Dissociating Landmark Stability from Orienting Value
Using Functional Magnetic Resonance Imaging
Stephen D. Auger and Eleanor A. Maguire
Abstrait
■ Retrosplenial cortex (RSC) plays a role in using environmental
landmarks to help orientate oneself in space. It has also been
consistently implicated in processing landmarks that remain fixed
in a permanent location. Cependant, it is not clear whether the
RSC represents the permanent landmarks themselves or instead
the orienting relevance of these landmarks. In previous functional
magnetic resonance imaging (IRMf) études, these features have
been conflated—stable landmarks were always useful for orient-
ing. Ici, we dissociated these two key landmark attributes to
investigate which one best reflects the function of the RSC.
Before scanning, participants learned the features of novel land-
marks about which they had no prior knowledge. During fMRI
scanning, we found that the RSC was more engaged when peo-
ple viewed permanent compared with transient landmarks and
was not responsive to the orienting relevance of landmarks. Ac-
tivity in RSC was also related to the amount of landmark perma-
nence information a person had acquired and, as knowledge
increased, the more the RSC drove responses in the anterior
thalamus while viewing permanent landmarks. In contrast, le
angular gyrus and the hippocampus were engaged by the orient-
ing relevance of landmarks, but not their permanence, avec le
hippocampus also sensitive to the distance between relevant
landmarks and target locations. We conclude that the coding
of permanent landmarks in RSC may drive processing in regions
like anterior thalamus, with possible implications for the efficacy
of functions such as navigation. ■
INTRODUCTION
Landmarks are an essential component of our spatial rep-
resentations of the environment (Burnett, Forgeron, & May,
2001; Siegel & Blanc, 1975; Lynch, 1960). Au cours des dernières années
there has been increased interest in characterizing the
features of landmarks, in particular seeking to identify
those traits that are helpful for building environmental
representations and that facilitate effective wayfinding
(Auger, Zeidman, & Maguire, 2017; Marchette, Vass,
Ryan, & Epstein, 2015; Auger & Maguire, 2013; Auger,
Mullally, & Maguire, 2012; Konkle & Oliva, 2012; Lew,
2011; Yoder, Clark, & Taube, 2011; Galati, Pelle, Berthoz,
& Committeri, 2010; Committeri et al., 2004; Janzen &
van Turennout, 2004). The brain areas that process these
landmark features have also begun to be scrutinized with
a view to understanding the neural evolution of environ-
mental representations and the mechanisms involved
(Alexander & Nitz, 2017; Auger et al., 2017; Chrastil, Sherrill,
Aselcioglu, Hasselmo, & Arrière, 2017; Mao, Kandler,
McNaughton, & Bonin, 2017; Vedder, Miller, Harrison, &
Forgeron, 2017; Shine, Valdés-Herrera, Hegarty, & Wolbers,
2016; Auger, Zeidman, & Maguire, 2015; Baumann &
Mattingley, 2013; Aggleton, 2010; Iaria, Chen, Guariglia,
Ptito, & Pétrides, 2007; Wolbers, Weiller, & Büchel, 2004).
Auger et al. (2012; see also Troiani, Stigliani, Forgeron, &
Epstein, 2014) examined features of everyday outdoor
University College London
landmarks, including their size, visual salience, et
whether the landmark had a stable and fixed location.
They found that, although the parahippocampal cortex
(PHC) processed visual features such as size and salience,
the retrosplenial cortex (RSC) responded to landmark
permanence, specifically only to landmarks that never
moved and were completely fixed in their location. En addition-
dition, Auger et al. (2012) showed that self-declared good
navigators were more consistent than poor navigators at
identifying the most permanent landmarks and had
increased engagement of the RSC and the anterior thal-
amus (AThal), a region heavily connected with the RSC
( Jankowski et al., 2013; Vann, Aggleton, & Maguire,
2009), when viewing those items. It has also been possi-
ble to decode how many permanent landmarks were in
view from functional magnetic resonance imaging (IRMf)
activity in the RSC, but not from other brain areas (Auger
& Maguire, 2013). This shows that the RSC was not sim-
ply engaged by the presence of permanence per se but
was mechanistically more nuanced, tracking the specific
number of permanent items.
When studying individual outdoor landmarks or envi-
ronments, fMRI studies, including those described above,
have typically used items that are already known to par-
ticipants from the real world (Tu et al., 2015; Auger &
Maguire, 2013; Auger et al., 2012; Baumann, Chan, &
Mattingley, 2010; Iaria et al., 2007; Spiers & Maguire,
2006; Wolbers et al., 2004). To understand how an
© 2018 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 3.0 unported (CC PAR 3.0) Licence.
Journal des neurosciences cognitives 30:5, pp. 698–713
est ce que je:10.1162/jocn_a_01231
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
un
_
0
1
2
3
1
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
environmental representation develops de novo,
Auger et al. (2015) devised a virtual environment that
was populated by entirely novel
landmarks about
which participants had no prior experience. Quelques
of these items moved every time they were seen,
whereas others stayed fixed in their locations. Quand
these landmarks were learned during the course of
fMRI scanning, Auger et al. (2015) found that the RSC
became selectively engaged by nonmoving, permanent
landmarks and not those which constantly changed their
location. A posterior part of the parieto-occipital sulcus
(POS) initially responded to the most memorable land-
marks, but as more was learned about them, it switched
to instead become engaged by the permanent items.
The hippocampus was eventually activated by the perma-
nent landmarks at the end of the learning phase. Also at
this point, hippocampal responses to permanent land-
marks emerged. De plus, the hippocampus showed
increased functional coupling with the RSC, and activity
patterns within the hippocampus mapped onto how
much participants knew about where the permanent
landmarks were located within the environment. Ce
study shows the significant influence of the RSC and
the key contribution of permanent landmarks in the for-
mation of environmental representations.
Dans l'ensemble, these findings suggest that the RSC plays a role
in representing landmarks and in particular their inherent
permanence. This could be linked to the presence of head
direction cells that have been identified within the rodent
RSC (Cho & Sharp, 2001; Chen, Lin, Vert, Barnes, &
McNaughton, 1994), perhaps suggesting a mechanism
whereby head direction cell firing is centered upon perma-
nent landmarks and this information is integrated within
RSC (Bicanski & Bourgeois, 2016; Auger et al., 2012). C'est
notable that the rodent AThal and the subiculum of the
hippocampus also contain head direction cells (Taube &
Muller, 1998; Taube, 1995; Taube, Muller, & Ranck, 1990)
and perhaps form a circuit with the RSC based around per-
manent environmental features (see also Marchette et al.,
2015). Recently, human RSC and AThal have also been
shown to contain a head direction signal (Shine et al., 2016).
Jacob et al. (2017) investigated how landmarks in-
fluence the brain’s computation of head direction in ro-
dents by dissociating local landmarks and global direction
in a bidirectionally symmetrical environment. They found
that some RSC neurons showed bidirectional firing pat-
terns, which may allow them to mediate both ways be-
tween visual landmarks and the global head direction
signal. In this way, the RSC could use landmarks to com-
pute head direction and, at the same time, use the head
direction signal to compute landmark stability. Le
authors suggest that this points to a mechanism for asso-
ciating landmarks to or dissociating them from the head
direction signal according to their directional stability
and/or their utility for orienting.
This latter study highlights an important point. Given
that stable environmental cues are normally the most
useful for orienting (Epstein & Vass, 2014; Galati et al.,
2010), it is unclear what precisely the RSC is processing—
the permanent landmarks themselves or the orienting
relevance of these useful landmarks. In all previous fMRI
études, these two features have been conflated—stable
landmarks were always useful for orienting. This issue
needs to be resolved if we are to truly understand the
mechanisms at play within the RSC and how this contrib-
utes to the formation of environmental representations
that permit navigation.
Auger et al. (2012) had participants rate the navigational
utility of real-world outdoor items, as well as their perma-
nence, size, and visual salience. Fait intéressant, in a factor
analysis they found that this navigational utility feature
loaded on a separate factor than permanence and one that
did not evoke RSC engagement. De plus, there was no
difference between good and poor navigators in their rat-
ings of navigational utility, whereas for ratings of perma-
nence the two groups diverged, as described above. Ce
subjective finding seems to link permanence rather than
navigational utility to the RSC, although clearly more
objective evidence is required.
Par conséquent, here we investigated whether RSC
codes for the absolute permanence of landmarks or if
it is in fact responsive to landmarks that can be used
for localizing targets. We dissociated these features
using a two-by-two factorial design where landmarks
were either permanent or transient and were either rel-
evant or irrelevant for finding a treasure chest. Nous avons utilisé
novel landmarks (from Auger et al., 2015) about which
participants had no prior knowledge. Participants
learned about the landmarks and were then tested
about their landmark knowledge during fMRI scanning.
