Memory Contextualization: The Role of Prefrontal Cortex
in Functional Integration across Item and
Context Representational Regions
Wei Zhang1,2, Vanessa A. van Ast3, Floris Klumpers1,2, Karin Roelofs1,2*,
and Erno J. Hermans1,4*
Abstrait
■ Memory recall is facilitated when retrieval occurs in the orig-
inal encoding context. This context dependency effect likely re-
sults from the automatic binding of central elements of an
experience with contextual features (c'est à dire., memory “contextuali-
zation”) during encoding. Cependant, despite a vast body of re-
search investigating the neural correlates of explicit associative
mémoire, the neural interactions during encoding that predict
implicit context-dependent memory remain unknown. Twenty-
six participants underwent fMRI during encoding of salient stim-
uli (faces), which were overlaid onto unique background images
(contexts). To index subsequent context-dependent memory,
face recognition was tested either in intact or rearranged con-
texts, after scanning. Enhanced face recognition in intact relative
to rearranged contexts evidenced successful memory contextu-
alization. Overall subsequent memory effects (brain activity pre-
dicting whether items were later remembered vs. forgotten)
were found in the left inferior frontal gyrus (IFG) and right
amygdala. Effective connectivity analyses showed that stronger
context-dependent memory was associated with stronger cou-
pling of the left IFG with face- and place-responsive areas, les deux
within and between participants. Our findings indicate an impor-
tant role for the IFG in integrating information across wide-
spread regions involved in the representation of salient items
and contextual features. ■
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
un
_
0
1
2
1
8
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
Context is essential for memory retrieval. It is well estab-
lished that memories are easier to recall when retrieval
occurs in a context that resembles the original encoding
contexte (van Ast, Cornelisse, Meeter, & Kindt, 2014; van
Ast, Cornelisse, Meeter, Joëls, & Kindt, 2013; Forgeron &
Vela, 2001; Godden & Baddeley, 1975; Tulving & Thomson,
1973). Memory contextualization, in which an event and
its context are automatically bound together into one inte-
grated representation during encoding, is vital for the
subsequent retrieval of relevant memories in specific situa-
tion (van Ast et al., 2013, 2014; Liberzon & Sripada, 2008).
Inversement, the inability to properly contextualize infor-
mation has been linked to fragmentation of episodic
memories and overgeneralization of (fear) memories that
are characteristic of, par exemple, posttraumatic stress dis-
order (PTSD; Quaedflieg et al., 2015; Acheson, Gresack, &
Risbrough, 2012; Meyer et al., 2012; Brewin, Gregory, Lipton,
& Bourgeois, 2010; Liberzon & Sripada, 2008; Ehlers & Clark,
2000) or schizophrenia (Talamini, de Haan, Nieman,
1Donders Institute for Brain, Cognition, and Behaviour, 2Behav-
ioural Science Institute, Radbound University, 3Université de
Amsterdam, 4Radboud university medical center
*These authors contributed equally to this article.
© 2017 Massachusetts Institute of Technology
Linszen, & Meeter, 2010). Cependant, despite its relevance
for general healthy memory function and clinical memory
overgeneralization, the brain mechanisms underlying mem-
ory contextualization—the process whereby automatic en-
coding of an item-in-context results in subsequent implicit
context effects on memory (c'est à dire., memory enhancement
by context similarity without testing for the explicit item–
context association)—are just beginning to be explored.
Evidence from rodents shows that selective hippo-
campal damage results in deficits in forming a memory
of the context (or location) where items were previously
encountered (Eichenbaum, 2004). Par exemple, rats with
hippocampal lesions fail to recognize a previously en-
countered object when contextual information relative
to encoding has been changed (Mumby, Gaskin, Glenn,
Schramek, & Lehmann, 2002). Other evidence for a role
of the hippocampus in implicit context effects on mem-
ory for cues comes from fear conditioning studies: Dans
addition to conditioned freezing to an auditory cue, rats
also exhibit freezing behavior when placed in the training
contexte, but hippocampal lesions eliminate such contex-
tual fear responses without affecting conditioned re-
sponses to the tone (Phillips & LeDoux, 1992). Recently,
using large-scale neuronal population recordings, new in-
sights into the nature of hippocampal context representa-
tions have emerged. Such studies show that hippocampal
Journal des neurosciences cognitives 30:4, pp. 579–593
est ce que je:10.1162/jocn_a_01218
ensemble context codes become associated with the
memories and behaviors that are appropriate for that con-
text. When confronted with a familiar context, the relevant
hippocampal context code is automatically reexpressed,
thereby priming the appropriate memories while reducing
interference from memories associated with other con-
texts (for a review, see Smith & Bulkin, 2014).
Analogous to this animal work, implicit context depen-
dency of memories has been demonstrated in human stud-
ies where context similarity between the original encoding
and retrieval context enhanced both recognition and re-
collection (Cox, Tijdens, Meeter, Sweegers, & Talamini,
2014; van Ast et al., 2013, 2014; Talamini & Gorree, 2012;
Talamini et al., 2010; Tsivilis, Otten, & Rugg, 2001). Le
consistent observation of highly context-dependent mem-
ories across this wide range of studies underscores context-
dependent memory as one of the hallmarks of human
episodic memory (Tulving, 1972). With respect to the neu-
ral correlates of such context-dependent memories, un
study using magnetoencephalography has shown that,
with an intact encoding-retrieval context, high theta power
during encoding predicted successful recognition, alors que
high theta power was detrimental when the retrieval con-
text was rearranged relative to encoding. En outre, cross-
frequency coupling analysis revealed a context-dependent
theta-to-gamma memory effect, which was assigned to the
left hippocampus using source localization (Staudigl &
Hanslmayr, 2013). An intracranial electroencephalography
(iEEG) study furthermore implicated the prefrontal cortex
(PFC) in successful item in context (temporal, in this case)
binding during memory encoding, resulting in implicit
context-dependent (c'est à dire., clustering) mémoire (Long &
Kahana, 2015). Cependant, given inherent uncertainty of
(deep) source localization of EEG and magnetoencepha-
lography signals and limited coverage of intracranial elec-
troencéphalographie, an extension of these findings using
techniques that allow for stronger spatial inferences, tel
as fMRI, is needed.
In contrast with such studies assessing (neural) encod-
ing mechanisms that can subsequently modulate implicit
context effects on recognition, previous fMRI studies have
almost exclusively focused on how subsequent explicit
memory of relations among cues is accomplished during
encoding (Davachi, 2006). “Context” in these studies re-
fers to scenes that were explicitly associated with objects:
The corresponding retrieval tests directly probed asso-
ciative memory among items and contexts. These studies
converge on the idea that encoding of items versus context
memories rely on distinct operations within the medial-
temporal lobe, in which “what” and “where” processing
streams function in parallel and converge within the hip-
pocampus (Eichenbaum, Sauvage, Fortin, Komorowski,
& Lipton, 2012; Ranganath, 2010b; Diana, Yonelinas, &
Ranganath, 2007; Davachi, 2006). In addition to the hippo-
campus, PFC has been shown to play an equally important
role in relational memory encoding when the integration
of contextual information with specific item features is re-
demandé (Murray & Ranganath, 2007; Summerfield et al.,
2006; Prince, 2005). En effet, lateral areas of PFC might be
involved in selecting task-relevant information and in
strategy implementation to find associations among items
during memory encoding (Ranganath, 2010un). When an
event is encoded in a particular context, PFC is therefore
likely to contribute to the process of integrating relevant
elements together. In agreement with this line of reason-
ing, some fMRI studies documented the involvement of
both the hippocampus and PFC in enhanced item (c'est à dire.,
word) memory in semantic versus nonsemantic contexts
(Wagner, 1998; Kapur et al., 1994) or maintaining a rep-
resentation of temporal context (Davachi & DuBrow, 2015;
Jenkins & Ranganath, 2010). Cependant, as these studies
employed either encoding instructions informing about a
subsequent explicit associative recognition test or used
very broad context manipulations, such observations do
not necessarily imply that the same brain regions are in-
volved when no instructions are given to memorize the
relevant items (c'est à dire., incidental encoding) or when sub-
sequent recognition is putatively modulated by implicit
context effects (c'est à dire., when memory enhancement by con-
text similarity is assessed on item recognition without test-
ing for the explicit item–context association). En effet, un
patient study suggests that the neural structures under-
lying explicit binding versus implicit contextual probing
may differ (Graf & Schacter, 1985; but see Chun & Phelps,
1999). Ainsi, previous fMRI studies did not unequivocally
address the question on which neural processes during
encoding contribute to subsequent implicit context effects
on memory.
Only two studies so far used an implicit context manip-
ulation during recognition (Hayes, Baena, Truong, &
Cabeza, 2010; Hayes, Nadel, & Ryan, 2007). In those
études, faces during encoding were presented against a
naturalistic scene (Hayes et al., 2007, 2010) or a mono-
colored white (Hayes et al., 2007) and black (Hayes
et coll., 2010) background. Both studies found a decrement
in recognition when a face encoded in a scene-context
was presented against a monocolored background during
reconnaissance, as compared with faces that were presented
against a monocolored background both during encod-
ing and recognition. This effect was associated with a
larger subsequent memory effect (SME), Par exemple,
in the hippocampus as well as enhanced connectivity be-
tween the hippocampus and visual association areas for
the face in scene-context (Hayes et al., 2010). Cependant,
the stronger involvement of the hippocampus and its as-
sociated connectivity with face-responsive areas can be
explained by the more complex and visually richer scene
processing during encoding, as simply perceiving visually
rich scenes has been associated with extensive hippo-
campal activation (Zeidman, Mullally, & Maguire, 2015).
Donc, it remains to be convincingly shown that
PFC and hippocampus are involved in memory contex-
tualization during encoding, resulting in subsequent im-
plicit context effects on memory.
580
Journal des neurosciences cognitives
Volume 30, Nombre 4
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
un
_
0
1
2
1
8
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
In the current study, we investigated the hypothesis
that information across neural circuits involved in item
and context representations would be integrated by
the hippocampus and prefrontal regions to subserve
memory contextualization, subsequently resulting in
context-dependent memories. To investigate this, nous
implemented a memory task using neutral face images
as items and scene images as contexts. During recogni-
tion, we asked our participants to indicate whether they
had seen a given face image during encoding or not.
Surtout, we manipulated context similarity during rec-
ognition relative to encoding by presenting faces against
either identical (intact) or different (rearranged) scènes.
By doing so, we ensured the presence of visually rich
background images at all time. En outre, any modu-
lation in memory performance by a shift in context can-
not be driven by a change in familiarity, as all items and
contexts have already been presented during encoding.
