Low Vision Impairs Implicit Sensorimotor Adaptation in
Response to Small Errors, But Not Large Errors
Jonathan S. Tsay , Steven Tan, Marlena A. Chu, Richard B. Ivry, and Emily A. Tonnelier
Abstrait
■ Successful goal-directed actions require constant fine-tuning of
the motor system. This fine-tuning is thought to rely on an implicit
adaptation process that is driven by sensory prediction errors (par exemple.,
where you see your hand after reaching vs. where you expected it
to be). Individuals with low vision experience challenges with
visuomotor control, but whether low vision disrupts motor adap-
tation is unknown. To explore this question, we assessed
individuals with low vision and matched controls with normal
vision on a visuomotor task designed to isolate implicit adaptation.
We found that low vision was associated with attenuated implicit
adaptation only for small visual errors, but not for large visual
errors. This result highlights important constraints underlying
how low-fidelity visual information is processed by the sensorimo-
tor system to enable successful implicit adaptation. ■
INTRODUCTION
Our ability to enact successful goal-directed actions
derives from multiple learning processes (McDougle
et coll., 2016; Bond & Taylor, 2015; Haith, Huberdeau, &
Krakauer, 2015; McDougle, Bond, & Taylor, 2015; Taylor,
Krakauer, & Ivry, 2014; Taylor & Ivry, 2011; Keisler &
Shadmehr, 2010). Among these processes, implicit motor
adaptation is key for ensuring that the sensorimotor sys-
tem remains well-calibrated in response to changes in
the body (par exemple., muscle fatigue) and in the environment
(par exemple., a heavy jacket). This adaptive process is driven by
a mismatch between the predicted sensory feedback
generated by the cerebellum and the actual sensory feed-
back arriving from the periphery—that is, sensory predic-
tion error (Kim, Avraham, & Ivry, 2021; Shadmehr, Forgeron,
& Krakauer, 2010).
Uncertainty in the sensory feedback has been shown to
reduce the system’s sensitivity to the feedback signal (c'est à dire.,
learning rate) et, as such, reduces the extent of implicit
adaptation (Ferrea, Franke, Morel, & Gail, 2022; Shyr &
Joshi, 2021; Samad, Chung, & Shams, 2015; van Beers,
2012; Wei & Conduite, 2010; Burge, Ernst, & Banks, 2008;
van Beers, Wolpert, & Haggard, 2002). This phenomenon
can be accounted for by an optimal integration model.
According to this model, the learning rate reflects a Bayes
optimal weighting between the sensory feedback and
feedforward prediction (Kawato, Ohmae, Hoang, &
Sanger, 2021; Shadmehr et al., 2010; Wei & Conduite,
2010; Burge et al., 2008; Conduite & Wolpert, 2004; Ito,
1986; Albus, 1971; Marr, 1969). When sensory noise is
haut, the model stipulates that this integration process
Université de Californie, Berkeley
lowers the weight given to the feedback signal, reduces
the strength of the resultant error, et, as such, attenuates
implicit adaptation for all error sizes.
Recent work has discovered an unappreciated con-
straint on this error integration process (Tsay, Avraham,
et coll., 2021). Uncertain visual feedback was found to only
attenuate adaptation when visual sensory prediction
errors were small, but not when they were large. Cependant,
sensory feedback noise was manipulated in a relatively
coarse, unnatural extrinsic environmental manipulation
(c'est à dire., a dispersed cloud of visual feedback). In the current
étude, we sought to examine how implicit motor adapta-
tion is affected by sensory uncertainty arising from intrin-
sic noise within the neural circuitry conveying sensory
feedback. Understanding how sensory uncertainty affects
implicit adaptation under a broad range of circumstances
serves to constrain our computational and neural models
of sensorimotor learning.
We used a Web-based visuomotor rotation task to assess
implicit adaptation in individuals with diverse forms of
visual impairments—that is, low vision because of reduced
visual acuity, reduced contrast sensitivity, or restricted visual
field. Although prior work has shown that low vision is
associated with impaired motor control (Cheong, Ling,
& Shehab, 2022; Lenoble, Corveleyn, Tran, Rouland, &
Boucart, 2019; Endo et al., 2016; Verghese, Tyson,
Ghahghaei, & Fletcher, 2016; Pardhan, Gonzalez-Alvarez,
& Subramanien, 2012; Timmis & Pardhan, 2012; Kotecha,
O’Leary, Melmoth, Grant, & Crabb, 2009; Jacko et al.,
2000), the effect of low vision on motor learning has not
been investigated. We hypothesized that low vision—a
heterogeneous set of visual impairments—would also
attenuate implicit adaptation for small, but not large, visuel
errors similar to the effect of extrinsic visual noise. Notre
© 2023 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 International (CC PAR 4.0) Licence.
Journal des neurosciences cognitives 35:4, pp. 736–748
https://doi.org/10.1162/jocn_a_01969
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
un
_
0
1
9
6
9
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
results support this hypothesis, providing converging
evidence for how low-fidelity visual information is proc-
essed by the sensorimotor circuitry to enable successful
implicit adaptation.
MÉTHODES
Ethics Statement
All participants gave written informed consent in accor-
dance with policies approved by the institutional review
board (protocol number: 2016-02-8439). Participation in
the study was in exchange for monetary compensation.
Participants
Individuals with impaired visual function that interferes
with the activities of daily life (c'est à dire., low vision) étaient
recruited through Meredith Morgan Eye Center and via
word of mouth. Potential participants were screened using
an on-line survey and were excluded if they did not have
a clinical diagnosis related to low vision (par exemple., macular
degeneration, glaucoma, Stargardt’s disease), or if their
self-reported visual acuity (c'est à dire., “Recall your visual acuity
results from your clinician-administered eye exam within
the last year”) in their best-seeing eye was better than
20/30 (c'est à dire., 0.2 logMAR). Participants also reported if their
low vision was related to peripheral and/or central vision, si
it was present since birth or acquired later in life (denoted
hereafter as early versus late onset), and if they had difficulty
seeing road signs; specifically, participants were prompted
with a Likert scale from 1 (road signs are very blurry) à 7
(road signs are very clear). This functional measure
correlates negatively with visual acuity (R = −.5, p = .04).
En outre, participants responded to five survey ques-
tions about whether their low vision condition affected
their day-to-day function. The questions were stated as
follows: (1) Do you use any mobility or navigational aids?
(2) Do you have difficulty detecting an edge of a step?
(3) Do you have difficulty pouring water into a cup? (4)
Do you have difficulty walking up and down stairs? (5) Do
you have difficulty detecting obstacles? Using these self-
report responses, we calculated a “visual impairment
index” by tallying the number of “yes” responses and
dividing this number by five (c'est à dire., the number of ques-
tion). A higher number denotes greater visual impair-
ment (max = 1; min = 0; Tableau 1).
To our knowledge, this is the first study to examine sen-
sorimotor learning in individuals with low vision. Ainsi, le
sample size was determined based on similar neuropsy-
chological studies examining sensorimotor learning in
different patient groups (par exemple., cerebellar degeneration,
Parkinson’s disease; Tsay, Najafi, Schuck, Wang, & Ivry,
2022; Tsay, Schuck, & Ivry, 2022; Tseng, Diedrichsen,
Krakauer, Shadmehr, & Bastian, 2007). Each participant
completed two sessions that were spaced at least 24 hr
apart to minimize any savings or interference (Avraham,
Morehead, Kim, & Ivry, 2021; Lerner et al., 2020; Krakauer,
Ghez, & Ghilardi, 2005). This amounted to a total of 40 sur-
line test sessions, with each session lasting approximately
45 min. Note that none of the participants with low vision
reported using special devices to augment their vision dur-
ing the experiment.
We also recruited 20 matched control participants via
Prolific, a Web site for online participant recruitment, à
match the low vision group based on age, sex, handed-
ness, and years of education. All control participants com-
pleted two sessions, which amounted to 40 on-line test
sessions, each lasting approximately 45 min. Participants
on Prolific have been vetted through a screening proce-
dure to ensure data quality. Two sessions from the control
data were incomplete because of technical difficulties and
thus not included in the analyses.
By design, the low vision and control groups did not dif-
fer significantly in age, t(36) = −0.6, p = .58, μ =−3.3,
[−15.4, 8.8], D = 0.2; control mean = 46.4, SD = 16.4
années; low vision mean = 49.7, SD = 21.1 années; handed-
ness, χ2(1) = 0, p = .93; both groups = 17 right-handers
and three ambidextrous individuals; sex, χ2(1) = 6.1,
p = .05; control = 14 female and six male participants,
low vision = 11 female and eight male participants, un
declined to specify; or years of education, t(38) = −0.1,
p = .89, μ = −0.1, [−1.5,1.3], D = 0; control = 17.0 ±
2.2, low vision = 17.1 ± 2.3 (Tableau 1). As expected, le
low vision group reported significantly more visual impair-
ments compared with the control group based on their
self-reported difficulty with reading road signs, t(25) =
16.0, p < .001,μ = 4.6, [4.0, 5.1], D = 5.1; control = 6.7 ±
0.5, low vision = 2.2 ± 1.2.
The participants with low vision completed the task
with the experimenter on the phone, and thus, available
to provide instructions and monitor performance. The
control participants completed the task on their own,
accessing the Web site at their convenience.
