Neural Correlates of Morphological Decomposition
during Visual Word Recognition
Brian T. Gold1 and Kathleen Rastle2
Abstrait
& Considerable behavioral research has demonstrated that
the visual word recognition system is sensitive to morphol-
ogical structure. It has typically been assumed that analysis of
morphologically complex words occurs only when the mean-
ing of these words can be derived from the meanings of their
constituents (par exemple., hunter = hunt + er). Cependant, results from
recent behavioral research using the masked priming tech-
nique have demonstrated that morphological analysis can occur
at an earlier orthographic level, in cases in which the meanings
of complex words cannot be derived from their constituents
(par exemple., corner = corn + er). Ici, we combine the logic of behav-
ioral masked priming with the neurophysiological phenomenon
of functional magnetic resonance imaging priming suppression
to look for evidence of nonsemantic morphological priming at
the neural level. Both behavioral and functional magnetic res-
onance imaging results indicated priming effects associated
with the mere appearance of morphological structure (corner–
CORN). En outre, these effects were distinguishable from
lexical–semantic effects (bucket–PAIL) and orthographic effects
(brothel–BROTH). Three left-lateralized occipito-temporal re-
gions showed sensitivity to early morphological components of
visual word recognition. Two of these regions also showed
orthographic priming ((cid:1)BA 37, peak: (cid:2)48 (cid:2)60 (cid:2)17; (cid:1)BA 19,
peak: (cid:2)40 (cid:2)77 (cid:2)1), whereas one was sensitive only to morpho-
logical similarity between primes and targets ((cid:1)BA 19, peak:
(cid:2)37 (cid:2)67 (cid:2)7). These findings provide a neurobiological basis
for a purely structural morphemic segmentation mechanism op-
erating at early stages of visual word recognition. &
INTRODUCTION
Two well-established components of visual word recog-
nition involve visual form (orthographic) and meaning-
based (semantic) processes. The orthographic component
involves sensitivity to visual features of letter strings such
as sequential dependencies (Olson, Sage, Conners, & Rack,
1990). The semantic component involves sensitivity to the
meanings of letter strings that have a lexical status (Strain,
Patterson, & Seidenberg, 1995). In addition to having a vi-
sual form and an associated meaning, some words have an
internal structure, or morphology. Morphology is a sub-
field of grammar. Morphologically complex words are
those that can be decomposed into simple meaningful sub-
units called morphemes. Morphemes consist of words or
meaningful parts of words (par exemple., affixes) that cannot be bro-
ken down further into smaller meaningful parts. For ex-
ample, the word ‘‘cleaner’’ can be segmented into the
stem ‘‘clean’’ and the suffix –er, providing an agentive func-
tion (a person who cleans). There is now considerable
agreement that morphologically complex words are some-
how decomposed in visual word recognition and analyzed
in terms of their constituent morphemes (voir, par exemple., Frost,
Grainger, & Rastle, 2005).
The dominant view of morphological processing in
visual word recognition asserts that it is a high-level phe-
1University of Kentucky, 2University of London
‘‘unclean,’’
nomenon constrained by semantic knowledge (Giraudo
& Grainger, 2000; Plaut & Gonnerman, 2000; Rueckl &
Raveh, 1999). Both localist and distributed-connectionist
expressions of this perspective are based on the notion
that morphological relationships provide an important
element of structure to the largely arbitrary orthography-
to-semantics mapping. These theories posit that in learn-
ing the orthography–semantics mapping,
letter strings
that consistently share orthography and meaning (tel
‘‘cleaner,’’
as morphological relatives, par exemple.,
‘‘cleanliness,’’ ‘‘cleanness’’) will become represented in
terms of their constituents (see Rastle, Davis, Marslen-
Wilson, & Tyler, 2000, for discussion). Morphological
decomposition is therefore proposed to occur only in
cases in which morphologically complex words are seman-
tically transparent (c'est à dire., in cases in which their meanings
can be derived from the meanings of their constituents;
par exemple., a ‘‘hunter’’ is someone who hunts). Decomposition
is not expected in pseudomorphological constructions in
which the relationship between the full form and the
stem is semantically opaque (par exemple., a ‘‘corner’’ is not some-
one who corns). These theories have been supported by
substantial research demonstrating that morphological
priming effects are obtained only when primes and targets
have a semantic relationship (par exemple., Meunier & Longtin,
2007; Rastle et al., 2000; Marslen-Wilson, Tyler, Waksler, &
Older, 1994). This support has been restricted, cependant,
D 2007 Massachusetts Institute of Technology
Journal des neurosciences cognitives 19:12, pp. 1983–1993
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
un
/
d
m
e
je
d
t
F
p
r
o
r
m
c
.
h
s
je
p
je
v
d
e
je
r
r
e
c
c
h
t
.
un
m
je
r
e
.
d
c
toi
o
m
o
/
c
j
n
o
un
c
r
n
t
je
c
/
un
e
r
–
p
t
d
je
c
1
je
9
e
1
–
2
p
1
d
9
F
8
/
3
1
9
1
/
9
1
3
6
2
1
/
2
1
8
9
8
o
3
c
/
n
1
2
7
0
5
0
6
7
5
1
0
9
1
/
1
j
2
o
1
c
9
n
8
.