Because stronger context-dependent memory is likely
observed with “deeper” encoding strategies (De Beni
& Pazzaglia, 1995; Graf & Schacter, 1985; Richardson,
1980), we instructed our participants to actively form a
vivid mental image of the face in context and to indicate
on a trial-by-trial basis how well they did in forming this
mental face-in-context image. We expected that this
“subjective memory contextualization index” during en-
coding would predict subsequent context-dependent
face memory. Surtout, we did not inform our par-
ticipants of the subsequent memory test, as to mimic
real-life memory formation, and to prevent deliberate
encoding of the explicit association between the face
and the context. Behaviorally, we expected to observe
enhanced face recognition in intact contexts versus re-
arranged contexts (Meyer, Krans, van Ast, & Smeets,
2017; Cox et al., 2014; van Ast et al., 2013, 2014; Talamini
et coll., 2010). At the neural level, regardless of context,
we expected to find SMEs for the faces in the face-
responsive region of the fusiform gyrus (FG; c'est à dire., fusi-
form face area [FFA]) and/or the amygdala, in line with
previous research (Kanwisher & Yovel, 2006; Kanwisher,
McDermott, & Chun, 1997). In response to the presented
contexts, we expected activity in the place-responsive
region in the parahippocampal gyrus (PHG; c'est à dire., para-
hippocampal place area [APP]), as this region has been
linked to the representation of contextual features
(Epstein, Harris, Stanley, & Kanwisher, 1999). La plupart
importantly, we predicted that the integration of face
(FFA/amygdala) and context (APP) representations
during effective memory contextualization would be
associated with stronger neural activity in PFC and
hippocampus. Enfin, as actively integrating infor-
mation likely requires functional connections among
these brain regions, we also expected to find stronger
neural coupling between PFC/ hippocampus and dis-
tributed areas involved in representations of face
(FFA/amygdala) and context (APP) to support memory
contextualization.
MÉTHODES
Participants
Thirty-four right-handed university students (âge moyen =
23.65 années) with no history of neurological or psychiatric
disease gave written informed consent. Because of tech-
nical failure (par exemple., MR scanner malfunction), data of eight
participants were lost. Aussi, in line with previous memory
recherche (Rimmele, Davachi, & Phelps, 2012), partici-
pants were excluded from the analyses if their memory
performance did not exceed chance level, leading to ex-
clusion of an additional five participants (see Behavioral
Data Analysis). Par conséquent, current analyses are based
sur 21 participants (âge moyen = 24.24 années, SD = 2.86;
15 femmes). All study procedures were approved by the
local institutional review board (Commissie Mensgebonden
Onderzoek Regio Arnhem-Nijmegen).
Experimental Tasks and Procedure
A localizer task and an encoding part of the memory con-
textualization task (MCT-encoding) were administered
during fMRI scanning. Recognition (MCT-recognition)
was assessed outside the scanner. In total, 280 color im-
ages of faces from three databases (Langner et al., 2010;
Tottenham et al., 2009; Lundqvist, Flykt, & Ohman, 1998)
et 160 color images from various sources that depict
indoor scenes, city landscapes, or natural scenes were
selected for these two tasks. All tasks were administered
using Presentation software ( Version 16.4, www.neurobs.
com).
Localizer Task
A functional localizer task was used to identify brain activa-
tion elicited specifically by faces and contexts. Forty face
images (out of total 280), 40 context images (out of total
160), and an equal number of those images that were
phase scrambled were presented in the task. Scrambling
was accomplished within MATLAB (The MathWorks, Inc.,
Natick, MA) by randomizing the Fourier-transformed
phase of R, G, and B layers of each stimulus image, lequel
was then added to the existing RGB phase structures in the
original images. Thereby, the relative phase of the RGB
layers in the scrambled images was identical to that in
the original images, and the color composition was kept
the same as in the original images as well.
During the task, the four categories (face, contexte,
scrambled face, and scrambled context) of image stimuli
were presented with a blocked design. The order of the
16 blocks was mirrored to avoid covariation of task
effects with linear trends. Within each block, 20 images
from one specific category (par exemple., face) were continuously
presented in 20 trials of 1-sec duration, without intertrial
interval (ITI). In each trial, the face or scrambled face
stimuli were presented within an oval shape in the middle
of the screen, whereas the context or scrambled context
Zhang et al.
581
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
un
_
0
1
2
1
8
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
stimuli were presented full screen. All images used in the
localizer task were presented twice, resulting in a total of
320 trials. To keep participants engaged, a small red dot
was presented in half of the trials that were randomly
selected. Participants were instructed to indicate their
detection of the red dot using a button press.
Memory Contextualization Task
During the memory contextualization task (MCT), faces
served as to-be recognized items, whereas background
scenes served as context. A total number of 240 face
images and 120 context images were used in MCT. Le
face images were divided into two sets, each of which
was randomly selected as either the target stimuli (c'est à dire.,
used in both the encoding [MCT-encoding] and recogni-
tion [MCT-recognition] phases) or as the lures (c'est à dire., used
only in MCT-recognition phase). Le 120 context images
used in the MCT-encoding were used again in the MCT-
reconnaissance.
MCT-encoding. To pair the face and context images
and to assign the paired face–context combinations to
different (retrieval) conditions that were later presented
in the MCT-recognition as either intact or rearranged, le
randomly selected 120 face images for encoding were di-
vided into two subsets with 60 gender-matched face im-
ages in each subset. De la même manière, all context stimuli were
also divided into two subsets that were matched on loca-
tion (indoors vs. outdoors) of contexts. For each partici-
pant, the face and the context stimuli subsets were
randomly assigned to the intact or rearranged retrieval
condition of MCT-recognition and those faces and con-
texts were then randomly paired (c'est à dire., into face–context
combinations), resulting in randomized pairing of face–
context combinations within each condition. Plus loin-
plus, restrictions were made so that no trials from one
condition (c'est à dire., intact) were presented more than twice
consecutively for each participant (see MCT-recognition).
All stimuli were presented in 120 trials with a jittered ITI
(average duration = 2 sec).
It is worth mentioning that the terms “intact” and “re-
arranged” have previously been used to refer to explicit
knowledge of item–item associations in some studies
(par exemple., Litman & Davachi, 2008; Giovanello, Schnyer, &
Verfaellie, 2004; Jackson & Schacter, 2004). Ici, nous
follow other recent studies that used the same terms to
investigate implicit context effects (Meyer et al., 2017;
van Ast et al., 2013, 2014; Hayes et al., 2010; Tsivilis, Otten,
& Rugg, 2001).
The MCT-encoding task was introduced as a test of
imagination ability to induce deep incidental encoding
(van Ast et al., 2013) since deeper or more vivid encod-
ing strategies have been shown to strengthen context-
dependent memory (De Beni & Pazzaglia, 1995; Richardson,
1980). Spécifiquement, participants were instructed to imagine
a scene where the person (face) interacts with the place
(contexte) as vividly as possible in each trial. We reasoned
that these instructions would aid deep encoding and
thereby promote the formation of an association between
faces and their unique contexts (De Beni & Pazzaglia,
1995; Richardson, 1980).
A 6-min long practice session, using the same face
and context stimuli as in the localizer task was carried
out before the MCT-encoding task (Figure 1A). This prac-
tice session allowed the participants to become familiar-
ized with the task and helped to attenuate primacy
effects. To further balance remaining primacy effects
across participants, 10 trials from each retrieval condition
(c'est à dire., to-be intact or to-be rearranged) were randomly
selected and presented in an intermixed way in the first
20 trials and the same items were tested at the begin-
ning of the MCT-recognition task (see below). Note that
ces 20 trials were not removed from analyses. Nous
aimed to minimize primacy effects because the current
study focused on the encoding process; primacy effects
are mainly due to, Par exemple, novelty effects during
encoding. We did not make specific restrictions to con-
trol recency effects, but because trials were presented
in a random order, no systematic influence on recency
effects can be expected.
Within each trial, a 1-sec presentation of a context
stimulus was followed by a 3-sec overlaid presentation
of an oval-shaped face stimulus to allow the encoding
of context alone without the interference from centrally
presented face stimuli. Participants then reported how
well they could imagine the person being part of this
scene within 1 sec, using a four-item scale (c'est à dire., depuis
“not vivid at all” to “perfectly vivid”). This trial-by-trial
index was used as subjective contextualization ability.
Total duration of the MCT-encoding task was 14 min
(7 sec × 120 trials).
MCT-recognition. Face recognition took place outside
the scanner approximately 20 min, with a range of 18–
22 min, after participants had finished the encoding task.
Le 120 old faces from encoding were intermixed with
120 new faces (c'est à dire., “lures”). Surtout, to assess context-
dependent memory, half of the old faces were presented
against the same context stimuli as in the encoding task
(intact retrieval condition), whereas the other half were
reshuffled and presented in different face–context com-
binations (rearranged retrieval condition). New faces
were randomly combined with the 120 old contexts.
Ainsi, during recognition, each context was presented
twice in total. As explained above, the first 20 trials from
the encoding phase were intermixed with 20 trials con-
taining new faces (c'est à dire., not presented in the encoding task)
and presented at the beginning of recognition for each
participant. The number of trials for each retrieval condi-
tion out of those 40 trials was counterbalanced: 10 trials
from the intact condition, 10 trials from the rearranged
condition (thus in total 20 trials with old faces), ainsi que
582
Journal des neurosciences cognitives
Volume 30, Nombre 4
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
un
_
0
1
2
1
8
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
20 trials from the “lure” condition. During the entire task,
the trial sequence was pseudorandomized individually,
whereby neither old nor new faces, nor the trials from
the same condition were presented on more than two trials
consecutively.
For each trial, the face stimulus was presented overlaid
onto the context. A 6-point confidence rating scale was
presented at the bottom of the screen (Figure 1B), avec
which participants indicated if the face was old or new
and to what extent they were confident about their judg-
ment (1 = absolutely sure it was a new face; 2 = some-
what sure it was a new face; 3 = guessing it was a new
face; 4 = guessing it was an old face; 5 = somewhat
sure it was an old face; 6 = absolutely sure it was an
old face). Trials were self-paced, and a fixed 2-sec ITI was
used. En moyenne, the MCT-recognition task took 22 min
(5.5 sec × 240 trials).
Data Acquisition and Analysis
fMRI Data Acquisition
All images were acquired using a 3-T Siemens Magnetom
Skyra (Erlangen, Allemagne) MRI scanner with a 32-channel
head coil at the Donders Institute for Brain, Cognition and
Behaviour in Nijmegen, the Netherlands. High-resolution
structural images (1 × 1 × 1 mm3) were acquired using a
T1-weighted magnetization-prepared rapid gradient echo
séquence (repetition time [TR] = 2300 msec, echo time
[TE] = 3.03 msec, flip angle = 8°, field of view [FOV] = 256 ×
256 × 192 mm3). During both localizer and encoding
tasks, T2*-weighted dual-echo EPI BOLD-fMRI images
were acquired using an interleaved ascending slice ac-
quisition sequence (slices = 40, TR = 2570 msec, TEs =
15/35.7 msec, flip angle = 90°, voxel size = 2 × 2 × 2 mm3,
slice gap = 0.34 mm, FOV = 212 × 212 mm2). Gradient-
echo field-map data were also acquired for EPI off-resonance
distortion correction (slices = 64, TR = 1020 msec, TEs =
10/12.46 msec, flip angle = 90°, FOV = 224 × 224 mm2,
slice thickness = 2 mm).
