Done That: Short-term Repetition Related Modulations of
Motor Cortex Activity as a Stable Signature for
Overnight Motor Memory Consolidation
Ella Gabitov1, David Manor1,2, and Avi Karni1,2
Abstrait
■ An almost universally accepted tacit expectation is that
learning and memory consolidation processes must be re-
flected in the average brain activity in brain areas relevant to
task performance. Motor cortex (M1) plasticity has been impli-
cated in motor skill acquisition and its consolidation. Never-
theless, no consistent pattern of changes in the average signal,
related to motor learning or motor memory consolidation fol-
lowing a single session of training, has emerged from imaging
études. Here we show that the pattern and magnitude of
short-term brain activity modulations in response to task repe-
tition, in M1, may provide a robust signature for effective motor
memory consolidation processes. We studied participants
during the paced performance of a finger-to-thumb opposition
séquence (FOS), intensively trained a day earlier, and a similarly
constructed untrained FOS. In addition to within-session “on-line”
gains, most participants expressed delayed, consolidation-phase
gains in the performance of the trained FOS. The execution of
the trained FOS induced repetition enhancements in the contra-
lateral M1 and bilaterally in the medial-temporal lobes, offsetting
novelty-related repetition suppression effects. De plus, the M1
modulations were positively correlated with the magnitude of
each participantʼs overnight delayed gains but not with absolute
performance levels. Our results suggest that short-term enhance-
ments of brain signals upon task repetition reflect the effective-
ness of overnight motor memory consolidation. We propose
that procedural memory consolidation processes may affect
the excitation–inhibition balance within cortical representations
of the trained movements; this new balance is better reflected in
repetition effects than in the average level of evoked neural
activité. ■
INTRODUCTION
The generation of fluent, errorless, and reliable movement
sequences (motor skills) requires practice (c'est à dire., repeated
experience; Adams, 1987). Practice-related gains in per-
formance may evolve not only during the actual training
experience on a novel sequence of movements but also
“off-line” after practice has ended (Krakauer & Shadmehr,
2006; Karni et al., 1998). Off-line, delayed gains (DGs) dans
performance presumably reflect procedural (skill) mémoire
consolidation processes that require time to evolve (Reis
et coll., 2013; Debas et al., 2010; Doyon et al., 2009; Song,
Howard, & Howard, 2007; Robertson, Pascual-Leone, &
Presse, 2004). Following practice on an explicitly introduced
motor sequence, the expression of DGs, in terms of fas-
ter and more accurate performance, was often depen-
dent on sleep (Barakat et al., 2013; Doyon et al., 2009;
Korman et al., 2007; Nishida & Walker, 2007; Korman, Raz,
Flash, & Karni, 2003; Fischer, Hallschmid, Elsner, & Born,
2002; Walker, Brakefield, Morgan, Hobson, & Stickgold,
2002; Maquet, 2001). Time and the affordance of sleep
may be necessary to complete cellular events subserving
long-term memory (Karni, 1996) such as experience-driven
1University of Haifa, 2C. Sheba Medical Center, Ramat Gan, Israel
© 2014 Massachusetts Institute of Technology
task-specific synaptic modifications (par exemple., Xu et al., 2009;
Lequel, Pan, & Gan, 2009).
There is consistent evidence from functional imaging
and TMS studies that motor learning is associated with
substantial changes within motor-related cortical and
subcortical areas (Barakat et al., 2013; Steele & Penhune,
2010; Hotermans, Peigneux, de Noordhout, Moonen, &
Maquet, 2008; Krakauer & Shadmehr, 2006; Forgeron
et coll., 2006; Doyon & Benali, 2005; Fischer, Nitschke,
Melchert, Erdmann, & Born, 2005; Floyer-Lea & Matthieu,
2005; Penhune & Doyon, 2005; Robertson, Presse, & Pascual-
Leone, 2005; Grafton, Hazeltine, & Ivry, 2002; Hikosaka,
Nakamura, Sakai, & Nakahara, 2002; Karni et al., 1995). Pour
example, the magnitude of the delayed sleep-dependent
improvement was linked to the recruitment of the primary
motor cortex (M1) during the initial training session (Steele
& Penhune, 2010). These findings support the idea that
training can initiate experience-driven changes in task
representations within M1 and that these changes may
be crucial for the triggering of consolidation processes
(Karni et al., 1995, 1998). Cependant, functional imaging
studies have provided apparently conflicting findings
regarding changes in the M1 average signal in relation to
motor memory consolidation following a single session
of training. The average magnitude of the BOLD signals
Journal des neurosciences cognitives 26:12, pp. 2716–2734
est ce que je:10.1162/jocn_a_00675
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
F
t
.
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
in M1 contralateral to the trained hand was found to be
either increased (Barakat et al., 2013), decreased (Fischer
et coll., 2005), or unchanged (Steele & Penhune, 2010).
En effet, it has been suggested that in the initial stages of
motor skill acquisition M1 may have a secondary role
and other representations of the task predominate (Doyon
& Benali, 2005; Hikosaka et al., 2002). Here we tested
whether the patterns of signal modulation in response to
task repetition, rather than the average signal intensity in
motor cortex per se, may provide a more reliable neural
signature for motor experience and specifically for effec-
tive motor memory consolidation.
The modulation of neural activity upon task repetition
is one of the most robust experience-related phenomena
(Grill-Spector, Henson, & Martine, 2006; Sayres & Grill-
Spector, 2006; Henson, 2003; Henson, Shallice, & Dolan,
2000; Grill-Spector et al., 1999; Karni et al., 1995, 1998;
Schacter & Buckner, 1998; Wiggs & Martine, 1998; Désimone,
1996; Miller & Désimone, 1994; Miller, Li, & Désimone,
1991, 1993). This modulation can occur even without
changes in the average evoked activity across trials, runs
or sessions, or the entire set of repeated stimuli. Reduced
physiological activity upon task repetition, repetition sup-
pression (RS) effects, presumably reflect the optimization
of processing and priming (Grill-Spector et al., 2006;
Henson, 2003; Schacter & Buckner, 1998; Wiggs & Martine,
1998; Désimone, 1996); cependant, these effects may be
performance independent (Sayres & Grill-Spector, 2006;
Miller & Désimone, 1994). RS effects are transient and tend
to saturate after a few iterations of the task. Repetition-
driven reduction in neural activity was shown to occur in
M1 in a variety of motor tasks (Valyear, Gallivan, McLean, &
Culham, 2012; Hamilton & Grafton, 2009; Dinstein, Hasson,
Rubin, & Heeger, 2007; Grafton & Hamilton, 2007) et était
found to be related to movement sequence learning (Karni
et coll., 1995, 1998). In the Karni et al. (1995) étude, when a
new sequence of movements was performed at a given rate
for the first time (the first performance block) and then
repeated (the second performance block) after a short rest
interval, the evoked activity in M1 contralateral to the per-
forming hand was found to be significantly decreased in
the second block (c'est à dire., was relatively suppressed). In sub-
sequent runs (sets), separated by short breaks, the mag-
nitude of neural activity recovered, as indicated by the first
performance block in the subsequent run. Nevertheless, le
evoked signal in M1 was again reduced during the second,
repeated performance block. Cependant, as practice con-
tinued, these RS effects underwent saturation, and the pat-
tern switched (about the seventh run) so that task repetition
induced a relative enhancement (RE). This latter pattern of
enhanced neural activity across performance blocks was
shown to be specific to the trained sequence and was
retained for at least a week (Karni et al., 1995, 1998). A pat-
tern of enhanced physiological signal to repeated expe-
rience (RE effects) was described in perceptual systems
and proposed to reflect increased attention, active working
mémoire, or the formation of new perceptual and even lex-
ical representation (Soldan, Particules, Hilton, & Arrière, 2008;
Henson et al., 2000; Désimone, 1996; Miller & Désimone,
1994). The within-session, experience-dependent switch
in the pattern of physiological activity to repetition across
performance blocks in a pair (c'est à dire., from RS to RE) était
proposed to reflect on-line learning of the specific move-
ment sequence (Karni et al., 1995). En particulier, it was hy-
pothesized that the within-session saturation of RS effects
within M1 may be related to familiarity with the compo-
nent movements of the sequence, which could be retained
in long-term memory, whereas RE effects may relate to
the recruitment of sequence-specific procedural memory
consolidation processes (Karni et al., 1998; see also
Hauptmann & Karni, 2002). Cependant, there is no evidence
that the persistence of RE within M1 in subsequent
sessions reflects off-line memory consolidation processes
rather than a certain level of familiarity and motor expe-
rience with the component movements of the sequence
per se.
The aim of the current study was to test the hypothesis
that the pattern of neural signal modulation by task repe-
tition, in motor cortex, constitutes a signature of motor
experience and, specifically, of off-line skill memory
consolidation.
MÉTHODES
Participants
Thirty-two healthy young adults participated in the cur-
rent study for payment: 17 participants (19–35 years,
mean = 25.7, SD = 4.4, five women) in the fMRI group
et 15 participants (n = 15, 20–35 years, mean = 25.47,
SD = 2.73, eight women) in the control group. Both
groups were trained and behaviorally tested in an iden-
tical protocol, whereas only participants of fMRI group
underwent the imaging session. The control group was
tested to evaluate the possible effects on subsequent
performance of the additional experience afforded dur-
ing the fMRI session. Two participants from the fMRI
group were not included in the analysis: One had diffi-
culties with executing the task in the scanner; another
withdrew from the fMRI session for personal reasons.
All participants reported no prior history of neurological
or psychiatric illness or brain injury and no addiction to
drugs, alcohol, or cigarettes (nonsmokers or occasional
smokers). Exclusion criteria included current or chronic
use of medication, any known learning disabilities, et
attention deficit disorder. Only individuals with little (less
que 2 années) or no formal music training participated in
the current study. Professional typists were excluded as
well. All participants affirmed that they had no sleep dis-
orders and reported at least 6 hr of proper night sleep
during the study period. Each participant was identified
as strongly right-handed using the Edinburgh Handedness
Inventory (Oldfield, 1971). Before the study, all par-
ticipants gave written informed consent according to a
Gabitov, Manor, and Karni
2717
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
/
.
t
F
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
protocol approved by the C. Sheba Medical Centerʼs
Ethics Committee.
Design and Procedures
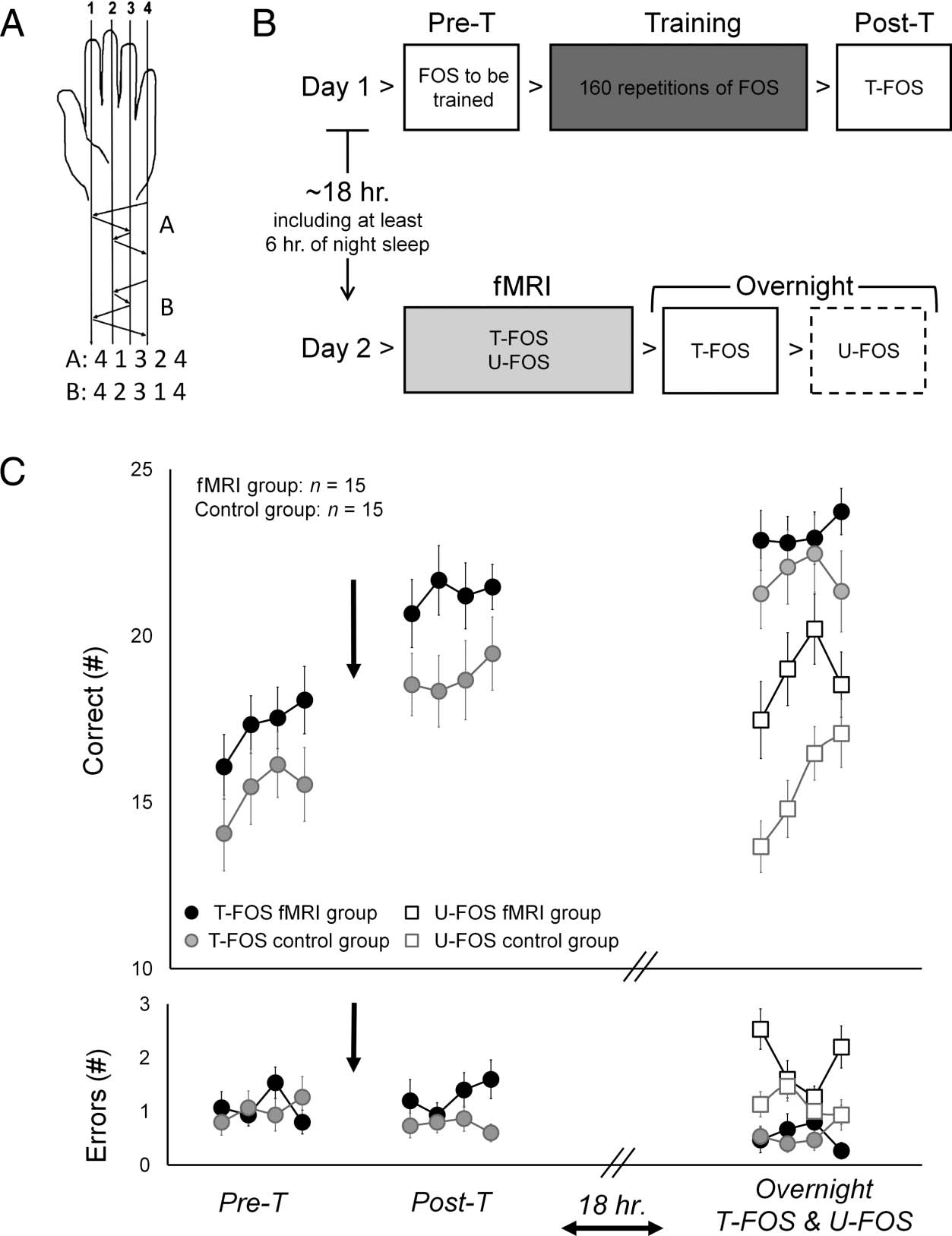
Participants were trained to accurately perform a given
five-element finger-to-thumb opposition sequence (FOS),
sequence A or sequence B, with their nondominant left
main (Figure 1A). The two FOSes consisted of identical
component movements and were mirror-reversed in rela-
tion to each other. Ainsi, the two sequences were matched
for the number of movements per digit and differed only
in their order. The movement sequence was randomly
assigned and then explicitly instructed. If the assigned
sequence for training was A (T-FOS), then the sequence
B was used as a novel untrained sequence (U-FOS) et
vice versa.