3
2
p
0
d
0
7
b
.
oui
1
g
9
toi
.
e
1
s
2
t
.
o
1
n
9
0
8
8
3
S
.
e
p
p
d
F
e
m
b
b
oui
e
r
g
2
toi
0
e
2
s
3
t
/
j
t
.
.
.
.
F
.
o
n
1
8
M.
un
oui
2
0
2
1
to paradigms thought to reflect lexical–semantic levels of
the language system such as cross-modal priming and vi-
sual priming with fully visible primes.
Very Early Morphological Decomposition
Recent results using the masked priming technique have
suggested a different view, cependant. In a typical masked
priming experiment, a briefly presented (<50 msec) lower-
case prime is sandwiched between a forward mask (e.g.,
######) and an uppercase target that acts as back-
ward (Forster & Davis, 1984). Several behavioral stud-
ies and, more recently, ERP studies using this paradigm
have reported robust priming for pseudomorphological
pairs (e.g., corner–CORN) illegal novel morpholog-
ical pairs spendical–SPEND) equivalent in
magnitude to the observed semantically
transparent morphological hunter–HUNT).
Critically, these effects cannot be ascribed to
simple orthographic similarity because they are not ob-
served prime–target have nonmorpho-
logical form relationship brothel–BROTH; –el never
functions suffix in English; Lavric, Clapp, Rastle,
2007; Longtin Meunier, 2005; Rastle, New,
2004; Longtin, Segui, Halle´, 2003; Rastle 2003).
These findings challenged dominant view of
morphological decomposition illustrate that
morphological necessarily dependent up-
on semantic transparency. Instead, results from these
masked suggested existence of
a mechanism within visual word recognition system
that operates at early sublexical level,
serving decompose any letter string appears mor-
phologically complex (Longtin Rastle
et al., 2000, 2004; et 2003). However, little is
known about neural bases of mor-
phological segmentation.
fMRI Priming Suppression
In present study, we combine logic behav-
ioral masked with neurophysiological phe-
nomenon functional magnetic resonance imaging
(fMRI) suppression look evidence pseudomor-
phological neural
level. fMRI
experiments reduced blood oxygen level-
dependent response (fMRI suppression) during trials in-
volving repeated compared stimuli (reviewed in
Wiggs Martin, 1998). Importantly, Dehaene al. (2001)
showed phenomenon fMRI also
accompanies short-term which tar-
get preceded immediately by identical (iden-
tity priming), even under conditions which
subjects were unaware primes. Despite short du-
ration primes (29 relatively small
behavioral effect size ((cid:1)16 msec), fMRI
suppression was extrastriate fusiform
cortices left hemisphere. Although identity
priming likely reflected shared contributions of
several different linguistic processes, study raised the
possibility correlates pro-
cesses could fractionated through suppression
studies vary kind be-
tween target.
Only one has explored
morphological recog-
nition. Devlin, Jamison, Matthews, Gonnerman (2004)
compared when targets preceded
by sharing (hunter–HUNT), orthographic
(craven–CRAVE), or (imitate–COPY). Neural entirely overlapped orthograph-
ic (in posterior occipito-temporal cortex) and
semantic lateral middle temporal cortex).
Devlin (2004) argued supported a
version theory process-
ing, namely, reflect conver-
gence similarity. However,
the condition Devlin consisted largely
of pseudomorphological corner–CORN). This
study did separate brothel–BROTH)
and into conditions
(as 2004), making its equally
consistent notion morphemic segmen-
tation procedure independently semantics.
Thus, despite mounting support form
of indepen-
dent lexical–semantic known about
the segmentation mechanism.
