FONCTION DE MISE AU POINT:
Connectivity, Cognition, and Consciousness
Functional network antagonism
and consciousness
Athena Demertzi1,2,3,4
, Aaron Kucyi5
, Adrián Ponce-Alvarez6
, Georgios A. Keliris7
,
Susan Whitfield-Gabrieli5,8*, and Gustavo Deco6,9,10,11*
1Physiology of Cognition, GIGA Consciousness Research Unit, GIGA Institute (B34), Sart Tilman,
University of Liège, Liège, Belgium
2Psychology and Neuroscience of Cognition (PsyNCog), Faculty of Psychology, Logopedics and
Educational Sciences, Sart Tilman, University of Liège, Liège, Belgium
3GIGA-CRC In Vivo Imaging, Sart Tilman, University of Liège, Liège, Belgium
4Fund for Scientific Research, FNRS, Bruxelles, Belgium
5Département de psychologie, Northeastern University, Boston, MA, Etats-Unis
6Center for Brain and Cognition, Computational Neuroscience Group, Department of Information and
Communication Technologies, Universitat Pompeu Fabra, Barcelona, Espagne
7Bio-Imaging Lab, Department of Biomedical Sciences, University of Antwerp, Wilrijk, Belgium
8Northeastern University Biomedical Imaging Center (NUBIC), Northeastern University
Interdisciplinary Science and Engineering Complex (ISEC), Boston, MA, Etats-Unis
9Institució Catalana de la Recerca i Estudis Avançats (ICREA), Barcelona, Espagne
10Department of Neuropsychology, Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Allemagne
11Turner Institute for Brain and Mental Health, School of Psychological Sciences, and Monash Biomedical Imaging,
Monash University, Clayton, Melbourne, VIC, Australia
*Equal contribution.
Mots clés: Anticorrelations, L'intégration, Segregation, Consciousness, Neural inhibition, Functional
connectivité
ABSTRAIT
Spontaneous brain activity changes across states of consciousness. A particular consciousness-
mediated configuration is the anticorrelations between the default mode network and other
brain regions. What this antagonistic organization implies about consciousness to date
remains inconclusive. In this Perspective Article, we propose that anticorrelations are the
physiological expression of the concept of segregation, namely the brain’s capacity to show
selectivity in the way areas will be functionally connected. We postulate that this effect is
mediated by the process of neural inhibition, by regulating global and local inhibitory activity.
While recognizing that this effect can also result from other mechanisms, neural inhibition
helps the understanding of how network metastability is affected after disrupting local and
global neural balance. In combination with relevant theories of consciousness, we suggest
that anticorrelations are a physiological prior that can work as a marker of preserved
consciousness. We predict that if the brain is not in a state to host anticorrelations, then most
likely the individual does not entertain subjective experience. We believe that this link
between anticorrelations and the underlying physiology will help not only to comprehend
how consciousness happens, but also conceptualize effective interventions for treating
consciousness disorders in which anticorrelations seem particularly affected.
RÉSUMÉ DE L'AUTEUR
The fMRI resting paradigm can quantify brain function by surpassing communication and
sophisticated setups, hence helping to infer consciousness in individuals who are unable to
un accès ouvert
journal
Citation: Demertzi, UN., Kucyi, UN., Ponce-
Álvarez, UN., Keliris, G. UN., Whitfield-
Gabriela, S., & Deco, G. (2022).
Functional network antagonism
and consciousness. Réseau
Neurosciences, 6(4), 998–1009.
https://doi.org/10.1162/netn_a_00244
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00244
Reçu: 1 Octobre 2021
Accepté: 6 Mars 2022
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Athena Demertzi
a.demertzi@uliege.be
Éditeur de manipulation:
Olaf Sporns
droits d'auteur: © 2022
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
Functional anticorrelations:
Negative functional connectivity that
certain brain regions show with
regard to the positive connectivity of
other regions as observed in zero-lag
correlations of BOLD activity.
communicate with their environment. A particular consciousness-mediated rsfMRI
configuration is that of functional anticorrelations, c'est, the antagonistic relationship
between a specific set of brain regions. We suggest that anticorrelations are a key physiological
prior, without which consciousness cannot be supported, because the brain cannot segregate
how regions get connected. We postulate that segregation is possible thanks to neural
inhibition, by regulating global and local inhibitory activity. We believe that the link between
anticorrelations and the underlying physiology can help not only to comprehend how
consciousness happens, but also conceptualize effective interventions for treating its disorders.
INTRODUCTION
Individuals during sleep, anesthesia, and in disorders of consciousness are unable to commu-
nicate intentionally with the environment. Par conséquent, their mental state needs to be inferred
by means of meaningful proxies. The fMRI resting paradigm has been a great asset to that
matter, as it quantifies brain function by surpassing the need for communication of experience
or behavioral output (Zhang et al., 2021). Dans l'ensemble, studies in such states of consciousness point
to lesser functional connectivity (FC) between regions that are within the same “network,” in
that they show positive FC during wakefulness (Heine et al., 2012). The positive coupling
between brain regions, especially those of the default mode network (DMN), was shown to
enable fast and accurate performance during higher order cognitive tasks concerning, pour
example, executive function (Shine et al., 2016) or working memory (Cohen & D'Esposito,
2016). Such integrative profiles reduce dramatically in states of unconsciousness—yet
within-network FC persists and does not entirely disappear (Boveroux et al., 2010; Di Perri
et coll., 2016). Donc, DMN correlations might be more about shaping connectivity interac-
tions rather than reflecting conscious mental activity (Boly et al., 2008).
A rather consciousness-sensitive connectivity profile is that of functional anticorrelations,
c'est, the negative FC that some regions show with the DMN. Depending on the state of
consciousness anticorrelations reduce in intensity, like after sleep deprivation (De Havas
et coll., 2012; Yeo et al., 2015), in slow wave sleep and REM (Chow et al., 2013), hypnosis
(Demertzi et al., 2011), and deep sedation (Luppi et al., 2019). Or they are undetectable, like
in deep anesthesia (Boveroux et al., 2010), and unresponsive brain-damaged patients (Di Perri
et coll., 2016; Threlkeld et al., 2018). Surtout, anticorrelations’ FC recovers during the
immediate postanesthetic period (Nir et al., 2020) and after emergence from a disorder of con-
sciousness (Di Perri et al., 2016; Threlkeld et al., 2018). Within the state of typical wakefulness,
the presence of anticorrelations was shown to contributes to cognitive function (J.. B. Keller et al.,
2015; Vanhaudenhuyse et al., 2011), with greater intensity leading to better within-subject
(par exemple., Kucyi et al., 2017) and between-subject performance (par exemple., Spreng et al., 2010). Aussi,
anticorrelations seem to contribute to life span, starting weak in children, strengthening during
adolescence, ending up fully anticorrelated in young adulthood (Chai et al., 2014), and getting
selectively decreased during healthy aging (J.. B. Keller et al., 2015). Considering that anti-
correlations are implicated in cognition and consciousness, an emerging question is what this
antagonistic configuration implies about the brain’s physiology and conscious experience. À
our knowledge, no such formulation has been suggested yet. To address this, we discuss con-
ceptual and methodological debates around anticorrelations, et, by tackling their physiolog-
ical underpinnings, we postulate a mechanistic link between micro- and macrocircuitry,
which may explain the function of anticorrelations in the context of conscious experience.
Neurosciences en réseau
999
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
Global signal:
BOLD signal time course averaged
across all brain voxels.