Each participant took part in two study phases that
took place on two consecutive days during the morning
or early afternoon hours and were separated by 18 hr
of interval. The interval between the two phases included
at least 6 hr of night sleep (self-report; Figure 1B). Sur
Day 1, each participant underwent a pretraining perfor-
mance test (Pre-T), a structured training session, et
an immediate posttraining performance test (Post-T).
On the next day, all participants were retested on the
performance of the trained sequence that was followed
by a test of the untrained sequence (Overnight: T-FOS
and U-FOS, respectivement) using the trained (gauche) aussi
as the untrained (droite) main. The results for the un-
trained hand will be reported elsewhere. Participants of
the fMRI group took part in the scanning session, lequel
immediately preceded the overnight performance tests.
Inside the scanner, participants repeatedly performed
either the trained or the untrained sequence at an iden-
tical auditory-paced rate. Ainsi, both the rate of the op-
position movements and the component movements
composing the sequences were matched. The logic of
Chiffre 1. Study design and
behavioral results. (UN) FOS. Le
two sequences were matched
for number of movements per
digit and mirror-reversed in
relation to each other (in terms
of order). (B) The overall study
conception. Day 1: a pre-training
performance test (Pre-T), un
structured training session
(Training), and an immediate
posttraining performance test
(Post-T). Day 2: performance
tests of the trained sequence
and the untrained sequence
(Overnight: T-FOS and U-FOS,
respectivement). Only participants
of the fMRI group took part in
the scanning session (IRMf),
immediately preceding
overnight performance tests.
The control (non-fMRI) group
was tested to evaluate the effect
of the additional experience
afforded during the fMRI
session on subsequent
performance. (C) Performance
of fMRI and control group.
Two measures of performance
are plotted: mean number of
correct sequences (top) et
mean number of errors
(bottom). Each data point
represents performance in
a 30-sec test-block; arrow,
entraînement (160 repetitions of
the FOS); bars, SEM. Note
that experience with the two
sequences inside the scanner
(fMRI group) did not result
in better performance of
the T-FOS (compared with
controls) in the subsequent test.
2718
Journal des neurosciences cognitives
Volume 26, Nombre 12
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
t
.
F
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
this design, using paced component movements, is that
differences in fMRI signals would not directly reflect dif-
ferences in performance speed that were expected to re-
sult from training on one but not the other sequence
(Korman et al., 2003; Karni et al., 1995). It has been
shown that the rate of finger movement execution can
significantly (and nonlinearly) affect the evoked BOLD
signals in M1 (Rao et al., 1996). In all sessions and tests,
the participants performed the instructed movement se-
quence lying supine. The executing hand was positioned
beside the trunk in direct view (palm-up) of a video cam-
era to allow the recording of all digit movements. Visual
feedback was not afforded at any time.
Training and Tests
The participants were trained and tested according to a
standard FOS training protocol (Korman et al., 2003,
2007). The training session consisted of 160 repetitions
of a given sequence divided into 10 training blocks. Dur-
ing the training, the beginning as well as the end of each
training block (10 blocks, 16 repetitions of a given se-
quence per block) were marked by a “READY” and a
“STOP” auditory signal, respectivement. To ensure that the
training experience afforded was identical to all partici-
pants, the initiation of each sequence during the training
was cued by another auditory signal at a rate of 0.4 Hz
(2.5 sec per sequence), which prior studies have shown
to be a comfortable rate for young adults (Korman et al.,
2003, 2007). Full explicit instruction of the sequence
and general encouragement to continue accurate per-
formance were given before each training block. Chaque
performance test (Pre-T, Post-T, Overnight T-FOS and
U-FOS) included four blocks of 30-sec duration. Chaque
test-block was followed by a rest interval of 30 sec. Before
each test-block, participants were asked to perform the
movement sequence, and the block was initiated only af-
ter the FOS was accurately reproduced three times in a
row. The great majority of participants successfully per-
formed a given sequence three times in a row in their
first attempt. The need for a second additional attempt
was very rare and was required only when an instructed
sequence was novel, c'est, during the Pre-T and U-FOS
performance tests. Each test-block was initiated by an
auditory “READY” signal, after which participants per-
formed the sequence continuously “as fast and as accu-
rately as possible” until a “STOP” signal was given.
Participants were instructed not to correct occasional er-
rors. In case an occasional error occurred, an instruction
“not to correct errors and to continue the task from the
initial movement component of a sequence as smoothly
as possible” was given. During the test, no feedback on
performance was provided. The participantsʼ perfor-
mance during the test-blocks was recorded by video
camera and scored off-line. For each test-block, deux
quantitative measures of performance were determined
from these recordings: (1) the number of correctly com-
pleted sequences as a measure of speed and (2) the num-
ber of sequences with ordering errors as a measure of
accuracy. The beginning of a sequence was identified
by the first component movement; all incorrect opposi-
tion movements within a single trial of a sequence were
counted as one error (c'est à dire., the measure reflected the
number of incorrect sequences).
fMRI Session
Participants were asked to perform one of the two possi-
ble sequences of finger movements, the sequence
trained the day before (T-FOS) and the novel sequence
(U-FOS; Figure 1A), using their trained (gauche) main. Le
untrained (droite) hand was subsequently tested as well;
the results to be reported elsewhere. The component
movements of the sequences were paced by an auditory
signal at a fixed rate of 1.66 Hz to control rate-related
changes in the BOLD signal (Rao et al., 1996). The paced
performance enabled the assessment of signal differ-
ences as a function of the order of the component move-
ments (sequence representation) minimizing potential
differences between the sequences as well as between
individuals driven by spontaneously generated perfor-
mance rates.
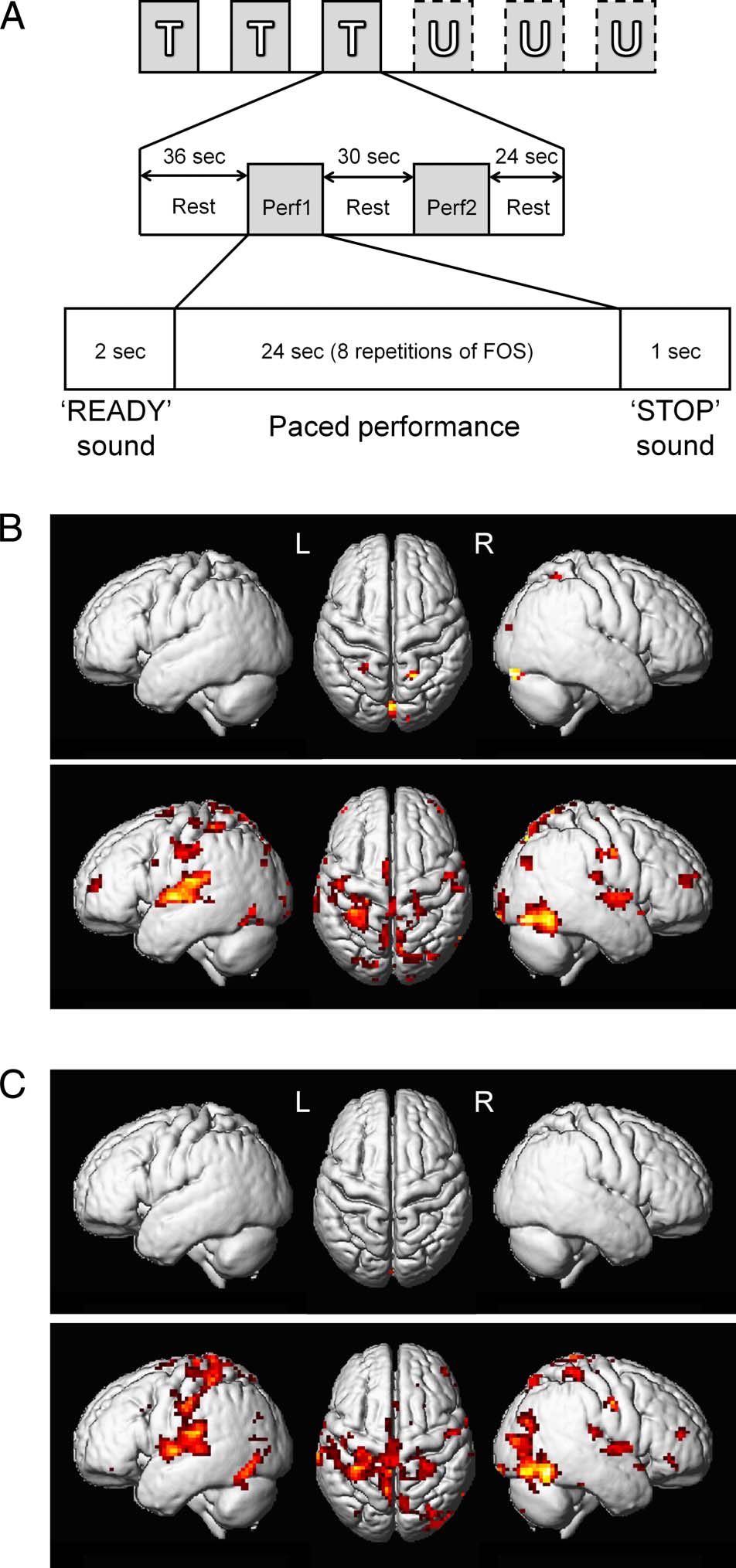
The imaging session consisted of three consecutive
runs for each sequence (T-FOS, U-FOS; Figure 3A). Dans
this way, potential effects of proactive interference and
contextual retrieval that could be caused by switching be-
tween the two sequences were minimized (Kiesel et al.,
2010; Cothros, Köhler, Dickie, Mirsattari, & Gribble,
2006). The order of sequences was counterbalanced
across participants. Experimental runs (chaque 144 sec
long) were separated by a 1.5- to 2-min break, which in-
cluded a verbal interaction with the participant. Partici-
pants were informed about the target sequence of
finger movements, either the T-FOS or the U-FOS, à
be performed in the next run, and the run was initiated
only after the target FOS was accurately reproduced
three times in a row by the participant. Each run con-
sisted of two performance blocks (Perf1 and Perf2) sep-
arated by a rest interval of 30 sec. Each block was initiated
by an auditory and visual “READY” cue (2 sec), after
which participants performed the required FOS continu-
ously in a paced manner for a total of eight repetitions of
the FOS (24 sec). The end of the performance was
marked by an auditory and visual “STOP” cue (1 sec).
Each run began and ended with a rest period of 36 et
24 sec, respectivement.
The participantsʼ performance during the fMRI session
was recorded by a video camera focused on the perform-
ing hand and evaluated by at least one trained observer,
on-line and off-line. Performance was scored for accuracy,
timing (c'est à dire., initiation and termination of FOS perfor-
mance), and performance rate. This experiment was realized
using Cogent 2000 developed by the Cogent 2000 team
at the FIL and the ICN and Cogent Graphics developed
Gabitov, Manor, and Karni
2719
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
/
t
.
F
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
by John Romaya at the LON, Wellcome Department of
Imaging Neuroscience, and implemented in Matlab (Le
Mathworks, Inc., Natick, MA).
Behavioral Data Analyses
For each participant, two performance measures were
calculated for each test-block: the number of correctly
completed sequences as a measure of speed and the
number of sequences with ordering errors as a measure
of accuracy. Note that throughout the experiment, partic-
ipants made very few errors. The detailed report of per-
formance gains and their sequence specificity refers to
analyses of the fMRI group data. The control group was
tested to evaluate the possible effects on subsequent per-
formance of the additional experience afforded during
the fMRI session. Unless otherwise stated, the analyses
were designed as within-subject comparisons. Separate
repeated-measures ANOVAs for each performance mea-
sure with Test and Test-block as within-subject factors
were run using Statistical Package for the Social Sciences
(SPSS Statistics for Windows, Version 19.0; IBM Corp.,
Armonk, New York). The results were corrected for nonspheric-
ity violation using the Greenhouse–Geisser adjustment.
To evaluate individual improvements in performance
speed, an average number of correct sequences (speed)
across the four test-blocks of each performance test was
calculated for each participant and converted to percents
relative to his or her average speed at Pre-T. Ainsi, speed
in percents achieved by each participant at Post-T is a
measure for individual within-session gains in speed. Dans-
dividual DGs in speed were calculated by subtracting
speed in percents achieved at Post-T from that achieved
overnight for the T-FOS (c'est à dire., Overnight T-FOS (%) −
Post-T (%)). To evaluate individual improvements in ac-
curacy, individual accuracy rates were determined for
each performance test as percent of correct sequences
(c'est à dire., [average correct (#)] / [average correct (#) + aver-
age errors (#)] × 100%). Within-session and DGs in accu-
racy were calculated accordingly as differences between
accuracy rates (c'est à dire., Post-T (%) − Pre-T (%) and Over-
night T-FOS (%) − Post-T (%) for within-session and
DGs, respectivement).
planar T2*-sequence (repetition time = 3000 msec, echo
time = 35 msec, flip angle = 90°, field of view = 220 ×
220 mm2, matrix size = 64 × 64 voxels, voxel size = 3.4 ×
3.4 × 3.4 mm3, no gap, ascending) avec 40 axial oblique
slices, covering the whole brain.
