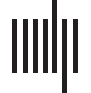FONCTION DE MISE AU POINT:
Connectivity, Cognition, and Consciousness
Allostasis as a core feature of hierarchical
gradients in the human brain
Yuta Katsumi1
, Jordan E. Theriault2, Karen S. Quigley2, and Lisa Feldman Barrett2,3,4
1Department of Neurology, Massachusetts General Hospital and Harvard Medical School, Boston, MA, Etats-Unis
2Département de psychologie, Northeastern University, Boston, MA, Etats-Unis
3Département de psychiatrie, Massachusetts General Hospital and Harvard Medical School, Boston, MA, Etats-Unis
4Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital and
Harvard Medical School, Boston, MA, Etats-Unis
un accès ouvert
journal
Mots clés: Traitement prédictif, Codage prédictif, Active inference, Interoception, Functional
réseaux, Energetics
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
t
/
.
ABSTRAIT
This paper integrates emerging evidence from two broad streams of scientific literature into
one common framework: (un) hierarchical gradients of functional connectivity that reflect the
brain’s large-scale structural architecture (par exemple., a lamination gradient in the cerebral cortex);
et (b) approaches to predictive processing and one of its specific instantiations called
allostasis (c'est à dire., the predictive regulation of energetic resources in the service of coordinating
the body’s internal systems). This synthesis begins to sketch a coherent, neurobiologically
inspired framework suggesting that predictive energy regulation is at the core of human brain
fonction, and by extension, psychological and behavioral phenomena, providing a shared
vocabulary for theory building and knowledge accumulation.
RÉSUMÉ DE L'AUTEUR
Allostasis refers to the process by which the brain anticipates the needs of the body and
attempts to meet those needs before they arise, and is one specific instantiation of a broader
predictive processing framework. In this perspective article, we propose that allostasis is a
basic function of the human brain subserved by an intrinsic architecture composed of two
hierarchical functional gradients. Our framework, based on a synthesis of multimodal and
multiscale evidence across species, begins to sketch a coherent, neurobiologically inspired
research program suggesting that predictive energy regulation is at the core of human brain
fonction, and by extension, psychological and behavioral phenomena, providing a shared
vocabulary for theory building and knowledge accumulation.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
A growing consensus in neuroscience is that progress in understanding the nature of mind and
behavior must begin by seriously considering the evolution and development of the human
brain (par exemple., Cisek, 2019, 2021). This perspective article extends hypotheses found in our earlier
work suggesting that one core function of a brain is to efficiently coordinate and regulate the
energetic requirements of its body, consistent with evidence from vertebrate brain evolution,
embryological development, and signal processing (Gee, 2018; Sterling & Laughlin, 2015;
Citation: Katsumi, Y., Theriault, J.. E.,
Quigley, K. S., & Barrett, L. F. (2022).
Allostasis as a core feature of
hierarchical gradients in the human
brain. Neurosciences en réseau, 6(4),
1010–1031. https://est ce que je.org/10.1162
/netn_a_00240
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00240
Reçu: 1 Octobre 2021
Accepté: 11 Février 2022
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Corresponding Authors:
Yuta Katsumi
ykatsumi@mgh.harvard.edu
Lisa Feldman Barrett
l.barrett@northeastern.edu
Éditeur de manipulation:
Olaf Sporns
droits d'auteur: © 2022
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
Allostasis as a core feature of hierarchical gradients in the brain
Allostasis:
The process of activating
physiological systems (tel que
hormonal, autonomic, or immune
systèmes) with the aim of returning the
body to homeostasis.
Allocortex:
Part of the cerebral cortex with
the simplest structure (two or
three layers). It comprises the
hippocampus and the primary
olfactory cortex (part of the cerebral
cortex that receives the projection
from the olfactory bulb).
Striedter & Northcutt, 2020). Spécifiquement, we synthesize theoretical and empirical evidence
emerging from two streams of literature: (un) hierarchical gradients of functional connectivity
that reflect the brain’s large-scale structural architecture (Par exemple, a lamination gradient
in the cerebral cortex; par exemple., Barbas, 2015; Hilgetag & Goulas, 2020; Margulies et al., 2016;
Zhang et al., 2019), et (b) predictive regulation of the body’s energy resources, called
allostasis (Schulkin & Sterling, 2019; Sennesh et al., 2021; Sterling, 2012; Sterling & Laughlin,
2015). Allostasis is the process by which the brain anticipates the needs of the body and
attempts to meet those needs before they arise, and is one specific instantiation of a broader
predictive processing framework that has developed to understand motor movements, percep-
tion, cognitions, emotions, and even consciousness (par exemple., Clark, 2013, 2016; Denève & Jardri,
2016; Friston, 2010; Friston et al., 2017; Hohwy, 2013; Hohwy & Seth, 2020; Hutchinson &
Barrett, 2019; Keller & Mrsic-Flogel, 2018; McNamee & Wolpert, 2019; Rao & Ballard, 1999;
Seth, 2015).
Building on prior theorizing and evidence on allostasis from our group and others (Barrett,
2017; Barrett & Simmons, 2015; Hutchinson & Barrett, 2019; Khalsa et al., 2018; Kleckner
et coll., 2017; Owens et al., 2018; Petzschner et al., 2021; Petzschner et al., 2017; Pezzulo
et coll., 2015; Pezzulo et al., 2021; Schulkin & Sterling, 2019; Seth & Friston, 2016; Seth &
Tsakiris, 2018; Stephan et al., 2016), as well as on papers dealing more generally with predic-
tive processing in relation to bodily regulation or bodily sensing (par exemple., Ainley et al., 2016;
Allen, 2020; Allen & Friston, 2018; Allen et al., 2019; Seth, 2013; Seth et al., 2012; Forgeron
et coll., 2017), we propose that two of the large-scale functional gradients identified in multiple
brain structures—in the cerebral cortex, including gradients in the isocortex and in the hippo-
campus (c'est à dire., allocortex), as well as in the cerebellum—can be meaningfully interpreted as an
intrinsic neural architecture that supports predictive processing, including allostasis. Our anal-
ysis extends the literature on bodily regulation by suggesting that allostasis is a whole-brain
phenomenon, rather than attempting to localize it to a small set of brain regions. Our frame-
work also extends the literature on hierarchical gradients by proposing that they may serve
domain-general functions in the brain, offering an opportunity to understand how cognition,
emotion, perception, and other psychological phenomena might emerge from a common set
of computational ingredients. Identifying allostasis as a key element in the state space of a
brain, which exists as a complex, nonlinear dynamical system that continually interacts with
its body and the surrounding world, offers new opportunities to build a unified science of
brain, body, and mind.
THE BRAIN’S INTRINSIC SYSTEM FOR ALLOSTASIS
Evolutionary, developmental, and anatomical studies of the vertebrate brain all suggest that its
fundamental job is to efficiently regulate the body’s internal systems as an animal navigates its
environmental niche. Predictive regulation is an improvement over reaction because reactive
systems adapt only in the face of error, but any mistake is potentially fatal (Sterling, 2012; Sterling
& Laughlin, 2015). Prediction also limits the extent to which incoming signals need to be
encoded (Shannon & Weaver, 1949/1964), which may save the metabolic costs of learning pre-
dictable information (Sengupta et al., 2013; Theriault, Shaffer, et coll., 2021; Theriault, Jeune,
et coll., 2021). Across the expanse of time, vertebrates evolved larger bodies, making new biolog-
ical systems necessary (Gee, 2018; Striedter & Northcutt, 2020). These include systems for waste
disposal, nutrient dispersal (par exemple., renal system, respiratory system, cardiovascular system), comme
well as systems to sense their expanding niche (par exemple., vision, audition, olfaction). As biological
systems proliferated, so did the need grow for a brain to actively coordinate and regulate them.
Accordingly, rudimentary neurons in a ganglion that sat atop the spinal cord became brain stem
Neurosciences en réseau
1011
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
.
/
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
structures (par exemple., the optic tectum, or the superior colliculi in mammals; the ventral hypothal-
amus), and eventually, novel structures such as the diencephalon and the telencephalon
(including the cerebral cortex) emerged (Gee, 2018; Striedter & Northcutt, 2020). Embryolog-
ical development is consistent with this evolutionary narrative, in that the brain, visceral sys-
thèmes, and exteroceptive systems all arise from adjacent locations on the neural plate (Feinberg
& Mallatt, 2013; Nieuwenhuys & Puelles, 2016). This narrative, and the embryological evi-
dence, suggests that the regulation of the body is a core consideration for brain evolution,
brain development, and brain function in organisms.
The viability of this hypothesis is bolstered by tract-tracing and cytoarchitectonic evidence
from mammalian brains (par exemple., Barbas, 2015; Carmichael & Prix, 1996; Evrard et al., 2014;
Mufson & Mesulam, 1982; Öngür et al., 2003; Vogt & Pandya, 1987; Vogt et al., 1987). Nous
summarized these findings in a recent paper showing their congruence with functional connec-
tivity of the human brain “at rest,” while the brain is not being probed by exogenous stimuli (par exemple.,
in an experimental task) but is still attached to body and regulating its internal systems (Kleckner
et coll., 2017). In primates and other mammals, cortical regions known as limbic cortices form a
ring around the thalamus, hypothalamus, and some brainstem regions (Willis, 1664) on the
medial wall of each hemisphere continuing into temporal and orbitofrontal cortices (Barbas,
2015; Chanes & Barrett, 2016). Limbic cortices send monosynaptic projections to subcortical
(par exemple., hypothalamus) and brainstem (par exemple., periaqueductal gray, parabrachial nucleus, nucleus of
the solitary tract) structures that regulate the internal systems of the body, including the
autonomic nervous system, immune system, and endocrine system. Limbic cortices traditionally
refer to the hippocampus, amygdala, olfactory cortex, and the most medial portions of the
cingulate cortices (and in some papers, portions of the basal ganglia). In some nomenclatures
(par exemple., Barbas, 2015), the “limbic” designation also includes areas that elsewhere are referred to
paralimbic regions of the brain, which are zones of cytoarchitectural transition between allocor-
tical tissue and more prototypic isocortex, such as ventral anterior insula/posterior orbitofrontal
cortex, temporal pole, cingulate cortices, and entorhinal cortex (see Mesulam, 2000). Dans ce
papier, we follow Barbas (2015) and refer to the entire ensemble as “limbic” (for a history of
the “limbic” concept in neuroanatomy, see Lautin, 2001; Roxo et al., 2011).
We identified homologous locations of these limbic cortices in the human brain based on
the coordinates from previous neuroimaging studies. Using functional magnetic resonance
imaging (IRMf) data collected at rest in more than 500 participants (split into discovery and
replication samples), we computed whole-brain functional connectivity maps for each cortical
limbic region, estimated as correlations in time courses of low-frequency blood oxygen level–
dependent (AUDACIEUX) signals for the voxels in each region with the voxels in the rest of the brain
(Kleckner et al., 2017). An unsupervised clustering analysis of both discovery and replication
maps revealed two spatially overlapping ensembles that are commonly referred to as the
“default mode network” and the “salience network,” which constitute the brain’s intrinsic sys-
tem for allostasis (Chiffre 1).
The interpretation of the default mode and salience networks and their overlap as an intrin-
sic allostatic neural system is consistent with connectivity profiles of the two networks. Le
cortical limbic nodes in both networks have extensive functional connectivity with subcortical
and brainstem structures that are thought to be important for allostasis (Figure 1C). These
limbic nodes also exhibit connectivity with the cerebellum and the hippocampus, consistent
with tract-tracing evidence identifying their connections to the structures that process outgoing
visceromotor and incoming interoceptive signals (Pisano et al., 2021; Suarez et al., 2018;
Vertes, 2015; Zhu & Wang, 2008). Functional imaging evidence also implicates activity in
the default mode and salience networks with bodily regulation. Par exemple, cardiac activity
Limbic cortices:
Part of the isocortex with agranular or
dysgranular structure. They are
sometimes referred to as
periallocortex (agranular) et
proisocortex (dysgranular) cortex.
Visceromotor cortices:
Agranular isocortical regions that
modulate the regulation of the
autonomic nervous system as well as
of the hormonal and immune
systèmes.
Neurosciences en réseau
1012
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
Chiffre 1. A schematic summary of findings from Kleckner et al. (2017) illustrating the intrinsic neural system for allostasis. (UN) Yellow high-
lights indicate areas of the cerebral cortex corresponding to the intrinsic allostatic system. (B) The intrinsic allostatic system consists of two
overlapping intrinsic functional networks, which are conventionally called the “default mode network” and the “salience network.” (C) Hip-
pocampal and subcortical connectivity of the two functional networks constituting the allostatic system; colors correspond to those used in
panel B; X, oui, and z values correspond to the MNI coordinates in millimeters. Panel A was modified from the figure originally published in
https://dana.org/article/interoception-the-secret-ingredient/.
at rest correlates with low-frequency BOLD signal fluctuations in these networks (Valenza
et coll., 2019), and tasks examining autonomic regulation of the cardiovascular system consis-
tently elicit activation within their cortical nodes and subcortical extents (Beissner et al., 2013;
Gianaros & Wager, 2015).
There are several notable observations that can be drawn from the intrinsic allostatic sys-
tem that provide opportunities for novel discoveries in the growing field of allostatic regula-
tion. D'abord, as the default mode and salience networks maintain allostasis, they play a role in
modeling the interoceptive consequences of allostatic regulation. Recent work suggests that
successful allostasis may require the predictive construction of interoceptive signals (antici-
pation of physiological changes in the body due to some future behavior; Barrett, 2017; Gu &
FitzGerald, 2014; Pezzulo et al., 2015). Regions in the default mode and salience networks
have been discussed in computational models of allostasis and other forms of bodily regula-
tion as well as interoception (Ainley et al., 2016; Allen, 2020; Allen et al., 2019; Barrett &
Simmons, 2015; Khalsa et al., 2018; Owens et al., 2018; Petzschner et al., 2017; Pezzulo
et coll., 2015; Seth, 2013; Seth & Friston, 2016; Seth et al., 2012; Smith et al., 2017; Stephan
et coll., 2016). These models propose that visceromotor regions (par exemple., anterior insula, anterior
midcingulate cortex, and subgenual anterior cingulate cortex) are broadly involved in issuing
signals to infer the causes of interoceptive signals and predict their trajectories into the future.
