these pairings via repeated three-alternative
forced-choice testing (with foils drawn from the set of
18) until they achieved a criterion-level performance of
72 consecutive correct trials. The learning task was com-
pleted in approximately five min for each participant.
Phase 3: Short-term Paired-associate Recognition
Immediately after learning the stimulus pairs, participants
returned to the scanner and performed paired-associate
recognition with those stimuli (Figure 1C). Each trial con-
sisted of a target stimulus (1 sec), a delay period (11 sec), un
probe stimulus (1 sec), a response period (3 sec) and an
ITI (10 sec) configured as in Phase 1. Participants indicated
with a yes–no button press whether the probe stimulus
was the correct associate of the target stimulus. Trials were
configured, such that there was a probability of .5 that the
probe stimulus was the correct associate of the target,
with foils drawn from the trial-irrelevant category (c'est à dire.,
the category to which neither the target nor its associate
belonged). The trial depicted in Figure 1C is an example
of a face–place trial: The target was a face, and its paired-
associate stimulus was a place. Randomly, on half of the
trials, four trial-irrelevant “distractor” pictures were pre-
sented during the delay period in rapid succession (0.5 sec
per stimulus, 2 sec in total). These stimuli were always se-
lected from the trial-irrelevant category (par exemple., object stimuli
on a face–place trial). Participants passively observed these
stimuli and were instructed not to divert their gaze from
the center of the screen when they appeared. Il y avait
144 trials (72 with distraction). One third (c'est à dire., 48) of the
trials involved face–place pairs, one third involved face–
object pairs, and the remaining one third involved object–
place pairs. For each pair, half of the trials presented one
stimulus as the target (par exemple., the face stimulus from a face–
place pair), and the other half of the trials presented its
associate as the target (par exemple., the place stimulus from the
same face–place pair). Each of the 18 unique pairs was pre-
sented in eight trials (four times in each direction). (Note
que, although this task requires LTM for stimulus pairings,
it is a test of STM, because the correct evaluation of the
probe requires memory for what was presented at the
beginning of the trial.)
Lewis-Peacock et al.
63
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
un
/
_
j
0
o
0
c
1
n
4
_
0
un
p
_
d
0
0
b
1
oui
4
g
0
u
.
e
p
s
d
t
o
F
n
b
0
oui
7
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
je
2
3
e
s
/
j
F
t
/
.
u
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Cognitive Strategies
In our previous study (Lewis-Peacock & Postle, 2008), nous
observed a large variability in the cognitive strategy em-
ployed by our participants to solve a short-term paired-
associate recognition task. Some participants favored a
retrospective strategy (c'est à dire., they thought about the stim-
ulus that was presented at the beginning of the trial),
others favored a prospective strategy (c'est à dire., they retrieved
from LTM the associate of the stimulus that was pre-
sented and thought about it for the remainder of the de-
lay period), and still others switched between the two
strategies across trials. In the Phase 3 task of Experi-
ment 1 in this study, we attempted to control for variabil-
ity in strategies by instructing half of our participants to
use a retrospective strategy on every trial (“hold the first
picture in mind and try not to think about its associate
until the probe appears”) and the other half to use a pro-
spective strategy (“as soon as you see the first picture,
quickly recall its associate and hold it in mind”). Ce
manipulation was designed to allow the independent ob-
servation of the effects of distraction on representations
derived from visual perception (in participants using the
retrospective strategy) and on representations recalled
from LTM (in participants using the prospective strat-
egy). In accordance with findings in the monkey (Takeda,
Naya, Fujimichi, Takeuchi, & Miyashita, 2005), we pre-
dicted that the neural representation in inferotemporal
cortex (ITC) of the target stimulus, but not its associate,
would be disrupted by the distractors. Assuming that
active neural representation is the neural basis for STM,
one would predict that the loss of the target representa-
tion would cause the participant to forget and thus be
forced to guess about the validity of the memory probe,
with a consequent decline in behavioral performance.
Data Collection
All tasks were implemented with E-Prime software ver-
sion 2.0 (Psychology Software Tools, Pittsburgh, Pennsylvanie)
and an Avotec goggle system (Avotec, Inc., Stuart, FL)
was used to display visual stimuli inside the scanner.
Des images du cerveau entier ont été acquises avec un scanner 3-T
(GE Signa VH/I). For all participants, we acquired high-
resolution T1-weighted images (30 tranches axiales, 0.9375 ×
0.9375 × 4 mm). We used a gradient-echo, echo-planar
séquence (time repetition = 2000 msec, echo time =
50 msec) to acquire data sensitive to the BOLD signal
dans un 64 × 64 matrice (30 axial slices coplanar with
the T1 acquisition, 3.75 × 3.75 × 5 mm). Eight blocks
of the Phase 1 short-term recognition task were obtained,
each scan consisting 15 trials lasting 5 min 50 sec, pour
46 min 40 sec in functional scans. All task runs were pre-
ceded by 20 sec of dummy pulses to achieve a steady
state of tissue magnetization. Eight blocks of the Phase
3 paired-associate recognition task were also obtained,
each scan consisting 18 trials lasting 8 min 8 sec, pour
65 min 4 sec in functional scans. Across both tasks, chaque
participant was tested for 111 min 45 sec.
Preprocessing
Preprocessing of the functional data was done with the
AFNI software package using the following preprocessing
mesures (in order): (1) correction for slice time acquisition
and rigid body realignment to the first volume from the
experimental task with 3dvolreg, (2) removal of signal
spikes with 3dDespike, (3) removal of the mean from
each voxel and linear and quadratic trends from within
each run with 3dDetrend, et (4) correction for mag-
netic field inhomogeneities (using in-house software).
Enfin, functional data from the second task were aligned
to data from the first task using 3dAllineate. Noter que
neither was spatial smoothing imposed nor were the
data spatially transformed into a common atlas space
before hypothesis testing. Plutôt, the data from each par-
ticipant were analyzed in that participantʼs unsmoothed,
native space.
For classification analyses, a feature selection ANOVA
was applied to the preprocessed images to select those
voxels whose activity varied significantly ( p < .05) between
face, place, and object categories over the course of the
Phase 1 task. This standard procedure reduces noise
in the classification analyses by removing uninformative
voxels. (Note that we repeated the analyses reported
here without prior feature selection, which produced
qualitatively similar, although quantitatively noisier, re-
sults.) The number of voxels passing feature selection
was 4540 (SD = 2255). Searchlight classification analyses
(with a sphere radius of 2 [7 voxels], 3 [19 voxels], or
4 [33 voxels]; see Kriegeskorte, Goebel, & Bandettini,
2006) were also applied to the Phase 1 data to assess the
extent of category-specific information throughout the
brain. Classifier decoding of Phase 3 data using voxels se-
lected by the searchlight technique produced qualitatively
similar results to those selected by the simpler ANOVA
procedure, and therefore, only results from the ANOVA-
based feature selection masks are reported. Many pre-
vious accounts have emphasized the importance of PFC
in supporting the temporary retention of information
across distraction. To address this idea, we divided the
feature-selected voxels into “no-PFC” and “PFC-only” masks.
Anatomically derived PFC masks were generated for
each participant in AFNI by backward transforming a
TT_Daemon atlas mask (consisting BA 8–11 and BA 44–
46) into that participantʼs native space. New “no-PFC”
masks were created by removing all PFC voxels from the
original feature-selected set. The number of voxels retained
in each condition was 3844 (SD = 1908) for the “no-PFC”
condition and 696 (SD = 347) for the “PFC-only” condi-
tion. An additional mask was created for each participant
covering the ITC, which consisted of the inferior temporal,
middle temporal, and fusiform gyri (403 voxels, SD = 156).
These masks were created in a similar fashion as PFC
64
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
masks. Voxels from these masks served as input nodes to
the pattern classifier for hypothesis testing.
MVPA: Training
A pattern classifier was trained, separately for each par-
ticipant, on data from the delay period of the Phase 1
task. The Princeton MVPA Toolbox (code.google.com/p/
princeton-mvpa-toolbox), in conjunction with the Matlab
Neural Network Toolbox, was used for all classification
analyses (see Pereira et al., 2009; Haynes & Rees, 2006;
Norman et al., 2006, for reviews). Data from the initial
8 sec, at intervals of 2-sec repetition time (TR), of each
trial from Phase 1 were used to train a two-layer (no
hidden layers) feedforward neural network via Matlabʼs
trainscg scaled conjugate gradient backpropagation algo-
rithm, with sigmoidal transfer functions between the
input layer (N voxels) and output layer (three stimulus
categories) of the network. The classifier was trained to
distinguish patterns of brain activity corresponding to the
short-term retention of faces, places, and objects. Note
that data from the ITI were not used as a baseline in train-
ing, because the interval between trials was filled with
a secondary task (arithmetic) that engaged the brain
more strongly than is characteristic of an unfilled ITI
(see Experiment 2). To assess empirically the inclusion
of the first TR of each trial (during which the visual stim-
ulus was on screen for the first 1 sec), we calculated the
classification accuracy at each time interval of the 8-sec
training window and found that category discrimination
was well above chance throughout the entire period.
Thus, we are confident that comparable stimulus-category-
specific activity was being evoked throughout the first
8 sec of the trial, despite contamination from the initial per-
ception and encoding of the target stimulus. A unique
classifier was created for each participant and applied only
to that participantʼs data. To reduce prediction error in
analyses involving the nondeterministic backpropagation
classifier algorithm, the reported results were the average
of 50 network iterations, each initialized with a different
set of random weights. All data used to train the classifiers
were shifted back in time by 4 sec to account for hemo-
dynamic lag of the BOLD signal. Therefore, the 8 sec of
fMRI data that were used from each trial were actually data
that were recorded between 4 and 12 sec after the begin-
ning of the trial. This adjustment, although crude, reason-
ably accommodates the slow hemodynamic response
and is standard practice in MVPA. As a check on validity,
we retrained the classifier using a 6-sec lag adjustment,
and this did not significantly alter the results. We evaluated
classifier training accuracy by using the method of k-fold
cross validation, that is, training on k − 1 blocks of data
and testing on the kth block and then rotating and repeat-
ing until all trials had been classified. For each 2-sec TR of
fMRI data, the classifier produced an estimate (from 0 to
1) of the extent to which the brain activity matched the
pattern of activity corresponding to the three categories
it had been trained on. These estimates reflected the clas-
sifierʼs evidence for each category. The classifierʼs predic-
tion at each TR corresponded to the category with the
most evidence. Prediction accuracy was calculated as the
proportion of TRs in which the classifier correctly pre-
dicted the actual category of the trial from which that TR
was sampled.
