Local Field Potentials and Spikes in the Human
Medial Temporal Lobe are Selective to
Image Category
Alexander Kraskov1, Rodrigo Quian Quiroga1,2, Leila Reddy3,
Itzhak Fried4,5, and Christof Koch1
D
o
w
n
je
o
un
d
e
d
F
r
o
m
Abstrait
& Local field potentials (LFPs) reflect the averaged dendro-
somatic activity of synaptic signals of large neuronal popula-
tion. Dans cette étude, we investigate the selectivity of LFPs and
single neuron activity to semantic categories of visual stimuli
in the medial temporal
lobe of nine neurosurgical patients
implanted with intracranial depth electrodes for clinical rea-
sons. Strong selectivity to the category of presented images
was found for the amplitude of LFPs in 8% of implanted mi-
croelectrodes and for the firing rates of single and multiunits
dans 14% of microelectrodes. There was little overlap between
the LFP- and spike-selective microelectrodes. Separate analy-
sis of the power and phase of LFPs revealed that the mean
phase was category-selective around the u frequency range
and that the power of the LFPs was category-selective for
high frequencies around the g rhythm. De la 36 microelec-
trodes with amplitude-selective LFPs, 30 were found in the
hippocampus. Enfin, it was possible to readout information
about the category of stimuli presented to the patients with
both spikes and LFPs. Combining spiking and LFP activity
enhanced the decoding accuracy in comparison with the ac-
curacy obtained with each signal alone, especially for short
time intervals. &
INTRODUCTION
Existing experimental approaches to investigate the
functions of the brain by recording electrical signals vary
from single-cell recordings in animals to surface electro-
encephalography (EEG) in humans. Animal electrophys-
iology usually relies on the analysis of spiking activity of
neurons, while placing less emphasis on local field
potentials (LFPs), a slow nonspiking component of the
recorded electrical signal. In contrast, studies of brain
electrical activity in humans typically use electrical po-
tentials recorded from the surface of the skull. Such EEG
signals represent the electrical activity of large fraction of
cortical and subcortical tissue but can still be linked to
different behavioral states or cognitive functions.
Certain neurological conditions,
in particular, phar-
macologically intractable epilepsy, require, on occasion,
the implantation of either subdural electrodes that rest
on the surface of the cortex, or of depth electrodes that
are implanted into the brain parenchyma (Ange, Moll,
Frit, & Ojemann, 2005; Lachaux, Rudrauf, & Kahane,
1California Institute of Technology, 2University of Leicester, ROYAUME-UNI,
3Massachusetts Institute of Technology, 4Université de Californie
at Los Angeles, 5Tel-Aviv University, Israel
2003; Bechtereva & Abdullaev, 2000). The signals ob-
tained with these intracranial electrodes represent aver-
age activity of the brain with temporal and spatial
resolution on the order of milliseconds and centimeters,
respectivement. The size and impedance of these clinically
used electrodes typically do not permit the recording of
spiking activity of neurons. Here we present data ob-
tained with microelectrodes implanted in the human
medial temporal lobe (MTL) of epilepsy patients. Their
impedance ((cid:1)0.5 M.(cid:1)) and size (diameter of the tip
(cid:1)40 Am) enabled us to record spiking activity of single
neurons as well as LFPs (Fried et al., 1999). The rela-
tionship between LFPs and spiking activity in nonpri-
mates was already addressed as early as the 1960s (voir
Haberly & Shepherd, 1973; John, 1967, 1972; Fromm,
1967; Buchwald, 1965 and Logothetis, 2003, for review).
There were also several studies dealing with the corre-
lation of spiking and epileptiform activity in epileptic
patients ( Wyler, Ojemann, & Ward, 1982; Verzeano,
Crandall, & Dymond, 1971). In monkey electrophysiol-
ogie, LFPs and their relationship to spiking activity have
been actively studied only in the last few years (Kreiman
et coll., 2006; Liu & Newsome, 2006; Henrie & Shapley,
2005; Scherberger, Jarvis, & Andersen, 2005; Mehring
et coll., 2003; Pesaran, Pezaris, Sahani, Mitra, & Andersen,
2002). Par exemple, the LFP activity in the parietal cortex
D 2007 Massachusetts Institute of Technology
Journal des neurosciences cognitives 19:3, pp. 479–492
je
je
/
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
un
/
d
m
e
je
d
t
F
r
p
o
r
m
c
.
h
s
je
p
je
v
d
e
je
r
r
e
c
c
h
t
.
m
un
je
r
e
.
d
c
toi
o
m
o
/
c
j
n
o
un
c
r
t
n
je
c
/
e
un
–
r
p
t
d
je
c
1
je
9
e
3
–
4
p
7
d
9
F
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
oui
7
g
.
toi
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
F
e
m
b
b
oui
e
r
g
2
toi
0
e
2
s
3
t
/
j
.
t
.
.
.
F
.
o
n
1
8
M.
un
oui
2
0
2
1
of macaque monkeys was found to discriminate between
preferred and antipreferred directions of reach plans
(Pesaran et al., 2002). The LFPs in monkey motor cortex
were also predictive of hand movements (Mehring et al.,
2003). Scherberger et al. (2005) reported that the mon-
key behavioral state can be decoded with LFPs better
than with the spiking activity. Increasing stimulus con-
trast was found to cause an increase in power,
dans
in gamma band, of the LFP recorded in
particular,
macaque V1 (Henrie & Shapley, 2005). De plus,
il
occurred over a contrast range within which the spike
rates of cortical cells were saturating. Recently, LFPs, comme
well as spiking activity recorded in the inferior tem-
poral cortex (IT) of macaque monkeys, were shown
to be object-selective but barely related to each other
(Kreiman et al., 2006). The aim of our study is to
investigate in humans, the selectivity of spiking and
LFP activity recorded simultaneously from the same
microelectrodes during a simple perceptual task, et
the relationship between their selectivities.
The selectivity of spiking activity recorded in the
human MTL to visual categories has already been re-
ported by our group (Kreiman, Koch, & Frit, 2000). Dans
this study, we investigated visual category selectivity
of the spiking activity and LFPs in a different group of
patients using the amplitude, pouvoir, and mean phase of
the LFPs. En outre, we compared selectivity properties
of the LFP and spiking activity and found a weak
correlation between them. We were also able to decode
information about the category of a presented stimulus
using the spiking and LFP activity separately and to-
gether. Decoding accuracy in a short time window was
found to be optimal using the LFPs and spiking activity
simultaneously as an input to the decoding algorithm.
The weak correlation of object selectivity properties of
the LFPs and spiking activity and augmented decoding
accuracy using both of them support the hypothesis that
the LFPs contain additional information about the cate-
gory of a visual stimulus.
MÉTHODES
Subjects and Recordings
The data came from 12 sessions in nine patients with
pharmacologically intractable epilepsy (all right-handed,
4 men, 17 à 47 ans). This set of patients overlaps
with the one used in a previous study (Quian Quiroga,
Reddy, Kreiman, Koch, & Frit, 2005), but the current
data corresponds to different experimental sessions.