MRI Analyses
Preprocessing
The structural and functional images were converted to
Neuroimaging Informatics Technology Initiative format
using MRIcron (University of South Carolina). Prepro-
cessing and statistical analysis of the data were carried
out with Statistical Parametric Mapping (SPM8; Wellcome
Department of Cognitive Neurology, Londres, ROYAUME-UNI) operating
under Matlab R2012a. For each run, the four initial scans
were discarded to allow for magnetic saturation and equili-
bration effects. D'abord, all images were reorientated to stereo-
tactic space. All functional volumes were realigned using a
least squares approach and a six-parameter (rigid body)
spatial transformation to remove movement-related vari-
ance. To correct for nonrigid distortion, realigned func-
tional volumes were unwarped, adjusting for interactions
between movement and local field inhomogeneity (Hutton,
Andersson, Deichmann, & Weiskopf, 2013; Andersson,
Hutton, Ashburner, Tourneur, & Friston, 2001). This dynamic
geometric distortion correction reduces motion-related
variance and improves temporal signal-to-noise ratio (Hutton
et coll., 2013; Andersson et al., 2001). Following segmen-
tation and skull-stripping of the structural data, functional
images were coregistered to the individual skull-stripped
3-D anatomical image and normalized to the Montreal
Neurological Institute (MNI) space using parameters ob-
tained from the segmentation procedure. The normalized
functional images were resampled to voxel dimensions of
3 mm3. Enfin, functional images were spatially smoothed
with a Gaussian kernel of 8 mm FWHM to improve the
signal-to-noise ratio. Before statistical analyses, head
motion artifact detection routine was applied on the pre-
processed data using the Artifact Detection Tools (Mazaika,
Hoeft, Glover, & Reiss, 2009). No significant head motion
artifacts were detected (z threshold = 2, mouvement
threshold = 2 mm, rotation threshold = 0.05 rad).
MRI Data Acquisition
Whole-brain and ROI Analyses
fMRI scanning was carried out at the C. Sheba Medical
Centre, Tel-Hashomer, using a 3T whole-body high-
definition system (GE EXCITE 3 HD) equipped with an
eight-channel head coil. High-resolution full-brain 3-D
structural images were acquired in the axial orientation
using a T1*-weighted echo-planar sequence (repetition
time = 7.3 msec, echo time = 3 msec, flip angle =
20°, field of view = 256 × 256 mm2, matrix size = 256 ×
256 voxels, voxel size = 1 × 1 × 1 mm3). BOLD-sensitive
functional images were obtained using a gradient echo-
Statistical analyses of BOLD signal changes were per-
formed using a general linear model (Friston et al., 1995).
Individual models were specified separately for each
séquence (T-FOS, U-FOS) using a multisession design,
and each session included data from a single run (three
runs). Regressors of interest (c'est à dire., Perf1 and Perf2) étaient
modeled as a boxcar function with a length of 24 sec
convolved with the canonical hemodynamic response
fonction. A high-pass filter of 128 sec was used to remove
low-frequency noise. For the block design, inclusion of
2720
Journal des neurosciences cognitives
Volume 26, Nombre 12
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
.
F
t
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
motion covariates has a deleterious impact on general
linear model sensitivity when even moderate correlation
existed between motion and the experimental design
( Johnstone et al., 2006). Donc, movement parameters
derived from realignment of the functional volumes were
not included as covariates. Following the model parameter
estimation, the linear contrasts for each sequence (T-FOS,
U-FOS) were defined as follows: Perf versus Rest (c'est à dire., main
effect of performance blocks) to assess task-related
changes in BOLD-fMRI signal; and Perf2 versus Perf1 to
assess changes in BOLD-fMRI signal upon task repetition.
To evaluate group effects of task-related activity and repe-
tition, contrast images were introduced into second-level
analyses treating participants as a random effect (un-
sample t test). To assess the main effect of learning,
comparison between the two sequences (T-FOS vs. U-FOS)
was performed on the task-related activity (Perf vs. Rest)
using a one-way within-subject ANOVA design.
Activation maps were thresholded at p ≤ .001, uncor-
rected, and overlaid on the mean structural image of all par-
ticipants using SPM8 and Functional Imaging Visualization
Environment (nmr.mgh.harvard.edu/harvardagingbrain/
People/AaronSchultz/OrthoView.html). Statistical infer-
ences were performed on the cluster level using p values
family-wise error rate (FWE)-corrected for multiple com-
parisons over the entire brain or on the peak level using
p values FWE-corrected over a small VOI. VOIs for small
volume corrections were defined for structures within
the motor-related (Halsband & Lange, 2006) and resting
state (Buckner, Andrews-Hanna, & Schacter, 2008) net-
works using Human Motor Area Template (HMAT; Mayka,
Corcos, Leurgans, & Vaillancourt, 2006) and anatomical
areas of Automated Anatomical Labeling (AAL; Tzourio-
Mazoyer et al., 2002). VOIs relevant to the reported statis-
tics are listed in Table 1.
dation, using the MarsBar toolbox for SPM (Brett, Anton,
Valabregue, & Poline, 2002). The ROIs were defined
as spheres with a radius of 6 mm centered within sig-
nificant clusters resulted from second-level analyses.
The extracted betas and contrast values were introduced
to SPSS for further analyses.
RÉSULTATS
Behavioral Results
The detailed report of performance gains and their se-
quence specificity refers to analyses of the fMRI group
data. The control group was tested to evaluate the possi-
ble effects on subsequent performance of the additional
experience afforded during the fMRI session.
Performance Gains for the Trained Sequence
Training on the assigned sequence of movements resulted
in both early (within-session) and delayed (“off-line,” time-
dependent) gains in performance (Figure 1C). Dans l'ensemble,
repeated-measures ANOVAs with Test (Pre-T, Post-T and
Overnight T-FOS) and Test-block (1–4) as within-subject
factors showed significant differences in performance
speed as well as in the number of errors across the three
tests (F(1.44, 20.14) = 87.48, p < .001; F(1.7, 23.85) =
4.82, p < .05, the number of correct and the number of
errors, respectively), indicating improvements of both
speed and accuracy. There was also a significant effect of
Test-block (F(2.00, 27.99) = 4.08, p < .05; F(2.51, 35.14) =
3.10, p < .05, the number of correct and the number of
errors, respectively), indicating that overall performance
tended to improve within the tests.
ROI analyses were performed to explore individual
differences in neural activity and its relevance to consoli-
To test for within-session gains, a post hoc comparison
between Pre-T and Post-T was perfomed. There were
Table 1. VOIs Used for Small Volume Corrections
N
1
2
3
4
5
6
7
8
9
VOI
Image Calculation
Right primary sensorimotor cortex
Left primary sensorimotor cortex
S1(R) + M1(R)
S1(L) + M1(L)
Supplementary motor area
SMA proper(L + R) + pre-SMA(L + R)
Right dorsal lateral premotor cortex
Left dorsal lateral premotor cortex
Right ventral lateral premotor cortex
Left ventral lateral premotor cortex
Right medial-temporal lobe
Left medial-temporal lobe
PMd(R)
PMd(L)
PMv(R)
PMv(L)
Hippocampus(R) + ParaHippocampal(R)
Hippocampus(L) + ParaHippocampal(L)
+ = union; R = right hemisphere; L = left hemisphere; S1 = primary sensory cortex (HMAT); M1 = primary motor cortex (HMAT); SMA proper =
supplementary motor area proper (HMAT); pre-SMA = pre-supplementary motor area (HMAT); PMd = dorsal lateral premotor cortex (HMAT); PMv =
ventral lateral premotor cortex (HMAT); Hippocampus (AAL), ParaHippocampal = parahippocampal gyrus (AAL).
Gabitov, Manor, and Karni
2721
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
a
/
_
j
0
o
0
c
6
n
7
5
_
a
p
_
d
0
0
b
6
y
7
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
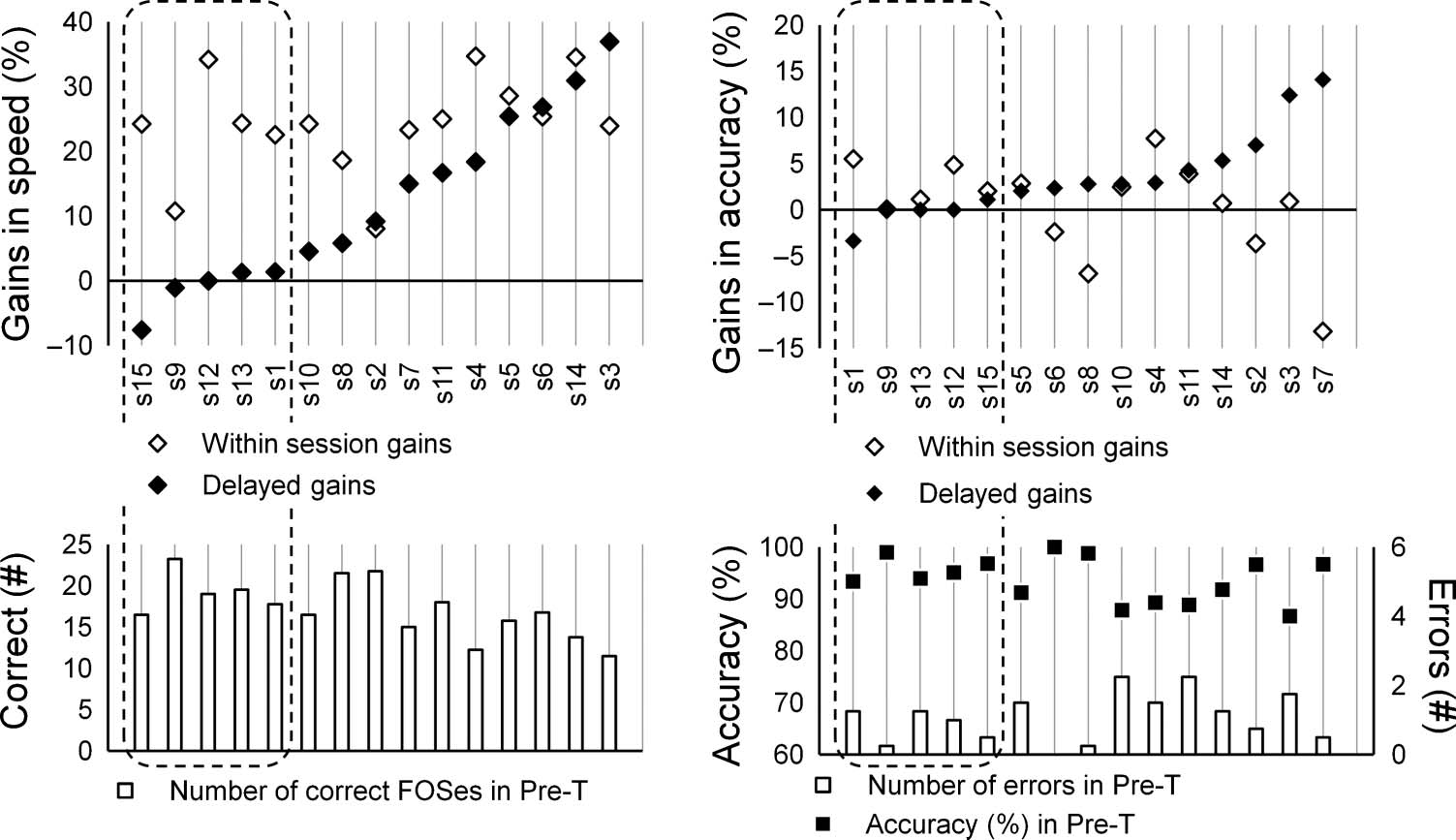
Figure 2. Individual
performance of fMRI group.
Each vertical line represents
data of a single participant
(s1–15). Dashed area,
participants who failed to
express DGs (<2% gains in
speed). Individual speeds for
each performance test were
converted into percents
(Pre-T = 100%). Gains in speed
(%): within-session gains =
Post-T (%), DGs = Overnight
T-FOS (%) − Post-T (%) (top
left plot). Mean number of
correct sequences (speed)
at Pre-T (bottom left plot).
Individual accuracy rates for
each performance test were
determined as percent of
correct sequences (i.e.,
[average correct (#)] /
[average correct (#) +
average errors (#)] × 100%).
Gains in accuracy (%): within-
session gains = Post-T (%) −
Pre-T (%), DGs = Overnight
T-FOS (%) − Post-T (%)
(top right plot). Mean number
of errors and accuracy (%) at
Pre-T (bottom right plot).
significant within-session gains in speed with no loss in
accuracy (F(1, 14) = 196.2, p < .001; F(1, 14) = 0.49,
p = .5, the number of correct and the number of errors,
respectively). For speed, the effect of test-block was signifi-
cant before but not immediately after the training (F(2.17,
30.41) = 4.53, p < .02; F(2.26, 31.60) = 1.09, p = .35 Pre-T
and Post-T, respectively), indicating that performance sta-
bilized as a result of practice; however, there was no signif-
icant test by test-block interaction. Importantly, additional
gains in performance speed developed overnight (Over-
night T-FOS compared to Post-T: F(1, 14) = 14.42, p <
.01). These DGs in speed were paralleled by a significant
reduction in the absolute number of errors (F(1, 14) = 9.18,
p < .01). The concurrent gains in both speed and accuracy
indicate skill acquisition rather than speed–accuracy trade-
off, that is, participants were not trading one aspect of
performance for the other. On average the overnight gains
(DGs) in speed were 1.83 ± 0.48 additional sequences per
test-block, the number of errors, that were very low
throughout, dropped from 1.8 ± 0.26 (mean ± SEM,
Post-T) to 0.55 ± 0.15 (Overnight T-FOS) per test-block.