The nature of our experimental manipulations required
participants to have a broad survey-type overview of the po-
sitions of landmarks and treasure chests. This would be dif-
ficult to achieve in a large-scale environment. Donc, à
enable learning about a sufficient number of landmarks in
the prescan phase, the stimuli were presented in locations
on a computer screen (Chiffre 1). This small-scale context
was qualitatively different from most previous work exam-
ining landmark permanence, where large-scale environ-
ments were used. Cependant, given that RSC permanence
responses were previously first demonstrated in relation
to single isolated objects on a computer screen (Auger
et coll., 2012), albeit real-world items, we predicted the cur-
rent approach would yield responses in RSC. That said, il
would still be an important proof-of-principle to establish
whether representations of landmark permanence or rele-
vance for orienting could be detected in this small-scale set-
ting. This could have an additional benefit, because a
simplified desktop version of tasks previously performed
in complex environments would be more useful for testing
patient populations and perhaps even non-humans. Notre
prediction, based on the limited subjective evidence de-
scribed earlier, was that RSC would be responsive to land-
mark permanence irrespective of orienting value.
Auger and Maguire
699
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
un
_
0
1
2
3
1
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
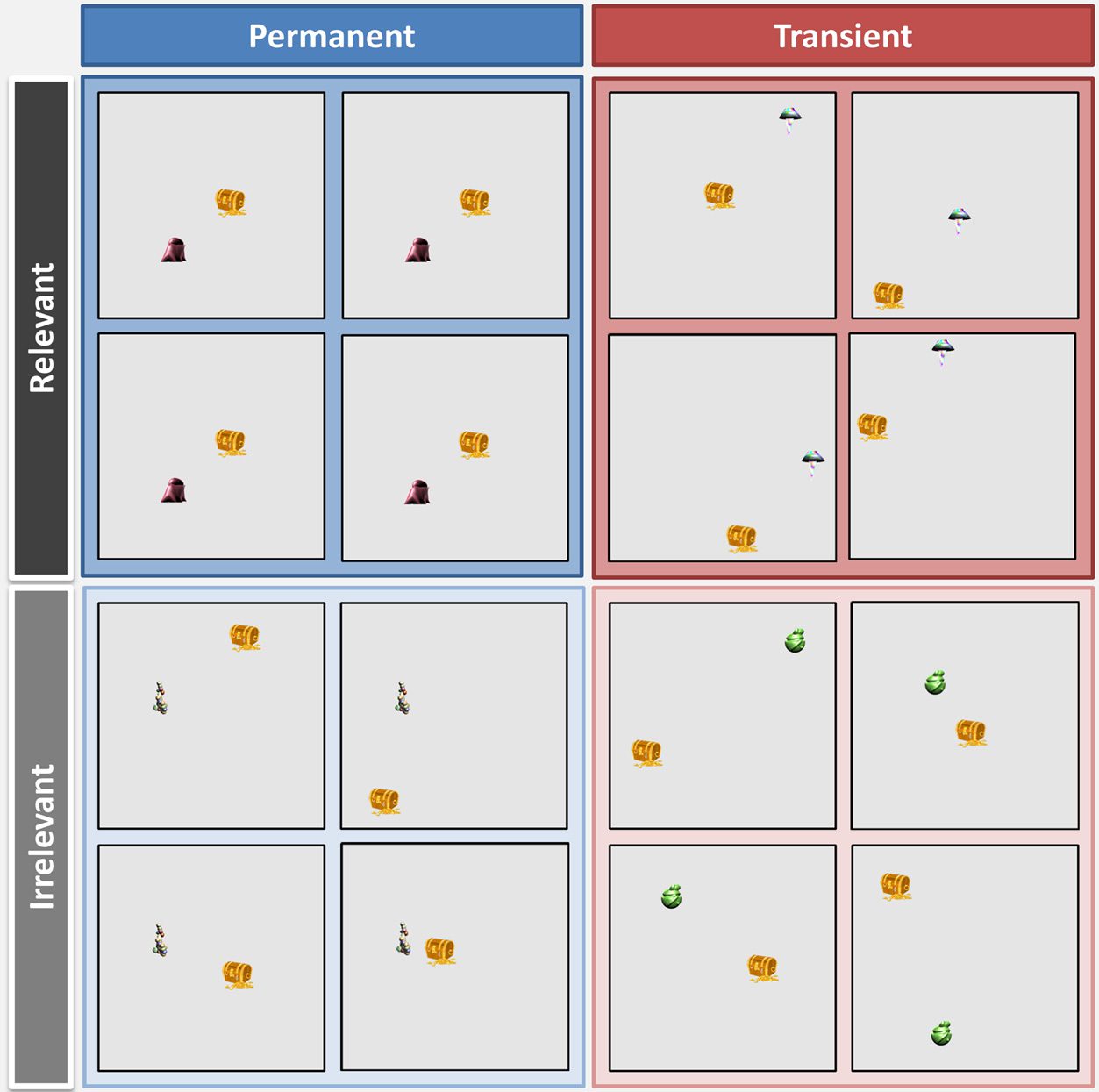
Chiffre 1. The four
experimental conditions.
Landmarks varied in terms of
their permanence and orienting
relevance. For each condition,
four example computer screens
are shown to represent four
different occasions when this
stimulus was presented during
learning. Permanent landmarks
(left/blue) were always
positioned in the exact same
screen location. Transient
landmarks (right/red) appeared
in a different place every time.
Relevant landmarks (top/
darker) could always be used to
locate where a treasure chest
would be, whereas irrelevant
landmarks (bottom/ lighter)
could not be used to locate the
treasure chest.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
un
_
0
1
2
3
1
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
This paradigm allowed us to address two additional
issues. D'abord, the hippocampus has been shown to process
the distance between specific spatial locations (Sherrill
et coll., 2013; Baumann, Chan, & Mattingley, 2012; Morgan,
MacEvoy, Aguirre, & Epstein, 2011; Spiers & Maguire,
2007). RSC has also been implicated in computing dis-
tance to a goal location (Sherrill et al., 2013; Baumann &
Mattingley, 2010; Wolbers & Büchel, 2005). These previ-
ous experiments have generally examined representations
of distance coding while people actively navigate within
large-scale, complex environments. Using the “relevant”
landmarks in our study, namely those which could be used
to locate a treasure chest, meant we could investigate
whether the RSC and/or hippocampus processed the dis-
tance between a relevant landmark and its associated tar-
get treasure location on a much smaller scale—the space
on a computer screen with people merely viewing land-
marks in isolation. The second additional point we could
examine related to individual differences. RSC responses
to landmark permanence have also been linked to a per-
son’s ability to navigate and acquire new spatial informa-
tion (Auger et al., 2012, 2017; Auger & Maguire, 2013).
AThal has also been shown to process permanent land-
marks differently, depending on a person’s spatial abili-
liens (Auger et al., 2012). We therefore also examined
how differences in the amount people learned about
the landmarks related to activity within their RSC and
its interactions with other connected brain regions, surtout-
cially those within the head direction circuit, tel que
AThal and the subiculum of the hippocampus (Shine
et coll., 2016; Yoder et al., 2011).
MÉTHODES
Participants
Thirty-two healthy, right-handed participants took part in
the experiment (16 femmes, âge moyen = 21.5 années, SD =
3.8). All had normal vision and gave written informed
consent in accordance with the approval of the local re-
search ethics committee. None had taken part in previ-
ous experiments involving these stimuli.
Stimuli
In a prescan learning session, participants were shown
numerous images, one at a time, on a computer screen.
Each image contained a single landmark and a treasure
chest (voir la figure 1 for examples). The landmarks came
from the set of unique, novel items, which had been cre-
ated for previous experiments (Auger et al., 2015, 2017)
and of which the participants had no prior experience.
The landmarks and treasure chests were viewed multiple
times and could appear in any 1 de 64 screen locations (dans
un 8 × 8 grid arrangement). Each landmark varied
according to two key features: (1) Permanence—a
landmark either always appeared in the exact same loca-
tion on the screen on every occasion it was presented
700
Journal des neurosciences cognitives
Volume 30, Nombre 5
(“permanent”) or in a different place every time (“tran-
sient”); (2) Relevance—a landmark was either “relevant”
for locating the treasure chest (and always appeared in
the exact same location relative to a treasure chest) ou
“irrelevant” for orienting to the treasure chest (the land-
mark and treasure chest were in completely different rel-
ative locations every time they appeared).