FUNCTIONAL ANTICORRELATIONS CONTAIN MEANINGFUL NEURAL ACTIVITY
Anticorrelations refer to brain regions showing negative FC in contrast to the positive FC within
intrinsic networks, such as the DMN. The regions showing negative FC with the DMN concern
primarily the intraparietal sulcus, the frontal eye fields, and the middle temporal + area (Fox
et coll., 2005). Historically, the areas showing anticorrelations were coined as “task-positive” in
contrast to a “task-negative” DMN (Fox et al., 2005). This connotation was given to highlight,
respectivement, activations and deactivations exhibited by these systems during task perfor-
mance, initially measured with positron emission tomography, and later confirmed by fMRI
(Raichle & Mintun, 2006). It has been proposed, cependant, that the dichotomization between
“task-positive” and “task-negative” might be misleading because it insinuates that the DMN is
not engaged actively in cognitive processes (Spreng, 2012). As the DMN indeed collaborates
with other task-related areas (Elton & Gao, 2015) and networks (Spreng, 2012) to promote
cognitive performance and mental flexibility (Spreng et al., 2014), we will here preferentially
utilize the term anticorrelations.
The discussion about anticorrelations very often goes hand in hand with the methodolog-
ical debate about correcting or not for the brain’s global signal (GS) during fMRI data prepro-
cessation. The GS can be obtained by averaging the resting-state time courses over the entire
brain (Desjardins et al., 2001). This whole-brain averaging implies the possibility that non-
neuronal sources can contribute to the GS along with neural signal. As most functional con-
nectivity studies are interested in identifying the neural counterparts of a task or a condition,
this implies that GS needs to be accounted for. GS correction can happen via linear regression,
subtraction, or normalization (T. T. Liu et al., 2017). Such a process, cependant, can lead to
systematically shifting the distribution of correlation values in the negative direction (Anderson
et coll., 2011; Murphy et al., 2009) et, donc, anticorrelations emerge. This implies that anticor-
relations are a matter of mathematical treatment, spurious, and not neuronally meaningful. À
date, cependant, there is support both for the nonneuronal and the neuronal significance of the
GS. D'une part, the GS is shown to reflect fMRI nuisance sources such as motion, scan-
ner artifacts, respiration (Power et al., 2017), cardiac rate (Chang & Glover, 2010), and vas-
cular activity (Colenbier et al., 2020; Zhu et al., 2015). On the other hand, GS is considered to
have a neuronal counterpart (Schölvinck et al., 2010) that promotes behavior (Li et al., 2019),
it was shown to correlate with spontaneous fluctuations in the local field potentials as mea-
sured with implanted electrodes in monkeys (Schölvinck et al., 2010), and it was associated
with vigilance (Wen & Liu, 2016; Wong et al., 2013) and arousal (X. Liu et al., 2018) comme
measured with EEG in humans. Ensemble, the debate about whether to employ GS correction
as a preprocessing step or not remains unresolved, while the choice can be driven by the
research question at hand (Murphy & Fox, 2017; Uddin, 2017).
Generally, we align with the view that the spontaneous anticorrelations are not mere arti-
facts and that they actually reflect neural activity. This is after considering that GS correction
does not preferentially affect only systems exhibiting positive correlations but also those which
show anticorrelations in the first place (Fox et al., 2009). Anticorrelations between the DMN
and the executive attention system can also be found using independent component analysis
(without GS correction), suggesting that the anticorrelations are not merely a mathematical
issue. Aussi, anticorrelations are shown to increase after caffeine intake, pointing to their phys-
iological dynamism (Wong et al., 2013). The anticorrelations between networks homologous
to DMN and the dorsal attention network (DAN) are also observed in rodents, dogs, and non-
human primates, confirming interspecies consistency (Belloy et al., 2018un; Gozzi & Schwarz,
2016; Hutchison & Everling, 2012; Szabó et al., 2019). En outre, the strength of anticor-
relations is shown to be predictive of disease phenotype (Adhikari et al., 2021; Belloy et al.,
Neurosciences en réseau
1000
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
Neural inhibition:
A physiological mechanism that
helps sustain balanced cortical
activity by orchestrating the
coordination between excitatory
pyramidal neurons and inhibitory
nonpyramidal cells.
Network metastability:
The ability of network transitioning
among variant functional states
au fil du temps.
2018b; Sripada et al., 2014) and is able to change by means of sensory stimulation, attention,
and neuromodulation (Hinz et al., 2019; Peeters et al., 2020). Enfin, the existence of spon-
taneous anticorrelated networks is evidenced in computational simulations in monkey and
human brains (Deco et al., 2009), as well as in neurophysiological studies. By means of simul-
taneous scalp EEG and fMRI in humans, a temporal relationship has been found between
increased alpha power and greater DMN-DAN anticorrelations (Chang et al., 2013), highlight-
ing their cognitive relevance. More direct evidence comes from intracranial recordings of local
field potentials (LFPs). In cats, simultaneous recordings of unit activity and LFPs showed that,
when attentional demands increased, LFP power in task-on (DAN-homologue) regions aug-
mented and task-off (DMN-homologue) regions decreased (Popa et al., 2009). Further human
intracranial EEG evidence suggests that during wakeful rest, areas of these networks exhibited
anticorrelated slow fluctuations of high gamma power (C. J.. Keller et al., 2013), ce qui était
correlated with neuronal firing rates (Manning et al., 2009). Aussi, high gamma power evoked
by tasks had opposing patterns in the DMN and antagonistic networks as measured by intra-
cranial EEG (Ossandón et al., 2011; Ramot et al., 2012). Enfin, intracranial EEG supports that
spontaneous, transient increases in high gamma (a proxy for neuronal spiking) regularly arose
within major nodes of the DMN versus DAN/salience networks and were time-locked to cog-
nitive and physiological events (Daitch & Parvizi, 2018; Kucyi & Parvizi, 2020). Collectively,
observational and interventional studies in humans and animals support that anticorrelations
have a physiological and neuronal importance.
NEURAL INHIBITION MEDIATES THE FORMULATION OF ANTICORRELATIONS
We postulate that anticorrelations may be emerging thanks to the process of neural inhibi-
tion. Neural inhibition is a pivotal mechanism for the brain to sustain balanced cortical activ-
ville (Isaacson & Scanziani, 2011). This is done by the orchestrated coordination between
excitatory pyramidal spiking neurons occupying 70%–80% of the cortex and the remaining
inhibitory nonpyramidal cells (DeFelipe & Fariñas, 1992), such that for every five excitatory
synapses there is approximately one inhibitory (Beaulieu & Colonnier, 1985). Excitation and
inhibition happen in a balanced way, leaving it unlikely to observe an increase in one with-
out observing an increase in the other; otherwise, no cell would reach firing threshold (Scannell
& Jeune, 1999). The recruitment of GABAergic inhibitory interneurons via the thalamus, corti-
cocortical, or other excitation pathways has been shown to assist cognition and motor behavior
(Swanson & Maffei, 2019). Aussi, it is via this synchronous activation of excitatory and inhibitory
activity that anticorrelated cortical network activity emerges (Arthurs & Boniface, 2002;
Logothetis, 2008).