However, although all of the participants gained in speed
within the session, five participants failed to show addi-
tional overnight improvements (DGs) in speed and accuracy
(Figure 2); in the control group, one participant did not
improve in speed overnight. The performance levels of
these participants were not exceptional at any of the per-
formance tests. Failure to express DGs overnight was not
related to the initial performance levels, the saturation of
within-test improvements in speed, or the absolute speed
achieved overnight for the T-FOS.
Specificity of Performance Gains
The gains in performance expressed after the training
were sequence specific (Figure 1C). Participants were un-
able to express the gains attained for the T-FOS in the
performance of the matching untrained sequence (U-FOS).
The performances of the T-FOS at both post-training and
overnight test were significantly faster than the performance
of the U-FOS (F(1, 14) = 12.33, p < .01; F(1, 14) = 39.47, p <
.001; Overnight U-FOS compared with Post-T and Overnight
T-FOS, respectively) as well as significantly more accurate
(F(1, 14) = 5.51, p < .05; F(1, 14) = 26.67, p < .001; Over-
night U-FOS compared with Post-T and Overnight T-FOS,
respectively). However, a day after the training, the fMRI
group performed the U-FOS significantly faster (F(1, 14) =
6.24, p < .05) but less accurate than the T-FOS before the
training (F(1, 14) = 11.04, p < .01).
Effects of Additional Experience in the Scanner
Comparison with the control group (trained and tested
with an identical protocol but without the added expe-
rience of the neuroimaging session afforded, on both
sequences, preceding the overnight performance tests)
showed that the fMRI group had no significant advantage
2722
Journal of Cognitive Neuroscience
Volume 26, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
a
/
_
j
0
o
0
c
6
n
7
5
_
a
p
_
d
0
0
b
6
y
7
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
in the performance of the T-FOS at any test (Figure 1C). A
repeated-measures ANOVA with Group (fMRI and control)
as a between-subject factor showed no significant Group
effect (F(1, 28) = 2.216, p = .148; F(1, 28) = 1.045, p =
.32, the number of correct and the number of errors,
respectively) and no significant Group × Test interaction
(F(1.56, 43.65) = 1.578, p = .22; F(1.79, 50.05) = 1.32,
p = .28, the number of correct and the number of errors,
respectively). There was, however, a significant main effect
of Test (F(1.56, 43.65) = 168.29, p < .001; F(1.79, 50.05) =
4.19, p < .05, the number of correct and the number of
errors, respectively) indicating robust gains in perfor-
mance, speed, as well as accuracy, across the three tests
for both groups. Significant differences in performance be-
tween the two groups were found only for the U-FOS with
a significantly faster but less accurate performance in the
fMRI group (F(1, 28) = 6.82, p = .01; F(1, 28) = 6.97, p =
.01, the number of correct and the number of errors, re-
spectively). Thus, experience with the two sequences inside
the scanner did not result in better performance of the
T-FOS in the subsequent test but had an effect on the per-
formance of the U-FOS.
fMRI Results
During the fMRI session, participants were instructed to
perform either the trained or the untrained sequence fol-
lowing paced auditory cue. Each run consisted of two
performance phases of the same sequence (T-FOS or
U-FOS) designed as blocks (Perf1 and Perf2) and separated
by a brief rest interval (Figure 3A).
Neural Correlates of Previous Experience
The second-level analyses were run twice, once on the
data from all participants (n = 15) and again on the data
from only those participants who expressed DGs in
speed overnight (DGs group, n = 10). Analyses referring
to both groups are presented in Figures 3, 4, 5, and 6 and
Tables 2 and 3. There were no significant differences in
task-related neural activity evoked by the two sequences
(T-FOS vs. U-FOS) in both analysis groups. However,
activity within extensive cortical areas was modulated dif-
ferentially across performance blocks for the trained and
the untrained sequences. Analyses of the data from all
participants showed that, during the T-FOS performance,
relative decreases in BOLD-fMRI signals from the first to
the second, repeated performance blocks (i.e., RS effects:
Perf1 > Perf2) were significant only within the calcarine
(Figure 3B, top; Tableau 2.1). Additional RS effects for the
T-FOS were evident within the right cerebellum (Crus1)
and small clusters located in the superior parietal gyri but
were not significant on the cluster level. The U-FOS, comment-
jamais, induced significant RS effects in multiple brain areas
(Figure 3B, bottom; Tableau 2.2). These RS effects were sig-
nificant within the SMA, the bilateral pre- and postcentral
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
t
.
/
F
Chiffre 3. fMRI session design and RS effects. (UN) The fMRI session
conception. T = T-FOS; U = U-FOS; Perf1, Perf2 = two blocks of FOS
performance. Note that both sequences were performed at an identical
auditory-paced rate of 1.66 Hz per movement. (B, C) Activation
maps showing RS effects for all participants (n = 15, B) et pour
participants expressing DGs (n = 10, C) for both sequences (top,
T-FOS; bottom, U-FOS). Activation maps are shown over the surface
rendered from the mean structural image of all participants,
thresholded at p < .001 (uncorrected).
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
gyri, as well as the inferior and superior parietal cortex.
Additional clusters with significant RS effects for the
U-FOS were located in the middle and posterior cingu-
late cortex as well as within the middle superior and pos-
terior inferior temporal gyri. Identical analyses of the
Gabitov, Manor, and Karni
2723
Table 2. RS Effects (All Participants)
Label
1. RS for T-FOS (Perf1 > Perf2)
Calcarine
Calcarine
R.
L
2. RS for U-FOS (Perf1 > Perf2)
Precentral
Postcentral*2
Parietal_Inf
Parietal_Sup
Precentral*5
Postcentral
Parietal_Inf*2
Precentral*6
Precentral
Postcentral
Postcentral*1
Cingulum_Mid
Cingulum_Mid
Supp_Motor_Area*3
Parietal
Precuneus
Precuneus
Temporal_Sup
Temporal_Mid
SupraMarginal
Insula
Temporal_Inf
Temporal_Mid
Occipital_Inf
Cerebellum_6
Rolandic_Oper
Temporal_Sup
Calcarine
Calcarine
Lingual
L
L
L
L
L
L
L
R.
R.
R.
R.
R.
R. + L
L
R.
R.
L
L
L
L
L
R.
R.
R.
R.
R.
R.
L
R.
R.
X
12
−6
−21
−30
−27
−33
−39
−45
−45
48
21
21
21
9
0
−3
21
6
–
−54
−51
−60
−36
51
60
42
21
63
57
−3
15
15
Coordonnées INM
oui
z
Size
( Voxels)
z Score
p
−88
−82
−28
−37
−46
−46
−7
−13
−25
−7
−19
−43
−28
−31
−37
2
−79
−76
−55
−10
−37
−28
−19
−58
−64
−76
−79
5
−4
−100
−94
−97
7
10
58
52
52
58
49
40
37
40
64
49
55
46
46
64
52
58
70
1
−2
22
4
−17
−2
−17
−17
4
1
−14
1
−11
149
204
108
70
271
59
239
425
367
95
123
.02FWE
.001FWE
.03FWE
.01
< .001FWE
.02
< .001FWE
< .001FWE
< .001FWE
.04FWE
.02FWE
4.48
3.92
3.51
4.32
3.82
3.50
3.90
3.47
3.77
3.97
3.29
4.20
4.02
4.17
3.87
3.81
4.34
3.70
3.56
4.51
3.39
3.55
3.23
4.31
3.39
4.01
3.39
4.00
3.99
3.93
3.87
3.38
Labeling clusters (the most significant local maxima for each area) obtained from activation maps thresholded at p < .001 (uncorrected) using AAL
(Tzourio-Mazoyer et al., 2002). *[N]Significant peak at p < .05 level FWE-corrected over a small VOI; [N] refers to a VOI used for small volume
correction as listed in Table 1; pFWE = cluster-level FWE-corrected over the entire brain; p = cluster-level uncorrected.
2724
Journal of Cognitive Neuroscience
Volume 26, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
a
/
_
j
0
o
0
c
6
n
7
5
_
a
p
_
d
0
0
b
6
y
7
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
DGs group showed no significant RS effects for the T-FOS
and similar RS effects during the U-FOS performance
(Figure 3C).
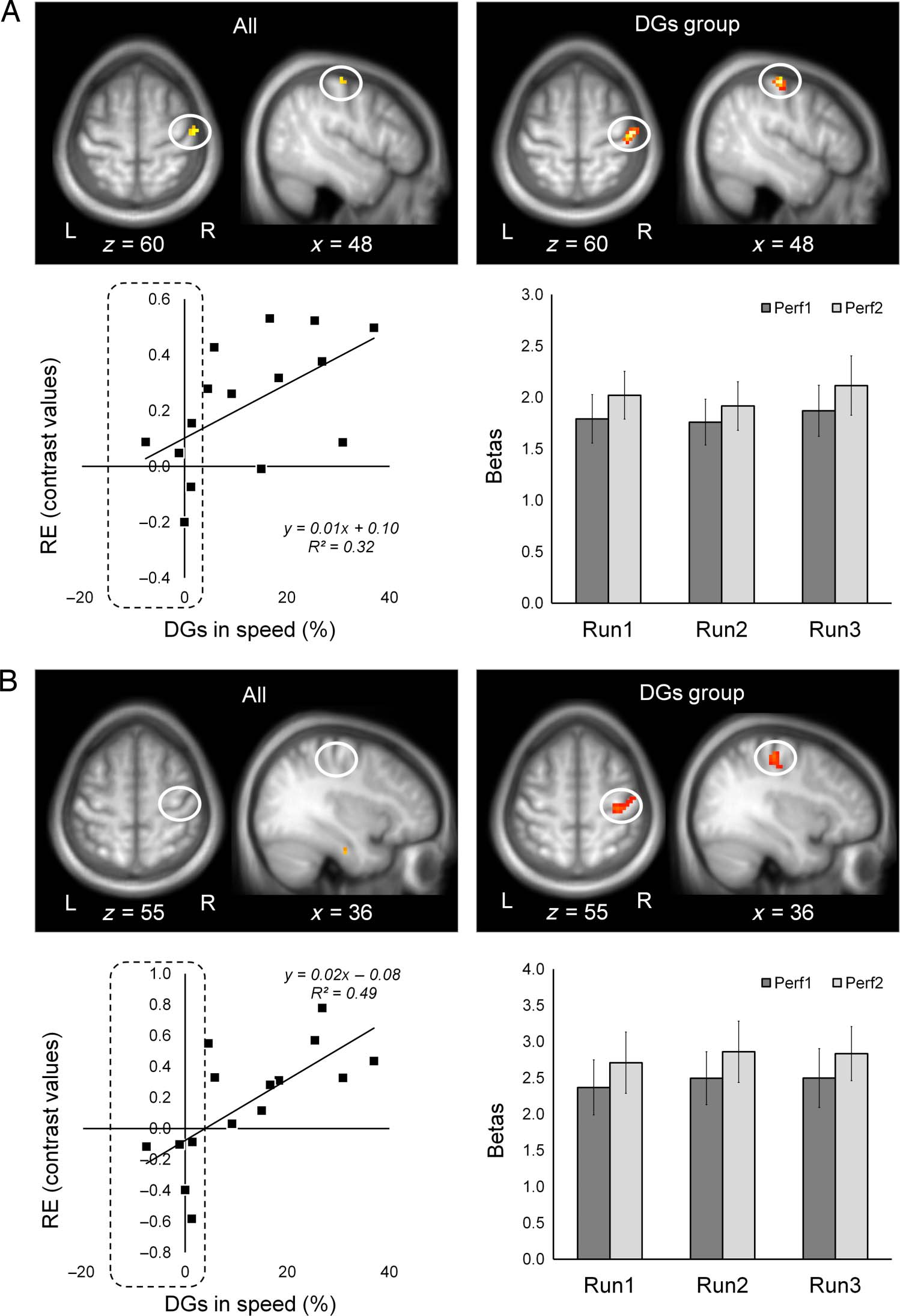
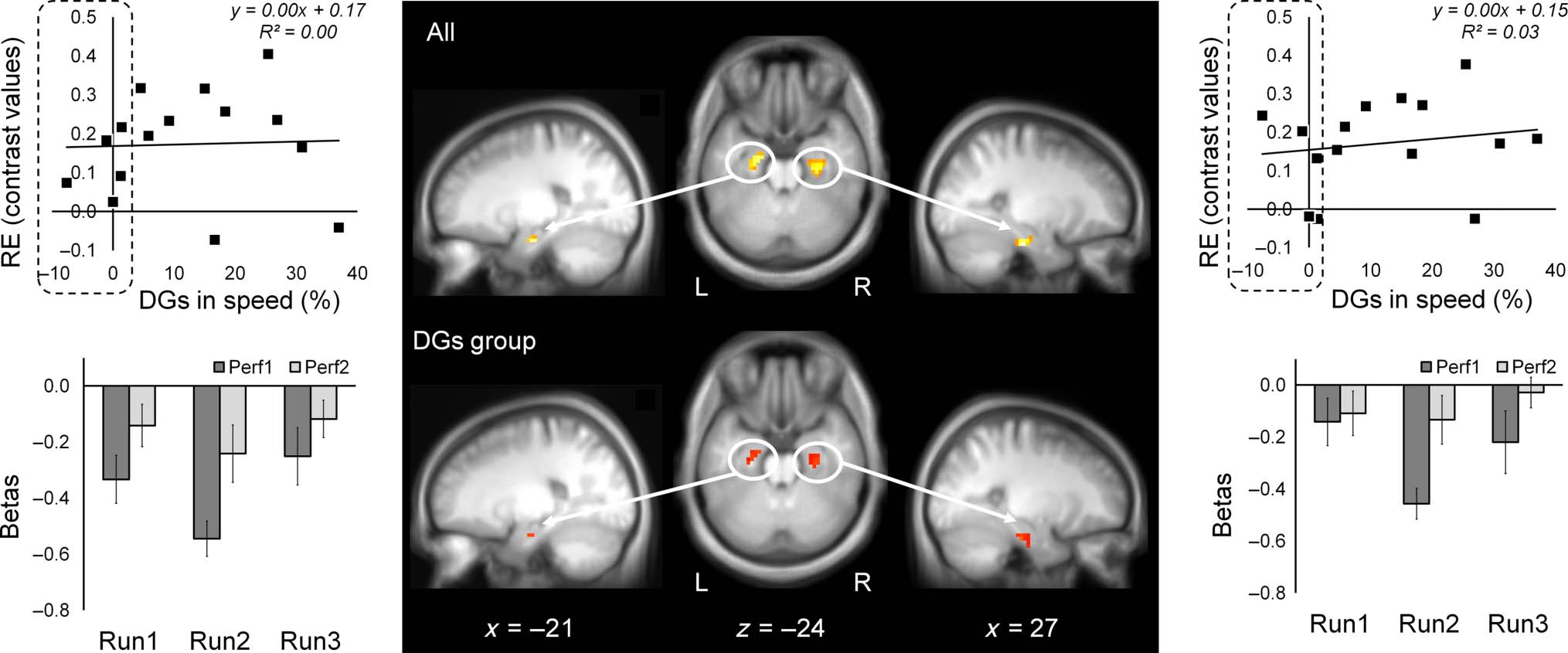
Importantly, relative increases in BOLD-fMRI signals
across performance blocks (i.e., RE effects: Perf1 <
Perf2) were found only for the T-FOS (Figures 4 and 5,
Table 3). The analysis of data from all participants
showed significant RE effects within a dorsal part of the
right (contralateral) sensory-motor cortex somewhat lat-
eral to the hand knob (lateral M1; Figure 4A) and bilat-
erally in the medial-temporal lobe (MTL)—an area
including the parahippocampal cortex and the hippo-
campus (Figure 5). In a similar analysis restricted to the
DGs group REs upon repeated performance of the T-FOS
occurred in the right sensory-motor cortex and included
the knob of the right precentral gyrus, that is, the M1
hand area (Yousry et al., 1997; Figure 4B). Note that
the RE effects in bilateral MTL reflected smaller negative
BOLD signals versus rest upon task repetition (Figure 5,
bottom plots). However, within the right sensory-motor
Figure 4. RE effects within
the right (contralateral) M1.