For these patients, extensive noninvasive monitoring
did not yield concordant data corresponding to a single
resectable epileptogenic focus. Donc, they were
implanted with chronic depth electrodes for 7–10 days
to determine the seizure focus for possible surgical
résection (Frit, MacDonald, & Wilson, 1997). Ici
we report data from microelectrodes in the hippocam-
pus, amygdala, entorhinal cortex, and parahippocampal
gyrus. All studies conformed to the guidelines of the
Medical Institutional Review Board at UCLA. The elec-
trode locations were based exclusively on clinical criteria
and were verified by magnetic resonance imaging (IRM)
or by computed tomography (CT) coregistered to pre-
operative MRI. Each electrode probe had a total of nine
microwires at its end, eight active recording channels
and one reference. The differential signal
from the
microwires was amplified using a 64-channel Neuralynx
système (Tucson, AZ), filtered between 1 et 9000 Hz
and sampled at 28 kHz. Spike detection and clustering
was done using recordings band-pass filtered between
300 et 3000 Hz. The LFPs were obtained by band-pass
filtering the same recordings between 1 et 100 Hz and
down-sampling them to 256 Hz.
Each recording session lasted about 30 min. Sujets
were sitting in bed, facing a laptop computer, on which
landmarks, or objects
pictures of individuals, animals,
were shown. The images covered about 1.58, were cen-
tered on a laptop screen, and were displayed six times
each in pseudorandom order for 1 sec. Images were
photos of animals, landmarks, and celebrities, which were
partially chosen according to the patients’ preferences,
as well as photos of people and places unknown to the
patients. More details about the stimulus set are availa-
ble in Quian Quiroga et al. (2005). The interstimulus in-
terval (ISI) was randomized with the minimum ISI equal
à 1.5 sec. In order to encourage subjects to attend to
the picture presentations, they had to respond whether
the pictures contained a face or something else by
pressing the ‘‘Y’’ and ‘‘N’’ keys, respectivement.
Data Analysis
We analyzed the signals recorded from 568 microelec-
trodes implanted in different locations of the human
MTL. For the LFP data, we initially applied a digital notch
filter at 60 Hz and the first two harmonics (4th-order
elliptic filter, 0.1 dB peak-to-peak ripple, 40 dB stopband
attenuation). Recordings that showed either peaks at
harmonics of 60 Hz on the power spectrum or high-
frequency noise were discarded, thus obtaining a final
set of 451 ‘‘clean’’ microelectrodes for LFP analysis, 384
of which showed spiking activity.
We discarded from the analysis trials having more
que 5 points outside of the mean ± 5 standard devia-
tion range. The mean and the standard deviation were
calculated across all trials for each sample point.
Analysis of the LFP Amplitude Selectivity
To quantify category selectivity of the LFP amplitude, nous
applied a sample-by-sample one-way analysis of variance
(ANOVA) (Blair & Karniski, 1993; Guthrie & Buchwald,
1991) with the category identity as a main factor to the
480
Journal des neurosciences cognitives
Volume 19, Nombre 3
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
un
/
d
m
e
je
d
t
F
r
p
o
r
m
c
.
h
s
je
p
je
v
d
e
je
r
r
e
c
c
h
t
.
m
un
je
r
e
.
d
c
toi
o
m
o
/
c
j
n
o
un
c
r
t
n
je
c
/
e
un
–
r
p
t
d
je
c
1
je
9
e
3
–
4
p
7
d
9
F
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
oui
7
g
.
toi
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
F
e
m
b
b
oui
e
r
g
2
toi
0
e
2
s
3
t
/
j
.
.
.
.
.
F
t
o
n
1
8
M.
un
oui
2
0
2
1
LFP values. The sample-by-sample ANOVA test gives a
time-resolved significance level of how the LFP values
are different across categories. We considered a micro-
electrode to be selective to a category if at least 15 con-
secutive points ((cid:1)60 msec) of the ANOVA trace crossed
the significance threshold of .001. To check whether the
ANOVA traces crossed the .001 threshold by chance, nous
applied two control tests. D'abord, we applied the ANOVA
test to the one-second interval preceding stimulus on-
ensemble. Deuxièmement, we applied a bootstrap procedure by shuf-
fling the pictures in-between categories. Such shuffling
destroys information about the category but preserves
time correlations of LFPs and correlations between
different presentations of the same picture. Because
the bootstrap procedure is time consuming, we ap-
plied it only to the selective microelectrodes (one-way
ANOVA, p < .001, 15 consecutive points). The goal of
the bootstrap test was to confirm that the significant
p values obtained with the ANOVA test were reflecting
differences in averaged LFPs for semantic categorization
but not for other possible ones, for example, the cases
when there was only one very strong response to a
single picture.
Latencies
The latency of the selectivity for the averaged LFPs was
defined as the time point when the LFPs for three
categories started to be significantly different from each
other, that is, when the ANOVA trace first crossed the
significance level of p = .001 for at least 15 consecutive
points. Analogously, for the definition of the latency of
responsiveness, we applied a sample-by-sample t test
comparing the distribution of LFP values for each cate-
gory and each time point during stimulus presentation,
with the distribution of all LFP values during the baseline
interval. The moment when the t test trace first crossed
significance level of p = .001 for at least 15 consecutive
points for one category was defined as the latency of
the responsiveness.
Phase and Power Analysis
To estimate the instantaneous phase and power, we
used the continuous wavelet transform. The LFP of each
trial was convoluted with complex Morlet wavelets (cid:2)( f0,
t) = (s2p)(cid:2)1/4 exp((cid:2)t2/2s2) exp(2pif0t), where f0 is the
frequency and s specifies the width of the
central
wavelet in time domain. The Morlet wavelet is a complex
sine wave whose amplitude is tapered by a Gaussian
function. A wavelet family is characterized by a constant
nc = 2pf0s, which we set equal to 6. The convolution
with a complex Morlet wavelet gives a series of complex
wavelet coefficients W( f0, t) = R (cid:2)( f0, t (cid:2) t) s(t)dt =
A( f0, t) exp(if( f0, t)). From the wavelet coefficients cor-
responding to each frequency and time point, it is pos-
sible to define the instantaneous power as |W( f0, t)|2,
and the instantaneous phase as f( f0, t). To resem-
ble the main EEG frequency bands, we used 11 frequen-
cies spaced on the approximately logarithmical scale
between 2.5 and 85 Hz. Because the distribution of the
power was found to be significantly different from
Gaussian, we used a nonparametric sample-by-sample
ANOVA (Kruskal–Wallis test) to test for selectivity of the
LFP power. To test for mean-phase selectivity of the LFP,
we used a sample-by-sample analysis of a common mean
direction. This test is a generalization of a t test analog
for circular data to more than two variables.
Because, in general, the correlation between consec-
utive time points is higher for the lower frequencies,
both for the power and phase analyses, we required that
the significant difference between categories (with
p < .001) lasted for at least two periods (Rizzuto et al.,
2003). Due to the fact that a two-period interval may be
relatively short, especially for high frequencies (e.g., for
85 Hz it is only about 23 msec, which corresponds to
6 points if the sampling frequency is 256 Hz), the test for
selectivity ( p < .001, two periods) was validated with a
bootstrap procedure where the pictures were shuffled
in-between the categories.