Considering this mechanism in relation to anticorrelations, we do not claim that neural
inhibition directly promotes anticorrelated patterns, c'est, by having networks straightly inhi-
biting one another (of note, most interareal connections are glutamatergic/excitatory). Nor do
we claim that neural inhibition always leads to anticorrelated profiles. Plutôt, we suggest that
neural inhibition mediates the rise of the anticorrelations indirectly, by breaking the local neu-
ral balance which affects network metastability and which eventually permits anticorrelations
to appear. More particularly, current computational whole-brain models assume that distant
brain regions establish connections between their corresponding excitatory neural populations
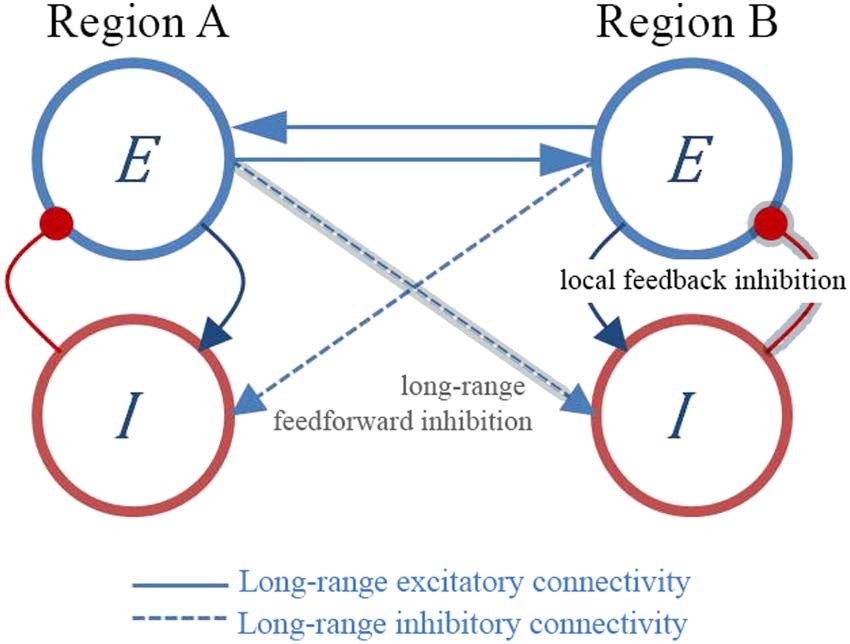
(Chiffre 1, E-E blue solid line connectivity). The resulting local increase of excitation produces
an increase of inhibition through the local E-I loop (feedback inhibition). Apart from the inter-
areal excitatory activity, une région du cerveau (UN) can also effectively inhibit the activity of a distal
brain region (B), by A targeting B’s inhibitory interneurons (Chiffre 1, E-I blue dashed line con-
nectivity), which in turn, locally connect to the pyramidal cells (Chiffre 1, I-E red connector;
Neurosciences en réseau
1001
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
Chiffre 1. Functional anticorrelations might emerge as a result of local and long-range inhibitory
activité. Brain regions are modeled by interconnected populations of excitatory (E) and inhibitory (je)
neurons. Long-range excitatory connections from brain region A to brain region B can target the
excitatory population (E → E, solid blue) or the inhibitory population (E → I, dashed blue). The level
of inhibition in region B depends i) on the local E-I loop (local feedback inhibition) and ii) on the
excitatory inputs from region A to the inhibitory neurons in region B, which in turn connect locally
to the excitatory cells (long-range feedforward inhibition, gray highlighted path). With this model,
we postulate that neural inhibition assists the emergence of the anticorrelations not by direct inter-
réseau, interareal inhibition (of note, most interareal connections around the brain are glutamater-
gic). Plutôt, anticorrelations appear indirectly after disrupting both local and global balance
between excitation and inhibition, eventually affecting network metastability.
c'est à dire., feedforward inhibition; Isaacson & Scanziani, 2011). Cependant, little attention has been
paid to this indirect long-range inhibition in modeling studies and how specifically feedforward
inhibition interacts with the local excitation-inhibition (E/I) ratio. Although previous theoretical
work has separately examined the effect of regulating the E/I ratio through feedback inhibition
or feedforward inhibition in whole-brain models (Deco et al., 2014), a model that takes both effects
into account has not been studied yet. Local heterogenous feedback inhibition, in particular, eux-mêmes-
nifies that the excitability of local population activity is achieved by variably determining each
region’s gain response function. Recent studies using heterogeneous and homogeneous whole-
brain modeling have mechanistically shown that, besides increasing the level of fitting of the
empirical data, an increase in ignition was observed (Deco et al., 2021). Based on this evidence,
one could speculate that ignition is related to the presence of regional heterogeneity. Bien que le
type of heterogeneity that is the most relevant for ignition remains open, one can nevertheless
say that thanks to heterogeneity in general, ignition-like dynamics may happen (Deco et al.,
2021), which are thought to support conscious experience (see next section).
Critique, it has been shown that, when feedback projections and randomization of the
connectivity weights were removed, dynamic network behavior was abolished, suggesting that
feedback connectivity and heterogeneity in connection strength gives rise to ignition-like activity
(Joglekar et al., 2018). We here hypothesize that the generation of the anticorrelations can be a
matter of both inhibitory effects (local feedback inhibition, long-range feedforward inhibition),
which will eventually affect network metastability, allowing the emergence of the anticorrelations.
À ce jour, the behavior of this model remains to be theoretically studied and empirically supported.
THE ANTICORRELATIONS’ SIGNIFICANCE TO CONSCIOUSNESS
On our quest to tackle the meaning of the anticorrelations to consciousness we lean toward
theoretical frameworks in order to see how they embrace the mechanism of inhibition. Nous
Gain response function:
A model parameter that can be set to
different values across brain regions
that impose various levels of
excitability.
Ignition:
The ability of a given brain area to
propagate feed-forward and
recurrent neuronal activity to other
régions.
Neurosciences en réseau
1002
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
Neuronal bistability:
The tendency of cortical neurons to
fall into a silent period (down state)
after an initial activation.
Segregation:
The brain’s capacity to show
selectivity in the way brain areas are
functionally connected.
notice that especially the global neuronal workspace theory (GNWT; Dehaene et al., 2003)
and the integrated information theory (IIT; Oizumi et al., 2014) are two such theories that uti-
lize inhibition when accounting for consciousness in their models.
The GNWT describes how reportable experience happens. The theory proposes a model
with various local processors that are all linked at a central executive. Its model suggests to
view the local processors as specialized modular cortical areas that process specific percep-
tual, moteur, mémoire, and evaluative information. The central executive (composed of widely
distributed excitatory neurons) can be considered as a second computational space, lequel
forms reciprocal connections to specific processor neurons. Based on this architecture, à
any moment, a piece of information within one or several processors can be selected, ampli-
fied, and broadcasted to all other processors, thus rendering it consciously accessible and
available for reporting. This global broadcasting happens thanks to ignition (Dehaene et al.,
2003; Mashour et al., 2020). Ignition is characterized by the sudden, coherent, and exclusive
activation of a subset of workspace neurons that codes the current conscious content, alors que
the remainder of the workspace neurons remain inhibited. Regardless of how ignition may be
triggered, simulations show that if feedforward connections are carefully balanced by local
inhibitory influences, incoming stimuli elicit a stable cascade of activity characterized by a
late and sudden ignition. Autrement dit, ignition of a global neural state coding for particular
conscious content leads to the active inhibition of other potential contents (as we also showed
before; Vanhaudenhuyse et al., 2011).
