Evidence for Integrated Visual Face and Body
Representations in the Anterior Temporal Lobes
Bronson B. Harry1,2, Katja Umla-Runge3, Andrew D. Lawrence3,
Kim S. Graham3, and Paul E. Downing2
Abstrait
■ Research on visual face perception has revealed a region in
the ventral anterior temporal lobes, often referred to as the
anterior temporal face patch (ATFP), which responds strongly
to images of faces. À ce jour, the selectivity of the ATFP has been
examined by contrasting responses to faces against a small
selection of categories. Ici, we assess the selectivity of the
ATFP in humans with a broad range of visual control stimuli
to provide a stronger test of face selectivity in this region. Dans
Experiment 1, participants viewed images from 20 stimulus
categories in an event-related fMRI design. Faces evoked more
activity than all other 19 categories in the left ATFP. In the right
ATFP, equally strong responses were observed for both faces
and headless bodies. To pursue this unexpected finding, dans
Experiment 2, we used multivoxel pattern analysis to examine
whether the strong response to face and body stimuli reflects a
common coding of both classes or instead overlapping but dis-
tinct representations. On a voxel-by-voxel basis, face and whole-
body responses were significantly positively correlated in the
right ATFP, but face and body-part responses were not. Ce
finding suggests that there is shared neural coding of faces and
whole bodies in the right ATFP that does not extend to individ-
ual body parts. In contrast, the same approach revealed distinct
face and body representations in the right fusiform gyrus. These
results are indicative of an increasing convergence of distinct
sources of person-related perceptual information proceeding
from the posterior to the anterior temporal cortex. ■
INTRODUCTION
fMRI studies of humans, Old World monkeys (macaques),
and New World monkeys (marmosets) have uncovered
several face-selective regions in the occipital and tem-
poral lobes (Hung et al., 2015; Tsao & Livingstone, 2008;
Kanwisher & Yovel, 2006; Haxby, Hoffman, & Gobbini,
2000). Although cross-species homology has not yet been
clearly established, this network of face-selective regions
shows a strikingly similar organization across human and
nonhuman primates (Hung et al., 2015; McMahon, Russ,
Elnaiem, Kurnikova, & Leopold, 2015; Rajimehr, Jeune,
& Cellule produit, 2009; Tsao, Moeller, & Friwald, 2008) et
consists of several ventral regions spanning the occipital
cortex, inferior temporal lobes, and STS.
An influential theoretical perspective (Haxby et al.,
2000), based on human functional imaging studies, di-
vides face-selective regions into a “core” system compris-
ing extrastriate nodes for the visual analysis of faces and
an “extended” system incorporating additional neural
regions that work in concert with the core system to ex-
tract various types of social information from faces. Le
core regions include the occipital face area (OFA; Pitcher,
Dilks, Saxe, Triantafyllou, & Kanwisher, 2011; Gauthier
et coll., 2000), the fusiform face area (FFA; Weiner &
1Western Sydney University, 2Bangor University, 3Cardiff University
Grill-Spector, 2010; Kanwisher & Yovel, 2006), et le
posterior STS (Pitcher et al., 2011; Puce, Allison, Bentin,
Sang, & McCarthy, 1998). OFA and FFA are proposed to
process static facial form, with the OFA more engaged in
part-based processing and the FFA more engaged in pro-
cessing the configuration of individual parts (Harris &
Aguirre, 2010; Schiltz, Dricot, Goebel, & Rossion, 2010;
Liu, Harris, & Kanwisher, 2009; Yovel & Kanwisher, 2005),
whereas posterior STS processes changeable aspects of
faces (par exemple., eye gaze; Haxby et al., 2000). In contrast, le
extended system is proposed to include regions such as
the amygdala and the anterior temporal cortex, domaines
that are argued to be important in appraising emotional
facial expressions (Calder, Lawrence, & Jeune, 2001; mais
see Mende-Siedlecki, Verosky, Turk-Browne, & Todorov,
2013) and encoding person-specific semantic knowledge
(Quiroga, Kreiman, Koch, & Frit, 2008; Thompson et al.,
2004), respectivement.
Recent evidence prompts consideration of whether
there are anterior temporal regions that should also be
included as a part of the core system (Duchaine & Yovel,
2015; Collins & Olson, 2014; Haxby & Gobbini, 2011). UN
number of reports (Ku, Tolias, Logothetis, & Goense, 2011;
Nestor, Plaut, & Behrmann, 2011; Pinsk et al., 2009;
Rajimehr et al., 2009; Tsao et coll., 2008) provide evidence
of at least one face-selective region in the anterior tempo-
ral lobes in humans and macaques. In macaques, electrical
© 2016 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 3.0 Unported (CC PAR 3.0) Licence
Journal des neurosciences cognitives 28:8, pp. 1178–1193
est ce que je:10.1162/jocn_a_00966
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
un
/
_
j
0
o
0
c
9
n
6
6
_
un
p
_
d
0
0
b
9
oui
6
g
6
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
.
/
F
t
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
stimulation of the anterior temporal face patches (ATFPs)
selectively induces activity in the posterior network
(Moeller, Friwald, & Tsao, 2008), suggesting that these
areas are functionally connected. De plus, the most
ATFP is unique in that neurons in this region respond
invariantly to different face views (Meyers, Borzello,
Friwald, & Tsao, 2015; Friwald & Tsao, 2010), suggérer-
ing that it forms higher-level representations that are
needed for identification (for similar evidence in human
participants, see Yang, Susilo, & Duchaine, 2016; Anzellotti,
Fairhall, & Caramazza, 2014). In support of the view that
human ATFP captures a similarly abstract representation,
Nasr and Tootell (2012) found in human participants that
fMRI activity in the ATFP closely mirrored changes in rec-
ognition performance brought about by image manipula-
tions such as face inversion and contrast reversal.
A limitation of previous studies examining visual selec-
tivity in the human ATFP has been the use of relatively
few visual control categories (par exemple., Nasr & Cellule produit, 2012;
Rajimehr et al., 2009; Tsao et coll., 2008). A wide assess-
ment of responses to items from a range of categories is
vital for determining the selectivity of a region’s response
profile (Désimone, Albright, Gross, & Bruce, 1984) et
thus for making inferences about its functional role(s).
There have been multiple-category surveys of the inferior
cortex temporal (Mur et al., 2012; Vul, Lashkari, Hsieh,
Golland, & Kanwisher, 2012; Downing, Chan, Peeling,
Dodds, & Kanwisher, 2006), but these studies only exam-
ined responses in posterior temporal regions.
This study attempts to resolve the aforementioned
limitations by measuring the profile of responses in the
functionally defined human ATFP to a wide range of visu-
ally presented stimulus categories. A further aim is to
compare this profile with that of more posterior face-
selective regions (OFA, FFA) in an effort to reveal how
categorical information (particularly about people)
emerges over the span of the temporal lobes.
To begin addressing these aims, in Experiment 1, nous
used a blocked-design functional localizer to first identify
the ATFP, as well as OFA and FFA, in individual partici-
pants. We used a simultaneous odd-one-out visual dis-
crimination (“oddity”) task as a localizer. This task was
selected on the grounds that it has been found to be
effective in previous fMRI research at selectively acti-
vating anterior temporal regions (Barense, Henson, Lee,
& Graham, 2010; O’Neil, Cate, & Köhler, 2009; Lee,
Scahill, & Graham, 2008) and that performance on this
paradigm is sensitive to selective lesions of anterior
temporal regions (c'est à dire., perirhinal cortex [PrC]) in mon-
keys (Buckley, Booth, Rolls, & Gaffan, 2001) and humans
(Barense, Gaffan, & Graham, 2007; Lee et al., 2005). Dans
the main experiment, the same participants were pre-
sented with images of items of 20 different kinds in an
event-related design, while they performed a 1-back task
to maintain attention to the stimuli. In this way, we were
able to assess in detail the selectivity profile of ATFP and
compare it with more posterior face-selective regions.
EXPERIMENT 1
Methods
Participants
Twenty healthy postgraduate volunteers (âge moyen =
25 années, range = 22–30 years; 13 femmes) were recruited
from Bangor University. All participants were screened
for MRI exclusion criteria and gave written informed
consent for participation in the experiment, ce qui était
approved by the research ethics committee of the School
of Psychology at Bangor University, United Kingdom.
Materials
Stimuli for the localizer runs (oddity task) consisted of
96 grayscale images of faces, scènes naturelles, and com-
mon handheld objects (Chiffre 1, top). Images were orga-
nized into 32 triplets for each category (Barense et al.,
2010). Each triplet was presented in a triangular forma-
tion, consisting of a pair of foil images and a target image,
on a 1200 × 840 pixel white background. The foil images
were two pictures of the same face, scène, or object taken
from different viewpoints. The target was another image
from the same category as the foil pair and was selected
to be highly similar in appearance to the other pictures.
Thirty-two triplets consisting of three black squares were
also constructed to appear as an active baseline condition.
One of the squares (the target) was slightly larger or smaller
than the other two shapes.
Stimuli for the main experimental runs (1-back task)
consisted of 48 color images from each of 20 different
catégories (Chiffre 1, bottom). Categories consisted of
birds, (headless) human bodies, cars, chairs, clothes,
crystals, faces, fish, flowers, fruit and vegetables, insects,
instruments, (nonhuman) mammals, prepared food, rep-
tiles, spiders, tools, weapons, indoor scenes, and outdoor
scènes (Downing et al., 2006). These stimuli were selected
to capture a range of object category distinctions (c'est à dire.,
animate vs. inanimate, large vs. petit, natural vs. manmade)
that modulate responses in the ventral temporal lobes
(Konkle & Oliva, 2012; Mahon & Caramazza, 2011). Stimuli
were centred on a white 400 × 400 pixel background,
except for scenes, which were cropped to completely fill
the image dimensions.
Procedure
To localize functional ROIs, participants completed four
runs of an oddity task, each comprising 21 blocks of 15 sec.
Blocks 1, 6, 11, 16, et 21 were fixation-only rest con-
ditions. Each of the four stimulus blocks (faces, scènes,
objets, et des formes) was presented once between each
pair of rest blocks. Stimulation blocks consisted of three
oddity trials, chaque 5 sec in duration. Participants indicated
the location of the target stimulus (the odd item out) par
pressing one of three buttons. Block order for each set of
stimulation conditions was randomly determined between
Harry et al.
1179
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
un
/
_
j
0
o
0
c
9
n
6
6
_
un
p
_
d
0
0
b
9
oui
6
g
6
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
t
.
/
F
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
and Events 22–24 were assigned to fixation-only rest
conditions. The full sequence was divided into eight
separate runs. For Runs 2–8, the final item from the pre-
ceding run was presented at the beginning of the next
run to reestablish sequence context (fabrication 145 events
per run). Participants were assigned to one of five counter-
balanced sequences. Stimulus and target events were pre-
sented for 300 msec followed by an ISI of 1200 msec that
consisted of a central fixation cross. For rest events, a fixa-
tion cross appeared for 1500 msec. Fixation-only rest
blocks (duration = 16 sec) were presented at the begin-
ning and end of each run.
Localizer runs were interspersed throughout the scan-
ning session so that participants completed one run of
the oddity task after every two runs of the main experi-
mental task.
Brain images were acquired with a Philips Achieva 3.0-T
scanner with a 32-channel head coil. BOLD contrast func-
tional images were collected with a T2*-weighted, gradient
EPI sequence (repetition time = 2000 msec, echo time =
35 msec, flip angle = 90°, field of view = 240 mm × 240 mm,
acquisition matrix = 96 × 96, in-plane resolution =
2.5 mm × 2.5 mm, slice thickness = 2.5 mm, no slice gap).