Decoding
A trial-by-trial decoding was done with a linear Fisher
algorithm (Duda, Hart, & Stork, 2001). We employed a
one versus all strategy, that is, for each trial, a decision
about its category was made based on the distributions
of all other trials. The decoding accuracy was defined as
the relative number of correct predictions. The chance
level was equal to the inverse number of categories
(.33). Time profiles of decoding accuracies were calcu-
lated using the number of spikes or mean LFP values
in sliding time windows as an input to the decoding
algorithm. The sliding windows had 50% overlap. To
increase the number of inputs to the classifier, we de-
creased the significance threshold to .01 in comparison
to .001, which was used for selectivity analysis. We also
calculated the decoding accuracies with a very loose sig-
nificance threshold of .1, which did not change signifi-
cantly the results.
RESULTS
We studied the spiking activity and LFPs recorded from
the same microelectrodes implanted in the MTL of
human subjects with pharmacologically intractable epi-
lepsy. The placement of the electrodes was determined
exclusively by clinical criteria (Fried et al., 1997). In 12
experimental sessions with nine patients, we recorded
activity from 568 microelectrodes. Only ‘‘clean’’ record-
ings (451 microelectrodes) were used for further analy-
sis (Methods). The microelectrodes were located in the
Kraskov et al.
481
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
3
-
4
p
7
d
9
f
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
.
t
f
o
n
1
8
M
a
y
2
0
2
1
amygdala (120), hippocampus (182), entorhinal cortex
(102), and parahippocampal gyrus (47). Anatomical
locations of microelectrodes were selectively estimated
from the fused image of structural MRI taken before
implantation of the electrodes and CT taken while the
electrodes were implanted (Fried et al., 1999).
Selectivity of the Amplitude of Local Field
Potentials to Categories
All images were divided into three semantic categories:
faces, places, and animals. LFPs were time-aligned to the
stimulus onset. Intervals of 1 sec before and 1 sec after
stimulus onset were used in the analysis. Examples of
averaged LFPs, raster plots, and poststimulus time histo-
grams for three categories are shown in Figure 1. Here the
spiking activity was clearly responsive and selective to the
category ‘‘places,’’ increasing from its background rate of
about 0.2 Hz to approximately 2 Hz. Yet surprisingly, the
amplitude of the averaged LFPs for the category ‘‘places’’
was the smallest among the three categories. The aver-
aged amplitudes of LFPs for the two other categories
(‘‘animals’’ and ‘‘faces’’) were significantly different from
baseline. Another example is shown in Figure 2. Here the
spiking activity, as well as LFPs, for the category ‘‘faces’’
was significantly different from the responses to images
from the other two categories. The LFPs for the three
categories from Figure 2 are plotted in Figure 3A. The
corresponding ANOVA trace (Methods) is shown in Fig-
ure 3B. Note that after stimulus onset, there are two in-
tervals corresponding to positive and negative reflections
of the averaged LFPs from baseline where the ANOVA
trace is far above the chosen significance value of .001. To
verify the category selectivity of the LFPs, we applied two
control tests (Methods). Note that for the example in
Figure 3, there was no single point before stimulus onset
where the amplitude of the LFPs was selective. In fact,
none of the 451 microelectrodes showed selectivity during
the baseline interval. In total, we found that 36 of the 451
microelectrodes (8.0%) produced LFPs with a significant
category selectivity (one way ANOVA, p < .001, 15 con-
secutive points, bootstrap 1000 shuffles; Methods). Thirty
of these microelectrodes were located in the hippocam-
pus, three in the amygdala, three in the entorhinal cortex,
and none in the parahippocampal gyrus (Figure 4). We
did not find any significant difference ( p > .05, binomial
test) in the number of LFP(cid:2) amplitude-selective micro-
electrodes between different hemispheres (gauche 19/251,
droite 17/200). An equal number of selective microelec-
trodes were found in the epileptogenic temporal lobe and
in the contralateral lobe (18/270 versus 18/181).
The latency of the selectivity (c'est à dire., the moment when
the LFPs of the three categories started to be significantly
Chiffre 1. Spike and LFP
responses from a microelectrode
in the left medial hippocampus.
For each of the three image
categories shown, (UN) raster
plots, (B) poststimulus time
histogram, et (C) average
LFPs (thick) ± 1 SEM across
trials (thick, thin) are plotted.
Vertical dashed lines indicate
début (zero on time axis) et
offset (1 sec) of the image. Le
number of pictures in a category
is specified by the number in
the brackets following the name
of the category. Each picture
was shown six times. Le
baseline firing rate was about
0.2 Hz for all three categories.
Only during presentation of
‘‘place’’ pictures did the firing
activity increase to 2 Hz. Dans
contraste, the average LFPs for
‘‘places’’ had the smallest
amplitude, whereas the
maximum amplitude of the
average LFPs for the categories
‘‘animals’’ and ‘‘faces’’ was
à propos 50 AV. The baseline
amplitudes for all three
categories were about 15 AV.
482
Journal des neurosciences cognitives
Volume 19, Nombre 3
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
un
/
d
m
e
je
d
t
F
r
p
o
r
m
c
.
h
s
je
p
je
v
d
e
je
r
r
e
c
c
h
t
.
m
un
je
r
e
.
d
c
toi
o
m
o
/
c
j
n
o
un
c
r
t
n
je
c
/
e
un
–
r
p
t
d
je
c
1
je
9
e
3
–
4
p
7
d
9
F
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
oui
7
g
.
toi
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
F
e
m
b
b
oui
e
r
g
2
toi
0
e
2
s
3
t
/
j
t
F
.
.
.
.
.
o
n
1
8
M.
un
oui
2
0
2
1
Chiffre 2. Spike and
LFP responses from a
microelectrode in the left
anterior hippocampus. Le
baseline rate of this unit
était 10.2 Hz. It decreased
significantly ( p < .001, t test)
upon presentation of animal
pictures, remained unchanged
for ‘‘places,’’ and increased
significantly ( p < .001, t test)
for ‘‘faces.’’ The LFPs for
‘‘faces’’ were significantly
different from baseline ( p <
.001, ANOVA, see Figure 3).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
different from each other) was 460 ± 15 msec (mean ±
SEM). For the selective microelectrodes, we also calcu-
lated the latency of the LFP responsiveness (i.e., when
LFPs of a category started to be significantly different from
the baseline [Methods] and found it to be 369 ± 53 msec
[mean ± SEM), which is significantly shorter than the
latency of the selectivity ( p < .01, t test). This difference is
explained by the observation that, very often, the LFPs of
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
3
-
4
p
7
d
9
f
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
f
.
.
t
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Kraskov et al.
483
Figure 3. Time-resolved
selectivity analysis. Selectivity
of the LFP from the
microelectrode of Figure 2. (A)
Averaged LFPs (thick line) for
the three categories ± 1 SEM
(thin lines). (B) Normalized
p values ((cid:2)log10( p)) obtained
from the ANOVA test. The
dash–dotted line corresponds
to a significance of p = .001.