The IIT begins the quest of consciousness by experience itself. The theory first identifies the
experience’s essential properties (axioms) and then infers what kind of properties a physical
system must have in order to account for these properties (postulates). The postulates specify
which conditions (such as neurons and their connections) must be satisfied by physical mech-
anisms to account for the phenomenology of experience. To generate consciousness, a phys-
ical system must be able to discriminate among a large repertoire of states (information), and it
must be doing so as a single system, therefore not decomposable into a collection of causally
independent parts (l'intégration). For integrated information to be high, a system must be con-
nected in such a way that information is generated by causal interactions among its parts,
rather than within them. In balanced states, the corticothalamic system is a fine example of
a functionally integrated and specialized network, able to generate high values of integrated
information peaks that are characterized by a complex spatiotemporal pattern of signal prop-
agations in different directions (feedforward, feedback, contralateral) and with variant weights,
including inhibitory. In unbalanced states, like those of low arousal, the cortical neurons are
inactivated due to the neuronal bistability of their membrane potential or active inhibition, et
thus cannot specify a conceptual structure. In that case, the system then collapses, leading to
low values of integrated information, and hence diminished conscious experience.
If we try to link the inhibitory mechanisms described in these models with functional anti-
correlations, we can observe that these promote the notion of functional segregation. In that
respect, anticorrelations can be considered as an FC segregated profile, which mediates various
states of consciousness. Dans le passé, Fox et al. (2005) similarly suggested that “while correlations
may serve as an integrative role in combining neuronal activity subserving similar goals or rep-
resentations, anticorrelations may serve as a differentiating role segregating neuronal processes,
subserving opposite goals or competing representations” (Fox et al., 2005, p. 9677). This view
on segregation refers to the ability of a system to distinguish information into distinct modules
that can perform specialized local computations (Shannon, 1948). This definition may slightly
deviate from how graph theory considers segregation, c'est, close to the notion of modularity
and as a measure of the relative strength between a graph’s nodes (Sporns, 2013). In that respect,
Neurosciences en réseau
1003
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
inhibition as described in the GNWT can be viewed as the segregative processes that hinder
widespread FC, leading to negative functional correlations. De la même manière, differentiation as described
in the IIT can be similarly considered as parallel to the processes of segregation, also expressed
as anticorrelated FC.
Taken together, our view is that anticorrelations are the physiological expression of segre-
gation, and we propose that neural inhibition is the mediating link. Our stance, bien sûr, does
not prevent other measures from being linked to unconsciousness. En effet, low values in sam-
ple entropy (which quantifies how unpredictable a signal is; Luppi et al., 2019) or “small-
worldeness” (which allows for a cost-efficient network organization; Uehara et al., 2014)
are also affected in reduced consciousness. We also remain mindful of the fact that anticorre-
lations can still result by fMRI preprocessing steps, as described above, or from changes in the
hemodynamics in some regions or even from time delays that produce phase lags. The DAN,
par exemple, was found to precede DMN deactivation by up to hundreds of milliseconds
(Kucyi et al., 2020; Raccah et al., 2018). One possible explanation for that is that the anti-
correlations are driven by sequences of spontaneous neuronal population events across the
DMN and antagonistic systems that involve interregional temporal delays (Kucyi et al.,
2020). The observed zero-lag anticorrelations could therefore be due to “blurring” of such
delays, meaning anticorrelations could be the result of transient events that are comprised
of spontaneous activations that are systematically coupled to subsequent DMN deactivations,
as observed in transient coactivation patterns at rest (Karahanoğlu & Van De Ville, 2015; X. Liu
& Duyn, 2013). Cependant, this hypothesis requires further investigation. These issues keep our
hypothesis about anticorrelations being a metric of consciousness in check and call for
experimental and mechanistic explanations.
Collectively, we consider that FC anticorrelations are an essential ingredient for conscious
mental activity and might work as another marker of preserved consciousness. Nous, donc,
predict that if the brain is not in a state to host the physiological prior of inhibitory activity, alors
most likely the individual does not entertain subjective experience. This stance opens new
avenues for the understating and treatment of clinical cases of consciousness alternations
by targeting anticorrelations specifically as the outcome measure, like it has been shown using
neurofeedback (Bauer et al., 2020) or meditation (Bauer et al., 2019) for psychiatric disorders.
We think that the introduction of yet another metric is justified by the difference this metric
makes and the pragmatic issues it addresses (Demertzi et al., 2017). Ainsi, we align with the
view that, when searching for consciousness, accumulative evidence stemming from multiple
nonoverlapping assessments with different modalities needs to be applied (Seth et al., 2008)—
ideally, those that receive theoretical framing in order to account for the mechanistic expla-
nations of the metric at hand. What this view of the anticorrelations essentially offers is a link
between the level of neuronal microcircuitry and the computational level, which starts gaining
support when attempting to describe how consciousness happens (Changeux, 2017).
CONCLUSIONS
We suggest that the FC anticorrelations emerge thanks to local and global neural inhibitory
activité, which leads to variant spatiotemporal configurations. Such rich network organization
was previously shown to characterize typical conscious conditions, while simpler interre-
gional connectivity was most frequently seen in anesthetized states and states of low report-
ability (Barttfeld et al., 2014; Demertzi et al., 2019; Huang et al., 2020). We eventually invite
researchers to view anticorrelations as the physiological expression of segregation via neural
inhibition, which can help us not only comprehend how consciousness happens, mais aussi
Neurosciences en réseau
1004
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
conceptualize and design effective interventions for treating consciousness disorders in which
anticorrelations seem particularly affected.
REMERCIEMENTS
We would like to thank Mr. Fort Larry, M.A., for proofreading the manuscript for grammar and
syntax.
CONTRIBUTIONS DES AUTEURS
Athena Demertzi: Conceptualisation; Enquête; Gestion de projet; Ressources;
Rédaction – ébauche originale; Rédaction – révision & édition. Aaron Kucyi: Enquête; Ressources;
Validation. Adrián Ponce-Alvarez: Méthodologie; Validation; Visualisation; Rédaction – révision
& édition. Georgios A. Keliris: Enquête; Méthodologie; Ressources; Validation. Susan
Whitfield-Gabrieli: Ressources; Surveillance; Validation. Gustavo Deco: Méthodologie;
Ressources; Surveillance; Validation; Visualisation.
INFORMATIONS SUR LE FINANCEMENT
Athena Demertzi, Fonds De La Recherche Scientifique – FNRS (https://dx.doi.org/10.13039
/501100002661).
RÉFÉRENCES
Adhikari, M.. H., Belloy, M.. E., Van der Linden, UN., Keliris, G. UN., &
Verhoye, M.. (2021). Resting-state co-activation patterns as prom-
ising candidates for prediction of Alzheimer’s disease in aged
mice. Frontiers in Neural Circuits, 14, 612529. https://doi.org
/10.3389/fncir.2020.612529, PubMed: 33551755
Anderson, J.. S., Druzgal, T. J., Lopez-Larson, M., Jeong, E. K., Desai,
K., & Yurgelun-Todd, D. (2011). Network anticorrelations, mondial
regression, and phase-shifted soft tissue correction. Cerveau humain
Cartographie, 32(6), 919–934. https://doi.org/10.1002/hbm.21079,
PubMed: 20533557
Arthurs, Ô. J., & Boniface, S. (2002). How well do we understand
the neural origins of the fMRI BOLD signal? Trends in Neurosci-
ences, 25(1), 27–31. https://doi.org/10.1016/s0166-2236(00)
01995-0, PubMed: 11801335
Barttfeld, P., Uhrig, L., Sitt, J.. D., Sigman, M., Jarraya, B., &
Dehaene, S. (2014). Signature of consciousness in the dynamics
of resting-state brain activity. Proceedings of the National Acad-
emy of Sciences, 112(3), 887–892. https://doi.org/10.1073/pnas
.1418031112, PubMed: 25561541
Bauer, C. C. C., Okano, K., Ghosh, S. S., Lee, Oui. J., Melero, H.,
Angeles, C. d. L., Nestor, P.. G., Del Re, E. C., Northoff, G.,
Niznikiewicz, M.. UN., & Whitfield-Gabrieli, S. (2020). Real-time
fMRI neurofeedback reduces auditory hallucinations and modu-
lates resting state connectivity of involved brain regions: Part 2:
Default mode network-preliminary evidence. Psychiatry
Research, 284, 112770. https://doi.org/10.1016/j.psychres.2020
.112770, PubMed: 32004893
Bauer, C. C. C., Whitfield-Gabrieli, S., Díaz, J.. L., Pasaye, E. H., &
( 2 0 1 9 ) . Fr o m s t a t e – t o – t r a i t m e d i t a t i o n :
B a r r i o s , F. UN .