Volumes consisted of 28 slices angled −30° from the AC–
PC plane to maximize signal over the medial-temporal
lobes. Volumes were positioned to completely cover the
temporal and occipital lobes at the expense of the dorsal
parietal cortex. A high-resolution T1-weighted anatomical
image was also acquired for each participant (3-D magne-
tization prepared rapid gradient-echo sequence; 175 slices,
voxel size = 1 mm isotropic, field of view = 256 mm ×
256 mm, repetition time = 8.4 msec, echo time = 3.8 msec,
flip angle = 8°). Stimuli were displayed on a Cambridge
Research Systems BOLDScreen located behind the
scanner bore and were viewed via a mirror fixed to the
head coil. Presentation of the stimuli was controlled by
Psychtoolbox (Brainard, 1997) running on MATLAB (Le
MathWorks, Natick, MA).
Chiffre 1. Example of stimuli presented in Experiment 1: localizer task
(top) and main experimental task (bottom).
Image Preprocessing and Analysis
runs and counterbalanced within runs according to a Latin
square design.
For the main experimental runs, all images from each
of the 20 stimulus categories were presented once in a
rapid, event-related design. Participants completed a
1-back task by pressing a button whenever a stimulus
was immediately repeated. Stimulus order was deter-
mined with a first-order counterbalanced, optimized,
n = 24, Type 1, Indice 1 séquence (Aguirre, 2007). Ce
procedure generated a sequence of 1153 events, inclure-
ing an initial event to establish sequence context. Le
20 stimulus categories were assigned to Event types
1–20 for each participant. Event 21 was assigned to tar-
get events, whereby the previous item was repeated,
Functional MRI data were preprocessed with SPM8
(Wellcome Department of Imaging Neuroscience, Londres,
ROYAUME-UNI; www.fil.ion.ucl.ac.uk/spm/software/spm8/ ) et en-
cluded rigid body realignment, coregistration, tissue seg-
mentation, normalization to the Montreal Neurological
Institut (MNI) 152 template with DARTEL (Ashburner,
2007) and spatial smoothing (6-mm FWHM Gaussian
kernel).
We localized face-selective regions for each individual
with data collected from the oddity task. Estimates of the
BOLD response in each voxel and category were derived
by entering the boxcar function of stimulation that was
convolved with the canonical hemodynamic response
into a fixed effects general linear model. Face selectivity
in each voxel was calculated by contrasting activity evoked
by faces against the average of scenes and objects.
1180
Journal des neurosciences cognitives
Volume 28, Nombre 8
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
un
/
_
j
0
o
0
c
9
n
6
6
_
un
p
_
d
0
0
b
9
oui
6
g
6
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
F
/
.
t
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Tableau 1. Mean MNI Peak Coordinates for Each ROI
Mean MNI Coordinates
X
42.4
−41.1
43.3
−41.9
37.3
−38.5
oui
−78.0
−80.9
−49.6
−51.6
−14.0
−13.2
z
−6.1
−7.63
−20.5
−21.0
−38.5
−33.0
X
6.7
6.4
3.8
3.1
5.0
4.3
SD
oui
6.1
4.8
5.7
4.9
2.5
5.8
z
4.2
5.9
4.2
2.7
7.6
7.4
rOFA
lOFA
rFFA
lFFA
rAFP
lAFP
Face-selective ROIs were localized by finding the most
face-selective voxel within expected regions of cortex
(OFA, inferior or mid-occipital gyrus; FFA, mid-fusiform
gyrus; ATFP, anterior occipito-temporal sulcus or anterior
collateral sulcus) near to typical MNI coordinates identi-
fied in previous studies ( Julian, Fedorenko, Webster, &
Kanwisher, 2012; right OFA [rOFA]: 44, −76, −12; gauche
OFA [lOFA]: −40, −76, −18; right FFA [rFFA]: 38,
−42, −22; left FFA: −40, −52, −18; Axelrod & Yovel,
2013; right ATFP [rATFP]: 34, −10, −39; left ATFP:
−34, −11, −35). ROIs were defined by selecting all sig-
nificant ( p < .001, uncorrected), contiguous voxels cen-
tered around the peak voxel closest to the coordinates
provided above. For analyses of response profiles in the
main experiment, ROI size was limited to 50 voxels be-
cause previous studies have shown that regions larger
than this do not fully capture category selectivity (Mur
et al., 2012).
Estimates of the response to each of the 20 categories
presented in the 1-back task were modeled separately as
instantaneous neural events (i.e., duration = 0 msec)
convolved with the canonical hemodynamic response. An
additional nuisance regressor of no interest was included
to model responses to the initial trial and to all target
trials. The values of the beta estimates for each category
were averaged over all voxels included in each ROI.
Results
Behavioral Performance
Average performance on the oddity task was 81% (SEM =
3%) correct for faces, 83% (SEM = 3%) correct for scenes,
82% (SEM = 3%) correct for objects, and 76% (SEM =
3%) correct for shapes. One-way ANOVA showed no sig-
nificant differences in performance between categories of
stimuli ( p > .2). Average performance on the 1-back task
était 83% (5%).
ized in 15 de 20 participants. Localizing the ATFP is
problematic because of signal loss in the anterior tem-
poral lobes (because of proximity to air-filled spaces such
as the ear canal and the sinus cavity), and finding this
region in 60–75% of participants is consistent with pre-
vious studies that used a single-session protocol (Axelrod
& Yovel, 2013; Rajimehr et al., 2009). FFA was localized
bilaterally in all 20 participants, whereas lOFA or rOFA
was localized in 19 de 20 participants (bilaterally in 18 par-
ticipants). Mean (±SD) peak coordinates for each ROI
are presented in Table 1. Individual MNI coordinates
for the peak voxels within the ATFP are provided in
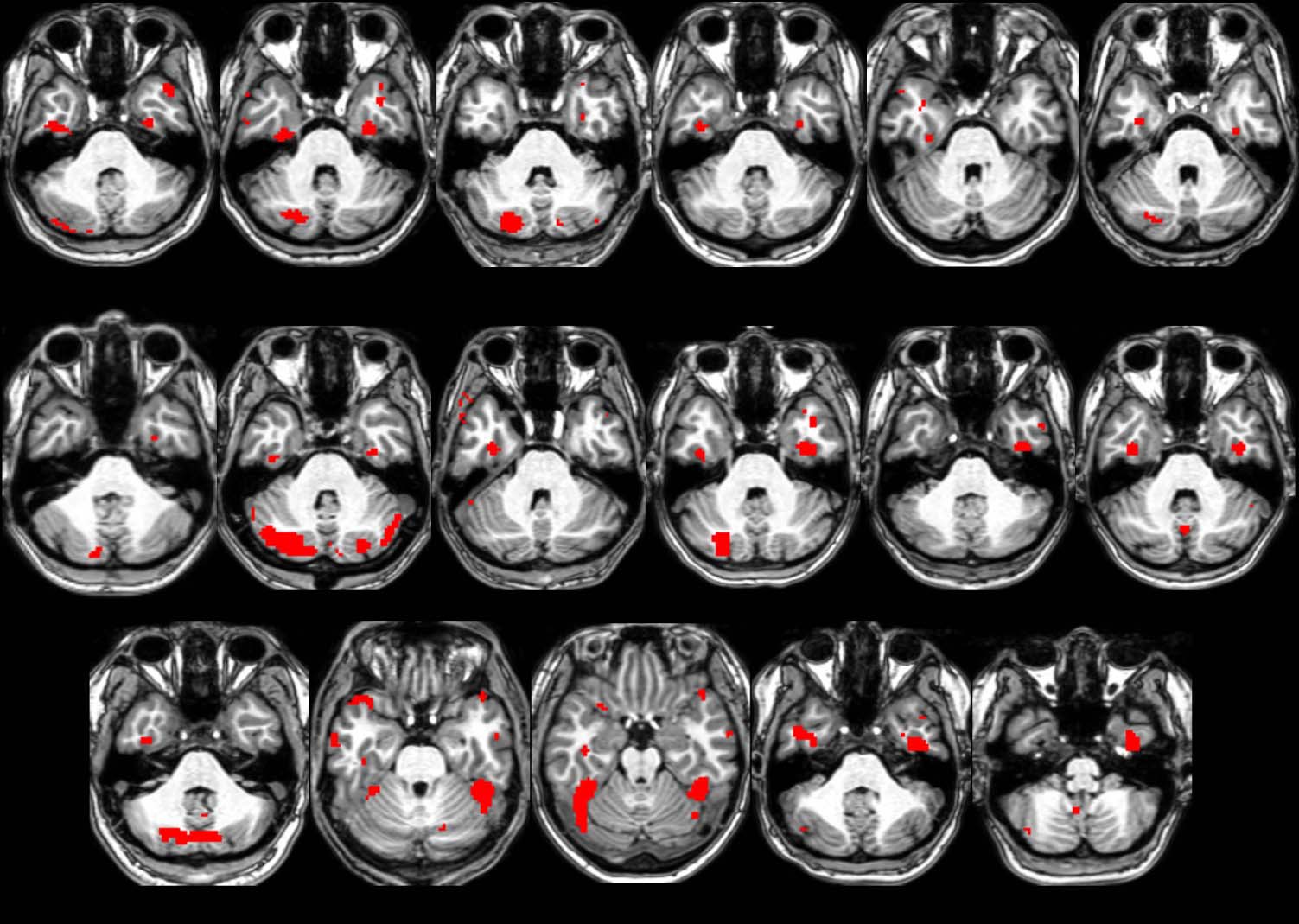
Tableau 2. Chiffre 2 illustrates the location of the ATFP in
four representative participants.