Some of these models consider regions that are part of the default mode network to be higher
in the processing hierarchy than regions of the salience network in the brain. Spécifiquement, le
former is hypothesized to infer the meaning of interoceptive signals based on past experience
(Smith et al., 2017) or issue predictions to visceromotor regions within the salience network
based on the brain’s beliefs about its capacity to successfully perform allostasis (c'est à dire., “meta-
cognition”; Stephan et al., 2016). These views are overall consistent with our framework in
which agranular areas (c'est à dire., an isocortical region with neurons that configure into a relatively
undifferentiated superficial layer [corresponding to layers II and III] and lacking a fully differ-
entiated layer IV) in the default mode network are thought to be crucial for the initiation of
Interoception:
The perception and integration of
autonomic, hormonal, visceral, et
immunological homeostatic signals
that collectively describe the
physiological state of the body.
Agranular cortex:
An isocortical region with neurons
that configure into a relatively
undifferentiated superficial layer
(corresponding to layers II and III)
and lacking a fully differentiated
layer IV.
Neurosciences en réseau
1013
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
prediction signals that help give meaning to incoming sensations by generalizing from similar
past experiences, which we have described as a process of continual ad hoc category con-
struction (see Prediction as a domain-general computational process in the brain, below).
The conceptual categories that are constructed as prediction signals constitute the brain’s
internal model of its body in the world (Barrett, 2017; Kleckner et al., 2017).
Deuxième, modeling of the sensory consequences of allostatic regulation also means model-
ing the expected changes in exteroceptive signals. C'est, the brain does not detect signals in
the world, but it models the features of those signals as they are transduced by the sensory
surfaces of the body. Some of these features are high-dimensional (closer in detail to the signals
from the sensory surfaces) and some are lower dimensional, compressed summaries (abstract
features) such as affective valence and arousal or other “psychological” features. Plus loin, to the
extent that interoceptive signals (as modeled by the brain) act as control signals for allostasis
(Sennesh et al., 2021), those signals may also play a role in the sampling of unanticipated
exteroceptive signals (c'est à dire., exteroceptive prediction errors). This hypothesis is supported by evi-
dence that sampling of visual, auditory, and other data from the sensory surfaces of the body is
statistically related to bodily signals such as heartbeats and respiration (par exemple., Al et al., 2020;
Aspell et al., 2013; Galvez-Pol et al., 2020; Grund et al., 2022; Kluger et al., 2021; Kunzendorf
et coll., 2019; Zelano et al., 2016).
Troisième, and relatedly, these findings suggest that allostasis may play a role in learning (c'est à dire.,
processing of prediction errors). Dorsal mid to posterior insula, which functions as primary inter-
oceptive cortex (Avery et al., 2015; Nieuwenhuys, 2012), is a point of overlap for the default
mode and salience networks (Figure 1B), suggesting that processing of unanticipated interocep-
tive and exteroceptive signals (c'est à dire., prediction errors) may be influenced by their predicted allo-
static relevance. This hypothesis is supported by observations that limbic cortices in both
networks show connectivity with brainstem nuclei that give rise to the neuromodulators
involved in attention and neural excitability (par exemple., ventral tegmental area, substantia nigra, dorsal
raphe nucleus, and locus coeruleus; Bär et al., 2016; Prix & Drevets, 2010). These findings sug-
gest that attention and levels of consciousness may also be influenced by allostatic regulation.
Fourth, further extending this hypothesis, our view is that allostasis is always operating con-
stantly regardless of whether an animal is awake (active) or at rest. A brain must always coor-
dinate internal bodily systems and anticipate upcoming energy needs (Schulkin & Sterling,
2019), although at rest, those needs differ from the energic needs during active periods. Pour
example, in contrast to active periods, digestion, waste secretion, and immune functions con-
tinue during rest (either asleep or quiescent). Plus loin, the primary source of metabolic fuel also
changes from active to resting periods (c'est à dire., a shift from greater carbohydrate utilization to
greater lipid utilization, respectivement), and during quiescent/dark periods, physiological pro-
cesses such as genomic replication that are especially sensitive to disruption by UV light
are prioritized (Asher & Schibler, 2011; Gerhart-Hines & Lazar, 2015). It has been suggested
that circadian cycling provides an important means by which allostatic (predictive) regulation
of metabolism can occur (Asher & Schibler, 2011), and there is an intimate and evolutionarily
long-standing coordination of circadian and metabolic control across tissues (par exemple., cardiac
muscle, skeletal muscle, gut microbiome; reviewed in Gerhart-Hines & Lazar, 2015) to mobi-
lize resources to the body’s internal systems where they would be needed the most (Schulkin
& Sterling, 2019). En outre, when the brain is perturbed with fewer exteroceptive prediction
errors (par exemple., in sleep), the intrinsic allostatic system may help refine the brain’s internal model
of its body in the world, par exemple, by removing redundancies in the model and thus reduc-
ing its complexity or generating “fictive” prediction error signals to train the model (for similar
arguments, see Barron et al., 2020; Pezzulo et al., 2021).
Neurosciences en réseau
1014
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
.
/
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
Enfin, the allostatic system may play a pivotal role in signal integration in the brain more
generally. The default mode and salience networks overlap in and contain the highest propor-
tion of “rich-club” hubs, defined as regions showing the densest anatomical connections
within the cerebral cortex. These rich-club hubs are interpreted as the brain’s backbone for
their central role in neural communication and synchrony (van den Heuvel & Sporns, 2011,
2013). Rich-club hubs that are limbic in structure (vs. non-limbic) exhibit topological proper-
ties more suited to function as “high-level” connectors, integrating already highly integrated
information across modules or communities of regions (Zhang et al., 2020). Taken together,
these findings support the existence of an intrinsic allostatic system in humans that is anatom-
ically central and contributes to information integration and coordination in the entire brain as
well as between the brain and the body.
PREDICTION AS A DOMAIN-GENERAL COMPUTATIONAL PROCESS IN THE BRAIN
That allostasis is one of the brain’s core tasks is further supported by converging evidence for
predictive processing models about bodily regulation and/or interoception. A variety of spe-
cific proposals abound (Ainley et al., 2016; Allen & Friston, 2018; Allen et al., 2019; Hohwy &
Seth, 2020; Hutchinson & Barrett, 2019; Khalsa et al., 2018; Owens et al., 2018; Parr et al.,
2018; Petzschner et al., 2021; Petzschner et al., 2017; Pezzulo et al., 2015; Pezzulo et al.,
2021; Schulkin & Sterling, 2019; Seth, 2013; Seth & Friston, 2016; Seth et al., 2012; Seth &
Tsakiris, 2018; Smith et al., 2021; Smith et al., 2017; Stephan et al., 2016), but they are united
by three components that are thought to be implemented in a hierarchical arrangement in the
brain’s architecture: (un) prediction signals that the brain generatively constructs using
memory—or alternatively, an “internal model” (par exemple., Berkes et al., 2011), “top-down” process-
ing (par exemple., Friston, 2010; Jordan & Keller, 2020; Rao & Ballard, 1999), a “forward model” (par exemple.,
Wolpert et al., 1998), or “feedback” signals (par exemple., Lamme & Roelfsema, 2000); (b) prediction
errors (or “bottom-up” processing, or “feedforward” signals) that encode the differences
between predicted sensory inputs and incoming sense data from the body’s sensory surfaces;
et (c) precision signals (or attention signals or executive control) that modulate the strength
and durability of predictions and prediction errors, and their ability to access motor control and
influence behavior (Feldman & Friston, 2010; Kanai et al., 2015). Prediction errors are potential
teaching signals, but their capacity to update predictions is thought to depend on how they are
weighted by precision signals, which are interpreted as the predicted value of the allostatic infor-
mation they will provide, or “salience” (Barrett, 2017; see also Parr & Friston, 2019, for a discus-
sion of salience). Prediction signals are also thought to be weighted by their estimated value to
explain the incoming sense data, which may correspond to their estimated prior probabilities
(Barrett, 2017; Feldman & Friston, 2010; Kanai et al., 2015). The advantage of predictive pro-
cessing for an organism is that learning accumulates to build an internal model of its body in the
monde, from which top-down predictions can be made and allostasis can be achieved.
It has been proposed that structural properties of the cerebral cortex support the flow of pre-
diction and prediction error signals. In macaque monkeys and other mammals, over 30 years of
tract-tracing evidence demonstrates a cytoarchitectural substrate for this flow of signals, où
predictions flow according to granular development (Chiffre 2; Barbas, 2015; García-Cabezas
et coll., 2019). Granular (or eulaminate) cortices have six definable cell layers, dysgranular areas
have only a rudimentary layer IV, and agranular areas lack it altogether (Barbas, 2015; García-
Cabezas et al., 2019). En outre, the size and connectivity of pyramidal neurons are different,
with granular cortices made up of many small neurons with few connections, whereas agranu-
lar cortices contain fewer, larger neurons with many more connections (for a discussion, voir
Finlay & Uchiyama, 2015). The central observation is that limbic cortices are either agranular or
Granular cortex:
An isocortical region with six
differentiated layers, including a
well-defined layer IV that contains
many stellate granule cells receiving
thalamocortical inputs.
Dysgranular cortex:
Part of the isocortex with a
rudimentary layer IV.
Neurosciences en réseau
1015
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
.
t
/
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
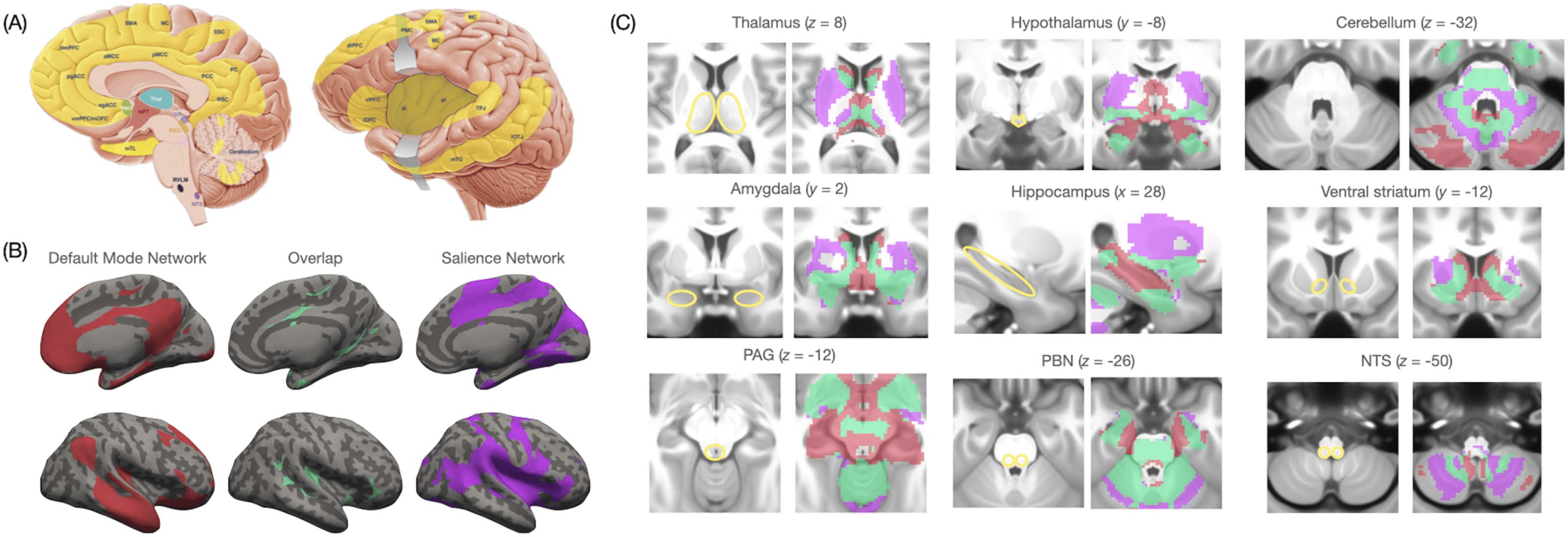
Chiffre 2. Hypothesized flow of prediction and prediction error signals in the human brain. (UN) Information flow between cortical regions is
predicted by the relative degree of laminar development in the communicating columns. Prediction signals flow from deep layers of less
granular cortices (par exemple., agranular areas with undifferentiated layers II and III and no layer IV) and terminate in superficial layers of more gran-
ular cortices (par exemple., dysgranular/granular cortices, where layers II and III are differentiated and layer IV is rudimentary [dysgranular] or well-
defined [granular]). Prediction error signals flow in the opposite direction, from superficial layers of more granular cortices to deep layers of less
granular cortices (Barbas, 2015; García-Cabezas et al., 2019; as discussed in Barrett, 2017; Hutchinson & Barrett, 2019). (B) This cytoarchi-
tectural gradient is thought to support lossy information compression in the cerebral cortex. C'est, sensory prediction errors ascend the
cortical hierarchy from superficial layers of (granular) sensory regions, first flowing to (less granular) heteromodal and motor cortex (Barbas
& García-Cabezas, 2015) to dysgranular/agranular limbic regions involved in visceromotor control. As sensory prediction errors ascend along
this laminar gradient, high-dimensional sensory signals are compressed and reduced in dimensionality (Barrett, 2017; Finlay & Uchiyama,
2015; for a related view, see Bastos et al., 2020) and become multimodal summaries (par exemple., Braga et al., 2013; Sepulcre et al., 2012; Szinte
& Knapen, 2020). The signals in limbic regions, alors, may implement some of the most compressed, multimodal representations in the cere-
bral cortex that guide the formation of prediction signals. Prediction signals may ultimately control action and construct perception and sub-
jective experience (Allen & Friston, 2018; Barrett, 2017; Fotopoulou & Tsakiris, 2017; Khalsa et al., 2018; Owens et al., 2018; Petzschner
et coll., 2017; Sennesh et al., 2021; Seth & Friston, 2016; Seth & Tsakiris, 2018; Stephan et al., 2016). Figure adapted from Hutchinson and
Barrett (2019), with permission.
dysgranular in laminar organization, and are thought to function as the most powerful feedback
(c'est à dire., prediction) regions in the brain (Joyce & Barbas, 2018). C'est, prediction signals stem
from the same neurons as those that are responsible for allostatic regulation of the body
(Chiffre 1).
In prior work (Barrett, 2017; Barrett & Simmons, 2015; Chanes & Barrett, 2016), nous
hypothesized that prediction signals begin as visceromotor control signals in agranular
and dysgranular limbic cortices, all of which are located in the default mode and salience
networks of the intrinsic allostatic system (par exemple., subgenual anterior cingulate cortex, entorhi-
nal cortex, anterior midcingulate cortex, ventral anterior insula/posterior orbitofrontal cortex).