Activation maps showing RE
effects for all participants (n =
15, top left) and for participants
expressing DGs (n = 10, top
right). Activation maps are
shown over the mean structural
image of all participants,
thresholded at p < .001
(uncorrected). Individual
contrast values for RE effects
(Perf1 < Perf2, y axis) versus
DGs (%, x axis) (left plots);
dashed area, participants who
failed to express DGs (<2%
gains in speed). Mean signals
(betas) for each performance
block of the T-FOS (right plots);
bars, SEM. Contrast values and
mean signals (betas) were
extracted from spherical ROIs
with a radius of 6 mm. (A)
lateral M1, sphere ROI centered
at [48 −16 60], mean signals
(betas) refer to the data from all
participants (right plot). (B) M1
hand area (knob), sphere ROI
centered at [36, −25, 55], mean
signals (betas) refer to the data
from DGs group (right plot).
Note that RE effects within the
knob of the central gyrus were
induced only in participants
expressing DGs.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
a
/
_
j
0
o
0
c
6
n
7
5
_
a
p
_
d
0
0
b
6
y
7
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Gabitov, Manor, and Karni
2725
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
a
/
_
j
0
o
0
c
6
n
7
5
_
a
p
_
d
0
0
b
6
y
7
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 5. RE effects within MTL. Activation maps showing RE effects for all participants (n = 15, top middle) and for participants expressing DGs
(n = 10, bottom middle). Activation maps are shown over the mean structural image of all participants, thresholded at p < .001 (uncorrected).
Individual contrast values for RE effects (Perf1 < Perf2, y axis) versus DGs (%, x axis) (top plots); dashed area, participants who failed to express DGs
(<2% gains in speed). Mean signals (betas) for each performance block of the T-FOS for all participants (bottom plots); bars, SEM. Contrast values
and mean signals (betas) were extracted from spherical ROIs with a radius of 6 mm. Left MTL, sphere ROI centered at [−21, −13, −23] (left);
right MTL, sphere ROI centered at [27, −22, −26] (right).
cortex, the RE effects were the result of enhanced positive
BOLD signals versus rest (Figure 4, right plots).
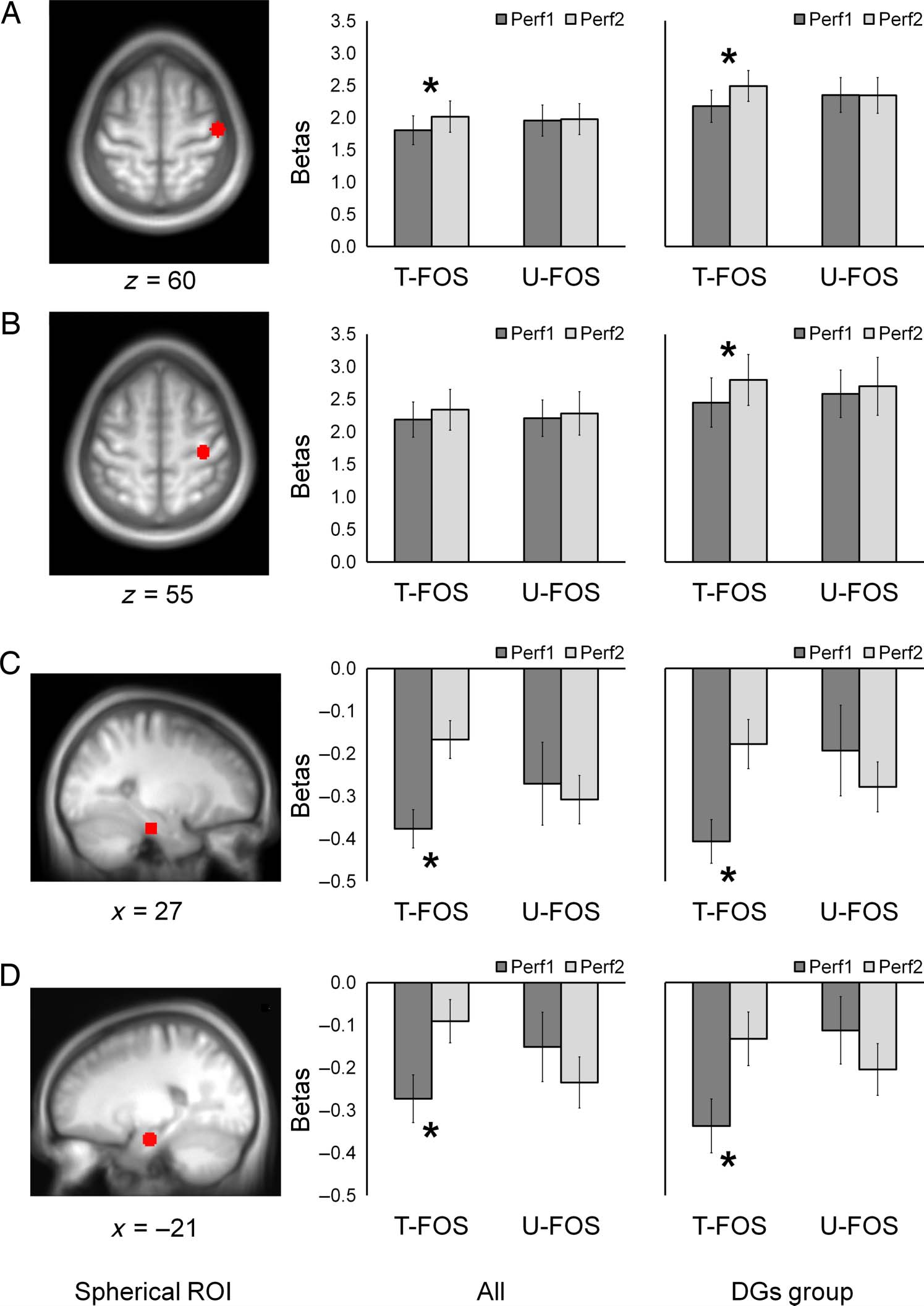
ROI and Correlation Analyses—The Expression of DGs
ROI analyses were performed on spheres centered within
the clusters showed significant RE effects (Figures 4 and
5). The choice of spheres was driven by the findings of
two separate local maxima within M1 in the group analy-
ses; note that the focus within the M1 hand knob was sig-
nificant only in the DGs group. To this end, mean signals
(betas) for each block as well as individual contrast values
for the main task effect (Perf > Rest) and repetition en-
hancement effect (Perf1 < Perf2) were extracted from
each ROI. Mean betas for each block for both sequences
(T-FOS and U-FOS) are shown in Figure 6. Repeated-
measures ANOVAs with Task (T-FOS and U-FOS) and
Repetition (Perf1 and Perf2) as within-subject factors,
Table 3. Repetition Enhancement Effects
All Participants (n = 15)
DGs Group (n = 10)
MNI Coordinates
MNI Coordinates
Label
1. RE for T-FOS (Perf1 < Perf 2)
Postcentral (+)
Precentral (+)
ParaHippocampal (−)
Hippocampus (−)
ParaHippocampal (−)
R
R
R
L
L
2. RE for U-FOS (Perf1 < Perf 2)
No significant results
–
x
51
–
27
−21
–
–
y
z
p
−19
–
−22
−13
–
–
64
–
−26
−23
–
–
.02*1
–
< .01*8
< .01*9
–
–
x
51
36
27
–
−18
y
z
p
−19
−25
−22
–
−13
64
55
−26
–
−23
.05FWE
∧
.05*8
–
.08*9
–
–
–
–
Labeling clusters obtained from activation maps thresholded at p < .001 (uncorrected) using AAL (Tzourio-Mazoyer et al., 2002). pFWE = cluster-level
FWE-corrected over the entire brain; p*[N] = peak-level FWE-corrected over a small VOI, [N] refers to a VOI used for small volume correction as listed
in Table 1; ∧ = for the cluster-level p values, areas within the same cluster as area listed above; (+) = increased task-related activity versus rest; (−) =
decreased task-related activity versus rest.
2726
Journal of Cognitive Neuroscience
Volume 26, Number 12
Figure 6. Mean betas extracted
from regions with RE effects.
Spherical ROIs with a radius of
6 mm centered within clusters
with significant RE effects are
shown over the mean structural
image of all participants. Mean
signals (betas) for each
performance block (Perf1 and
Perf2) and each sequence
(T-FOS and U-FOS) averaged
across three runs for all
participants (left plots) and
DGs group (right plots): bars,
SEM. *p ≤ .01 level. (A) Rght
(contralateral) lateral M1,
sphere ROI centered at
[48 −16 60]. (B) Right
(contralateral) M1 hand area
(knob), sphere ROI centered
at [36, −25, 55]. (C) Left
MTL, sphere ROI centered at
[−21, −13, −23]. (D) Right
MTL, sphere ROI centered
at [27, −22, −26].
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
a
/
_
j
0
o
0
c
6
n
7
5
_
a
p
_
d
0
0
b
6
y
7
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
in all participants, showed significant RE effect within
the lateral M1 with a trend toward Repetition × Task inter-
action (F(1, 14) = 7.22, p < .05; F(1, 14) = 3.64, p = .08,
Repetition and Repetition × Task interaction, respectively)
as well as significant Repetition × Task interaction within
the MTL bilaterally (F(1, 14) = 7.03, p < .05; F(1, 14) =
9.07, p < .01, left and right MTL, respectively). Post hoc
analyses performed separately for each movement se-
quence, in line with the whole-brain analyses, showed that
RE effects within the lateral M1 and MTL were significant
for the T-FOS (F(1, 14) = 12.10, p < .01; F(1, 14) = 42.52,
p < .001; F(1, 14) = 30.69, p < .001, lateral M1, left and
right MTL, respectively) but were not significant for the
U-FOS (F(1, 14) = 0.20, p = .65; F(1, 14) = 0.18, p =
.68; F(1, 14) = 1.33, p = .27, lateral M1, left and right
MTL, respectively). Significant RE effects within the M1
hand area were found only for the DGs group with signif-
icant Repetition × Task interaction (Repetition: F(1, 9) =
10.31, p = .01; Repetition × Task: F(1, 9) = 4.87, p = .055).
Post hoc analyses performed separately for each sequence
showed that RE effects within the M1 hand area were
restricted to the T-FOS within the DGs group (F(1, 9) =
23.05, p = .001; F(1, 9) = 1.25, p = .29, T-FOS and U-FOS,
respectively).