Apparatus
Participants used their own computer to access a Web
page hosted on Google Firebase. The task was created
using the OnPoint platform (Tsay, Lee, Ivry, & Avraham,
2021), and the task progression was controlled by
JavaScript code running locally in the participant’s Web
browser. The size and position of stimuli were scaled
based on each participant’s screen size/resolution (height =
920 ± 180 px, width = 1618 ± 433 px), which was auto-
matically detected. As such, any differences in screen size
and screen magnification were accounted for between
individuals. For ease of exposition, the stimulus parame-
ters reported below reflect the average screen resolution
in our participant population. Importantly, before starting
the experiment, the experimenter verified that partici-
pants were seated at a comfortable distance away from
the screen (20–30 in.) and were able to comfortably see
the various visual stimuli on the screen (e.g., the blue tar-
get and the white feedback cursor). In our prior validation
Tsay et al.
737
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
7
3
8
J
o
u
r
n
a
l
o
f
C
o
g
n
i
t
i
v
e
N
e
u
r
o
s
c
i
e
n
c
e
V
o
l
u
m
e
3
5
,
N
u
m
b
e
r
4
Table 1. Participants’ Reponses to Questions about Their Low Vision
#
Age
Hand
Etiology
Visual Acuity
in Better Eye
Diff. Seeing
Road Signs
Peripheral
Visual Field
Central
Visual Field
Low Vision
Onset
Visual Impairment
Index
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
31
34
60
60
86
70
34
61
26
60
55
86
29
59
31
57
24
78
24
28
R
R
A
R
R
R
R
A
R
R
R
R
R
R
R
A
R
R
R
R
Retinitis pigmentosa
Achromatopsia
Cerebral visual impairment
Infection
Glaucoma
Macular degeneration
Pseudotumor cerebri
Rieger syndrome
Genetic
Infection
Stargardt disease
Glaucoma
Glaucoma
Diabetic retinopathy
Glaucoma
Glaucoma
Albinism
Macular degeneration
Optic nerve atrophy
Nystagmus
0.3
0.9
0.4
1.7
1.0
0.9
1.3
0.2
0.9
1.2
1.0
1.0
0.7
1.2
1.7
1.0
1.2
0.6
0.8
0.5
2
1
3
1
2
2
2
2
3
2
1
1
3
2
1
5
1
4
1
4
Impaired
Intact
Impaired
Intact
Intact
Intact
Intact
Impaired
Impaired
Intact
Intact
Impaired
Impaired
Intact
Intact
Impaired
Impaired
Impaired
Impaired
Impaired
Intact
Impaired
Intact
Intact
Intact
Impaired
Impaired
Intact
Intact
Impaired
Impaired
Intact
Intact
Intact
Intact
Intact
Impaired
Impaired
Intact
Intact
Late
Early
Late
Late
Late
Late
Late
Late
Late
Early
Late
Late
Late
Late
Early
Early
Early
Late
Late
Early
0.8
0.6
1
0.6
1
0.2
1
1
0.8
0.6
0.8
0.6
1
0.8
0
1
0.4
1
0.2
0.2
Age is reported in years, and handedness is reported as right (R) or ambidextrous (A) (no participants in this group were left-handed). Self-reports of visual acuity of the better seeing eye (logMAR) and
peripheral/central visual field loss are provided. Difficulty with reading road signs is self-reported on a scale from 1 to 7. Low vision onset was self-reported as early onset, late onset, or unknown (no one
responded unknown). Visual impairment scores denoted the degree to which low vision impacted activities of daily living (1 = most severe impairment; 0 = least severe impairment).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
work using this on-line interface and procedure, the exact
movement and the exact device used did not impact mea-
sures of performance or learning on visuomotor adapta-
tion tasks (Tsay, Lee, et al., 2021). We note that, unlike
our laboratory-based setup in which we occlude vision of
the reaching hand, this was not possible with our on-line
testing protocol. That being said, we have found that mea-
sures of implicit and explicit adaptation are similar
between in-person and on-line settings (Tsay, Lee, et al.,
2021). Moreover, based on our informal observations, par-
ticipants remain focused on the screen during the experi-
ment (to see the target and how well they are doing), so
vision of the hand would be limited to the periphery.
Reaching Task Stimuli and General Procedure
During the task, the participant performed small reaching
movements by moving their computer cursor with their
trackpad or mouse. The participant’s mouse or trackpad
sensitivity (gain) was not modified, but rather left at the
setting the participant was familiar with. On each trial, par-
ticipants made a center-out planar movement from the
center of the workspace to a visual target. A white annulus
(1% of screen height: 0.24 cm in diameter) indicated the
start location at the center of the screen, and a red circle
(1% of screen height: 0.24 cm in diameter) indicated the
target location. The radial distance of the target from the
start location was 8 cm (40% of screen height). The target
could appear at one of three directions from the center.
Measuring angles counterclockwise and defining right-
ward as 0°, these directions were: 30° (upper right quad-
rant), 150° (upper left quadrant), and 270° (straight down).
Within each experiment, triads of trials (i.e., a cycle) con-
sisted of one trial to each of the three targets. The order in
which the three targets were presented was randomized
within each cycle. Note that participants with color vision
deficits could still do the task because position informa-
tion also indicated the difference between the start loca-
tion and target location.
At the beginning of each trial, participants moved their
cursor to the start location at the center of their screen.
Cursor position feedback, indicated by a white dot (0.6%
of screen height: 0.1 cm in diameter), was provided when
the cursor was within 2 cm of the start location (10% of
screen height). After maintaining the cursor in the start
location for 500 msec, the target appeared at one of three
locations (see above). Participants were instructed to
move rapidly to the target, attempting to “slice” through
the target. If the movement was not completed within
400 msec, the message “too slow” was displayed in red
20 pt. Times New Roman font at the center of the screen
for 400 msec.
Feedback during this movement phase could take one
of the following forms: veridical feedback, no-feedback, or
rotated noncontingent (“clamped”) feedback. During
veridical feedback trials, the movement direction of the
cursor was veridical with respect to the movement
direction of the hand. That is, the cursor moved with their
hand as would be expected for a normal computer cursor.
During no-feedback trials, the cursor was extinguished as
soon as the hand left the start annulus and remained off for
the entire reach. During rotated clamped feedback trials,
the cursor moved along a fixed trajectory relative to the
position of the target—a manipulation shown to isolate
implicit motor adaptation (Tsay, Parvin, & Ivry, 2020; R.
Morehead, Taylor, Parvin, & Ivry, 2017). The clamp was
temporally contingent on the participant’s movement,
matching the radial distance of the hand from the center
circle, but noncontingent on the movement in terms of
its angular offset relative to the visual target. The fixed angu-
lar offset (with respect to the target) was either 3° or 30°
(see below). The participant was instructed to “ignore the
visual feedback and reach directly to the target.”
For all feedback trials, the radial position of the cursor
corresponded to that of the hand up to 8 cm (the radial
distance of the target), at which point the cursor position
was frozen for 50 msec, before disappearing. After com-
pleting a trial, participants moved the cursor back to the
starting location. The visual cursor remained invisible until
the participant moved within 2 cm of the start location, at
which point the cursor appeared without any rotation.
The Impact of Low Vision on Implicit Motor
Adaptation for Small and Large Errors
Participants with low vision and control participants (n =
20 per group) were tested in two sessions, with clamped
feedback used to induce implicit adaptation. Numerous
studies have observed that the degree of implicit adapta-
tion saturates for visual errors greater than 5° (Hayashi,
Kato, & Nozaki, 2020; Kim, Morehead, Parvin, Moazzezi,
& Ivry, 2018); thus, we examined implicit adaptation
in response to 3° errors (an error before the saturation
zone) and 30° errors (an error within the saturation zone;
Figure 1A). The session order and direction (clockwise or
counterclockwise) of the clamped rotation were counter-
balanced across individuals. Each session consisted of
75 cycles (225 trials total), distributed across three blocks:
baseline veridical feedback block (15 cycles), rotated
clamped feedback (50 cycles), and a no-feedback after-
effect block (10 cycles).
Before the baseline block, the instruction “Move
directly to the target as fast and accurately as you can”
appeared on the screen. Before the clamped feedback
block, the instructions were modified to read: “The white
cursor will no longer be under your control. Please ignore
the white cursor and continue to aim directly towards the
target.” To clarify the invariant nature of the clamped feed-
back, three demonstration trials were provided. On all
three trials, the target appeared directly above the start
location on the screen (90° position), and the participant
was told to reach to the left (Demo 1), to the right (Demo
2), and downward (Demo 3). On all three of these demon-
stration trials, the cursor moved in a straight line, 90° offset
Tsay et al.