Reconfiguration of central executive and default mode networks.
ENeuro, 6(6). https://doi.org/10.1523/ENEURO.0335-18.2019,
PubMed: 31694816
Beaulieu, C., & Colonnier, M.. (1985). A laminar analysis of the
number of round-asymmetrical and flat-symmetrical synapses
on spines, dendritic trunks, and cell bodies in area 17 of the
cat. Journal of Comparative Neurology, 231(2), 180–189.
https://doi.org/10.1002/cne.902310206, PubMed: 3968234
Belloy, M.. E., Naeyaert, M., Abbas, UN., Shah, D., Vanreusel, V., van
Audekerke, J., Keilholz, S. D., Keliris, G. UN., Van der Linden, UN.,
& Verhoye, M.. (2018un). Dynamic resting state fMRI analysis in
mice reveals a set of quasi-periodic patterns and illustrates their
relationship with the global signal. NeuroImage, 180(Pt B),
463–484. https://doi.org/10.1016/j.neuroimage.2018.01.075,
PubMed: 29454935
Belloy, M.. E., Shah, D., Abbas, UN., Kashyap, UN., Roßner, S., Van der
Linden, UN., Keilholz, S. D., Keliris, G. UN., & Verhoye, M.. (2018b).
Quasi-periodic patterns of neural activity improve classification
of Alzheimer’s disease in mice. Rapports scientifiques, 8(1), 10024.
https://doi.org/10.1038/s41598-018-28237-9, PubMed:
29968786
Boly, M., Phillips, C., Tshibanda, L., Vanhaudenhuyse, UN., Schabus,
M., Dang-Vu, T. T., Moonen, G., Hustinx, R., Maquet, P., & Laureys,
S. (2008). Intrinsic brain activity in altered states of consciousness:
How conscious is the default mode of brain function? Annals of the
New York Academy of Sciences, 1129, 119–129. https://doi.org
/10.1196/annals.1417.015, PubMed: 18591474
Boveroux, P., Vanhaudenhuyse, UN., Bruno, M.. UN., Noirhomme, Q.,
Lauwick, S., Luxen, UN., Degueldre, C., Plenevaux, UN., Schnakers,
Neurosciences en réseau
1005
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
C., Phillips, C., Brichant, J.. F., Bonhomme, V., Maquet, P.,
Greicius, M.. D., Laureys, S., & Boly, M.. (2010). Breakdown of
within- and between-network resting state functional magnetic
resonance imaging connectivity during propofol-induced loss of
consciousness. Anesthesiology, 113(5), 1038–1053. https://est ce que je
.org/10.1097/ALN.0b013e3181f697f5, PubMed: 20885292
Chaï, X. J., Ofen, N., Gabriela, J.. D. E., & Whitfield-Gabrieli, S.
(2014). Selective development of anticorrelated networks in the
intrinsic functional organization of the human brain. Journal de
Neurosciences cognitives, 26(3), 501–513. https://est ce que je.org/10
.1162/jocn_a_00517, PubMed: 24188367
Chang, C., & Glover, G. H. (2010). Time–frequency dynamics of
resting-state brain connectivity measured with fMRI. Neuro-
Image, 50(1), 81–98. https://doi.org/10.1016/j.neuroimage.2009
.12.011, PubMed: 20006716
Chang, C., Liu, Z., Chen, M.. C., Liu, X., & Duyn, J.. H. (2013). EEG
correlates of time-varying BOLD functional connectivity. Neuro-
Image, 72, 227–236. https://doi.org/10.1016/j.neuroimage.2013
.01.049, PubMed: 23376790
Changeux, J.-P. (2017). Climbing brain levels of organisation from
genes to consciousness. Tendances des sciences cognitives, 21(3),
168–181. https://doi.org/10.1016/j.tics.2017.01.004, PubMed:
28161289
Chow, H. M., Horovitz, S. G., Carr, W. S., Picchioni, D.,
Coddington, N., Fukunaga, M., Xu, Y., Balkin, T. J., Duyn,
J.. H., & Brun, UN. R.. (2013). Rhythmic alternating patterns of
brain activity distinguish rapid eye movement sleep from
other states of consciousness. Actes de la Nationale
Academy of Sciences of the United States of America, 110(25),
10300–10305. https://doi.org/10.1073/pnas.1217691110,
PubMed: 23733938
Cohen, J.. R., & D'Esposito, M.. (2016). The segregation and
integration of distinct brain networks and their relationship
to cognition. Journal des neurosciences, 36(48), 12083–12094.
https://doi.org/10.1523/ JNEUROSCI.2965-15.2016, PubMed:
27903719
Colenbier, N., Van de Steen, F., Uddin, L. Q., Poldrack, R.. UN.,
Calhoun, V. D., & Marinazzo, D. (2020). Disambiguating the
role of blood flow and global signal with partial information
decomposition. NeuroImage, 213, 116699. https://est ce que je.org/10
.1016/j.neuroimage.2020.116699, PubMed: 32179104
Daitch, UN. L., & Parvizi, J.. (2018). Spatial and temporal heterogene-
ity of neural responses in human posteromedial cortex. Proceed-
ings of the National Academy of Sciences of the United States of
America, 115(18), 4785–4790. https://doi.org/10.1073/pnas
.1721714115, PubMed: 29666262
De Havas, J.. UN., Parimal, S., Soon, C. S., & Chee, M.. W. L. (2012).
Sleep deprivation reduces default mode network connectivity
and anti-correlation during rest and task performance. Neuro-
Image, 59(2), 1745–1751. https://doi.org/10.1016/j.neuroimage
.2011.08.026, PubMed: 21872664
Deco, G., Jirsa, V., McIntosh, UN. R., Sporns, O., & Kötter, R.. (2009).
Key role of coupling, delay, and noise in resting brain fluctua-
tion. Proceedings of the National Academy of Sciences of the
United States of America, 106(25), 10302–10307. https://est ce que je
.org/10.1073/pnas.0901831106, PubMed: 19497858
Deco, G., Kringelbach, M.. L., Arnatkeviciute, UN., Oldham, S.,
Sabaroedin, K., Rogasch, N. C., Aquino, K. M., & Fornito, UN.