Behavioral Data Analysis
Participants were tested in a within-subject factorial design
with subsequent memory (later remembered vs. later for-
gotten) and retrieval context (intact vs. rearranged) as main
experimental factors.
To ensure memory performance was above chance
level, we conducted binomial tests to investigate the sta-
tistical significance of the observed deviations (number
of remembered vs. forgotten trials) from the null distri-
bution (c'est à dire., random performance). We defined above-
chance level performance as a number of correct trials
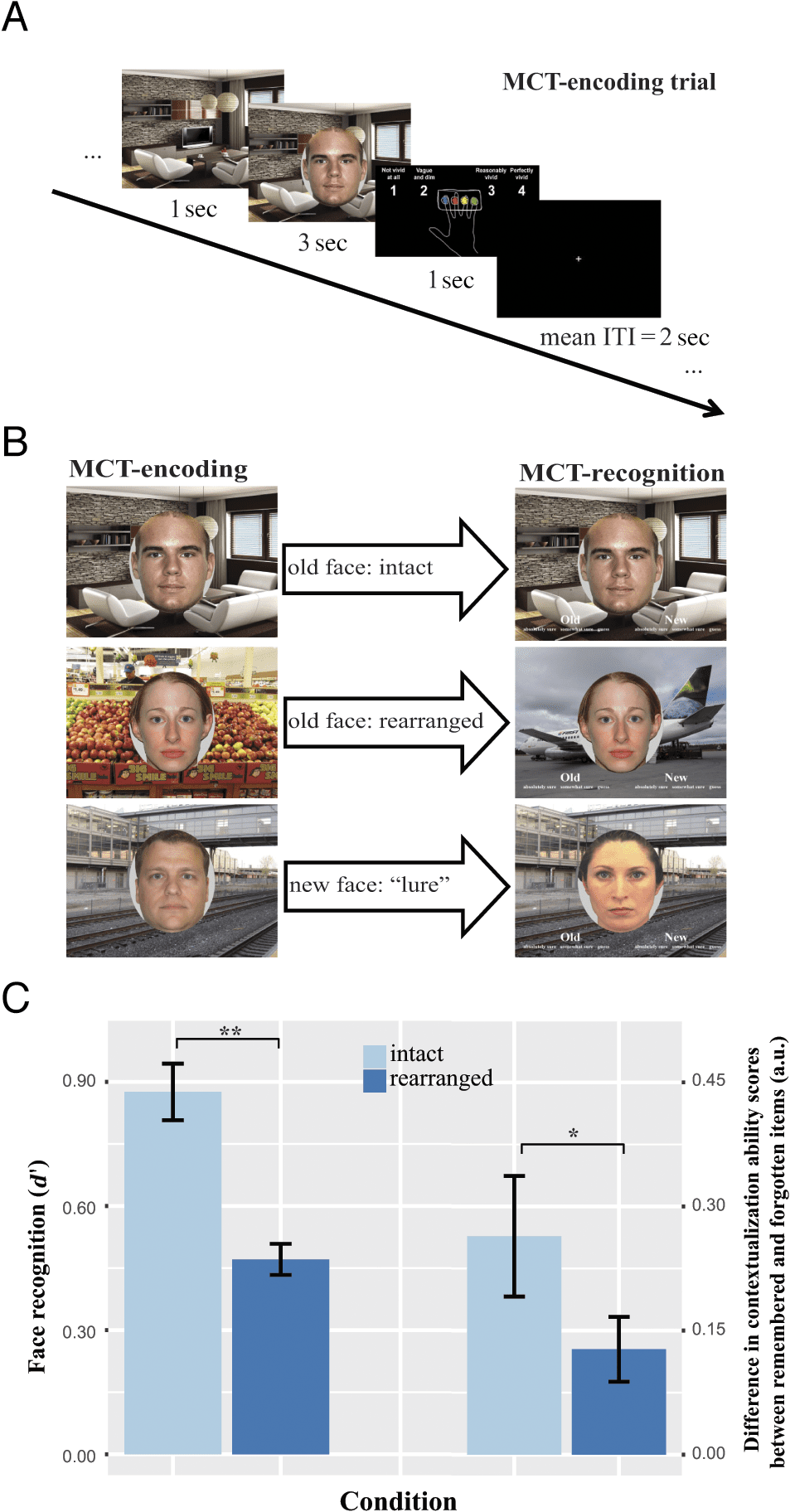
Chiffre 1. Experimental paradigm of MCT and the validation of the
experiment. (UN) MCT-encoding trial: For each trial of the encoding task,
the context stimulus was presented for 1 sec, followed by the combined
presentation of the face and context stimuli for 3 sec. Participants
were instructed to imagine the person interacting with the context.
Participants then reported how well they could imagine this interactive
scene on a 4-point scale (1 = not vivid at all, 4 = perfectly vivid )
presented for 1 sec. Trials were separated by a jittered ITI with an
average of 2 sec. (B) MCT-encoding and recognition: During recognition,
previously presented face stimuli from the encoding task (gauche) étaient
presented against either their original encoding context stimuli (“intact”)
or against different ones (“rearranged”). New face stimuli (not seen
during encoding) were presented in “lure” trials against context stimuli
(seen during encoding and recognition; bottom right). The presentation
of face and context stimuli in the recognition task was self-paced.
Participants were instructed to indicate if the presented face was old
or new and to what extent they were confident about their judgment
0 was found
based on a 6-point scale. (C) A significant difference in d
between intact and rearranged trials, which validated the experimental
manipulation of context dependency of face memory (gauche); higher
subjective contextualization ability scores were found for remembered
versus forgotten items in intact versus rearranged conditions (droite).
Error bars represent standard errors of the mean. **p < .001, two-tailed;
*p < .05, one-tailed.
Zhang et al.
583
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
a
_
0
1
2
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
that has a chance of p < .05 of arising from this null dis-
tribution. This criterion led to a threshold of 134 correct
trials ( p = .041) out of 240 faces presented in the rec-
ognition task. Based on this procedure, data from five
participants were excluded from all analyses.
To assess memory recognition, hit rates (i.e., propor-
tion of correct responses to “old” faces) and false alarm
rates (i.e., proportion of incorrect responses to “new”
0 (the
faces) were calculated and then converted to d
sensitivity index used in signal detection theory that
takes into account response bias (Emmerich, 1967) as a
function of retrieval context and of subjective contex-
0
between intact and
tualization ability. The difference of d
0) quantified context
rearranged conditions (i.e., delta d
0 indicating
dependency of memory, with a larger delta d
a stronger contextualization effect.
Subjective contextualization ability was derived from
the trial-by-trial vividness scores that participants reported
for their imagined scenarios involving the face-in-context
stimuli. These contextualization ability scores were ana-
lyzed using a repeated-measures ANOVA with Subsequent
memory performance (i.e., hits vs. misses) and Retrieval
conditions (i.e., intact vs. rearranged) as within-subject
factors. When the analysis returned a significant inter-
action effect, we used a (one-tailed) t test to test our pre-
diction that the difference between contextualization
ability scores for hits and misses would be larger in intact
than in rearranged trials. Furthermore, hit rates corrected
for false alarm rates (i.e., hit rates minus false alarm rates)
were modeled as a function of confidence level (i.e., low,
middle and high) and retrieval conditions (i.e., intact vs.
rearranged) to investigate the association between mem-
ory performance and confidence rating-based memory
strength (Kirwan, Wixted, & Squire, 2008; Slotnick &
Dodson, 2005). A repeated-measures ANOVA was used
for these models.
fMRI Data Analysis
Data preprocessing and statistical analyses were carried
out using Statistical Parametric Mapping Software (SPM8,
Wellcome Trust Centre for Neuroimaging). Before pre-
processing, dual-echo images were corrected for geomet-
ric distortions caused by magnetic field inhomogeneity
(Hutton et al., 2002) separately for each echo using field
map images. The corrected single-echo images were then
recombined using the parallel-acquired inhomogeneity-
desensitized method (Poser, Versluis, Hoogduin, & Norris,
2006). Preprocessing and further analyses were carried
out on the combined images. The first five recombined
EPI volumes were discarded to allow for T1 equilibration.
Preprocessing of the fMRI data included coregistration of
functional and structural images using mutual informa-
tion maximization, spatial normalization with the Montreal
Neurological Institute (MNI) template using nonlinear
warping, and spatial smoothing using an 8-mm FWHM
Gaussian kernel.
Functional localizer. To localize the brain regions re-
sponsive to faces and contexts respectively, brain activa-
tion in response to faces in contrast to scrambled faces,
and to contexts in contrast to scrambled contexts, were
estimated using a general linear model with 24 additional
motion parameters as nuisance regressors (six realign-
ment parameters, six squared realignment parameters,
six first derivatives of realignment parameters, and six
squared first derivatives of realignment parameters). Voxel-
level whole-brain family-wise error (FWE) corrections were
used as multiple comparison correction. Because the FG
(including FFA) and the amygdala have been implicated
in facial feature processing (Mende-siedlecki, Said, &
Todorov, 2013; Todorov, 2012) whereas the PHG (in-
cluding PPA) has been associated with context represen-
tations, we a priori hypothesized that these regions
would be involved in face and context processing, re-
spectively. In line with these hypotheses, we then created
three spherical ROIs with an 8-mm radius centered at the
peak voxels of aforementioned regions.
Memory contextualization. To investigate brain mech-
anisms underlying memory contextualization, all trials
during encoding were sorted based on whether faces
were later remembered or forgotten (i.e., SMEs; Paller
& Wagner, 2002; Kirchhoff, Wagner, Maril, & Stern, 2000;
Brewer, Zhao, Desmond, Glover, & Gabrieli, 1998; Wagner,
1998; Gabrieli, Brewer, Desmond, & Glover, 1997; Nyberg,
Cabeza, & Tulving, 1996). To ensure that neural correlates
truly related to successful memory formation, we checked
whether the subjective confidence levels as assessed dur-
ing memory recognition related to successful subsequent
memory. Hit rates differed from false alarm rates for
high, t(18) = 5.45, p < .001, middle, t(19) = 10.23, p <
.001, and low confidence levels, t(19) = 2.15, p = .045.
However, a closer examination revealed that, at the
lowest confidence level, participants on average recog-
nized old faces correctly in 21 out of a total 38 of trials
(hit rate = 55%), whereas they misjudged new faces as
“old” (i.e., false alarm) in 19 trials out of a total of 40 (false
alarm rate = 48%). Based on the observed false alarm rate
(19/40), one can estimate the number of correct guesses as
(38 × 19/40 =) 18 trials out of a total of 38 low-confidence
trials. Therefore, only (21 − 18 =) three additional correct
recognitions on average can be attributed to a perfor-
mance benefit due to memory. We therefore excluded all
low-confidence correct trials from the SME analyses. The
“remembered” trials for the SME analyses therefore only
included trials where participants recognized faces with
middle to high confidence level. Consequently, the average
trial number for each regressor (i.e., hits/misses in intact/
rearranged conditions) in the fMRI statistical model ranged
from 23 to 37 across participants.