This gave rise to four different types of landmarks
(Chiffre 1): Permanent Relevant (both the landmark and
treasure chest always appeared in the exact same location
on the screen whenever they were seen), Transient Rel-
evant (the landmark and treasure always appeared in dif-
ferent locations on the screen whenever they were seen,
but their location relative to each other was fixed), Per-
manent Irrelevant (the landmark always appeared in the
same location whenever it was seen, but the treasure
chest was in a different place every time), and Transient
Irrelevant (both the landmark and treasure appeared in
constantly changing locations). Il y avait 15 stimuli in
each condition, giving a total of 60 landmarks.
The landmarks in each condition were matched for a
number of other perceptual features based on ratings
provided in a separate study with a different set of partic-
ipants (Auger et al., 2015); these included the following:
salience—to what extent does this item grab your atten-
tion? 1/Not at all to 5/ Very much, F(3, 56) = 0.350, p =
.8; associations with other items—does this remind you
of anything? Yes/No, F(3, 56) = 0.502, p = .7; strength of
association with other items—how strongly does it remind
you of this? 1/Only slightly to 5/ Very much, F(3, 56) =
0.439, p = .7; how likeable the landmark was—how do
you feel about this item? Like/Dislike, F(3, 56) = 0.886,
p = .5; animateness—does this item look like it could
be alive or not? Alive/Not alive, F(3, 56) = 0.414, p =
.7; memorableness—memory of having seen the items
after answering all other questions about them Yes/No,
F(3, 56) = 0.039, p = 1.0. The landmarks were all the
same size, and the locations that appeared on the screen
were matched so that an equal number from each of the
four conditions appeared in all four quarters of the
screen. The locations that treasure chests were positioned
relative to landmarks were also matched, so that an equal
number of treasure chests appeared above/below and
left/right of the four different types of landmark.
The experiment comprised two parts: a learning phase
outside the MRI scanner, followed by a testing phase
while participants underwent fMRI scanning.
Prescan Learning Phase
Before starting the learning phase, participants had the
task explained to them. They were instructed that they
had to view the images of landmarks and treasure chests
and concentrate on learning the two key features for
each landmark, c'est, whether or not it was permanent
(always appearing in the exact same place each time it
was seen) and whether it could be used to find treasure.
They were told that the task inside the MRI scanner
would require them to use landmarks to help find trea-
sure. No indication was given about precisely how their
knowledge of the landmarks would be tested, just that
they needed to focus on learning the two key properties
for each landmark.
The learning phase had 15 learning sessions. In each
session, all of the 60 landmarks were presented (avec un
treasure chest) once for 3.5 sec without any intertrial in-
terval in a different randomized order to the other learn-
ing sessions. At the end of Sessions 2, 4, 6, 8, 10, 12, 14,
et 15, there were “mini-test” periods. On each trial in
these mini-tests, an image of a single landmark was
shown on a gray background for 2 sec in the center of
the screen. Separate screens then immediately asked par-
ticipants to rate the permanence (is this landmark
Permanent/Transient) and relevance (could you use this
landmark to find the treasure Yes/No) of that landmark.
As soon as they gave their response, a screen showed
participants whether it was correct or incorrect for
1 sec before moving to the next trial. Each mini-test
had eight trials, except for the final mini-test at the end
of Session 15, which had four trials. In this way, each of
le 60 landmarks was rated once in the mini-tests. Ce
ensured that exposure to all the landmarks in the learn-
ing phase was identical. The mini-tests served two main
purposes—they ensured participants remained focused
on learning the two key features of each landmark and
allowed us to gauge the amount they had learned through-
out the learning phase.
The number of landmarks, learning sessions, and mini-
tests used were optimized based on a series of pilot ex-
periments to ensure that people could learn and retain
sufficient new information about the permanence and
relevance of the landmarks without rendering them too
fatigued to proceed to the testing phase in the scanner.
fMRI Testing Phase
At the end of the learning phase, participants were pre-
pared for fMRI scanning and had the testing phase task
explained to them. On each trial, they were presented
with an image of a single landmark for 3 sec in the center
of the screen on a gray background (Chiffre 2). Immedi-
ately after viewing this landmark image, they rated the
permanence and relevance of that landmark before mov-
ing on to the next trial. Between trials, il y avait un 2- à
4-sec jittered interval in which a small black cross was
presented in the center of a gray background. Partici-
pants were instructed to fixate on this cross during the
intertrial interval. The order that the participants were
asked to rate the permanence and relevance of landmarks
was randomized to ensure they could not anticipate which
feature they would need to consider first while the land-
mark image was on screen. The way in which the perma-
nence and relevance questions were asked also varied to
keep participants attending carefully; there were three
Auger and Maguire
701
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
un
_
0
1
2
3
1
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
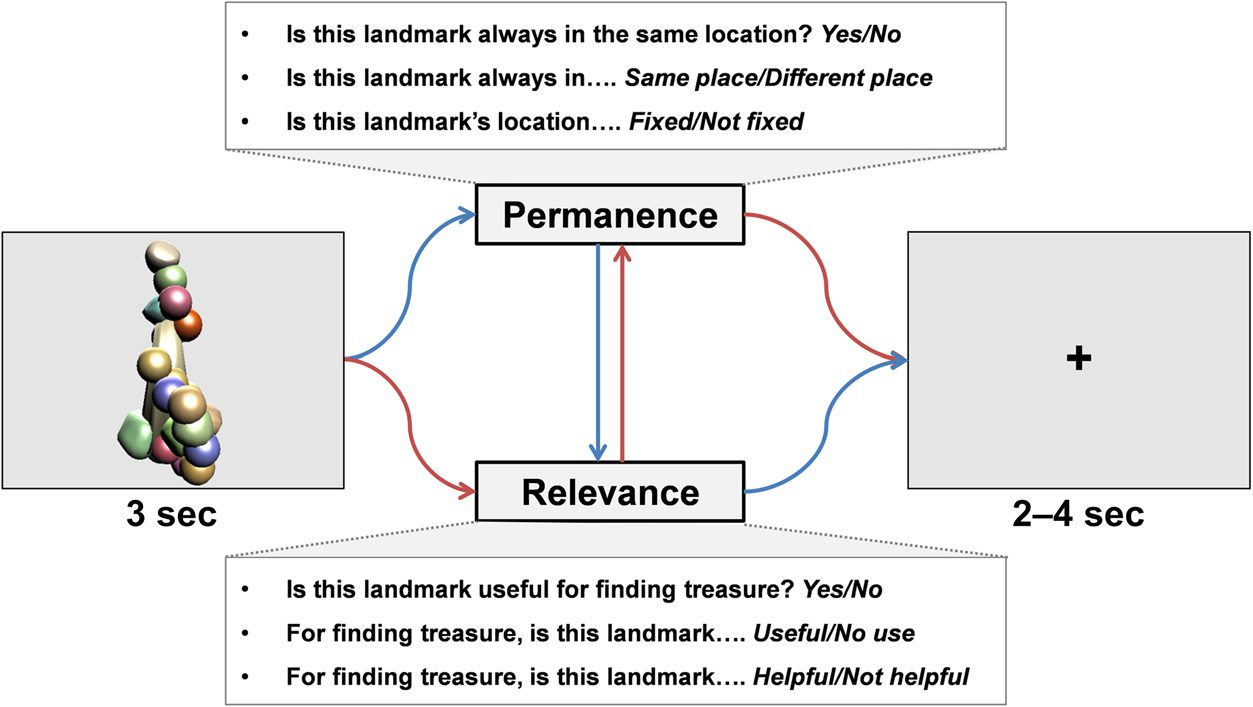
Chiffre 2. The testing phase
during fMRI scanning. An item
appeared on the screen, et
participants were asked about
its permanence and also its
relevance for locating the
treasure chest. The order of
questions was randomized, et
the way in which the
permanence and relevance
questions were asked also
varied. There was then a jittered
2- to 4-sec interval before the
next trial.
varieties for each feature: Permanence—(1) Is this land-
mark always in the same location? Yes/No, (2) Is this land-
mark always in….Same place/Different place, (3) Est-ce
landmark’s location…Fixed/Not fixed; Relevance—(1) Is
this landmark relevant for finding treasure? Yes/No, (2)
For finding treasure, is this landmark…Useful/No use,
(3) For finding treasure, is this landmark…Helpful/Not
helpful.
Participants rated the permanence and relevance of
each landmark on three separate occasions, using each
of the three question variations once, in a randomized
order. This gave a total of 180 trials (60 landmarks each
rated three times), which were split into three scanning
runs of 60 trials each. Within each scanning run, every
landmark was viewed and then rated once, in a random-
ized order.