The extent morphological
decomposition overlaps>
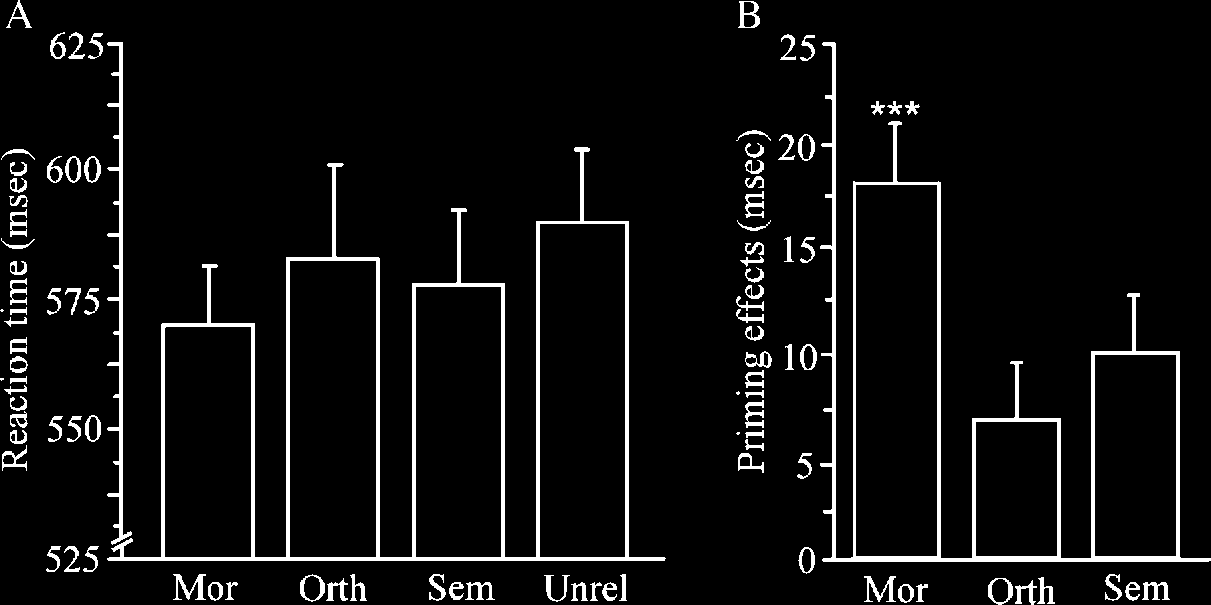
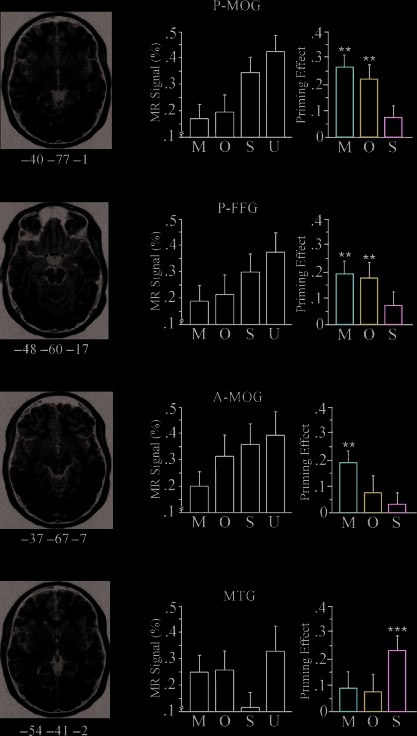
related) for the M, Ô, and S conditions. Note: **p < .01, ***p < .001.
Gold and Rastle
1989
In contrast, orthographic priming was observed in two
more posterior left hemisphere regions: a posterior
portion of the fusiform gyrus (P-FFG; (cid:1)BA 37), and an
extrastriate region in a posterior portion of the middle
occipital gyrus (P-MOG; (cid:1)BA 19). Both of these regions
have been implicated in the processing of visual form.
For example, neuropsychological evidence indicates that
damage to large portions of the left occipito-temporal
cortex can be associated with pure alexia, a unimodal
deficit of word reading, with sparing of writing and
auditory word comprehension (Binder & Mohr, 1992;
Damasio & Damasio, 1983; Dejerine, 1892). More spe-
cifically, recent fMRI results have demonstrated a prom-
inent P-FFG response in tasks associated with the
processing of abstract letter units (Cohen et al., 2000,
2002) and have yielded neural priming effects in the
P-FFG for an orthographic priming condition that includ-
ed opaque morphological pairs (corner–CORN; Devlin
et al., 2004). Similarly, the extrastriate region showing
orthographic priming in our study (P-MOG) has shown
greater response to alphabetic stimuli compared to
checkerboards (Cohen et al., 2003) and significant acti-
vation related to orthographic components of visual
word recognition (Rumsey et al., 1997). In addition,
neural orthographic priming using partial word primes
(ATL–ATLAS) has previously been demonstrated in the
P-MOG (Gold et al., 2006).