739
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
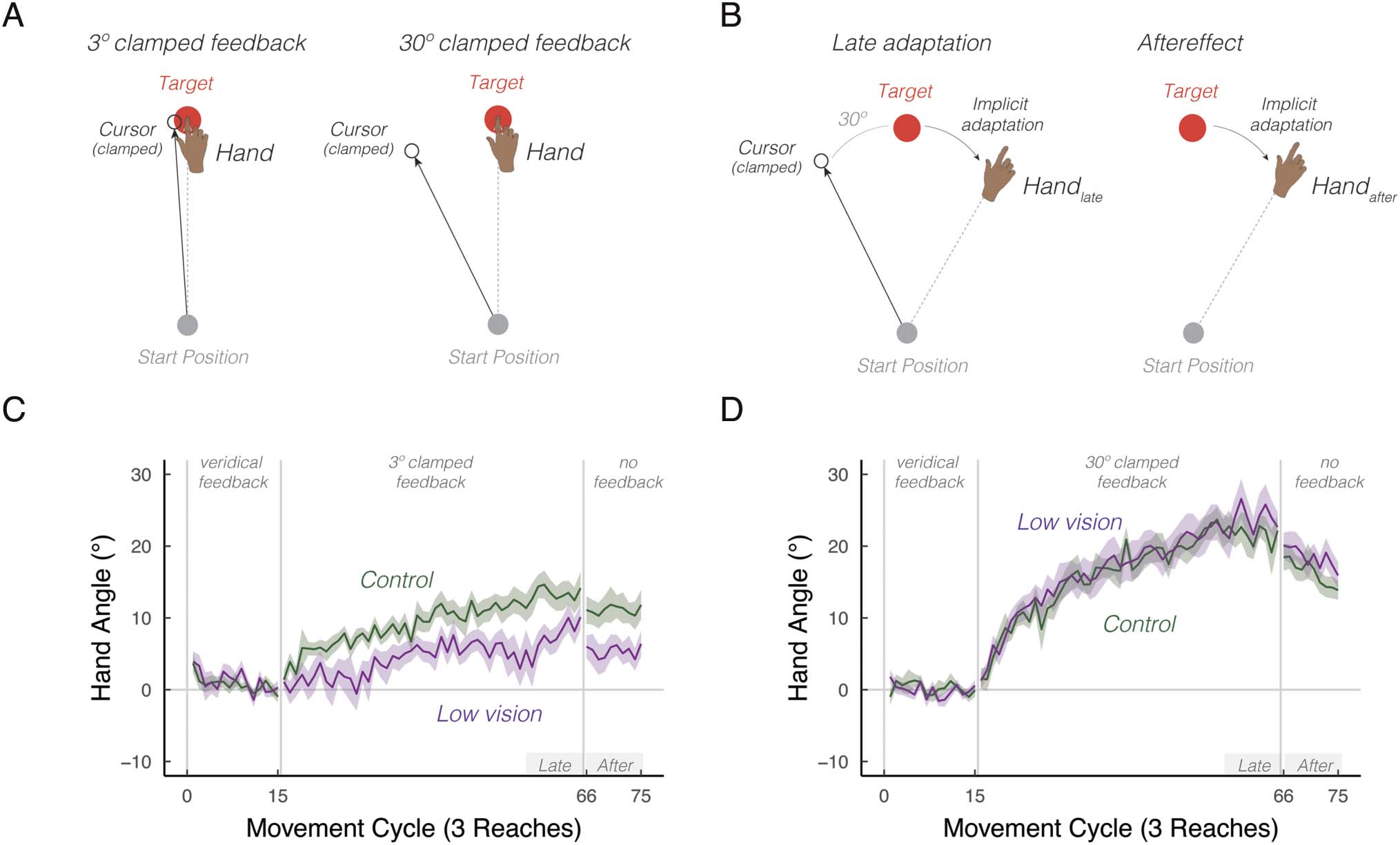
Figure 1. Intrinsic visual feedback uncertainty attenuates implicit adaptation in response to small, but not large errors. (A) Schematic of the clamped
feedback task. The sensory prediction error—the difference between the predicted visual feedback location (i.e., the target, red circle) and visual
feedback location (i.e., the cursor, white circle)—can either be small (3°) or large (30°). The cursor feedback follows a constant trajectory rotated
relative to the target, independent of the angular position of the participant’s hand. The rotation size remains invariant throughout the rotation
block. The participant was instructed to move directly to the target and ignore the visual feedback. A robust aftereffect is observed when the visual
cursor is removed during the no feedback block, implying that the clamp-induced adaptation is implicit. Note that participants reached toward three
targets: 30° (upper right quadrant), 150° (upper left quadrant), and 270° (straight-down). Only one target is shown in the schematic for ease of
illustration. (B) We defined two summary measures of learning: late adaptation (handlate) and aftereffect (handafter). Late adaptation reflects the
average hand angle relative to the target at the end of the clamped feedback block. Aftereffect reflects the average hand angle during the subsequent
no-feedback block. (C–D) Mean time courses of hand angle for 3° (C) and 30° (D) visual clamped feedback, for both the low vision (dark magenta)
and matched control (green) groups. Hand angle is presented relative to the baseline hand angle (i.e., last five cycles of the veridical feedback block).
Shaded region denotes SEM. Gray horizontal bars labeled Late and After indicate late and aftereffect phases of the experiment.
from the target. In this way, the participant could see that
the spatial trajectory of the cursor was unrelated to their
own reach direction. Before the no-feedback aftereffect
block, the participants were reminded to “Move directly
to the target as fast and accurately as you can.”
and ignore the white dot. Press ‘b’: I will aim directly
towards the target location and ignore the white dot.”
The experiment was terminated if participants did not
press the correct key (i.e., press “b”). Note that no partic-
ipants in either group were excluded based on these
attention and instruction checks.
Attention and Instruction Checks
It is difficult in on-line studies to verify that participants
fully attend to the task. To address this issue, we sporadi-
cally instructed participants to make specific keypresses:
“Press the letter ‘b’ to proceed.” If participants did not
press the correct key, the experiment was terminated.
These attention checks were randomly introduced within
the first 50 trials of the experiment. We also wanted to ver-
ify that the participants understood the clamped rotation
manipulation. To this end, we included one instruction
check after the three demonstration trials: “Identify the
correct statement. Press ‘a’: I will aim away from the target
Data Analysis
The primary dependent variable of reach performance was
hand angle, defined as the angle of the participant’s move-
ment location relative to the target when movement
amplitude reached an 8-cm radial distance from the start
position. Specifically, we measured the angle between a
line connecting the start position to the target and a line
connecting the start position to the position the partici-
pant moved to. Given that there is little generalization of
learning between target locations spaced more than 120°
apart (Morehead et al., 2017; Krakauer et al., 2005), the
740
Journal of Cognitive Neuroscience
Volume 35, Number 4
data are graphed by cycles. For visualization purposes, the
hand angles were flipped for blocks in which the clamp
was counterclockwise with respect to the target.
Outlier responses were defined as trials in which the
hand angle deviated by more than 3 SDs from a moving
five-trial window or if the hand angle was greater than
90° from the target (median percent of trials removed
per participant ± interquartile range (IQR): control = 0.1 ±
1.0%, low vision = 0.1 ± 1.0%).
The hand angle data were baseline corrected on an indi-
vidual basis to account for idiosyncratic angular biases in
reaching to the three target locations (Morehead & Ivry,
2015; Vindras, Desmurget, Prablanc, & Viviani, 1998).
These biases were estimated based on heading angles
during the last five veridical-feedback baseline cycles
(Trials 31–45), with these bias measures then subtracted
from the data for each cycle. We defined two summary
measures of
late adaptation and aftereffect
(Figure 1B). Late adaptation was defined as the mean
hand angle over the last 10 movement cycles of the rota-
tion block (Trials 166–195). The aftereffect was opera-
tionalized as the mean angle over all movement cycles
of the no-feedback aftereffect block (Trials 196–225).
learning:
These data were submitted to a linear mixed effects
model, with hand angle measures as the dependent vari-
able. We included experiment phase (late adaptation,
aftereffect), group (low vision or control), and error size
(3°, 30°) as fixed effects and participant ID as a random
effect. A priori, we hypothesized that the low vision group
would differ from the controls in their response to the
small errors.
We employed F tests with the Satterthwaite method to
evaluate whether the coefficients obtained from the linear
mixed effects model were statistically significant (R func-
tions: lmer, lmerTest, anova). Pairwise post hoc t tests
(two-tailed) were used to compare hand angle measures
between the low vision and control groups (R function:
emmeans). p Values were adjusted for multiple comparisons
using the Tukey method. The degrees of freedom were
also adjusted when the variances between groups were
not equal. Ninety-five percent confidence intervals for
group comparisons (t tests) obtained from the linear
mixed effects model are reported in squared brackets.
Standard effect size measures are also provided (D for
between-participants comparisons; D z for within-
participant comparisons; η2
p for between-subjects ANOVA;
Lakens, 2013).
RESULTS
Consistent with numerous prior studies, participants in
both groups showed a gradual change in hand angle in
the opposite direction of the clamped feedback, trending
toward an asymptotic level (Figure 1C–D; Tsay, Haith, Ivry,
& Kim, 2022; Kim, Parvin, & Ivry, 2019; Morehead et al.,
2017). Late adaptation was significant in all four condi-
tions, indicating robust implicit adaptation generated by
the clamped feedback, regardless of error size or partici-
pant vision level (3° controls: t(17) = 7.8, p < .001, μ =
13.4°, [9.9°, 17.1°], D = 1.8; 30° controls: t(17) = 13.9,
p < .001, μ = 21.6°, [18.4°, 24.9°], D = 3.1; 3° low vision:
t(19)= 5.6, p < .001, μ = 7.5°, [4.7°, 10.2°], D = 1.3); 30°
low vision: t(19) = 10.0, p < .001, μ = 23.8°, [18.8°, 28.8°],
D = 2.2). The main effect of Phase was not significant, F(1,
110) = 1.0, p = .32, ηp
2 = .0, indicating that implicit adap-
tation exhibited minimal decay back to baseline when
visual feedback was removed. Comparing the left and right
panels of Figure 1C and 1D, the learning functions were
higher when the error was 30° compared with when the
error was 3°, F(1, 112) = 21.6, p < .001, ηp
2 = .6, corrobo-
rating previous reports showing that implicit adaptation
increases with the size of the error (Kim et al., 2018;
Marko, Haith, Harran, & Shadmehr, 2012).