(2021). Dynamical consequences of regional heterogeneity in
the brain’s transcriptional landscape. Science Advances, 7(29),
eabf4752. https://doi.org/10.1126/sciadv.abf4752, PubMed:
34261652
Deco, G., Ponce-Alvarez, UN., Hagmann, P., Romani, G. L., Mantini,
D., & Corbetta, M.. (2014). How local excitation-inhibition ratio
impacts the whole brain dynamics. Journal des neurosciences,
34(23), 7886–7898. https://doi.org/10.1523/JNEUROSCI.5068
-13.2014, PubMed: 24899711
DeFelipe, J., & Fariñas, je. (1992). The pyramidal neuron of the cere-
bral cortex: Morphological and chemical characteristics of the
synaptic inputs. Progrès en neurobiologie, 39(6), 563–607. https://
doi.org/10.1016/0301-0082(92)90015-7, PubMed: 1410442
Dehaene, S., Sergent, C., & Changeux, J.-P. (2003). A neuronal net-
work model linking subjective reports and objective physiologi-
cal data during conscious perception. Actes du
National Academy of Sciences of Sciences of the United States
of America, 100(14), 8520–8525. https://doi.org/10.1073/pnas
.1332574100, PubMed: 12829797
Demertzi, UN., Sitt, J.. D., Sarasso, S., & Pinxten, W. (2017). Measuring
states of pathological (et)consciousness: Research dimensions,
clinical applications and ethics. Neuroscience of Consciousness,
3(1), 1–13. https://doi.org/10.1093/nc/nix010, PubMed:
30042843
Demertzi, UN., Soddu, UN., Faymonville, M., Bahri, M.. UN., Gosseries,
O., Vanhaudenhuyse, UN., Phillips, C., Maquet, P., Noirhomme,
(2011). Hypnotic modulation
Q., Luxen, UN., & Laureys, S.
of resting state fMRI default mode and extrinsic network con-
nectivity. Progress in Brain Research, 193, 309–322. https://est ce que je
.org/10.1016/ B978-0-444-53839-0.00020-X, PubMed:
21854971
Demertzi, UN., Tagliazucchi, E., Dehaene, S., Deco, G., Barttfeld, P.,
Raimondo, F., Martial, C., Fernández-Espejo, D., Rohaut, B.,
Voss, H. U., Schiff, N. D., Owen, UN. M., Laureys, S., Naccache,
L., & Sitt, J.. D. (2019). Human consciousness is supported by
dynamic complex patterns of brain signal coordination. Science
Advances, 5(2), eaat7603. https://doi.org/10.1126/sciadv
.aat7603, PubMed: 30775433
Desjardins, UN. E., Kiehl, K. UN., & Liddle, P.. F. (2001). Removal of
confounding effects of global signal in functional MRI analyses.
NeuroImage, 13(4), 751–758. https://doi.org/10.1006/nimg.2000
.0719, PubMed: 11305902
Di Perri, C., Bahri, M.. UN., Amico, E., Thibaut, UN., Heine, L.,
Antonopoulos, G., Charland-Verville, V., Wannez, S., Gomez, F.,
Hustinx, R., Tshibanda, L., Demertzi, UN., Soddu, UN., & Laureys, S.
(2016). Neural correlates of consciousness in patients who have
emerged from a minimally conscious state: A cross-sectional mul-
timodal imaging study. La neurologie du Lancet, 15(8), 830–842.
https://est ce que je.org/10.1016/S1474-4422(16)00111-3, PubMed:
27131917
Elton, UN., & Gao, W. (2015). Task-positive functional connectivity
of the default mode network transcends task domain. Journal de
Neurosciences cognitives, 27(12), 2369–2381. https://est ce que je.org/10
.1162/jocn_a_00859, PubMed: 26244722
Fox, M.. D., Snyder, UN. Z., Vincent, J.. L., Corbetta, M., Van Essen,
D. C., & Raichle, M.. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Proceedings of the National Academy of Sciences of the United
Neurosciences en réseau
1006
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
States of America, 102(27), 9673–9678. https://doi.org/10.1073
/pnas.0504136102, PubMed: 15976020
Fox, M.. D., Zhang, D., Snyder, UN. Z., & Raichle, M.. E. (2009). Le
global signal and observed anticorrelated resting state brain net-
travaux. Journal de neurophysiologie, 101, 3270–3283. https://est ce que je
.org/10.1152/jn.90777.2008, PubMed: 19339462
Gozzi, UN., & Schwarz, UN. J.. (2016). Large-scale functional connec-
tivity networks in the rodent brain. NeuroImage, 127, 496–509.
https://doi.org/10.1016/j.neuroimage.2015.12.017, PubMed:
26706448
Heine, L., Soddu, UN., Gómez, F., Vanhaudenhuyse, UN., Tshibanda,
L., Thonnard, M., Charland-Verville, V., kirsch, M., Laureys, S., &
Demertzi, UN. (2012). Resting state networks and consciousness:
Alterations of multiple resting state network connectivity in phys-
iological, pharmacological, and pathological consciousness
states. Frontiers in Psychology, 3, 295. https://doi.org/10.3389
/fpsyg.2012.00295, PubMed: 22969735
Hinz, R., Peeters, L. M., Shah, D., Missault, S., Belloy, M., Vanreusel,
V., Malekzadeh, M., Verhoye, M., Van der Linden, UN., & Keliris,
G. UN. (2019). Bottom-up sensory processing can induce negative
BOLD responses and reduce functional connectivity in nodes
of the default mode-like network in rats. NeuroImage, 197,
167–176. https://doi.org/10.1016/j.neuroimage.2019.04.065,
PubMed: 31029872
Huang, Z., Zhang, J., Wu, J., Mashour, G. UN., & Hudetz, UN. G. (2020).
Temporal circuit of macroscale dynamic brain activity supports
human consciousness. Science Advances, 6(11), eaaz0087.
https://doi.org/10.1126/sciadv.aaz0087, PubMed: 32195349
Hutchison, R.. M., & Everling, S. (2012). Monkey in the middle:
Why non-human primates are needed to bridge the gap in
resting-state investigations. Frontiers in Neuroanatomy, 6, 29.
https://doi.org/10.3389/fnana.2012.00029, PubMed: 22855672
Isaacson, J.. S., & Scanziani, M.. (2011). How inhibition shapes cor-
tical activity. Neurone, 72(2), 231–243. https://est ce que je.org/10.1016/j
.neuron.2011.09.027, PubMed: 22017986
Joglekar, M.. R., Mejias, J.. F., Lequel, G. R., & Wang, X.-J. (2018).
Inter-areal balanced amplification enhances signal propagation
in a large-scale circuit model of the primate cortex. Neurone,
98(1), 222–234. https://doi.org/10.1016/j.neuron.2018.02.031,
PubMed: 29576389
Karahanoğlu, F. JE., & Van De Ville, D. (2015). Transient brain activ-
ity disentangles fMRI resting-state dynamics in terms of spatially
and temporally overlapping networks. Communications naturelles,
6, 7751. https://doi.org/10.1038/ncomms8751, PubMed:
26178017
Keller, C. J., Bickel, S., Honey, C. J., Groppe, D. M., Entz, L.,
Craddock, R.. C., Lado, F. UN., Kelly, C., Milham, M., &
Mehta, UN. D. (2013). Neurophysiological investigation of
spontaneous correlated and anticorrelated fluctuations of the
BOLD signal. Journal des neurosciences, 33(15), 6333–6342.
https://doi.org/10.1523/ JNEUROSCI.4837-12.2013, PubMed:
23575832
Keller, J.. B., Hedden, T., Thompson, T. W., Anteraper, S. UN.,
Gabriela, J.. D. E., & Whitfield-Gabrieli, S. (2015). Resting-state
anticorrelations between medial and lateral prefrontal cortex:
Association with working memory, aging, and individual differ-
ences. Cortex, 64, 271–280. https://doi.org/10.1016/j.cortex
.2014.12.001, PubMed: 25562175
Kucyi, UN., Daitch, UN., Raccah, O., Zhao, B., Zhang, C., Esterman,
M., Zeineh, M., Halperne, C. H., Zhang, K., Zhang, J., & Parvizi, J..