Prediction signals descend from the deep layers of these cortices to subcortical and brain
stem nuclei, which proceed to the spinal cord to coordinate and regulate the body’s internal
systèmes. En même temps, collateral axons carry efferent copies of visceromotor predictions
both to primary motor cortex (as skeletomotor prediction signals) and to primary interocep-
tive, visuel, auditory, and somatosensory cortices as sensory prediction signals, either
directly or via polysynaptic connections (for similar views of skeletomotor efferent signals,
see Adams et al., 2013; Bastos et al., 2012). Primary motor cortex has a definable layer IV,
but it is less well developed than most primary sensory areas (Barbas & García-Cabezas,
2015), meaning that primary motor cortex is hypothesized to send sensory prediction signals
(Barrett, 2017). The same is true of primary interoceptive cortex, which has less laminar dif-
ferentiation and likely sends sensory prediction signals to primary visual, auditory, et
Neurosciences en réseau
1016
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
t
/
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
somatosensory cortices (Barrett, 2017; Chanes & Barrett, 2016). The exceptions to this
hypothesis are primary olfactory and gustatory cortices, which are dysgranular in laminar
organization, and therefore are at the same level of the predictive hierarchy as the viscer-
omotor regions issuing allostatic control signals.
In this view, behavior and mental events involve a coordination of predictions: visceromo-
tor predictions that regulate the internal milieu to make energetic resources available for ske-
leotomotor movements and experience, skeletomotor prediction signals that prepare the body
for movement, interoceptive prediction signals related to affective features (c'est à dire., faible-
dimensional representation of interoceptive sensations; Barrett & Bliss-Moreau, 2009), et
exteroceptive sensory prediction signals that prepare the sensory surfaces of the body to
receive upcoming sensory signals. Incoming interoceptive and exteroceptive signals either
confirm or constrain these predictions. Interoceptively, ascending viscerosensory signals are
carried along the vagus nerve and small diameter C and Aδ fibers (Craig, 2002), via various
thalamic nuclei, and salient, unanticipated sensory inputs will be encoded as interoceptive
prediction errors (par exemple., reward or aversive prediction errors; Seth et al., 2012). Salient extero-
ceptive signals arriving from the sensory surfaces are also encoded as prediction errors, lequel,
together with interoceptive prediction errors, modify the internal model and future predictions
that constitute skeletomotor and visceromotor action plans, optimized to deal with particular
sensory events. Both prediction and prediction error signals are subject to modulation by pre-
cision signals. This suggests that, from birth to death, the intrinsic allostatic system may be
sending a cascade of reference signals to the body and predict the sensory consequences of
those reference signals as sensory prediction errors that ascend the hierarchical arrangement of
neurons along the cerebral cortex.
HIERARCHICAL GRADIENTS IN THE CEREBRAL CORTEX AND THEIR ROLE IN
PREDICTIVE PROCESSING
The hypotheses discussed so far propose that signal propagation in predictive processing is
coordinated along a cytoarchitectural gradient in the cerebral cortex. Recent neuroimaging
research examining intrinsic functional connectivity in the human brain suggests that the
cerebral cortex is functionally organized along multiple gradients (Bethlehem et al., 2020;
Margulies et al., 2016; Paquola et al., 2020; Paquola et al., 2019; Shafiei et al., 2020). In a
recent series of studies, we have proposed that two of these gradients represent components
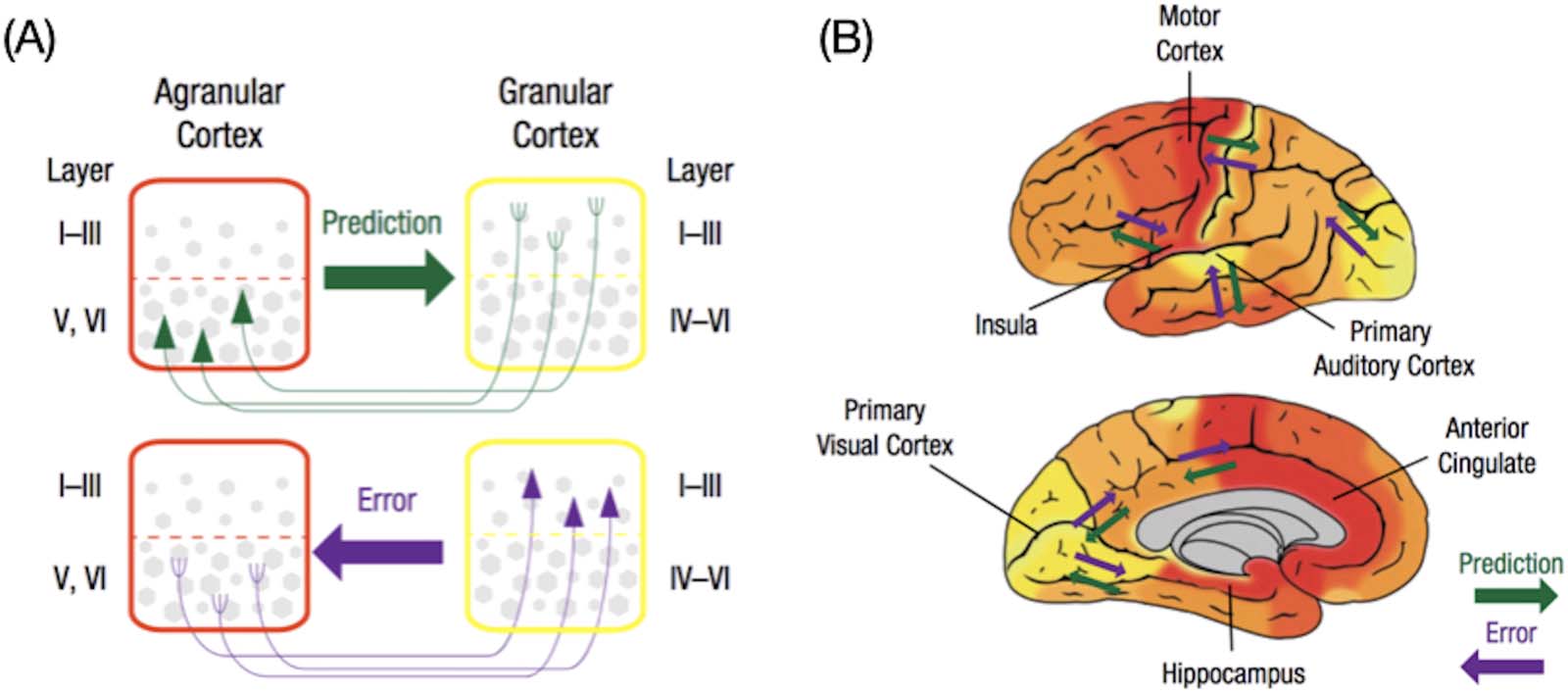
of predictive processing (Katsumi et al., 2021; Zhang et al., 2019).
Typiquement, functional connectivity gradients are computed by constructing an affinity matrix,
which describes the similarity of connectivity profiles in a set of data points (par exemple., voxels, sur-
face vertices, or parcels). A dimensionality reduction technique is then used to decompose this
matrix into principal eigenvectors describing axes of largest variance (for additional technical
details, see Vos de Wael et al., 2020). Each eigenvector can be used to identify gradual tran-
sitions in the pattern of functional connectivity within a given structure, thus yielding a con-
nectivity “gradient.” We have recently proposed that two of the dominant and commonly
identified connectivity gradients in the cerebral cortex are consistent with the role of cortical
ensembles in predictive processing (as hypothesized in Barrett, 2017; Chiffre 3), which are
discussed in detail below.
The principal gradient (explaining the largest amount of variance in connectivity profiles) est
anchored at one end by the default mode and “frontoparietal” networks, and at the other end
by the exteroceptive sensory and motor (par exemple., somatomotor, visuel) as well as salience net-
travaux. Ici, we refer to this gradient as the association-sensorimotor gradient, although it
(Functional) connectivity gradients:
Low-dimensional representations of
similarity in connectivity profiles
across a set of data points (par exemple.,
voxels, surface vertices, or regions of
interest).
Neurosciences en réseau
1017
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
t
.
/
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
Chiffre 3. Two of the dominant functional connectivity gradients identified in the cerebral cortex. (UN) For each connectivity gradient, le
proximity of colors can be interpreted as greater similarity of connectivity patterns between a pair of cortical vertices. (B) Boxplots show
medians and interquartile ranges of gradient values in canonical functional networks (Yeo et al., 2011), ordered by median values. Conven-
tional network labels are used here, but note that “default” and “limbic” networks are not always distinguished in the literature (par exemple., Kong
et coll., 2019) and both contain agranular, limbic tissue (Kleckner et al., 2017). (C) A scatterplot illustration of the two connectivity gradients.
Colors correspond to those used in panel B to depict functional networks.
has also been called a “transmodal-unimodal” gradient (par exemple., Margulies et al., 2016). Converg-
ing evidence from neuroimaging, histological, transcriptomic, and electrophysiological studies
identifies this gradient as a dominant axis of feature organization in the cerebral cortex, haut-
lighting its role as a domain-general organizational motif (reviewed in Sydnor et al., 2021). Nous
propose that this gradient can be interpreted through the lens of predictive processing, tel
that it separates regions involved in representing prediction signals on one end, and prediction
error signals on the other.
At the “association” end, the default mode network is thought to construct highly com-
pressed multimodal representations, which enable the initiation of prediction signals that con-
stitute the brain’s internal model of its body in the world, guiding motor actions and making
perception possible (Barrett, 2017; Buckner, 2012; Smith et al., 2017; Stawarczyk et al., 2021).
Topographical features unique to the default mode network may facilitate these computations:
Regions of the default mode network are, in certain cases, multiple synapses from primary
exteroceptive sensory areas, allowing ample opportunity for multimodal compression—that
est, the construction of abstract features (for a similar view, see Chanes & Barrett, 2016; Margulies
et coll., 2016; Smallwood et al., 2021). This is consistent with the role of this network in conceptual
traitement (Fernandino et al., 2016) and ad hoc category construction that gives meaning to
sensory inputs (as discussed in Barrett, 2017).
The frontoparietal network is thought to have two major roles: (un) estimating the precision of
prediction signals by suppressing predictions whose priors are very low, and when necessary,
(b) sculpting and maintaining predictions longer than the several hundred milliseconds it takes
to process imminent prediction errors (Barrett, 2017; see Smith et al., 2017, for a similar view).
This is consistent with other accounts of the frontoparietal network’s function that its subnet-
work acts as an extension of the default mode network and regulates complex introspective
processes (Dixon et al., 2018). This may also explain why the default mode and frontoparietal
Neurosciences en réseau
1018
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
t
.
/
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
networks exhibit longer timescales of cortical processing than any other functional network in
the cerebral cortex (Hasson et al., 2015; Raut et al., 2020).
At the “sensorimotor” end of the association-sensorimotor gradient, exteroceptive sensory
networks process sensory inputs that continually confirm or refine predictions made from the
brain’s internal model of its body in the world. The salience network is hypothesized to esti-
mate the precision of prediction error signals by altering the gain on prediction error signals as
they propagate from the sensory periphery, reflecting confidence in the reliability and quality
of incoming sensory information and its predicted relevance for allostasis. The salience net-
travail, alors, may help the brain adjust its internal model to the energetic conditions of the body
(Barrett, 2017). This proposal is consistent with accounts of salience network function in both
attention regulation (Power et al., 2011; Touroutoglou et al., 2012; Uddin, 2015; Ullsperger
et coll., 2014) and multisensory integration (Craig, 2009; Sepulcre et al., 2012), and with com-
putational models of interoception that highlight the role of salience network regions in esti-
mating the precision of ascending interoceptive prediction error signals (par exemple., Ainley et al.,
2016; Allen, 2020; Allen et al., 2019; Seth et al., 2012). Our interpretation of the
association-sensorimotor gradient provides an opportunity to integrate evidence from multi-
modal and multiscale approaches into a common framework. Spécifiquement, this functional gra-
dient is consistent with a cytoarchitectural gradient discussed above, which is hypothesized to
support the flow of prediction and prediction error signals (Chiffre 2). It is also overall consis-
tent with findings from recent studies investigating cortical myeloarchitecture, which identified
a similar principal gradient spanning primary sensory and limbic regions (Huntenburg et al.,
2017; Paquola et al., 2019).
The second gradient consistently identified in the literature is anchored at one end by the
default mode and exteroceptive sensory networks and at the other end by the salience and
frontoparietal networks. We refer to this gradient as a representation-modulation gradient, sep-
arating ensembles involved in the representation of low-dimensional multimodal summaries of
brain states or more precise sensorimotor signals (Fernandino et al., 2016) from those involved
in modulating these representations (par exemple., via attention regulation, goal maintenance, strategy
selection, or performance monitoring; Corbetta & Shulman, 2002; Dosenbach et al., 2007;
Miller & Cohen, 2001; Uddin, 2015). This gradient has also been described as a “multiple
demand” gradient (Genon et al., 2021; Paquola et al., 2020; Valk et al., 2021), as modulatory
networks are often engaged in the face of task-based cognitive demands (Assem et al., 2020;
Duncan, 2010; Fedorenko et al., 2013). Interpreted in terms of predictive processing, this gra-
dient distinguishes regions that represent prediction and prediction error signals from regions
that implement attentional modulation to compute the precision of these signals.
Another gradient commonly reported in studies of functional connectivity gradients is
anchored at one end by the visual network and at the other end by the somatomotor network
(Bethlehem et al., 2020; Margulies et al., 2016; Mckeown et al., 2020), suggesting a segrega-
tion of exteroceptive sensory systems. The anatomical and multiscale features of this gradient
are not yet clear and the functional implications of this segregation remain unknown. The role
of this gradient in predictive processing, donc, awaits further evidence about its anatom-
ical and functional features. Current evidence shows that this gradient appears dominant in
newborn infant brains, explaining the largest magnitude of variance in intrinsic functional con-
nectivity (Larivière et al., 2020). This gradient seems to remain dominant until the transition to
adolescence, at which point the association-sensorimotor gradient supersedes (Dong et al.,
2021). It is unclear what this means, cependant, given that both the visual and the somatomotor
networks are still in development during infancy and make substantial advances in neuroty-
pical development within the first year of life (Hadders-Algra, 2018; Johnson, 2013).
Neurosciences en réseau
1019
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
Integration of the visual-somatomotor gradient with the current predictive processing frame-
work is an opportunity for future research.