To assess the relative importance of different brain areas
to the classification of the stimulus categories, we deter-
mined, from the trained pattern classifier, which voxels
were important for identifying patterns of brain activity
corresponding to each of the three categories. We applied
the voxel importance formula (from Polyn et al., 2005):
impij
¼ 100 (cid:1) wij (cid:1) avgij;
where wij is the weight between input unit i and output unit
j and avgij is the average activity of input i during the short-
term retention of category j. Importance maps for the three
categories were calculated for each participant, transformed
into standardized space using AFNIʼs @auto_tlrc and
adwarp, blurred with a 4-mm FWHM and averaged across
all participants with 3dmerge, thresholded at an importance
score of 2.0, and overlaid on an inflated anatomical version
of the N27 brain data set (Holmes et al., 1998) using AFNIʼs
surface mapping utility for display purposes.
MVPA: Testing
A trained pattern classifier for each participant, trained on
all eight blocks of Phase 1 data, was used to assess the
extent to which category-specific patterns of brain activity
reappeared during the delay period of the Phase 3 task.
Preprocessed fMRI data at intervals of 2-sec TR were clas-
sified from the initial 20 sec of each trial (Figure 1C), cor-
responding to target presentation (1 sec), delay period
(11 sec), probe presentation (1 sec), and the first 7 sec
of the ITI (which was not rest, but filled with an arith-
metic task). Pattern classification of these data allowed
us to distinguish brain activity corresponding to the tar-
get, its associate, and the trial-irrelevant category. If, for
example, a face-like delay period activity pattern was
identified on a face–place trial, this would suggest that
the brain was actively maintaining, via persistent brain ac-
tivity, a representation of the face stimulus presented at
the beginning of the trial, consistent with a retrospective
strategy. Delay period activity reflecting a prospective
strategy would consist brain activity patterns identified
as corresponding to the category of the targetʼs associate
(in the example, places). This could only occur if, upon
seeing the target stimulus, the participant retrieved from
LTM the representation of its associate and actively re-
tained this representation. The amount of distraction-
induced brain activity during the delay period would be
indicated by the classifierʼs evidence for the category of
the distractors (in the example, objects). Importantly,
Lewis-Peacock et al.
65
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the continuous decoding of data from these trials al-
lowed for a complete characterization of the evolution
of category-specific representations throughout each trial,
allowing for the detection of transitions between target-,
distractor-, and associate-related activity within the same
brain regions. Note that possible contamination of de-
lay period activity because of perceptual processing of
the probe stimulus was not a concern, as this process-
ing would be expected to introduce noise, not coherent
category-specific activity. This follows from the fact that
the stimulus presented as the probe was from the same
category as the associate of the target on only half of the
trials, the remaining trials presented foils drawn from a
different category.
Searching for Distraction Resistance
An additional analysis was designed to search the brain
for any evidence of distraction-resistant STM representa-
tions. The purpose of this analysis was to identify voxels
whose activity in the Phase 3 task, after being decoded by
the classifier, would show that a task-relevant stimulus
representation was sustained in the face of distraction.
We selected voxels whose activity appeared to be the least
responsive to the presentation of the distractors and then
assessed whether decoding the brain activity from these
regions produced interpretable and reliable evidence of
distraction resistance. If this analysis failed, we reasoned
that it would be unlikely to find such representations any-
where else in the brain. We applied a modified version
of the searchlight classification technique (Kriegeskorte
et al., 2006). To search for distraction-resistant activity in
the prospective strategy group, we identified spheres of
voxels (separately using a radius of 2, 3, or 4 voxels) that
both (1) coded for the associate stimulus and (2) were
least responsive to the distractors. We recorded, for all
spheres, the proportion of post-distraction data (i.e., data
from distraction-present trials between the onset of dis-
traction and the onset of the probe, 6–12 sec), during
which the classifierʼs evidence for the associateʼs category
was higher than its evidence for all other categories. This
proportion was assigned to the center voxel of the sphere,
then the sphere was shifted, and this procedure was re-
peated until all spheres had been tested. A complementary
algorithm implemented a search for distraction-resistant
activity for the target stimulus in the retrospective strategy
group. The resulting statistical voxel maps were thresh-
olded (at scores of 0.45) using estimates from a χ2 distri-
bution test with df = 2, using a strict alpha of 2 × 10−6 as
a Bonferroni correction for multiple comparisons. (Note
that these maps were also thresholded using an uncor-
rected alpha, which produced qualitatively similar results.)
Voxels from all suprathreshold spheres were combined
into one mask and used as input to the classifier for re-
training on Phase 1 and retesting on Phase 3. For a sphere
radius of 3, the average number of voxels in the prospec-
tive strategy group was 240 (SD = 243), and the average
number of voxels in the retrospective strategy group was
190 (SD = 56).
Results (Phase 1)
Behavior
The mean accuracy and RT across all participants in the
Phase 1 task were 94% (SEM = 1%) and 650 msec (SEM =
10 msec). RTs from trials with an incorrect response were
excluded. A three-way repeated measures ANOVA on re-
sponse accuracy with Stimulus Category (face/place/object)
as a within-subject factor revealed a significant main ef-
fect of Stimulus Category (F(2, 24) = 3.50, p = .046), and
follow-up pairwise comparisons (with Bonferroni correc-
tion) indicated that the accuracy on object trials (96%,
SEM = 1%) was marginally higher ( p = .053) than the accu-
racy on place trials (91%, SEM = 2%). An identical ANOVA
on RT revealed a significant main effect of Stimulus Cate-
gory (F(2, 24) = 9.36, p < .001), but follow-up pairwise com-
parisons (both with or without Bonferroni correction)
indicated that there were no reliable differences between
any category pairs.
MVPA
Brain data from all Phase 1 trials were used to train a clas-
sifier separately for each participant. Group-averaged
classification performance showed that brain activity
from the delay period of the Phase 1 task was reliably
classified as consistent with the appropriate category of
the trial (Figure 2A). The classifierʼs prediction accuracy
for each category was significantly above chance (33%)
based on one-tailed, independent-sample t tests across
participants, with p < .005, for all three categories. The
mean classifier evidence for each category showed strong
category selectivity (e.g., the face evidence was selectively
high for face trials), supported by a significant interaction
of Trial Type (face/place/object) × Evidence Type (face/
place/object) from a 3 × 3 repeated measures ANOVA on
the classifier evidence values (F(4, 48) = 220.09, p < .001).
For clarity, only data from the “no-PFC” condition are
shown here. However, training the classifier on voxel
activity from the whole brain or from voxels restricted
only to PFC or ITC was also successful (but performance
in PFC was considerably closer to chance-level predic-
tion than in the other regions). Although established
category-selective areas contributed to the classification
of the three categories (e.g., the midfusiform gyrus for
faces, the parahippocampal gyrus for places, and the lat-
eral occipital cortex for objects), multiple, distributed brain
regions were also identified as important for each cate-
gory (Figure 2B). This replicates previous findings when
famous faces, famous places, and common objects were
evaluated in a test of LTM (Lewis-Peacock & Postle, 2008;
Polyn et al., 2005).
66
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
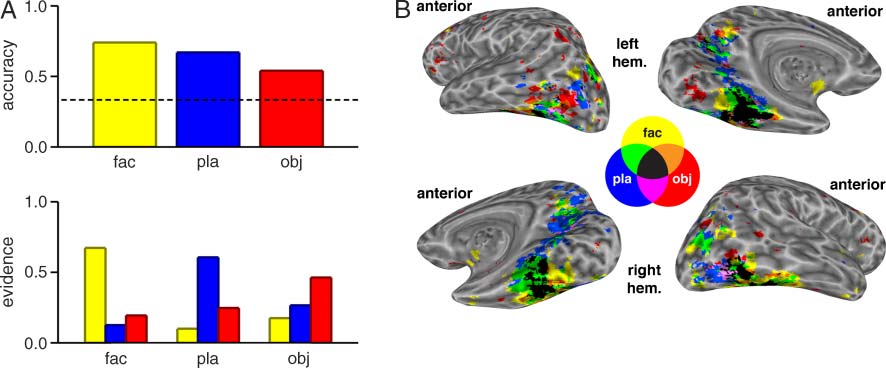
Figure 2. Classifier training for Experiment 1. (A) Classification results for the Phase 1 data are shown separately for all three categories on
which it was trained: face ( fac, yellow), place ( pla, blue), and object (obj, red). Prediction accuracy is shown on the top graph (chance level
accuracy of 0.33 is indicated by the dashed line), and average classifier evidence is shown on the bottom graph. The evidence values reflect
reliable category discrimination (e.g., for face trials, the classifierʼs evidence for face was much higher than its evidence for either place or
object). (B) Classifier-derived voxel importance maps show voxels whose activity exerted a strong influence on the classifierʼs identification
of a particular category. Group-averaged data are displayed on an inflated brain (left hemisphere in top row, right hemisphere in bottom row).
Brain areas are colored according to the Venn diagram in the center (e.g., black represents an overlap of all three categories).
Results (Phase 3)
Behavior
The mean accuracy and RT across all participants in the
Phase 3 task were 96% (SEM < 1%) and 778 msec (SEM =
11 msec). A 2 × 2 × 6 mixed ANOVA on response accu-
racy, with Instructed Strategy (retrospective/prospective)
as a between-subject factor and Distraction Condition
(absent/present) and Trial Type (six pairwise combinations
of face, object, and scene) as within-subject factors, revealed
a marginally significant main effect of Instructed Strategy
(F(1, 11) = 4.18, p = .065), indicating a trend that the
prospective strategy (98%, SEM < 1%) produced better
accuracy than the retrospective strategy (95%, SEM = 1%).