(2020). Electrophysiological dynamics of antagonistic brain net-
works reflect attentional fluctuations. Communications naturelles,
11(1), 325. https://doi.org/10.1038/s41467-019-14166-2,
PubMed: 31949140
Kucyi, UN., Hove, M.. J., Esterman, M., Hutchison, R.. M., & Valera,
E. M.. (2017). Dynamic brain network correlates of spontaneous
fluctuations in attention. Cortex cérébral, 27(3), 1831–1840.
https://doi.org/10.1093/cercor/bhw029, PubMed: 26874182
Kucyi, UN., & Parvizi, J.. (2020). Pupillary dynamics link sponta-
neous and task-evoked activations recorded directly from
Journal des neurosciences, 40(32), 6207–6218.
human insula.
https://doi.org/10.1523/ JNEUROSCI.0435-20.2020, PubMed:
32631937
Li, J., Kong, R., Liégeois, R., Orban, C., Tan, Y., Sun, N., Holmes,
UN. J., Sabuncu, M.. R., Ge, T., & Yeo, B. T. T. (2019). Global signal
regression strengthens association between resting-state func-
tional connectivity and behavior. NeuroImage, 196, 126–141.
https://doi.org/10.1016/j.neuroimage.2019.04.016, PubMed:
30974241
Liu, T. T., Nalci, UN., & Falahpour, M.. (2017). The global signal in
IRMf: Nuisance or Information? NeuroImage, 150, 213–229.
https://doi.org/10.1016/j.neuroimage.2017.02.036, PubMed:
28213118
Liu, X., de Zwart, J.. UN., Schölvinck, M.. L., Chang, C., Ye, F. Q.,
Leopold, D. UN., & Duyn, J.. H. (2018). Subcortical evidence for
a contribution of arousal to fMRI studies of brain activity.
Communications naturelles, 9(1), 395. https://est ce que je.org/10.1038
/s41467-017-02815-3, PubMed: 29374172
Liu, X., & Duyn, J.. H. (2013). Time-varying functional network
information extracted from brief instances of spontaneous brain
activité. Proceedings of the National Academy of Sciences of the
United States of America, 110(11), 4392–4397. https://est ce que je.org/10
.1073/pnas.1216856110, PubMed: 23440216
Logothetis, N. K. (2008). What we can do and what we cannot do
with fMRI. Nature, 453(7197), 869–878. https://est ce que je.org/10.1038
/nature06976, PubMed: 18548064
Luppi, UN. JE., Craig, M.. M., Pappas, JE., Finoia, P., Williams, G. B.,
Allanson, J., Pickard, J.. D., Owen, UN. M., Naci, L., Menon,
D. K., & Stamatakis, E. UN. (2019). Consciousness-specific
dynamic interactions of brain integration and functional diver-
ville. Communications naturelles, 10(1), 4616. https://est ce que je.org/10
.1038/s41467-019-12658-9, PubMed: 31601811
Manning, J.. R., Jacobs, J., Frit, JE., & Kahana, M.. J.. (2009). Broad-
band shifts in local field potential power spectra are correlated
with single-neuron spiking in humans. Journal des neurosciences,
29(43), 13613–13620. https://doi.org/10.1523/JNEUROSCI.2041
-09.2009, PubMed: 19864573
Mashour, G. UN., Roelfsema, P., Changeux, J.-P., & Dehaene, S.
(2020). Conscious processing and the global neuronal work-
space hypothesis. Neurone, 105(5), 776–798. https://est ce que je.org/10
.1016/j.neuron.2020.01.026, PubMed: 32135090
Murphy, K., Birn, R.. M., Handwerker, D. UN., Jones, T. B., &
Bandettini, P.. UN. (2009). The impact of global signal regression
on resting state correlations: Are anti-correlated networks intro-
duced? NeuroImage, 44(3), 893–905. https://est ce que je.org/10.1016/j
.neuroimage.2008.09.036, PubMed: 18976716
Neurosciences en réseau
1007
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
Murphy, K., & Fox, M.. D. (2017). Towards a consensus regarding
global signal regression for resting state functional connectivity
IRM. NeuroImage, 154, 169–173. https://est ce que je.org/10.1016/j
.neuroimage.2016.11.052, PubMed: 27888059
Nir, T., Jacob, Y., Huang, K.-H., Schwartz, UN. E., Brallier, J.. W., Ahn,
H., Kundu, P., Tang, C. Y., Delman, B. N., McCormick, P.. J.,
Sano, M., Deiner, S., Baxter, M.. G., & Mincer, J.. S. (2020).
Resting-state functional connectivity in early postanaesthesia
recovery is characterised by globally reduced anticorrelations.
British Journal of Anaesthesia, 125(4), 529–538. https://doi.org
/10.1016/j.bja.2020.06.058, PubMed: 32800503
Oizumi, M., Albantakis, L., & Tononi, G. (2014). From the phenom-
enology to the mechanisms of consciousness: Integrated informa-
tion theory 3.0. Biologie computationnelle PLoS, 10(5), e1003588.
https://doi.org/10.1371/journal.pcbi.1003588, PubMed:
24811198
Ossandón, T., Jerbi, K., Vidal, J.. R., Bayle, D. J., Henaff, M.-A., Jung,
J., Minotti, L., Bertrand, O., Kahane, P., & Lachaux, J.-P. (2011).
Transient suppression of broadband gamma power in the
default-mode network is correlated with task complexity and sub-
ject performance. Journal des neurosciences, 31(41), 14521–14530.
https://doi.org/10.1523/ JNEUROSCI.2483-11.2011, PubMed:
21994368
Peeters, L. M., van den Berg, M., Hinz, R., Majumdar, G., Pintelon,
JE., & Keliris, G. UN. (2020). Cholinergic modulation of the default
mode like network in rats. IScience, 23(9), 101455. https://est ce que je
.org/10.1016/j.isci.2020.101455, PubMed: 32846343
Popa, D., Popescu, UN. T., & Paré, D. (2009). Contrasting activity
profile of two distributed cortical networks as a function of atten-
tional demands. Journal des neurosciences, 29(4), 1191–1201.
https://doi.org/10.1523/ JNEUROSCI.4867-08.2009, PubMed:
19176827
Power, J.. D., Plitt, M., Laumann, T. O., & Martine, UN. (2017). Sources
and implications of whole-brain fMRI signals in humans. Neuro-
Image, 146, 609–625. https://doi.org/10.1016/j.neuroimage
.2016.09.038, PubMed: 27751941
Raccah, O., Daitch, UN. L., Kucyi, UN., & Parvizi, J.. (2018). Direct cor-
tical recordings suggest temporal order of task-evoked responses
in human dorsal attention and default networks. Journal of Neu-
roscience, 38(48), 10305–10313. https://est ce que je.org/10.1523
/JNEUROSCI.0079-18.2018, PubMed: 30315126
Raichle, M.. E., & Mintun, M.. UN. (2006). Brain work and brain imag-
ing. Revue annuelle des neurosciences, 29(1), 449–476. https://est ce que je
.org/10.1146/annurev.neuro.29.051605.112819, PubMed:
16776593
Ramot, M., Fisch, L., Harel, M., Kipervasser, S., Andelman, F.,
Neufeld, M.. Y., Kramer, U., Frit, JE., & Malach, R.. (2012). UN
widely distributed spectral signature of task-negative electrocor-
ticography responses revealed during a visuomotor task in the
human cortex. Journal des neurosciences, 32(31), 10458–10469.
https://doi.org/10.1523/ JNEUROSCI.0877-12.2012, PubMed:
22855795
Scannell, J.. W., & Jeune, M.. P.. (1999). Neuronal population activ-
ity and functional imaging. Proceedings of the Royal Society B:
Sciences biologiques, 266(1422), 875–881. https://est ce que je.org/10
.1098/rspb.1999.0718, PubMed: 10380677
Schölvinck, M.. L., Maier, UN., Ye, F. Q., Duyn, J.. H., & Leopold,
D. UN. (2010). Neural basis of global resting-state fMRI activity.
Proceedings of the National Academy of Sciences of the United
States of America, 107(22), 10238–10243. https://est ce que je.org/10
.1073/pnas.0913110107, PubMed: 20439733
Seth, UN. K., Dienes, Z., Cleeremans, UN., Overgaard, M., & Personne, L.