Neural morphological priming did not overlap lexical–
semantic priming but did overlap substantially with
orthographic priming. Morphological priming was ob-
served in both of the regions showing orthographic
priming: P-FFG and P-MOG. There were voxels that
showed unique morphological priming effects within
each of these two regions. However, these morpholog-
ical effects consisted of voxel clusters that were spatially
contiguous with orthographic priming effects. To be
conservative, these spatially contiguous clusters are re-
ported as a single cluster, common to orthographic and
morphological priming (see Table 1). The finding that
morphological priming did not overlap lexical–semantic
priming but did show substantial overlap with ortho-
graphic priming provides direct functional neuroanato-
mic evidence supporting the existence of a morphological
segmentation mechanism that occurs independently of
lexical–semantic processes, at an early structural
level
(Rastle et al., 2004; Longtin et al., 2003; Rastle & Davis,
2003). In addition, results also provide evidence that this
early morphological decomposition process cannot be
explained on the basis of orthographic (letter) similarity
alone because there was one region that showed priming
unique to the morphological condition.
Unique morphological priming was observed in the
A-MOG ((cid:1)BA 19). Unlike the spatially contiguous ortho-
graphic and morphological voxel clusters observed in
the more posterior extrastriate region (P-MOG), the de-
activation cluster in the A-MOG was clearly spatially
distinct from that in the P-MOG, with peak decreases
between the two regions separated by 1 cm. The A-MOG
has been activated during many tasks involving high-
level visual processing,
including, but not limited to,
object processing (Grill-Spector et al., 1999) and face
processing (Gorno-Tempini et al., 1998). The A-MOG
has also been activated during the naming of very rapidly
presented words (Dehaene et al., 2001). The region there-
fore appears to be involved in numerous high-level visual
processes and is not specialized for morphological pro-
cessing. The important point with respect to the goals
of the present study is that this region showed neural
priming unique to the morphological condition in a sin-
gle group of subjects who completed morphological, or-
thographic, and lexical–semantic conditions.
It is interesting to consider why the A-MOG showed
morphological but not orthographic priming in the pres-
ent study. One possibility is that this region did not
show orthographic priming due to the small behavioral
priming effect observed in this condition. However, the
correlational analyses that we reported, along with the
fact that robust neural orthographic priming was ob-
served in two other regions, would seem to rule this
possibility out. More likely is the possibility that the
posterior-to-anterior, orthographic–morphological gra-
dient of neural priming effects observed in this study
reflects a general organizing principle of the occipito-
temporal cortex: that the processing stream tends to
proceed in the anterior direction as visual linguistic op-
erations become more complex and abstract (McCarthy,
Nobre, Bentin, & Spencer, 1995; Halgren, Baudena, Heit,
Clarke, & Marinkovic, 1994; Nobre, Allison, & McCarthy,
1994). This notion is consistent with evidence from
intracranial recordings that have suggested sensitivity
to orthographic form in the posterior occipito-temporal
cortex and sensitivity to lexical–semantic material in the
mid-to-anterior occipito-temporal cortex (Nobre et al.,
1994). Morphemes are clusters of letters that serve as
functional units within words, and can therefore be re-
garded in a hierarchical
framework as occupying a
greater level of abstraction than letters themselves.
One interesting aspect of our data is that no fMRI
suppression effects were observed in the left inferior pre-
frontal cortex (LIPC), despite neuropsychological and func-
tional neuroimaging research suggesting that this region
may be involved in morphological processing (Tyler,
Marslen-Wilson, & Stamatakis, 2005; Tyler, Stamatakis,
Post, Randall, & Marslen-Wilson, 2005; Miceli et al., 2002).
However, these studies all employed tasks that permit
strategic processing, such as explicit morphological deci-
sion tasks or priming designs in which primes were sub-
ject to conscious appreciation, whereas our study used
a masked priming paradigm in which participants were
unaware of primes. Together, these findings raise the
possibility that the LIPC contributes more prominently
to later more strategic components of morphological
analysis than earlier more automatic components. Future
research will be required to explore this issue directly,
1990
Journal of Cognitive Neuroscience
Volume 19, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
9
e
1
-
2
p
1
d
9
f
8
/
3
1
9
1
/
9
1
3
6
2
1
/
2
1
8
9
8
o
3
c
/
n
1
2
7
0
5
0
6
7
5
1
0
9
1
/
1
j
2
o
1
c
9
n
8
.