The main effect of Distraction Condition was also marginally
significant (F(1, 11) = 4.36, p = .061), indicating a trend that
participants responded more accurately to distraction-
present trials (97%, SEM = 1%) than to distraction-absent
trials (96%, SEM = 1%). However, neither of these main
effects were statistically reliable at the 5% standard alpha cut-
off. The main effect of Trial Type (F(1, 11) = 0.35, p = .882)
and all interactions between the factors were nonsignifi-
cant. An identical 2 × 2 × 6 mixed ANOVA on RTs re-
vealed a significant main effect of Distraction Condition
(F(1, 11) = 46.86, p < .001), indicating that participants re-
sponded faster on trials with distraction (734 msec, SEM =
14 msec) than on trials without distraction (823 msec,
SEM = 16 msec). This difference likely reflected a gen-
eral attentional enhancement for distraction-present trials
because of the processing of additional stimuli during the
otherwise long, unfilled delay period (see also Postle,
Idzikowski, Della Salla, Logie, & Baddeley, 2006). A related
possibility is that because the distractors were always from
a different category than the target and its associate, the
presentation of distractors during the delay period may
have served to reduce uncertainty about the category of
the targetʼs associate, thus narrowing the retrieval space
and facilitating performance. The main effect of Trial Type
was significant (F(5, 55) = 2.44, p = .045), but follow-up
pairwise comparisons (both with and without Bonferroni
correction) revealed no reliable differences between any
pair of trial types. The main effect of Instructed Strategy
(F(1, 11) = 1.92, p = .193) and all interactions between
factors were nonsignificant.
MVPA
Brain data from all Phase 3 trials were decoded, separately
for each participant, using a classifier that was trained on
data from all Phase 1 trials. For clarity, we present only
results from the “no-PFC” ROI.2 For participants who were
instructed to retain the perceptual stimulus during the de-
lay (“retrospective strategy”), a sustained representation
of this stimulus was identified on distraction-absent trials,
as indicated by relatively greater evidence for the target cat-
egory throughout the delay (Figure 3, top left). Although
strong evidence for the target category was also observed
during the early portion of the delay period on distraction-
present trials, it was sharply attenuated and replaced by
evidence for the trial-irrelevant category following the
onset of the distractors (Figure 3, bottom left). This re-
sult indicates that the active neural representation of the
target stimulus (as assessed by MVPA) was replaced by
perceptual representations of the distractors. For par-
ticipants instructed to retrieve the targetʼs associate and
Lewis-Peacock et al.
67
retain it in anticipation of the probe (“prospective strat-
egy”), sustained representation of the category of the
associate was identified on distraction-absent trials, indi-
cated by a transition from strong evidence for the tar-
get to strong evidence for its associate during the delay
(Figure 3, top right). Because the probe stimulus had not
yet been presented, any brain activity classified as con-
sistent with the associateʼs category must have been
reinstated from LTM. It has been proposed that the in-
formation that is retrieved from LTM and then actively
retained in STM is more robust to distraction than percep-
tually derived information (Takeda et al., 2005). Contrary
to this proposal, however, our results show that sustained
category-specific information related to the LTM-derived
associate stimulus was disrupted by the distractors. The
classifierʼs evidence for the associate was attenuated (and
became indistinguishable from the estimates of the task-
irrelevant stimulus category) when distractors were pre-
sented during the delay, accompanied by a significant
increase in evidence for the distractors (Figure 3, bottom
right).
A 2 × 2 × 3 × 10 mixed ANOVA on classifier evidence
values with Instructed Strategy (retrospective/prospective)
as a between-subject factor and Distraction Condition
(absent/present), Stimulus Type (target/associate/irrelevant),
and Time (TRs 1–10) as within-subject factors revealed
a significant three-way Strategy × Stimulus × Time in-
teraction (F(18, 198) = 1.77, p = .031). This result sup-
ports the qualitative interpretation, suggested in Figure 3,
that task instruction had a differential effect on the trial-
averaged classifier evidence values for the two groups
of participants. The three-way Distraction × Stimulus ×
Time interaction was also significant (F(18, 198) = 11.11,
p < .001), confirming that the distraction manipulation
had a statistically reliable effect on the classifier evidence
values across the duration of the trials. The four-way in-
teraction of Strategy × Distraction × Stimulus × Time
was nonsignificant (F(18, 198) = 1.33, p = .174). Taken
together, the results from both groups indicate that the
active task-relevant representation was disrupted following
distraction.
An additional analysis using a voxel searchlight tech-
nique identified, in each participant, a small set of voxels
that exhibited a relatively weaker response to the dis-
tractor stimuli (see Methods). However, retraining a classi-
fier on Phase 1 data from only these voxels failed to find
any reliable evidence for distraction-resistant represen-
tations in the Phase 3 data (data not shown). Any brain
region we tested that showed evidence of sustained repre-
sentation of the task-relevant stimulus during the first half
of the delay period also showed a robust neural response
to the trial-irrelevant distractors, which in turn suppressed
the activity pattern associated with the former. Therefore,
despite applying two different classification approaches
(from large ROIs that included thousands of voxels and
from small searchlight spheres that included tens of
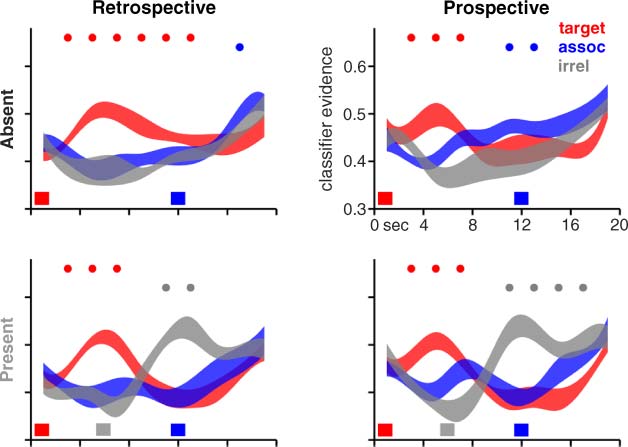
Figure 3. Classifier decoding
for Experiment 1. Mean
classifier evidence values are
shown separately for the
retrospective strategy group
(left column, Retrospective)
and the prospective strategy
group (right column,
Prospective), and separately
for the distraction-absent
trials (top row, Absent) and
the distraction-present trials
(bottom row, Present).
Evidence values for the face,
place, and object categories
were relabeled and collapsed
across all trials into three
new categories: target (red,
the category of the target
stimulus on a given trial),
assoc (blue, the category
of the targetʼs associate
stimulus), and irrel (gray,
the trial-irrelevant category).
Data for each category are shown as ribbons whose thickness indicate ±1 SEM across participants, interpolated across the 10 discrete data
points in the trial-averaged data. The colored bars along the horizontal axis indicate the onset of the target (red, 0 sec), the distractors (gray,
6 sec; distraction-present trials only), and the probe (blue, 12 sec). Statistical comparisons of evidence values for the three categories focused
on within-subject differences. For every 2-sec interval throughout the trial, color-coded circles at the top of each graph indicate the category
whose evidence was greater ( p < .05, based on repeated measures t tests) than the average evidence for the other two categories. Unlike
the data from Phase 1 that was used to train the classifier, these data were not shifted in time, and therefore the peak response to a trial
event appears approximately 4–6 sec after the onset of the event.
68
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
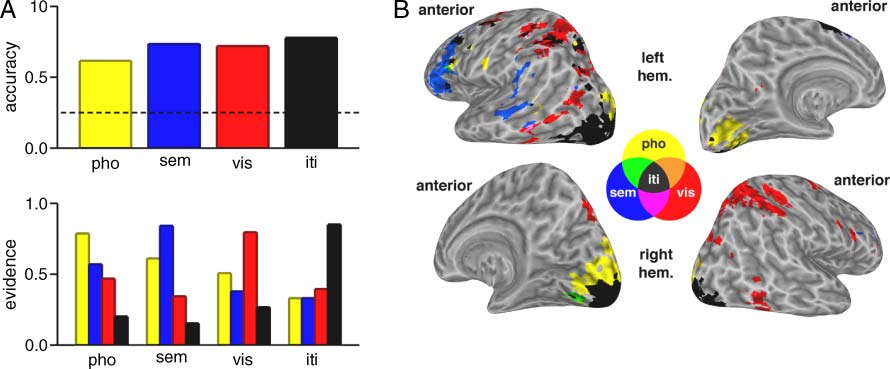
Figure 4. Task diagrams for
Experiment 2. (A) In the first
phase, participants performed
short-term recognition of a
pseudoword (phonological
STM), a word (semantic STM),
or two lines (visual STM).
(B) In the second phase,
during the same scanning
session, participants performed
short-term recognition with
two stimuli (between-category
combinations of pseudowords,
words, and lines). On half of the
trials, the same memory item
was selected as behaviorally
relevant by the first and second
cues (repeat trials), and on the
other half of trials the second
cue selected the previously
uncued item (switch trials).
voxels), we were unable to find any reliable evidence
for distraction-resistant representations of trial-relevant
information in the fMRI data.
Discussion
The effects of visual distraction during the delay period of
the Phase 3 task were twofold: The pattern of distributed
brain activity corresponding to a representation of the
trial-relevant stimulus dropped to baseline, and yet there
was no loss of recognition accuracy compared with trials
without the distraction. This result is intriguing because
classifier estimates of category-specific activity have been
shown to accurately reflect the strength of neural rep-
resentation of a specific stimulus (Kuhl, Rissman, Chun,
& Wagner, 2011; Newman & Norman, 2010; Quamme,
Weiss, & Norman, 2010). A strong interpretation of our
results is that the short-term retention of information does
not depend on persistent activation of representations of
the remembered material. Two methodological issues that
may cause concern with this interpretation are as follows:
(1) It is unclear whether the pattern classifier was capable
of identifying multiple, concurrently active STM represen-
tations (if they existed) or whether the results merely
reflected a winner-take-all classification outcome. (2) Be-
cause the classifier was trained on delay period activity
from the Phase 1 data, it may have been unfair to directly
compare decoding results for on-screen stimuli (the dis-
tractors) with decoding results for remembered stim-
uli (the targets and their associates), because perceptual
stimulation engages the brain more strongly than does
STM retention (Serences, Ester, Vogel, & Awh, 2009; Sheth
& Shimojo, 2003). Experiment 2, however, was not sus-
ceptible to either of these concerns.
EXPERIMENT 2
Methods
Participants
Nine (five men, ages 21–30 years) healthy, right-handed
adults were recruited from the undergraduate and medical
campuses of the University of Wisconsin—Madison. None
reported any medical, neurological, or psychiatric illness,
and all gave informed consent.