Event-related Response Profiles
De la 20 stimulus categories tested in the main experi-
ment, faces evoked the maximal response in all of the
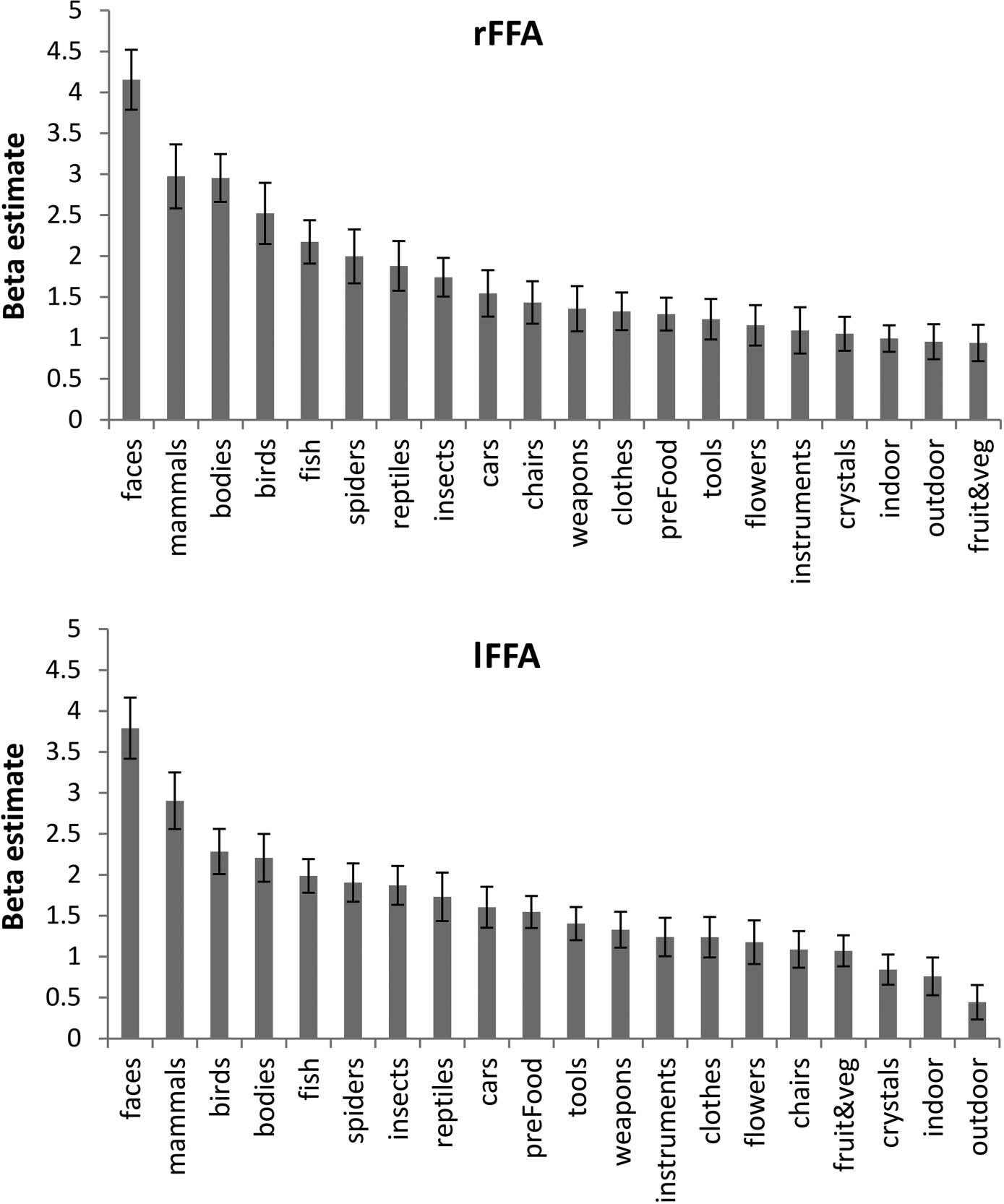
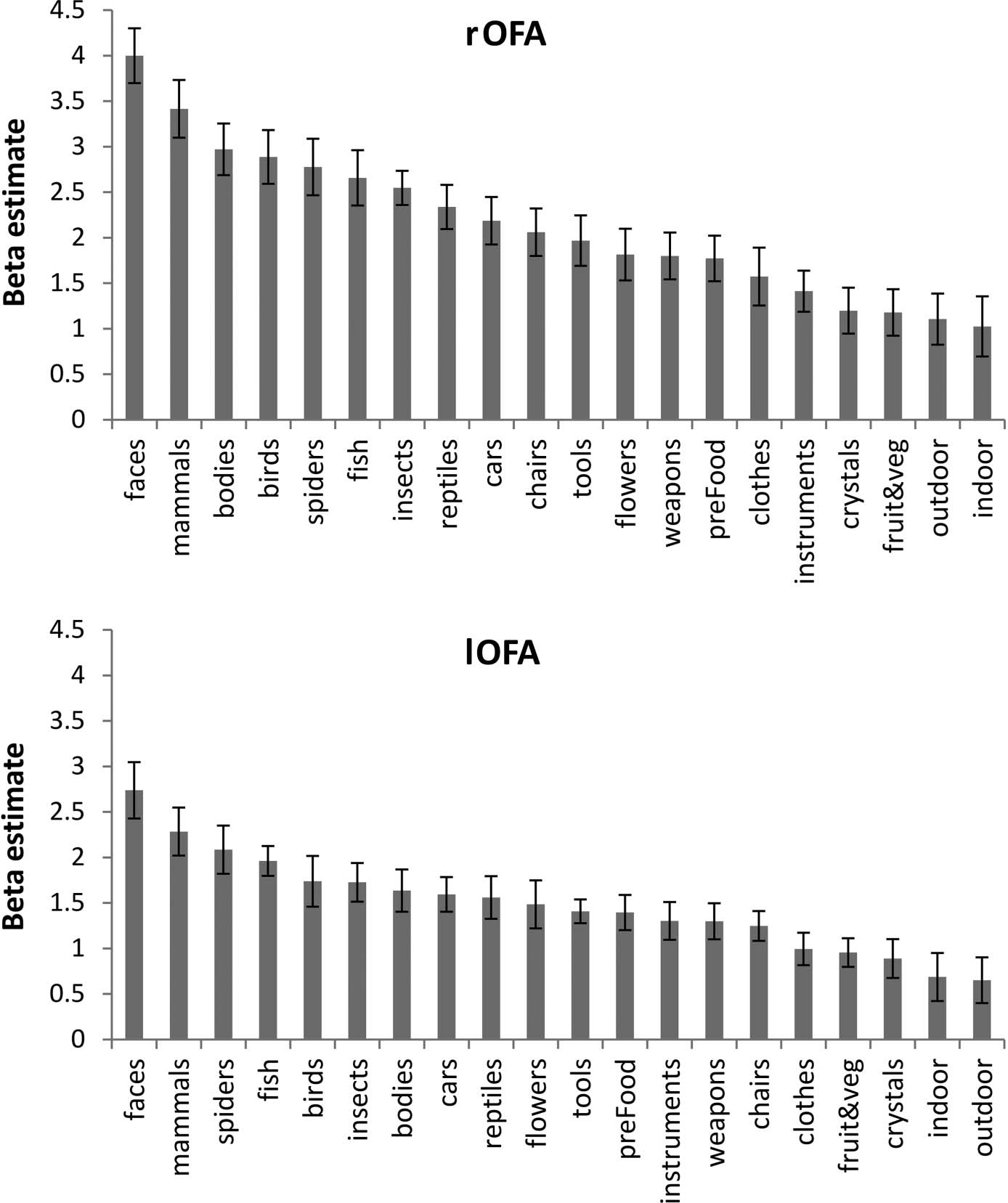
independently defined functional ROIs (OFA, Chiffre 3;
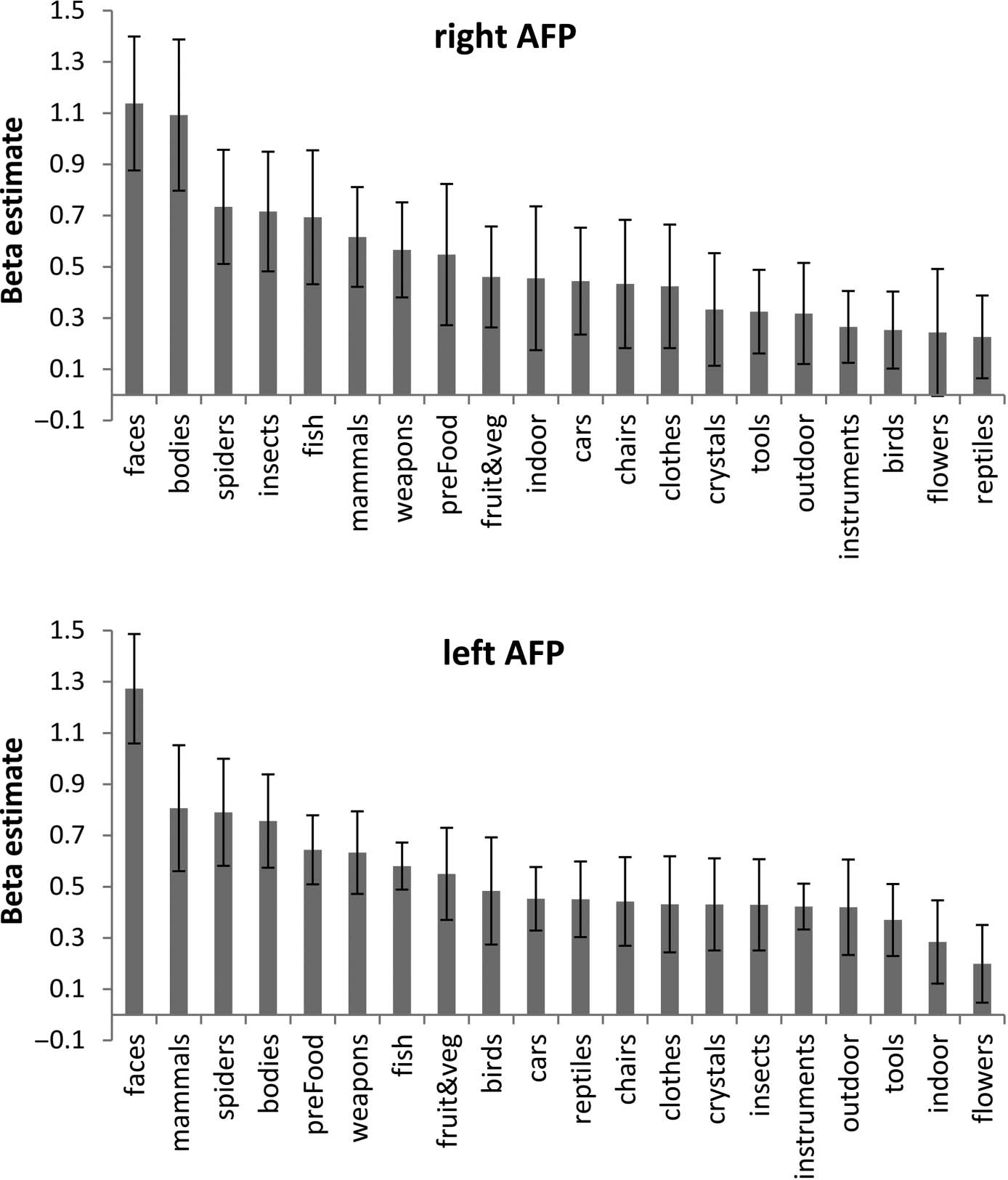
FFA, Chiffre 4; ATFP, Chiffre 5). In line with previous fMRI
studies of category selectivity (Mur et al., 2012; Downing
Tableau 2. Individual MNI Peak Coordinates for the Left and
Right AFP
Left AFP
Right AFP
Participant
X
oui
z
1
2
3
4
5
6
8
11
12
13
14
15
16
17
18
19
20
−40
−12.5 −35
−37.5 −20
−22.5
−
−
X
40
30
45
oui
z
−17.5 −22.5
−15
−37.5
−17.5 −22.5
−12.5 −30
32.5 −12.5 −42.5
−22.5 −32.5
−7.5 −32.5
−37.5 −17.5 −25
−12.5 −45
−
−
35
45
−
−
−15
−15
−
−
−37.5
−42.5
−10
−32.5
37.5 −12.5 −50
−17.5 −32.5
37.5 −10
−40
−5
−47.5
37.5 −12.5 −42.5
−37.5 −12.5 −32.5
−37.5
−7.5 −42.5
40
−
−12.5 −40
−
−
−
−
−
27.5 −12.5 −40
−42.5 −17.5 −22.5
−5
−35
−
40
−
−
−12.5 −40
−17.5 −27.5
37.5 −17.5 −42.5
−
−40
−30
−35
−45
−35
−40
−35
−45
−40
Definition of ROIs
The right-hemisphere ATFP was localized in 13 de 20 par-
ticipants, and the left anterior face patch (AFP) was local-
Mean
SD
−38.5 −13.2 −33.0
37.3 −14.0 −38.5
4.3
5.8
7.4
5
2.5
7.6
Harry et al.