The significance values are in
logarithmic scale, significance
less than .001 corresponds to
the values on y-axis larger than
(cid:2)log10(.001) = 3. The ANOVA
test showed a significant
difference among categories
in two intervals of about
50 and 100 msec duration,
corresponding to positive
and negative reflections of
the averaged LFPs for the
category ‘‘faces.’’
Figure 4. Localization of
selective microelectrodes. (A)
Total number of category-
selective microelectrodes
across different brain
regions [hippocampus
(Hipp), amygdala (Am),
parahippocampal gyrus (PG),
entorhinal cortex (EC)]. (B)
The number of selective
microelectrodes normalized
to the number of electrodes
implanted in the area.
The vast majority of the
amplitude-selective
microelectrodes were found
in the hippocampus. Spike-
selective microelectrodes
were more evenly distributed
across these four regions.
all three categories first start to deviate from the baseline
and only later from each other (Figure S2).
Selectivity of the Mean Phase and Power
of the LFPs
In addition to the selectivity of the LFP amplitudes, we
studied whether their phase and power were category-
selective. The instantaneous phase and power of each
category were defined for every time point in the
interval [(cid:2)1:2] sec using a complex Morlet wavelet
transform (Grossmann, Kronland-Martinet, & Morlet,
1989), widely used in EEG analysis (TallonBaudry,
Bertrand, Delpuech, & Pernier, 1997) (Methods). We
applied a sample-by-sample, one-way, nonparametric
ANOVA to the values of the power in different frequency
bands. This gave a nonparametric ANOVA trace for
each microelectrode and frequency band. Similarly, to
quantify the category selectivity of the mean phases,
we applied a sample-by-sample test for a common mean
direction (Fisher, 1995) (Methods). In Figure 5, an ex-
ample of a power-selective microelectrode is presented.
The upper panel shows the averaged log-transformed
power in the g band (the central frequency of the
Morlet wavelet was 45 Hz) for the three categories.
Around 320 msec, the power of the category ‘‘faces’’
starts to be clearly different from the baseline power
and from the power in the other two categories, the
latter being reflected in the ANOVA trace plot (lower
panel). An example of a mean phase-selective micro-
electrode is presented in Figure 6. The averaged LFPs
filtered in u band (the central frequency of the Morlet
wavelet was 6 Hz) for the three categories are shown in
Figure 6A. Figure 6B shows the significance of the
sample-by-sample test for a common mean direction.
The test reached its maximum significance around
300 msec. The phase distributions and their mean di-
rection at this particular time are shown in Figure 6C.
The length of the mean direction vector is proportional
to the difference between the phase distribution for a
given category from the uniform distribution (Rayleigh
test). Figure 7A plots the total number of the micro-
electrodes showing selectivity to a category with the
power and Figure 7B with the mean phase across differ-
ent frequency bands. The percentage of microelectrodes
that showed selectivity with power (24; 5.3%) or mean
phase (27; 6.0%) was relatively small. Comparing the data
presented in Figure 7A and B, one can see a higher per-
centage of mean phase-selective LFPs for the lower fre-
quencies and an opposite trend for the power-selective
LFPs, namely, higher percentage of power-selective LFPs
for higher frequencies (30–100 Hz). It leads to the
intriguing hypothesis that there are two possible differ-
ent mechanisms for the selectivity of LFPs. One involves
phase locking in the lower frequencies and another one
engages power increase in higher frequencies.
Half (13 microelectrodes) of the mean phase-selective
microelectrodes were also selective for the amplitude of
the LFPs (dark blue bars in Figure 7B), whereas only four
microelectrodes showed selectivity for both power and the
amplitude of the LFPs (dark blue bars in Figure 7A). This is
not very surprising because the activity phase-locked to
the stimulus onset is mostly preserved in the averaged
LFPs, whereas the induced, nonstimulus-locked activity is
averaged out and is revealed only in the averaged power.
484
Journal of Cognitive Neuroscience
Volume 19, Number 3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
3
-
4
p
7
d
9
f
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
f
.
.
.
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 5. Example of a
microelectrode with selective
LFP power. (A) Averaged
log-transformed power in the g
band with a central frequency
of the complex Morlet wavelet
at 45 Hz (thick line) for
three categories ± 1 SEM
(thin lines). (B) Normalized
p values obtained from ANOVA
test. The dash–dotted line
corresponds to the significance
level p = .001. The averaged
instantaneous power is larger
for the category ‘‘faces’’
than the power for the other
two image categories for
about 100 msec starting at
(cid:1)300 msec.
Selectivity of Spiking Activity to Categories
The spiking activity recorded with the same microelec-
trodes used to record the LFPs was preprocessed using a
novel spike sorting algorithm (Quian Quiroga, Nadasdy,
& Ben-Shaul, 2004). To quantify the category selectivity
of the spiking activity, we applied an one-way ANOVA
with the category identity as a main factor, and the
number of spikes in the interval [300:1000] msec (Quian
Quiroga et al., 2005) as repeated measures. A t test
comparison with the baseline interval [(cid:2)1000:(cid:2)300]
msec was used as post hoc test to define the responsive
category. Additionally, we performed a bootstrap test.
The pictures were randomly shuffled between catego-
ries and an ANOVA test was applied. We found that 66
out of the 591 recorded units (11.2%) had a spiking
response with significant category selectivity (ANOVA,
p < .001, t test, p < .001, bootstrap 1000). Fourteen
units showed a significant decrease in firing rate, and
eight units showed a significant increase to one category
and a significant decrease in firing rate to another one.
Units selective to at least one of the three semantic
categories were recorded from 56 of the 384 micro-
electrodes (14.6%) used in the analysis. These numbers
are comparable with those reported in Kreiman et al.
(2000). As a control, the baseline and stimulus presen-
tation intervals were exchanged and the same analysis
was repeated. In this case, only one significant response
was found. The distribution of the spike-selective micro-
electrodes across different brain regions was more uni-
form in comparison to that of the LFPs. We found 18
selective microelectrodes in hippocampus, 25 in the
amygdala, 8 in entorhinal cortex, and 5 in parahippo-
campal gyrus (Figure 4).
To compare the latency of the selective spiking and
LFP activities, we convolved each spike train with a
Gaussian kernel (100 msec width at half height) and
repeated the same analysis used for the selectivity of the
LFPs amplitudes (sample-by-sample one-way ANOVA,
p < .001, 15 consecutive points). The latency of spike-
selectivity was defined as the first time point when there
was a significant difference between the categories. The
spike-responsiveness latency was found equivalent to the
latency of selectivity. The average value for the latency of
selectivity was found to be 341 msec (14 msec SEM). It
was significantly ( p < .001, t test) earlier then the latency
of selectivity of the LFPs. Yet, the latency of the spiking
responses was found to be not significantly different from
the latency of the LFP responses (t > 0.3, t test).