3
2
p
0
d
0
7
b
.
y
1
g
9
u
.
e
1
s
2
t
.
o
1
n
9
0
8
8
3
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
perhaps by studying morphological priming effects at
different SOAs.
The pattern of fMRI suppression effects observed in
the present study provides some neurophysiological
constraints on theories of morphological segmentation
during visual word recognition. Prime–target pairs with a
pseudomorphological relationship (e.g., corner–CORN)
yielded significant neural priming effects that (a) did not
overlap at all with neural priming effects observed for
pairs with a lexical–semantic relationship; (b) over-
lapped considerably with neural priming effects ob-
served for pairs with an orthographic relationship; and
(c) were unique to one region of the extrastriate cortex.
These data are inconsistent with any morpho-semantic
theory (whether localist or connectionist) in which mor-
phology is described as a characterization of the form-
meaning mapping (e.g., Giraudo & Grainger, 2000; Plaut
& Gonnerman, 2000; Rueckl & Raveh, 1999) and in which
morphological decomposition arises only in cases in
which morphologically complex words are semantically
related to their stems (Marslen-Wilson et al., 1994). In-
stead, these data support morpho-orthographic theo-
ries in which the sublexical orthographic representations
used in visual word processing are themselves morpho-
logically structured (Rastle et al., 2004; see also Taft,
1994; Seidenberg, 1987).
The early form of morphological segmentation that
we have observed has been characterized in both classical-
localist and distributed-connectionist frameworks, so it
is worthwhile to consider briefly whether our data favor
one or the other of these perspectives. Localist theories
account for this type of decomposition in terms of an
explicit level of morphological representation that re-
sides between representations of letters and ortho-
graphic representations of whole words (Rastle et al.,
2004; Taft, 1994). According to these theories, represen-
tations of ‘‘corner’’ would overlap those of ‘‘CORN’’ in
both letter units and morpheme units, whereas repre-
sentations of ‘‘brothel’’ would overlap those of ‘‘BROTH’’
only in letter units. This theory would therefore predict
that morphological neural priming effects should overlap
with pure orthographic neural priming effects in some
brain regions due to shared letter representations of
prime–target pairs in each condition. However, classical-
localist theories would also predict morphological neural
priming effects in brain regions distinct from those
showing pure orthographic neural priming effects be-
cause morphological prime–target pairs share overlap in
higher-level morphemic representations, whereas ortho-
graphic prime–target pairs do not. In general, this is the
pattern of neural priming effects that we observed.
By contrast, distributed-connectionist theories of non-
semantic morphological segmentation eschew the no-
tion of explicit morphological representations. These
theories argue instead that representations of ortho-
graphic form may develop a morphological structure as
a result of the distinctive bigram and trigram frequency
contours that characterize morphologically complex
words (Rastle et al., 2004; Seidenberg, 1987).1 Networks
sensitive to this characteristic of morphologically com-
plex words may come to represent these words compo-
nentially at the orthographic level, such that learned
orthographic representations of ‘‘corner’’ and ‘‘CORN’’
would overlap to a greater degree than learned ortho-
graphic representations of ‘‘brothel’’ and ‘‘BROTH.’’ As
with localist theories, then, connectionist theories
would predict that morphological neural priming effects
should overlap with pure orthographic neural priming
effects.
However, at least some (but not necessarily all) con-
nectionist theories would also predict that neural mor-
phological priming should be of a greater magnitude
than neural orthographic priming in ‘‘orthographic re-
gions’’ due to the greater orthographic similarity of
morphological pairs (corner and CORN) than pure
orthographic pairs (brothel and BROTH) that these
theories posit. We found no evidence for such graded
neural priming effects in ‘‘orthographic regions’’ in the
present study. Rather, similar sized neural morphologi-
cal and orthographic effects were observed in regions
that showed overlapping neural priming effects. Finally,
connectionist theories would also seem to have difficulty
explaining neural morphological priming in a region not
showing orthographic priming, a finding that was ob-
served in the present study. On balance, therefore, it
would appear that the localist theory of nonsemantic
morphological segmentation provides a closer descrip-
tion to the pattern of neural priming effects that we
observed.