(2008). Measuring consciousness: Relating behavioural and neu-
rophysiological approaches. Tendances des sciences cognitives, 12(8),
314–321. https://doi.org/10.1016/j.tics.2008.04.008, PubMed:
18606562
Shannon, C. E. (1948). A mathematical theory of communication.
Bell System Technical Journal, 27(3), 379–423. https://est ce que je.org/10
.1002/j.1538-7305.1948.tb01338.x
Shine, J.. M., Bissett, P.. G., Cloche, P.. T., Koyejo, O., Balsters, J.. H.,
Gorgolewski, K. J., Moodie, C. UN., & Poldrack, R.. UN. (2016). Le
dynamics of functional brain networks: Integrated network states
during cognitive task performance. Neurone, 92(2), 544–554.
https://doi.org/10.1016/j.neuron.2016.09.018, PubMed: 27693256
Sporns, Ô. (2013). Network attributes for segregation and integra-
tion in the human brain. Opinion actuelle en neurobiologie, 23(2),
162–171. https://doi.org/10.1016/j.conb.2012.11.015, PubMed:
23294553
Spreng, R.. N. (2012). The fallacy of a “task-negative” network.
Frontiers in Psychology, 3, 145. https://doi.org/10.3389/fpsyg
.2012.00145, PubMed: 22593750
Spreng, R.. N., DuPre, E., Selarka, D., Garcia, J., Gojkovic, S.,
Mildner, J., Luh, W.-M., & Tourneur, G. R.. (2014). Goal-congruent
default network activity facilitates cognitive control. Journal de
Neurosciences, 34(42), 14108–14114. https://est ce que je.org/10.1523
/JNEUROSCI.2815-14.2014, PubMed: 25319706
Spreng, R.. N., Stevens, W. D., Chamberlain, J.. P., Gilmore, UN. W., &
Schacter, D. L. (2010). Default network activity, coupled with the
frontoparietal control network, supports goal-directed cognition.
NeuroImage, 53(1), 303–317. https://est ce que je.org/10.1016/j
.neuroimage.2010.06.016, PubMed: 20600998
Sripada, C. S., Kessler, D., & Angstadt, M.. (2014). Lag in maturation
of the brain’s intrinsic functional architecture in attention-deficit/
hyperactivity disorder. Proceedings of the National Academy of
Sciences of Sciences of the United States of America, 111(39),
14259–14264. https://doi.org/10.1073/pnas.1407787111,
PubMed: 25225387
Swanson, Ô. K., & Maffei, UN. (2019). From hiring to firing: Activa-
tion of inhibitory neurons and their recruitment in behavior. Fron-
tiers in Molecular Neuroscience, 12, 168. https://doi.org/10.3389
/fnmol.2019.00168, PubMed: 31333413
Szabó, D., Czeibert, K., Kettinger, Á., Gácsi, M., Andics, UN.,
Miklósi, Á., & Kubinyi, E. (2019). Resting-state fMRI data of
awake dogs (Canis familiaris) via group-level independent com-
ponent analysis reveal multiple, spatially distributed resting-state
réseaux. Rapports scientifiques, 9(1), 15270. https://est ce que je.org/10.1038
/s41598-019-51752-2, PubMed: 31649271
Threlkeld, Z. D., Bodien, Oui. G., Rosenthal, E. S., Giacino, J.. T.,
Nieto-Castanon, UN., Wu, O., Whitfield-Gabrieli, S., & Edlow,
B. L. (2018). Functional networks reemerge during recovery of
consciousness after acute severe traumatic brain injury. Cortex,
106, 299–308. https://doi.org/10.1016/j.cortex.2018.05.004,
PubMed: 29871771
Uddin, L. Q. (2017). Mixed signals: On separating brain signal from
bruit. Tendances des sciences cognitives, 21(6), 405–406. https://est ce que je
.org/10.1016/j.tics.2017.04.002, PubMed: 28461113
Neurosciences en réseau
1008
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Anticorrelations and consciousness
J.-I., & Tobimatsu, S.
Uehara, T., Yamasaki, T., Okamoto, T., Koike, T., Kan, S., Miyauchi,
S., Kira,
(2014). Efficiency of a
“small-world” brain network depends on consciousness level:
A resting-state fMRI study. Cortex cérébral, 24(6), 1529–1539.
https://doi.org/10.1093/cercor/bht004, PubMed: 23349223
Vanhaudenhuyse, UN., Demertzi, UN., Schabus, M., Noirhomme, Q.,
Bredart, S., Boly, M., Phillips, C., Soddu, UN., Luxen, UN., Moonen,
G., & Laureys, S. (2011). Two distinct neuronal networks mediate
the awareness of environment and of self. Journal of Cognitive
Neurosciences, 23(3), 570–578. https://doi.org/10.1162/jocn
.2010.21488, PubMed: 20515407
Wen, H., & Liu, Z. (2016). Broadband electrophysiological dynam-
ics contribute to global resting-State fMRI signal. Journal of Neu-
roscience, 36(22), 6030–6040. https://est ce que je.org/10.1523
/JNEUROSCI.0187-16.2016, PubMed: 27251624
Wong, C. W., Olafsson, V., Tal, O., & Liu, T. T. (2013). Le
amplitude of the resting-state fMRI global signal is related to
EEG vigilance measures. NeuroImage, 83, 983–990. https://
doi.org/10.1016/j.neuroimage.2013.07.057, PubMed:
23899724
Yeo, B. T. T., Tandi, J., & Chee, M.. W. L. (2015). Functional connec-
tivity during rested wakefulness predicts vulnerability to sleep
deprivation. NeuroImage, 111, 147–158. https://est ce que je.org/10.1016
/j.neuroimage.2015.02.018, PubMed: 25700949
Zhang, J., Kucyi, UN., Raya, J., Nielsen, UN. N., Nomi, J.. S., Damoiseaux,
J.. S., Vert, D. J., Horovitz, S. G., Uddin, L. Q., & Whitfield-
Gabriela, S. (2021). What have we really learned from functional
connectivity in clinical populations? NeuroImage, 242, 118466.
https://doi.org/10.1016/j.neuroimage.2021.118466, PubMed:
34389443
Zhu, D. C., Tarumi, T., Khan, M.. UN., & Zhang, R.. (2015). Vascular
coupling in resting-state FMRI: Evidence from multiple modalities.
Journal of Cerebral Blood Flow & Metabolism, 35(12), 1910–1920.
https://doi.org/10.1038/jcbfm.2015.166, PubMed: 26174326
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
6
4
9
9
8
2
0
5
6
2
5
2
n
e
n
_
un
_
0
0
2
4
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurosciences en réseau
1009