For the spiking activity, we found 24/315 category selec-
tive units in the left hemisphere and 42/276 in the right
hemisphere, et 20/343 category selective units in the
epileptogenic hemisphere and 46/248 on the contralat-
eral one. We found more category-selective units in the
contralateral side. Because we do not have extensive pa-
tient statistics (9 patients), we cannot make any conclusive
claims about lateralization of the category selectivity effect.
In total, we found 85 microelectrodes which pro-
duced either selective LFPs or selective spiking re-
sponses and which passed the bootstrap test which
Kraskov et al.
485
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
un
/
d
m
e
je
d
t
F
r
p
o
r
m
c
.
h
s
je
p
je
v
d
e
je
r
r
e
c
c
h
t
.
m
un
je
r
e
.
d
c
toi
o
m
o
/
c
j
n
o
un
c
r
t
n
je
c
/
e
un
–
r
p
t
d
je
c
1
je
9
e
3
–
4
p
7
d
9
F
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
oui
7
g
.
toi
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
F
e
m
b
b
oui
e
r
g
2
toi
0
e
2
s
3
t
/
j
F
.
t
.
.
.
.
o
n
1
8
M.
un
oui
2
0
2
1
Chiffre 6. Example of a
microelectrode with selective
LFP mean phase. (UN) Averaged
LFPs filtered in the u band with
the central frequency of the
complex Morlet wavelet at 6 Hz
(thick line) for three categories
± 1 SEM (thin lines). (B)
Normalized p values retrieved
from a test for a common
mean direction. The black
vertical line indicates the
time point at which the
mean phases were most
different. The dash–dotted
line corresponds to the
significance level p = .001.
(C) Phase distributions across
trials taken at the moment of
the largest difference among
catégories, as shown in (UN)
et (B) with the solid vertical
line. Colored vectors show
the mean phase direction.
The length of the vector
is proportional to the
significance of the difference
of the phase distribution for a
given category from a uniform
circular distribution (Rayleigh
test). p values are given next
to the category names. Ici,
the phase distribution of the
category ‘‘faces’’ significantly
differs from a uniform circular
distribution ( p < 10(cid:2)7).
shuffles pictures between categories. However, only
seven of them were selective for both the LFPs and
the spiking activity. Six microelectrodes showed mean
phase and spiking selectivity and 10 spike-selective
microelectrodes showed power selectivity. The distribu-
tion of these channels across the different frequency
bands is indicated with red bars in Figure 7.
Decoding with LFPs and Spiking Activity
We applied a linear decoding algorithm to the LFPs and
the spiking activity recorded simultaneously from many
microelectrodes in the MTL in order to ascertain how
much information pertaining to the semantic category of
the images can be inferred readout from the neural data.
Here we use the term ‘‘decoding’’ in the computational
sense, namely, we studied how reliably one can predict
in each single trial the category identity of the stimulus
given the firing of the neurons or the LFP activity. We
studied the time profile of the decoding accuracy
(Methods) in the time interval [(cid:2)1:2] sec. The inputs
to the decoding algorithm were the number of spikes
for each category or the mean value of the amplitude of
the LFP defined in sliding windows of different sizes with
50% overlap. Only the activity of amplitude-selective
microelectrodes (one-way ANOVA, p < .01, 15 consec-
utive points) was taken as an input.
We found that the time profile of the decoding ac-
curacy using spiking data increased with the length of
the moving window (Figure 8B) and saturated for
windows longer then 200 msec. For LFPs, the time pro-
file remained approximately at the same level (Figure 8A)
for different window sizes. Both for spiking and LFP
data, and for all durations of the moving window,
decoding accuracy during the baseline interval did not
differ from chance (t test, p > .05). LFPs slightly out-
performed the spiking activity only for very small win-
dow sizes of 10 et 20 msec. De plus, for 10-msec
windows, the classifier could barely distinguish between
the categories using only spikes. This can be explained
486
Journal des neurosciences cognitives
Volume 19, Nombre 3
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
un
/
d
m
e
je
d
t
F
r
p
o
r
m
c
.
h
s
je
p
je
v
d
e
je
r
r
e
c
c
h
t
.
m
un
je
r
e
.
d
c
toi
o
m
o
/
c
j
n
o
un
c
r
t
n
je
c
/
e
un
–
r
p
t
d
je
c
1
je
9
e
3
–
4
p
7
d
9
F
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
oui
7
g
.
toi
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
F
e
m
b
b
oui
e
r
g
2
toi
0
e
2
s
3
t
/
j
F
.
.
.
.
.
t
o
n
1
8
M.
un
oui
2
0
2
1
Chiffre 7. Number of
microelectrodes with showing
selectivity with LFP power
and mean phase. Le
distribution of the (UN) pouvoir
et (B) mean phase-selective
microelectrodes across
different frequency bands
of the LFPs. A total of
20 microelectrodes out
de 475 showed category
selectivity using the
power analysis and 29
microelectrodes showed
category selectivity using mean
phase analysis. En principe,
the same microelectrodes
could be selective in different
frequency bands. For the
total number of selective
microelectrodes, we count
such electrodes only once.
A large overlap is observed
in selective microelectrodes
according to LFP amplitude
analysis and the mean phase
analyse (15 microelectrodes).
by the typically low firing rates of these neurons, con-
sidering that they may not produce a single spike during
a 10-msec window. Combining the LFP and spiking
activities increased the decoding accuracy for all window
sizes between 10 et 200 msec (Figure 8C). The traces
in Figure 8D show the decoding accuracies obtained
with the LFPs, with the spikes and with both the LFPs
and spikes for a window length of 20 msec. The red
crosses indicate time points when decoding using the
combination of the LFPs and spikes was significantly
better (paired t test, p < .01) than the decoding
accuracy using only the amplitude of the LFPs, whereas
the blue circles show the comparison between accuracy
of the combined spikes and LFPs signal with the one
using only spikes. For longer windows, decoding accu-
racy with combined LFPs and spikes input was also
slightly better than accuracy obtained with each signal
alone. It is notable that the time when the decoding
accuracy became significantly different from chance was
very similar for the LFPs and the spiking activity, around
300 msec after stimulus onset. The fact that we can infer
category identity by applying a classifier to LFPs does not
imply that the brain makes use of this information, but
only that this information is present in the MTL and
could be used by postsynaptic processes.
DISCUSSION
In previous studies, the spiking activity of single neurons
in the human MTL was found to be selective to different
categories of visual stimuli (Kreiman et al., 2000) and even
invariant to different views of the same person or object
(Quian Quiroga et al., 2005). The analysis of the LFPs
recorded from the surface of the brain revealed face-
selective LFPs in the extrastriate cortex (Allison, Ginter,
et al., 1994; Allison, McCarthy, Nobre, Puce, & Belger,
1994). Later studies using intracranial depth electrodes lo-
calized the source of the face-selective LFPs near the fusi-
form gyrus (Lachaux et al., 2005; Allison, Puce, Spencer,
& McCarthy, 1999), in good agreement with fMRI find-
ings (Kanwisher, McDermott, & Chun, 1997). In this arti-
cle, we undertook a combined study of the LFPs and
spiking activity recorded by the same microelectrodes
to assess their object- and face-selectivity properties. We
found that the spiking activity recorded from 56 mi-
croelectrodes was selective to semantic categories. The
amplitude of the LFPs from a smaller number of micro-
electrodes (36) also showed category selectivity, but not
necessarily to the category ‘‘faces.’’