Scanning Parameters
T2*-weighted echo-planar images with BOLD contrast were
acquired on a 3-T whole-body MRI scanner (Magnetom TIM
Trio, Siemens Healthcare, Erlangen, Allemagne) operated
with the standard radio frequency transmit body coil and
a 32-channel head receive coil. Scanning parameters
were selected to achieve whole-brain coverage but opti-
mized for the hippocampus and surrounding tissue: 48
oblique axial slices angled at −45° from the axial to cor-
onal plane (as defined in Weiskopf, Hutton, Joseph, &
Deichmann, 2006), 2.5 mm thickness (with interslice dis-
tance factor 20%), repetition time = 3.36 sec (slice rep-
etition time = 70 msec), excitation flip angle = 90°, echo
temps (TE) = 30 msec, in-plane resolution = 3 mm ×
3 mm, field of view = 192 mm × 192 mm, 64 × 64 ma-
trix, phase encoding in the anterior–posterior direction,
13% oversampling in the phase encoding direction, echo
spacing = 500 μsec. For reduction of signal loss in the
hippocampal region, slices were angulated, and a z-shim
gradient moment of +0.6 mT/m msec was applied
( Weiskopf et al., 2006). To allow for T1 equilibration
effects, the first six “dummy” volumes from each scan-
ning run were discarded. Field maps were acquired
using a standard manufacturer’s double-echo gradient-
echo field map sequence (short TE = 10 msec, long
LE = 12.46 msec; 64 axial slices with 2 mm thickness
et 1 mm gap yielding whole-brain coverage; in-plane
resolution = 3 mm × 3 mm). A 3-D MDEFT T1-weighted
structural scan (Deichmann, Schwarzbauer, & Tourneur,
2004) was acquired for each participant with 1 mm iso-
tropic resolution.
MRI Data Preprocessing
fMRI data were analyzed with SPM8 (www.fil.ion.ucl.ac.
uk/spm). Images were bias-corrected, realigned, et-
warped (using the field maps), and normalized to a stan-
dard EPI template in MNI space with a resampled voxel
size of 3 × 3 × 3 mm. For all the whole-brain univariate
and connectivity analyses, images were then smoothed
using a Gaussian kernel with FWHM of 8 mm. For multi-
voxel pattern analysis (MVPA), unsmoothed images were
used to facilitate the detection of information present
across patterns of voxels.
Behavioral Analyses
We compared the rates that participants learned land-
mark permanence and orienting value during the prescan
learning phase. This was achieved by performing sepa-
rate linear regression analyses for participants’ learning
of landmark permanence and orienting relevance and
then directly comparing the slopes with a t test. We also
compared the accuracy of participants’ ratings of perma-
nence and orienting relevance in the test phase within
the scanner. In the testing phase task, landmark perma-
nence and orienting relevance was asked in three different
ways (Chiffre 2). The ordering of the permanence/relevance
questions was also randomized. We therefore additionally
analyzed the responses using one-way ANOVAs to deter-
mine whether the way or order in which the permanence
and relevance questions were asked had any impact on the
702
Journal des neurosciences cognitives
Volume 30, Nombre 5
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
un
_
0
1
2
3
1
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
accuracy of responses. A threshold of p < .05 was used
throughout. All statistical analyses were performed using
SPSS version 20 (SPSS, Inc., Chicago, IL).
Whole-brain fMRI Univariate Analyses
To assess fMRI responses in relation to the permanence
and orienting value of landmarks across the whole brain,
we first performed an interaction analysis. The main
effects of each condition (landmark permanence and
orienting relevance) were then analyzed. Namely, we
compared whole-brain BOLD responses for permanent
versus transient, transient versus permanent, relevant
versus irrelevant, and irrelevant versus relevant land-
marks. For each contrast, we created regressors for each
condition of interest and convolved them with the canon-
ical hemodynamic response function.
Each testing phase trial was modeled from the time of
onset of a landmark image for 1.5 sec (the first half of the
landmark image presentation time) given previous find-
ings suggesting that this time frame ensures responses
corresponded to automatic and rapid processing of land-
marks (Auger et al., 2012). Separate participant-specific
movement regressors were treated as covariates of no in-
terest. Subject-specific parameter estimates pertaining to
each regressor of interest (betas) were calculated for each
voxel. Second-level random effects analyses were then run
using one-sample t tests on the parameter estimates
(collapsing across the three scanning runs of the task).
We also performed additional univariate analyses, which
included regressors at the second level relating to each in-
dividual’s accuracy in answering questions about landmark
features. For all contrasts, we report any activation that
survived a whole-brain family-wise error (FWE)-corrected
threshold of p < .05, unless otherwise stated.
The hippocampus has been reported to process the
distance between specific spatial locations (Sherrill
et al., 2013; Baumann et al., 2012; Morgan et al., 2011;
Spiers & Maguire, 2007). Therefore, we also looked for
BOLD responses related to the distance between a land-
mark and its associated treasure location. For every rele-
vant landmark, we calculated the distance between it and
its target treasure location. These values were used to
create parametric regressors for a whole-brain general
linear model fMRI analysis. Specifically, we looked for ac-
tivity that was linearly modulated by a target location,
which was closer or farther away from its associated land-
mark. We report any fMRI activations that survived a
whole-brain FWE-corrected threshold of p < .05, except
for the hippocampus where, given our prior hypotheses
regarding this specific region, we report activations at a
whole-brain uncorrected threshold of p < .001.
Connectivity Analyses
We were also interested in RSC interactions with other
brain regions and how this may relate to how well partic-
ipants had learned information about the landmarks. As
such, for any landmark features to which RSC was
responsive in the whole-brain univariate analyses (a so-
called “feature-of-interest”), we also investigated its inter-
actions with other brain areas and how these interactions
varied depending on how well participants had learned
that feature. Specifically, we used a generalized psycho-
physiological interaction (gPPI) analysis (McLaren, Ries,
Xu, & Johnson, 2012; Friston et al., 1997) to examine
the functional coupling between RSC and the rest of the
brain while people viewed landmarks possessing the par-
ticular feature-of-interest. We then also added the partici-
pants’ accuracy scores for that landmark feature during the
in-scanner testing phase as a second-level covariate of in-
terest. We performed additional gPPI analyses for any
other regions shown to be responsive to landmark per-
manence or relevance from the whole-brain univariate
analysis.
The PPI analyses were performed using the General-
ized Form of Context-Dependent Psychophysiological
Interactions SPM toolbox (McLaren et al., 2012). As seed
regions, we used clusters from the corresponding
whole-brain univariate fMRI contrasts with a specific
focus on RSC (i.e., the clusters within RSC, which were
responsive to a particular feature-of-interest). For all the
gPPI analyses, we report any significant activation that
survived a whole-brain FWE-corrected threshold of p <
.05, unless otherwise stated.
For any functional connectivity identified by the gPPI
analyses, we used dynamic causal modeling (DCM;
Stephan, Penny, Daunizeau, Moran, & Friston, 2009;
Friston, Harrison, & Penny, 2003) to investigate the
nature of the information flow between the regions.
The gPPI analyses specifically indicated which regions in-
creased their interaction in connection with more knowl-
edge of the feature-of-interest. We therefore compared
how the nature of the interaction between the regions
may differ between participants who had learned the in-
formation particularly well and those who had learned
less well. To do this, we created a regressor for the
feature-of-interest for use in a parametric empirical Bayes
(PEB) DCM analysis (Friston et al., 2016).
For each participant, we created a design matrix with
two main regressors of interest: one modeling all land-
marks, to be used as the input for each DCM model
(C matrix), and another for just those with the specific
feature-of-interest (e.g., amount of learning about perma-
nent landmarks), to be used as the models’ modulatory
input (B matrix). Each model assumed the presence of
endogenous self-connections and reciprocal connectivity
between the two brain regions (A matrix) that emerged
from the gPPI analyses. We used DCM12 to fit each
model to the fMRI data and also modeled stochastic fluc-
tuations in the state equations to account for neural
noise, which is particularly relevant for these endoge-
nously driven interactions (Daunizeau, Stephan, &
Friston, 2012).
Auger and Maguire
703
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
We created a full PEB model, with all connections men-
tioned above present (i.e., complete bilateral reciprocal
connectivity between the two brain regions), and a
second-level regressor for the feature-of-interest identi-
fied in the gPPI analysis to act as the modulatory B matrix.
We then compared this full PEB model with two nested
PEB models. These two nested models each removed
one of the two B matrix modulatory inputs so that we could
compare the modulatory effect of Region 1 upon Region 2
and vice versa, in relation to the feature-of-interest
(Figure 5B shows the precise model architectures that were
compared).