We next turned to our main question, asking how low
vision impacts implicit adaptation in response sensory
prediction errors. There was a significant interaction
between Group and Error Size, F(1, 111) = 10.5, p =
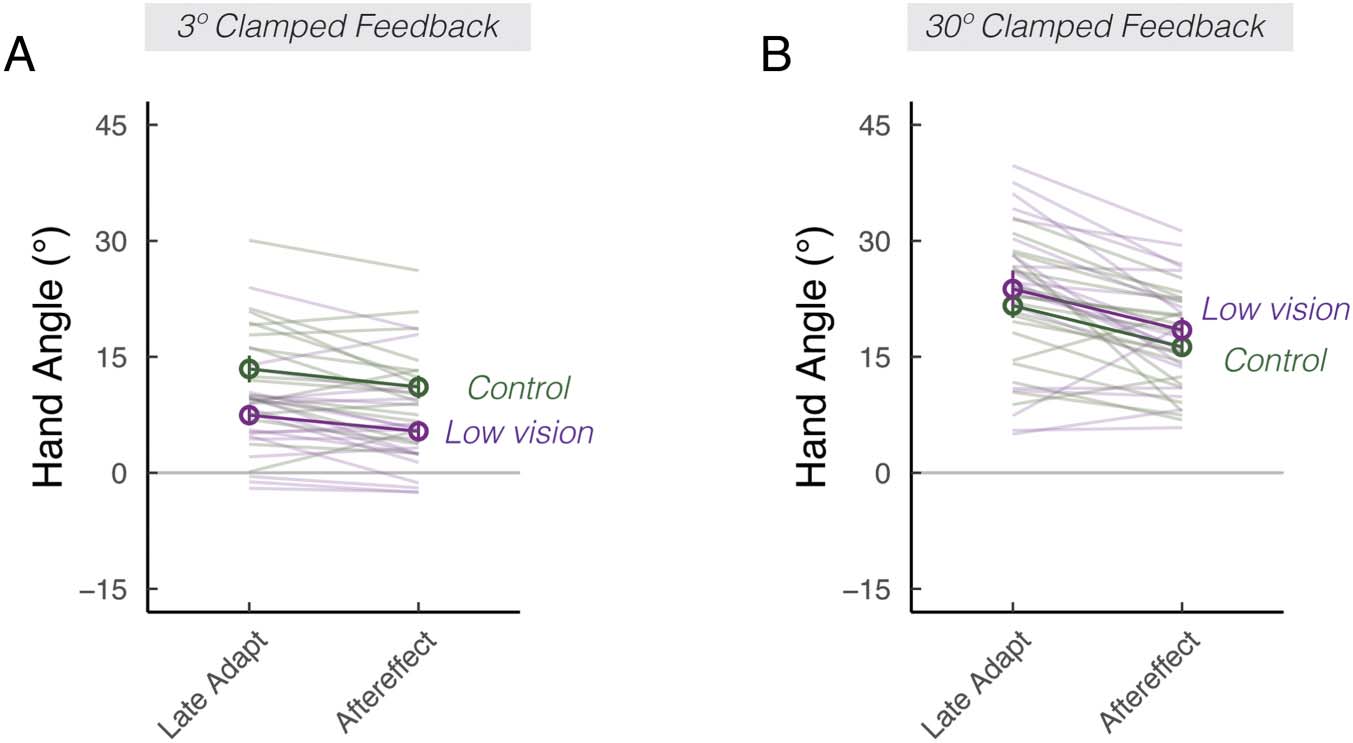
Figure 2. Visual uncertainty
attenuates implicit adaptation
in response to small, but not
large errors. Mean hand angles
± SEM during the late phase of
the clamped-feedback block,
and during the no-feedback
aftereffect block, for 3° (A) and
30° (B) clamped rotation
sessions. Thin lines denote
individual participants.
Tsay et al.
741
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
.002, ηp
2 = .2: Whereas the learning function between the
two groups were indistinguishable in response to a 30°
error (Figures 1D and 2B), the learning function in the
low vision group was attenuated compared with controls
in response to a 3° error (Figures 1C and 2A). This assess-
ment was confirmed by post hoc t tests, revealing that low
vision was associated with attenuated implicit adaptation
in response to the small error, t(60) = −3.0, p = .02,
μ = −5.8, [−9.6, −1.9], D = 0.9, but not the large error,
t(57) = 1.1, p = .67, μ = 2.2, [−1.6, 5.9], D = 0.2.
Together, these results underscored an unappreciated
implicit adaptation impairment associated with low vision,
but only when the size of the visual error was small.
Session Order Effect
Although the session order (3° or 30°) was fully counter-
balanced across participants, one potential concern in a
within-participant design of learning is that there may be
an effect of transfer or interference between sessions
(Avraham et al., 2021; Lerner et al., 2020; Krakauer et al.,
2005). For instance, experiencing a 30° clamped feedback
in the first session may interfere with learning in the sec-
ond session, resulting in attenuated learning. We did not
observe a significant Session Order effect on the extent of
motor aftereffects, although the effect was marginal, F(1,
36) = 3.7, p = .06, ηp
2 = .1. The key interaction between
Group and Error Size remained significant even when
Session Order was entered into the model as a covariate,
F(1, 36) = 10.5, p = .003, ηp
2 = .2, driven by a selective
attenuating effect of low vision on small errors, t(71) =
2.9, p = .03, μ = −5.8, [−11.1, −0.5], D = 1.1, but not large
errors, t(11) = 2.1, p = .66, μ = 2.3, [−2.9, 7.5], D = 0.5.
Kinematic Effects
There were no group differences in movement time (MT),
that is, the time between the start of the movement (i.e.,
1 cm from the center) and end of the movement (i.e., 8 cm
from the center; MT: t(34) = 0.6, p = .58, μ = 17.5, [−46.9,
81.9], D = 0.2; median MT ± IQR, low vision = 140.0 ±
106.5 msec; control = 106.4 ± 163.9 msec). In contrast,
RT, the interval between target presentation to the start
of movement, was, on average, slower in the low vision
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
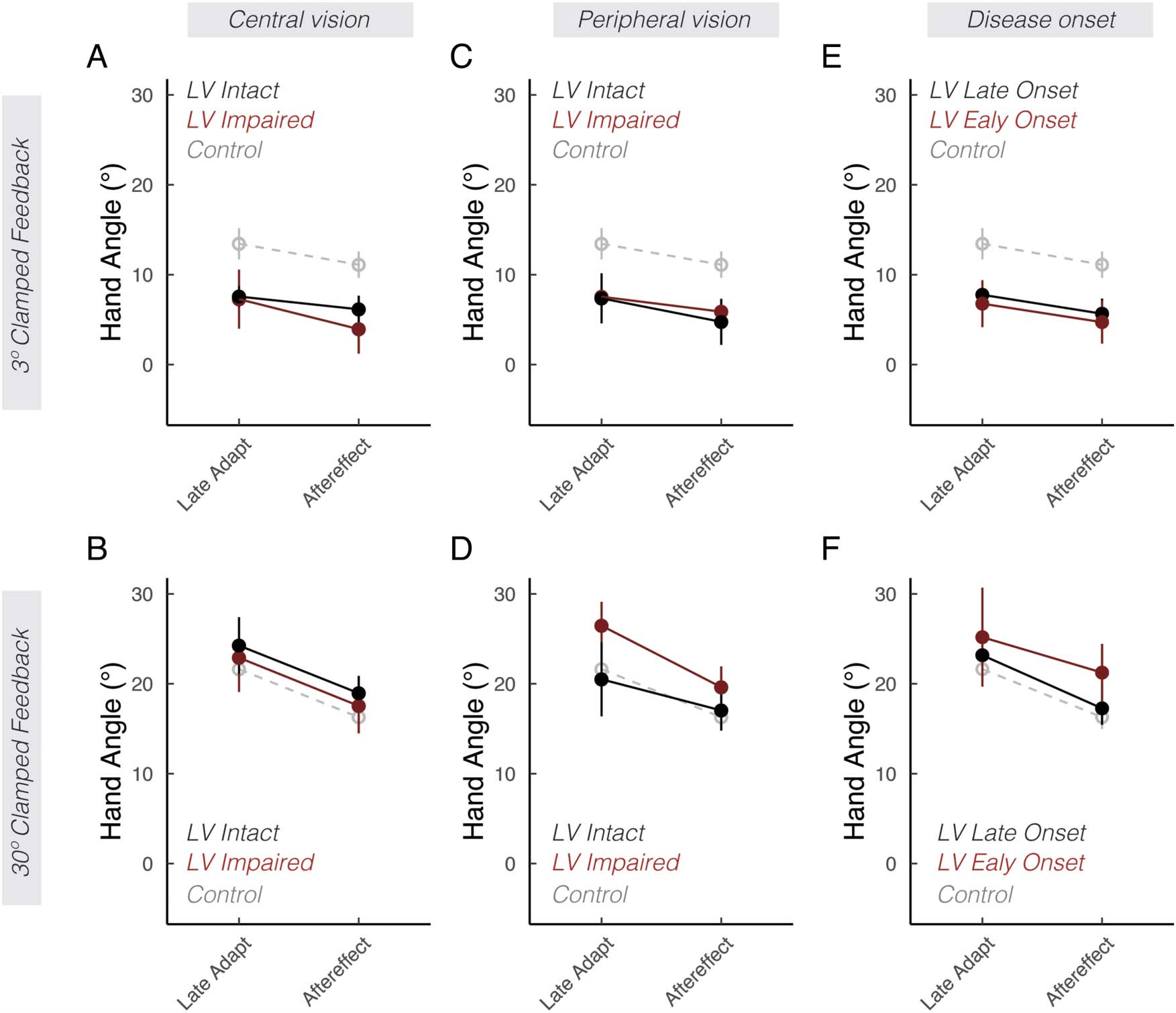
Figure 3. Low vision subgroup analyses. Mean hand angles ± SEM during late adaptation and aftereffect phases. Each column divides the low vision
(LV) group based on a different performance or clinical variable: central vision loss (A, B), peripheral vision loss (C, D), or disease onset (E, F). The
control group is shown in gray dashed lines.