Phase 1: Short-term Recognition
Participants performed 72 trials of short-term recogni-
tion of a stimulus selected randomly from one of three
categories—English words, pronounceable pseudowords,
and line segments—with 24 trials drawn from each cate-
gory (Figure 4A). Each trial consisted of a category cue
(2 sec), a target presentation (0.5 sec), a delay period
(7.5 sec), a probe presentation (0.5 sec), a response pe-
riod (1.5 sec), followed by a blank screen (10 sec) that
preceded the next trial. Participants indicated with a but-
ton press whether the probe stimulus matched the item
in memory according to a category-specific criterion. Trials
were configured such that there was a probability of .5
that the probe stimulus satisfied the criterion. A synonym
judgment was required for words, a rhyme judgment was
required for pseudowords, and a visual orientation judg-
ment was required for line segments. Foils (to-be-rejected
probes) for the three categories were conceptually un-
related words, single-syllable pseudowords with a non-
matching vowel sound, and line segments in which one
of the segments differed in orientation by at least 30°.
Although phonological, semantic, and visual encoding
processes were likely involved in the processing of all
Lewis-Peacock et al.
69
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
target items (Wickens, 1970), the stimuli and task were
designed to encourage encoding in one primary domain
of representation on each trial. That is, we attempted to
elicit the short-term retention of information in a semantic
(i.e., conceptual) form on trials that required a synonym
judgment, in a phonological form on trials that required
a rhyme judgment, and in a visual form on trials that re-
quired a line orientation judgment. Words were presented
in white (on black background) to indicate that the stimu-
lus was to be primarily encoded based on its semantic
characteristics. Pseudowords were presented in cyan to
indicate that the stimulus was to be primarily encoded
based on its phonological characteristics. Line segments
were always presented in white (on black background)
and were to be primarily encoded in a visual form. The
domain-specific comparison criteria used here were mod-
eled after a rich literature highlighting dissociations be-
tween verbal and visual processes in STM (Baddeley,
1986), as well as more recent studies dissociating semantic
and phonological components (Cameron, Haarmann,
Grafman, & Ruchkin, 2005; Shivde & Thompson-Schill,
2004; Martin, Wu, Freedman, Jackson, & Lesch, 2003;
Haarmann & Usher, 2001).
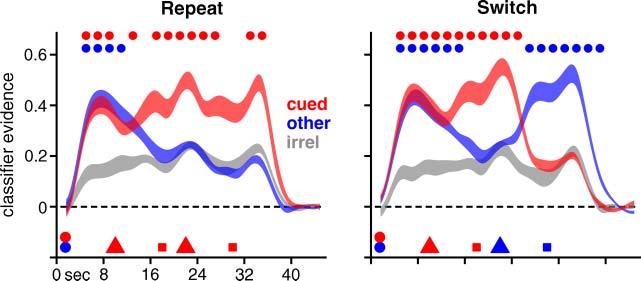
Phase 2: Short-term Recognition with Relevance Cues
Participants performed a second short-term recognition
task in the scanner immediately after completing the
Phase 1 task. This task was modeled on a modified version
of the Sternberg recognition task (Oberauer, 2005). At the
beginning of each trial, one stimulus was presented on
the top half of the screen, and another was presented on
the bottom half (Figure 4B). The two stimuli for each trial
were always selected from separate categories, such that
two of the three stimulus categories were represented in
every trial. Stimulus offset was followed by a brief delay
and then a cue indicating which memory item was relevant
for the first recognition probe. The cues consisted of two
inward-facing red arrows, centered on either the top or
bottom half of the screen, the location of which cor-
responded to the location where a stimulus had been
presented at the beginning of the trial. After the probe
(and response), a second cue appeared which indicated
the relevant memory item for a second recognition probe,
with equal probability of cuing either item. Thus, until the
onset of the second cue, both stimuli from the beginning
of the trial needed to be retained for successful task per-
formance. Trials in which the same memory item was se-
lected by both cues are referred to as “repeat” trials, and
the other trials are referred to as “switch” trials. Similar to
the Phase 1 task, trials in Phase 2 were configured such
that there was a probability of .5 that the probe stimulus
satisfied the category-specific criterion, with foils chosen
as before. There were 72 trials, one third of which involved
stimuli representing each of the three category combina-
tions (i.e., words and pseudowords, words and lines, and
pseudowords and lines).
Stimuli
Words were nouns, verbs, and adjectives selected from an
on-line psycholinguistic database (www.psy.uwa.edu.au/
mrcdatabase/uwa_mrc.htm) with concreteness, imageabil-
ity, and frequency of each within one standard deviation
of the mean of the entire database. Pseudowords consisted
of pronounceable single-syllable letter strings that were
created for this study. Intended pronunciation of the
pseudowords was based on standard English (i.e., a string
ending with the letter “e” indicated a long vowel sound and
a string ending with a double consonant indicated a short
vowel sound). No compound vowels (e.g., “ou”) were used.
Line stimuli consisted of a pair of line segments, each line
tilted between 10° and 170°, at intervals of 10°, away from
vertical. Tilt angles of 0°, 90°, and 180° were excluded to
discourage participants from recoding the stimuli into
categorical codes (e.g., “vertical” or “horizontal“).
Data Collection and Preprocessing
The collection and preprocessing of MRI data were iden-
tical to the procedures described for Experiment 1. Four
blocks of the Phase 1 task were obtained, each consisting
18 trials (six trials from each stimulus category) lasting
6 min 56 sec, for a total of 27 min 44 sec in functional
scans. In the same scanning session, eight blocks of the
Phase 2 task were also obtained, each consisting nine
trials lasting 7 min 14 sec, for a total of 57 min 52 sec in
functional scans. Across both tasks, each participant per-
formed memory tasks for 85 min 19 sec. A feature selection
ANOVA was applied to the training data, as in Experiment 1,
to remove uninformative voxels. The average number
of voxels selected across participants was 11,184 (SD =
2648). Voxels from these masks served as input nodes to
the pattern classifier for hypothesis testing.
MVPA: Training
A pattern classifier was trained, separately for each par-
ticipant, on data from the delay period of the Phase 1 task.
Data from the final 6 sec of the 7.5-sec delay period, at
intervals of 2-sec TR, were used to train a classifier to dis-
tinguish patterns of brain activity corresponding to the
short-term retention of information encoded primarily in
a phonological (pseudoword trials), semantic (word trials),
or visual (line trials) form. As in Experiment 1, all data were
shifted back in time by 4 sec to account for hemodynamic
lag of the BOLD signal. Therefore, the 6 sec of fMRI data
that were used from each trial were actually data that were
recorded between 8 and 14 sec after the beginning of the
trial. To improve the interpretability of the whole-trial de-
coding of the Phase 2 data, we also trained the classifier
on resting state brain activity during the unfilled ITI. Rest-
ing activity served as a “ground reference” for the classifier,
analogous to how the Earth serves as a zero-voltage ground
reference for electrical circuits. Training the classifier with
70
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
rest activity did not alter the classifierʼs assessment of the
relative differences between the three stimulus categories
during the task portion of the trial. It did, however, nor-
malize the classifierʼs assessment such that evidence for
the stimulus categories was low during the rest periods,
consistent with the fact that participants were not per-
forming a memory task during these periods of the ex-
periment. Data from the ITI was randomly sampled so
that, within each block of trials, the classifier was trained
on the same number of exemplars for all four categories
(72 total TRs each of phonological, semantic, visual, and
ITI across the whole experiment). A unique classifier was
created for each participant and applied only to that par-
ticipantʼs data. Classifier training accuracy was assessed,
and voxel importance maps (thresholded at an importance
value of 0.075) were calculated as described above for
Experiment 1.
Classification for Experiment 2 was carried out using
penalized logistic regression, using L2 regularization with
a penalty parameter of 50. Regularization prevents over-
fitting by punishing large weights during classifier training
(Duda, Hart, & Stork, 2001). Note that the classification
for both Experiments 1 and 2 was initially carried out
using backpropagation (see Experiment 1 under Methods)
but was also rerun using penalized logistic regression. Clas-
sification performance for Experiment 1 did not change
(and therefore, we report the initial results), but classifier
performance was significantly improved for Experiment 2.
We believe that L2 regularization was particularly impor-
tant for Experiment, 2 because the classifier was also
trained on resting state activity between trials, and there-
fore, it partially learned to discriminate the three task con-
ditions based on features that were in common to the
three stimulus categories. Overfitting was less problematic
in Experiment 1, because the classifier was not trained on
resting activity (the intertrial intervals were filled with an
arithmetic task).
MVPA: Testing
A pattern classifier for each participant, trained on all
four blocks of Phase 1 data, was used to assess the extent
to which category-specific patterns of brain activity re-
appeared during the Phase 2 task. Preprocessed fMRI data
at intervals of 2-sec TR were classified from every trial. Be-
cause the classifier was also trained on resting state activ-
ity, the evidence values at the beginning and end of each
trial for the three stimulus categories were equally low,
but nonzero. For display purposes, this low-level evidence
was removed from all classifier evidence values so that
the trial-averaged decoding traces would begin at zero.
The continuous decoding of data from the entirety of the
trials allowed for a complete characterization of the evolu-
tion of brain states corresponding to category-specific
information inside and outside the focus of attention. If
sustained brain activity reflected the contents of the focus
of attention, but not all of STM, one would expect that
the category information decoded by the classifier would
track only that information which is held in the focus of
attention. During the initial delay period, both memory
items would be maintained in the focus because both were
potentially relevant for the first response. Following the
first cue, removal of task-irrelevant information from the
focus would be indicated by an attenuation of classifier
evidence for that memory item. Whether the strength of
classifier evidence were to drop to an intermediate level
or to baseline would have implications for what it means
for information to be in “in” STM but outside the focus
of attention. On switch trials, retrieval of information
from “activated LTM” back into the focus of attention
would be indicated by the restrengthening of classifier
evidence for the memory item cued as relevant for the
second decision. In contrast, if sustained brain activity
reflected the full contents of STM, we would expect that,
regardless of cueing, evidence for the categories of both
target items should be detected by the classifier through-
out the trial (at least until the second cue, because both
stimuli had to be remembered up to that point).
Results (Phase 1)
Behavior
The mean accuracy and RT across all participants in the
Phase 1 task were 94% (SEM = 1%) and 933 msec (SEM =
22 msec). RTs from trials with an incorrect response were
excluded. A three-way repeated measures ANOVA on Re-
sponse Accuracy with Stimulus Category (phonological|
semantic|visual) as a within-subject factor revealed a
significant main effect of Stimulus Category (F(2, 16) =
4.06, p = .037), and follow-up pairwise comparisons
(with Bonferroni correction) indicated that the accuracy
on semantic trials (98%, SEM = 1%) was reliably higher
( p = .037) than the accuracy on phonological trials
(91%, SEM = 3%). An identical ANOVA on RT revealed a
significant main effect of Stimulus Category (F(2, 16) =
4.11, p = .036), but follow-up pairwise comparisons (both
with or without Bonferroni correction) indicated that there
were no reliable differences between any pair of stimulus
categories.