The relevance of the association-sensorimotor and representation-modulation gradients for
allostasis is further substantiated by multimodal evidence identifying similar gradients within
specific regions of the cerebral cortex. Spécifiquement, existing computational models of bodily
regulation (including allostasis) and interoception highlight the contribution of a functional
gradient within the insular cortex, such that the posterior and mid insula represents interocep-
tive (and exteroceptive) information, whereas the anterior insula integrates multimodal infor-
mation and exerts visceromotor control (Allen et al., 2019; Allen & Friston, 2018; Barrett &
Simmons, 2015; Smith et al., 2017; Stephan et al., 2016). Extending this view, a recent study
examining myeloarchitectural gradients within the entire insular cortex showed that the prin-
cipal gradient captures gradual transitions from the posterior to anterior insula; this gradient
corresponded with a shift in functional connectivity profiles from primarily sensorimotor to
greater affiliation with the salience network (Royer et al., 2020). This is consistent with the
cytoarchitectural gradient from granular (posterior) to agranular (anterior) insula, correspond-
ing to a hypothesized flow of prediction and prediction error signaling within this cortical area
(as discussed in Barrett & Simmons, 2015). This posterior–anterior insular gradient, alors,
appears to correspond to the cortical association-sensorimotor gradient, with posterior insula
at the sensorimotor end, and the anterior insula at the association end. En plus, le
second most dominant myeloarchitectural gradient in the insula showed transitions from the
posterior and ventral subregions to the dorsal anterior insula; this gradient corresponded
with a shift in functional connectivity patterns from uniquely sensorimotor to uniquely
modulatory/attentional (c'est à dire., salience, dorsal attention, and frontoparietal networks; Royer
et coll., 2020), suggesting the correspondence of this insular cytoarchitectural gradient with
the cortical representation-modulation gradient. This evidence is consistent with the hypoth-
esis that the precision of ascending sensory prediction errors is computed along intra-insular
gradients (Ainley et al., 2016; Allen, 2020; Allen et al., 2019) or more generally by the salience
réseau (as discussed in Barrett, 2017).
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
ROLE OF HIERARCHICAL HIPPOCAMPAL AND CEREBELLAR GRADIENTS IN
PREDICTIVE PROCESSING
Beyond the isocortex, the association-sensorimotor and representation-modulation gradients
also appear to govern the functional organization of other structures in the brain, tel que
the cerebellum (Guell, Schmahmann, et coll., 2018) and the hippocampus (c'est à dire., allocortex;
Vos de Wael et al., 2018). We built on these prior studies to assess the extent to which the
functional connectivity gradients in these structures corresponded with the two isocortical gra-
dients (Katsumi et al., 2021). In the cerebellum, the principal gradient captured a bilateral dis-
sociation of lobules IV, V, and VI and lobule VIII from the posterior part of Crus I and II and the
medial part of lobule IX, whereas the second dominant gradient distinguished bilaterally the
anterior parts of Crus I and Crus II along with lobule VIIb from the rest of the cerebellar cortex
(Figure 4A). In the hippocampus, the principal gradient revealed a functional dissociation
along the longitudinal axis, whereas the second most dominant gradient additionally captured
variation along the transverse (c'est à dire., medial-lateral) axis (Figure 4B).
To characterize these cerebellar and hippocampal connectivity gradients in terms of their
relations to the isocortex, we calculated intrinsic functional connectivity maps for each of these
isocortical structures weighted as a factor of voxel-wise gradient values. Par exemple, to char-
acterize how a given cerebellar gradient related to the isocortex, we computed a cerebello-
Neurosciences en réseau
1020
Allostasis as a core feature of hierarchical gradients in the brain
Chiffre 4. Functional connectivity gradients of the cerebellum and the hippocampus. Gradient-weighted functional connectivity maps rep-
resent, for a given gradient, the relationship between a given pair of structures in terms of their functional connectivity profiles. Par exemple,
voxels in Crus I and Crus II of the cerebellum anchoring one end of its Gradient 1 (depicted in red in a flat map above) showed relatively
greater (positive) functional connectivity with the default mode network in the isocortex than did cerebellar voxels anchoring the other end of
the same gradient.
isocortical connectivity map for each cerebellar voxel and multiplied it by the corresponding
gradient value for that particular voxel. In this way, the pattern of functional connectivity
between each cerebellar voxel and all isocortical vertices was weighted by the position of
the voxel on the cerebellar gradient. These voxel-wise, gradient-weighted cerebello-isocortical
connectivity values were summed over all cerebellar voxels, resulting in a single isocortical
projection of the cerebellar gradient. We repeated this procedure for each gradient derived
for the cerebellum and the hippocampus. This procedure allowed us to project cerebellar
and hippocampal gradients onto the isocortex, thus revealing the extent to which they spatially
corresponded with the isocortical association-sensorimotor and representation-modulation gra-
dients. We found that the principal cerebellar gradient strongly corresponded with the
association-sensorimotor gradient, whereas its second most dominant gradient corresponded
with the representation-modulation gradient. In contrast, in the hippocampus, the principal gra-
dient strongly corresponded with the representation-modulation gradient, whereas its second
most dominant gradient corresponded with the association-sensorimotor gradient (see cortical
surface maps in each panel, Chiffre 4).
The observed correspondence between connectivity gradients in the isocortex, the cerebel-
lum, and the hippocampus suggests a starting point for developing one unified, integrative
view of brain function, where allostatic regulation may be one of the core computational fea-
photos. This view extends earlier research examining computational capacities of these struc-
photos, which have been described in terms of predictive processing (Barron et al., 2020; Ito,
2008; Liu et al., 2018; Pezzulo et al., 2017; Wolpert et al., 1998). Converging empirical evi-
dence supports the hypothesis that both the cerebellum and the hippocampus are involved in
allostasis and interoception. The cerebellum exhibits direct or indirect anatomical connections
with various subcortical structures implicated in allostasis, including the hypothalamus, peri-
aqueductal gray, nucleus of solitary tract, and amygdala (Zhu & Wang, 2008). Of note, direct
and bidirectional connections between the cerebellum and the hypothalamus are thought to
be critical for the regulation of the body’s internal systems, including cardiovascular, respira-
tory, gastrointestinal, and immune systems (Zhu et al., 2006). The hippocampus also receives
Neurosciences en réseau
1021
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
rich inputs from subcortical structures, including the medial septum, amygdala, anterior tha-
lamic nuclei, supramammillary nucleus of the hypothalamus, and brain stem nuclei such as
ventral tegmental area, periaqueductal gray, and locus coeruleus (Amaral & Cowan, 1980;
Insausti & Amaral, 2012). En outre, the proportion of endocrine receptor expression in
le (mouse) hippocampus exceeds anything that has been observed in all comparable brain
regions with the exception of the hypothalamus (Lathe et al., 2020). These findings demon-
strate a clear link between hippocampal processing and allostatic concerns; par exemple, le
hippocampus may be key for the integration of interoceptive information with previous expe-
riences and exteroceptive sensory signals (Quigley et al., 2021).
It remains to be discovered to what extent, if any, functional gradients in the cerebellum
and the hippocampus map onto allostatic processing in the manner proposed for the gradients
of the isocortex. Although speculative, one possible hypothesis emerging from current evi-
dence is that the isocortex, the cerebellum, and the hippocampus might integrate over the
same information to modulate one another as they construct prediction, prediction error,
and precision signals in the service of allostasis, owing to their extensive connectivity with
one another and with the subcortical structures implicated in processing of these signals.
In sensorimotor coordination, Par exemple, the cerebellum has been traditionally thought to
estimate the sensory state of the body by anticipating the consequences of motor commands
(Shadmehr et al., 2010; Sokolov et al., 2017; Wolpert et al., 1998), possibly as a means to
compensate for delays in sensory feedback (Sultan et al., 2012; Tanaka et al., 2020)—for
instance, as signals physically travel from fingertips, through the periphery and spinal cord,
and into the cerebral cortex. Extrapolating from this evidence, we hypothesize that the cere-
bellum may compute sensory prediction errors to tune signals of various cortical ensembles
faster than sensory prediction errors computed in the cerebral cortex. This view is consistent
with the ability of granule cells (the majority of cerebellar neurons) to generate action poten-
tials that are relatively short-lived and at much higher frequencies than cerebral cortical neu-
rons (Sultan et al., 2012). Anatomical evidence has also identified polysynaptic interconnections
between the cerebellar cortex and nonprimary sensorimotor areas of the cerebral cortex (par exemple.,
parietal association, parahippocampal, occipitotemporal, and prefrontal cortices; Apps &
Watson, 2013; Kelly & Strick, 2003; Schmahmann, 1996), further supporting a domain-
general view of cerebellar influence on cortical processing.
The hippocampus is thought to generate prediction signals (Barron et al., 2020; Buzsáki &
Tingley, 2018; Lisman & Redish, 2009; Pezzulo et al., 2017) and facilitate reweighting of sig-
nals in the isocortex (Kumaran et al., 2016). En particulier, the hippocampus may help ensure
that the subsequent prediction signals generated based on the isocortical internal model are
not slaves to the statistics of the external sensory environment and instead more in line with the
goals of the animal (c'est à dire., weighted for the current and predicted conditions of the body’s inter-
nal environment; Kumaran et al., 2016). By interfacing with isocortical ensembles at many
levels of the predictive hierarchy, the hippocampus too may intervene at multiple points of
this hierarchy to modulate cortical signaling.
These hypotheses are consistent with the increasing consensus that the cerebellum (par exemple.,
Guell, Gabriela, et coll., 2018; King et al., 2019) and the hippocampus (par exemple., Grady, 2020) sont
functionally heterogeneous. The cerebellum and the hippocampus may serve domain-general
functions within a predictive hierarchy in the service of efficient bodily regulation, plutôt que
supporting particular functional domains exclusively (par exemple., related to motor coordination, epi-
sodic memory, or spatial cognition). This domain-general perspective may help advance both
an understanding of cerebello-isocortical (Shadmehr et al., 2010; Sokolov et al., 2017),
Neurosciences en réseau
1022
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
hippocampo-isocortical (Buzsáki & Tingley, 2018; Kumaran et al., 2016), and even cerebello-
hippocampal (par exemple., Babayan et al., 2017; Onuki et al., 2015; Watson et al., 2019) interactions,
and a computational understanding of their implications for various psychological processes.
Specific computational hypotheses concerning the role of hierarchical functional gradients
in predictive processing still need to be directly tested. Néanmoins, the existence of the
common axes of functional organization across brain structures suggests that it may be fruit-
ful to study the human brain organization in terms of domain-general functions in the service
of allostasis.
CONCLUSIONS
Allostasis is the brain’s predictive regulation of the body’s internal milieu (Sterling, 2012).
Coordination and regulation of the body’s internal systems has been hypothesized to be a
basic function of the brain, either as allostasis (Barrett, 2017; Barrett & Simmons, 2015;
Hutchinson & Barrett, 2019; Khalsa et al., 2018; Kleckner et al., 2017; Owens et al., 2018;
Petzschner et al., 2021; Petzschner et al., 2017; Pezzulo et al., 2015; Pezzulo et al., 2021;
Schulkin & Sterling, 2019; Seth & Friston, 2016; Seth & Tsakiris, 2018; Stephan et al., 2016) ou
as predictive regulation of the body more generally (Ainley et al., 2016; Allen & Friston, 2018;
Allen et al., 2019; Seth, 2013; Seth et al., 2012; Smith et al., 2017), and therefore may be a
basic feature of the mind. The brain’s modeling of the sensory consequences of allostasis, les deux
interoceptive and exteroceptive, may translate into consciously experienced feelings of
valence and arousal as basic features of consciousness (par exemple., Lindquist et al., 2016; Satpute
et coll., 2015; for discussion, see Barrett, 2017). In this paper, we connected these hypotheses
with hierarchical functional gradients that appear to organize whole-brain function (Katsumi
et coll., 2021; Zhang et al., 2019), which are grounded in biological structure (thus far estab-
lished for the cerebral cortex). Our claim is not that allostasis is the exclusive function of the
brain; rather, we suggest that behavior is always planned and executed—and mental events
always emerge—in the context of regulating the body’s internal systems, and the entire brain is
engaged in this regulatory function at the same time as it governs behavior and mental activity.
Although speculative, one final intriguing hypothesis emerging from this view is that all
psychological phenomena (par exemple., cognition, emotion, and perception) may be whole-brain phe-
nomena with allostatic features, rather than separate states arising from unique computations
that are localized to specific regions. This idea is consistent with a growing body of anatomical
and functional evidence. Par exemple, as mentioned above, exteroceptive sensory processing
is statistically associated with processing of bodily signals. Primary motor cortex contains vis-
ceromotor maps (Levinthal & Strick, 2012, 2020), suggesting intimate integration of skeleto-
motor and visceromotor functions. The anterior cingulate cortex (visceromotor cortex) sends
direct projections to neurons in V1 (Zhang et al., 2014), which may carry top-down prediction
signals (Leinweber et al., 2017). En effet, a substantial fraction of activity in the visual cortex
does not depend on incoming visual input (Keck et al., 2013; Muckli et coll., 2015), et le
majority of synapses in V1 originate from top-down sources (Sillito & Jones, 2002). Such evi-
dence runs counter to traditional assumptions that psychological functions can be uniquely
localized to specific brain regions or networks and is consistent with the hypothesis of a
domain-general computational architecture of the brain (par exemple., see Barrett, 2017, for how this
approach is applied to understanding the nature of emotions). This “whole-brain” view is
increasingly gaining empirical support in human neuroimaging studies that are designed to
be sensitive to such observations (par exemple., Gonzalez-Castillo et al., 2015; Gonzalez-Castillo
et coll., 2012; Liang et al., 2013) and in nonhuman animal research (reviewed in Kaplan &
Zimmer, 2020). An allostatically oriented whole-brain framework has the potential to unify
Neurosciences en réseau
1023
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
our understanding of brain, esprit, and body. Our approach offers the basis for a coherent,
neurobiologically inspired research program that attempts to explain how a variety of phys-
ical and mental events emerge from the same biological mechanisms. Data-driven
approaches reveal low-dimensional gradients that summarize the organizational features
du cerveau; biology will help us interpret them.
REMERCIEMENTS
The views, opinions, and/or findings contained in this review are those of the authors and shall
not be construed as an official Department of the Army position, politique, or decision, unless
so designated by other documents; nor do they necessarily reflect the views of the Elizabeth
R.. Koch Foundation. The authors thank Nada Kamona and Liz Cory for their assistance with
figure creation.