742
Journal of Cognitive Neuroscience
Volume 35, Number 4
group compared with control group (RT: t(32) = 3.3, p =
.002, μ = 116.8, [47.6, 185.9], D = 1.1; median RT ± IQR,
low vision = 425.0 ± 206.5 msec; control = 317.0 ±
133.6 msec). On an individual level, RTs did not signifi-
cantly correlate with the degree of visual acuity (R = −.1,
p = .75), ability to see road signs (R = −.3, p = .14), or
visual impairment indices (R = .3, p = .18) in the low vision
group. These findings are consistent with the notion that
the group of individuals with low vision was impaired in
their ability to visually detect targets relative to the controls,
but that the ways in which low vision can impact target
acquisition in a visuomotor task like ours are multifaceted
and not necessarily predictable from low-dimensional
measures of visual function. The group-level RT difference
prompted us to include RT as a covariate in our analyses.
We found that implicit adaptation was not significantly
modulated by RT (main effect of RT: F(1, 95) = 1.2, p =
.27, ηp
2 = .0; 3° aftereffect correlated with baseline RT:
R = .01, p = .96; 30° aftereffect correlated with baseline
RT: R = .00, p = .99). The interaction between Group
and Error Size remained significant, F(1, 112) = 11.4, p =
.001, ηp
2 = .2, with the low vision group exhibiting attenu-
ated implicit adaptation in response to small errors, t(60) =
−2.4, p = .04, μ = −5.0, [0.9, 9.1], D = 0.9, but not large
errors, t(60) = −1.4, p = .47, μ = −3.1, [−7.2, 1.1], D = 0.2.
Subgroup Analysis
We also explored whether various subgroups of the partic-
ipants with low vision exhibited differences in implicit
adaptation. As shown in Figure 3, there were no apprecia-
ble differences between participants with and without
central vision loss (Figure 3A and B; F(1, 18) = 0.3, p =
.62, ηp
2 = .0), with and without peripheral vision loss
(Figure 3C and D; F(1, 18)= 1.0, p = .32, ηp
2 = .1), or
early versus late onset of low vision (Figure 3E and F;
F(1, 17) = 0.7, p = .51, ηp
2 = .1). Furthermore, there
were also no appreciable association between partici-
pants’ self-reports of visual acuity (Figure 4A and D),
ability to perceive road signs (Figure 4B and E), visual
impairment scores (Figures 4C and F), and clinical diag-
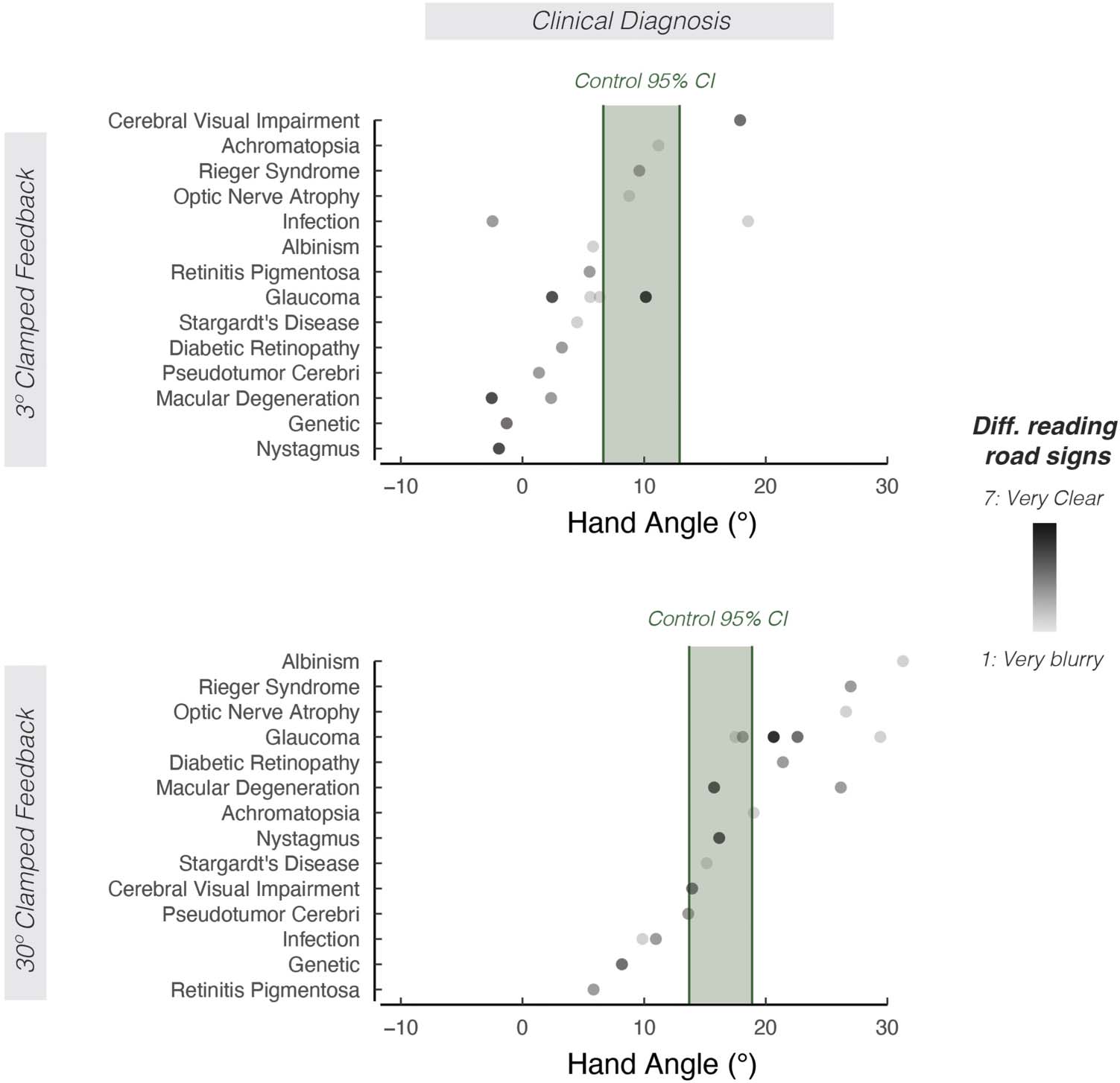
noses (Figure 5) with the extent of implicit adaptation. In
summary, we did not identify additional features among
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. The association between visual acuity and visual function and motor aftereffects. Correlation between visual acuity of the less impaired eye
and motor aftereffects (A, D). Correlation between how clearly participants report seeing road signs (1 = very clear; 7 = very blurry) and motor
aftereffects (B, E). Correlation between participant’s visual impairment index and motor aftereffects (C, F). The solid line indicates the regression
line, and the shaded region indicates SEM. The Spearman correlation is noted by R.
Tsay et al.
743
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. The effect of clinical diagnosis on motor aftereffects. Mean aftereffects sorted by clinical diagnoses involving low vision. Shading of the dot
indicates how well participants report seeing road signs (light shading = road signs are very blurry; dark shading = road signs are very clear). The
95% confidence interval for the control group is indicated by the green shaded region.
individuals in the low vision group that impacted implicit
adaptation.