MVPA
Brain data from all Phase 1 trials were used to train a classi-
fier separately for each participant. Group-averaged classifi-
cation performance showed that brain activity from the
retention interval of the Phase 1 task was reliably classified
as matching the stimulus category of the trial (Figure 5A).
The classifierʼs prediction accuracy for each category was
significantly above chance (25%) based on one-tailed,
independent-sample t tests across participants, with p <
.01. The mean classifier evidence for each category showed
strong category selectivity (e.g., the phonological classi-
fier evidence was selectively high for phonological trials),
Lewis-Peacock et al.
71
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 5. Classifier training for Experiment 2. Graph conventions are described in Figure 2. The classifier training performance (A) and
voxel importance maps (B) are shown for phonological ( pho), semantic (sem), visual (vis), and resting state brain activity from the intertrial
interval (iti). Chance level predication accuracy was 0.25 and is indicated by the dashed line on the top graph in (A). Unlike in Figure 2, the
voxels on the inflated brain hemispheres in (B) correspond to an overlap between one or more trial categories with the iti category. There
were no important voxels that overlapped for pho, sem, and vis but not iti, and there were very few important voxels for the iti alone, and
so all iti-related voxels were painted black.
supported by a significant interaction of Trial Type (phono-
logical/semantic/visual/ITI) × Evidence Type (phonological/
semantic/visual/ ITI) from a 4 × 4 repeated measures
ANOVA on the classifier evidence values (F(9, 72) = 66.14,
p < .001). Because each stimulus category putatively
required short-term retention in one primary domain of
representation, this result indicates that the classifier suc-
cessfully differentiated visual from phonological (Baddeley,
1986) from semantic (Cameron et al., 2005; Shivde &
Thompson-Schill, 2004; Martin et al., 2003; Haarmann
& Usher, 2001) STM, and all three from the resting state
activity recorded during the ITI. A distributed network of
voxels throughout the brain was identified as important for
supporting the classification of each category of stimulus
(Figure 5B).
Results (Phase 2)
Behavior
The mean accuracy and RT across all participants in the
Phase 2 task were 91% (SEM = 1%) and 936 msec (SEM =
10 msec). RTs from trials with an incorrect response were
excluded. A 2 × 2 × 6 repeated measures ANOVA on
response accuracy with Cue Type (repeat/switch), Probe
Type (first/second), and Stimulus Type (six pairwise com-
binations of phonological, semantic, and visual) as within-
subject factors revealed a significant main effect of Cue
Type (F(1, 8) = 27.18, p < .001), indicating that partici-
pants were more accurate on repeat trials (93%, SEM =
1%) than on switch trials (88%, SEM = 1%). The main
effect of Probe Type was nonsignificant (F(1, 8) = 0.30,
p = .597), but the main effect of Stimulus Type was sig-
nificant (F(5, 40) = 3.80, p = .007), and follow-up pair-
wise comparisons (with Bonferroni correction) indicated
that participants responded less accurately to visual–
phonological trials (i.e., trials in which the first stimulus
that was cued was a line, and the other stimulus presented
at the beginning of the trial was a pseudoword; 84%,
SEM = 2%) than to both phonological–semantic trials
(94%, SEM = 2%; p = .022) and semantic–phonological
trials (94%, SEM = 2%; p = .013). Finally, the Probe Type ×
Stimulus Type interaction was significant (F(5, 40) = 2.56,
p = .042), and the three-way interaction of Cue Type ×
Probe Type × Stimulus Type was also significant (F(5,
40) = 3.94, p = .005). An identical 2 × 2 × 6 repeated
measures ANOVA on RTs revealed a significant main effect
of Cue Type (F(1, 8) = 7.86, p = .023), indicating that par-
ticipants were faster to respond on repeat trials (924 msec,
SEM = 14 msec) than on switch trials (948 msec, SEM =
15 msec), and a significant main effect of Probe Type
(F(1, 8) = 25.23, p = .001), indicating that participants
were faster to respond to the second probe (898 msec,
SEM = 13 msec) than to the first probe (975 msec, SEM =
15 msec). All two-way interactions and the three-way inter-
action were nonsignificant.
MVPA
Brain data from every time point in all Phase 2 trials were
decoded, separately for each participant, using a classifier
that was trained on specific time points (i.e., delay and
rest periods) from all Phase 1 trials. Group-averaged classi-
fication results for both repeat and switch trials (Figure 6)
revealed an initial rise in classifier evidence for all three
categories in concert with the onset of the trial, although
the waveforms quickly diverged as a function of whether
72
Journal of Cognitive Neuroscience
Volume 24, Number 1
the category was relevant on that trial. Classifier evidence
for the trial-irrelevant category (say, for phonological in-
formation on trials that presented lines and a noun) quickly
peaked at a low level and sustained this in a tonic manner
until the end of the trial. The waveform deviated from
this square wave-like shape only for slight increases cor-
responding to the onset of the two probes. Thus, the
irrelevant category provided a baseline reference against
which we could quantitatively assess evidence for repre-
sentation of trial-relevant information. In all trials, classifier
evidence for both trial-relevant categories rose precipi-
tously at trial onset and remained at the same elevated
level until the onset of the first cue. This indicated that
both items were encoded and sustained in the focus of
attention across the initial memory delay, while it was equi-
probable that either would be relevant for the first mem-
ory response. Following onset of the first cue, however,
classifier evidence for the two memory items diverged.
Postcue brain activity patterns were classified as highly
consistent with the category of the cued item, whereas
evidence for the uncued item dropped precipitously,
becoming indistinguishable from the classifierʼs evidence
for the stimulus category not presented on that trial (i.e.,
not different from baseline). If the second cue was a repeat
cue, classifier evidence for the already-selected memory
item remained elevated and that of the uncued item re-
mained indistinguishable from baseline (Figure 6, Repeat).
If, in contrast, the second cue was a switch cue, classifier
evidence for the previously uncued item was reinstated,
while evidence for the previously cued item dropped to
baseline (Figure 6, Switch).
A 2 × 3 × 10 repeated measures ANOVA on classifier
evidence values from the first half of all trials (before the
onset of the second cue) with Cue Type (repeat/switch),
Stimulus Type (cued/other/irrel), and Time (TRs 1–10) as
within-subject factors revealed a significant interaction of
Stimulus Type × Time (F(18, 144) = 23.71, p < .001), con-
firming the validity of the pairwise comparisons between
classifier evidence values (shown at the top of each graph
in Figure 6 for every 2-sec time interval) which indicate
strong evidence for both memory items after encoding,
followed by selective evidence for the cued item after the
first cue. The three-way interaction of Cue Type × Stimulus
Type × Time was nonsignificant (F(18, 144) = 0.37, p =
.991), indicating that there was no discernible difference
between classifier evidence for repeat and switch trials be-
fore the second cue (confirming that the task demands of
both trial types were identical up to this point). To assess
the impact of the second cue on classifier evidence, a 2 ×
3 × 13 repeated measures ANOVA was performed on the
classifier evidence values from the second half of the trials
(posterior to the onset of the second cue) with Cue Type
(repeat/switch), Stimulus Type (cued/other/irrel), and Time
(TRs 11–23) as within-subject factors. Unlike the results
from the first half of the trials, this analysis revealed a
significant three-way Cue Type × Stimulus Type × Time
interaction (F(24, 192) = 25.42, p < .001). This analysis con-
firms that repeat and switch cues had different effects on
the classifierʼs assessment of brain activity following
the second cue, such that the classifier identified persistent
evidence only for the item that was cued for the second
response.
Discussion
Together, these results suggest that, across the 8-sec delay
periods, only the immediately behaviorally relevant STM
item, putatively in the focus of attention, was supported
by persistent patterns of category-specific delay period ac-
tivity. Notably, classifier evidence for the uncued category
did not maintain an intermediate level of activation, de-
spite the fact that it remained “in” STM. One explanation
Figure 6. Classifier decoding
for Experiment 2. Results are
shown separately for repeat
(left) and switch (right) trials.
Classifier evidence values
for phonological, semantic,
and visual were relabeled
and collapsed across all trials
into three new categories:
cued (red, the category of
the memory item selected by
the first cue), other (blue, the
category of the other memory
item), and irrel (gray, the
trial-irrelevant category). The
colored shapes along this
horizontal axis indicate the onset of the targets (red and blue circles, 0 sec), the first cue (red triangle, 10 sec), the first recognition probe (red
square, 18 sec), the second cue (red or blue triangle, 22 sec), and the final recognition probe (red or blue square, 30 sec). Data for each category
are shown as ribbons whose thickness indicate ±1 SEM across participants, interpolated across the 23 discrete data points in the trial-averaged
data. Statistical comparisons of evidence values focused on within-subject differences. For every 2-sec interval throughout the trial, color-coded
circles along the top of each graph indicate that the classifierʼs evidence for the cued or other categories, respectively, was reliably stronger
( p < .002, based on repeated measures t tests, corrected for multiple comparisons) than the evidence for the trial-irrelevant category (irrel ).
Lewis-Peacock et al.
73
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
for this finding, consistent with the results from Experi-
ment 1, is that only information that is in the focus of at-
tention is held in an active state. An alternative explanation
is that the representation of the uncued stimulus may not
have disappeared, but rather it changed following the cue.
A related possibility is that item-specific representations
(to which our category-specific classification methods
were insensitive) may have survived despite the loss of
category-level representations. We believe that these al-
ternatives are unlikely because no current theories, to
our knowledge, allow for the instantaneous, contextually
dependent recoding of STM representations into some al-
ternate form of active representation (including a form
devoid of category information). Nonetheless, we tested
the first of these alternatives by running a follow-up analy-
sis in which we trained and tested a classifier with only
postcue brain activity from the Phase 2 task. K-fold cross
validation (k = 8; see Methods) was used so that the clas-
sifier was trained and tested on separate data. The subset
of fMRI data used for this analysis consisted of the three
TRs (trial time = 10–16 sec) following the onset of first
cue from all trials. As in the original analysis, the data were
shifted by 4 sec to account for hemodynamic lag. For each
trial, the brain data were labeled according to the category
of the uncued stimulus (e.g., if the semantic stimulus was
cued on a semantic–visual trial, the data would be labeled
visual). Across all participants, this classification analysis
failed to produce above-chance decoding of the category
of the uncued stimulus. Although a null result, this finding
indicates that in our data an alternative state of active rep-
resentation of the uncued stimulus, if it was to exist, could
not be readily identified using the same measurement and
analysis techniques that successfully identified the active
representation of cued stimulus.