It should be noted that gPPI and DCM analyses differ in
the precise data that are modeled. DCM models are fit
on the main task model, whereas gPPI adds a regressor
to the univariate model and is therefore fit on the resid-
uals of the task activity model.
Multivoxel Pattern Analysis
Previous studies have used MVPA as a potentially more
sensitive measure for detecting neural representations
compared with mass univariate analyses (e.g., Bonnici
et al., 2012). We therefore also used MVPA to examine
subject-specific neural representations of the two land-
mark features (Chadwick, Bonnici, & Maguire, 2012;
Haynes & Rees, 2006; Norman, Polyn, Detre, & Haxby,
2006). Separate regressors were created for each of the
180 trials. Participant-specific parameter estimates per-
taining to each trial regressor were then calculated and
used in the MVPA analyses. We selected ROIs to use for
the MVPA analyses from brain areas shown in this and
previous studies (Auger & Maguire, 2013; Auger et al.,
2012) to process permanence and related landmark fea-
tures, namely, RSC, hippocampus, and PHC, as well as
additional brain regions identified in this study’s whole-
brain univariate or PPI analyses. ROIs were defined ana-
tomically for RSC, hippocampus, and PHC using bilateral
masks, which were delineated by an experienced re-
searcher, not involved in this project, on an averaged
structural MRI brain scan from an independent group
of participants (n = 30) and guided by Duvernoy
(1999), Insausti et al. (1998), and Vann et al. (2009).
The other areas were defined functionally from the out-
put of the univariate and gPPI analyses.
MVPA analyses were performed for every subject to
ascertain whether or not it was possible to decode the
type of landmark being viewed based on patterns of acti-
vation in each of the ROIs. All MVPA analyses used a linear
support vector machine (SVM) implemented through
LIBSVM (Chang & Lin, 2011) with fixed regularization hy-
perparameter C = 1. We used a standard cross-validation
procedure throughout (Hsu & Lin, 2002; Duda, Hart, &
Stork, 2001) whereby a single trial is assigned as the test
trial and all others as training trials. In each instance, an
SVM was trained using the training trial data. The SVM
was then presented with the fMRI data from the test trial
to “guess” what type of trial it was (i.e., what type of land-
mark was being viewed while that specific fMRI activation
pattern was elicited). This process was repeated, chang-
ing the test trial each time until all trials had been tested
once. Classifier accuracy is defined as the proportion of
the SVM guesses that were correct.
In line with previous studies (Auger & Maguire, 2013;
Bonnici et al., 2012; Chadwick et al., 2012), we used fea-
ture selection to first identify the voxels within each ROI,
which were most likely to carry relevant information per-
taining to what was being classified (Guyon & Elisseeff,
2003). Only the most informative participant-specific
voxels within each ROI were then used for a separate
final classification to establish a region’s classifier accuracy
value. This process in effect increases the signal-to-noise
ratio. The feature selection used a multivariate searchlight
strategy (Kriegeskorte, Goebel, & Bandettini, 2006). This
aims to determine the information present within a “local
environment” surrounding each voxel within an ROI. We
used a series of classifications (as described above) to
assess the amount of relevant information present within
a sphere (of radius 3 voxels) surrounding each voxel
within the ROIs. Only voxels with responses carrying
the most information were then selected for use in the
final classification.
To avoid any form of circular analysis, we used inde-
pendent data sets for the feature selection and final clas-
sifications (Kriegeskorte, Simmons, Bellgowan, & Baker,
2009). The testing phase consisted of three scanning
runs; we therefore used two runs for feature selection
and the independent data set from the remaining run
for the final classification. This was repeated twice more,
changing the scanning run that was used for the final clas-
sification on each occasion. The classifier accuracy values
from these three repetitions were then averaged to pro-
vide an overall threefold cross-validation. This produced
a single participant-specific classification accuracy value
for each ROI. We then performed t tests on these values
to assess whether or not accuracy across all subjects was
significantly above chance (i.e., t tests were all one tailed).
We first used this MVPA procedure to assess the ROIs’
response patterns associated with representations of the
features with a four-way classification of landmark type
(Permanent Relevant vs. Transient Relevant vs. Perma-
nent Irrelevant vs. Transient Irrelevant; chance = 25%).
Similar to the gPPI connectivity analysis, we then looked
for any relationship between these results and people’s
knowledge about the landmarks. For ROIs implicated in
coding for landmark features by the four-way classifica-
tion, we also performed separate two-way classifications
of permanence and relevance to assess representations
of the two properties independently.
Finally, we investigated whether it was possible to classify
the distance between a relevant landmark and its associated
treasure location. We took a median split of the relevant
landmarks’ distances from their related treasure location
to define “close” and “far” groups. We then performed an
704
Journal of Cognitive Neuroscience
Volume 30, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
MVPA analysis to determine whether the activation patterns
elicited while viewing these landmarks might contain infor-
mation about the proximity of their target location.
RESULTS
Behavioral Data
The participants successfully learned both the perma-
nence and relevance of the landmarks, and there was
no difference in their accuracy scores for the two features
in the final prescan learning phase mini-test (mean per-
manence accuracy = 93.0%, SD = 15.9; mean relevance
accuracy = 86.7%, SD = 16.8; t31 = 1.761, p = .09).
There was also no difference in the rate at which they
learned landmark permanence or relevance (mean differ-
ence between slopes of the linear learning regressors =
0.05, SD = 0.26; t31 = 1.084, p = .3).
Responses made by participants during the in-scanner
testing phase task also indicated that there were no dif-
ferences in how well subjects knew the permanence and
orienting relevance of landmarks (mean permanence ac-
curacy = 90.1%, SD = 10.7; mean relevance accuracy =
87.7%, SD = 12.5; t31 = 0.426, p = .4).
The three different ways in which the questions were
asked for the task inside the scanner also had no impact
on the accuracy of responses for permanence (question
one mean accuracy = 90.6%, SD = 9.8; question two
mean accuracy = 89.2%, SD = 12.2; question three mean
accuracy = 90.4%, SD = 10.6; F(2, 93) = 0.155, p = .9) or
relevance (question one mean accuracy = 87.6%, SD =
13.2; question two mean accuracy = 87.3%, SD = 13.0;
question three mean accuracy = 88.3%, SD = 11.9; F(2,
93) = 0.054, p = .9).
The order of the questions also had no impact on the ac-
curacy of participant responses for either permanence
(mean accuracy if first question = 89.7%, SD = 11.1; mean
accuracy if second question = 90.5%, SD = 10.3; t31 = 0.324,
p = .8) or relevance (mean accuracy if first question =
88.0%, SD = 13.4; mean accuracy if second question =
87.2%, SD = 12.0; t31 = 0.132, p = .9). Overall, therefore,
any differences in fMRI responses could not be attributed
to disparity in the extent that participants knew the two
landmark features or how this information was elicited.
Areas Responding to Properties of the Landmarks
across the Whole Brain
We first performed whole-brain univariate contrasts to
look for regions that were more engaged by permanent
and/or relevant landmarks. An interaction analysis found
no regions where activity was influenced by a combina-
tion of both landmark properties. We then performed
separate analyses to assess the main effects of each indi-
vidual condition.