In summary, the present study provides the first
functional neuroanatomic evidence supporting the exis-
tence of an early form of morphological decomposition
in visual word processing that operates on any letter
string that has the appearance of morphological com-
plexity. Three left-lateralized occipito-temporal regions
showed sensitivity to early morphological components
of visual word recognition. Two of these regions also
showed orthographic priming ((cid:1)BA 37, peak: (cid:2)48 (cid:2)60
(cid:2)17; (cid:1)BA 19, peak: (cid:2)40 (cid:2)77 (cid:2)1), whereas one was
sensitive only to morphological similarity between
primes and targets ((cid:1)BA 19, peak: (cid:2)37 (cid:2)67 (cid:2)7). Brain
regions sensitive to morphological structure were dis-
tinct from those sensitive to lexical–semantic similarity
((cid:1)BA 21, peak: (cid:2)54 (cid:2)41 (cid:2)2). These findings provide a
neurobiological basis for a purely structural morpholog-
ical segmentation mechanism operating at early stages of
visual word recognition, providing support for morpho-
orthographic theories of morphology (Rastle et al., 2004;
Longtin et al., 2003).
Acknowledgments
This research was supported by National Institutes of Health
for help with data
grant DC007315. We thank Jeff Covell
Gold and Rastle
1991
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
9
e
1
-
2
p
1
d
9
f
8
/
3
1
9
1
/
9
1
3
6
2
1
/
2
1
8
9
8
o
3
c
/
n
1
2
7
0
5
0
6
7
5
1
0
9
1
/
1
j
2
o
1
c
9
n
8
.
3
2
p
0
d
0
7
b
.
y
1
g
9
u
.
e
1
s
2
t
.
o
1
n
9
0
8
8
3
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
f
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
collection and Drs. Anders Andersen, David Powell, and two
anonymous reviewers for helpful comments.
Reprint requests should be sent to Brian T. Gold, Department
of Anatomy and Neurobiology, University of Kentucky School
of Medicine, Lexington, KY 40536-0298, or via e-mail: brian.
gold@uky.edu.
Note
1. These words typically exhibit a ‘‘trough’’ pattern in which
bigram and trigram frequencies are much higher within mor-
phemic units than across morphemic boundaries (see Rastle
et al., 2004).
REFERENCES
Dejerine, J. (1892). Contribution a` l’e´tude anatomo-
pathologique et clinique des diffe´rentes varie´te´s de ce´cite´
verbale. Me´moires de la Socie´te´ de Biologie, 4, 61–90.
Devlin, J. T., Jamison, H. L., Matthews, P. M., & Gonnerman,
L. M. (2004). Morphology and the internal structure of
words. Proceedings of the National Academy of Sciences,
U.S.A., 101, 14984–14988.
Forster, K. I., & Davis, C. (1984). Repetition priming and
frequency attenuation in lexical access. Journal of
Experimental Psychology: Learning, Memory, & Cognition,
10, 680–698.
Frost, R., Grainger, J., & Rastle, K. (2005). Current issues in
morphological processing. New York: Psychology Press.
Giraudo, H., & Grainger, J. (2000). Effects of prime word
frequency and cumulative root frequency in masked
morphological priming. Language and Cognitive Processes,
15, 421–444.
Glover, G. H. (1999). Deconvolution of impulse response in
event-related BOLD fMRI. Neuroimage, 9, 416–429.
Baayen, R. H., Piepenbrock, R., & van Run, H. (1993). The
Gold, B. T., Balota, D. A., Jones, S. J., Powell, D. K., Smith,
CELEX lexical database [CD-ROM]. Philadelphia: University
of Pennsylvania, Linguistic Data Consortium.
Binder, J. R., Frost, J. A., Hammeke, T. A., Cox, R. W., Rao, S. M.,
& Prieto, T. (1997). Human brain language areas identified
by functional magnetic resonance imaging. Journal of
Neuroscience, 17, 353–362.
Binder, J. R., & Mohr, J. P. (1992). The topography of callosal
reading pathways. A case–control analysis. Brain, 115,
1807–1826.
Burgess, C., & Livesay, K. (1998). The effect of corpus size in
predicting reaction time in a basic word recognition task:
Moving on from Kucera and Francis. Behavior Research
Methods, Instruments & Computers, 30, 272–277.
Cohen, L., Dehaene, S., Naccache, L., Lehericy, S., Dehaene-
Lambertz, G., Henaff, M., et al. (2000). The visual word form
area: Spatial and temporal characterization of an initial stage
of reading in normal subjects and posterior split-brain
patients. Brain, 123, 291–307.
Cohen, L., Lehericy, S., Chochon, F., Lemer, C., Rivard, S.,
& Dehaene, S. (2002). Language-specific tuning of visual
cortex? Functional properties of the visual word form area.
Brain, 125, 1054–1069.
Cohen, L., Martinaud, O., Lemer, C., Lehericy, S., Samson, Y.,
Obadia, M., et al. (2003). Visual word recognition in the
left and right hemispheres: Anatomical and functional
correlates of peripheral alexias. Cerebral Cortex, 12,
1313–1333.