We also found that the power in the g band was dis-
criminative between categories but only in a small number
of microelectrodes. This finding is similar to one reported
by Oya, Kawasaki, Howard, and Adolphs (2002), who
showed selectivity of the LFP g power in the amygdala
in response to emotional faces. Although we did not find
any power selectivity in lower frequency bands for a small
number of microelectrodes, the mean phases of three
categories were different in the d (1–4 Hz) and u (4–8 Hz)
bands, but not in the g band (higher than 30 Hz following
the definition in Engel, Fries, & Singer, 2001). This
suggests two possible mechanisms for selectivity of LFPs:
via phase-locking in the lower frequency bands and/or via
power increase at high frequencies. Because the power of
d and u oscillations is significantly larger than the power
of g oscillations (see the average LFP power spectra Fig-
ure S2), it is more efficient (i.e., less energy consuming
Kraskov et al.
487
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
3
-
4
p
7
d
9
f
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
f
t
.
o
n
1
8
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
3
-
4
p
7
d
9
f
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
f
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 8. Time profile of decoding accuracy, using the spiking activity, the LFPs, and both signals together. Accuracy varies between perfect
(1) and chance level (1/3). Decoding accuracy using (A) the average value of LFPs amplitude, (B) firing rates (SPK), and (C) their combination
(SPK/LFP). The different curves on the (A,B,C) subplots correspond to window sizes of 10, 20, 50, 100, and 200 msec. Decoding accuracy with LFPs
is nearly independent of the window size in this range, whereas accuracy with spikes (with or without LFP) increased with the window size. All
curves were smoothed with a 5-point moving average. Dash–dotted lines show the confidence intervals obtained for the time points from a t test
comparison with chance level (1/3, black line), p < .01. Decoding with LFPs, spikes, and their combination is shown in (D) for a window of size
20 msec. Optimal accuracy is achieved with the combination of LFPs and spiking activity. Crosses (circles) indicate time points where decoding
for combination of the LFPs and spiking activity was significantly better than decoding using only LFPs (only spikes) (paired t test, p < .01).
and faster) to transmit information by modulation of g
power. On the other hand, g oscillations are much faster
than d or u ones, therefore, a small jitter in g oscillations
will destroy their synchronization but would hardly influ-
ence slow d and u oscillations. Previous studies with rats
performing spatial tasks also found a phase locking of
spikes with the ongoing activity in the u band (Siapas,
Lubenov, & Wilson, 2005). Synchronization of g oscilla-
tions was suggested as possible mechanism for infor-
mation processing (Singer, 1999). The vast majority of
the LFP amplitude-selective microelectrodes were found
in the hippocampus (30 out of 36). The relative number
of hippocampal-selective microelectrodes (16%) was three
times larger than in all other areas (Figure 4). At the same
time, the relative number of spike-selective electrodes was
evenly distributed across all four investigated areas. This
discrepancy suggests that the category-selective LFPs are
either the result of local processing within the hippocam-
pus or that the hippocampus receives a category-specific
input from adjacent areas.
Only a small overlap was observed between spikes
and LFP selective microelectrodes. This lack of correla-
tion supports the view that neurons in the MTL are only
weakly spatially clustered in terms of the semantic
categories (faces, places, animals). This result is in line
with the weak correlation between the object-selective
LFPs and object-selective spiking activity recently re-
ported in monkey IT (Kreiman et al., 2006). It also
suggests that spiking activity and LFPs contain different
information about stimulus category.
Halgren et al. (1980) recorded LFPs in the human MTL
during an ‘‘oddball’’ paradigm and found a P300 evoked
potential well known from surface EEG measurements.
Some studies argue for a generation of the P300 in the hip-
pocampal formation and amygdala (Halgren, Marinkovic,
& Chauvel, 1998; McCarthy, Wood, Williamson, & Spencer,
488
Journal of Cognitive Neuroscience
Volume 19, Number 3
(Logothetis, 2003; Mitzdorf, 1985). Therefore, the ob-
served selectivity of LFPs in the MTL might be caused by
specific pattern of dendritic activity arising, for example,
from the prefrontal cortex, which has been reported to
be involved in categorical representation of visual stimuli
(Freedman, Riesenhuber, Poggio, & Miller, 2001) or by lo-
cal synaptic circuitry, which is differentially activated for
different categories of visual stimuli. It is not possible to
distinguish among these possibilities with the current data.
The weak correlation in the selectivity properties of
LFPs and spikes suggests that they reflect two different
aspects of brain activity. A similar disassociation between
LFPs and spikes was also observed in macaque V1
(Henrie & Shapley, 2005). These authors hypothesized
that the network activity captured by LFPs originates
from inhibitory interneurons, whereas single-unit activ-
ity is largely biased toward pyramidal neurons.
This relative independence observed in the selectivity
of the spiking and LFPs activities is compatible with our
decoding analysis. Reading out both the mean number of
spikes as well as the mean amplitude of the LFP allowed
us to infer the identity of the category of the visual stimu-
lus significantly better than using either measure by itself.
This effect was more pronounced when information
from short time windows was used for classification.
SUPPLEMENTARY MATERIAL
Signal-to-noise Ratio in Different Brain Areas
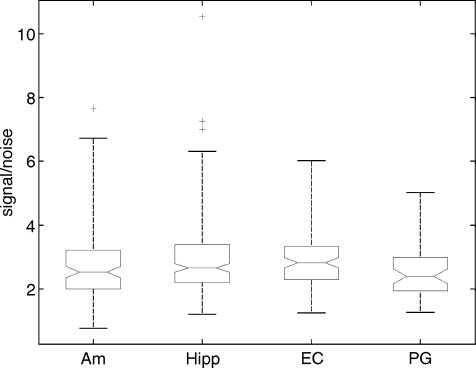
Figure S1. Signal-to-noise ration indifferent brain areas. Boxplot of
the signal-to-noise ratios in different brain areas. Horizontal middle
lines represent medians of distributions of signal-to-noise ratios.
1989). To check whether the evoked potentials which we
observed were task-dependent, we repeated our experi-
ment in one patient without a task (i.e., the patient
was asked to passively look at the pictures presented on
the screen for 500 msec). We found two (out of 24 micro-
electrodes analyzed for this session) LFP amplitude-
selective microelectrodes (Figure S4). This corresponds
to the 8% of LFP-selective microelectrodes found with task
and argues in favor of task independence.
The LFPs represent the average dendrosomatic activ-
ity of presynaptic signals of large neuronal populations
One possible explanation for the fact that 30 out of 36
LFP-selective microelectrodes were found in the hippo-
Figure S2. Average LFPs
power spectrum. Magenta
(blue) solid line shows a
power spectrum averaged
across stimulus presentation
[0:1] sec (baseline [(cid:2)1:0] sec)
interval for all analyzed LFPs.