1181
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
un
/
_
j
0
o
0
c
9
n
6
6
_
un
p
_
d
0
0
b
9
oui
6
g
6
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
/
F
.
t
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Chiffre 2. Location of the AFP
(red; faces > scenes + objets,
p < .001).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
et al., 2006), we expected significantly stronger responses
to faces compared with all other categories. To test the
selectivity of each ROI, we compared the response to faces
against the response to the next most effective stimulus
category in a 3 × 2 × 2 repeated-measures ANOVA with
ROI, Hemisphere, and Stimulus category as factors. This
analysis revealed a significant three-way interaction be-
tween ROI, Hemisphere, and Stimulus category (F(2, 18) =
4.5, p < .05), a significant two-way interaction between
ROI and Stimulus category (F(2, 18) = 6.03, p < .01),
and significant main effects for ROI (F(2, 18) = 29.83, p <
.001), Hemisphere (F(1, 9) = 11.70, p < .01), and Stimulus
category (F(1, 9) = 8.19, p < .05).
To interpret the three-way interaction, we carried out
two separate 3 × 2 repeated-measures ANOVAs for each
hemisphere with ROI (OFA, FFA, ATFP) and Stimulus
category (face vs. next best category) as factors. For the
left hemisphere, this analysis found only significant main
effects of ROI (F(2, 26) = 22.75, p < .001, Bonferroni-
corrected) and Stimulus category (F(1, 13) = 9.34, p <
.05, Bonferroni-corrected). Analysis of the right hemi-
sphere revealed a significant interaction between ROI
and Stimulus category (F(2, 24) = 8.1, p < .01, Bonferroni-
corrected) and significant main effects of ROI (F(2, 24) =
17.97, p < .001, Bonferroni-corrected) and Stimulus cate-
gory (F(1, 12) = 10.18, p < .01, Bonferroni-corrected).
Simple effects analysis revealed a main effect of stimulus
category in the rOFA (F(1, 18) = 7.2, p < .05, Bonferroni-
corrected) and FFA (F(1, 19) = 7.2, p < .05, Bonferroni-
corrected), but not in the rATFP (F(1, 13) = 0.15).
Post hoc tests revealed that the response averaged across
faces and bodies in the rATFP was significantly higher
than the next most effective category (spiders; F(1, 13) =
9.3, p < .01).
A final analysis examined whether the structure of
nonpreferred responses in the ATFP was similar to that
found in the posterior face-selective regions. It is a
known property of the latter regions that they generally
respond more strongly to animate compared with inani-
mate categories of stimuli (Wiggett, Pritchard, & Downing,
2009; Downing et al., 2006). Thus, for each ROI, we com-
pared the response averaged over all nonhuman ani-
mate categories (birds, fish, insects, mammals, reptiles)
against the response averaged over all inanimate categories
(cars, chairs, clothes, crystals, flowers, fruit and vegetables,
instruments, prepared foods, tools, weapons). To test the
preference of each ROI, we compared the response to
animates and inanimates in a 3 × 2 × 2 repeated-measures
ANOVA with ROI, Hemisphere, and Animacy (animate,
inanimate) as factors. This analysis revealed significant two-
way interactions between ROI and Animacy (F(2, 18) = 18.7,
p < .001) and between Hemisphere and Category (F(2,
9) = 10.4, p = .01) and significant main effects for ROI
(F(2, 18) = 29.01, p < .001), Hemisphere (F(1, 9) = 6.8,
p < .05), and Animacy (animates > inanimates, F(1, 9) =
32.6, p < .001).
To interpret the interaction effects, we carried out three
separate 2 × 2 repeated-measures ANOVAs for each ROI
(OFA, FFA, ATFP) with Hemisphere and Animacy as fac-
tors. This analysis found only significant main effects for
Animacy in the FFA (F(1, 19) = 59.13, p < .001, Bonferroni-
corrected) and ATFP (F(1, 10) = 8.7, p < .05, Bonferroni-
corrected). In contrast, analysis of the OFA revealed a
significant main effect of Hemisphere (F(1, 18) = 11.84,
1182
Journal of Cognitive Neuroscience
Volume 28, Number 8
p < .01, Bonferroni-corrected) and Animacy (F(1, 18) =
62.3, p < .001, Bonferroni-corrected) and a two-way inter-
action between Hemisphere and Animacy (F(1, 18) =
11.13, p < .01, Bonferroni-corrected). Simple effects anal-
ysis revealed a significant effect of Animacy in both the
rOFA (F(1, 18) = 54.6, p < .001, Bonferroni-corrected)
and lOFA (F(1, 18) = 40.0, p < .001, Bonferroni-corrected).
Discussion
Face-selective regions across the temporal lobes showed
a similar profile of activity, across a wide range of stimu-
lus kinds, consistent with a model in which these regions
cooperate functionally (Moeller et al., 2008). There was
variation, however, in the pattern of responses across the
regions tested; in particular, the right-hemisphere ATFP
showed a strong response to both faces and bodies (statis-
tically different from the response to spiders, the next most
effective category) despite the fact that the ROI was local-
ized independently on the basis of a contrast of faces
versus scenes and objects. This pattern of colocalized sig-
nificant response to bodies and faces, along with a weaker
response to other kinds of objects, is highly similar to that
found in the right fusiform gyrus. In that region, strong
fMRI responses to faces and bodies have been found to
overlap closely ( Weiner & Grill-Spector, 2010; Peelen &
Downing, 2005). One account of this general finding is
that it reflects the common co-occurrence of faces and
bodies in the visual input and the need to jointly process
the socially relevant information they provide (Peelen &
Downing, 2007).
Several studies have attempted to determine whether
the fusiform face- and body-selective responses reflect a sin-
gle neural system or rather two distinct ones. Schwarzlose,
Baker, and Kanwisher (2005) used high-resolution imag-
ing to show that, in many participants, alongside “shared”
voxels that respond to both categories, it is possible to
identify distinct, but adjacent, highly selective patches
for faces and bodies, referring to these accordingly as
FFA and “fusiform body area” (FBA). Another approach
Figure 3. Selectivity profile for
rFFA (top) and lFFA (bottom).
Error bars show the SEM.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Harry et al.
1183
Figure 4. Selectivity profile for
rOFA (top) and lOFA (bottom).
Error bars show the SEM.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
tests for distinct neural systems at the pattern level, with-
out requiring that they be identified in a binary fashion
with separate sets of voxels. The logic of this method is
that overlapping voxels (at whatever resolution) need
not reflect shared neural processes—an assumption com-
monly made in fMRI research (Peelen & Downing, 2007).
For example, in a region where there are overlapping
but functionally distinct face and body representations,
local patterns of selectivity to these two categories should
be uncorrelated (or negatively correlated). That is, con-
sidered across a set of voxels, variability in the selectivity
for bodies would not be expected to relate systematically
to variability in the selectivity for faces. In contrast, where
there are two overlapping and integrated representa-
tions, the variability in selectivity to these two categories
would be expected to be related across voxels: Strong
selectivity to one category should tend to predict strong
selectivity to the other. This would result in a positive
correlation between the local patterns of selectivity
evoked by each category. Studies taking this approach
have found evidence for independent fusiform face- and
body-selective representations (Kim, Lee, Erlendsdottir,
& McCarthy, 2014; Weiner & Grill-Spector, 2010; Peelen,
Wiggett, & Downing, 2006; see also Downing, Wiggett, &
Peelen, 2007).
Thus, motivated by these previous findings in the extra-
striate cortex and by our present results in the right-
hemisphere ATFP, in Experiment 2, we used the multivoxel
approach described above to examine whether faces and
bodies recruit overlapping or segregated representations
in the anterior temporal lobes and in the fusiform gyrus.
EXPERIMENT 2
We localized face- and body-selective regions (FFA,
ATFP, FBA) in the right-hemisphere mid-fusiform and
anterior temporal regions with a blocked 1-back design.
Only responses in the right hemisphere were examined,
as the rATFP demonstrated strong responses to both
faces and bodies in Experiment 1. We opted for a block
1184
Journal of Cognitive Neuroscience
Volume 28, Number 8
design 1-back localizer task in Experiment 2 because a
pilot oddity task including headless bodies was too dif-
ficult for participants to complete. Then, we assessed
the functional responses in each ROI to six different
conditions (faces, whole bodies, body parts, mammals,
foods, and tools) with an event-related design. These
categories were selected to assess responses across
animate, natural, and manmade objects. Moreover, body
parts were included to examine the breadth of body
responses in ATFP. We improved the experimental pro-
tocol with a coronal slice orientation, which, compared
with axial orientation (as used in Experiment 1), has been
shown to maximize signal over the anterior temporal
lobes (Axelrod & Yovel, 2013), and we obtained higher-
resolution images (voxel size = 2 mm3) to mitigate partial
voluming effects.
Bangor University. All participants were screened for MRI
exclusion criteria, after which they gave written informed
consent for participation in the experiment, which was
approved by the research ethics committee of the School
of Psychology at Bangor University, United Kingdom.
Materials
Stimuli for the localizer consisted of 40 images each of
faces, bodies, and chairs (Downing et al., 2006). Stimuli
for the main experiment consisted of 24 images each of
faces, (headless) bodies, body parts, (nonhuman) mam-
mals, food, and tools (Figure 6). All images were pre-
pared in a similar manner as Experiment 1. None of the
images presented in the localizer task appeared in the
event-related runs.
Methods
Procedure
Ten healthy postgraduate volunteers (mean age = 25 years,
range = 24–29 years; six women) were recruited from
Participants completed five runs of a 1-back localizer task,
each consisting of 25 blocks. Blocks 1, 5, 9, 13, 17, 21,
Figure 5. Selectivity profile for
right (top) and left (bottom)
AFP. Error bars show the SEM.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Harry et al.
1185
Figure 6. Example stimuli
presented in Experiment 2.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
and 25 were fixation-only rest conditions lasting 10 sec in
duration. Each of the three stimulus blocks (faces, bod-
ies, and chairs) was presented once between each pair of
rest blocks. Stimulation blocks were composed of 15
stimulus exemplars drawn from a pool of 40 images.
Stimuli were presented sequentially and appeared for
300 msec followed by a 700-msec ISI. Repetitions oc-
curred twice per block.
For the main experimental runs, all images from each
of the six stimulus categories were presented in an
event-related design; each stimulus was presented once
per run. Participants performed a 1-back task. Stimulus
order for each run was determined with a first-order
counterbalanced, optimized, n = 8, Type 1, Index 1 se-
quence. This procedure generated a sequence of 193
events of eight types, including an initial event to estab-
lish sequence context. Event types 1–6 were assigned to
the six stimulus classes, Event type 7 was assigned to
target events (1-back, stimulus repetition), and Event
type 8 was assigned to fixation-only rest condition. Par-
ticipants completed three runs of the main experiment,
resulting in 72 trials per condition. Stimulus and target
events were presented for 300 msec followed by an ISI
of 1700 msec consisting of a fixation cross. For rest
events, a fixation cross was presented for 2000 msec.
Fixation blocks (duration = 16 sec) were presented at
the beginning and end of each run. Runs of the localizer
and the main experiment were completed in an alter-
nating sequence.
Brain images were acquired with a Philips Achieva
3.0-T scanner with a 32-channel head coil. BOLD contrast
functional images were collected with a T2*-weighted,
gradient EPI sequence (repetition time = 2500 msec,
echo time = 35 msec, flip angle = 90°, field of view =
240 mm × 240 mm, acquisition matrix = 120 × 120,
in-plane resolution = 2 mm × 2 mm, slice thickness =
2 mm, no slice gap). Volumes were composed of 28
slices in coronal orientation that were split into two sep-
arate stacks of 14 slices to cover the anterior temporal
lobes and the mid-fusiform gyrus (Axelrod & Yovel,
2013). A high-resolution T1-weighted anatomical image
was also acquired for each participant. All other as-
pects of the experimental setup were the same as in
Experiment 1.