CONTRIBUTIONS DES AUTEURS
Yuta Katsumi: Enquête; Rédaction – ébauche originale. Jordan E. Theriault: Rédaction – révision &
édition. Karen S. Quigley: Rédaction – révision & édition. Lisa Feldman Barrett: Conceptualisation;
Acquisition de financement; Enquête; Surveillance; Rédaction – révision & édition.
INFORMATIONS SUR LE FINANCEMENT
Lisa Feldman Barrett, National Science Foundation, Award ID: BCS 1947972. Lisa Feldman
Barrett, the US Army Research Institute for the Behavioral and Social Sciences, Award ID:
W911NF-16-1-019. Lisa Feldman Barrett, Elizabeth R. Koch Foundation, Unlikely Collabora-
tors Fund. Lisa Feldman Barrett, National Institutes of Health (https://dx.doi.org/10.13039
/100000002), Award ID: R01 MH113234. Lisa Feldman Barrett, National Institutes of Health
(https://dx.doi.org/10.13039/100000002), Award ID: R01 MH109464. Lisa Feldman Barrett,
National Institutes of Health (https://dx.doi.org/10.13039/100000002), Award ID: U01
CA193632.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
.
t
/
RÉFÉRENCES
Adams, R.. UN., Shipp, S., & Friston, K. J.. (2013). Predictions not com-
mands: Active inference in the motor system. Brain Structure and
Function, 218(3), 611–643. https://doi.org/10.1007/s00429-012
-0475-5, PubMed: 23129312
Ainley, V., Apps, M.. UN. J., Fotopoulou, UN., & Tsakiris, M.. (2016).
‘Bodily precision’: A predictive coding account of individual
differences in interoceptive accuracy. Transactions philosophiques
of the Royal Society B: Sciences biologiques, 371(1708), 20160003.
https://doi.org/10.1098/rstb.2016.0003, PubMed: 28080962
Al, E., Iliopoulos, F., Forschack, N., Nierhaus, T., Grund, M.,
Motyka, P., Gaebler, M., Nikulin, V. V., & Villringer, UN. (2020).
Heart–brain interactions shape somatosensory perception and
evoked potentials. Proceedings of the National Academy of Sci-
ences, 117(19), 10575–10584. https://doi.org/10.1073/pnas
.1915629117, PubMed: 32341167
Allen, M.. (2020). Unravelling the neurobiology of interoceptive
inference. Tendances des sciences cognitives, 24(4), 265–266.
https://doi.org/10.1016/j.tics.2020.02.002, PubMed: 32160563
Allen, M., & Friston, K. J.. (2018). From cognitivism to autopoiesis:
Towards a computational framework for the embodied mind.
Synthese, 195(6), 2459–2482. https://doi.org/10.1007/s11229
-016-1288-5, PubMed: 29887647
Allen, M., Levy, UN., Parr, T., & Friston, K. J.. (2019). In the body’s
eye: The computational anatomy of interoceptive inference.
bioRxiv. https://doi.org/10.1101/603928
Amaral, D. G., & Cowan, W. M.. (1980). Subcortical afferents to the
hippocampal formation in the monkey. Journal of Comparative
Neurologie, 189(4), 573–591. https://doi.org/10.1002/cne
.901890402, PubMed: 6769979
Apps, R., & Watson, T. C. (2013). Cerebro-cerebellar connections.
En M. Manto, J.. D. Schmahmann, F. Rossi, D. L. Gruol, & N.
Koibuchi (Éd.), Handbook of the cerebellum and cerebellar
disorders (pp. 1131–1153). Springer Netherlands. https://doi.org
/10.1007/978-94-007-1333-8_48
Asher, G., & Schibler, U. (2011). Crosstalk between components of
circadian and metabolic cycles in mammals. Cell Metabolism,
13(2), 125–137. https://doi.org/10.1016/j.cmet.2011.01.006,
PubMed: 21284980
Aspell, J.. E., Heydrich, L., Marillier, G., Lavanchy, T., Herbelin, B.,
& Blanke, Ô. (2013). Turning body and self inside out: Visualized
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Neurosciences en réseau
1024
Allostasis as a core feature of hierarchical gradients in the brain
heartbeats alter bodily self-consciousness and tactile perception.
Sciences psychologiques, 24(12), 2445–2453. https://est ce que je.org/10
.1177/0956797613498395, PubMed: 24104506
Assem, M., Glasser, M.. F., Van Essen, D. C., & Duncan, J.. (2020). UN
domain-general cognitive core defined in multimodally parcel-
lated human cortex. Cortex cérébral, 30(8), 4361–4380. https://
doi.org/10.1093/cercor/bhaa023, PubMed: 32244253
Avery, J.. UN., Kerr, K. L., Ingeholm, J.. E., Burrows, K., Bodurka, J., &
Simmons, W. K. (2015). A common gustatory and interoceptive
representation in the human mid-insula. Human Brain Mapping,
36(8), 2996–3006. https://doi.org/10.1002/hbm.22823, PubMed:
25950427
Babayan, B. M., Watilliaux, UN., Viejo, G., Paradis, A.-L., Girard, B.,
& Rondi-Reig, L. (2017). A hippocampo-cerebellar centred net-
work for the learning and execution of sequence-based naviga-
tion. Rapports scientifiques, 7(1), 17812. https://est ce que je.org/10.1038
/s41598-017-18004-7, PubMed: 29259243
Bär, K.-J., de la Cruz, F., Schumann, UN., Koehler, S., Sauer, H.,
Critchley, H., & Wagner, G. (2016). Functional connectivity
and network analysis of midbrain and brainstem nuclei. Neuro-
Image, 134, 53–63. https://doi.org/10.1016/j.neuroimage.2016
.03.071, PubMed: 27046112
Barbas, H. (2015). General cortical and special prefrontal connec-
tion: Principles from structure to function. Annual Review of
Neurosciences, 38(1), 269–289. https://est ce que je.org/10.1146/annurev
-neuro-071714-033936, PubMed: 25897871
Barbas, H., & García-Cabezas, M.. Á. (2015). Motor cortex layer 4:
Moins c'est plus. Trends in Neurosciences, 38(5), 259–261. https://
doi.org/10.1016/j.tins.2015.03.005, PubMed: 25868984
Barrett, L. F. (2017). The theory of constructed emotion: An active
inference account of interoception and categorization. Social
Cognitive and Affective Neuroscience, 12(1), 1–23. https://est ce que je
.org/10.1093/scan/nsw154, PubMed: 27798257
Barrett, L. F., & Bliss-Moreau, E. (2009). Affect as a psychological
primitive. In Advances in experimental social psychology
(Vol. 41, pp. 167–218). Academic Press. https://est ce que je.org/10.1016
/S0065-2601(08)00404-8, PubMed: 20552040
Barrett, L. F., & Simmons, W. K. (2015). Interoceptive predictions in
the brain. Nature Revues Neurosciences, 16(7), 419–429. https://
doi.org/10.1038/nrn3950, PubMed: 26016744
Barron, H. C., Auksztulewicz, R., & Friston, K. (2020). Prediction
and memory: A predictive coding account. Progress in Neurobi-
ology, 192, 101821. https://doi.org/10.1016/j.pneurobio.2020
.101821, PubMed: 32446883
Bastos, UN. M., Lundqvist, M., Waite, UN. S., Kopell, N., & Miller, E. K.
(2020). Layer and rhythm specificity for predictive routing.
bioRxiv. https://doi.org/10.1101/2020.01.27.921783
Bastos, UN. M., Usrey, W. M., Adams, R.. UN., Mangun, G. R., Fries, P.,
& Friston, K. J.. (2012). Canonical microcircuits for predictive
coding. Neurone, 76(4), 695–711. https://est ce que je.org/10.1016/j
.neuron.2012.10.038, PubMed: 23177956
Beissner, F., Meissner, K., Bär, K.-J., & Napadow, V. (2013). Le
autonomic brain: An activation likelihood estimation meta-
analysis for central processing of autonomic function. Journal de
Neurosciences, 33(25), 10503–10511. https://est ce que je.org/10.1523
/JNEUROSCI.1103-13.2013, PubMed: 23785162
Berkes, P., Orbán, G., Lengyel, M., & Fiser, J.. (2011). Spontaneous
cortical activity reveals hallmarks of an optimal internal model of
l'environnement. Science, 331(6013), 83–87. https://est ce que je.org/10
.1126/science.1195870, PubMed: 21212356
Bethlehem, R.. UN. JE., Paquola, C., Seidlitz, J., Ronan, L., Bernhardt,
B., Cam-CAN Consortium, & Tsvetanov, K. UN. (2020). Dispersion
of functional gradients across the adult lifespan. NeuroImage,
222, 117299. https://doi.org/10.1016/j.neuroimage.2020
.117299, PubMed: 32828920
Braga, R.. M., Sharp, D. J., Leeson, C., Sage, R.. J.. S., & Leech, R..
(2013). Echoes of the brain within default mode, association,
and heteromodal cortices. Journal des neurosciences, 33(35),
14031–14039. https://doi.org/10.1523/ JNEUROSCI.0570-13
.2013, PubMed: 23986239
Buckner, R.. L. (2012). The serendipitous discovery of the brain’s
default network. NeuroImage, 62(2), 1137–1145. https://doi.org
/10.1016/j.neuroimage.2011.10.035, PubMed: 22037421
Buzsáki, G., & Tingley, D. (2018). Space and time: The hippocampus
as a sequence generator. Tendances des sciences cognitives, 22(10),
853–869. https://doi.org/10.1016/j.tics.2018.07.006, PubMed:
30266146
Carmichael, S. T., & Prix, J.. je. (1996). Connectional networks
within the orbital and medial prefrontal cortex of macaque mon-
keys. Journal of Comparative Neurology, 371(2), 179–207.
https://doi.org/10.1002/(SICI)1096-9861(19960722)371:2<179::
AID-CNE1>3.0.CO;2-#, PubMed: 8835726
Chanes, L., & Barrett, L. F. (2016). Redefining the role of limbic
areas in cortical processing. Tendances des sciences cognitives, 20(2),
96–106. https://doi.org/10.1016/j.tics.2015.11.005, PubMed:
26704857
Cisek, P.. (2019). Resynthesizing behavior through phylogenetic
refinement. Attention, Perception, and Psychophysics, 81(7),
2265–2287. https://doi.org/10.3758/s13414-019-01760-1,
PubMed: 31161495
Cisek, P.. (2021). An evolutionary perspective on embodiment. Dans
M.. D. Robinson & L. E. Thomas (Éd.), Handbook of embodied
psychologie: Pensée, feeling, and acting (pp. 547–571). Springer
International Publishing. https://doi.org/10.1007/978-3-030
-78471-3_24
Clark, UN. (2013). Quelle que soit la suite? Des cerveaux prédictifs, situated agents,
et l'avenir des sciences cognitives. Behavioral and Brain Sciences,
36(3), 1–24. https://doi.org/10.1017/S0140525X12000477,
PubMed: 23663408
Clark, UN. (2016). Surfing uncertainty: Prediction, action, et le
embodied mind. Presse universitaire d'Oxford. https://est ce que je.org/10
.1093/acprof:oso/9780190217013.001.0001
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed and
stimulus-driven attention in the brain. Nature Reviews Neurosci-
ence, 3(3), 201–215. https://doi.org/10.1038/nrn755, PubMed:
11994752
Craig, UN. D. (2002). How do you feel? Interoception: The sense of
the physiological condition of the body. Nature Reviews Neuro-
science, 3(8), 655–666. https://doi.org/10.1038/nrn894,
PubMed: 12154366
Craig, UN. D. (2009). How do you feel—now? The anterior insula
and human awareness. Nature Revues Neurosciences, 10(1),
59–70. https://doi.org/10.1038/nrn2555, PubMed: 19096369
Denève, S., & Jardri, R.. (2016). Circular inference: Mistaken belief,
misplaced trust. Current Opinion in Behavioral Sciences, 11, 40–48.
https://doi.org/10.1016/j.cobeha.2016.04.001
Neurosciences en réseau
1025
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
t
/
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
Dixon, M.. L., Vega, UN. D. L., Mills, C., Andrews-Hanna, J., Spreng,
R.. N., Cole, M.. W., & Christoff, K. (2018). Heterogeneity within
the frontoparietal control network and its relationship to the
default and dorsal attention networks. Actes du
Académie nationale des sciences, 115(7), E1598–E1607. https://
doi.org/10.1073/pnas.1715766115, PubMed: 29382744
Dong, H.-M., Margulies, D. S., Zuo, X.-N., & Holmes, UN. J..
(2021). Shifting gradients of macroscale cortical organization
mark the transition from childhood to adolescence. Proceed-
i n g s o f t h e N a t i o n a l A c a d e m y o f S c i e n c e s , 11 8 ( 2 8 ) ,
e2024448118. https://doi.org/10.1073/pnas.2024448118,
PubMed: 34260385
Dosenbach, N. U. F., Fair, D. UN., Miezin, F. M., Cohen, UN. L., Wenger,
K. K., Dosenbach, R.. UN. T., Fox, M.. D., Snyder, UN. Z., Vincent, J.. L.,
Raichle, M.. E., Schlaggar, B. L., & Petersen, S. E. (2007). Distinct
brain networks for adaptive and stable task control in humans.
Actes de l'Académie nationale des sciences, 104(26),
11073–11078. https://doi.org/10.1073/pnas.0704320104,
PubMed: 17576922
Duncan, J.. (2010). The multiple-demand (MARYLAND) system of the pri-
mate brain: Mental programs for intelligent behaviour. Trends
in Cognitive Sciences, 14(4), 172–179. https://est ce que je.org/10.1016/j
.tics.2010.01.004, PubMed: 20171926
Evrard, H. C., Logothetis, N. K., & (Bud) Craig, UN. D. (2014). Mod-
ular architectonic organization of the insula in the macaque
monkey. Journal of Comparative Neurology, 522(1), 64–97.
https://doi.org/10.1002/cne.23436, PubMed: 23900781
Fedorenko, E., Duncan, J., & Kanwisher, N. (2013). Broad domain
generality in focal regions of frontal and parietal cortex. Proceed-
ings of the National Academy of Sciences, 110(41), 16616–16621.
https://doi.org/10.1073/pnas.1315235110, PubMed: 24062451
Feinberg, T., & Mallatt, J.. (2013). The evolutionary and genetic ori-
gins of consciousness in the Cambrian Period over 500 million
years ago. Frontiers in Psychology, 4, 667. https://est ce que je.org/10
.3389/fpsyg.2013.00667, PubMed: 24109460
Feldman, H., & Friston, K. J.. (2010). Attention, uncertainty, et
free-energy. Frontiers in Human Neuroscience, 4, 215. https://
doi.org/10.3389/fnhum.2010.00215, PubMed: 21160551
Fernandino, L., Binder, J.. R., Desai, R.. H., Pendl, S. L., Humphries,
C. J., Gross, W. L., Conant, L. L., & Seidenberg, M.. S. (2016).
Concept representation reflects multimodal abstraction: A frame-
work for embodied semantics. Cortex cérébral, 26(5), 2018–2034.
https://doi.org/10.1093/cercor/bhv020, PubMed: 25750259
Finlay, B. L., & Uchiyama, R.. (2015). Developmental mechanisms
channeling cortical evolution. Trends in Neurosciences, 38(2),
69–76. https://doi.org/10.1016/j.tins.2014.11.004, PubMed:
25497421
Fotopoulou, UN., & Tsakiris, M.. (2017). Mentalizing homeostasis:
The social origins of interoceptive inference. Neuropsychoana-
lysis, 19(1), 3–28. https://doi.org/10.1080/15294145.2017
.1294031
Friston, K. (2010). The free-energy principle: A unified brain theory?