For statistical analysis of the encoding task, event-related
trial responses were modeled with 4-sec box-car functions
in a first-level general linear model analysis and separate re-
gressors were created for “remembered” and “forgotten”
584
Journal of Cognitive Neuroscience
Volume 30, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
a
_
0
1
2
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
trials in intact versus rearranged conditions. In addition,
subjective contextualization ability scores were added in
the model as linear parametric modulators to remembered
and forgotten trials, respectively, which we expected to
predict the subsequent memory of faces. Mean time series
of white matter and CSF as well as 24 motion parameters
were included as nuisance regressors. The main effects of
subsequent memory and their interactions with retrieval
context (i.e., memory contextualization where SMEs differ
between conditions) were tested at the group level. A
priori ROIs, including the amygdala and face- and context-
responsive regions, were used for small volume correction
(SVC). In addition to those functional ROI masks derived
from the localizer task, we used the standardized bilateral
hippocampus parcellation from the Automated Anatomical
Labeling template (Tzourio-Mazoyer et al., 2002).
Finally, a psychophysiological interaction (PPI) analysis
(Gitelman, Penny, Ashburner, & Friston, 2003) was con-
ducted to investigate functional connectivity associated
with memory contextualization. The brain regions that
were not only activated in response to the overall SME con-
trast (remembered vs. forgotten trials) but also associated
with the subjective contextualization ability score were
taken as the seed region for the analysis. We first extracted
the BOLD time courses of the seed region and calculated
the first eigenvariate. We then deconvolved this time
course using the canonical hemodynamic response func-
tion to obtain the estimated time course of neural activity,
which was used as physiological component. The inter-
action of the subsequent memory (remembered vs. for-
gotten) and retrieval context (intact vs. rearranged) was
used to define the psychological component. We then
created the PPI term by multiplying the psychological
and physiological components. This interaction time
course, which is used to test for stronger connectivity asso-
ciated with subsequent memory in intact versus rearranged
trials, was reconvolved with the canonical hemodynamic
response function and then included in new first-level
models alongside the first eigenvariate of the seed region
time course. We then calculated the parameter estimate
maps for the PPI regressor for each participant and used
a one-sample t test at second level to test this interaction
at the group level. We further used the effective connectiv-
ity coefficients of this PPI analysis to behaviorally predict
context dependency of memories that was indicated by
between intact and rearranged conditions
the delta d
across all participants via an ANCOVA. We further checked
0 based on the calculation of
the distribution of delta d
Mahalanobis distance for potential outliers and then used
nonparametric permutation tests (Nichols & Holmes,
2001) for verification of results if any data point deviated
from the mean more than two standard deviations.
0
All statistical analyses of fMRI data have used voxel-level
whole-brain FWE corrections or SVCs for a priori ROIs
with p < .05.
RESULTS
Memory Performance
As expected, we found stronger context dependency of
0 in intact versus
memory for faces, indicated by higher d
rearranged trials, F(1, 20) = 64.59, ηp
2 = .76, p < .001
(see Figure 1C; also see hits/misses per condition, per
confidence level in Table 1). Confidence ratings during
recognition, which can be seen as a measure of memory
strength (Kirwan et al., 2008; Slotnick & Dodson, 2005),
were associated with memory performance, F(2, 26) =
156.11, η p
2 = .92, p < .001, as indicated by a main effect
of Confidence level on memory performance, with better
memory performance (i.e., higher hit rate minus false alarm
rate) at higher confidence levels: high level > middle level,
t(14) = 11.7, p < .001; middle level > low level, t(16) =
7.99, p < .001. This effect was also modulated by context,
F(1, 13) = 7.84, ηp
2 = .38, p < .005, as indicated by an
interaction effect of Confidence level and Retrieval condition,
with all three levels of confidence ratings associated with
better memory performance in intact versus rearranged
Table 1. Descriptives [Mean Number of Trials and Accuracy (SEM )] of Memory Performance
Stimulus
Retrieval Context
Memory Performance
Hit
Confidence level
Intact
Miss
Target
Rearranged
Lure
n.a.
HR-FAR
Hit
Miss
HR-FAR
CR
FA
Overall
High
Middle
Low
0
d
39.57 (1.41) 20.86 (1.43)
0.16 (0.03)
30.05 (1.08) 29.52 (1.04)
0.03 (0.02)
80.86 (2.18) 39.14 (2.18)
16.67 (2.14)
5.71 (1.36)
0.54 (0.03)
8.19 (1.28)
7.57 (1.69)
0.33 (0.05)
24.71 (4.63)
6.95 (1.33)
13.14 (1.43)
8.24 (0.79)
0.10 (0.03)
11.57 (1.13) 11.76 (1.14)
0 (0.03)
35.62 (3.10) 13.38 (1.52)
9.76 (1.28)
6.90 (1.08) −0.17 (0.04) 10.29 (1.08) 10.19 (1.28) −0.24 (0.03) 20.52 (2.29) 18.81 (1.83)
0.88 (0.07)
n.a.
0.47 (0.04)
n.a.
n.a.
Note that false alarm rates are assessed in lure trials and can therefore not be calculated separately for intact versus rearranged trials. CR = correct rejection;
FA = false alarm; HR-FAR = hit rate minus false alarm rate; n.a. = not applicable.
Zhang et al.
585
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
a
_
0
1
2
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 2. Descriptives [Mean Scores (SEM )] of Subjective Contextualization Ability
Retrieval Context
Memory Performance
Overall
High confidence level
Middle confidence level
Low confidence level
Intact
Rearranged
Hit
2.90 (.075)
3.09 (.11)
2.65 (.17)
2.41 (.20)
Miss
2.70 (.087)
2.13 (.33)
2.54 (.17)
2.42 (.20)
Hit
2.88 (.080)
2.79 (.18)
2.65 (.17)
2.67 (.18)
Miss
2.75 (.087)
2.31 (.25)
2.47 (.18)
2.67 (.18)
Note that subjective contextualization ability score was not tested statistically as a function of confidence level.
trials: intacthigh > rearrangedhigh, t(15) = 6.14, p < .001;
intactmid > rearrangedmid, t(17) = 3.84, p < .005; intactlow >
rearrangedlow, t(16) = 2.29, p < .05 (also see Table 1).
Higher subjective contextualization ability scores dur-
ing encoding were associated with trials that were later re-
membered versus forgotten, F(1, 20) = 19.55, ηp
2 = .49,
p < .001 (mHit = 2.84, SEM = 0.44; mMiss = 2.64, SEM =
0.46). In agreement with our expectations, this difference
in subjective contextualization ability score for remem-
bered versus forgotten items was enhanced by context
similarity, t(20) = 1.79, p = .045, one-tailed (see Figure 1C
and Table 2), suggesting a predictive effect of contextuali-
zation ability on the degree to which context aids retrieval.
These behavioral results demonstrate that the MCT re-
sulted in context-dependent memories, allowing to then
investigate the neural mechanisms of these effects.
Definition of ROIs
Brain activity during the localizer task was investigated
using the [face > scrambled face] et [context > scram-
bled context] contrasts. Among other regions, the FFA,
the amygdala, and the inferior frontal gyrus (IFG) étaient
identified for the contrast [Face > Scrambled Face]. UN
cluster of suprathreshold voxels lying at the junction of
the FG and PHG (FG–PHG junction) for the contrast
[Context > Scrambled Context] was detected as well
(see Table 3 for full results). Spécifiquement, as one of the
most active clusters in response to contexts resided in
the FG and extended to the adjacent regions (c'est à dire., PHG),
the peak voxel in the FG–PHG junction was defined
as the voxel with strongest activity within the PHG that
was closest to the boundaries of two regions based on
Automated Anatomical Labeling atlas. For later analyses
on these ROIs, spherical functional ROI masks for FFA,
amygdala, and FG–PHG junction were created on the basis
of these results, in addition to the anatomical template of
the hippocampus (see Memory Contextualization section).