Image Preprocessing and Analysis
Preprocessing was similar to Experiment 1 except that, to
better preserve the local spatial patterns of brain activity,
images were not normalized and spatial smoothing was
performed with a 3-mm Gaussian kernel. Responses to
each category in the blocked localizer and event-related
runs were derived in a similar manner as Experiment 1.
To evaluate mean univariate response profiles in selective
regions similar to Experiment 1, the FFA, ATFP, and FBA
were defined from the localizer by contrasting each cate-
gory against chairs (faces > chairs, bodies > chairs; p <
.001, uncorrected).
For the pattern analysis, localizer data were further
used to identify two functional ROIs in both the mid-
fusiform and anterior temporal lobes for the purpose of
pattern analyses. First, a broad “human form” selective
ROI was defined as the union of all face- and body-
selective voxels within the mid-fusiform and collateral
sulcus. This combined ROI was examined to ensure that
the results of the pattern analysis would not be biased
toward one stimulus category owing to unbalanced voxel
selection. Second, we examined only the face-selective
voxels corresponding to the ATFP. For this analysis, we
defined the ATFP, as in Experiment 1, as the 30 most
face-selective voxels (i.e., faces > bodies + chairs, p <
.001) that were contiguous with the peak voxel residing
within the collateral sulcus. For both ROIs, an indepen-
dent measure of face, whole body, and body part selec-
tivity was calculated from the main experiment data by
contrasting the response of each of these conditions
against the average of mammals, food, and tools. The
resultant t values were extracted for all voxels residing
within each ROI. For each participant and ROI, the ex-
tracted pattern of t values quantifying face selectivity was
correlated with the corresponding pattern of t values for
whole bodies and body parts.
1186
Journal of Cognitive Neuroscience
Volume 28, Number 8
Results
Behavioral Performance
Average performance in the localizer scans was 75.1%
(SEM = 4.2%), and average performance in the main ex-
periment was 78.8% (SEM = 7.5%).
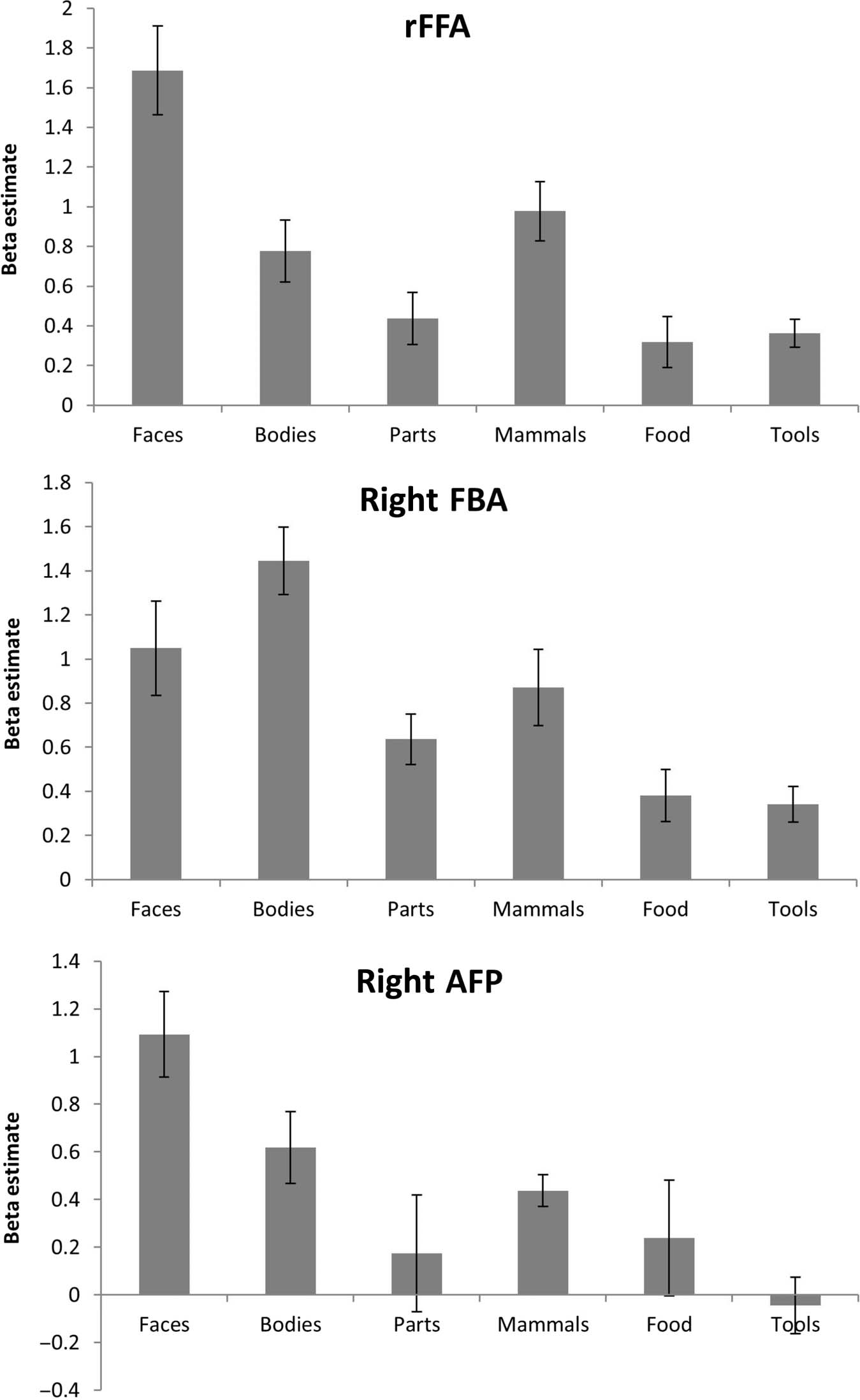
Univariate Analysis of Main Experiment
The three independently defined ROIs (rFFA, right FBA,
and rATFP) all showed maximal responses to the ex-
pected categories: faces in FFA and ATFP and whole
bodies in FBA (Figure 7). To examine whether the pre-
ferred category evoked significantly more activity than
all other stimuli, the responses to the preferred category
and the next most effective category were entered into a
3 × 2 repeated-measures ANOVA with ROI (FFA, FBA,
ATFP) and Stimulus category (preferred category vs. next
most effective) as factors. For the FFA and the ATFP, re-
sponses to faces were compared against mammals and
bodies, respectively, whereas in the FBA, bodies were
Figure 7. Profile of responses
to six stimulus categories,
for the rFFA (top), right FBA
(center), and right AFP
(bottom). Error bars show
the SEM.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Harry et al.
1187
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
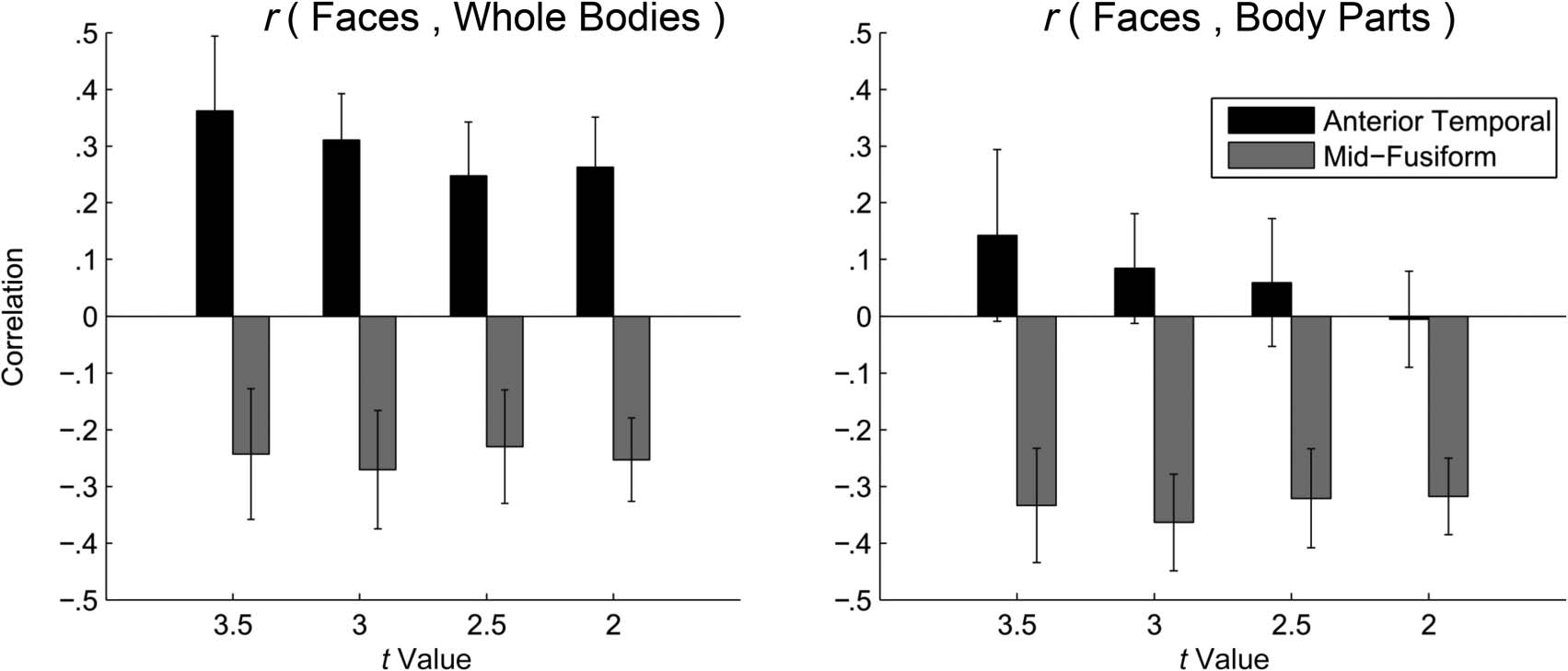
Figure 8. Correlation between face and body selectivity (left) and face and body-part selectivity (right) for the union of all independently defined
face- and body-selective voxels in the mid-fusiform and anterior temporal lobes. Correlations shown were obtained for a range of thresholds for
defining face- and body-selective voxels (x axis). These findings suggest that, in the anterior temporal cortex, face and body representations are
integrated, in contrast to the fusiform gyrus, where they appear to remain distinct. Error bars show the SEM.
compared with faces. This analysis revealed only a signif-
icant main effect of Stimulus category (F(1, 7) = 53.69,
p < .001), indicating that faces and whole bodies evoked
more activity than the next most effective stimulus cate-
gory in face- and body-selective regions, respectively.
Pattern Analysis
We performed a pattern analysis to examine the relation-
ship of face- and body-selective populations in both the
right-hemisphere mid-fusiform and anterior temporal
lobes. In the first analysis, we examined all voxels that
showed a preference for either faces or bodies by taking
the union of the face- and body-selective regions as de-
fined by the localizer runs. These regions were defined
for a range of thresholds (tROI > 3.5, 3, 2.5, 2) to ensure
that the results of the pattern analysis were not depen-
dent on how stringently voxels were selected. Separately
for the mid-fusiform and anterior temporal ROIs, voxel-
wise patterns of selectivity in the main experiment for
faces and for whole human bodies (faces > mammals +
nourriture + tools, bodies > mammals + nourriture + tools) étaient
correlated for each participant. Single-sample t tests
comparing the Fisher-transformed correlations in each
region (Chiffre 8, gauche) showed that face and body selec-
tivity were negatively correlated in the mid-fusiform ROI
(significantly at thresholds tROI > 3, p = .0321; TROI > 2,
p < .01, Bonferroni-corrected). This suggests, consistent
with previous findings (Kim et al., 2014; Peelen et al.,
2006), that face and body representations remain distinct
in the fusiform gyrus—faces and bodies elicit distinct
patterns of local activity. In contrast, significantly positive
correlations between face and body selectivity were
observed in the anterior temporal lobes (rs range =
0.24–0.36, all ps at different selection thresholds < .05,
Bonferroni-corrected), suggestive of a shared face and
body representation.