Dash–dotted lines depict
least-squares logarithmic fit,
f (cid:2)a. For stimulus presentation
interval average a was found
to be equal to 2.01 ± (cid:2)0.02,
(mean ± SEM) and for
the baseline interval to be
2.04 ± 0.02, (mean ± SEM).
Note that the magenta and
blue curves practically almost
overlap. Drop at 60 Hz is
due to the digital notch
filter applied at 60 Hz.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
3
-
4
p
7
d
9
f
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
f
.
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Kraskov et al.
489
Figure S3. Latency of
selectivity versus latency of
responsiveness. (A) Averaged
LFPs (thick line) for the three
categories ± 1 SEM (thin
lines) corresponding to an
electrode implanted in the
right amygdala. (B) Normalized
p values obtained with the
ANOVA test of selectivity (black
thick line) and the t tests or
responsiveness for each
category (color lines). A
sample-by-sample t test was
applied to the distribution
of LFPs values for each category
and each time point during
stimulus presentation with
the distribution of all LFPs
values during the baseline
interval. The dash–dotted line
corresponds to a significance
of .001. The ANOVA trace first
crosses the significance level
of p = .001 at (cid:1)300 msec,
whereas the t test reached
significance earlier at
(cid:1)230 msec.
campus could be that given its anatomical structure, the
signal-to-noise ratio in the hippocampus is better than in
the amygdala. We statistically compared signal-to-noise
ratios for evoked potentials in different brain areas (see
Figure S1). Signal-to-noise ratio was estimated for each
microelectrode as the ratio of the maximum amplitude
of the evoked potential to the standard error of the
mean. Mean values of signal-to-noise ratios in the hippo-
campus and entorhinal cortices were found to be higher
than in the amygdala and parahippocampal gyrus. But
Figure S4. Time-resolved
selectivity analysis. Selectivity
analysis of LFPs recorded
during a passive viewing task.
Each stimulus was presented
for 500 msec. (A) Averaged
LFPs (thick line) for the three
categories ± 1 SEM (thin
lines). (B) Normalized p values
obtained with the ANOVA
test. The dash–dotted line
corresponds to a significance
of p = .001. All three
categories show significant
response but there is a clear
difference between the
category ‘‘faces’’ and two
other categories.
490
Journal of Cognitive Neuroscience
Volume 19, Number 3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
3
-
4
p
7
d
9
f
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
f
.
.
.
.
t
.
o
n
1
8
M
a
y
2
0
2
1
the difference was not statistically significant (t test, p >
.05 for hippocampus vs. amygdala, and hippocampus vs.
entorhinal cortex), and was only slightly significant for
the hippocampus versus parahippocampal gyrus (.01 <
p < .05). Therefore, the difference between signal-to-
noise ratios in different brain areas cannot explain the
larger selectivity of LFPs found in the hippocampus than
in any other area.
Numbers of Responsive LFPs and Units to
Different Categories
For the spiking activity a t test comparison between the
firing rate during the poststimulus [300:1000] msec and
baseline [1000:(cid:2)300] msec intervals was used to define
the responsive category. For LFPs we applied a sample-
by-sample t test comparing the distribution of LFP values
for each category and each time point during stimulus
presentation, with the distribution of all LFP values
during the baseline interval. The LFPs were defined to
be responsive to a category if the t test trace crossed a
significance level of p = .001 for at least 15 consecutive
points (see Figure S3). The following tables provide
information about the relative number of responsive
units and LFPs across different categories as well as
different brain areas.
LFP
All
AM
Hipp
EC
PG
Spikes
All
AM
Hipp
EC
PG
Animals
Faces
Places
24 (67%)
35 (97%)
14 (39%)
3
21
0
0
2
30
3
0
1
13
0
0
32 (48%)
31 (47%)
26 (39%)
17
10
1
4
15
7
7
2
12
8
1
5
Note that the same unit might show a significant
increase in firing rate for one category and a significant
decrease for another one (see Figure 2). For the LFP, we
use a time-resolved measure, therefore, the LFP from
the same microelectrode can respond to several catego-
ries but can be different at different time points. For
example, the LFPs in Figure S3 are responsive for all
three image categories. Yet at the same time, an ANOVA
test shows that the LFP differs among the three image
in this
categories at somewhat different times. Thus,
example, the LFP can discriminate among the three
categories but at different times.
Acknowledgments
We thank all patients for their participation and Eric Behnke,
Tony Fields, Emily Ho, Eve Isham, and Charles Wilson for
technical assistance. We also would like to thank Gabriel
Kreiman and Daniel Rizzuto for helpful discussions. This work
was supported by grants from the NINDS, NIMH, NSF, DARPA,
EPSRC, the Office of Naval Research, the W.M. Keck Founda-
tion Fund for Discovery in Basic Medical Research, the Gordon
Moore Foundation, the Sloan Foundation, and the Swartz
Foundation for Computational Neuroscience.
Reprint requests should be sent to Dr. Alexander Kraskov,
Sobell Department of Motor Neuroscience, UCL Institute of
Neurology, Queen Square, London WC1N 3BG, UK, or via
e-mail: a.kraskov@ion.ucl.ac.uk.
REFERENCES
Allison, T., Ginter, H., McCarthy, G., Nobre, A. C., Puce, A.,
Luby, M., et al. (1994). Face recognition in human
extrastriate cortex. Journal of Neurophysiology, 71, 821–825.
Allison, T. T., McCarthy, G. G., Nobre, A. A., Puce, A. A., &
Belger, A. A. (1994). Human extrastriate visual cortex and
the perception of faces, words, numbers, and colors.
Cerebral Cortex, 4, 544.
Allison, T. T., Puce, A. A., Spencer, D. D. D., & McCarthy,
G. G. (1999). Electrophysiological studies of human face
perception: I. Potentials generated in occipitotemporal
cortex by face and non-face stimuli. Cerebral Cortex,
9, 415.
Bechtereva, N. P., & Abdullaev, Y. G. (2000). Depth electrodes
in clinical neurophysiology: Neuronal activity and human
cognitive function. International Journal of
Psychophysiology, 37, 11–29.
Blair, R. C., & Karniski, W. (1993). An alternative method
for significance testing of wave-form difference potentials.
Psychophysiology, 30, 518–524.
Buchwald, J. S. (1965). Comparison of multiple-unit and
electroencephalogram activity recorded from same brain
sites during behavioural conditioning. Nature, 205, 1012.
Duda, O. H., Hart, P. E., & Stork, D. G. (2001). Pattern
classification (2nd ed.). New York: Wiley.
Engel, A. K., Fries, P., & Singer, W. (2001). Dynamic
predictions: Oscillations and synchrony in top-down
processing. Nature Reviews Neuroscience, 2, 704–716.
Engel, A. K., Moll, C. K. E., Fried, I., & Ojemann, G. A.
(2005). Invasive recordings from the human brain: Clinical
insights and beyond. Nature Reviews Neuroscience, 6,
35–47.
Fisher, N. I. (1995). Statistical analysis of circular data.
Cambridge: Cambridge University Press.
Freedman, D. J., Riesenhuber, M., Poggio, T., & Miller, E. K.