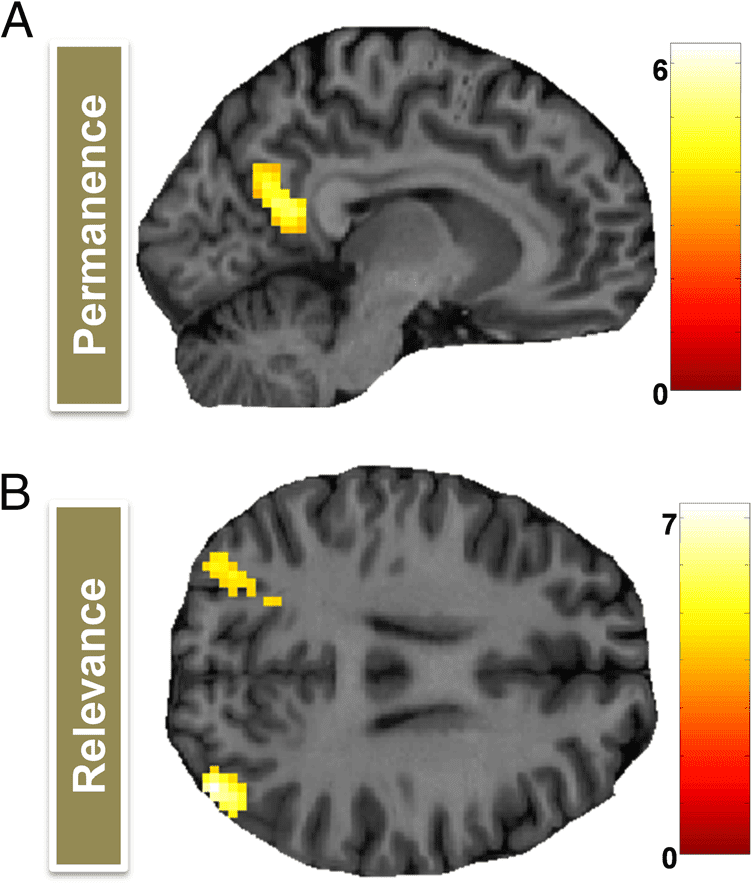
Comparing fMRI responses when participants viewed
permanent and transient landmarks, there was increased
activity for permanent items within the right RSC
(Figure 3A; 15, −52, 19, Z = 5.86; the left RSC was also
active just below threshold), which extended into poste-
rior parts of the POS, as well as additional bilateral clus-
ters in posterior occipital cortex (left: −18, −88, −8, Z =
6.34; right: 18, −91, −2, Z = 6.31). A contrast comparing
landmarks relevant for localizing the treasure chest with
those which were irrelevant produced no significant acti-
vation in RSC, but there were bilateral clusters of activa-
tion in the region of the angular gyrus (Figure 3B; right:
39, −82, 25, Z = 7.26; left: −33, −82, 34, Z = 6.05). No
brain areas were more responsive to transient than per-
manent landmarks or irrelevant than relevant landmarks.
No brain regions were responsive to the accuracy of
answering the questions about landmark features.
For relevant landmarks, we also looked for fMRI
responses related to the distance between them and
their associated treasure location. A larger distance be-
tween a landmark and its target location was associated
with a greater BOLD response in right hippocampus
(Figure 4A; 30, −28, −11, Z = 4.89). No region was
more engaged by landmarks associated with closer
treasure locations.
RSC Interactions with Other Brain Areas and the
Effect of Landmark Knowledge
We then looked for brain areas with which the permanence-
responsive RSC was interacting and how this may be
Figure 3. Brain areas responsive to landmark permanence and
relevance—whole-brain univariate analysis. (A) The RSC and posterior
parts of POS were more engaged by permanent than transient
landmarks. (B) Bilateral clusters in the angular gyrus were more active
when people viewed a relevant than an irrelevant landmark. Activations
are displayed on a sagittal (A) and axial (B) section of a single
representative participant’s structural MRI brain scan. The color bars
indicate each voxel’s associated Z score.
Auger and Maguire
705
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
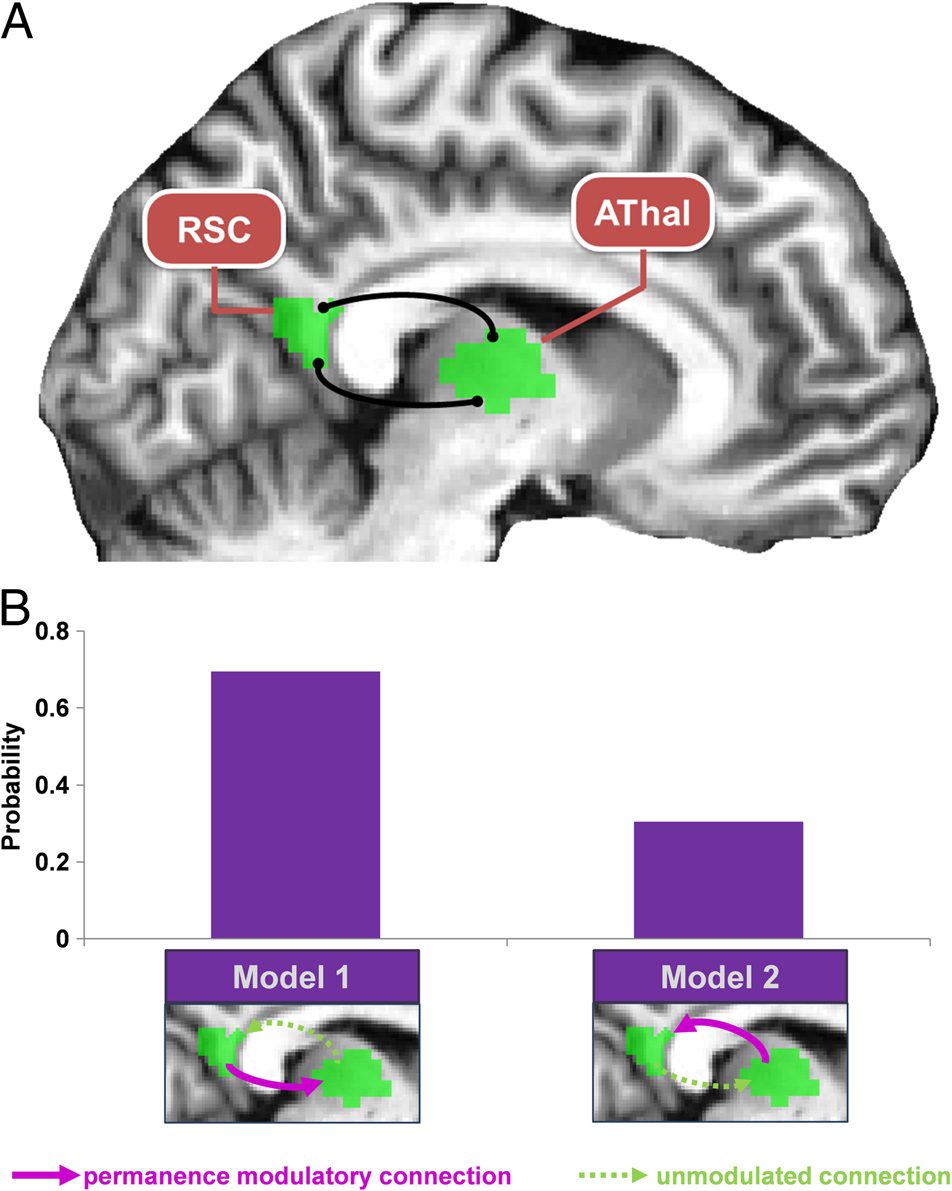
Next, we investigated the nature of the RSC–AThal
interaction using DCM. We compared a full PEB model
of connectivity between the two regions with two nested
PEB models. The first model excluded the connection for
individuals’ permanent landmark learning from AThal to
RSC, and the second excluded the modulatory connec-
tion in the opposite direction. Model 1 therefore repre-
sented the effect of RSC driving responses in AThal
according to how well participants had learned about
permanent landmarks, and Model 2 represented the
effect of AThal driving responses in RSC in line with the
level of permanence learning (see Figure 5B). Model 1
was the clear winner (Model 1 posterior probability =
.695, Model 2 posterior probability = .305). This indicates
that the increased interaction between RSC and AThal re-
vealed by the gPPI analysis likely reflected an increase in
RSC driving responses in AThal, the better people knew
the permanence of landmarks.
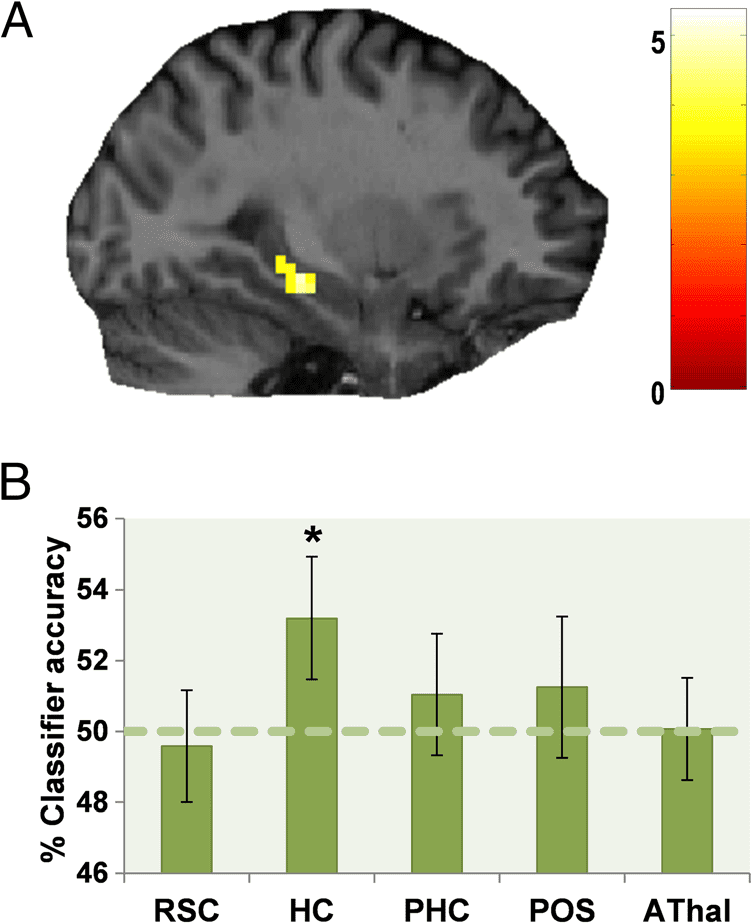
Multivoxel Pattern Analysis
To explore the representations of landmark features in
greater detail, we used MVPA. We first investigated
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. RSC connectivity associated with individual differences in
permanence knowledge. RSC = retrosplenial cortex; AThal = anterior
thalamus. (A) A gPPI analysis showed that, when participants viewed
an image of a permanent landmark, the better they had learned
landmark permanence, then the more their RSC interacted with AThal.