An important question to consider when evaluating the
results from Experiment 2 is how to interpret the baseline,
which we operationalized as the classifierʼs estimates for
trial-irrelevant information (i.e., for the stimulus category
that was not presented in the trial). A likely explanation
is that this low-level of elevated classifier evidence reflects
a task set or context that is not specific to any trial stimulus,
but is engaged with the onset of each trial, and disengaged
at the offset. This idea is compatible with accounts of
proactive interference (e.g., Gardiner, Craik, & Birstwistle,
1972). It may be that the classifier identified activity cor-
responding to the trial-irrelevant category because neural
representations of stimuli from that category (which were
presented in previous trials) were incidentally reactivated
at the beginning of each trial. This possibility arises from
the assumption that memory is accomplished in part
by associating stimulus items to their encoding contexts
(Polyn, Norman, & Kahana, 2009; Sederberg, Howard, &
Kahana, 2008; Howard & Kahana, 2002; Nairne, 2002).
Accordingly, when a context representation is activated—
either to associate a new item to it or to retrieve an old
item from it—this reactivation leads also to the reactiva-
tion of other items associated to it and, to some degree,
also to the reactivation of items associated to similar con-
texts. This process could provide an explanation for a
key piece of evidence for the idea of “activated LTM” in
the embedded component model: Recognition probes
matching the uncued contents in the modified Sternberg
task (Oberauer, 2001) or matching list elements from
recent previous trials (so-called “recent negative lures”;
Monsell, 1978; DʼEsposito, Postle, Jonides, & Smith,
1999) are harder to reject than novel probes not encoun-
tered during the last few trials. The difficulty with rejecting
this kind of lure might not come from persistent activa-
tion of their representations in LTM, but from their re-
activation by the current retrieval context, which overlaps
substantially with the context in which they have last been
experienced.
GENERAL DISCUSSION
How does the brain retain information across brief peri-
ods? The embedded component framework (Oberauer,
2002; Cowan, 1995; Ericsson & Kintsch, 1995) suggests
a distinction between retention within the focus of atten-
tion and retention outside the focus in a presumably ac-
tivated state of LTM. Although a link between attention
and STM has been widely acknowledged for some time,
the importance of internally directed attention for select-
ing subsets of information within STM (Chun, Golomb, &
Turk-Browne, 2011; Bays & Husain, 2008; Cowan, 1988)
has only recently been recognized by neuroscience re-
searchers (Chun, 2011; Cowan, 2011; Gazzaley, 2011;
Ikkai & Curtis, 2011; Lepsien, Thornton, & Nobre, 2011;
Nee & Jonides, 2008, 2011; Olivers & Eimer, 2011; Stokes,
2011; Vandenbroucke, Sligte, & Lamme, 2011; Esterman,
Chiu, Tamber-Rosenau, & Yantis, 2009; Woltz & Was,
2006; Griffin & Nobre, 2003). This study provides con-
verging neurophysiological evidence for the distinction
of two states of representations within STM by demon-
strating that the moment-to-moment information content
of delay period activity reflects items in the focus of at-
tention, but not those retained in memory outside the
focus. Intriguingly, this was true whether the information
in the focus was stimulating sensory receptors (as in
Experiment 1) or, instead, was itself already in STM (as in
Experiment 2).
Attention and memory were unconfounded by causing
either an external shift of attention to trial-irrelevant stimuli
or by causing an internal shift to a subset of information
already being remembered. Experiment 1 showed that,
following the presentation of trial-irrelevant stimuli during
a delay period, ongoing brain activity carried information
about the distractors on the screen and therefore pre-
sumably in the focus of attention and not about the items
that were not on the screen but yet retained in memory (as
verified by near-perfect recognition performance). One
possibility is that our analysis methods were insufficiently
sensitive to detect unattended STM representations in
the presence of perceptual distraction. An alternative,
74
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
however, is that sustained delay period activity reflects
only that information that is currently in the focus of atten-
tion rather than the full contents of STM. Experiment 2
provided evidence for the latter interpretation. It dem-
onstrated that temporarily irrelevant items in STM were
quickly removed from the focus of attention to a point at
which their signature in ongoing brain activity vanished
completely. However, these items could re-enter the focus
and have their active neural signature restored, if they
were cued as relevant for behavior a few seconds later.
These results, therefore, support the distinction between
two functional states of representations in STM: inside
and outside the focus of attention (Oberauer, 2002; Cowan,
1995). Both serve STM, but only representations inside
the focus are detectable in the moment-to-moment pat-
terns of delay period brain activity.
We will now discuss, in more detail, a series of concerns,
methodological and theoretical, that relate to our present
findings. Assuming that ongoing neural activity is accom-
panied by a correlated pattern of regional CBF, there are
two classes of explanation for the finding of STM (outside
the focus of attention) without persistent neural activity.
The first is methodological. The short-term retention of
information may have been accomplished via a reduced
level of sustained firing that was not detectable with our
fMRI protocol. A related possibility is that retention was sup-
ported by some other type of sustained activity to which
BOLD is less sensitive, such as coherent low-frequency
oscillations among task-specific neural populations. Note,
however, that MVPA is much more sensitive than tradi-
tional measures of BOLD (Kriegeskorte et al., 2006; Norman
et al., 2006). This is seen, for example, in the ability to re-
cover stimulus-related information in V1 during the delay
period of STM tasks despite the absence of above-baseline
activity (Harrison & Tong, 2009; Serences et al., 2009) and
in the ability to discriminate patterns of activity represent-
ing individual faces (Kriegeskorte et al., 2007). Also, the
results from Experiment 2 in this study demonstrate that
the classification procedure was sensitive enough to detect
superimposed patterns of brain activity corresponding to
the active representations of two memory items from dif-
ferent stimulus categories. It was only after one item was
cued during the delay period that the classifierʼs evidence
for the items diverged.
Another methodological concern is that the experimen-
tal design may have been too insensitive to test our hy-
potheses. Training a classifier on brain activity from one
task and using it to decode brain activity from another task
(with a different set of task demands) may not succeed if
the STM representations were qualitatively different as a
result of the different cognitive demands of the two tasks.
However, the successful detection of task-relevant stimulus
representations in distraction-absent trials (Experiment 1)
and in precue delay periods (Experiment 2) validates that
patterns of active stimulus representations were similar
across the training and testing phases in each experiment.
The possibility that the qualitative form of active represen-
tation changed, rather than disappeared, for items outside
the focus of attention seems unlikely and is not antici-
pated by any existing theories with which we are familiar.
Furthermore, a follow-up analysis in Experiment 2 that
considered brain activity from only the Phase 2 task failed
to find evidence for an alternative form of active represen-
tation of the unattended memory items.
A second class of explanation for our results arises from
an alternative to activation-based accounts of short-term
retention. One mechanism that could accomplish short-
term retention without persistent activity is weight-based
retention via changes in synaptic potentiation. During the
delay period, the memory traces are not actively main-
tained in the sense of elevated firing rates or metabolic
demands. Rather, they are passively retained by short-term
increases in the strength of synaptic connections between
neurons that represent the information. Synaptic weights
can be temporarily modified via transient elevation of the
concentration of presynaptic calcium ions (Mongillo, Barak,
& Tsodyks, 2008) or by GluR1-dependent short-term
potentiation (Erickson, Maramara, & Lisman, 2010). The
information coded in these synaptic weight changes can
be translated back into active neural firing if the memory
is later reactivated by a retrieval cue (Nairne, 2002).
The idea that memory representations can be reactivated
during short-delay tests of STM is anticipated in neural-
network models of serial order recall (Botvinick & Plaut,
2006; Burgess & Hitch, 2006; Farrell & Lewandowsky,
2002; Burgess & Hitch, 1999) and in retrieved context mod-
els of memory search (Polyn et al., 2009; Sederberg et al.,
2008; Howard & Kahana, 2002). These models suggest an
interaction between two cognitive representations: a repre-
sentation of the memory item and a representation of the
encoding context. These two representations can influence
one another through synaptic weight changes in bidirec-
tional associations between the item and its context. When
an item is studied, an episodic memory is formed by linking
the item features to the currently active pattern of contex-
tual activity. The associations formed on the context-to-
item weights allow the context representation to serve as
a retrieval cue: If a particular context representation is re-
activated, it can then be used to revive the item represen-
tation(s) that co-occurred with that particular context state.
The reverse interaction, driven by the item-to-context asso-
ciations, provides retrieval of the context representation
that prevailed when that memory item was originally en-
countered. This latter process, described as mental time
travel (Tulving, 2002), is crucial for the perpetuation of the
free recall process but is incidental to the cued recall pro-
cess required by many tests of STM. Although these models
arose in an attempt to explain variability in free recall per-
formance, our present findings suggest that the memory
retrieval mechanisms that they propose may also provide
useful explanations for variability in cued recall performance
at short memory delays.
Another objection that could be raised against our con-
clusions is that they seem to be contradicted by the findings
Lewis-Peacock et al.
75
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
of sustained activity observed with electrophysiological
recordings from individual neurons in monkeys, the loss
of which has been thought to indicate disruption of STM
(Miller, Erickson, & Desimone, 1996; Miller & Desimone,
1994; Miller, Li, & Desimone, 1993). In contrast, our results
from MVPA of fMRI recordings in humans indicate that per-
sistent neural activation is not required for STM. One way
to reconcile the two sets of findings is to appeal to the as-
sumption that contents of STM are represented in the brain
by highly distributed and overlapping patterns of activity
(e.g., Haxby et al., 2001). Thus, the activity of individual
neurons is unlikely to accurately reflect a representation
that only exists in the distributed pattern of activity across
many neurons. A second consideration is that these previ-
ous studies confounded attention and STM, such that the
information to be remembered was the most task-relevant
information throughout the memory interval and therefore
was likely to be continuously attended to. The persistent
activity of individual neurons, which correlates with per-
formance in STM tasks, might instead reflect sustained
attention, a reinterpretation which would be consistent with
the present results.
The suggestion that LTM mechanisms support perfor-
mance during a test of short-term retention is not novel.