(2001). Categorical representation of visual stimuli in the
primate prefrontal cortex. Science, 291, 312–316.
Fried, I., MacDonald, K. A., & Wilson, C. L. (1997). Single
neuron activity in human hippocampus and amygdala
during recognition of faces and objects. Neuron, 18,
753–765.
Fried, I., Wilson, C. L., Maidment, N. T., Engel, J., Behnke, E.,
Fields, T. A., et al. (1999). Cerebral microdialysis combined
Kraskov et al.
491
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
3
-
4
p
7
d
9
f
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
t
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
with single-neuron and electroencephalographic recording
in neurosurgical patients [Technical note]. Journal of
Neurosurgery, 91, 697–705.
Fromm, G. H. (1967). Relationship between neuron activity
and cortical steady potentials. Electroencephalography
and Clinical Neurophysiology, 22, 159.
Grossmann, A., Kronland-Martinet, R., & Morlet, J. (1989).
Reading and understanding continuous wavelet transforms.
In J. Combes, A. Grossmann, & P. Tchamitchian (Eds.),
Wavelets: Time–frequency methods and phase space.
Berlin: Springer.
Guthrie, D., & Buchwald, J. S. (1991). Significance testing
of difference potentials. Psychophysiology, 28, 240–244.
Haberly, L. B., & Shepherd, G. M. (1973). Current–density
analysis of summed evoked potentials in opossum
prepyriform cortex. Journal of Neurophysiology, 36,
789–802.
Halgren, E., Marinkovic, K., & Chauvel, P. (1998). Generators
of the late cognitive potentials in auditory and visual
oddball tasks. Electroencephalography and Clinical
Neurophysiology, 106, 156–164.
Halgren, E., Squires, N. K., Wilson, C. L., Rohrbaugh, J. W.,
Babb, T. L., & Crandall, P. H. (1980). Endogenous potentials
generated in the human hippocampal-formation and
amygdala by infrequent events. Science, 210, 803–805.
Henrie, J. A., & Shapley, R. (2005). LFP power spectra in V1
cortex: The graded effect of stimulus contrast. Journal of
Neurophysiology, 94, 479–490.
John, E. R. (1967). Effects of visual form on evoked response.
Science, 155, 1439.
Logothetis, N. K. (2003). The underpinnings of the BOLD
functional magnetic resonance imaging signal. Journal
of Neuroscience, 23, 3963–3971.
McCarthy, G., Wood, C. C., Williamson, P. D., & Spencer,
D. D. (1989). Task-dependent field potentials in human
hippocampal-formation. Journal of Neuroscience, 9,
4253–4268.
Mehring, C., Rickert, J., Vaadia, E., de Oliveira, S. C., Aertsen, A.,
& Rotter, S. (2003). Inference of hand movements from
local field potentials in monkey motor cortex. Nature
Neuroscience, 6, 1253–1254.
Mitzdorf, U. (1985). Current source–density method and
application in cat cerebral cortex: Investigation of evoked
potentials and EEG phenomena. Physiological Reviews,
65, 37–100.
Oya, H., Kawasaki, H., Howard, M. A., & Adolphs, R. (2002).
Electrophysiological responses in the human amygdala
discriminate emotion categories of complex visual stimuli.
Journal of Neuroscience, 22, 9502–9512.
Pesaran, B., Pezaris, J. S., Sahani, M., Mitra, P. P., & Andersen,
R. A. (2002). Temporal structure in neuronal activity
during working memory in macaque parietal cortex.
Nature Neuroscience, 5, 805–811.
Quian Quiroga, R., Nadasdy, Z., & Ben-Shaul, Y. (2004).
Unsupervised spike detection and sorting with wavelets
and superparamagnetic clustering. Neural Computation,
16, 1661–1687.
Quian Quiroga, R., Reddy, L., Kreiman, G., Koch, C., &
Fried, I. (2005). Invariant visual representation by single
neurons in the human brain. Nature, 435, 1102–1107.
John, E. R. (1972). Switchboard versus statistical theories of
Rizzuto, D. S., Madsen, J. R., Bromfield, E. B., Schulze-Bonhage,
learning and memory. Science, 177, 850.
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). The
fusiform face area: A module in human extrastriate cortex
specialized for face perception. Journal of Neuroscience,
17, 4302–4311.
Kreiman, G., Hung, C. P., Kraskov, A., Quian Quiroga, R.,
Poggio, T., & DiCarlo, J. (2006). Object selectivity of local
field potentials and spikes in the macaque inferior temporal
cortex. Neuron, 49, 433–445.
Kreiman, G., Koch, C., & Fried, I. (2000). Category-specific
visual responses of single neurons in the human medial
temporal lobe. Nature Neuroscience, 3, 946–953.
Lachaux, J. P., George, N., Tallon-Baudry, C., Martinerie, J.,
Hugueville, L., Minotti, L., et al. (2005). The many faces
of the gamma band response to complex visual stimuli.
Neuroimage, 25, 491–501.
Lachaux, J. P., Rudrauf, D., & Kahane, P. (2003). Intracranial
EEG and human brain mapping. Journal of Physiology—
Paris, 97, 613–628.
Liu, J., & Newsome, W. T. (2006). Local field potential in
cortical area MT: Stimulus tuning and behavioral
correlations. Journal of Neuroscience, 26, 7779–7790.
A., Seelig, D., Aschenbrenner-Scheibe, R., et al. (2003).
Reset of human neocortical oscillations during a working
memory task. Proceedings of the National Academy of
Sciences, U.S.A., 100, 7931–7936.
Scherberger, H., Jarvis, M. R., & Andersen, R. A. (2005).
Cortical local field potential encodes movement intentions
in the posterior parietal cortex. Neuron, 46, 347–354.
Siapas, A. G., Lubenov, E. V., & Wilson, M. A. (2005). Prefrontal
phase locking to hippocampal theta oscillations. Neuron,
46, 141–151.
Singer, W. (1999). Neuronal synchrony: A versatile code for
the definition of relations? Neuron, 24, 49–65.
TallonBaudry, C., Bertrand, O., Delpuech, C., & Pernier, J.
(1997). Oscillatory gamma-band (30–70 Hz) activity
induced by a visual search task in humans. Journal of
Neuroscience, 17, 722–734.
Verzeano, M., Crandall, P. H., & Dymond, A. (1971). Neuronal
activity of amygdala in patients with psychomotor
epilepsy. Neuropsychologia, 9, 331–344.
Wyler, A. R., Ojemann, G. A., & Ward, A. A. (1982). Neurons
in human epileptic cortex—Correlation between unit and
EEG activity. Annals of Neurology, 11, 301–308.
492
Journal of Cognitive Neuroscience
Volume 19, Number 3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
3
-
4
p
7
d
9
f
/
1
1
9
9
3
6
/
2
3
1
/
7
4
7
o
9
c
/
n
1
2
7
0
5
0
6
7
6
1
0
9
4
/
3
j
4
o
7
c
9
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
3
o
.
n
4
0
7
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
t
f
.
o
n
1
8
M
a
y
2
0
2
1