In principle, however, the positive pattern correlation
between face and body selectivity in the anterior tem-
poral lobe could be because of general factors affecting
responses in this region (i.e., signal limitations, use of
common baseline), rather than a specific property of face
and body representations. To exclude this possibility, we
performed the same analysis comparing patterns of face
and body-part selectivity (Figure 8, right). This analysis
revealed that face and body-part selectivity were nega-
tively correlated in the mid-fusiform region (all ps <
.01, Bonferroni-corrected). Critically, face and body-part
selectivity were not significantly correlated in the anterior
temporal lobes (all ts < 1). Furthermore, in the anterior
temporal region, the correlation between face and whole-
body selectivity was significantly greater than the correla-
tion between face and body-part selectivity ( ps < .05,
Bonferroni-corrected). Therefore, it is not simply the
case that response patterns evoked by any and all visual
stimuli are positively correlated within the anterior tem-
poral region.
Although examining the union of all face- and body-
selective voxels is an unbiased method of voxel selection,
this approach is less directly comparable with previous
studies of the human ATFP. Therefore, in a second pat-
tern analysis, we limited the analyzed region to only the
30 most face-selective voxels (selected from the localizer
data) located within the collateral sulcus. Overall, the cor-
relations observed for the ATFP were similar to those
found in the first pattern analysis. The mean correlation
between face and whole-body selectivity was significantly
above zero (rmean = .38, t = 4.73, p < .01, Bonferroni-
corrected), whereas the mean correlation between faces
and body parts was not (rmean = .08, t = 0.86). Moreover,
1188
Journal of Cognitive Neuroscience
Volume 28, Number 8
the correlation between face and whole-body selectivity
was greater than between faces and body parts (t =
8.05, p < .0001, Bonferroni-corrected), indicating that
the spatial organization of face- and body-selective re-
sponses observed for the combined ROI is also present
in the ATFP.
Discussion
In Experiment 2, we did not replicate the finding of
Experiment 1 that faces and headless bodies drive activity
in the rATFP equally well, perhaps because, in this study,
we scanned at a higher spatial resolution and optimized
slice selection, resulting in more precise localization of
highly face-selective voxels in the anterior temporal cor-
tex. The multivoxel pattern analysis showed, however,
that, in this region, there is a significant positive correla-
tion between the activity patterns evoked by face and
whole-body stimuli. Importantly, this was significantly
greater than the relationship between faces and indi-
vidual body parts, which was not greater than chance,
indicating that the positive correlation was not because
of general properties of the responses observed in this
region. Furthermore, these results were distinct from
the findings from the mid-fusiform region, where patterns
of face and body selectivity were negatively correlated,
indicating independent (or at least less integrated) encod-
ing of these stimuli. Taken together, these results suggest
that part of the right anterior temporal lobe, in contrast to
the posterior fusiform gyrus, encodes an integrated rep-
resentation of the visual appearance of the face and of
the whole body—a representation that does not extend
to isolated body parts.
GENERAL DISCUSSION
Our study aimed to survey the response profile of the
ATFP (Rajimehr et al., 2009) to different categories of
stimuli. The profile of ATFP across multiple categories
appeared, to a large extent, to mirror that of the FFA
and OFA. A notable finding from Experiment 1 was the
high response to human bodies (without faces) in the
independently defined right-hemisphere ATFP ROI. In
previous work on human anterior temporal face repre-
sentations, Tsao and colleagues (2008) found a reliable
response to bodies that was nonetheless significantly
lower than the response to faces. These findings of colo-
cated strong responses to faces and bodies are another
potentially important similarity between the visual rep-
resentations of the anterior and posterior inferior tem-
poral lobes, where face- and body-evoked activations
are intertwined in the fusiform gyrus (Kim et al., 2014;
Weiner & Grill-Spector, 2010; Peelen & Downing, 2005;
Schwarzlose et al., 2005).
Therefore, in Experiment 2, we used a pattern analysis
to examine the spatial organization of face and whole-
body representations in the anterior and posterior tem-
poral regions. In the fusiform gyrus, the pattern of face
and body selectivity was negatively correlated, consistent
with functionally distinct representations. In contrast, in
the anterior temporal region, the correlation between
patterns evoked by bodies and faces was positive, sug-
gestive of integrated, whole-person representations. This
interpretation is supported by the finding that face-
evoked activity in the rATFP was more similar to whole
bodies compared with isolated body parts, suggesting
that anterior person-selective responses are primarily
driven by whole-person form information.
These findings, considered alongside a recent study
showing super-additive responses to combined face and
whole-body stimuli in the ATFP of the macaque (Fisher
& Freiwald, 2015), provide support for the existence of
integrated whole-agent processing regions in the primate
visual system. In principle, integrated processing is not
required to form whole-agent representations (Afraz,
2015), as whole-agent information is also present in the
distributed response across face- and body-selective re-
gions in the mid-fusiform. Evidence of super-additive re-
sponses and joint selectivity, however, confirms that
integrated representations are indeed formed in the ante-
rior temporal cortex and likely play a key role in visual
recognition (Lehky & Tanaka, 2016).
More specifically, these and other findings provide
some evidence for a hierarchically organized chain of
face- and body-form representations along the length of
the ventral occipito-temporal cortex (Grill-Spector &
Weiner, 2014; Taylor & Downing, 2011; Minnebusch &
Daum, 2009) that become progressively more integrated
with each other (Figure 9). In the posterior occipito-
temporal cortex, face and body representations (OFA,
EBA) are anatomically distinct (Pitcher et al., 2009) and
appear to emphasize the representation of component
parts (e.g., Schiltz et al., 2010; Liu et al., 2009; Taylor,
Wiggett, & Downing, 2007). More anteriorly, the FFA
and FBA encode more holistic properties of their pre-
ferred stimuli (Brandman & Yovel, 2016; Harris & Aguirre,
2010; Schiltz et al., 2010; Liu et al., 2009; Taylor et al., 2007;
Schiltz & Rossion, 2006). A question under active inves-
tigation is whether, in the fusiform regions, there is fur-
ther integrated processing across faces and bodies
(Bernstein, Oron, Sadeh, & Yovel, 2014; Kaiser, Strnad,
Seidl, Kastner, & Peelen, 2014; Song, Luo, Li, Xu, & Liu,
2013; Schmalzl, Zopf, & Williams, 2012), with some evi-
dence both for (e.g., Bernstein et al., 2014) and against
(Fisher & Freiwald, 2015; Kaiser et al., 2014) this proposal.
The current results argue for even closer integration of
face and whole-body representation in the anterior-most
reaches of the temporal cortex. On this view, the anterior
temporal cortex can be seen as a core region in a broadly
defined person-form processing pathway that combines
domain-specific representations that are constructed in
the extrastriate cortex.
This perspective integrates the current findings with
previous behavioral work and suggests how the very
Harry et al.
1189
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
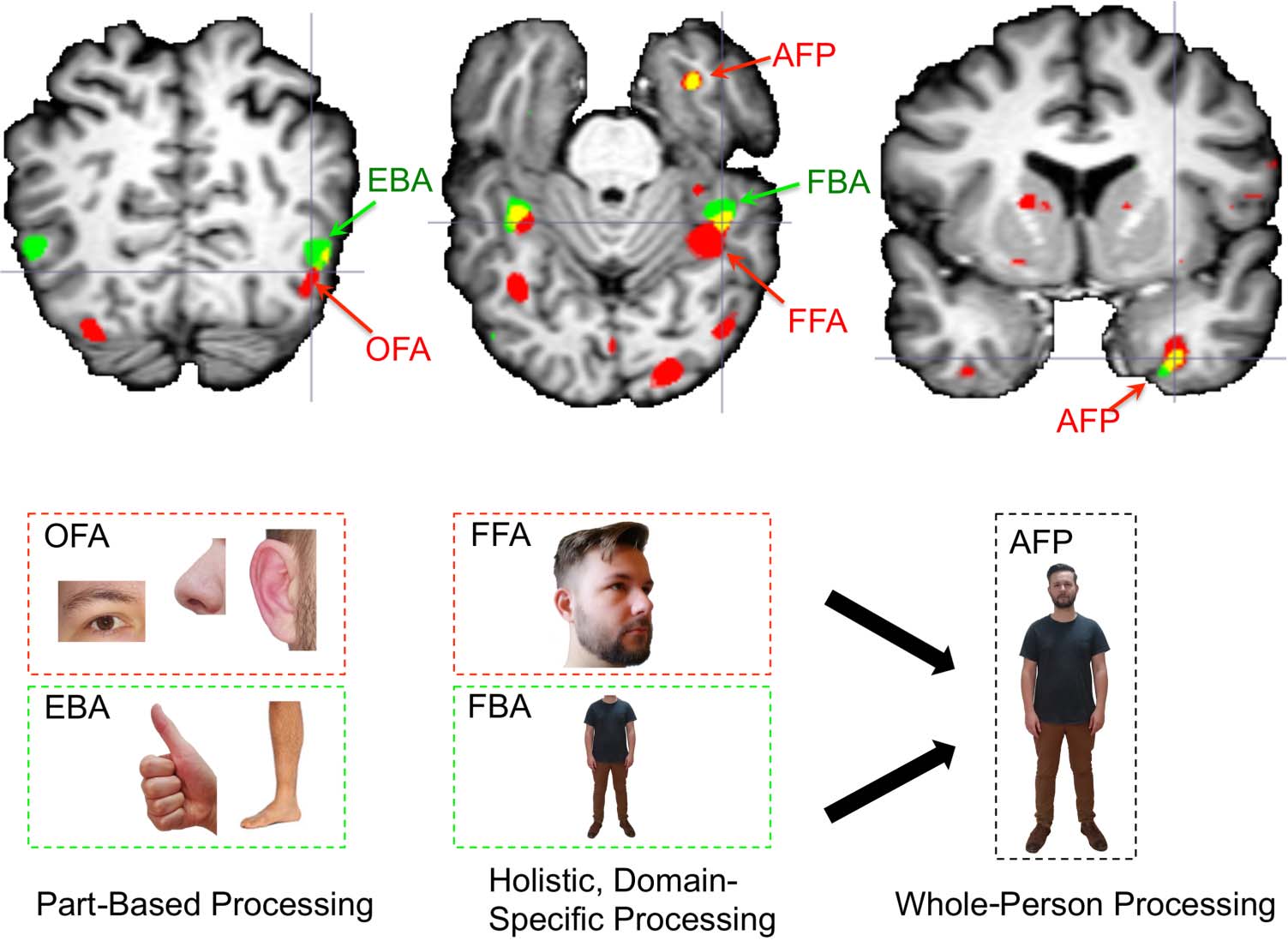
Figure 9. Schematic illustration
of a proposed hierarchical
organization of human form
information in the ventral
temporal lobes. EBA and OFA,
which are anatomically distinct,
engage in domain-specific
part-based processing of their
preferred categories. FFA and
FBA are closely overlapping
and form domain-specific
holistic representations of
faces and bodies, respectively.