(B) To examine the nature of this RSC–AThal interaction in relation to
learning of landmark permanence, we performed a PEB DCM analysis.
The winning model (Model 1) indicates that RSC drove activity in AThal
in accordance with individuals’ learning of landmark permanence.
Figure 4. Hippocampal processing of distance to the treasure and
orienting relevance. (A) The hippocampus increased its engagement for
landmarks that were associated with a more distant target treasure
location. Activations are displayed on sagittal sections of a single
representative participant’s structural MRI brain scan. The color bar
indicates each voxel’s Z score. (B) Only the hippocampus had patterns
of fMRI activity, which could be used to decode whether a relevant
landmark’s associated treasure was nearby or farther away. The green
dashed line indicates the chance level (50%) for this two-way
classification; error bars show the SEM, and * denotes classifications
that are significantly above chance ( p < .05). RSC = retrosplenial
cortex; HC = hippocampus; PHC = parahippocampal cortex; POS =
parieto-occipital sulcus; AThal = anterior thalamus.
related to how well participants had learned landmark
permanence. An initial gPPI analysis across the whole
participant group found no significant interaction be-
tween regions. However, a further gPPI analysis, which
took into account individuals’ learning of landmark
features, revealed that, when participants viewed perma-
nent landmarks, the better they had learned the land-
mark permanence, then the more their RSC displayed
functional coupling with the AThal (−12, −19, 7, Z =
5.80; Figure 5A). There was also an additional significant
cluster in the cerebellum (the posterior part of the qua-
drangular lobe: −9, −70, −11, Z = 6.59). At a reduced
threshold ( p < .001 whole brain uncorrected), there
was also increased activity in the left hippocampus, in-
cluding the subiculum (−21, −22, −8, Z = 4.34). This
is particularly interesting, given that the RSC, AThal, and
subiculum are known to contain head direction cells in
rodents (Sharp, Blair, & Cho, 2001). Further gPPI anal-
yses were run using the other brain areas shown to be
responsive to landmark permanence or relevance in the
whole-brain univariate analyses (as described above).
No other region displayed any differences in functional
connectivity relating to the amount of information
learned about landmarks.
706
Journal of Cognitive Neuroscience
Volume 30, Number 5
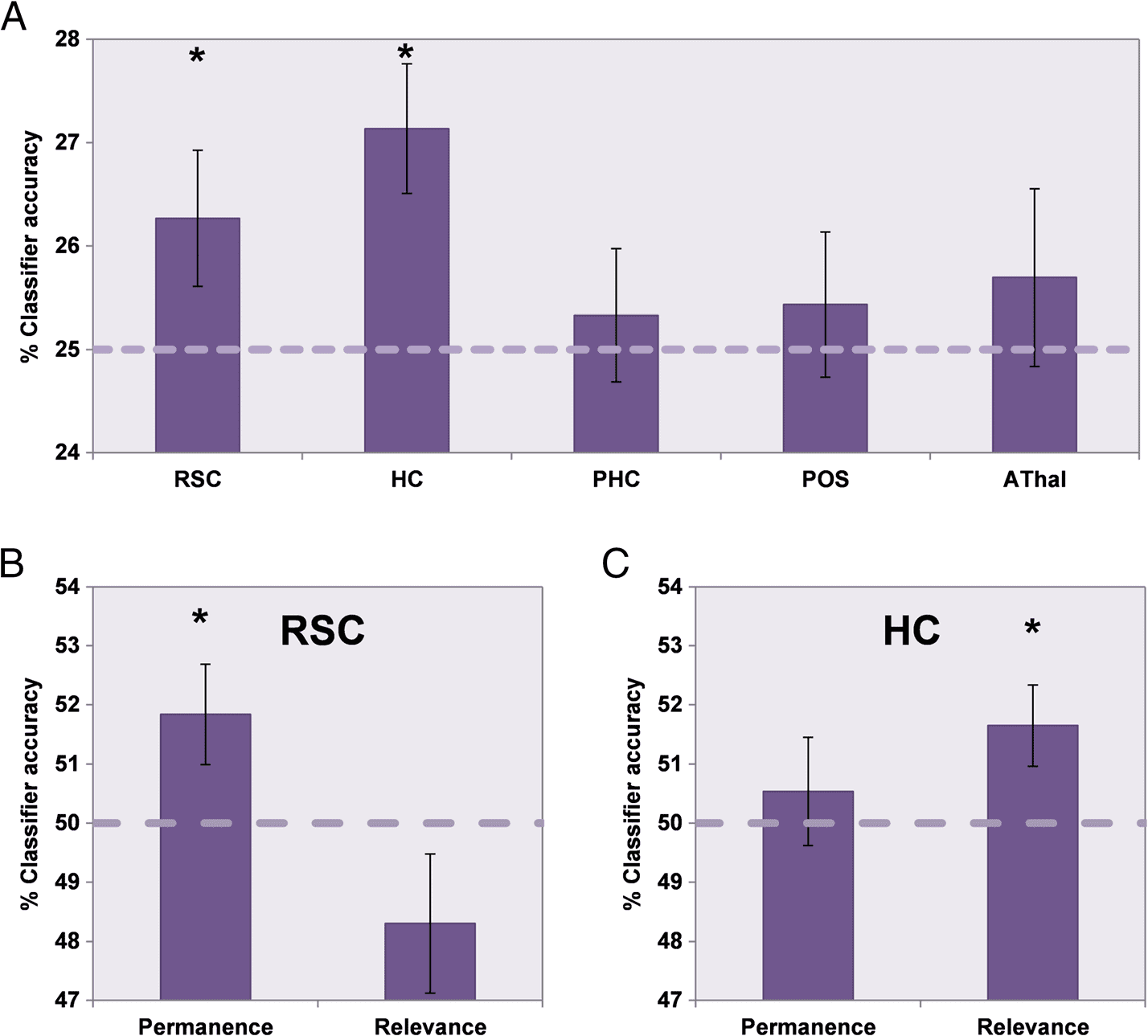
Figure 6. MVPA analysis of
landmark permanence and
relevance. (A) The classification
accuracy for decoding between
the four types of landmark
in each of the ROIs.
RSC = retrosplenial cortex;
HC = hippocampus;
PHC = parahippocampal
cortex; POS = parieto-occipital
sulcus; AThal = anterior
thalamus. Above chance
classification was only possible
for RSC and hippocampus. To
determine which feature each
region was particularly sensitive
to, additional two-way
classifications of landmark
permanence and relevance were
performed in RSC (B) and
hippocampus (C). RSC responses
could be used to classify
landmark permanence but not
relevance, whereas hippocampal
activity could be used to classify
orienting relevance but not
permanence of landmarks.
Dashed lines indicate each
classification’s chance level, error
bars show the SEM, and * denotes
classifications that are significantly
above chance ( p < .05).
whether it was possible to decode which of the four land-
mark types (Permanent Relevant, Transient Relevant, Per-
manent Irrelevant, or Transient Irrelevant) a participant
was viewing based on the multivoxel patterns of activity
in RSC, hippocampus, and PHC (defined anatomically),
as well as the two other regions implicated in this
study—the POS (the parts of the functional cluster re-
sponding more to permanent than transient landmarks
in the univariate analysis, see Figure 3A, excluding the
parts in RSC) and AThal (as found in the PPI analysis;
see Figure 5A). Figure 6A illustrates results of this four-
way MVPA analysis. Landmark type could be classified
above chance (25%) based on responses in RSC (mean
accuracy = 26.3%, SD = 3.7; t31 = 1.924, p = .03) and
hippocampus (mean accuracy = 27.1%, SD = 3.5; t31 =
3.410, p < .001), but not in PHC (mean accuracy =
25.3%, SD = 3.6; t31 = 0.512, p = .3), POS (mean accu-
racy = 25.4%, SD = 4.0; t31 = 0.618, p = .3), or AThal
(mean accuracy = 25.7%, SD = 4.9; t31 = 0.808, p = .2).