Memory Contextualization
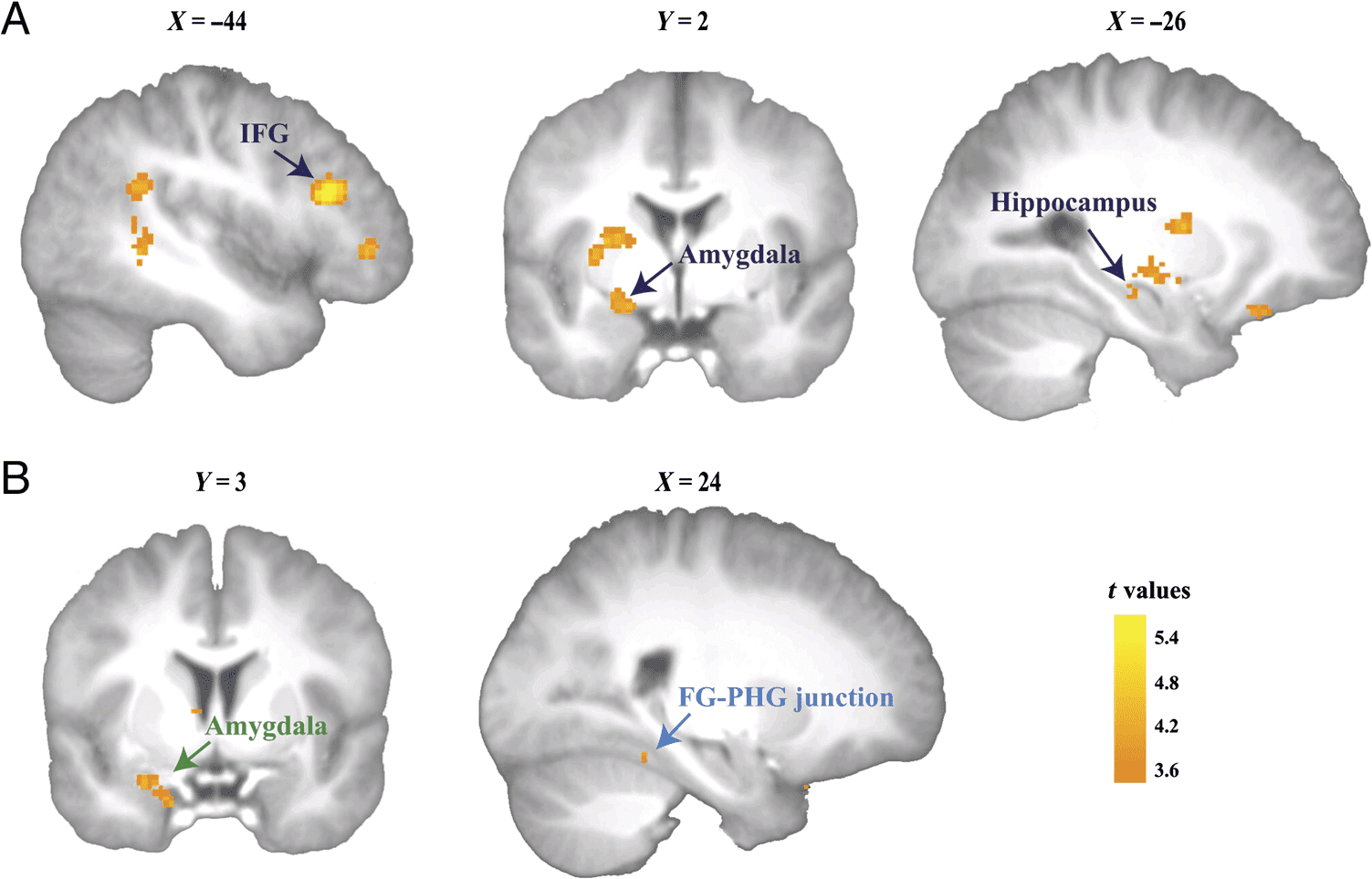
Brain regions associated with SMEs independent of con-
text (c'est à dire., all remembered > all forgotten) were identified
in the left IFG ( p < .05, whole-brain FWE) and left amyg- dala ( p < .001, SVC), but not in FFA and the FG–PHG junction (not even at a more liberal threshold of p < .005, uncorrected). A cluster of suprathreshold voxels in the left hippocampus showed marginally significant activation ( p = .058, SVC; see Figure 2A and Table 3). We then checked which neural mechanisms at encoding might have mediated the subsequent contextualization memory benefit. We defined memory contextualization at the neural level as SMEs as a function of retrieval context (i.e., interaction between subsequent memory and re- trieval context). No significant voxels emerged for this contrast. However, higher trial-by-trial subjective contex- tualization ability score, which was tested orthogonally to the memory effect (i.e., remembered vs. forgotten), was associated with enhanced activity in a cluster of supra- threshold voxels in the same left IFG region as revealed in the main SME ( p < .05, FWE; see Table 3). These find- ings reveal involvement of left IFG both in general memory performance as well as in subjective contextualization ability. These results raised the question whether IFG ac- tivity would still predict subsequent memory when control- ling for subjective contextualization ability. We tested this effect in a separate model and observed the same IFG clus- ter (peak voxel: −46, 26, 20) showed stronger activity in remembered versus forgotten trials. Given our expectation that PFC plays a role in informa- tion integration as well as the association between the left IFG and both objective subsequent memory and sub- jective contextualization ability, we performed a left IFG- based PPI analysis to investigate if the left-IFG based functional connectivity could potentially explain context dependency of memories. Enhanced connectivity be- tween the left IFG and left amygdala indeed predicted stronger subsequent memory for trials in the intact versus the rearranged condition ( p < .05, SVC; see Figure 2B). Connectivity with the right FG–PHG junction showed a trend in the same direction ( p = .054, SVC; see Figure 2B). We then investigated whether the effective connectivity between these regions would also predict interindividual differences in context-dependent face memory performance. The two PPI-derived neural cou- pling estimates were added as covariates in separate 586 Journal of Cognitive Neuroscience Volume 30, Number 4 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u / j / o c n a r t i c e - p d l f / / / / 3 0 4 5 7 9 1 7 8 7 2 1 0 / j o c n _ a _ 0 1 2 1 8 p d . f b y g u e s t t o n 0 8 S e p e m b e r 2 0 2 3 Table 3. Peak Voxel Coordinates in MNI Space and t Statistics Contrast Region Localizer Face > scrambled face
Amygdala
Fusiform gyrus
Inferior occipital cortex
IFG (opercular)
IFG (orbital)
IFG (triangularis)
Mid occipital cortex
Mid temporal lobe
Superior temporal lobe
Supramarginal gyrus
Context > scrambled context
Calcarine sulcus
Cerebellum
Fusiform gyrus
Inferior occipital cortex
Lingual gyrus
Mid occipital cortex
FG–PHG junction
Encoding
SMEs (remembered > forgotten)
Left IFG
Left amygdala
Left hippocampus
Left Hemisphere
Right Hemisphere
X
oui
z
Peak (t)
X
oui
z
Peak (t)
22
44
46
40
36
44
32
46
54
24
30
44
16
36
26
−2
−44
−80
12
28
24
−86
−52
−42
−58
−42
−82
−52
−82
−38
−18
−22
−10
26
−18
28
6
0
16
20
−8
−8
4
18
−10
5.29*
11.73
15.87
5.54
6.27
6.31
5.55
6.19
5.93
7.98
13.8
7.18
6.77
11.27
7.3un
−26
−36
−44
−40
−34
−50
−2
−48
−80
20
−86
−64
−64
−50
−18
−30
−26
−42
−16
−34
−30
−44
−20
−26
−62
−40
−44
−76
−50
−88
−42
24
−4
−12
−16
−20
−8
24
−8
2
28
12
−24
−10
−10
4
12
−8
20
−14
−10
4.02*
11.23
10.24
5.52
6.26
5.75
5.48
8.49
6.23
11.68
6.56
5.47
12.04
11.01un
5.73
4.2**
3.75*
Subjective contextualization ability (main effect)
Left IFG (triangularis)
−46
24
18
3.75
Memory contextualization (PPI, SME in intact > SME in rearranged)
(left IFG-) Left amygdala
(left IFG-) FG/PHG junction
−26
24
4
−42
−22
−16
4.85**
3.99*
All statistical values reported here were significant at p < .05, whole-brain FWE-corrected, unless indicated otherwise.
aIdentified based on voxel intensity and distance to the cluster peak (see Results: Definition of ROIs).
*p < .05, small volume corrected.
**p < .01, small volume corrected.
Zhang et al.
587
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
a
_
0
1
2
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Brain activity
associated with SMEs and
contextualization. (A) SMEs
were found in the left IFG (left)
and left amygdala (middle); a
small cluster of suprathreshold
voxels in the left hippocampus
(right) also showed marginally
significant activity. (B) Stronger
neural coupling between the
left IFG (seed region) and
the left amygdala (left) was
associated with memory
contextualization (greater
SME in intact vs. rearranged
conditions); neural coupling
between the left IFG (seed
region) and the right FG–PHG
junction was marginally
significant for the same contrast
(right). The images are
thresholded at p < .001
uncorrected, for visualization
purposes. Peak voxels of clusters
assigned to the left IFG, the
amygdala, and the hippocampus
(see Table 3) fell within these
regions as defined by the
Automatic Anatomical Labeling
template. The left IFG cluster is
located in the triangular part.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
a
_
0
1
2
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
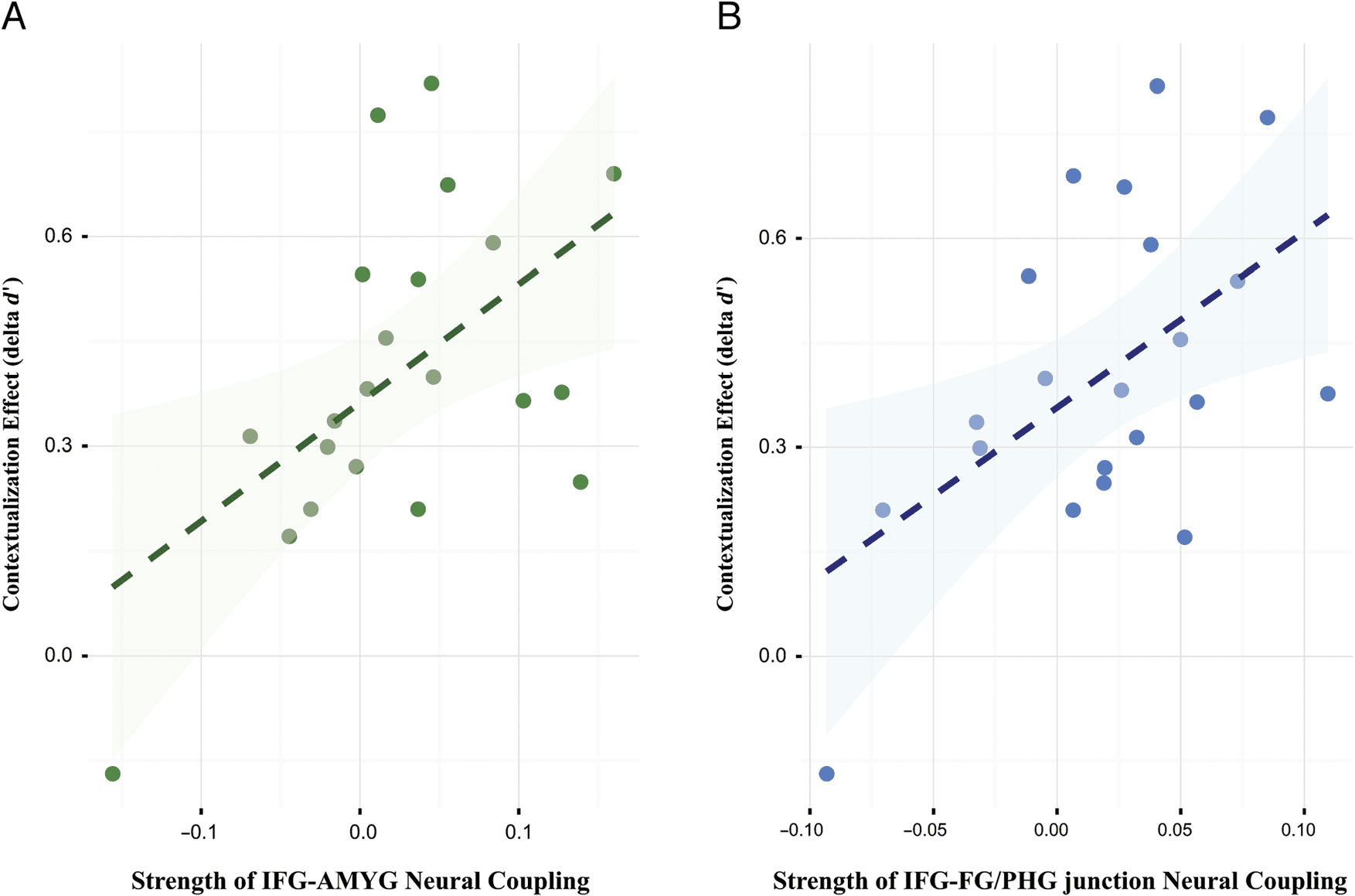
Figure 3. Neural coupling predictive of context-dependent memories. (A) The strength of neural coupling between the left IFG and amygdala predicted
interindividual differences in context dependency of memory at the behavioral level, as indicated by differences in d
retrieval conditions (delta d
in context dependency of memory.
0). (B) The strength of neural coupling between the left IFG and FG–PHG junction also predicted interindividual differences
0 between intact and rearranged
588
Journal of Cognitive Neuroscience
Volume 30, Number 4
ANCOVAs with Retrieval context as within-subjects factor
and Memory performance as dependent variable. We
found significant interactions between Retrieval context
and the left IFG-amygdala connectivity, F(1, 19) = 7.85,
ηp
2 = .29, p < .05 (see Figure 3A), and between Retrieval
context and the left IFG-FG/PHG junction connectivity,
F(1, 19) = 7.57, ηp
2 = .29, p < .05 (see Figure 3B). Non-
parametric correlation tests with 100,000 random permu-
tations further confirmed these findings (IFG-amygdala:
r(19) = 0.54, p = .012; IFG-FG/PHG junction: r(19) =
0.53, p = .013). Together, these additional tests confirm
that stronger left IFG-based coupling with the amygdala
and with the FG–PHG junction, all associated with memory
contextualization processes, predict stronger context-
dependent memory, both within and between participants.