Nature Revues Neurosciences, 11(2), 127–138. https://est ce que je.org/10
.1038/nrn2787, PubMed: 20068583
Friston, K., Fitzgerald, T., Rigoli, F., Schwartenbeck, P., & Pezzulo,
G. (2017). Active inference: A process theory. Neural Computa-
tion, 29(1), 1–49. https://doi.org/10.1162/ NECO_a_00912,
PubMed: 27870614
Galvez-Pol, UN., McConnell, R., & Kilner, J.. M.. (2020). Active sam-
pling in visual search is coupled to the cardiac cycle. Cognition,
196, 104149. https://doi.org/10.1016/j.cognition.2019.104149,
PubMed: 31786324
García-Cabezas, M.. Á., Zikopoulos, B., & Barbas, H. (2019). Le
structural model: A theory linking connections, plasticity, pathol-
ogy, development and evolution of the cerebral cortex. Cerveau
Structure and Function, 224(3), 985–1008. https://est ce que je.org/10
.1007/s00429-019-01841-9, PubMed: 30739157
Gee, H. (2018). Across the bridge: Understanding the origin of the
vertebrates. University of Chicago Press. https://doi.org/10.7208
/chicago/9780226403199.001.0001
Genon, S., Bernhardt, B. C., La Joie, R., Amunts, K., & Eickhoff, S. B.
(2021). The many dimensions of human hippocampal organization
et (dys)fonction. Trends in Neurosciences, 44(12), 977–989.
https://doi.org/10.1016/j.tins.2021.10.003, PubMed: 34756460
Gerhart-Hines, Z., & Lazar, M.. UN. (2015). Circadian metabolism in
the light of evolution. Endocrine Reviews, 36(3), 289–304.
https://doi.org/10.1210/er.2015-1007, PubMed: 25927923
Gianaros, P.. J., & Wager, T. D. (2015). Brain-body pathways linking
psychological stress and physical health. Current Directions in
Sciences psychologiques, 24(4), 313–321. https://est ce que je.org/10.1177
/0963721415581476, PubMed: 26279608
Gonzalez-Castillo, J., Hoy, C. W., Handwerker, D. UN., Roopchansingh,
V., Il a dit, S. J., Saad, Z. S., Cox, R.. W., & Bandettini, P.. UN. (2015).
Task dependence, tissue specificity, and spatial distribution of
widespread activations in large single-subject functional MRI
datasets at 7T. Cortex cérébral, 25(12), 4667–4677. https://est ce que je
.org/10.1093/cercor/bhu148, PubMed: 25405938
Gonzalez-Castillo, J., Saad, Z. S., Handwerker, D. UN., Il a dit, S. J.,
Brenowitz, N., & Bandettini, P.. UN. (2012). Whole-brain,
time-locked activation with simple tasks revealed using massive
averaging and model-free analysis. Actes de la Nationale
Académie des Sciences, 109(14), 5487–5492. https://est ce que je.org/10
.1073/pnas.1121049109, PubMed: 22431587
Grady, C. L. (2020). Meta-analytic and functional connectivity evi-
dence from functional magnetic resonance imaging for an ante-
rior to posterior gradient of function along the hippocampal axis.
Hippocampus, 30(5), 456–471. https://doi.org/10.1002/ hipo
.23164, PubMed: 31589003
Grund, M., Al, E., Pabst, M., Dabbagh, UN., Stephani, T., Nierhaus,
T., Gaebler, M., & Villringer, UN. (2022). Respiration, heartbeat,
and conscious tactile perception. Journal des neurosciences, 42(4),
643–656. https://doi.org/10.1523/ JNEUROSCI.0592-21.2021,
PubMed: 34853084
Gu, X., & FitzGerald, T. H. B. (2014). Interoceptive inference:
Homeostasis and decision-making. Tendances des sciences cognitives,
18(6), 269–270. https://doi.org/10.1016/j.tics.2014.02.001,
PubMed: 24582825
Guell, X., Gabriela, J.. D. E., & Schmahmann, J.. D. (2018). Triple
representation of language, working memory, social and emotion
processing in the cerebellum: Convergent evidence from task
and seed-based resting-state fMRI analyses in a single large
cohort. NeuroImage, 172, 437–449. https://est ce que je.org/10.1016/j
.neuroimage.2018.01.082, PubMed: 29408539
Guell, X., Schmahmann, J.. D., Gabriela, J.. D., & Ghosh, S. S. (2018).
Functional gradients of the cerebellum. eLife, 7, e36652. https://
doi.org/10.7554/eLife.36652, PubMed: 30106371
Neurosciences en réseau
1026
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
.
t
/
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
Hadders-Algra, M.. (2018). Early human motor development: Depuis
variation to the ability to vary and adapt. Neuroscience and Bio-
behavioral Reviews, 90, 411–427. https://est ce que je.org/10.1016/j
.neubiorev.2018.05.009, PubMed: 29752957
Hasson, U., Chen, J., & Honey, C. J.. (2015). Hierarchical process
mémoire: Memory as an integral component of information pro-
cessation. Tendances des sciences cognitives, 19(6), 304–313. https://est ce que je
.org/10.1016/j.tics.2015.04.006, PubMed: 25980649
Hilgetag, C. C., & Goulas, UN. (2020). ‘Hierarchy’ in the organiza-
tion of brain networks. Philosophical Transactions of the Royal
Society B: Sciences biologiques, 375(1796), 20190319. https://
doi.org/10.1098/rstb.2019.0319, PubMed: 32089116
Hohwy,
J..
(2013). The predictive mind. Oxford University
Presse. https://doi.org/10.1093/acprof:oso/9780199682737
.001.0001
Hohwy, J., & Seth, UN. (2020). Predictive processing as a systematic
basis for identifying the neural correlates of consciousness. Phi-
losophy and the Mind Sciences, 1(II). https://doi.org/10.33735
/phimisci.2020.II.64
Huntenburg, J.. M., Bazin, P.-L., Goulas, UN., Tardif, C. L., Villringer,
UN., & Margulies, D. S. (2017). A systematic relationship between
functional connectivity and intracortical myelin in the human
cerebral cortex. Cortex cérébral, 27(2), 981–997. https://doi.org
/10.1093/cercor/bhx030, PubMed: 28184415
Hutchinson, J.. B., & Barrett, L. F. (2019). The power of predictions:
An emerging paradigm for psychological research. Current Direc-
tions in Psychological Science, 28(3), 280–291. https://est ce que je.org/10
.1177/0963721419831992, PubMed: 31749520
Insausti, R., & Amaral, D. G. (2012). Hippocampal formation. Dans
The human nervous system (pp. 896–942). Elsevier. https://est ce que je
.org/10.1016/B978-0-12-374236-0.10024-0
Ito, M.. (2008). Control of mental activities by internal models in the
cerebellum. Nature Revues Neurosciences, 9(4), 304–313.
https://doi.org/10.1038/nrn2332, PubMed: 18319727
Johnson, S. P.. (2013). Development of the visual system. In Neural
circuit development and function in the brain (pp. 249–269).
Elsevier. https://doi.org/10.1016/B978-0-12-397267-5.00033-9
Jordan, R., & Keller, G. B. (2020). Opposing influence of top-down
and bottom-up input on excitatory layer 2/3 neurons in mouse
primary visual cortex. Neurone, 108(6), 1194–1206. https://est ce que je
.org/10.1016/j.neuron.2020.09.024, PubMed: 33091338
Joyce, M.. K. P., & Barbas, H. (2018). Cortical connections position
primate area 25 as a keystone for interoception, emotion, et
mémoire. Journal des neurosciences, 38(7), 1677–1698. https://
doi.org/10.1523/ JNEUROSCI.2363-17.2017, PubMed:
29358365
Kanai, R., Komura, Y., Shipp, S., & Friston, K. (2015). Cérébral
hierarchies: Traitement prédictif, precision and the pulvinar.
Philosophical Transactions of the Royal Society B: Biological
les sciences, 370(1668), 20140169. https://doi.org/10.1098/rstb
.2014.0169, PubMed: 25823866
Kaplan, H. S., & Zimmer, M.. (2020). Brain-wide representations of
ongoing behavior: A universal principle? Current Opinion in
Neurobiology, 64, 60–69. https://doi.org/10.1016/j.conb.2020
.02.008, PubMed: 32203874
Katsumi, Y., Kamona, N., Zhang, J., Bunce, J.. G., Hutchinson, J.. B.,
Yarossi, M., Tunik, E., Quigley, K. S., Dickerson, B. C., & Barrett,
L. F. (2021). Functional connectivity gradients as a common
neural architecture for predictive processing in the human brain.
bioRxiv. https://doi.org/10.1101/2021.09.01.456844
Keck, T., Keller, G. B., Jacobsen, R.. JE., Eysel, U. T., Bonhoeffer, T.,
& Hübener, M.. (2013). Synaptic scaling and homeostatic plastic-
ity in the mouse visual cortex in vivo. Neurone, 80(2), 327–334.
https://doi.org/10.1016/j.neuron.2013.08.018, PubMed:
24139037
Keller, G. B., & Mrsic-Flogel, T. (2018). Traitement prédictif: UN
canonical cortical computation. Neurone, 100(2), 424–435. https://
doi.org/10.1016/j.neuron.2018.10.003, PubMed: 30359606
Kelly, R.. M., & Strick, P.. L. (2003). Cerebellar loops with motor
cortex and prefrontal cortex of a nonhuman primate. Journal de
Neurosciences, 23(23), 8432–8444. https://est ce que je.org/10.1523
/JNEUROSCI.23-23-08432.2003, PubMed: 12968006
Khalsa, S. S., Adolphs, R., Cameron, Ô. G., Critchley, H. D.,
Davenport, P.. W., Feinstein, J.. S., Feusner, J.. D., Garfinkel, S. N.,
voie, R.. D., Mehling, W. E., Meuret, UN. E., Nemeroff, C. B.,
Oppenheimer, S., Petzschner, F. H., Pollatos, O., Rhudy, J.. L.,
Schramm, L. P., Simmons, W. K., Stein, M.. B., … Zucker, N.
(2018). Interoception and mental health: A roadmap. Biological
Psychiatry: Cognitive Neuroscience and Neuroimaging, 3(6),
501–513. https://doi.org/10.1016/j.bpsc.2017.12.004, PubMed:
29884281
King, M., Hernandez-Castillo, C. R., Poldrack, R.. UN., Ivry, R.. B., &
Diedrichsen, J.. (2019). Functional boundaries in the human cer-
ebellum revealed by a multi-domain task battery. Nature Neuro-
science, 22(8), 1371–1378. https://doi.org/10.1038/s41593-019
-0436-X, PubMed: 31285616
Kleckner, je. R., Zhang, J., Touroutoglou, UN., Chanes, L., Xia, C., Simmons,
W. K., Quigley, K. S., Dickerson, B. C., & Barrett, L. F. (2017). Evi-
dence for a large-scale brain system supporting allostasis and
interoception in humans. Nature Human Behaviour, 1(5), 1–14.
https://doi.org/10.1038/s41562-017-0069, PubMed: 28983518
Kluger, D. S., Balestrieri, E., Busch, N. UN., & Gross, J.. (2021). Respi-
ration aligns perception with neural excitability. eLife, 10, e70907.
https://doi.org/10.7554/eLife.70907, PubMed: 34904567
Kong, R., Li, J., Orban, C., Sabuncu, M.. R., Liu, H., Schaefer, UN.,
Sun, N., Zuo, X.-N., Holmes, UN. J., Eickhoff, S. B., & Yeo,
B. T. T. (2019). Spatial topography of individual-specific cortical
networks predicts human cognition, personality, and emotion.
Cortex cérébral, 29(6), 2533–2551. https://est ce que je.org/10.1093
/cercor/bhy123, PubMed: 29878084
Kumaran, D., Hassabis, D., & McClelland, J.. L. (2016). What learn-
ing systems do intelligent agents need? Complementary learning
systems theory updated. Tendances des sciences cognitives, 20(7),
512–534. https://doi.org/10.1016/j.tics.2016.05.004, PubMed:
27315762
Kunzendorf, S., Klotzsche, F., Akbal, M., Villringer, UN., Ohl, S., &
Gaebler, M.. (2019). Active information sampling varies across
the cardiac cycle. Psychophysiology, 56(5), e13322. https://est ce que je
.org/10.1111/psyp.13322, PubMed: 30620083
Lamme, V. UN. F., & Roelfsema, P.. R.. (2000). The distinct modes of
vision offered by feedforward and recurrent processing. Trends in
Neurosciences, 23(11), 571–579. https://doi.org/10.1016/S0166
-2236(00)01657-X, PubMed: 11074267
Larivière, S., Vos de Wael, R., Hong, S.-J., Paquola, C., Tavakol, S.,
Lowe, UN. J., Schrader, D. V., & Bernhardt, B. C. (2020). Multiscale
structure–function gradients in the neonatal connectome.
Neurosciences en réseau
1027
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
.
t
/
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
Cortex cérébral, 30(1), 47–58. https://doi.org/10.1093/cercor
/bhz069, PubMed: 31220215
Lathe, R., Singadia, S., Jordan, C., & Riedel, G. (2020). The intero-
ceptive hippocampus: Mouse brain endocrine receptor expres-
sion highlights a dentate gyrus (DG)–cornu ammonis (Californie)
challenge–sufficiency axis. PLoS ONE, 15(1), e0227575. https://
doi.org/10.1371/journal.pone.0227575, PubMed: 31940330
Lautin, UN. (2001). The limbic brain. Kluwer Academic/ Plenum
Publishers.