DISCUSSION
Low vision can cause difficulty in discriminating the posi-
tion of visual objects (Timmis & Pardhan, 2012; Massof &
Fletcher, 2001). This impairment impacts motor perfor-
mance, resulting in slower and less accurate goal-directed
movements (Cheong et al., 2022; Lenoble et al., 2019;
Endo et al., 2016; Verghese et al., 2016; Pardhan et al.,
2012; Timmis & Pardhan, 2012; Kotecha et al., 2009; Jacko
et al., 2000). Here, we asked how low vision impacts motor
learning using a visuomotor adaptation task that isolates
implicit adaptation. The results revealed that low vision
was associated with attenuated implicit adaptation when
the sensory prediction error was small, but not when the
error was large. The error size by intrinsic visual uncer-
tainty interaction converges with a recent in-laboratory
study, in which sensory uncertainty was artificially
increased using different cursor patterns (Tsay, Avraham,
et al., 2021). Together, these results point to a strong
convergence between the effect of extrinsic uncertainty
in the visual stimulus (e.g., a foggy day) and intrinsic
uncertainty induced by low vision (e.g., damage to or
pathology of the visual system).
Potential Neural Learning Mechanisms that
May Give Rise to the Error Size by Visual
Uncertainty Interaction
An optimal integration hypothesis posits that intrinsic
uncertainty induced by low vision would be associated
with decreased sensitivity to errors and attenuate implicit
adaptation for all error sizes. Therefore, an optimal inte-
gration hypothesis cannot account for our results. That
being said, this error by uncertainty interaction can be
explained by a modified Bayesian perspective, which
posits that the nervous system performs causal inference
(Hong, Badde, & Landy, 2021; Shams & Beierholm, 2010;
Wei & Körding, 2009): Small errors, attributed to a misca-
librated movement (e.g., not reaching far enough to
retrieve a glass of water because of muscle fatigue), are
“relevant” and thus require implicit adaptation to nullify
744
Journal of Cognitive Neuroscience
Volume 35, Number 4
these sensorimotor errors; the weight given to these small
errors will fall off with increasing uncertainty. On the other
hand, large errors are more likely attributed to “irrelevant”
external sources from the environment (e.g., a missed
basketball shot because of a sudden gust of wind) and will
therefore get discounted by the sensorimotor system.
Paradoxically, the weight given to these large errors will
increase with uncertainty, because uncertainty can
obscure the attribution of large errors to an external
source. As such, the causal inference model predicts a
crossover point, where implicit adaptation will be higher
for small certain errors (compared with small uncertain
errors) but be lower for large certain errors (compared
with large uncertain errors; Wei & Körding, 2009).
A recent theory of implicit adaptation proposes an alter-
native possibility: The kinesthetic re-alignment hypothesis
centers on the notion that implicit adaptation is driven to
reduce a kinesthetic error, the mismatch between the per-
ceived and desired position of the hand, rather than a
visual error (Tsay, Kim, Haith, & Ivry, 2022). Note that in
the original exposition of this model (Tsay, Kim, et al.,
2022), we used the phrase “proprioceptive re-alignment.”
However, moving forward, we will adopt the term “kines-
thetic re-alignment” given that this better captures the
idea that the perceived position of the hand is an inte-
grated signal composed of multisensory inputs from vision
and peripheral proprioceptive afferents, as well as predic-
tive information from efferent signals (Proske & Gandevia,
2012). According to the kinesthetic re-alignment hypoth-
esis, visual uncertainty indirectly affects implicit adapta-
tion by influencing the magnitude of the kinesthetic shift,
that is, the degree to which visual feedback recalibrates
(biases) the perceived position of the hand (Cressman &
Henriques, 2011). When the visual error is small, visual
uncertainty attenuates the size of kinesthetic shifts and,
therefore, attenuates the degree of implicit adaptation.
When the visual error is larger than ∼10°, kinesthetic shifts
saturate and are therefore invariant to uncertainty (Tsay,
Kim, Parvin, Stover, & Ivry, 2021; ’t Hart, Ruttle, &
Henriques, 2020; Tsay et al., 2020). As such, visual uncer-
tainty has no impact on implicit adaptation when the
visual error is large. The mechanism driving kinesthetic
shifts (i.e., the extent to which vision biases/attracts the
perceived hand position) remains an active area for
research. Some suggest that these shifts are because of
mechanisms like causal inference (Hong et al., 2021; Wei
& Körding, 2009), whereas others hypothesize that these
mechanisms may follow a simple, fixed ratio rule (Zaidel,
Turner, & Angelaki, 2011). Our data motivate future
studies to directly evaluate the impact of visual uncertainty
on kinesthetic shifts and probe the neural correlates that
support this learning process.
Importantly, visual uncertainty was characterized rather
coarsely in the current study via recalling the results of a
recent clinician-administered eye exam and via self-
reporting how low vision impacted ability to carry out
activities of daily living. We acknowledge that without
more fine-grained psychophysical tests of visual acuity,
contrast sensitivity, and visual field loss, it is challenging
to evaluate quantitative differences in visual uncertainty
between the two study groups, or to examine potential
effects of individual differences. Thus, it is possible that
the effects observed between our two groups derive from
other differences between them that do not reflect differ-
ent levels of visual function. For example, the two groups
may have viewed their screens at slightly different dis-
tances or with differing levels of brightness. To mitigate
this concern, we have tried our best to standardize our
setup. That is, we verified that participants all viewed the
screen at a similar, comfortable distance and were all able
to see the visual stimuli without using any compensatory
viewing strategies. In addition, we note that possible
setup differences would likely result in main effects
between groups (e.g., participants with low vision paying
less attention to the visual feedback, and therefore atten-
uating adaptation for all visual error sizes; Parvin, Dang,
Stover, Ivry, & Morehead, 2022), rather than result in an
interaction between error size and visual uncertainty.
Detecting this interaction in a diverse sample of people
with low vision who have performed the task in a natural-
istic environment in fact highlights the robustness in our
results.
The Impact of Low Vision on Sensorimotor Control
and Learning
Previous work has shown that individuals with low vision
move slower and make more errors when performing
goal-directed movements ( Verghese et al., 2016; Pardhan
et al., 2012; Timmis & Pardhan, 2012; Pardhan, Gonzalez-
Alvarez, & Subramanian, 2011; Kotecha et al., 2009).
Although these deficits are observed in people with both
central and peripheral vision loss, reductions in central
vision appear to be the key limiting factor (Pardhan
et al., 2011, 2012). Central vision loss, which can result
in lower acuity and contrast sensitivity, likely worsens
the ability to precisely locate the intended visual target
as well as respond to the sensory predictions conveying
motor performance, an impairment that would be espe-
cially pronounced when the target and error are small
(Legge, Parish, Luebker, & Wurm, 1990; Tomkinson,
1974). Interestingly, in our Web-based studies, we did
not observe strong associations between subjective mea-
sures of visual ability and implicit adaptation (see
Figures 3–4). That being said, we readily acknowledge that
our Web-based approach (adopted to continue research
during the global pandemic) offers preliminary evidence
for the impact of low vision on sensorimotor learning.
We opted to recruit a diverse cohort of low vision partici-
pants, one that is largely representative of the diversity
inherent to low vision. By administering a more detailed
psychophysical battery, future follow-up studies would
be able to home in on how different visual impairments
Tsay et al.
745
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
(e.g., contrast sensitivity, color sensitivity) may jointly
impact the extent of implicit adaptation.
From a practical perspective, our results provide the
first characterization of how low vision affects not only
motor performance, but also motor learning. Specifically,
when the sensory inputs to the sensorimotor system can-
not be clearly disambiguated because of low vision (i.e.,
small and uncertain visual errors), the extent of implicit
adaptation is attenuated. However, when visual errors
are clearly disambiguated despite having low fidelity
(i.e., large and uncertain errors), the extent of implicit
adaptation is not impacted by low vision. This dissociation
underscores how the underlying learning mechanism per
se is not compromised by low vision and may be exploited
to enhance motor outcomes during clinical rehabilitation
(Tsay & Winstein, 2020). For example, clinicians and prac-
titioners could use nonvisual feedback (e.g., auditory or
tactile) to enhance the saliency and possibly reduce local-
ization uncertainty of small visual error signals (Endo et al.,
2016; Patel, Park, Bonato, Chan, & Rodgers, 2012). More-
over, rehabilitative specialists could provide explicit
instructions to highlight the presence of small errors,
such that individuals may learn to rely more on explicit
re-aiming strategies to compensate for these errors
(Merabet, Connors, Halko, & Sánchez, 2012). Future work
could examine which of these techniques is most effective
to enhance motor learning when errors are small.
Reprint requests should be sent to Jonathan Tsay, Department
of Psychology, University of California, Berkeley, 2121 Berkeley
Way West, Berkeley, CA 96704, United States, or via e-mail:
xiaotsay2015@gmail.com.
Data Statement Availability
Raw data and code can be accessed at https://datadryad.org/stash
/share/thhAGiZyDdcHQMqQYa6iwLITFNUpxwJbOqWnvmNHqsU.
Author Contributions
Jonathan S. Tsay: Conceptualization; Data curation; Formal
analysis; Funding acquisition; Investigation; Methodology;
Software; Supervision; Validation; Visualization; Writing—
Original draft; Writing—Review & editing. Steven Tan:
Data curation; Investigation. Marlena A. Chu: Resources;
Writing—Review & editing. Richard B. Ivry: Funding
acquisition; Methodology; Writing—Review & editing.
Emily A. Cooper: Conceptualization; Methodology; Super-
vision; Visualization; Writing—Review & editing.