Finally, face and body
information is functionally
integrated in the anterior
temporal lobes (including the
region typically identified as
the AFP) to contribute to
whole-person representation.
Red clusters, faces > chairs,
p < .001; green clusters,
bodies > chairs, p < .001.
different kinds of visual cues provided by bodies and
faces are brought together to represent what must be
the real object of interest for social perception: whole
people (Macrae, Quinn, Mason, & Quadflieg, 2005). Evi-
dence from perceptual studies shows that judgments of
identity, emotion, and gender from faces can be strongly
influenced by the state of the body (Rice, Phillips, Natu,
An, & O’Toole, 2013; Aviezer, Trope, & Todorov, 2012).
Moreover, adaptation to pictures of bodies presented in
isolation can alter the perception of subsequently viewed
faces (Ghuman, McDaniel, & Martin, 2010), suggesting
that faces and bodies share processing mechanisms. A
very important open question is to what extent the
pathways described here contribute to the integrated
processing of different facial and bodily cues. Notably,
extensive work by de Gelder and colleagues (2006) points
to other, largely subcortical routes that are involved in
rapidly extracting and integrating emotional information
from faces and bodies (Meeren, van Heijnsbergen, &
de Gelder, 2005). Here, we propose that the ATFP forms
part of a ventral temporal pathway involved in person
perception and identification that integrates static form
cues from across the face and the body.
Evidence of joint face and whole-body selectivity in the
ATFP also aligns with paired object associative responses
found in the PrC (Fujimichi et al., 2010). PrC is an ante-
rior temporal region located in the anterior collateral
sulcus (rhinal sulcus; Suzuki & Naya, 2014), which has
been argued to occupy the highest point in the ventral
visual processing pathway (Murray, Bussey, & Saksida,
2007). PrC receives dense input from visual area TE
(Suzuki & Naya, 2014) and also receives input from
regions of STS containing body-form selective neurons
(Suzuki & Naya, 2014; Oram & Perrett, 1994). In the
macaque, paired-associate learning involving two objects
alters the selectivity of neurons in the PrC such that neu-
rons that are selective for a particular object also become
selective for a paired associate when repeatedly pre-
sented together (Fujimichi et al., 2010). Similar to the
present findings, these “unitized” representations also
show a hierarchical organization, with a gradient of in-
creasingly overlapping responses found spanning area
TE and regions within the PrC (areas A36 and A35;
Hirabayashi et al., 2014; Fujimichi et al., 2010). Given that
the ATFP was primarily observed in the anterior collateral
sulcus, it is possible that the current evidence of integrated
face and body processing reflects a specific instance of a
general PrC process that forms unitized representations
of highly relevant objects via paired-associate coding
mechanisms.
Such a hierarchical scheme accords with proposals
arising from the literature on memory and conceptual
representations. These hold that the PrC is involved in
forming complex, conjunctive object representations by
combining feature-based representations derived in the
extrastriate cortex (Clarke & Tyler, 2014; Graham,
Barense, & Lee, 2010; Barense et al., 2005, 2007; Bartko,
Winters, Cowell, Saksida, & Bussey, 2007; Buckley &
Gaffan, 2006; Lee et al., 2005). According to this account,
functionally integrating domain-specific representations
serve to help buffer the perceptual system against inter-
ference, which has a larger impact on the simpler, part-
based representations found in the extrastriate cortex
( Watson & Lee, 2013; Barense et al., 2012; Bartko,
1190
Journal of Cognitive Neuroscience
Volume 28, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Cowell, Winters, Bussey, & Saksida, 2010; Fujimichi et al.,
2010).
To summarize, this study demonstrated that the ATFP
shares several similarities with other face-selective re-
gions found in the extrastriate cortex. Differences were
evident, however, in the way that face- and body-selective
responses are organized in these distributed temporal
lobe brain areas, with evidence for integrated selectivity
of faces and whole bodies found only in the ATFP. In that
sense, the present findings are consistent with models
that posit a posterior-to-anterior gradient in perceptual
representations of objects that is built on increasingly
complex combinations of features (Lehky & Tanaka,
2016).
Acknowledgments
We thank Morgan Barense for providing the stimuli used in the
oddity task and Richard Ramsey for comments on an earlier
draft. This work was supported by the Biotechnology and
Biological Sciences Research Council grant BB/1007091/1.
Reprint requests should be sent to Bronson B. Harry or Paul E.
Downing, School of Psychology, Bangor University, Brigantia
Building, Bangor, Gwynedd LL57 2AS, United Kingdom, or via
e-mail: b.harry@westernsydney.edu.au, p.downing@bangor.ac.uk.
REFERENCES
Afraz, A. (2015). Head to toe, in the head. Proceedings of the
National Academy of Sciences, U.S.A., 112, 15004–15005.
Aguirre, G. K. (2007). Continuous carry-over designs for fMRI.
Neuroimage, 35, 1480–1494.
Anzellotti, S., Fairhall, S. L., & Caramazza, A. (2014). Decoding
representations of face identity that are tolerant to rotation.
Cerebral Cortex, 24, 1988–1995.
Ashburner, J. (2007). A fast diffeomorphic image registration
algorithm. Neuroimage, 38, 95–113.
Aviezer, H., Trope, Y., & Todorov, A. (2012). Body cues, not
facial expressions, discriminate between intense positive and
negative emotions. Science, 338, 1225–1229.
Axelrod, V., & Yovel, G. (2013). The challenge of localizing
the anterior temporal face area: A possible solution.
Neuroimage, 81, 371–380.
Barense, M. D., Bussey, T. J., Lee, A. C. H., Rogers, T. T., Davies,
R. R., Saksida, L. M., et al. (2005). Functional specialization in
the human medial temporal lobe. Journal of Neuroscience,
25, 10239–10246.
Barense, M. D., Gaffan, D., & Graham, K. S. (2007). The human
medial temporal lobe processes online representations of
complex objects. Neuropsychologia, 45, 2963–2974.
Barense, M. D., Groen, I. I. A., Lee, A. C. H., Yeung, L.-K., Brady,
S. M., Gregori, M., et al. (2012). Intact memory for irrelevant
information impairs perception in amnesia. Neuron, 75,
157–167.
Barense, M. D., Henson, R. N. A., Lee, A. C. H., & Graham, K. S.
(2010). Medial temporal lobe activity during complex
discrimination of faces, objects, and scenes: Effects of
viewpoint. Hippocampus, 20, 389–401.
Bartko, S. J., Cowell, R. A., Winters, B. D., Bussey, T. J., &
Saksida, L. M. (2010). Heightened susceptibility to
interference in an animal model of amnesia: Impairment in
encoding, storage, retrieval—Or all three? Neuropsychologia,
48, 2987–2997.
Bartko, S. J., Winters, B. D., Cowell, R. A., Saksida, L. M., &
Bussey, T. J. (2007). Perceptual functions of perirhinal cortex
in rats: Zero-delay object recognition and simultaneous
oddity discriminations. Journal of Neuroscience, 27,
2548–2559.
Bernstein, M., Oron, J., Sadeh, B., & Yovel, G. (2014). An
integrated face–body representation in the fusiform gyrus
but not the lateral occipital cortex. Journal of Cognitive
Neuroscience, 26, 2469–2478.
Brainard, D. H. (1997). The psychophysics toolbox. Spatial
Vision, 10, 433–436.
Brandman, T., & Yovel, G. (2016). Bodies are represented as
wholes rather than their sum of parts in the occipital–temporal
cortex. Cerebral Cortex, 26, 530–543.
Buckley, M. J., Booth, M. C. A., Rolls, E. T., & Gaffan, D. (2001).
Selective perceptual impairments after perirhinal cortex
ablation. Journal of Neuroscience, 21, 9824–9836.
Buckley, M. J., & Gaffan, D. (2006). Perirhinal cortical
contributions to object perception. Trends in Cognitive
Sciences, 10, 100–107.
Calder, A. J., Lawrence, A. D., & Young, A. W. (2001).
Neuropsychology of fear and loathing. Nature Reviews
Neuroscience, 2, 352–363.
Clarke, A., & Tyler, L. K. (2014). Object-specific semantic coding
in human perirhinal cortex. Journal of Neuroscience, 34,
4766–4775.
Collins, J. A., & Olson, I. R. (2014). Beyond the FFA: The role
of the ventral anterior temporal lobes in face processing.
Neuropsychologia, 61, 65–79.
de Gelder, B. (2006). Towards the neurobiology of emotional
body language. Nature Reviews Neuroscience, 7, 242–249.
Desimone, R., Albright, T. D., Gross, C. G., & Bruce, C.
(1984). Stimulus-selective properties of inferior temporal
neurons in the macaque. Journal of Neuroscience, 4,
2051–2062.
Downing, P. E., Chan, A. W.-Y., Peelen, M. V., Dodds, C. M., &
Kanwisher, N. (2006). Domain specificity in visual cortex.
Cerebral Cortex, 16, 1453–1461.
Downing, P. E., Wiggett, A. J., & Peelen, M. V. (2007). Functional
magnetic resonance imaging investigation of overlapping
lateral occipitotemporal activations using multi-voxel pattern
analysis. Journal of Neuroscience, 27, 226–233.
Duchaine, B., & Yovel, G. (2015). A revised neural framework
for face processing. Annual Review of Vision Science, 1,
393–416.
Fisher, C., & Freiwald, W. A. (2015). Whole-agent selectivity
within the macaque face-processing system. Proceedings
of the National Academy of Sciences, U.S.A., 112,
14717–14722.
Freiwald, W. A., & Tsao, D. Y. (2010). Functional
compartmentalization and viewpoint generalization
within the macaque face-processing system. Science,
330, 845–851.
Fujimichi, R., Naya, Y., Koyano, K. W., Takeda, M., Takeuchi, D.,
& Miyashita, Y. (2010). Unitized representation of paired
objects in area 35 of the macaque perirhinal cortex.
European Journal of Neuroscience, 32, 659–667.
Gauthier, I., Tarr, M. J., Moylan, J., Skudlarski, P., Gore, J. C.,
& Anderson, A. W. (2000). The fusiform “face area” is
part of a network that processes faces at the individual
level. Journal of Cognitive Neuroscience, 12, 495–504.
Ghuman, A. S., McDaniel, J. R., & Martin, A. (2010). Face
adaptation without a face. Current Biology, 20, 32–36.
Graham, K. S., Barense, M. D., & Lee, A. C. H. (2010). Going
beyond LTM in the MTL: A synthesis of neuropsychological
and neuroimaging findings on the role of the medial
temporal lobe in memory and perception. Neuropsychologia,
48, 831–853.
Harry et al.
1191
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Grill-Spector, K., & Weiner, K. S. (2014). The functional
architecture of the ventral temporal cortex and its role in
categorization. Nature Reviews Neuroscience, 15, 536–548.
Harris, A., & Aguirre, G. K. (2010). Neural tuning for face wholes
and parts in human fusiform gyrus revealed by fMRI
adaptation. Journal of Neurophysiology, 104, 336–345.
Haxby, J. V., & Gobbini, M. I. (2011). Distributed neural systems
for face perception. In A. J. Calder, G. Rhodes, M. H. Johnson,
& J. V. Haxby (Eds.), The Oxford handbook of face
perception (pp. 93–110). Oxford, UK: OUP.
Haxby, J. V., Hoffman, E. A., & Gobbini, M. I. (2000). The
distributed human neural system for face perception. Trends
in Cognitive Sciences, 4, 223–233.