Copland, D. A., de Zubicaray, G. I., McMahon, K., Wilson, S. J.,
Eastburn, M., & Chenery, H. J. (2003). Brain activity during
automatic semantic priming revealed by event-related functional
magnetic resonance imaging. Neuroimage, 20, 302–310.
Cox, R. W. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers
and Biomedical Research, 29, 162–173.
Cox, R. W., & Jesmanowicz, A. (1999). Real-time 3D image
registration for functional MRI. Magnetic Resonance in
Medicine, 42, 1014–1018.
Dale, A. M. (1999). Optimal experimental design for event-
related fMRI. Human Brain Mapping, 8, 109–114.
Damasio, A. R., & Damasio, H. (1983). The anatomic basis of
pure alexia. Neurology, 33, 1573–1583.
Davis, M. H. (2004). Units of representation in visual word
recognition. Proceedings of the National Academy of
Sciences, 101, 14687–14688.
Dehaene, S., Naccache, L., Cohen, L., Le Bihan, D., Mangin,
J. F., Poline, J. B., et al. (2001). Cerebral mechanisms of word
masking and unconscious repetition priming. Nature
Neuroscience, 4, 752–758.
C. D., & Andersen, A. H. (2006). Dissociation of automatic
and strategic lexical–semantics: Functional magnetic
resonance imaging evidence for differing roles of multiple
frontotemporal regions. Journal of Neuroscience, 26,
6523–6532.
Gold, B. T., Balota, D. A., Kirchhoff, B. A., & Buckner, R. L.
(2005). Common and dissociable activation patterns
associated with controlled semantic and phonological
processing: Evidence from fMRI adaptation. Cerebral
Cortex, 15, 1438–1450.
Gold, B. T., & Buckner, R. L. (2002). Common prefrontal
regions co-activate with dissociable posterior regions during
controlled semantic and phonological tasks. Neuron, 35,
803–812.
Gorno-Tempini, M. L., Price, C. J., Josephs, O., Vandenberghe,
R., Cappa, S. F., Kapur, N., et al. (1998). The neural systems
sustaining face and proper-name processing. Brain, 121,
2103–2118.
Grill-Spector, K., Kushnir, T., Edelman, S., Avidan, G., Itzchak,
Y., & Malach, R. (1999). Differential processing of objects
under various viewing conditions in the human lateral
occipital complex. Neuron, 24, 187–203.
Halgren, E., Baudena, P., Heit, G., Clarke, J. M., & Marinkovic,
K. (1994). Spatio-temporal stages in face and word
processing: 1. Depth-recorded potentials in the human
occipital, temporal and parietal lobes. Journal of Physiology
(Paris), 88, 1–50.
Hart, J., Kraut, M. A., Kremen, S., Soher, B., & Gordon, B.
(2000). Neural substrates of orthographic lexical access as
demonstrated by functional brain imaging. Neuropsychiatry,
Neuropsychology, and Behavioral Neurology, 13, 1–7.
Lavric, A., Clapp, A., & Rastle, K. (2007). ERP evidence of
morphological analysis from orthography: A masked
priming study. Journal of Cognitive Neuroscience, 19,
866–877.
Longtin, C. M., & Meunier, F. (2005). Morphological
decomposition in early visual word processing. Journal of
Memory and Language, 53, 26–41.
Longtin, C. M., Segui, J., & Halle´, P. A. (2003). Morphological
priming without morphological relationship. Language and
Cognitive Processes, 18, 313–334.
Marslen-Wilson, W. D., Tyler, L. K., Waksler, R., & Older, L.
(1994). Morphology and meaning in the English mental
lexicon. Psychological Review, 101, 3–33.
McCarthy, G., Nobre, A. D., Bentin, S., & Spencer, D. D. (1995).
Language-related field potentials in the anterior–medial
temporal lobe: I. Intracranial distribution and neural
generators. Journal of Neuroscience, 15, 1080–1089.
1992
Journal of Cognitive Neuroscience
Volume 19, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
9
e
1
-
2
p
1
d
9
f
8
/
3
1
9
1
/
9
1
3
6
2
1
/
2
1
8
9
8
o
3
c
/
n
1
2
7
0
5
0
6
7
5
1
0
9
1
/
1
j
2
o
1
c
9
n
8
.
3
2
p
0
d
0
7
b
.
y
1
g
9
u
.
e
1
s
2
t
.
o
1
n
9
0
8
8
3
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Meunier, F., & Longtin, C. M. (2007). Morphological
decomposition and semantic integration in word processing.
Journal of Memory and Language, 56, 457–471.