DISCUSSION
The ability to store memories in conjunction with a rep-
resentation of their encoding context may protect against
subsequent dysfunctional memory generalization. There-
fore, the current study aimed to reveal the functional
neurobiology by which the brain contextualizes biologically
meaningful items during encoding, resulting in subsequent
context-dependent memories. Indeed, recognition per-
formance was enhanced when the retrieval context was
identical to the original encoding context, suggesting
effective memory contextualization processes during en-
coding. Context-dependent memories were associated
with stronger neural coupling during encoding between
the left IFG and the amygdala, as well as between the left
IFG and a region at the junction of FG and PHG. Impor-
tantly, the strength of these neural connections also pre-
dicted the extent of the context dependency of memories
when tested across participants, providing additional evi-
dence that contextualization processes during encoding
are mediated by these regions.
In the current study, we used an incidental memory
test to investigate how item recognition memory, in this
case for faces, is facilitated by contextual information.
This contrasts our study with previous work on relational
and associative memory, which has provided important
insight into neural mechanisms underlying item binding
but has commonly used explicit memory tests involving
recognition of pairs of items (Davachi, 2006). The advan-
tage of testing implicit context effects on item memory
during recognition, combined with incidental encoding
of items-in-contexts, is that it most closely mimics real-life
memory function: Context-dependent memory is con-
sidered a hallmark of human memory function (Tulving
& Thomson, 1973). Notably, contemporary animal re-
search shows that, when confronted with a familiar con-
text, hippocampal context codes are automatically
reexpressed, thereby priming the relevant memories
and reducing the interference from memories associated
with other contexts (for a review, see Smith & Bulkin,
2014): There seems to be no need to explicitly encode or
assess the association of the item with its context. Thus,
our paradigm builds on a long-standing tradition of both
animal and human behavioral work showing the power
of context to (implicitly) aid memory function.
The two studies by Hayes et al. (2007, 2010) did use a
similar experimental setup as ours. However, their find-
ings of the hippocampal and parahippocampal involve-
ment could be explained by processing of more complex
visual features due to unbalanced visual input in the con-
trast. To account for this, we adopted a task that was used
in previous behavioral studies into the context dependency
of memories. In this task, all items are encoded against
context backgrounds, but 50% of contexts are rearranged
during retrieval (van Ast et al., 2013, 2014). Unlike these
studies, which used words as items, here we used faces
because we specifically intended to investigate brain
mechanisms by which emotional or biologically salient
items are bound to contexts during encoding. An addi-
tional motivation was that faces are known to consistently
activate a face-responsive region within the FG (Kanwisher
et al., 1997) and the amygdala (Costafreda, Brammer,
David, & Fu, 2008; Sergerie, Chochol, & Armony, 2008),
a brain structure that is well known for its involvement in
processing emotion and salience (Sergerie et al., 2008;
Liberzon, Phan, Decker, & Taylor, 2003). Because another
area, located within the PHG (i.e., PPA), responds to the
presentation of spatial scenes (Epstein et al., 1999), our
design allowed us to distinguish item (i.e., face) and con-
text (i.e., scene) representations at the neural level.
Behaviorally, our results revealed that memory recog-
nition was strongly facilitated when the encoding and re-
trieval contexts were identical. This observation of
context dependency of memories aligns well with previ-
ous studies using highly similar experimental paradigms
(Meyer et al., 2017; Cox et al., 2014; van Ast et al.,
2013, 2014; Staudigl & Hanslmayr, 2013; Talamini et al.,
2010). New was that we assessed the subjective ability to
contextualize items on a trial-by-trial basis during encod-
ing. We observed that better trial-by-trial contextualiza-
tion ability yielded better subsequent item recognition
performance. More importantly, the degree to which the
original encoding context could help later retrieval was
positively associated with this contextualization ability. This
is in line with the notion that information presented in a
more vivid and detailed manner helps to enhance future
recall of that information (Buchsbaum, Lemire-Rodger,
Fang, & Abdi, 2012). Individual imagery ability, indicated
by measures of the vividness of visual imagery, has like-
wise been suggested to positively affect memory (Baddeley
& Andrade, 2000; McKelvie, 1984; McKelvie & Demers,
1979). The current findings are in line with these stud-
ies but also extend these by showing a predictive effect
of contextualization ability on context dependency of
memories. This finding suggests that stronger associative
imagery during encoding particularly benefits later recogni-
tion memory performance in the presence of contextual
cues.
Zhang et al.
589
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
a
_
0
1
2
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
With respect to the neural correlates of memory con-
textualization, we could not identify regions in which the
amplitude of regional BOLD responses was predictive of
context-dependent memory. It thus appears that contex-
tualization is not strongly predicted by the mere magni-
tude of neural responses in any specific brain region.
Rather, functional coupling between the left IFG and
the amygdala, as well as between left IFG and the FG–
PHG junction, was positively related to face recognition
as a function of retrieval context. In addition, the neural
coupling strength of these regions predicted individual
differences in the extent of context-dependent memory.
The encoding-related activity of the IFG, part of PFC, has
been consistently reported as being associated with SMEs
(Kim, 2011). More specifically, it seems to process both re-
lational and item-specific information as part of the ventro-
lateral PFC (Blumenfeld, Parks, Yonelinas, & Ranganath,
2011; Ranganath, 2010b; Murray & Ranganath, 2007), to
generate associations between items (Addis & McAndrews,
2006; Uncapher & Rugg, 2005), and to support the dura-
bility of episodic memories (Uncapher & Rugg, 2005).
Our findings broaden understanding of the mnemonic
functions of this region by providing compelling evidence
that the left IFG is a key region enabling effective memory
contextualization.
The observation that IFG connectivity rather than re-
gional activity was associated with contextualization
points toward a role for the IFG in actively integrating
information across distributed regions. These distant re-
gions included the amygdala, a core face-responsive re-
gion (Mende-siedlecki et al., 2013; Costafreda et al.,
2008) that processes information forwarded by more
face-selective regions such as the FFA (Todorov, 2012).
Here, we found left IFG-amygdala rather than IFG–FFA
connectivity, suggesting that memory contextualization
of biologically meaningful stimuli such as faces might re-
quire more salient feature processing that involves the
amygdala. Left IFG connectivity with a region at the junc-
tion of PHG and FG was also predictive of memory con-
textualization. We first identified this region as responsive
to the spatial contexts employed in our localizer task.
Previous studies have found that an adjacent region within
the PHG, the PPA, responds selectively to images of houses
or buildings (Aminoff, Kveraga, & Bar, 2013; Epstein &
Kanwisher, 1998). The difference between these findings
and ours may be explained by the fact that we chose scene
images with more complex features to increase distinc-
tiveness. Together, these findings indicate that strongly
contextualized memories result from an IFG-based coordi-
nation of mnemonic processes across distant regions re-
presenting distinct aspects of a memory, likely including
perceptual features, spatiotemporal context, and motiva-
tional salience.
The hippocampus is also known to play a critical role in
memory formation (Eichenbaum, 2000; Scoville & Milner,
2000; Gabrieli, 1998; Squire & Zola-Morgan, 1991) and has
been highlighted as the “binding” center where features
or elements of episodes and environments that are essen-
tial for recollection are bound together ( Yonelinas, 2013).
Here, we observed a marginally significant main effect of
subsequent memory (SME) in the hippocampus, which is
consistent with previous studies on its critical role in
memory formation ( Wagner, 1998). However, we did
not find hippocampal involvement associated with mem-
ory contextualization. Perhaps the current experimental
design was more likely to invoke an active and complex
process of information integration that strongly depends
on higher-order cognitive processes supported by pre-
frontal regions such as the IFG. The hippocampus, on
the other hand, may become more important during
memory consolidation, a process that persists well be-
yond the time of initial encoding (Knowlton & Fanselow,
1998; McClelland, McNaughton, & O’Reilly, 1995; Squire,
1992) that was not targeted by the current design. Alter-
natively, the absence of evidence for hippocampal in-
volvement might result from decreased power due to
interindividual variability in functional specialization
along the longitudinal axis of the hippocampus (Strange,
Witter, Lein, & Moser, 2014; Poppenk, Evensmoen,
Moscovitch, & Nadel, 2013). Future studies should use
tailored tasks focusing also on the post-encoding period
to elucidate hippocampal involvement in memory con-
textualization with higher anatomical precision.
Inappropriate memory contextualization is considered
a hallmark of traumatic memories (Brewin et al., 2010).
For instance, low-level memory representations that are
improperly contextualized are thought to contribute to
memory flashbacks in PTSD (Acheson et al., 2012; Liberzon
& Sripada, 2008; Ehlers & Clark, 2000). The brain structures
we identified as being involved in memory contextualiza-
tion are especially sensitive to stress-related neuromodula-
tory changes (Arnsten, 2009, 2015; Hermans, Henckens,
Joëls, & Fernández, 2014). Stress levels of noradrenergic
activation lead to occupation of lower-affinity alpha-1
adrenoceptors in PFC, thus impairing functioning of this
region (Birnbaum, Gobeske, Auerbach, Taylor, & Arnsten,
1999). Acute stress has an opposite effect on functioning
of the amygdala, where beta-1 adrenoceptors become en-
gaged at elevated levels of noradrenergic activity (Arnsten,
2000). Such dual effects are thought to be amplified by gluco-
corticoid activation (Roozendaal, McEwen, & Chattarji,
2009). In agreement, a previous study by van Ast et al.
(2013) using a similar task as the current study indeed
found impaired memory contextualization after a phar-
macological elevation of glucocorticoid levels. Notably,
reduced memory contextualization, as assessed by the
same task as used in this study, has been shown to predict
more traumatic memory intrusions and related distress
upon seeing a “trauma movie” (trauma analogue in the
lab; Meyer et al., 2017). These studies, together with the
present findings, suggest that extreme stress asso-
ciated with traumatic events could lead to unusually strong
amygdala-based representations, whereas a transient
suppression of PFC functioning may give rise to a lack of
590
Journal of Cognitive Neuroscience
Volume 30, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
a
_
0
1
2
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
mnemonic integration, resulting in decontextualization,
fragmentation, and generalization of memories. This inter-
pretation is in line with the revised dual representation
theory of PTSD, which states that traumatic experiences
can be stored as sensory-bound memories isolated from
original encoding contexts, which would allow for the re-
trieval of traumatic memories triggered by perceptual cues
reminiscent of the original trauma without retrieval of the
appropriate context (Bisby, Horner, Hørlyck, & Burgess,
2016; Brewin et al., 2010). Our data, however, indicate that
this model may place too much emphasis on hippocampal
instead of PFC dysfunction as central factor (Diamond,
Campbell, Park, Halonen, & Zoladz, 2007). It should be
noted that, in the current study, we only used neutral faces
as item stimuli to investigate memory contextualization.