Funding Information
Jonathan S. Tsay, National Institute of Neurological Disor-
ders and Stroke (https://dx.doi.org/10.13039/100000065),
grant number: 1F31NS120448; Richard B. Ivry, National
Institute of Neurological Disorders and Stroke (https://dx
. d o i . o r g / 1 0 . 1 3 0 3 9 / 1 0 0 0 0 0 0 6 5 ) , g r a n t n u m b e r :
R35NS116883-01; Emily A. Cooper, NSF, grant number:
2041726. The funders had no role in study design, data
collection and analysis, decision to publish, or preparation
of the manuscript.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
REFERENCES
Albus, J. S. (1971). A theory of cerebellar function. Mathematical
Biosciences, 10, 25–61. https://doi.org/10.1016/0025-5564(71)
90051-4
Avraham, G., Morehead, R., Kim, H. E., & Ivry, R. B. (2021).
Reexposure to a sensorimotor perturbation produces
opposite effects on explicit and implicit learning processes.
PLoS Biology, 19, e3001147. https://doi.org/10.1371/journal
.pbio.3001147, PubMed: 33667219
Bond, K. M., & Taylor, J. A. (2015). Flexible explicit but rigid
implicit learning in a visuomotor adaptation task. Journal of
Neurophysiology, 113, 3836–3849. https://doi.org/10.1152/jn
.00009.2015, PubMed: 25855690
Burge, J., Ernst, M. O., & Banks, M. S. (2008). The statistical
determinants of adaptation rate in human reaching. Journal
of Vision, 8, 20.1–20.19. https://doi.org/10.1167/8.4.20,
PubMed: 18484859
Cheong, Y., Ling, C., & Shehab, R. (2022). An empirical
comparison between the effects of normal and low vision
on kinematics of a mouse-mediated pointing movement.
International Journal of Human–Computer Interaction, 38,
562–572. https://doi.org/10.1080/10447318.2021.1952802
Cressman, E. K., & Henriques, D. Y. P. (2011). Motor adaptation
and proprioceptive recalibration. Progress in Brain
Research, 191, 91–99. https://doi.org/10.1016/B978-0-444
-53752-2.00011-4, PubMed: 21741546
Endo, T., Kanda, H., Hirota, M., Morimoto, T., Nishida, K., &
Fujikado, T. (2016). False reaching movements in localization
test and effect of auditory feedback in simulated ultra-low
vision subjects and patients with retinitis pigmentosa. Graefe’s
Archive for Clinical and Experimental Ophthalmology, 254,
947–956. https://doi.org/10.1007/s00417-015-3253-2, PubMed:
26743752
Ferrea, E., Franke, J., Morel, P., & Gail, A. (2022). Statistical
determinants of visuomotor adaptation in a virtual reality
three-dimensional environment. Scientific Reports, 12, 10198.
https://doi.org/10.1038/s41598-022-13866-y
746
Journal of Cognitive Neuroscience
Volume 35, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Haith, A. M., Huberdeau, D. M., & Krakauer, J. W. (2015). The
influence of movement preparation time on the expression
of visuomotor learning and savings. Journal of Neuroscience,
35, 5109–5117. https://doi.org/10.1523/JNEUROSCI.3869-14
.2015, PubMed: 25834038
Hayashi, T., Kato, Y., & Nozaki, D. (2020). Divisively normalized
integration of multisensory error information develops motor
memories specific to vision and proprioception. Journal
of Neuroscience, 40, 1560–1570. https://doi.org/10.1523
/JNEUROSCI.1745-19.2019, PubMed: 31924610
Hong, F., Badde, S., & Landy, M. S. (2021). Causal inference
regulates audiovisual spatial recalibration via its influence
on audiovisual perception. PLoS Computational Biology,
17, e1008877. https://doi.org/10.1371/journal.pcbi.1008877,
PubMed: 34780469
Ito, M. (1986). Long-term depression as a memory process in
the cerebellum. Neuroscience Research, 3, 531–539. https://
doi.org/10.1016/0168-0102(86)90052-0, PubMed: 3534647
Jacko, J. A., Barreto, A. B., Marmet, G. J., Chu, J. Y. M., Bautsch,
H. S., Scott, I. U., et al. (2000). Low vision: The role of visual
acuity in the efficiency of cursor movement. In Proceedings
of the fourth international ACM conference on assistive
technologies (pp. 1–8).
Kawato, M., Ohmae, S., Hoang, H., & Sanger, T. (2021). 50 years
since the Marr, Ito, and Albus models of the cerebellum.
Neuroscience, 462, 151–174. https://doi.org/10.1016/j
.neuroscience.2020.06.019, PubMed: 32599123
Keisler, A., & Shadmehr, R. (2010). A shared resource between
declarative memory and motor memory. Journal of
Neuroscience, 30, 14817–14823. https://doi.org/10.1523
/JNEUROSCI.4160-10.2010, PubMed: 21048140
Kim, H. E., Avraham, G., & Ivry, R. B. (2021). The psychology of
reaching: Action selection, movement implementation, and
sensorimotor learning. Annual Review of Psychology, 72,
61–95. https://doi.org/10.1146/annurev-psych-010419-051053,
PubMed: 32976728
Kim, H. E., Morehead, R., Parvin, D. E., Moazzezi, R., & Ivry, R. B.
(2018). Invariant errors reveal limitations in motor correction
rather than constraints on error sensitivity. Communications
Biology, 1, 19. https://doi.org/10.1038/s42003-018-0021-y,
PubMed: 30271906
Kim, H. E., Parvin, D. E., & Ivry, R. B. (2019). The influence of
task outcome on implicit motor learning. eLife, 8, e39882.
https://doi.org/10.7554/eLife.39882, PubMed: 31033439
Körding, K. P., & Wolpert, D. M. (2004). Bayesian integration in
sensorimotor learning. Nature, 427, 244–247. https://doi.org
/10.1038/nature02169, PubMed: 14724638
Kotecha, A., O’Leary, N., Melmoth, D., Grant, S., & Crabb, D. P.
(2009). The functional consequences of glaucoma for
eye-hand coordination. Investigative Ophthalmology &
Visual Science, 50, 203–213. https://doi.org/10.1167/iovs.08
-2496, PubMed: 18806294
Krakauer, J., Ghez, C., & Ghilardi, M. F. (2005). Adaptation
to visuomotor transformations: Consolidation, interference,
and forgetting. Journal of Neuroscience, 25, 473–478.
https://doi.org/10.1523/JNEUROSCI.4218-04.2005, PubMed:
15647491
Lakens, D. (2013). Calculating and reporting effect sizes to
facilitate cumulative science: A practical primer for t tests and
ANOVAs. Frontiers in Psychology, 4, 863. https://doi.org/10
.3389/fpsyg.2013.00863, PubMed: 24324449
degeneration and glaucoma patients. Visual Cognition, 27,
732–739. https://doi.org/10.1080/13506285.2019.1661319
Lerner, G., Albert, S., Caffaro, P. A., Villalta, J. I., Jacobacci, F.,
Shadmehr, R., et al. (2020). The origins of anterograde
interference in visuomotor adaptation. Cerebral Cortex, 30,
4000–4010. https://doi.org/10.1093/cercor/bhaa016, PubMed:
32133494
Marko, M. K., Haith, A. M., Harran, M. D., & Shadmehr, R.
(2012). Sensitivity to prediction error in reach adaptation.
Journal of Neurophysiology, 108, 1752–1763. https://doi.org
/10.1152/jn.00177.2012, PubMed: 22773782
Marr, D. (1969). A theory of cerebellar cortex. Journal of
Physiology, 202, 437–470. https://doi.org/10.1113/jphysiol
.1969.sp008820, PubMed: 5784296
Massof, R. W., & Fletcher, D. C. (2001). Evaluation of the NEI
visual functioning questionnaire as an interval measure of
visual ability in low vision. Vision Research, 41, 397–413.
https://doi.org/10.1016/S0042-6989(00)00249-2, PubMed:
11164454
McDougle, S. D., Boggess, M. J., Crossley, M. J., Parvin, D., Ivry,
R. B., & Taylor, J. A. (2016). Credit assignment in movement-
dependent reinforcement learning. Proceedings of the
National Academy of Sciences, U.S.A., 113, 6797–6802.
https://doi.org/10.1073/pnas.1523669113, PubMed: 27247404
McDougle, S. D., Bond, K. M., & Taylor, J. A. (2015). Explicit and
implicit processes constitute the fast and slow processes
of sensorimotor learning. Journal of Neuroscience, 35,
9568–9579. https://doi.org/10.1523/JNEUROSCI.5061-14
.2015, PubMed: 26134640
Merabet, L. B., Connors, E. C., Halko, M. A., & Sánchez, J.
(2012). Teaching the blind to find their way by playing video
games. PLoS One, 7, e44958. https://doi.org/10.1371/journal
.pone.0044958, PubMed: 23028703
Morehead, J. R., & Ivry, R. (2015). Intrinsic biases systematically
affect visuomotor adaptation experiments. Neural Control
of Movement. https://ivrylab.berkeley.edu/uploads/4/1/1/5
/41152143/morehead_ncm2015.pdf
Morehead, R., Taylor, J. A., Parvin, D. E., & Ivry, R. B. (2017).