Miceli, G., Turriziani, P., Caltagirone, C., Capasso, R.,
Tomaiuolo, F., & Caramazza, A. (2002). The neural correlates
of grammatical gender: An fMRI investigation. Journal of
Cognitive Neuroscience, 15, 618–628.
Naeser, M. A., Helm-Estabrooks, N., Haas, G., Auerbach, S., &
Srinivasan, M. (1987). Relationship between lesion extent in
‘‘Wernicke’s area’’ on computed tomographic scan and
predicting recovery of comprehension in Wernicke’s
aphasia. Archives of Neurology, 44, 73–82.
Nobre, A. C., Allison, T., & McCarthy, G. (1994). Word
recognition in the human inferior temporal lobe. Nature,
372, 260–263.
Olson, R., Wise, B., Conners, F., & Rack, J. (1990).
Organization, heritability, and remediation of component
word recognition and language skills in disabled readers. In
T. H. Carr & B. A. Levy (Eds.), Reading and its development:
Component skills approaches (pp. 261–322). San Diego:
Academic Press.
Plaut, D. C., & Gonnerman, L. M. (2000). Are non-semantic
morphological priming effects incompatible with a
distributed connectionist approach to language processing?
Language and Cognitive Processes, 15, 445–485.
Rastle, K., & Davis, M. H. (2003). Reading morphologically-
complex words: Some thoughts from masked priming. In
S. Kinoshita & S. J. Lupker (Eds.), Masked priming: State of
the art. Hove: Psychology Press.
Rastle, K., Davis, M. H., Marslen-Wilson, W. D., & Tyler, L. K.
(2000). Morphological and semantic effects in visual word
recognition: A time-course study. Language and Cognitive
Processes, 15, 507–537.
Rastle, K., Davis, M. H., & New, B. (2004). The broth in my
brother’s brothel: Morpho-orthographic segmentation in
visual word recognition. Psychonomic Bulletin & Review,
11, 1090–1098.
Rueckl, J. G., & Raveh, M. (1999). The influence of
morphological regularities on the dynamics of a
connectionist network. Brain and Language, 68, 110–117.
Rumsey, J. M., Horwitz, B., Donohue, B. C., Nace, K., Maisog,
J. M., & Andreason, P. (1997). Phonological and orthographic
components of word recognition. Brain, 120, 739–759.
Seidenberg, M. S. (1987). Sublexical structures in visual word
recognition: Access units or orthographic redundancy? In M.
Coltheart (Ed.), Attention and performance: 12. The
psychology of reading. Hillsdale, NJ: Erlbaum.
Strain, E., Patterson, K. E., & Seidenberg, M. S. (1995).
Semantic influences on word recognition. Journal of
Experimental Psychology: Learning, Memory, & Cognition,
21, 1140–1154.
Taft, M. (1994). Interactive activation as a framework for
understanding morphological processing. Language and
Cognitive Processes, 9, 271–294.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic
atlas of the human brain. Stuttgart: Thieme.
Tyler, L. K., Marslen-Wilson, W. D., & Stamatakis, E. A. (2005).
Differentiating lexical form, meaning, and structure in the
neural language system. Proceedings of the National
Academy of Sciences, 102, 8375–8380.
Tyler, L. K., Stamatakis, E. A., Post, B., Randall, B., & Marslen-
Wilson, W. D. (2005). Temporal and frontal systems involved
in speech processing: An fMRI study of the past tense
processing. Neuropsychologia, 43, 1963–1974.
Vandenberghe, R., Price, C., Wise, R., Josephs, O., & Frackowiak,
R. S. J. (1996). Functional anatomy of a common semantic
system for words and pictures. Nature, 383, 254–256.
Van Essen, D. C., Dickson, J., Harwell, J., Hanlon, D., Anderson,
C. H., & Drury, H. A. (2001). An integrated software system
for surface-based analyses of cerebral cortex. Journal of the
American Medical Informatics Association, 41, 1359–1378.
Wiggs, C. L., & Martin, A. (1998). Properties and mechanisms
of perceptual priming. Current Opinion in Neurobiology,
8, 227–233.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
9
e
1
-
2
p
1
d
9
f
8
/
3
1
9
1
/
9
1
3
6
2
1
/
2
1
8
9
8
o
3
c
/
n
1
2
7
0
5
0
6
7
5
1
0
9
1
/
1
j
2
o
1
c
9
n
8
.
3
2
p
0
d
0
7
b
.
y
1
g
9
u
.
e
1
s
2
t
.
o
1
n
9
0
8
8
3
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
f
.
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Gold and Rastle
1993