Gabitov, Manor, and Karni
2727
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
The linear correlation analyses revealed the existence
of a strong positive relationship between overnight im-
provements in speed (DGs) and RE effects during the
T-FOS performance within the right M1 hand area (Fig-
ure 4B, left plot; Table 4). Moreover, in line with the
results of the whole-brain and ROI analyses on the group
level, only participants who expressed DGs were charac-
terized by increased BOLD signal upon repetition within
this region (i.e., positive values for RE effects within the
M1 hand area; Figure 4B, left plot). There was also posi-
tive relationship between DGs and RE effects within the
right lateral M1, but this effect failed to be significant after
a Bonferroni correction (Figure 4A, left plot; Table 4).
However, the magnitude of RE effects during the T-FOS
performance within the MTL was not correlated with
the expression of DGs (Figure 5, top plots; Table 4). No
significant correlations were found between repetition
effects (Perf1 < Perf2) and participantsʼ absolute speed
achieved overnight in any of the ROIs tested (Table 4).
There were also no significant correlations between the
neural activity evoked in the four ROIs (Perf > Rest)
and either the DGs or the absolute speed achieved
overnight for the T-FOS; there was a positive correlation
of activity within the lateral M1 and the DGs, but this
effect failed to be significant after a Bonferroni correction
(Tableau 4).
DISCUSSION
The current results suggest that the pattern of neural
activity modulation upon task repetition (across per-
formance blocks in a pair), rather than the magnitude
of the average evoked signal per se, reflected the effec-
tiveness of overnight memory consolidation following
motor learning. A clear pattern of RE in the evoked
BOLD-fMRI signals characterized the paced performance
of the sequence of finger-to-thumb opposition move-
ments intensively trained a day earlier. These RE effects
were observed within the sensory-motor cortex contra-
lateral to the performing hand as well as in the MTL bilat-
erally. De plus, the magnitude of RE effects within the
contralateral M1 correlated with the DGs in the perfor-
mance of the T-FOS. Taken together these results sug-
gest that the modulation of the BOLD signal upon task
repetition constitute a reliable neural signature for off-line
memory consolidation processes.
The performance of the trained sequence a day after
the initial training was significantly faster and more accu-
rate than the performance of the untrained sequence,
composed of identical opposition movements (Figure 1C).
During the scanning session these differences in the rate
of sequence executions were minimized by requiring
participants to perform the component movements at
a comfortable but externally paced rate. Ainsi, the differ-
ences in neural activity were not directly related to faster
task execution but rather reflected differences in the
order of the component movements—trained versus
untrained (Karni et al., 1995). The brain areas activated
for the T-FOS were not different from those activated for
the U-FOS. On the other hand, upon repetition after the
brief rest interval, the U-FOS induced significant reduc-
tions in the BOLD signal within extensive brain areas,
whereas reduced activity upon repeated performance
of the T-FOS was significant only within the calcarine
(presumably reflecting the fact that the visual cues for
task performance were novel, c'est à dire., presented to the
Tableau 4. Correlations between Contrast Values and Behavioral Measures for the T-FOS
ROI
RE vs. DGs (%)
RE vs. Correct (#)
Act vs. DGs (%)
Act vs. Correct (#)
Right lateral M1 centered at [48, −16, 60]
Right M1 hand area centered at [36, −25, 55]
Right MTL centered at [27, −22, −26]
Left MTL centered at [−21, −13, −23]
r = .56
p = .03*
pcor = .11
r = .70
p < .01**
pcor = .01**
r = .17
p = .55
r = .45
p = .87
r = .02
p = .95
r = .12
p = .68
r = .24
p = .40
r = .02
p = .95
r = .61
p = .02*
pcor = .08
r = .47
p = .08
r = .47
p = .08
r = .24
p = .39
r = .18
p = .54
r = .04
p = .89
r = .20
p = .47
r = .42
p = .12
RE = repetition enhancement effects (Perf1 < Perf2); Act = task-related activity (Perf > Rest); DGs (%) = DGs in speed (Overnight T-FOS (%) −
Post-T (%)); correct (#) = the average number of correct sequences achieved overnight during the T-FOS performance test; pcor = values corrected
for multiple tests (four correlation analyses for each ROI) using Bonferroni adjustments.
*p ≤ .05 level.
**p ≤ .01 level.
2728
Journal des neurosciences cognitives
Volume 26, Nombre 12
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
t
F
.
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
participants for the first time during the imaging session;
Grill-Spector et al., 1999). RS for the untrained sequence
was apparent in most of the task-related areas, in line
with previous studies (Hamilton & Grafton, 2009; Grill-
Spector et al., 1999; Buckner et al., 1998). RS is consid-
ered a neural correlate of behavioral priming and novelty
(Grill-Spector et al., 2006; Henson, 2003; Schacter &
Buckner, 1998; Wiggs & Martine, 1998; Désimone, 1996)
and was observed in both block design (Grill-Spector
et coll., 1999; Karni et al., 1995; Raichle et al., 1994; Squire
et coll., 1992) and event-related (Sayres & Grill-Spector,
2006; Henson et al., 2000; Buckner et al., 1998) études
across various time intervals (Grill-Spector et al., 1999;
Fahy, Riches, & Brun, 1993; Miller et al., 1991). Repetition-
related reductions in neural activity may reflect a process
of selection whereby neurons that poorly represent the
task-relevant features of the initial experience drop out
upon repeated experience (Désimone, 1996; Miller et al.,
1993). From this point of view, only cells carrying critical
information for task performance continue to produce
robust activation at later stages of practice. As practice
continues, task-relevant units may actually be enhanced
after a certain amount of experience (pratique; Karni et al.,
1995, 1998) offsetting the initial suppression–selection
processus (Désimone & Duncan, 1995). Novelty and priming
effects in both behavior and the brain are of a transient
nature and, as a rule, saturate after a rather limited number
of repetitions (Hauptmann & Karni, 2002; Karni et al., 1995,
1998; Raichle et al., 1994; Karni & Sagi, 1993; Miller et al.,
1991). Training beyond the saturation of repetition priming
was proposed as a critical requirement for the expression
of DGs in performance (Hauptmann & Karni, 2002; Karni
et coll., 1998).
In the current study, all participants were trained accord-
ing to a standard FOS training protocol to ensure adequate
pratique (Korman et al., 2003, 2007). Rapid changes in per-
formance were observed before the training (Pre-T); ces
effects were saturated as a result of practice (Post-T). Given
the identical structure of the two sequences, the counter-
balancing and previous imaging studies (Karni et al., 1995,
1998), it is reasonable to assume that the initial perfor-
mance of the to-be-trained sequence was characterized
by RS effects. These effects were likely to be saturated by
the end of the training session (Karni et al., 1995, 1998).
The imaging results of the current study clearly indicate,
donc, that on the day after the initial training session
the saturation of RS for the T-FOS was maintained in all
participants. Ainsi, the training afforded in the current
study was sufficient for saturating RS effects, for the T-FOS,
irrespective to the ability to express DGs. Nevertheless, five
participants failed to express DGs in performance over-
night (Chiffre 2). Failure to express DGs overnight was
not related to the initial performance levels, the saturation
of within-test improvements in speed or the absolute
speed achieved overnight for the T-FOS. Ainsi, insufficient
training or relatively slow or inaccurate performance can-
not explain the lack of additional gains overnight. The mag-
nitude of DGs in motor sequence tasks was shown to be
sleep-dependent regardless to the time of day during
which training, retest, and sleep were afforded (Korman
et coll., 2003, 2007; Nishida & Walker, 2007; Fischer et al.,
2002; Walker et al., 2002). Failures to express DGs may
have resulted from individual differences in the recruit-
ment of the motor network in general and of M1 in par-
ticular during the training experience (Steele & Penhune,
2010). Cependant, individual differences in posttraining
experience (Tibi, Eviatar, & Karni, 2013; Balas, Roitenberg,
Giladi, & Karni, 2007; Brun & Robertson, 2007) et en
susceptibility to interference (Korman et al., 2007;
Brashers-Krug, Shadmehr, & Bizzi, 1996) as well as dif-
ferences in posttraining sleep (Korman et al., 2003, 2007;
Fischer et al., 2002; Walker et al., 2002) and sleep structure
(Barakat et al., 2013; Karni, Tanne, Rubenstein, Askenasy, &
Sagi, 1994; Maquet, 2001) may critically affect the expres-
sion of DGs.
The current results indicate that the failure to express
overnight DGs was related to the absence of RE within
the M1 hand representation area across performance
blocks of the T-FOS. There is evidence for focal increased
excitability, paralleled by surround inhibition within the
motor system that contributes to the selection of volun-
tary movements (Sohn & Hallett, 2004). Horizontal intra-
cortical axon collaterals that interconnect the entire M1
hand representation area (Huntley & Jones, 1991) may
be involved in the coordination of patterns of motor out-
put to multiple muscles. Although much shorter time
intervals (compared with the rest interval within each
run afforded in the current study) were used in animal
études, there is evidence for summation (enhancement)
effects suggesting local modulations of interconnections
under some repeated activation protocols (Boulanger, Olivier,
& Lemon, 1998). Given the notion of sleep-dependent
resetting of the overall excitatory–inhibitory balance dur-
ing the consolidation phase (Tononi & Cirelli, 2006), nous
conjecture that the local circuitry underlying the execu-
tion of the trained movement sequence may undergo
such resetting during a successful consolidation interval
that includes sleep. Given our behavioral and imaging
résultats, this resetting is presumably specific for the
trained and successfully consolidated sequence of move-
ments. Nevertheless, there were no RS effects in the contra-
lateral motor hand area for the execution of the same
movements arranged in a different, novel, order (U-FOS;
Figures 3, bottom, and 6B). This absence of RS effects
within the contralateral motor hand area a day after training
may reflect the prior experience with movement com-
ponents of this sequence. The results of Karni et al.
(1995, 1998) have shown that, during the initial phase
in the motor skill acquisition, some of the repetition-
dependent BOLD signal modulations (RS) in M1 contra-
lateral to the performing hand could be ascribed to the
overlap in the representation of the component move-
ments. Ainsi, RS effects may reflect the identity of the
component movements irrespective of their order when
Gabitov, Manor, and Karni
2729
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
t
F
.
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
both sequences are introduced concurrently (Karni et al.,
1998). In the current study, the untrained sequence was
introduced to the participants after the trained sequence
was extensively practiced a day earlier. Ainsi, the novelty
of the component movements of the U-FOS in terms of
M1 activity may have been reduced.
It is not known whether the modulation of neural
activity upon task repetition is dependent on the length
and nature of the rest interval inserted between the two
performance intervals. The number and rate of task iter-
ations (the block length) may also be important factors
in the modulation of BOLD signal to task repetition.
Adaptation studies in the visual system showed that the
magnitude of repetition effects was increased with longer
exposures to the stimuli (Grill-Spector et al., 1999) et
decreased with longer ISIs (Henson et al., 2000; Grill-
Spector et al., 1999). Single cell recordings in studies
using the DMS task, with multiple intervening items
between the sample and matching test stimulus, showed
that activity within visual processing regions exhibited RS
as well as match enhancement (Miller & Désimone, 1994;
Miller et al., 1991, 1993). These effects bridged up to
six intervening stimuli (∼8 sec) within a trial (Miller
et coll., 1991). Cependant, there was no modulation of re-
sponses when a sample on one trial was repeated on
the next trial following the intertrial interval (1–2 sec),
ruling out temporal contiguity alone as the explanation
of the sample-match effects (Miller & Désimone, 1994;
Miller et al., 1993). It has been suggested that the failure
of repetition effects to carry across trials in the DMS task
indicates the presence of an active reset mechanism that
reboots the memory traces to avoid cross-trial inter-
ference or to prime neural cells with the memory of a
sample (Miller et al., 1993). In the current study, repe-
tition enhancement effects across blocks in a pair could
be reproduced in successive runs, which were separated
by breaks (1.5–2 min) dedicated to verbal interaction
with participants (Figures 4 et 5). This suggests that
such an interval was sufficient to recover the repetition
effects in line with previously reported results (Karni et al.,
1995, 1998). One cannot rule out that the verbal inter-
action with the participants during the break between runs
was also a factor in the recovery of the repetition effects.
There is consistent evidence from animal and human
studies that M1 plays a central role in the long-term re-
tention of motor skills following multisession training
(Mandelblat-Cerf et al., 2011; Matsuzaka, Picard, & Strick,
2007; Floyer-Lea & Matthieu, 2005; Kleim et al., 2004;
Penhune & Doyon, 2002; Karni et al., 1995, 1998; Kleim,
Barbay, & Nudo, 1998; Nudo, Milliken, Jenkins, & Merzenich,
1996). De plus, it has been recently shown that extended
practice of the motor skill stabilizes the M1 activity pattern
(Huang et al., 2013) and is associated with reduced meta-
bolic activity in M1 (Picard, Matsuzaka, & Strick, 2013).
Cependant, a number of studies suggest that regions other
than M1, Par exemple, cerebellum and BG (Lehéricy
et coll., 2005; Doyon et al., 2002), as well as the hippocampus
(Albouy et al., 2008), contribute to early phases of acqui-
sition of motor skill. Although some studies reported
an increase in M1 activation even after a single session of
entraînement (Albouy et al., 2012; Orban et al., 2010; Honda
et coll., 1998), these effects can be ascribed to enhance task
execution, Par exemple, an increase in speed of perfor-
mance, rather than to the effects of sequence learning
per se (Orban et al., 2010; Honda et al., 1998). In the cur-
rent study, the rate of sequence execution during scanning
was identical in all participants and for both sequences
(paced performance). De plus, there was no significant
correlation between the individualsʼ actual performance
speed for the T-FOS overnight and the magnitude of either
average activity or repetition effects in M1.