In dual-store models (Atkinson & Shiffrin, 1968; Waugh &
Norman, 1965), the contribution of LTM is thought to sup-
plement (and not replace) a STM system that is capable of
holding several items. Neural evidence for this idea comes
from neuroimaging and neuropsychological studies, which
have demonstrated that medial-temporal lobe structures
(known to be essential for LTM) also contribute to perfor-
mance on tests of short-term retention ( Jeneson, Mauldin,
Hopkins, & Squire, 2011; Jeneson, Mauldin, & Squire, 2010;
Hannula, Tranel, & Cohen, 2006; Nichols, Kao, Verfaellie,
& Gabrieli, 2006; Olson, Moore, Stark, & Chatterjee, 2006;
Olson, Page, Moore, Chatterjee, & Verfaellie, 2006). All
theories of STM assume a capacity of more than one item,
and typical estimates are around four (Cowan, 2001; Luck
& Vogel, 1997). In this study, we deliberately held the
overall memory load so small (two items at maximum)
that the capacity limits of STM would not be exceeded.
Therefore, on the basis of the ubiquitous assumption that
sustained activity is the neural correlate of maintenance
in STM, we would expect to observe persistent neural
representations for all memory items in our tasks. However,
our results demonstrate that only the item in the focus
of attention retained its active representation during the
delay period. In Experiment 2, the focus of attention de-
monstrably held two items at the same time, as shown by
high classifier evidence for both target items after encod-
ing, so it was not for lack of attentional capacity that only
one item was actively represented after the cue. Rather it
was the behavioral relevance of the item that determined
its activity status.
The present research was motivated by the family of
embedded component theories of STM, which charac-
terize the system enabling the short-term retention of
information as consisting of a central component of STM
(referred to here as the focus of attention) and a more
peripheral component (commonly referred to as “acti-
vated LTM”). However, the results that we have presented
here suggest that the label for the retention of information
outside the focus of attention might be a misnomer—it is
perhaps more accurately labeled “privileged LTM” because
this information is in a privileged state (i.e., it affects on-
going processing more strongly than does dormant infor-
mation in LTM) but is not supported by an active neural
trace. This study makes two important contributions to
the further refinement of these theories: (1) It provides
some of the first evidence (see also Nee & Jonides, 2008,
2011; Nee, Berman, Moore, & Jonides, 2008) that the dis-
tinction between the two components, which has been
proposed on the basis of behavioral evidence (Oberauer,
2002; Cowan, 1988), has a neural basis. (2) It maps the
time course of the neural signature of the removal of task-
irrelevant information from the focus of attention, showing
that it corresponds to the time course of the behavioral
signature of these processes (Oberauer, 2001, 2005). Inde-
pendent of the embedded component model, this study
demonstrates that the active neural signature of informa-
tion held in STM can be disrupted by redirecting attention
externally or internally, without sacrificing the short-term
retention of that information. These results raise questions
about the common view that persistent maintenance of
neural activity is required for short-term retention and
support an alternative interpretation: Delay period activ-
ity reflects the focus of attention, rather than the contents
of STM.
Acknowledgments
This research was funded by the National Institutes of Mental
Health grants R01 MH064498 (B. P.) and F31 MH085444 (J. L.-P.).
Reprint requests should be sent to Jarrod A. Lewis-Peacock,
Green Hall, Princeton University, Princeton, NJ 08540, or via
e-mail: jalewpea@princeton.edu.
Notes
1. What we refer to as the “focus of attention” is the broad focus
of attention (Cowan, 1995) that has a capacity limit of about four
items. This contrasts with a narrow focus of attention, consisting
of a single item, that is differentiated from the “direct access re-
gion” which can hold about four items (Oberauer, 2002). Our
data do not address the distinction between these constructs,
and therefore, we consistently imply the broader definition.
2. Decoding with voxels from the whole brain or only those
restricted to ITC produced qualitatively similar results. How-
ever, although classifier training on Phase 1 data in the PFC was
successful, decoding of Phase 3 data from this region failed to
produce interpretable results. PFC is thought to be a critical
neural substrate for cognitive control and the representation of
task demands (Miller & Cohen, 2001). Although the stimulus
materials were identical between the training task (Phase 1) and
the testing task (Phase 3), the cognitive demands of each task
were not (short-term recognition vs. short-term paired-associate
76
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
recognition). This may underlie the classifierʼs inability to gener-
alize from the training data to the testing data in PFC.
REFERENCES
Atkinson, R. C., & Shiffrin, R. M. (1968). Human memory:
A proposed system and its control processes. The
Psychology of Learning and Motivation: Advances
in Research and Theory, 2, 89–195.
Baddeley, A. D. (1986). Working memory. London:
Oxford University Press.
Bays, P. M., & Husain, M. (2008). Dynamic shifts of limited
working memory resources in human vision. Science,
321, 851–854.
Botvinick, M. M., & Plaut, D. C. (2006). Short-term memory
for serial order: A recurrent neural network model.
Psychological Review, 113, 201–233.
Burgess, N., & Hitch, G. J. (1999). Memory for serial order:
A network model of the phonological loop and its timing.
Psychological Review, 106, 551.
Burgess, N., & Hitch, G. J. (2006). A revised model of
short-term memory and long-term learning of verbal
sequences. Journal of Memory and Language, 55,
627–652.
Cameron, K. A., Haarmann, H. J., Grafman, J., & Ruchkin, D. S.
(2005). Long-term memory is the representational basis
for semantic verbal short-term memory. Psychophysiology,
42, 643–653.
Chun, M. M. (2011). Visual working memory as visual
attention sustained internally over time. Neuropsychologia,
49, 1407–1409.
Chun, M. M., Golomb, J. D., & Turk-Browne, N. B. (2011).
A taxonomy of external and internal attention. Annual
Review of Psychology, 62, 73–101.
Courtney, S. M., Ungerleider, L. G., Keil, K., & Haxby, J. V.
(1997). Transient and sustained activity in a distributed
neural system for human working memory. Nature,
386, 608–611.
Cowan, N. (1988). Evolving conceptions of memory storage,
selective attention, and their mutual constraints within
the human information-processing system. Psychological
Bulletin, 104, 163–191.
Cowan, N. (1995). Attention and memory: An integrated
framework. New York: Oxford University Press.
Cowan, N. (2001). The magical number 4 in short-term
memory: A reconsideration of mental storage capacity.
Behavioral and Brain Sciences, 24, 87–114.
Cowan, N. (2011). The focus of attention as observed in
visual working memory tasks: Making sense of competing
claims. Neuropsychologia, 49, 1401–1406.
Curtis, C. E., & DʼEsposito, M. (2003). Persistent activity in
the prefrontal cortex during working memory. Trends
in Cognitive Sciences, 7, 415–423.
DʼEsposito, M., Postle, B. R., Jonides, J., & Smith, E. E.
(1999). The neural substrate and temporal dynamics
of interference effects in working memory as revealed
by event-related functional MRI. Proceedings of the
National Academy of Sciences, U.S.A., 96, 7514–7519.
di Pellegrino, G., & Wise, S. P. (1993). Visuospatial versus
visuomotor activity in the premotor and prefrontal
cortex of a primate. Journal of Neuroscience, 13,
1227–1243.
Druzgal, T. J., & DʼEsposito, M. (2003). Dissecting contributions
of prefrontal cortex and fusiform face area to face working
memory. Journal of Cognitive Neuroscience, 15, 771–784.
Duda, R. O., Hart, P. E., & Stork, D. G. (2001). Pattern
classification (2nd ed.). New York: Wiley.
Erickson, M. A., Maramara, L. A., & Lisman, J. (2010).
A single brief burst induces glur1-dependent associative
short-term potentiation: A potential mechanism for
short-term memory. Journal of Cognitive Neuroscience,
22, 2530–2540.
Ericsson, K. A., & Kintsch, W. (1995). Long-term working
memory. Psychological Review, 102, 211–244.
Esterman, M., Chiu, Y. C., Tamber-Rosenau, B. J., & Yantis, S.
(2009). Decoding cognitive control in human parietal cortex.
Proceedings of the National Academy of Sciences, U.S.A.,
106, 17974.
Farrell, S., & Lewandowsky, S. (2002). An endogenous
distributed model of ordering in serial recall. Psychonomic
Bulletin & Review, 9, 59–79.
Fuster, J. M., & Alexander, G. E. (1971). Neuron activity
related to short-term memory. Science, 173, 652–654.
Gardiner, J. M., Craik, F. I. M., & Birstwistle, J. (1972). Retrieval
cues and release from proactive inhibition. Journal of
Verbal Learning and Verbal Behavior, 11, 778–783.
Gauthier, I., Skudlarski, P., Gore, J. C., & Anderson, A. W.
(2000). Expertise for cars and birds recruits brain areas
involved in face recognition. Nature Neuroscience, 3,
191–197.
Gazzaley, A. (2011). Influence of early attentional modulation
on working memory. Neuropsychologia, 49, 1410–1424.
Griffin, I. C., & Nobre, A. C. (2003). Orienting attention to
locations in internal representations. Journal of Cognitive
Neuroscience, 15, 1176–1194.
Haarmann, H., & Usher, M. (2001). Maintenance of semantic
information in capacity-limited item short-term memory.
Psychonomic Bulletin & Review, 8, 568–578.
Hannula, D. E., Tranel, D., & Cohen, N. J. (2006). The long
and the short of it: Relational memory impairments in
amnesia, even at short lags. Journal of Neuroscience, 26,
8352–8359.
Harrison, S. A., & Tong, F. (2009). Decoding reveals the
contents of visual working memory in early visual areas.
Nature, 458, 632–635.
Haxby, J. V., Gobbini, M. I., Furey, M. L., Ishai, A., Schouten,
J. L., & Pietrini, P. (2001). Distributed and overlapping
representations of faces and objects in ventral temporal
cortex. Science, 293, 2425–2430.
Haxby, J. V., Petit, L., Ungerleider, L. G., & Courtney, S. M.
(2000). Distinguishing the functional roles of multiple
regions in distributed neural systems for visual working
memory. Neuroimage, 11, 380–391.
Haynes, J. D., & Rees, G. (2006). Decoding mental states from
brain activity in humans. Nature Reviews Neuroscience, 7,
523–534.
Hebb, D. O. (1949). The organization of behavior:
A neuropsychological theory. New York: Wiley.
Holmes, C. J., Hoge, R., Collins, L., Woods, R., Toga, A. W.,
& Evans, A. C. (1998). Enhancement of magnetic resonance
images using registration for signal averaging. Computer
Assisted Tomography, 22, 324–333.