Given the aversive and negatively arousing properties of
traumatic events, studies directly manipulating the emo-
tional valence and arousal of items are required to further
explore the clinical implications of our findings regarding
the neural substrates of memory contextualization. In addi-
tion, some studies have shown that emotional valence of
the encoding context can influence memory retrieval that
seems to recruit similar neural circuits activated during
encoding (Hofstetter, Achaibou, & Vuilleumier, 2012;
Sterpenich et al., 2006; Erk, Martin, & Walter, 2005; Erk
et al., 2003). It is therefore important to investigate how
emotional valence of contexts could contribute to in-
appropriate memory contextualization as well.
To conclude, our results indicate that memory contex-
tualization depends on the integration of information
across neural circuits that are involved in item and con-
text representations. Our findings in particular highlight a
key role for the left IFG in coordinating this mnemonic
process. Given the vulnerability of the prefrontal regions
to the effects of acute stress, our findings thereby provide
a novel framework for understanding the pathogenesis of
traumatic memories that are often seen in stress-related
disorders such as PTSD.
Acknowledgments
This research was supported by a grant from the Netherlands
Organization for Scientific Research (NWO) awarded to K. R.
(016 085 354). E. J. H. was supported by a grant from the
European Research Council (ERC-2015-CoG 682591), and V. v. A.
was supported by a VENI NWO grant (451-16-021). We are espe-
cially thankful to Milou Sep for her help in piloting the MCT.
Reprint requests should be sent to Wei Zhang, Centre for Cogni-
tive Neuroimaging, Donders Institute for Brain Cognition and
Behaviour, Radboud Universiteit, Kapittelweg 29, 6525 EN
Nijmegen, The Netherlands, or via e-mail: w.zhang@donders.ru.nl.
REFERENCES
Acheson, D. T., Gresack, J. E., & Risbrough, V. B. (2012).
Hippocampal dysfunction effects on context memory: Possible
etiology for posttraumatic stress disorder. Neuropharmacology,
62, 674–685.
Addis, D. R., & McAndrews, M. P. (2006). Prefrontal and
hippocampal contributions to the generation and binding
of semantic associations during successful encoding.
Neuroimage, 33, 1194–1206.
Aminoff, E. M., Kveraga, K., & Bar, M. (2013). The role of the
parahippocampal cortex in cognition. Trends in Cognitive
Sciences, 17, 379–390.
Arnsten, A. F. T. (2000). Through the looking glass: Differential
noradenergic modulation of prefrontal cortical function.
Neural Plasticity, 7, 133–146.
Arnsten, A. F. T. (2009). Stress signalling pathways that impair
prefrontal cortex structure and function. Nature Reviews
Neuroscience, 10, 410–422.
Arnsten, A. F. T. (2015). Stress weakens prefrontal networks:
Molecular insults to higher cognition. Nature Neuroscience,
18, 1376–1385.
Baddeley, A. D., & Andrade, J. (2000). Working memory and the
vividness of imagery. Journal of Experimental Psychology:
General, 129, 126–145.
Birnbaum, S., Gobeske, K. T., Auerbach, J., Taylor, J. R., &
Arnsten, A. F. T. (1999). A role for norepinephrine in stress-
induced cognitive deficits: Alpha-1-adrenoceptor mediation
in the prefrontal cortex. Biological Psychiatry, 46, 1266–1274.
Bisby, J. A., Horner, A. J., Hørlyck, L. D., & Burgess, N. (2016).
Opposing effects of negative emotion on amygdalar and
hippocampal memory for items and associations. Social
Cognitive and Affective Neuroscience, 11, 981–990.
Blumenfeld, R. S., Parks, C. M., Yonelinas, A. P., & Ranganath, C.
(2011). Putting the pieces together: The role of dorsolateral
prefrontal cortex in relational memory encoding. Journal of
Cognitive Neuroscience, 23, 257–265.
Brewer, J. B., Zhao, Z., Desmond, J. E., Glover, G. H., &
Gabrieli, J. D. (1998). Making memories: Brain activity that
predicts how well visual experience will be remembered.
Science, 281, 1185–1187.
Brewin, C. R., Gregory, J. D., Lipton, M., & Burgess, N. (2010).
Intrusive images in psychological disorders: Characteristics,
neural mechanisms, and treatment implications.
Psychological Review, 117, 210–232.
Buchsbaum, B. R., Lemire-Rodger, S., Fang, C., & Abdi, H. (2012).
The neural basis of vivid memory is patterned on perception.
Journal of Cognitive Neuroscience, 24, 1867–1883.
Chun, M. M., & Phelps, E. (1999). Memory deficits for implicit
contextual information in amnesic subjects with hippocampal
damage. Nature Neuroscience, 2, 844–847.
Costafreda, S. G., Brammer, M. J., David, A. S., & Fu, C. H. Y.
(2008). Predictors of amygdala activation during the processing
of emotional stimuli: A meta-analysis of 385 PET and fMRI
studies. Brain Research Reviews, 58, 57–70.
Cox, R., Tijdens, R. R., Meeter, M. M., Sweegers, C. C. G., &
Talamini, L. M. (2014). Time, not sleep, unbinds contexts
from item memory. PLoS ONE, 9, 1–6.
Davachi, L. (2006). Item, context and relational episodic encoding
in humans. Current Opinion in Neurobiology, 16, 693–700.
Davachi, L., & DuBrow, S. (2015). How the hippocampus
preserves order: The role of prediction and context. Trends
in Cognitive Sciences, 19, 92–99.
De Beni, R., & Pazzaglia, F. (1995). Memory for different kinds
of mental images: Role of contextual and autobiographic
variables. Neuropsychologia, 33, 1359–1371.
Diamond, D. M., Campbell, A. M., Park, C. R., Halonen, J., &
Zoladz, P. R. (2007). The temporal dynamics model of
emotional memory processing: A synthesis on the
neurobiological basis of stress-induced amnesia, flashbulb
and traumatic memories, and the Yerkes–Dodson law.
Neural Plasticity, 2007, 60803.
Diana, R. A., Yonelinas, A. P., & Ranganath, C. (2007). Imaging
recollection and familiarity in the medial temporal lobe: A
Zhang et al.
591
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
a
_
0
1
2
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
three-component model. Trends in Cognitive Sciences, 11,
379–386.
Ehlers, A., & Clark, D. M. (2000). A cognitive model of
posttraumatic stress disorder. Behaviour Research and
Therapy, 38, 319–345.
Eichenbaum, H. (2000). A cortical-hippocampal system for
declarative memory. Nature Reviews Neuroscience, 1, 41–50.
Eichenbaum, H. (2004). Hippocampus: Cognitive processes
and neural representations that underlie declarative memory.
Neuron, 44, 109–120.
Eichenbaum, H., Sauvage, M., Fortin, N., Komorowski, R., &
Lipton, P. (2012). Towards a functional organization of
episodic memory in the medial temporal lobe. Neuroscience
and Biobehavioral Reviews, 36, 1597–1608.
Emmerich, D. S. (1967). Signal detection theory and
psychophysics. David M. Green, John A. Swets. The
Quarterly Review of Biology, 42, 578.
Jackson, O., & Schacter, D. L. (2004). Encoding activity in
anterior medial temporal lobe supports subsequent
associative recognition. Neuroimage, 21, 456–462.
Jenkins, L. J., & Ranganath, C. (2010). Prefrontal and medial
temporal lobe activity at encoding predicts temporal context
memory. Journal of Neuroscience, 30, 15558–15565.
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). The fusiform
face area: A module in human extrastriate cortex specialized for
face perception. Journal of Neuroscience, 17, 4302–4311.
Kanwisher, N., & Yovel, G. (2006). The fusiform face area: A
cortical region specialized for the perception of faces.
Philosophical Transactions of the Royal Society of London,
Series B, Biological Sciences, 361, 2109–2128.
Kapur, S., Craik, F. I., Tulving, E., Wilson, A. A., Houle, S., &
Brown, G. M. (1994). Neuroanatomical correlates of encoding
in episodic memory: Levels of processing effect. Proceedings of
the National Academy of Sciences, U.S.A., 91, 2008–2011.
Epstein, R., Harris, A., Stanley, D., & Kanwisher, N. (1999).
Kim, H. (2011). Neural activity that predicts subsequent
The parahippocampal place area: Recognition, navigation,
or encoding? Neuron, 23, 115–125.
Epstein, R., & Kanwisher, N. (1998). A cortical representation
of the local visual environment. Nature, 392, 598–601.
Erk, S., Kiefer, M., Grothe, J., Wunderlich, A. P., Spitzer, M., &
Walter, H. (2003). Emotional context modulates subsequent
memory effect. Neuroimage, 18, 439–447.
Erk, S., Martin, S., & Walter, H. (2005). Emotional context
during encoding of neutral items modulates brain activation
not only during encoding but also during recognition.
Neuroimage, 26, 829–838.
Gabrieli, J. D. E. (1998). Cognitive neuroscience of human
memory. Annual Review of Psychology, 49, 87–115.
Gabrieli, J. D. E., Brewer, J. B., Desmond, J. E., & Glover, G. H.
(1997). Separate neural bases of two fundamental memory
processes in the human medial temporal lobe. Science, 276,
264–266.
Giovanello, K. S., Schnyer, D. M., & Verfaellie, M. (2004). A
critical role of the anterior hippocampus in relational
memory: Evidence from an fMRI study comparing associative
and item recognition. Hippocampus, 14, 5–8.
Gitelman, D. R., Penny, W. D., Ashburner, J., & Friston, K. J.
(2003). Modeling regional and psychophysiologic
interactions in fMRI: The importance of hemodynamic
deconvolution. Neuroimage, 19, 200–207.
Godden, D. R., & Baddeley, A. D. (1975). Context dependent
memory in two natural environments: On land and
underwater. British Journal of Psychology, 66, 325–331.
Graf, P., & Schacter, D. L. (1985). Implicit and explicit memory
for new associations in normal and amnesic subjects. Journal
of Experimental Psychology: Learning, Memory, and
Cognition, 11, 501–518.
Hayes, S. M., Baena, E., Truong, T.-K., & Cabeza, R. (2010).
Neural mechanisms of context effects on face recognition:
Automatic binding and context shift decrements. Journal of
Cognitive Neuroscience, 22, 2541–2554.
Hayes, S. M., Nadel, L., & Ryan, L. (2007). The effect of scene
context on episodic object recognition: Parahippocampal
cortex mediates memory encoding and retrieval success.
Hippocampus, 17, 873–889.
Hermans, E. J., Henckens, M. J. A. G., Joëls, M., & Fernández, G.
(2014). Dynamic adaptation of large-scale brain networks in
response to acute stressors. Trends in Neurosciences, 37,
304–314.
Hofstetter, C., Achaibou, A., & Vuilleumier, P. (2012). Reactivation
of visual cortex during memory retrieval: Content specificity
and emotional modulation. Neuroimage, 60, 1734–1745.
Hutton, C., Bork, A., Josephs, O., Deichmann, R., Ashburner, J.,
& Turner, R. (2002). Image distortion correction in fMRI:
A quantitative evaluation. Neuroimage, 16, 217–240.
memory and forgetting: A meta-analysis of 74 fMRI studies.