Hirabayashi, T., Tamura, K., Takeuchi, D., Takeda, M., Koyano,
K. W., & Miyashita, Y. (2014). Distinct neuronal interactions
in anterior inferotemporal areas of macaque monkeys
during retrieval of object association memory. Journal of
Neuroscience, 34, 9377–9388.
Hung, C.-C., Yen, C. C., Ciuchta, J. L., Papoti, D., Bock, N. A.,
Leopold, D. A., et al. (2015). Functional mapping of
face-selective regions in the extrastriate visual cortex of
the marmoset. Journal of Neuroscience, 35, 1160–1172.
Julian, J. B., Fedorenko, E., Webster, J., & Kanwisher, N. (2012).
An algorithmic method for functionally defining regions
of interest in the ventral visual pathway. Neuroimage, 60,
2357–2364.
Kaiser, D., Strnad, L., Seidl, K. N., Kastner, S., & Peelen, M. V.
(2014). Whole person-evoked fMRI activity patterns in
human fusiform gyrus are accurately modeled by a linear
combination of face- and body-evoked activity patterns.
Journal of Neurophysiology, 111, 82–90.
Kanwisher, N., & Yovel, G. (2006). The fusiform face area: A
cortical region specialized for the perception of faces.
Philosophical Transactions of the Royal Society of London,
Series B, Biological Sciences, 361, 2109–2128.
Kim, N. Y., Lee, S. M., Erlendsdottir, M. C., & McCarthy, G.
(2014). Discriminable spatial patterns of activation for faces
and bodies in the fusiform gyrus. Frontiers in Human
Neuroscience, 8, 632.
Konkle, T., & Oliva, A. (2012). A real-world size organization of
object responses in occipitotemporal cortex. Neuron, 74,
1114–1124.
Ku, S.-P., Tolias, A. S., Logothetis, N. K., & Goense, J. (2011).
fMRI of the face-processing network in the ventral temporal
lobe of awake and anesthetized macaques. Neuron, 70,
352–362.
Lee, A. C. H., Buckley, M. J., Pegman, S. J., Spiers, H., Scahill,
V. L., Gaffan, D., et al. (2005). Specialization in the medial
temporal lobe for processing of objects and scenes.
Hippocampus, 15, 782–797.
Lee, A. C. H., Scahill, V. L., & Graham, K. S. (2008). Activating
the medial temporal lobe during oddity judgment for faces
and scenes. Cerebral Cortex, 18, 683–696.
Lehky, S. R., & Tanaka, K. (2016). Neural representation for
object recognition in inferotemporal cortex. Current
Opinion in Neurobiology, 37, 23–35.
Liu, J., Harris, A., & Kanwisher, N. (2009). Perception of face
parts and face configurations: An fMRI study. Journal of
Cognitive Neuroscience, 22, 203–211.
Macrae, C. N., Quinn, K. A., Mason, M. F., & Quadflieg, S.
(2005). Understanding others: The face and person
construal. Journal of Personality and Social Psychology,
89, 686–695.
Mahon, B. Z., & Caramazza, A. (2011). What drives the
organization of object knowledge in the brain? Trends in
Cognitive Sciences, 15, 97–103.
vision: Diversity, consistency, and spatial sensitivity among
AF face patch neurons. Journal of Neuroscience, 35,
5537–5548.
Meeren, H. K. M., van Heijnsbergen, C. C. R. J., & de Gelder, B.
(2005). Rapid perceptual integration of facial expression and
emotional body language. Proceedings of the National
Academy of Sciences, U.S.A., 102, 16518–16523.
Mende-Siedlecki, P., Verosky, S. C., Turk-Browne, N. B., &
Todorov, A. (2013). Robust selectivity for faces in the human
amygdala in the absence of expressions. Journal of Cognitive
Neuroscience, 25, 2086–2106.
Meyers, E. M., Borzello, M., Freiwald, W. A., & Tsao, D. (2015).
Intelligent information loss: The coding of facial identity,
head pose, and non-face information in the macaque face
patch system. Journal of Neuroscience, 35, 7069–7081.
Minnebusch, D. A., & Daum, I. (2009). Neuropsychological
mechanisms of visual face and body perception.
Neuroscience & Biobehavioral Reviews, 33, 1133–1144.
Moeller, S., Freiwald, W. A., & Tsao, D. Y. (2008). Patches with
links: A unified system for processing faces in the macaque
temporal lobe. Science, 320, 1355–1359.
Mur, M., Ruff, D. A., Bodurka, J., Weerd, P. D., Bandettini, P. A., &
Kriegeskorte, N. (2012). Categorical, yet graded—Single-image
activation profiles of human category-selective cortical
regions. Journal of Neuroscience, 32, 8649–8662.
Murray, E. A., Bussey, T. J., & Saksida, L. M. (2007). Visual
perception and memory: A new view of medial temporal lobe
function in primates and rodents. Annual Review of
Neuroscience, 30, 99–122.
Nasr, S., & Tootell, R. B. H. (2012). Role of fusiform and
anterior temporal cortical areas in facial recognition.
Neuroimage, 63, 1743–1753.
Nestor, A., Plaut, D. C., & Behrmann, M. (2011). Unraveling
the distributed neural code of facial identity through
spatiotemporal pattern analysis. Proceedings of the National
Academy of Sciences, U.S.A., 108, 9998–10003.
O’Neil, E. B., Cate, A. D., & Köhler, S. (2009). Perirhinal
cortex contributes to accuracy in recognition memory and
perceptual discriminations. Journal of Neuroscience, 29,
8329–8334.
Oram, M. W., & Perrett, D. I. (1994). Responses of anterior
superior temporal polysensory (STPa) neurons to “biological
motion” stimuli. Journal of Cognitive Neuroscience, 6,
99–116.
Peelen, M. V., & Downing, P. E. (2005). Selectivity for the human
body in the fusiform gyrus. Journal of Neurophysiology,
93, 603–608.
Peelen, M. V., & Downing, P. E. (2007). The neural basis of
visual body perception. Nature Reviews Neuroscience, 8,
636–648.
Peelen, M. V., Wiggett, A. J., & Downing, P. E. (2006).
Patterns of fMRI activity dissociate overlapping functional
brain areas that respond to biological motion. Neuron,
49, 815–822.
Pinsk, M. A., Arcaro, M., Weiner, K. S., Kalkus, J. F., Inati,
S. J., Gross, C. G., et al. (2009). Neural representations of
faces and body parts in macaque and human cortex: A
comparative fMRI study. Journal of Neurophysiology, 101,
2581–2600.
Pitcher, D., Charles, L., Devlin, J. T., Walsh, V., & Duchaine, B.
(2009). Triple dissociation of faces, bodies, and objects in
exastriate cortex. Current Biology, 19, 319–324.
Pitcher, D., Dilks, D. D., Saxe, R. R., Triantafyllou, C., &
Kanwisher, N. (2011). Differential selectivity for dynamic
versus static information in face-selective cortical regions.
Neuroimage, 56, 2356–2363.
McMahon, D. B. T., Russ, B. E., Elnaiem, H. D., Kurnikova, A. I.,
& Leopold, D. A. (2015). Single-unit activity during natural
Puce, A., Allison, T., Bentin, S., Gore, J. C., & McCarthy, G.
(1998). Temporal cortex activation in humans viewing eye
1192
Journal of Cognitive Neuroscience
Volume 28, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
and mouth movements. Journal of Neuroscience, 18,
2188–2199.
Quiroga, R. Q., Kreiman, G., Koch, C., & Fried, I. (2008). Sparse
but not “grandmother-cell” coding in the medial temporal
lobe. Trends in Cognitive Sciences, 12, 87–91.
Rajimehr, R., Young, J. C., & Tootell, R. B. H. (2009). An anterior
temporal face patch in human cortex, predicted by macaque
maps. Proceedings of the National Academy of Sciences,
U.S.A., 106, 1995–2000.
Rice, A., Phillips, P. J., Natu, V., An, X., & O’Toole, A. J.
(2013). Unaware person recognition from the body
when face identification fails. Psychological Science, 24,
2235–2243.
Schiltz, C., Dricot, L., Goebel, R., & Rossion, B. (2010). Holistic
perception of individual faces in the right middle fusiform
gyrus as evidenced by the composite face illusion. Journal of
Vision, 10, 25.
Schiltz, C., & Rossion, B. (2006). Faces are represented
holistically in the human occipito-temporal cortex.
Neuroimage, 32, 1385–1394.
Schmalzl, L., Zopf, R., & Williams, M. A. (2012). From head to
toe: Evidence for selective brain activation reflecting visual
perception of whole individuals. Frontiers in Human
Neuroscience, 6, 108.
Schwarzlose, R. F., Baker, C. I., & Kanwisher, N. (2005).
Separate face and body selectivity on the fusiform gyrus.
Journal of Neuroscience, 25, 11055–11059.
Song, Y., Luo, Y. L. L., Li, X., Xu, M., & Liu, J. (2013).
Representation of contextually related multiple objects in
the human ventral visual pathway. Journal of Cognitive
Neuroscience, 25, 1261–1269.
Suzuki, W. A., & Naya, Y. (2014). The perirhinal cortex. Annual
Review of Neuroscience, 37, 39–53.
Taylor, J. C., & Downing, P. E. (2011). Division of labor
between lateral and ventral extrastriate representations of
faces, bodies, and objects. Journal of Cognitive Neuroscience,
23, 4122–4137.
Taylor, J. C., Wiggett, A. J., & Downing, P. E. (2007). Functional
MRI analysis of body and body part representations in the
extrastriate and fusiform body areas. Journal of
Neurophysiology, 98, 1626–1633.
Thompson, S. A., Graham, K. S., Williams, G., Patterson, K.,
Kapur, N., & Hodges, J. R. (2004). Dissociating person-specific
from general semantic knowledge: Roles of the left and
right temporal lobes. Neuropsychologia, 42, 359–370.
Tsao, D. Y., & Livingstone, M. S. (2008). Mechanisms of face
perception. Annual Review of Neuroscience, 31, 411–437.
Tsao, D. Y., Moeller, S., & Freiwald, W. A. (2008). Comparing
face patch systems in macaques and humans. Proceedings of
the National Academy of Sciences, U.S.A., 105, 19514–19519.
Vul, E., Lashkari, D., Hsieh, P.-J., Golland, P., & Kanwisher, N.
(2012). Data-driven functional clustering reveals dominance
of face, place, and body selectivity in the ventral visual
pathway. Journal of Neurophysiology, 108, 2306–2322.
Watson, H. C., & Lee, A. C. H. (2013). The perirhinal cortex and
recognition memory interference. Journal of Neuroscience,
33, 4192–4200.
Weiner, K. S., & Grill-Spector, K. (2010). Sparsely-distributed
organization of face and limb activations in human ventral
temporal cortex. Neuroimage, 52, 1559–1573.
Wiggett, A. J., Pritchard, I. C., & Downing, P. E. (2009). Animate
and inanimate objects in human visual cortex: Evidence for
task-independent category effects. Neuropsychologia, 47,
3111–3117.
Yang, H., Susilo, T., & Duchaine, B. (2016). The anterior
temporal face area contains invariant representations of face
identity that can persist despite the loss of right FFA and
OFA. Cerebral Cortex, 26, 1096–1107.
Yovel, G., & Kanwisher, N. (2005). The neural basis of the
behavioral face-inversion effect. Current Biology, 15, 2256–2262.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
8
2
8
1
/
1
8
7
/
8
1
1
1
9
7
5
8
1
/
6
1
3
7
3
8
o
5
c
2
n
0
_
8
a
/
_
j
0
o
0
c
9
n
6
6
_
a
p
_
d
0
0
b
9
y
6
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Harry et al.
1193