Howard, M. W., & Kahana, M. J. (2002). A distributed
representation of temporal context 1. Journal of
Mathematical Psychology, 46, 269–299.
Ikkai, A., & Curtis, C. E. (2011). Common neural mechanisms
supporting spatial working memory, attention and motor
intention. Neuropsychologia, 49, 1428–1434.
Jeneson, A., Mauldin, K. N., Hopkins, R. O., & Squire, L. R.
(2011). The role of the hippocampus in retaining relational
information across short delays: The importance of memory
load. Learning & Memory, 18, 301–305.
Jeneson, A., Mauldin, K. N., & Squire, L. R. (2010). Intact working
memory for relational information after medial-temporal
lobe damage. Journal of Neuroscience, 30, 13624–13629.
Lewis-Peacock et al.
77
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Kriegeskorte, N., Formisano, E., Sorger, B., & Goebel, R.
(2007). Individual faces elicit distinct response patterns
in human anterior temporal cortex. Proceedings of the
National Academy of Sciences, U.S.A., 104, 20600–20605.
Kriegeskorte, N., Goebel, R., & Bandettini, P. (2006).
Information-based functional brain mapping. Proceedings of
the National Academy of Sciences, U.S.A., 103, 3863–3868.
Kubota, K., & Niki, H. (1971). Prefrontal cortical unit activity
and delayed alternation performance in monkeys. Journal
of Neurophysiology, 34, 337–347.
Kuhl, B. A., Rissman, J., Chun, M. M., & Wagner, A. D.
(2011). Fidelity of neural reactivation reveals competition
between memories. Proceedings of the National Academy
of Sciences, U.S.A., 108, 5903–5908.
Lepsien, J., Thornton, I., & Nobre, A. C. (2011). Modulation
of working-memory maintenance by directed attention.
Neuropsychologia, 49, 1569–1577.
Lewis-Peacock, J. A., & Postle, B. R. (2008). Temporary
activation of long-term memory supports working memory.
Journal of Neuroscience, 28, 8765–8771.
Luck, S. J., & Vogel, E. K. (1997). The capacity of visual
working memory for features and conjunctions. Nature,
390, 279–280.
Martin, R. C., Wu, D., Freedman, M., Jackson, E. F., &
Lesch, M. (2003). An event-related fmri investigation
of phonological versus semantic short-term memory.
Journal of Neurolinguistics, 16, 341–360.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of
prefrontal cortex function. Annual Review of Neuroscience,
24, 167–202.
Miller, E. K., & Desimone, R. (1994). Parallel neuronal
mechanisms for short-term memory. Science, 263,
520–522.
Miller, E. K., Erickson, C. A., & Desimone, R. (1996).
Neural mechanisms of visual working memory in prefrontal
cortex of the macaque. Journal of Neuroscience, 16,
5154–5167.
Miller, E. K., Li, L., & Desimone, R. (1993). Activity of neurons
in anterior inferior temporal cortex during a short-term
memory task. Journal of Neuroscience, 13, 1460–1478.
Mongillo, G., Barak, O., & Tsodyks, M. (2008). Synaptic
theory of working memory. Science, 319, 1543–1546.
Monsell, S. (1978). Recency, immediate recognition memory,
and reaction-time. Cognitive Psychology, 10, 465–501.
Nairne, J. S. (2002). Remembering over the short-term:
The case against the standard model. Annual Review
of Psychology, 53, 53–81.
Nee, D. E., Berman, M. G., Moore, K. S., & Jonides, J. (2008).
Neuroscientific evidence about the distinction between
short- and long-term memory. Current Directions in
Psychological Science, 17, 102–106.
Nee, D. E., & Jonides, J. (2008). Neural correlates of access to
short-term memory. Proceedings of the National Academy
of Sciences, U.S.A., 105, 14228–14233.
Nee, D. E., & Jonides, J. (2011). Dissociable contributions
of prefrontal cortex and the hippocampus to short-term
memory: Evidence for a 3-state model of memory.
Neuroimage, 54, 1540–1548.
Newman, E. L., & Norman, K. A. (2010). Moderate excitation
leads to weakening of perceptual representations. Cerebral
Cortex, 20, 2760–2770.
Nichols, E. A., Kao, Y. C., Verfaellie, M., & Gabrieli, J. D.
(2006). Working memory and long-term memory for faces:
Evidence from fMRI and global amnesia for involvement
of the medial temporal lobes. Hippocampus, 16, 604–616.
Norman, K. A., Polyn, S. M., Detre, G. J., & Haxby, J. V.
(2006). Beyond mind-reading: Multi-voxel pattern analysis
of fMRI data. Trends in Cognitive Sciences, 10, 424–430.
Oberauer, K. (2001). Removing irrelevant information from
working memory: A cognitive aging study with the modified
Sternberg task. Journal of Experimental Psychology:
Learning, Memory, & Cognition, 27, 948–957.
Oberauer, K. (2002). Access to information in working
memory: Exploring the focus of attention. Journal
of Experimental Psychology: Learning, Memory, &
Cognition, 28, 411–421.
Oberauer, K. (2005). Control of the contents of working
memory—A comparison of two paradigms and two age
groups. Journal of Experimental Psychology: Learning,
Memory, & Cognition, 31, 714–728.
Olivers, C. N., & Eimer, M. (2011). On the difference between
working memory and attentional set. Neuropsychologia,
49, 1553–1558.
Olson, I. R., Moore, K. S., Stark, M., & Chatterjee, A. (2006).
Visual working memory is impaired when the medial
temporal lobe is damaged. Journal of Cognitive
Neuroscience, 18, 1087–1097.
Olson, I. R., Page, K., Moore, K. S., Chatterjee, A., &
Verfaellie, M. (2006). Working memory for conjunctions
relies on the medial temporal lobe. Journal of Neuroscience,
26, 4596–4601.
Pereira, F., Mitchell, T., & Botvinick, M. (2009). Machine
learning classifiers and fMRI: A tutorial overview.
Neuroimage, 45(1 Suppl.), S199–S209.
Poldrack, R. A. (2006). Can cognitive processes be inferred
from neuroimaging data? Trends in Cognitive Sciences,
10, 64–69.
Polyn, S. M., Natu, V. S., Cohen, J. D., & Norman, K. A.
(2005). Category-specific cortical activity precedes retrieval
during memory search. Science, 310, 1963–1966.
Polyn, S. M., Norman, K. A., & Kahana, M. J. (2009). A context
maintenance and retrieval model of organizational processes
in free recall. Psychological Review, 116, 129–156.
Postle, B. R. (2006). Working memory as an emergent property
of the mind and brain. Neuroscience, 139, 23–38.
Postle, B. R., Druzgal, T. J., & DʼEsposito, M. (2003). Seeking
the neural substrates of working memory storage. Cortex;
A Journal Devoted to the Study of the Nervous System
and Behavior, 39, 927–946.
Postle, B. R., Idzikowski, C., Della Salla, S., Logie, R. H., &
Baddeley, A. D. (2006). The selective disruption of spatial
working memory by eye movements. Quarterly Journal
of Experimental Psychology, 59, 100–120.
Quamme, J. R., Weiss, D. J., & Norman, K. A. (2010). Listening
for recollection: A multi-voxel pattern analysis of recognition
memory retrieval strategies. Frontiers in Human
Neuroscience, 4, 1–17.
Ranganath, C., Cohen, M. X., Dam, C., & DʼEsposito, M.
(2004). Inferior temporal, prefrontal, and hippocampal
contributions to visual working memory maintenance
and associative memory recall. Journal of Neuroscience,
24, 3917–3925.
Ranganath, C., DeGutis, J., & DʼEsposito, M. (2004).
Category-specific modulation of inferior temporal activity
during working memory encoding and maintenance.
Cognitive Brain Research, 20, 37–45.
Ruchkin, D. S., Grafman, J., Cameron, K., & Berndt, R. S.
(2003). Working memory retention systems: A state of
activated long-term memory. Behavioral and Brain
Sciences, 26, 709–777.
Sederberg, P. B., Howard, M. W., & Kahana, M. J. (2008).
A context-based theory of recency and contiguity in free
recall. Psychological Review, 115, 893–912.
Serences, J. T., Ester, E. F., Vogel, E. K., & Awh, E. (2009).
Stimulus-specific delay activity in human primary visual
cortex. Psychological Science, 20, 207–214.
78
Journal of Cognitive Neuroscience
Volume 24, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Sheth, B. R., & Shimojo, S. (2003). Signal strength determines
the nature of the relationship between perception and working
memory. Journal of Cognitive Neuroscience, 15, 173–184.
Shivde, G. S., & Thompson-Schill, S. L. (2004). Dissociating
semantic and phonological maintenance using fMRI. Cognitive,
Affective, & Behavioral Neuroscience, 4, 10–19.
Stokes, M. G. (2011). Top–down visual activity underlying VSTM
and preparatory attention. Neuropsychologia, 49, 1425–1427.
Takeda, M., Naya, Y., Fujimichi, R., Takeuchi, D., & Miyashita, Y.
(2005). Active maintenance of associative mnemonic signal
in monkey inferior temporal cortex. Neuron, 48, 839–848.
Tulving, E. (2002). Episodic memory: From mind to brain.
Annual Review of Psychology, 53, 1–25.
Vandenbroucke, A. R., Sligte, I. G., & Lamme, V. A. (2011).
Manipulations of attention dissociate fragile visual short-term
memory from visual working memory. Neuropsychologia,
49, 1559–1568.
Vogel, E. K., McCollough, A. W., & Machizawa, M. G. (2005).
Neural measures reveal individual differences in controlling
access to working memory. Nature, 438, 500–503.
Waugh, N. C., & Norman, D. A. (1965). Primary memory.
Psychological Review, 72, 89.
Wickens, D. D. (1970). Encoding categories of words-Empirical
approach to meaning. Psychological Review, 77, 1–15.
Woltz, D. J. (1996). Perceptual and conceptual priming in a
semantic reprocessing task. Memory and Cognition, 24,
429–440.
Woltz, D. J., & Was, C. A. (2006). Availability of related
long-term memory during and after attention focus in
working memory. Memory & Cognition, 34, 668–684.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
1
2
4
6
/
1
1
1
/
9
6
4
1
3
/
3
1
1
7
9
8
o
0
c
3
n
2
_
3
a
/
_
j
0
o
0
c
1
n
4
_
0
a
p
_
d
0
0
b
1
y
4
g
0
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Lewis-Peacock et al.
79