Neuroimage, 54, 2446–2461.
Kirchhoff, B. A., Wagner, A. D., Maril, A., & Stern, C. E. (2000).
Prefrontal-temporal circuitry for episodic encoding and
subsequent memory. Journal of Neuroscience, 20, 6173–6180.
Kirwan, C. B., Wixted, J. T., & Squire, L. R. (2008). Activity in the
medial temporal lobe predicts memory strength, whereas
activity in the prefrontal cortex predicts recollection. Journal
of Neuroscience, 28, 10541–10548.
Knowlton, B. J., & Fanselow, M. S. (1998). The hippocampus,
consolidation and on-line memory. Current Opinion in
Neurobiology, 8, 293–296.
Langner, O., Dotsch, R., Bijlstra, G., Wigboldus, D. H. J., Hawk,
S. T., & van Knippenberg, A. (2010). Presentation and
validation of the Radboud Faces Database. Cognition &
Emotion, 24, 1377–1388.
Liberzon, I., Phan, K. L., Decker, L. R., & Taylor, S. F. (2003).
Extended amygdala and emotional salience: A PET activation
study of positive and negative affect. Neuropsychopharmacology,
28, 726–733.
Liberzon, I., & Sripada, C. S. (2008). The functional neuroanatomy
of PTSD: A critical review. Progress in Brain Research, 167,
151–169.
Litman, L., & Davachi, L. (2008). Distributed learning enhances
relational memory consolidation. Learning & Memory, 15,
711–716.
Long, N. M., & Kahana, M. J. (2015). Successful memory
formation is driven by contextual encoding in the core memory
network. Neuroimage, 119, 332–337.
Lundqvist, D., Flykt, A., & Ohman, A. (1998). The
Karolinska directed emotional faces (KDEF). CD ROM
from Department of Clinical Neuroscience, Psychology
Section, Karolinska Institutet, 91–630. https://doi.org/10.1017/
S0048577299971664.
McClelland, J. L., McNaughton, B. L., & O’Reilly, R. C. (1995).
Why there are complementary learning systems in the
hippocampus and neocortex: Insights from the successes and
failures of connectionist models of learning and memory.
Psychological Review, 102, 419–457.
McKelvie, S. J. (1984). Reported visual imagery for faces and facial
recognition memory. Perceptual and Motor Skills, 59, 825–826.
McKelvie, S. J., & Demers, E. G. (1979). Individual differences in
reported visual imagery and memory performance. British
Journal of Psychology, 70, 51–57.
Mende-siedlecki, P., Said, C. P., & Todorov, A. (2013). The
social evaluation of faces: A meta-analysis of functional
neuroimaging studies. Social Cognitive and Affective
Neuroscience, 8, 285–299.
Meyer, T., Krans, J., van Ast, V., & Smeets, T. (2017). Visuospatial
context learning and configuration learning is associated with
592
Journal of Cognitive Neuroscience
Volume 30, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
a
_
0
1
2
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
analogue traumatic intrusions. Journal of Behavior Therapy
and Experimental Psychiatry, 54, 120–127.
Meyer, T., Smeets, T., Giesbrecht, T., Quaedflieg, C. W. E. M.,
Girardelli, M. M., Mackay, G. R. N., et al. (2012). Individual
differences in spatial configuration learning predict the
occurrence of intrusive memories. Cognitive, Affective, &
Behavioral Neuroscience, 13, 186–196.
Mumby, D. G., Gaskin, S., Glenn, M. J., Schramek, T. E., &
Lehmann, H. (2002). Hippocampal damage and exploratory
preferences in rats: Memory for objects, places, and
contexts. Learning & Memory, 9, 49–57.
Murray, L. J., & Ranganath, C. (2007). The dorsolateral
prefrontal cortex contributes to successful relational memory
encoding. Journal of Neuroscience, 27, 5515–5522.
Nichols, T. E., & Holmes, A. P. (2001). Nonparametric permutation
tests for {PET} functional neuroimaging experiments: A primer
with examples. Human Brain Mapping, 15, 1–25.
Nyberg, L., Cabeza, R., & Tulving, E. (1996). PET studies of
encoding and retrieval: The HERA model. Psychonomic
Bulletin & Review, 3, 135–148.
Paller, K. A., & Wagner, A. D. (2002). Observing the
transformation of experience into memory. Trends in
Cognitive Sciences, 6, 93–102.
Phillips, R. G., & LeDoux, J. E. (1992). Differential contribution
of amygdala and hippocampus to cued and contextual fear
conditioning. Behavioral Neuroscience, 106, 274–285.
Poppenk, J., Evensmoen, H. R., Moscovitch, M., & Nadel, L.
(2013). Long-axis specialization of the human hippocampus.
Trends in Cognitive Sciences, 17, 230–240.
Smith, S. M., & Vela, E. (2001). Environmental context-dependent
memory: A review and meta-analysis. Psychonomic Bulletin
& Review, 8, 203–220.
Squire, L. (1992). Memory and the hippocampus: A synthesis
from findings with rats, monkeys, and humans. Psychological
Review, 99, 195–231.
Squire, L., & Zola-Morgan, S. (1991). The medial temporal
lobe memory system. Science, 253, 1380–1386.
Staudigl, T., & Hanslmayr, S. (2013). Theta oscillations at
encoding mediate the context-dependent nature of human
episodic memory. Current Biology, 23, 1101–1106.
Sterpenich, V., D’Argembeau, A., Desseilles, M., Balteau, E.,
Albouy, G., Vandewalle, G., et al. (2006). The locus ceruleus
is involved in the successful retrieval of emotional memories
in humans. Journal of Neuroscience, 26, 7416–7423.
Strange, B., Witter, M. P., Lein, E. S., & Moser, E. I. (2014).
Functional organization of the hippocampal longitudinal axis.
Nature Reviews Neuroscience, 15, 655–669.
Summerfield, C., Greene, M., Wager, T., Egner, T., Hirsch, J., &
Mangels, J. (2006). Neocortical connectivity during episodic
memory formation. PLoS Biology, 4, 855–864.
Talamini, L. M., de Haan, L., Nieman, D. H., Linszen, D. H., &
Meeter, M. (2010). Reduced context effects on retrieval in
first-episode schizophrenia. PloS One, 5, e10356.
Talamini, L. M., & Gorree, E. (2012). Aging memories:
Differential decay of episodic memory components.
Learning & Memory, 19, 239–246.
Todorov, A. (2012). The role of the amygdala in face perception
and evaluation. Motivation and Emotion, 36, 16–26.
Poser, B. A., Versluis, M. J., Hoogduin, J. M., & Norris, D. G.
Tottenham, N., Tanaka, J. W., Leon, A. C., McCarry, T., Nurse,
(2006). BOLD contrast sensitivity enhancement and
artifact reduction with multiecho EPI: Parallel-acquired
inhomogeneity-desensitized fMRI. Magnetic Resonance in
Medicine, 55, 1227–1235.
Prince, S. E. (2005). Neural correlates of relational memory:
Successful encoding and retrieval of semantic and perceptual
associations. Journal of Neuroscience, 25, 1203–1210.
Quaedflieg, C. W. E. M., van de Ven, V., Meyer, T., Siep, N.,
Merckelbach, H., & Smeets, T. (2015). Temporal dynamics of
stress-induced alternations of intrinsic amygdala connectivity
and neuroendocrine levels. PLoS One, 10, e0124141.
Ranganath, C. (2010a). A unified framework for the functional
organization of the medial temporal lobes and the phenomenology
of episodic memory. Hippocampus, 20, 1263–1290.
Ranganath, C. (2010b). Binding items and contexts: The
cognitive neuroscience of episodic memory. Current
Directions in Psychological Science, 19, 131–137.
Richardson, J. T. E. (1980). Mental imagery and human
memory. London: Macmillan.
Rimmele, U., Davachi, L., & Phelps, E. A. (2012). Memory for
time and place contributes to enhanced confidence in
memories for emotional events. Emotion, 12, 834–846.
Roozendaal, B., McEwen, B. S., & Chattarji, S. (2009). Stress, memory
and the amygdala. Nature Reviews Neuroscience, 10, 423–433.
Scoville, W. B., & Milner, B. (2000). Loss of recent memory
after bilateral hippocampal lesions. 1957. Journal of
Neuropsychiatry and Clinical Neurosciences, 12, 103–113.
Sergerie, K., Chochol, C., & Armony, J. L. (2008). The role of
the amygdala in emotional processing: A quantitative
meta-analysis of functional neuroimaging studies.
Neuroscience and Biobehavioral Reviews, 32, 811–830.
Slotnick, S. D., & Dodson, C. S. (2005). Support for a
continuous (single-process) model of recognition memory
and source memory. Memory & Cognition, 33, 151–170.
Smith, D. M., & Bulkin, D. A. (2014). The form and function
of hippocampal context representations. Neuroscience
and Biobehavioral Reviews, 40, 52–61.
M., Hare, T. A., et al. (2009). The NimStim set of facial
expressions: Judgments from untrained research participants.
Psychiatry Research, 168, 242–249.
Tsivilis, D., Otten, L. J., & Rugg, M. D. (2001). Context effects on
the neural correlates of recognition memory: An
electrophysiological study. Neuron, 31, 497–505.
Tulving, E. (1972). Episodic and semantic memory. In E.
Tulving & W. Donaldson (Eds.), Organization of memory
(pp. 381–403). New York: Academic Press.
Tulving, E., & Thomson, D. M. (1973). Encoding specificity and
retrieval processes in episodic memory. Psychological
Review, 80, 352–373.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D.,
Crivello, F., Etard, O., Delcroix, N., et al. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject
brain. Neuroimage, 15, 273–289.
Uncapher, M. R., & Rugg, M. D. (2005). Encoding and the
durability of episodic memory: A functional magnetic resonance
imaging study. Journal of Neuroscience, 25, 7260–7267.
van Ast, V., Cornelisse, S., Meeter, M., Joëls, M., & Kindt, M.
(2013). Time-dependent effects of cortisol on the
contextualization of emotional memories. Biological
Psychiatry, 74, 809–816.
van Ast, V., Cornelisse, S., Meeter, M., & Kindt, M. (2014). Cortisol
mediates the effects of stress on the contextual dependency of
memories. Psychoneuroendocrinology, 41, 97–110.
Wagner, A. (1998). Prefrontal cortex and recognition memory.
Functional-MRI evidence for context-dependent retrieval
processes. Brain, 121, 1985–2002.
Yonelinas, A. P. (2013). The hippocampus supports high-
resolution binding in the service of perception, working
memory and long-term memory. Behavioural Brain
Research, 254, 34–44.
Zeidman, P., Mullally, S. L., & Maguire, E. A. (2015). Constructing,
perceiving, and maintaining scenes: Hippocampal activity
and connectivity. Cerebral Cortex, 25, 3836–3855.
Zhang et al.
593
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
4
5
7
9
1
7
8
7
2
1
0
/
j
o
c
n
_
a
_
0
1
2
1
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3