Leinweber, M., Ward, D. R., Sobczak, J.. M., Attinger, UN., & Keller,
G. B. (2017). A sensorimotor circuit in mouse cortex for visual
flow predictions. Neurone, 95(6), 1420–1432. https://est ce que je.org/10
.1016/j.neuron.2017.08.036, PubMed: 28910624
Levinthal, D. J., & Strick, P.. L. (2012). The motor cortex com-
municates with the kidney.
Journal des neurosciences, 32(19),
6726–6731. https://doi.org/10.1523/ JNEUROSCI.0406-12
.2012, PubMed: 22573695
Levinthal, D. J., & Strick, P.. L. (2020). Multiple areas of the cerebral
cortex influence the stomach. Proceedings of the National Acad-
emy of Sciences, 117(23), 13078–13083. https://doi.org/10.1073
/pnas.2002737117, PubMed: 32434910
Liang, M., Mouraux, UN., Hu, L., & Iannetti, G. D. (2013). Primary
sensory cortices contain distinguishable spatial patterns of activ-
ity for each sense. Communications naturelles, 4(1), 1979. https://
doi.org/10.1038/ncomms2979, PubMed: 23752667
Lindquist, K. UN., Satpute, UN. B., Wager, T. D., Weber, J., & Barrett,
L. F. (2016). The brain basis of positive and negative affect: Evi-
dence from a meta-analysis of the human neuroimaging litera-
ture. Cortex cérébral, 26(5), 1910–1922. https://est ce que je.org/10
.1093/cercor/bhv001, PubMed: 25631056
Lisman, J., & Redish, UN. D. (2009). Prediction, sequences and the
hippocampus. Philosophical Transactions of the Royal Society B:
Sciences biologiques, 364(1521), 1193–1201. https://est ce que je.org/10
.1098/rstb.2008.0316, PubMed: 19528000
Liu, K., Sibille, J., & Dragoi, G. (2018). Generative predictive codes
by multiplexed hippocampal neuronal tuplets. Neurone, 99(6),
1329–1341. https://doi.org/10.1016/j.neuron.2018.07.047,
PubMed: 30146305
Margulies, D. S., Ghosh, S. S., Goulas, UN., Falkiewicz, M., Huntenburg,
J.. M., Langs, G., Bezgin, G., Eickhoff, S. B., Castellanos, F. X.,
Pétrides, M., Jefferies, E., & Smallwood, J.. (2016). Situating the
default-mode network along a principal gradient of macroscale
cortical organization. Proceedings of the National Academy of
les sciences, 113(44), 12574–12579. https://doi.org/10.1073/pnas
.1608282113, PubMed: 27791099
Mckeown, B., Strawson, W. H., Wang, H.-T., Karapanagiotidis, T.,
Vos de Wael, R., Benkarim, O., Turnbull, UN., Margulies, D.,
Jefferies, E., McCall, C., Bernhardt, B., & Smallwood, J.. (2020).
The relationship between individual variation in macroscale
functional gradients and distinct aspects of ongoing thought.
NeuroImage, 220, 117072. https://doi.org/10.1016/j.neuroimage
.2020.117072, PubMed: 32585346
McNamee, D., & Wolpert, D. M.. (2019). Internal models in biolog-
ical control. Annual Review of Control, Robotics, and Autono-
mous Systems, 2, 339–364. https://est ce que je.org/10.1146/annurev
-control-060117-105206, PubMed: 31106294
Mesulam, M.. (Ed.). (2000). Principles of behavioral and cognitive
neurology (2nd ed). Presse universitaire d'Oxford.
Miller, E. K., & Cohen, J.. D. (2001). An integrative theory of prefrontal
cortex function. Revue annuelle des neurosciences, 24(1), 167–202.
https://doi.org/10.1146/annurev.neuro.24.1.167, PubMed:
11283309
Muckli, L., De Martino, F., Visioli, L., Petro, L. S., Forgeron, F. W., Voiture d'Ugur,
K., Goebel, R., & Yacoub, E. (2015). Contextual feedback to
superficial layers of V1. Biologie actuelle, 25(20), 2690–2695.
https://doi.org/10.1016/j.cub.2015.08.057, PubMed: 26441356
Mufson, E. J., & Mesulam, M.-M. (1982). Insula of the old world
monkey. II: Afferent cortical input and comments on the claus-
trum. Journal of Comparative Neurology, 212(1), 23–37. https://
doi.org/10.1002/cne.902120103, PubMed: 7174906
Nieuwenhuys, R.. (2012). The insular cortex: A review. In Progress
in brain research (Vol. 195, pp. 123–163). Elsevier. https://doi.org
/10.1016/B978-0-444-53860-4.00007-6, PubMed: 22230626
Nieuwenhuys, R., & Puelles, L. (2016). Towards a new neuromor-
phology. Springer International Publishing. https://est ce que je.org/10
.1007/978-3-319-25693-1
Öngür, D., Ferry, UN. T., & Prix, J.. L. (2003). Architectonic subdivi-
sion of the human orbital and medial prefrontal cortex. Journal de
Comparative Neurology, 460(3), 425–449. https://est ce que je.org/10
.1002/cne.10609, PubMed: 12692859
Onuki, Y., Van Someren, E. J.. W., De Zeeuw, C. JE., & Van der Werf,
Oui. D. (2015). Hippocampal–cerebellar interaction during spatio-
temporal prediction. Cortex cérébral, 25(2), 313–321. https://est ce que je
.org/10.1093/cercor/bht221, PubMed: 23968839
Owens, UN. P., Allen, M., Ondobaka, S., & Friston, K. J.. (2018). Inter-
oceptive inference: From computational neuroscience to clinic.
Neuroscience and Biobehavioral Reviews, 90, 174–183. https://
doi.org/10.1016/j.neubiorev.2018.04.017, PubMed: 29694845
Paquola, C., Benkarim, O., DeKraker, J., Larivière, S., Frässle, S.,
Royer, J., Tavakol, S., Valk, S., Bernasconi, UN., Bernasconi, N.,
Khan, UN., Evans, UN. C., Razi, UN., Smallwood, J., & Bernhardt,
B. C. (2020). Convergence of cortical types and functional motifs
in the human mesiotemporal lobe. eLife, 9, e60673. https://est ce que je
.org/10.7554/eLife.60673, PubMed: 33146610
Paquola, C., Wael, R.. V. D., Wagstyl, K., Bethlehem, R.. UN. JE., Hong,
S.-J., Seidlitz, J., Bullmore, E. T., Evans, UN. C., Misic, B., Margulies,
D. S., Smallwood, J., & Bernhardt, B. C. (2019). Microstructural
and functional gradients are increasingly dissociated in transmo-
dal cortices. PLoS Biology, 17(5), e3000284. https://est ce que je.org/10
.1371/journal.pbio.3000284, PubMed: 31107870
Parr, T., & Friston, K. J.. (2019). Attention or salience? Current Opin-
ion in Psychology, 29, 1–5. https://doi.org/10.1016/j.copsyc
.2018.10.006, PubMed: 30359960
Parr, T., Rees, G., & Friston, K. J.. (2018). Computational neuropsy-
chology and Bayesian inference. Frontiers in Human Neurosci-
ence, 12, 61. https://doi.org/10.3389/fnhum.2018.00061,
PubMed: 29527157
Petzschner, F. H., Garfinkel, S. N., Paulus, M.. P., Koch, C., &
Khalsa, S. S. (2021). Computational models of interoception
and body regulation. Trends in Neurosciences, 44(1), 63–76.
https://doi.org/10.1016/j.tins.2020.09.012, PubMed: 33378658
Petzschner, F. H., Weber, L. UN. E., Gard, T., & Stephan, K. E. (2017).
Computational psychosomatics and computational psychiatry:
Toward a joint framework for differential diagnosis. Biological
Psychiatry, 82(6), 421–430. https://doi.org/10.1016/j.biopsych
.2017.05.012, PubMed: 28619481
Neurosciences en réseau
1028
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
t
.
/
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
Pezzulo, G., Kemere, C., & der Meer, M.. UN. UN. (2017). Internally
generated hippocampal sequences as a vantage point to probe
future-oriented cognition. Annals of the New York Academy of
les sciences, 1396(1), 144–165. https://doi.org/10.1111/nyas
.13329, PubMed: 28548460
Pezzulo, G., Rigoli, F., & Friston, K. (2015). Active inference,
homeostatic regulation and adaptive behavioural control. Prog-
ress in Neurobiology, 134, 17–35. https://est ce que je.org/10.1016/j
.pneurobio.2015.09.001, PubMed: 26365173
Pezzulo, G., Zorzi, M., & Corbetta, M.. (2021). The secret life of
predictive brains: What’s spontaneous activity for? Trends in Cog-
nitive Sciences, 25(9), 730–743. https://doi.org/10.1016/j.tics
.2021.05.007, PubMed: 34144895
Pisano, T. J., Dhanerawala, Z. M., Kislin, M., Bakshinskaya, D.,
Ange, E. UN., Hansen, E. J., Hoag, UN. T., Lee, J., de Oude, N. L.,
Umadevi, K., Verpeut, J.. L., Hoebeek, F. E., Richardson, B. D., &
Boele, H.-J. (2021). Homologous organization of cerebellar path-
ways to sensory, moteur, and associative forebrain. Cell Reports,
36(12), 109721. https://doi.org/10.1016/j.celrep.2021.109721,
PubMed: 34551311
Power, J.. D., Cohen, UN. L., Nelson, S. M., Wig, G. S., Barnes, K. UN.,
Church, J.. UN., Vogel, UN. C., Laumann, T. O., Miezin, F. M.,
Schlaggar, B. L., & Petersen, S. E. (2011). Functional network orga-
nization of the human brain. Neurone, 72(4), 665–678. https://est ce que je
.org/10.1016/j.neuron.2011.09.006, PubMed: 22099467
Prix, J.. L., & Drevets, W. C. (2010). Neurocircuitry of mood disor-
ders. Neuropsychopharmacology, 35(1), 192–216. https://doi.org
/10.1038/npp.2009.104, PubMed: 19693001
Quigley, K. S., Kanoski, S., Grill, W. M., Barrett, L. F., & Tsakiris, M..
(2021). Functions of interoception: From energy regulation to
experience of the self. Trends in Neurosciences, 44(1), 29–38.
https://doi.org/10.1016/j.tins.2020.09.008, PubMed: 33378654
Rao, R.. P.. N., & Ballard, D. H. (1999). Predictive coding in the
visual cortex: A functional interpretation of some extra-classical
receptive-field effects. Neurosciences naturelles, 2(1), 79–87. https://
doi.org/10.1038/4580, PubMed: 10195184
Raut, R.. V., Snyder, UN. Z., & Raichle, M.. E. (2020). Hierarchical
dynamics as a macroscopic organizing principle of the human
brain. Actes de l'Académie nationale des sciences, 117(34),
20890–20897. https://doi.org/10.1073/pnas.2003383117,
PubMed: 32817467
Roxo, M.. R., Franceschini, P.. R., Zubaran, C., Kleber, F. D., &
Sander, J.. W. (2011). The limbic system conception and its his-
torical evolution. The Scientific World Journal, 11, 2427–2440.
https://doi.org/10.1100/2011/157150, PubMed: 22194673
Royer, J., Paquola, C., Larivière, S., Vos de Wael, R., Tavakol, S.,
Lowe, UN. J., Benkarim, O., Evans, UN. C., Bzdok, D., Smallwood,
J., Frauscher, B., & Bernhardt, B. C. (2020). Myeloarchitecture
gradients in the human insula: Histological underpinnings and
association to intrinsic functional connectivity. NeuroImage,
216, 116859. https://doi.org/10.1016/j.neuroimage.2020
.116859, PubMed: 32325211
Satpute, UN. B., Kang, J., Bickart, K. C., Yardley, H., Wager, T. D., &
Barrett, L. F. (2015). Involvement of sensory regions in affective
experience: A meta-analysis. Frontiers in Psychology, 6, 1860.
https://doi.org/10.3389/fpsyg.2015.01860, PubMed: 26696928
Schmahmann, J.. D. (1996). From movement to thought: Anatomic
substrates of the cerebellar contribution to cognitive processing.
Human Brain Mapping, 4(3), 174–198. https://doi.org/10.1002
/(SICI )1097-0193(1996)4:3<174::AID-HBM3>3.0.CO;2-0,
PubMed: 20408197
Schulkin, J., & Sterling, P.. (2019). Allostasis: A brain-centered, pre-
dictive mode of physiological regulation. Trends in Neurosci-
ences, 42(10), 740–752. https://doi.org/10.1016/j.tins.2019.07
.010, PubMed: 31488322
Sengupta, B., Stemmler, M.. B., & Friston, K. J.. (2013). Information
and efficiency in the nervous system—A synthesis. PLoS Compu-
tational Biology, 9(7), e1003157. https://doi.org/10.1371/journal
.pcbi.1003157, PubMed: 23935475
Sennesh, E., Theriault, J., Brooks, D., Meent, J.-W. van de, Barrett,
L. F., & Quigley, K. S. (2021). Interoception as modeling, allosta-
sis as control. Biological Psychology, 167, 108242. https://doi.org
/10.1016/j.biopsycho.2021.108242, PubMed: 34942287
Sepulcre, J., Sabuncu, M.. R., Yeo, T. B., Liu, H., & Johnson, K. UN.