Time-dependent changes in performance, such as the
expression of DGs, were hypothesized to reflect neuron-
level memory consolidation processes (Karni, 1996; Karni
& Sagi, 1993). Recent studies, using in vivo transcranial
two-photon microscopy to examine changes within pyra-
midal neurons in the mouse motor cortex, showed that
synaptic connections rapidly respond to motor experi-
ence in a novel task with the formation of new dendritic
spines (Xu et al., 2009; Yang et al., 2009). The extent of
spine remodeling in motor cortex correlated with delayed
behavioral improvement (Xu et al., 2009; Yang et al.,
2009). De plus, new spines that were induced during
the early stage of learning tended to stabilize (Xu et al.,
2009), suggesting lasting experience-specific synaptic re-
organization within motor cortex.
Studies of repetitive TMS indicate that M1 is implicated
in the initiation of long-term motor memory (Richardson
et coll., 2006), including presumably early stages (5–30 min
after training) of motor skill consolidation following train-
ing (Hotermans et al., 2008; Baraduc, Lang, Rothwell, &
Wolpert, 2004; Muellbacher et al., 2002). Cependant, after
practice on the motor sequence task inhibition to the
contralateral M1, induced by repetitive TMS, abolished
only the daytime improvements (Robertson et al.,
2005) and did not fully abolish the early posttraining
boost effect in performance (Hotermans et al., 2008).
Brain structures other than M1 may have a role in off-line
memory processes: importantly, the corticostriatal net-
travaux (Doyon & Benali, 2005; Lehéricy et al., 2005)
and even interactions with the hippocampus (Albouy
et coll., 2008, 2013; Hobson & Pace-Schott, 2002).
A recent study (Steele & Penhune, 2010) suggests that
the recruitment of M1 during the initial training session
predicts the overnight improvement of performance.
Cependant, Steele and Penhune (2010) found no signifi-
cant difference in the average M1 activation overnight,
c'est, before and after the consolidation phase following
initial training—a result that is in line with the current
findings. Ainsi, the magnitude or extent of BOLD signals
within M1 may not reflect familiarity with a movement
sequence or procedural memory consolidation processes;
both conditions, cependant, are reflected in the pattern of
neural activity upon task repetition.
2730
Journal des neurosciences cognitives
Volume 26, Nombre 12
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
/
F
.
t
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
There were T-FOS-specific RE effects in the MTL. Un
should note that the MTL modulation pattern is somewhat
different from that of M1 because the evoked signals were
negative (compared with rest). Ainsi, the repetition of the
T-FOS induced less suppression of the MTL activity. Le
MTL is part of the resting state (default) réseau, exhibit-
ing consistent activity decreases during task performance
compared with rest (Buckner et al., 2008; Raichle et al.,
2001). The suppression of the resting state network has
been shown to be reduced when a task becomes more famil-
iar (McKiernan, Kaufman, Kucera-Thompson, & Binder,
2003), less difficult (McKiernan et al., 2003), and well prac-
ticed (Kincses et al., 2008). The decrease in task-induced
deactivation as a function of task repetition was previously
reported during repeated exposure to visual stimuli of
familiar and unfamiliar objects and may reflect reduced re-
quirements for attention and cognitive control (Soldan
et coll., 2008). It has been suggested that cognitive control
may impede procedural mnemonic processes (Brun &
Robertson, 2007). Recent studies suggest that hippocampal
activity may be correlated with the ability to engage pro-
cedural memory processes (Albouy et al., 2008, 2013). Nous
propose that the emergence of RE within M1 contralateral
to the performing hand and the decreased deactivations
upon task repetition within the MTL reflect two aspects of
procedural memory: the formation of a differential neural
representation for the trained sequence and the decreased
demand on control and attention processes in the execution
of the trained sequence, respectivement.
Altogether, our results suggest that delayed perfor-
mance gains, but not absolute performance levels, sont
related to changes in the way the brain repeatedly gener-
ates a movement sequence after its consolidation in
mémoire. We propose that procedural memory consoli-
dation processes may affect the excitation–inhibition
balance within cortical representations of the trained
mouvements; this new balance is better reflected in repe-
tition effects than in the average level of evoked neural
activité. We conjecture that the local circuitry underlying
the execution of the trained movement sequence may
undergo a resetting during a successful consolidation
interval. This resetting relates to the movement sequence
rather than the component movements. Ainsi, the mode
of the neural activity in motor cortex to repeated task
iterations constitutes a neural signature for motor expe-
rience and specifically for the effectiveness of motor
memory consolidation processes.
Remerciements
E. G. was partially supported by a fellowship from the E. J.. Safra
Brain Research Center for the Study of Learning Disabilities. Nous
thank Tammi Kushnir, Ph.D., director of research at the C. Sheba
Medical Center, for her technical and administrative support.
Reprint requests should be sent to Ella Gabitov, Université de
Haifa, 199 Abba Hushi Blvd., Mount Carmel, Haifa 3498838,
Israel, ou par e-mail: gabitovella@gmail.com.
RÉFÉRENCES
Adams, J.. UN. (1987). Historical review and appraisal of research
on the learning, retention, and transfer of human motor
skills. Psychological Bulletin, 101, 41–74.
Albouy, G., Sterpenich, V., Balteau, E., Vandewalle, G.,
Desseilles, M., Dang-Vu, T., et autres. (2008). Both the
hippocampus and striatum are involved in consolidation of
motor sequence memory. Neurone, 58, 261–272.
Albouy, G., Sterpenich, V., Vandewalle, G., Darsaud, UN., Gais, S.,
Rauchs, G., et autres. (2012). Neural correlates of performance
variability during motor sequence acquisition. Neuroimage,
60, 324–331.
Albouy, G., Sterpenich, V., Vandewalle, G., Darsaud, UN.,
Gais, S., Rauchs, G., et autres. (2013). Interaction between
hippocampal and striatal systems predicts subsequent
consolidation of motor sequence memory. PloS One,
8, e59490.
Andersson, J.. L. R., Hutton, C., Ashburner, J., Tourneur, R., &
Friston, K. J.. (2001). Modeling geometric deformations in
EPI time series. Neuroimage, 13, 903–919.
Boulanger, S. N., Olivier, E., & Lemon, R.. N. (1998). An investigation
of the intrinsic circuitry of the motor cortex of the monkey
using intra-cortical microstimulation. Experimental Brain
Research, 123, 397–411.
Balas, M., Roitenberg, N., Giladi, N., & Karni, UN. (2007). Quand
practice does not make perfect: Well-practiced handwriting
interferes with the consolidation phase gains in learning a
movement sequence. Experimental Brain Research, 178,
499–508.
Baraduc, P., Lang, N., Rothwell, J.. C., & Wolpert, D. M.. (2004).
Consolidation of dynamic motor learning is not disrupted
by rTMS of primary motor cortex. Biologie actuelle, 14,
252–256.
Barakat, M., Carrier, J., Debas, K., Lungu, O., Fogel, S.,
Vandewalle, G., et autres. (2013). Sleep spindles predict neural
and behavioral changes in motor sequence consolidation.
Human Brain Mapping, 34, 2918–2928.
Brashers-Krug, T., Shadmehr, R., & Bizzi, E. (1996). Consolidation
in human motor memory. Nature, 382, 252–255.
Brett, M., Anton, J.-L., Valabregue, R., & Poline, J.-B. (2002).
Region of interest analysis using an SPM toolbox
[abstract]. Presented at the 8th International Conference
on Functional Mapping of the Human Brain, June 2–6, 2002,
Sendai, Japan. Available on CD-ROM in Neuroimage,
16 (2).
Brun, R.. M., & Robertson, E. M.. (2007). Offline processing:
Reciprocal interactions between declarative and procedural
memories. Journal des neurosciences, 27, 10468–10475.
Buckner, R.. L., Andrews-Hanna, J.. R., & Schacter, D. L. (2008).
The brainʼs default network: Anatomy, fonction, et
relevance to disease. Annals of the New York Academy of
les sciences, 1124, 1–38.
Buckner, R.. L., Homme bon, J., Burock, M., Rotte, M., Koutstaal,
W., Schacter, D. L., et autres. (1998). Functional-anatomic
correlates of object priming in humans revealed by
rapid presentation event-related fMRI. Neurone, 20,
285–296.
Cothros, N., Köhler, S., Dickie, E. W., Mirsattari, S. M., &
Gribble, P.. L. (2006). Proactive interference as a result of
persisting neural representations of previously learned motor
skills in primary motor cortex. Journal of Cognitive
Neurosciences, 18, 2167–2176.
Debas, K., Carrier, J., Orban, P., Barakat, M., Lungu, O.,
Vandewalle, G., et autres. (2010). Brain plasticity related to the
consolidation of motor sequence learning and motor
adaptation. Proceedings of the National Academy of
les sciences, USA., 107, 17839–17844.
Gabitov, Manor, and Karni
2731
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
t
.
F
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Désimone, R.. (1996). Neural mechanisms for visual memory
and their role in attention. Actes de la Nationale
Académie des Sciences, USA., 93, 13494–13499.
Désimone, R., & Duncan, J.. (1995). Neural mechanisms of
selective visual attention. Revue annuelle des neurosciences,
18, 193–222.
Dinstein, JE., Hasson, U., Rubin, N., & Heeger, D. J..
(2007). Brain areas selective for both observed and
executed movements. Journal de neurophysiologie, 98,
1415–1427.
Doyon, J., & Benali, H. (2005). Reorganization and plasticity
in the adult brain during learning of motor skills. Actuel
Opinion in Neurobiology, 15, 161–167.
Doyon, J., Korman, M., Morin, UN., Dostie, V., Hadj Tahar, UN.,
Benali, H., et autres. (2009). Contribution of night and day sleep
vs. simple passage of time to the consolidation of motor
sequence and visuomotor adaptation learning. Experimental
Brain Research, 195, 15–26.
Doyon, J., Song, UN. W., Karni, UN., Lalonde, F., Adams, M.. M., &
Ungerleider, L. G. (2002). Experience-dependent changes in
cerebellar contributions to motor sequence learning.
Actes de l'Académie nationale des sciences, USA.,
99, 1017–1022.
Fahy, F. L., Riches, je. P., & Brun, M.. W. (1993). Neuronal
activity related to visual recognition memory: Long-term
memory and the encoding of recency and familiarity
information in the primate anterior and medial inferior
temporal and rhinal cortex. Experimental Brain Research,
96, 457–472.
Fischer, S., Hallschmid, M., Elsner, UN. L., & Born, J.. (2002). Sleep
forms memory for finger skills. Actes de la Nationale
Académie des Sciences, USA., 99, 11987–11991.
Fischer, S., Nitschke, M.. F., Melchert, U. H., Erdmann, C., &
Born, J.. (2005). Motor memory consolidation in sleep shapes
more effective neuronal representations. Journal de
Neurosciences, 25, 11248–11255.
Floyer-Lea, UN., & Matthieu, P.. M.. (2005). Distinguishable
brain activation networks for short- and long-term
motor skill learning. Journal de neurophysiologie, 94,
512–518.
Friston, K. J., Holmes, UN. P., Worsley, K. J., Poline, J.-B., Frith,
C. D., & Frackowiak, R.. S. J.. (1995). Statistical parametric
maps in functional imaging: A general linear approach. Human
Brain Mapping, 2, 189–210.
Grafton, S. T., & Hamilton, UN. F. de C. (2007). Evidence for a
distributed hierarchy of action representation in the brain.
Human Movement Science, 26, 590–616.
Grafton, S. T., Hazeltine, E., & Ivry, R.. B. (2002). Motor
sequence learning with the nondominant left hand. A PET
functional imaging study. Experimental Brain Research,
146, 369–378.
Grill-Spector, K., Henson, R.. N. UN., & Martine, UN. (2006).
Repetition and the brain: Neural models of stimulus-specific
effects. Tendances des sciences cognitives, 10, 14–23.
Grill-Spector, K., Kushnir, T., Edelman, S., Avidan, G., Itzchak,
Y., & Malach, R.. (1999). Differential processing of objects
under various viewing conditions in the human lateral occipital
complexe. Neurone, 24, 187–203.
Halsband, U., & Lange, R.. K. (2006). Motor learning in man:
A review of functional and clinical studies. Journal de
Physiology, 99, 414–424.
Hamilton, UN. F. de C., & Grafton, S. T. (2009). Repetition
suppression for performed hand gestures revealed by fMRI.
Human Brain Mapping, 30, 2898–2906.
Hauptmann, B., & Karni, UN. (2002). From primed to learn:
The saturation of repetition priming and the induction of
long-term memory. Brain Research, Cognitive Brain Research,
13, 313–322.
Henson, R.. N. UN. (2003). Neuroimaging studies of priming.
Progrès en neurobiologie, 70, 53–81.
Henson, R.. N. UN., Shallice, T., & Dolan, R.. (2000). Neuroimaging
evidence for dissociable forms of repetition priming. Science,
287, 1269–1272.
Hikosaka, O., Nakamura, K., Sakai, K., & Nakahara, H. (2002).
Central mechanisms of motor skill learning. Current Opinion
in Neurobiology, 12, 217–222.
Hobson, J.. UN., & Pace-Schott, E. F. (2002). The cognitive
neuroscience of sleep: Neuronal systems, consciousness and
learning. Nature Revues Neurosciences, 3, 679–693.