Characteristics of implicit sensorimotor adaptation revealed
by task-irrelevant clamped feedback. Journal of Cognitive
Neuroscience, 29, 1061–1074. https://doi.org/10.1162/jocn_a
_01108, PubMed: 28195523
Pardhan, S., Gonzalez-Alvarez, C., & Subramanian, A. (2011).
How does the presence and duration of central visual
impairment affect reaching and grasping movements?
Ophthalmic & Physiological Optics, 31, 233–239. https://doi
.org/10.1111/j.1475-1313.2010.00819.x, PubMed: 21410742
Pardhan, S., Gonzalez-Alvarez, C., & Subramanian, A. (2012).
Target contrast affects reaching and grasping in the visually
impaired subjects. Optometry and Vision Science, 89,
426–434. https://doi.org/10.1097/OPX.0b013e31824c1b89,
PubMed: 22366708
Parvin, D. E., Dang, K. V., Stover, A. R., Ivry, R. B., & Morehead,
J. R. (2022). Implicit adaptation is modulated by the relevance
of feedback. bioRxiv. https://doi.org/10.1101/2022.01.19
.476924
Patel, S., Park, H., Bonato, P., Chan, L., & Rodgers, M. (2012).
A review of wearable sensors and systems with application
in rehabilitation. Journal of Neuroengineering and
Rehabilitation, 9, 21. https://doi.org/10.1186/1743-0003-9-21,
PubMed: 22520559
Legge, G. E., Parish, D. H., Luebker, A., & Wurm, L. H. (1990).
Proske, U., & Gandevia, S. C. (2012). The proprioceptive
Psychophysics of reading. XI. Comparing color contrast
and luminance contrast. Journal of the Optical Society
of America, 7, 2002–2010. https://doi.org/10.1364/josaa.7
.002002, PubMed: 2231110
senses: Their roles in signaling body shape, body position
and movement, and muscle force. Physiological Reviews,
92, 1651–1697. https://doi.org/10.1152/physrev.00048.2011,
PubMed: 23073629
Lenoble, Q., Corveleyn, X., Tran, T. H. C., Rouland, J.-F., & Boucart,
M. (2019). Can I reach it? A study in age-related macular
Samad, M., Chung, A. J., & Shams, L. (2015). Perception of body
ownership is driven by Bayesian sensory inference. PLoS One,
Tsay et al.
747
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
10, e0117178. https://doi.org/10.1371/journal.pone.0117178,
PubMed: 25658822
Shadmehr, R., Smith, M. A., & Krakauer, J. (2010). Error
correction, sensory prediction, and adaptation in motor
control. Annual Review of Neuroscience, 33, 89–108. https://
doi.org/10.1146/annurev-neuro-060909-153135, PubMed:
20367317
Shams, L., & Beierholm, U. R. (2010). Causal inference in
perception. Trends in Cognitive Sciences, 14, 425–432.
https://doi.org/10.1016/j.tics.2010.07.001, PubMed: 20705502
Shyr, M. C., & Joshi, S. S. (2021). Validation of the Bayesian
sensory uncertainty model of motor adaptation with a
remote experimental paradigm. In 2021 IEEE 2nd
international conference on human-machine systems
(ICHMS) (pp. 1–6).
Taylor, J. A., & Ivry, R. B. (2011). Flexible cognitive strategies
during motor learning. PLoS Computational Biology, 7,
e1001096. https://doi.org/10.1371/journal.pcbi.1001096,
PubMed: 21390266
Taylor, J. A., Krakauer, J. W., & Ivry, R. B. (2014). Explicit and
implicit contributions to learning in a sensorimotor
adaptation task. Journal of Neuroscience, 34, 3023–3032.
https://doi.org/10.1523/JNEUROSCI.3619-13.2014, PubMed:
24553942
’t Hart, B. M., Ruttle, J. E., & Henriques, D. Y. P. (2020).
Proprioceptive recalibration generalizes relative to hand
position. https://deniseh.lab.yorku.ca/files/2020/05/tHart_SfN
_2019.pdf?x64373
Timmis, M. A., & Pardhan, S. (2012). The effect of central visual
impairment on manual prehension when tasked with
transporting-to-place an object accurately to a new location.
Investigative Ophthalmology & Visual Science, 53,
2812–2822. https://doi.org/10.1167/iovs.11-8860, PubMed:
22467581
Tomkinson, C. R. (1974). Accurate assessment of visual acuity in
low vision patients. Optometry and Vision, 51, 321–324.
https://doi.org/10.1097/00006324-197405000-00004, PubMed:
4830429
Tsay, J. S., Avraham, G., Kim, H. E., Parvin, D. E., Wang, Z., &
Ivry, R. B. (2021). The effect of visual uncertainty on implicit
motor adaptation. Journal of Neurophysiology, 125, 12–22.
https://doi.org/10.1152/jn.00493.2020, PubMed: 33236937
Tsay, J. S., Haith, A. M., Ivry, R. B., & Kim, H. E. (2022).
Interactions between sensory prediction error and task error
during implicit motor learning. PLoS Computational Biology,
18, e1010005. https://doi.org/10.1371/journal.pcbi.1010005,
PubMed: 35320276
Tsay, J. S., Kim, H., Haith, A. M., & Ivry, R. B. (2022).
Understanding implicit sensorimotor adaptation as a process
of proprioceptive re-alignment. eLife, 11, e76639. https://doi
.org/10.7554/eLife.76639, PubMed: 35969491
Tsay, J. S., Kim, H. E., Parvin, D. E., Stover, A. R., & Ivry, R. B.
(2021). Individual differences in proprioception predict the
extent of implicit sensorimotor adaptation. Journal of
Neurophysiology, 125, 1307–1321. https://doi.org/10.1152/jn
.00585.2020, PubMed: 33656948
Tsay, J. S., Lee, A., Ivry, R. B., & Avraham, G. (2021). Moving
outside the lab: The viability of conducting sensorimotor
learning studies online. Neurons, Behavior, Data Analysis,
and Theory. https://doi.org/10.51628/001c.26985
Tsay, J. S., Najafi, T., Schuck, L., Wang, T., & Ivry, R. B. (2022).
Implicit sensorimotor adaptation is preserved in Parkinson’s
disease. Brain Communications, 4, fcac303. https://doi.org
/10.1093/braincomms/fcac303, https://doi.org/10.1101/2022
.03.11.484047, PubMed: 36531745
Tsay, J. S., Parvin, D. E., & Ivry, R. B. (2020). Continuous reports
of sensed hand position during sensorimotor adaptation.
Journal of Neurophysiology, 124, 1122–1130. https://doi.org
/10.1152/jn.00242.2020, PubMed: 32902347
Tsay, J. S., Schuck, L., & Ivry, R. B. (2022). Cerebellar
degeneration impairs strategy discovery but not strategy
recall. Cerebellum. https://doi.org/10.1007/s12311-022-01500-6,
PubMed: 36464710
Tsay, J. S., & Winstein, C. J. (2020). Five features to look for in
early-phase clinical intervention studies. Neurorehabilitation
and Neural Repair, 35, 3–9. https://doi.org/10.1177
/1545968320975439, PubMed: 33243083
Tseng, Y.-W., Diedrichsen, J., Krakauer, J. W., Shadmehr, R., &
Bastian, A. J. (2007). Sensory prediction errors drive
cerebellum-dependent adaptation of reaching. Journal of
Neurophysiology, 98, 54–62. https://doi.org/10.1152/jn.00266
.2007, PubMed: 17507504
van Beers, R. J. (2012). How does our motor system determine
its learning rate? PLoS One, 7, e49373. https://doi.org/10.1371
/journal.pone.0049373, PubMed: 23152899
van Beers, R. J., Wolpert, D. M., & Haggard, P. (2002). When
feeling is more important than seeing in sensorimotor
adaptation. Current Biology, 12, 834–837. https://doi.org/10
.1016/S0960-9822(02)00836-9, PubMed: 12015120
Verghese, P., Tyson, T. L., Ghahghaei, S., & Fletcher, D. C.
(2016). Depth perception and grasp in central field loss.
Investigative Ophthalmology & Visual Science, 57, 1476–1487.
https://doi.org/10.1167/iovs.15-18336, PubMed: 27031841
Vindras, P., Desmurget, M., Prablanc, C., & Viviani, P. (1998).
Pointing errors reflect biases in the perception of the initial
hand position. Journal of Neurophysiology, 79, 3290–3294.
https://doi.org/10.1152/jn.1998.79.6.3290, PubMed: 9636129
Wei, K., & Körding, K. (2009). Relevance of error: What drives
motor adaptation? Journal of Neurophysiology, 101, 655–664.
https://doi.org/10.1152/jn.90545.2008, PubMed: 19019979
Wei, K., & Körding, K. (2010). Uncertainty of feedback and state
estimation determines the speed of motor adaptation.
Frontiers in Computational Neuroscience, 4, 11. https://doi
.org/10.3389/fncom.2010.00011, PubMed: 20485466
Zaidel, A., Turner, A. H., & Angelaki, D. E. (2011). Multisensory
calibration is independent of cue reliability. Journal of
Neuroscience, 31, 13949–13962. https://doi.org/10.1523
/JNEUROSCI.2732-11.2011, PubMed: 21957256
748
Journal of Cognitive Neuroscience
Volume 35, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
4
7
3
6
2
0
7
6
0
3
9
/
j
o
c
n
_
a
_
0
1
9
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3