(2012). Stepwise connectivity of the modal cortex reveals the
multimodal organization of the human brain. Journal of Neuro-
science, 32(31), 10649–10661. https://est ce que je.org/10.1523
/JNEUROSCI.0759-12.2012, PubMed: 22855814
Seth, UN. K. (2013). Interoceptive inference, emotion, et le
embodied self. Tendances des sciences cognitives, 17(11), 565–573.
https://doi.org/10.1016/j.tics.2013.09.007, PubMed: 24126130
Seth, UN. K. (2015). The cybernetic Bayesian brain—From interocep-
tive inference to sensorimotor contingencies. In T. Metzinger &
J.. M.. Windt (Éd.), Open MIND. https://doi.org/10.15502
/9783958570108
Seth, UN. K., & Friston, K. J.. (2016). Active interoceptive inference
and the emotional brain. Philosophical Transactions of the Royal
Society of London B: Sciences biologiques, 371(1708), 20160007.
https://doi.org/10.1098/rstb.2016.0007, PubMed: 28080966
Seth, UN. K., Suzuki, K., & Critchley, H. D. (2012). An interoceptive
predictive coding model of conscious presence. Frontiers in Psy-
cologie, 2, 395. https://doi.org/10.3389/fpsyg.2011.00395,
PubMed: 22291673
Seth, UN. K., & Tsakiris, M.. (2018). Being a beast machine: Le
somatic basis of selfhood. Tendances des sciences cognitives, 22(11),
969–981. https://doi.org/10.1016/j.tics.2018.08.008, PubMed:
30224233
Shadmehr, R., Forgeron, M.. UN., & Krakauer, J.. W. (2010). Error correc-
tion, sensory prediction, and adaptation in motor control. Annual
Review of Neuroscience, 33(1), 89–108. https://doi.org/10.1146
/annurev-neuro-060909-153135, PubMed: 20367317
Shafiei, G., Markello, R.. D., de Wael, R.. V., Bernhardt, B. C.,
Fulcher, B. D., & Misic, B. (2020). Topographic gradients of
intrinsic dynamics across neocortex. bioRxiv. https://est ce que je.org/10
.1101/2020.07.03.186916
Shannon, C., & Weaver, W. (1964). The mathematical theory of
communication (10th ed.). University of Illinois Press. (Original
work published 1949)
Sillito, UN. M., & Jones, H. E. (2002). Corticothalamic interactions in
the transfer of visual information. Philosophical Transactions of
the Royal Society of London B: Sciences biologiques, 357(1428),
1739–1752. https://doi.org/10.1098/rstb.2002.1170, PubMed:
12626008
Smallwood, J., Bernhardt, B. C., Leech, R., Bzdok, D., Jefferies, E.,
& Margulies, D. S. (2021). The default mode network in cogni-
tion: A topographical perspective. Nature Revues Neurosciences,
Neurosciences en réseau
1029
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
22, 503–513. https://doi.org/10.1038/s41583-021-00474-4,
PubMed: 34226715
Forgeron, R., Badcock, P., & Friston, K. J.. (2021). Recent advances in the
application of predictive coding and active inference models within
clinical neuroscience. Psychiatry and Clinical Neurosciences, 75(1),
3–13. https://doi.org/10.1111/pcn.13138, PubMed: 32860285
Forgeron, R., Thayer, J.. F., Khalsa, S. S., & voie, R.. D. (2017). The hier-
archical basis of neurovisceral integration. Neuroscience and
Biobehavioral Reviews, 75, 274–296. https://est ce que je.org/10.1016/j
.neubiorev.2017.02.003, PubMed: 28188890
Sokolov, UN. UN., Miall, R.. C., & Ivry, R.. B. (2017). The cerebellum:
Adaptive prediction for movement and cognition. Trends in
Cognitive Sciences, 21(5), 313–332. https://est ce que je.org/10.1016/j
.tics.2017.02.005, PubMed: 28385461
Stawarczyk, D., Bezdek, M.. UN., & Zacks, J.. M.. (2021). Event repre-
sentations and predictive processing: The role of the midline
default network core. Topics in Cognitive Science, 13(1), 164–186.
https://doi.org/10.1111/tops.12450, PubMed: 31486286
Stephan, K. E., Manjaly, Z. M., Mathys, C. D., Weber, L. UN. E.,
Paliwal, S., Gard, T., Tittgemeyer, M., Fleming, S. M., Haker, H.,
Seth, UN. K., & Petzschner, F. H. (2016). Allostatic self-efficacy: UN
metacognitive theory of dyshomeostasis-induced fatigue and
depression. Frontiers in Human Neuroscience, 10, 550. https://
doi.org/10.3389/fnhum.2016.00550, PubMed: 27895566
Sterling, P.. (2012). Allostasis: A model of predictive regulation.
Physiology and Behavior, 106(1), 5–15. https://est ce que je.org/10.1016
/j.physbeh.2011.06.004, PubMed: 21684297
Sterling, P., & Laughlin, S. (2015). Principles of neural design. AVEC
Presse. https://doi.org/10.7551/mitpress/9780262028707.001
.0001
Striedter, G. F., & Northcutt, R.. G. (2020). Brains through time: UN
natural history of vertebrates. Presse universitaire d'Oxford. https://
doi.org/10.1093/oso/9780195125689.001.0001
Suarez, UN. N., Hsu, T. M., Liu, C. M., Noble, E. E., Cortella, UN. M.,
Nakamoto, E. M., Hahn, J.. D., de Lartigue, G., & Kanoski, S. E.
(2018). Gut vagal sensory signaling regulates hippocampus
function through multi-order pathways. Communications naturelles,
9(1), 2181. https://doi.org/10.1038/s41467-018-04639-1,
PubMed: 29872139
Sultan, F., Augath, M., Hamodeh, S., Murayama, Y., Oeltermann,
UN., Rauch, UN., & Thier, P.. (2012). Unravelling cerebellar path-
ways with high temporal precision targeting motor and extensive
sensory and parietal networks. Communications naturelles, 3(1),
924. https://doi.org/10.1038/ncomms1912, PubMed: 22735452
Sydnor, V. J., Larsen, B., Bassett, D. S., Alexander-Bloch, UN., Fair,
D. UN., Liston, C., Mackey, UN. P., Milham, M.. P., Pines, UN., Roalf,
D. R., Seidlitz, J., Xu, T., Raznahan, UN., & Satterthwaite, T. D.
(2021). Neurodevelopment of the association cortices: Motifs,
mechanisms, and implications for psychopathology. Neurone,
109(18), 2820–2846. https://doi.org/10.1016/j.neuron.2021.06
.016, PubMed: 34270921
Szinte, M., & Knapen, T. (2020). Visual organization of the default
réseau. Cortex cérébral, 30(6), 3518–3527. https://est ce que je.org/10
.1093/cercor/bhz323, PubMed: 32031204
Tanaka, H., Ishikawa, T., Lee, J., & Kakei, S. (2020). Le
cerebro-cerebellum as a locus of forward model: A review. Fron-
tiers in Systems Neuroscience, 14, 19. https://doi.org/10.3389
/fnsys.2020.00019, PubMed: 32327978
Theriault, J., Shaffer, C., Dienel, G. UN., Sander, C. Y., Hooker, J.. M.,
Dickerson, B. C., Barrett, L. F., & Quigley, K. (2021). Aerobic gly-
colysis, the efficiency tradeoff hypothesis, and the biological
basis of neuroimaging: A solution to a metabolic mystery at the
heart of neuroscience. PsyArXiv. https://doi.org/10.31234/osf.io
/pkzr8
Theriault, J., Jeune, L., & Barrett, L. F. (2021). The sense of should:
A biologically-based framework for modeling social pressure.
Physics of Life Reviews, 36, 100–136. https://est ce que je.org/10.1016/j
.plrev.2020.01.004, PubMed: 32008953
Touroutoglou, UN., Hollenbeck, M., Dickerson, B. C., & Barrett, L. F.
(2012). Dissociable large-scale networks anchored in the right
anterior insula subserve affective experience and attention.
NeuroImage, 60(4), 1947–1958. https://est ce que je.org/10.1016/j
.neuroimage.2012.02.012, PubMed: 22361166
Uddin, L. Q. (2015). Salience processing and insular cortical
function and dysfunction. Nature Revues Neurosciences, 16(1),
55–61. https://doi.org/10.1038/nrn3857, PubMed: 25406711
Ullsperger, M., Danielmeier, C., & Jocham, G. (2014). Neurophys-
iology of performance monitoring and adaptive behavior. Physi-
ological Reviews, 94(1), 35–79. https://doi.org/10.1152/physrev
.00041.2012, PubMed: 24382883
Valenza, G., Sclocco, R., Duggento, UN., Passamonti, L., Napadow,
V., Barbieri, R., & Toschi, N. (2019). The central autonomic
network at rest: Uncovering functional MRI correlates of
time-varying autonomic outflow. NeuroImage, 197, 383–390.
https://doi.org/10.1016/j.neuroimage.2019.04.075, PubMed:
31055043
Valk, S. L., Kanske, P., Parc, B., Hong, S. J., Böckler-Raettig, UN.,
Trautwein, F.-M., Bernhardt, B. C., & Chanteur, T. (2021). Changing
the social brain: Plasticity along macro-scale axes of functional
connectivity following social mental training. bioRxiv. https://est ce que je
.org/10.1101/2020.11.11.377895
van den Heuvel, M.. P., & Sporns, Ô. (2011). Rich-club organization
of the human connectome. Journal des neurosciences, 31(44),
15775–15786. https://doi.org/10.1523/ JNEUROSCI.3539-11
.2011, PubMed: 22049421
van den Heuvel, M.. P., & Sporns, Ô. (2013). An anatomical sub-
strate for integration among functional networks in human cor-
tex. Journal des neurosciences, 33(36), 14489–14500. https://est ce que je
.org/10.1523/JNEUROSCI.2128-13.2013, PubMed: 24005300
Vertes, R.. P.. (2015). Major diencephalic inputs to the hippocampus:
Supramammillary nucleus and nucleus reuniens. Circuitry and
fonction. In S. O’Mara & M.. Tsanov (Éd.), Progress in brain
recherche (Vol. 219, pp. 121–144). Elsevier. https://est ce que je.org/10
.1016/bs.pbr.2015.03.008, PubMed: 26072237
Vogt, B. UN., & Pandya, D. N. (1987). Cingulate cortex of the rhesus
monkey: II. Cortical afferents. Journal of Comparative Neurology,
262(2), 271–289. https://doi.org/10.1002/cne.902620208,
PubMed: 3624555
Vogt, B. UN., Pandya, D. N., & Rosene, D. L. (1987). Cingulate cortex
of the rhesus monkey: je. Cytoarchitecture and thalamic afferents.
Journal of Comparative Neurology, 262(2), 256–270. https://est ce que je
.org/10.1002/cne.902620207, PubMed: 3624554
Vos de Wael, R., Benkarim, O., Paquola, C., Lariviere, S., Royer, J.,
Tavakol, S., Xu, T., Hong, S.-J., Langs, G., Valk, S., Misic, B.,
Milham, M., Margulies, D., Smallwood, J., & Bernhardt, B. C.
(2020). BrainSpace: A toolbox for the analysis of macroscale
Neurosciences en réseau
1030
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Allostasis as a core feature of hierarchical gradients in the brain
gradients in neuroimaging and connectomics datasets. Commu-
nications Biology, 3(1), 1-dix. https://doi.org/10.1038/s42003
-020-0794-7, PubMed: 32139786
Vos de Wael, R., Larivière, S., Caldairou, B., Hong, S.-J., Margulies,
D. S., Jefferies, E., Bernasconi, UN., Smallwood, J., Bernasconi, N.,
& Bernhardt, B. C. (2018). Anatomical and microstructural deter-
minants of hippocampal subfield functional connectome embed-
ding. Actes de l'Académie nationale des sciences, 115(40),
10154–10159. https://doi.org/10.1073/pnas.1803667115,
PubMed: 30249658
Watson, T. C., Obiang, P., Torres-Herraez, UN., Watilliaux, UN., Coulon,
P., Rochefort, C., & Rondi-Reig, L. (2019). Anatomical and physio-
logical foundations of cerebello-hippocampal interaction. eLife, 8,
e41896. https://doi.org/10.7554/eLife.41896, PubMed: 31205000
Willis, T. (1664). Cerebri anatome, cui accessit nervorum descripto
et usus (p. [26], 240 p., [12] leaves of plates). Typis Tho.
Roycroft, Impensis Jo. Martyn & Ja. Allestry. Retrived from
https://www.proquest.com/docview/2248499302/citation
/2D60B5EA8904437BPQ/1
Wolpert, D. M., Miall, R.. C., & Kawato, M.. (1998). Internal models
in the cerebellum. Tendances des sciences cognitives, 2(9), 338–347.
https://est ce que je.org/10.1016/S1364-6613(98)01221-2, PubMed:
21227230
Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M.. R., Lashkari,
D., Hollinshead, M., Roffman, J.. L., Smoller, J.. W., Zöllei, L., Polimeni,
J.. R., Fischl, B., Liu, H., & Buckner, R.. L. (2011). The organization
of the human cerebral cortex estimated by intrinsic functional
connectivité. Journal de neurophysiologie, 106(3), 1125–1165.
https://est ce que je.org/10.1152/jn.00338.2011, PubMed: 21653723
Zelano, C., Jiang, H., Zhou, G., Arora, N., coquilles, S., Rosenow, J.,
& Gottfried, J.. UN. (2016). Nasal respiration entrains human limbic
oscillations and modulates cognitive function. Journal of Neuro-
science, 36(49), 12448–12467. https://est ce que je.org/10.1523
/JNEUROSCI.2586-16.2016, PubMed: 27927961
Zhang, J., Abiose, O., Katsumi, Y., Touroutoglou, UN., Dickerson,
B. C., & Barrett, L. F. (2019). Intrinsic functional connectivity is
organized as three interdependent gradients. Rapports scientifiques,
9(1), 15976. https://doi.org/10.1038/s41598-019-51793-7,
PubMed: 31685830
Zhang, J., Scholtens, L. H., Wei, Y., van den Heuvel, M.. P., Chanes,
L., & Barrett, L. F. (2020). Topography impacts topology: Anatom-
ically central areas exhibit a “high-level connector” profile in the
human cortex. Cortex cérébral, 30(3), 1357–1365. https://doi.org
/10.1093/cercor/bhz171, PubMed: 31504277
Zhang, S., Xu, M., Kamigaki, T., Do, J.. P.. H., Chang, W.-C., Jenvay,
S., Miyamichi, K., Luo, L., & Dan, Oui. (2014). Long-range and
local circuits for top-down modulation of visual cortical process-
ing. Science, 345(6197), 660–665. https://doi.org/10.1126
/science.1254126, PubMed: 25104383
Zhu, J.-N., & Wang, J.-J. (2008). The cerebellum in feeding control:
Possible function and mechanism. Cellular and Molecular Neu-
robiology, 28(4), 469–478. https://doi.org/10.1007/s10571-007
-9236-z, PubMed: 18027085
Zhu, J.-N., Yung, W.-H., Kwok-Chong Chow, B., Chan, Y.-S., &
Wang, J.-J. (2006). The cerebellar-hypothalamic circuits: Potential
pathways underlying cerebellar involvement in somatic-visceral
l'intégration. Brain Research Reviews, 52(1), 93–106. https://est ce que je
.org/10.1016/j.brainresrev.2006.01.003, PubMed: 16497381
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
6
4
1
0
1
0
2
0
5
6
2
7
4
n
e
n
_
un
_
0
0
2
4
0
p
d
t
.
/
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Neurosciences en réseau
1031