Honda, M., Deiber, M.-P., Ibáñez, V., Pascual-Leone, UN., Zhuang,
P., & Hallett, M.. (1998). Dynamic cortical involvement in
implicit and explicit motor sequence learning. A PET study.
Cerveau, 121, 2159–2173.
Hotermans, C., Peigneux, P., de Noordhout, UN. M., Moonen, G.,
& Maquet, P.. (2008). Repetitive transcranial magnetic
stimulation over the primary motor cortex disrupts early
boost but not delayed gains in performance in motor
sequence learning. European Journal of Neuroscience, 28,
1216–1221.
Huang, Y., Zhen, Z., Song, Y., Zhu, Q., Wang, S., & Liu, J.. (2013).
Motor training increases the stability of activation patterns
in the primary motor cortex. PLoS One, 8, e53555.
Huntley, G. W., & Jones, E. G. (1991). Relationship of intrinsic
connections to forelimb movement representations in monkey
motor cortex: A correlative anatomic and physiological study.
Journal of Physiology, 66, 390–413.
Hutton, C., Andersson, J., Deichmann, R., & Weiskopf, N. (2013).
Phase informed model for motion and susceptibility. Human
Brain Mapping, 34, 3086–3100.
Johnstone, T., Ores Walsh, K. S., Greischar, L. L., Alexander,
UN. L., Fox, UN. S., Davidson, R.. J., et autres. (2006). Motion
correction and the use of motion covariates in multiple-
subject fMRI analysis. Human Brain Mapping, 27,
779–788.
Karni, UN. (1996). The acquisition of perceptual and motor skills:
A memory system in the adult human cortex. Brain Research,
Cognitive Brain Research, 5, 39–48.
Karni, UN., Meyer, G., Jezzard, P., Adams, M.. M., Tourneur, R., &
Ungerleider, L. G. (1995). Functional MRI evidence for adult
motor cortex plasticity during motor skill learning. Nature,
377, 155–158.
Karni, UN., Meyer, G., Rey-Hipolito, C., Jezzard, P., Adams, M.. M.,
Tourneur, R., et autres. (1998). The acquisition of skilled motor
performance: Fast and slow experience-driven changes in
primary motor cortex. Proceedings of the National Academy
of Sciences, USA., 95, 861–868.
Karni, UN., & Sagi, D. (1993). The time course of learning a visual
skill. Nature, 365, 250–252.
Karni, UN., Tanne, D., Rubenstein, B. S., Askenasy, J.. J., &
Sagi, D. (1994). Dependence on REM sleep of overnight
improvement of a perceptual skill. Science, 265, 679–682.
Kiesel, UN., Steinhauser, M., Wendt, M., Falkenstein, M., Jost, K.,
Philipp, UN. M., et autres. (2010). Control and interference in
task switching—A review. Psychological Bulletin, 136,
849–874.
Kincses, Z. T., Johansen-Berg, H., Tomassini, V., Bosnell, R.,
Matthieu, P.. M., & Beckmann, C. F. (2008). Model-free
characterization of brain functional networks for motor
sequence learning using fMRI. Neuroimage, 39,
1950–1958.
Kleim, J.. UN., Barbay, S., & Nudo, R.. J.. (1998). Functional
reorganization of the rat motor cortex following motor skill
learning. Journal de neurophysiologie, 80, 3321–3325.
Kleim, J.. UN., Hogg, T. M., VandenBerg, P.. M., Tonnelier, N. R.,
Bruneau, R., & Remple, M.. (2004). Cortical synaptogenesis
and motor map reorganization occur during late, but not
2732
Journal des neurosciences cognitives
Volume 26, Nombre 12
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
.
F
/
t
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
early, phase of motor skill learning. Journal des neurosciences,
24, 628–633.
Korman, M., Doyon, J., Doljansky, J., Carrier, J., Dagan, Y., &
Karni, UN. (2007). Daytime sleep condenses the time course of
motor memory consolidation. Neurosciences naturelles, 10,
1206–1213.
Korman, M., Raz, N., Flash, T., & Karni, UN. (2003). Multiple
shifts in the representation of a motor sequence during
the acquisition of skilled performance. Actes du
Académie nationale des sciences, USA., 100, 12492–12497.
Krakauer, J.. W., & Shadmehr, R.. (2006). Consolidation of motor
mémoire. Trends in Neurosciences, 29, 58–64.
Lehéricy, S., Benali, H., Van de Moortele, P.-F., Pélégrini-Issac,
M., Waechter, T., Voiture d'Ugur, K., et autres. (2005). Distinct basal
ganglia territories are engaged in early and advanced motor
sequence learning. Proceedings of the National Academy
of Sciences, USA., 102, 12566–12571.
Mandelblat-Cerf, Y., Novick, JE., Paz, R., Link, Y., Freeman, S.,
& Vaadia, E. (2011). The neuronal basis of long-term
sensorimotor learning. Journal des neurosciences, 31,
300–313.
Maquet, P.. (2001). The role of sleep in learning and memory.
Science, 294, 1048–1052.
Matsuzaka, Y., Picard, N., & Strick, P.. L. (2007). Skill
representation in the primary motor cortex after long-term
pratique. Journal de neurophysiologie, 97, 1819–1832.
Mayka, M.. UN., Corcos, D. M., Leurgans, S. E., & Vaillancourt,
D. E. (2006). Three-dimensional locations and boundaries
of motor and premotor cortices as defined by functional
brain imaging: A meta-analysis. Neuroimage, 31,
1453–1474.
Mazaika, P.. K., Hoeft, F., Glover, G. H., & Reiss, UN. L. (2009).
Methods and software for fMRI analysis of clinical
sujets. Annual Meeting of the Organization for Human
Brain Mapping.
McKiernan, K. UN., Kaufman, J.. N., Kucera-Thompson, J., &
Binder, J.. R.. (2003). A parametric manipulation of factors
affecting task-induced deactivation in functional neuroimaging.
Journal des neurosciences cognitives, 15, 394–408.
Miller, E. K., & Désimone, R.. (1994). Parallel neuronal mechanisms
for short-term memory. Science, 263, 520–522.
Miller, E. K., Li, L., & Désimone, R.. (1991). Un mécanisme neuronal
for working and recognition memory in inferior temporal
cortex. Science, 254, 1377–1379.
Miller, E. K., Li, L., & Désimone, R.. (1993). Activity of neurons
in anterior inferior temporal cortex during a short-term
memory task. Journal des neurosciences, 13, 1460–1478.
Muellbacher, W., Ziemann, U., Wissel, J., Dang, N., Kofler, M.,
Facchini, S., et autres. (2002). Early consolidation in human
primary motor cortex. Nature, 415, 640–644.
Nishida, M., & Walker, M.. P.. (2007). Daytime naps, moteur
memory consolidation and regionally specific sleep spindles.
PloS One, 2, e341.
Nudo, R.. J., Milliken, G. W., Jenkins, W. M., & Merzenich, M.. M..
(1996). Use-dependent alterations of movement
representations in primary motor cortex of adult squirrel
monkeys. Journal des neurosciences, 16, 785–807.
Oldfield, R.. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsychologie,
9, 97–113.
Orban, P., Peigneux, P., Lungu, O., Albouy, G., Breton, E.,
Laberenne, F., et autres. (2010). The multifaceted nature of the
relationship between performance and brain activity in motor
sequence learning. Neuroimage, 49, 694–702.
Penhune, V. B., & Doyon, J.. (2002). Dynamic cortical and
subcortical networks in learning and delayed recall of
timed motor sequences. Journal des neurosciences, 22,
1397–1406.
Penhune, V. B., & Doyon, J.. (2005). Cerebellum and M1
interaction during early learning of timed motor sequences.
Neuroimage, 26, 801–812.
Picard, N., Matsuzaka, Y., & Strick, P.. L. (2013). Extended
practice of a motor skill is associated with reduced
metabolic activity in M1. Neurosciences naturelles, 16,
1340–1347.
Raichle, M.. E., Fiez, J.. UN., Videen, T. O., MacLeod, A.-M. K.,
Pardo, J.. V., Fox, P.. T., et autres. (1994). Practice-related changes
in human brain functional anatomy during nonmotor
learning. Cortex cérébral, 4, 8–26.
Raichle, M.. E., MacLeod, A.-M. K., Snyder, UN. Z., Powers, W. J.,
Gusnard, D. UN., & Shulman, G. L. (2001). A default mode of
brain function. Proceedings of the National Academy of
les sciences, USA., 98, 676–682.
Rao, S. M., Bandettini, P.. UN., Binder, J.. R., Bobholz, J.. UN.,
Hammeke, T. UN., Stein, E. UN., et autres. (1996). Relationship
between finger movement rate and functional magnetic
resonance signal change in human primary motor cortex.
Journal of Cerebral Blood Flow & Metabolism, 16,
1250–1254.
Reis, J., Fischer, J.. T., Prichard, G., Weiller, C., Cohen, L. G.,
& Fritsch, B. (2013). Time- but not sleep-dependent
consolidation of tDCS-enhanced visuomotor skills. Cérébral
Cortex. est ce que je:10.1093/cercor/bht208.
Richardson, UN. G., Overduin, S. UN., Valero-Cabré, UN., Padoa-
Schioppa, C., Pascual-Leone, UN., Bizzi, E., et autres. (2006).
Disruption of primary motor cortex before learning impairs
memory of movement dynamics. Journal des neurosciences,
26, 12466–12470.
Robertson, E. M., Pascual-Leone, UN., & Presse, D. Z. (2004).
Awareness modifies the skill-learning benefits of sleep.
Biologie actuelle, 14, 208–212.
Robertson, E. M., Presse, D. Z., & Pascual-Leone, UN. (2005).
Offline learning and the primary motor cortex. Journal de
Neurosciences, 25, 6372–6378.
Sayres, R., & Grill-Spector, K. (2006). Object-selective cortex
exhibits performance-independent repetition suppression.
Journal de neurophysiologie, 95, 995–1007.
Schacter, D. L., & Buckner, R.. L. (1998). Priming and the brain
revoir. Neurone, 20, 185–195.
Forgeron, J.. F., Chen, K., Johnson, S., Morrone-Strupinsky, J.,
Reiman, E. M., Nelson, UN., et autres. (2006). Analyse de réseau
of single-subject fMRI during a finger opposition task.
Neuroimage, 32, 325–332.
Sohn, Oui. H., & Hallett, M.. (2004). Surround inhibition in
human motor system. Experimental Brain Research, 158,
397–404.
Soldan, UN., Particules, E., Hilton, H. J., & Arrière, Oui. (2008). Global
familiarity of visual stimuli affects repetition-related neural
plasticity but not repetition priming. Neuroimage, 39,
515–526.
Song, S., Howard, J.. H., Jr., & Howard, D. V. (2007). Sleep does
not benefit probabilistic motor sequence learning. Journal de
Neurosciences, 27, 12475–12483.
Squire, L. R., Ojemann, J.. G., Miezin, F. M., Petersen, S. E.,
Videen, T. O., & Raichle, M.. E. (1992). Activation of the
hippocampus in normal humans: A functional anatomical
study of memory. Proceedings of the National Academy of
les sciences, USA., 89, 1837–1841.
Steele, C. J., & Penhune, V. B. (2010). Specific increases within
global decreases: A functional magnetic resonance imaging
investigation of five days of motor sequence learning.
Journal des neurosciences, 30, 8332–8341.
Tibi, R., Eviatar, Z., & Karni, UN. (2013). Fact retrieval and memory
consolidation for a movement sequence: Bidirectional effects
of “unrelated” cognitive tasks on procedural memory
(F. Di Russo, Ed.). PLoS One, 8, e80270.
Gabitov, Manor, and Karni
2733
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
t
.
F
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Tononi, G., & Cirelli, C. (2006). Sleep function and synaptic
homeostasis. Sleep Medicine Reviews, 10, 49–62.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., et autres. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
Neuroimage, 15, 273–289.
Valyear, K. F., Gallivan, J.. P., McLean, D. UN., & Culham, J.. C.
(2012). fMRI repetition suppression for familiar but not
arbitrary actions with tools. Journal des neurosciences, 32,
4247–4259.
Walker, M.. P., Brakefield, T., Morgan, UN., Hobson, J.. UN., &
Stickgold, R.. (2002). Practice with sleep makes perfect:
Sleep-dependent motor skill learning. Neurone, 35, 205–211.
Wiggs, C. L., & Martine, UN. (1998). Properties and mechanisms of
perceptual priming. Opinion actuelle en neurobiologie, 8,
227–233.
Xu, T., Yu, X., Perlik, UN. J., Tobin, W. F., Zweig, J.. UN., Tennant,
K. UN., et autres. (2009). Rapid formation and selective stabilization
of synapses for enduring motor memories. Nature, 462,
915–919.
Lequel, G., Pan, F., & Gan, W.-B. (2009). Stably maintained
dendritic spines are associated with lifelong memories.
Nature, 462, 920–924.
Yousry, T. UN., Schmid, U. D., Alkadhi, H., Schmidt, D., Peraud,
UN., Buettner, UN., et autres. (1997). Localization of the motor hand
area to a knob on the precentral gyrus. A new landmark.
Cerveau, 120, 141–157.
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
6
/
1
2
2
6
/
2
1
7
2
1
/
6
2
1
7
9
1
4
6
8
/
4
1
0
9
7
8
o
2
c
3
n
8
_
9
un
/
_
j
0
o
0
c
6
n
7
5
_
un
p
_
d
0
0
b
6
oui
7
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
.
F
/
t
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
2734
Journal des neurosciences cognitives
Volume 26, Nombre 12