To establish whether the significant four-way classifi-
cation in RSC and hippocampus was driven by repre-
sentations of landmark permanence or relevance, we
performed independent two-way classifications for each
feature based on responses in the two regions. A 2 × 2
ANOVA comparing classifier accuracy in RSC and hippo-
campus for permanence and relevance demonstrated a
significant interaction, F(1, 31) = 7.550, p = .01. t tests
demonstrated that, in RSC (Figure 6B), response pat-
terns could be used to classify the permanence (mean
accuracy = 51.8%, SD = 4.8; t31 = 2.168, p = .02) but
not relevance (mean accuracy = 48.3%, SD = 6.7; t31 =
−1.444, p = below chance classification) of landmarks
significantly above chance. The opposite was true of
hippocampus (Figure 6C), where relevance (mean
accuracy = 51.6%, SD = 3.9; t31 = 2.398, p = .01)
but not permanence (mean accuracy = 50.5%, SD =
5.2; t31 = 0.586, p = .3) could be decoded. We also
performed post hoc two-way classifications in PHC,
POS, and AThal for both permanence and orienting
relevance. Unsurprisingly, given the results of the
four-way classification reported above, it was not pos-
sible to classify either feature in any of the regions
with above chance accuracy.
Similar to the univariate analysis of fMRI responses re-
lating to the distance between a relevant landmark and its
associated treasure location, we also examined whether it
was possible to decode this distance using MVPA. The
multivoxel response pattern elicited in hippocampus
while people viewed an image of a landmark could be
used to classify whether or not it was relevant for finding
treasure that was nearby or farther away (Figure 4B;
mean accuracy = 53.2%, SD = 9.8; t31 = 1.852, p =
.04). This was not the case in the other ROIs (RSC mean
accuracy = 49.6%, SD = 8.9; t31 = −0.264, below chance
classification; PHC mean accuracy = 51.0%, SD = 9.7; t31 =
0.605, p = .3; POS mean accuracy = 51.3%, SD = 11.3;
Auger and Maguire
707
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
t31 = 0.626, p = .3; AThal mean accuracy = 50.1%,
SD = 8.2; t31 = 0.048, p = .5).
DISCUSSION
The RSC has been linked with the processing of perma-
nent landmarks in a number of previous studies (Auger
et al., 2012, 2015, 2017; Marchette, Vass, Ryan, & Epstein,
2014; Troiani et al., 2014; Auger & Maguire, 2013). How-
ever, it is unclear whether these RSC responses truly re-
flect representations of a landmark’s permanence or
instead the fact that permanent landmarks tend to be
the most useful cues for orienting. Here we dissociated
these two important landmark properties. When people
viewed permanent landmarks, there was increased activ-
ity in RSC extending posteriorly into the POS. The same
was not true, however, for landmarks that could be used
to locate target treasure chests. We also found that RSC
contained subject-specific patterns of activity associated
with knowledge of permanence, but not relevance, of
landmarks. This was not the case for any other brain re-
gion. In contrast, the orienting relevance of landmarks
was associated with engagement of the angular gyrus
and the hippocampus, with the hippocampus also repre-
senting the distance between a treasure chest and its
associated landmark. These distinct patterns of brain ac-
tivity associated with landmark permanence and orient-
ing relevance could not be attributed to differences in
the rate at which the two features were learned or the
overall amount of information that was acquired. There-
fore, a landmark’s permanence seems to be a primary
feature processed by RSC, with orienting value of land-
marks and their spatial relationships coded by other
regions.
Landmark Permanence
RSC has been implicated in a diverse range of complex
cognitive functions including navigation, scene process-
ing, episodic memory, and imagination of future and fic-
titious events ( Vann et al., 2009; Spreng, Mar, & Kim,
2009; Epstein, 2008). However, there is limited evidence
to indicate what specific role it might contribute to these
processes. We have previously proposed that a key func-
tion of the RSC is to identify permanent stable landmarks,
which might then be used to build environmental repre-
sentations (Auger et al., 2012, 2015, 2017; Auger &
Maguire, 2013). However, the permanent landmarks in
these previous experiments were always inherently more
relevant and reliable cues for orienting. Here, by dissoci-
ating these two properties, we were able to confirm that
RSC does indeed appear to primarily process landmark
permanence independent of any utility for making spatial
judgements. This accords with our previous factor analy-
sis finding that navigational utility loaded onto a separate
factor than permanence and did not evoke RSC engage-
ment (Auger et al., 2012). Moreover, in the same study,
there was no difference between good and poor navigators
in their ratings of navigational utility, whereas poor naviga-
tors were significantly less consistent in rating permanence.
Previous work has demonstrated that experiencing
landmarks from first-person and survey-type perspectives
can give rise to differences in the associated neural re-
sponses (Boccia, Guariglia, Sabatini, & Nemmi, 2016;
Boccia, Nemmi, & Guariglia, 2014; Shelton & Gabrieli,
2002). The current survey-type paradigm is qualitatively
different from previous first-person perspective work,
which related RSC activity to landmark permanence
(Auger et al., 2012, 2015). Nevertheless, we found that
RSC was specifically responsive to the permanence of land-
marks even on this small scale. This suggests that the scope
of RSC permanence processing may be broad, encompass-
ing situations involving items in space in different frames of
reference. Our results also indicate that tasks need not in-
volve complex virtual spaces to study the RSC, which could
be useful for testing patients and perhaps nonhumans.
The dominance of permanence, rather than relevance,
representations in RSC and POS seems to be inconsistent
with the suggestion that a key function of these regions
lies in using landmarks to localize and orientate within
space (Epstein & Vass, 2014). The role RSC plays in these
more complex processes could merely reflect the fact
that they are usually centered upon permanent environ-
mental features. RSC has also been suggested to assist in
translating between and integrating egocentric and allo-
centric spatial information (Sherrill et al., 2013; Sulpizio,
Committeri, Lambrey, Berthoz, & Galati, 2013; Vann
et al., 2009; Byrne, Becker, & Burgess, 2007), but again
this could similarly reflect the reliance of these processes
on manipulating mainly permanent cues.
RSC has also been implicated in processing informa-
tion relating to the distance and direction to a goal loca-
tion (Vedder et al., 2017; Sherrill et al., 2013; Baumann &
Mattingley, 2010; Wolbers & Büchel, 2005). However, this
did not appear to be the case in this study; only the hip-
pocampus showed any sensitivity to the distance be-
tween a landmark and its associated treasure location.
This may relate to our use of small-scale space and not
navigation within an environment or the recent finding
that goal proximity coding switches from hippocampus
to RSC over time (Patai et al., preprint). It could also
be the case that the RSC’s apparent involvement in these
processes in previous studies may in fact have reflected
their reliance upon the use of permanent, stable environ-
mental cues. From the perspective of navigation system
robustness, it may be advantageous that the RSC re-
sponds specifically to permanence, as that is perhaps less
likely to change than orienting relevance.
Orienting Relevance
The hippocampus has been found to be responsive to
permanent landmarks (Auger et al., 2015). This might
seem at odds with the current result where there was
708
Journal of Cognitive Neuroscience
Volume 30, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
no evidence of hippocampal activity relating to landmark
permanence. However, the reason for this may be be-
cause the hippocampus in that previous study was ob-
served to process permanent landmarks when they
were associated with a precise location (Auger et al.,
2015) and not for landmarks that were devoid of specific
spatial connections (Auger & Maguire, 2013; Auger et al.,
2012). This points to the hippocampus perhaps playing a
role in processing the spatial relationships between land-
marks and other parts of the environment, rather than
the permanence of landmarks themselves. This is consis-
tent with the results of this study, where a landmark’s
permanence was dissociable from other spatial relation-
ships and consequently the hippocampus was not
engaged.
Further weight is added to this account of hippocam-
pal function by the fact that the hippocampus was re-
sponsive to the distance between a landmark and its
related treasure. This accords with previous reports of
the hippocampus representing the distances between
items in larger, three-dimensional environments (Sherrill
et al., 2013; Baumann et al., 2012; Morgan et al., 2011;
Spiers & Maguire, 2007). Here we demonstrate that this
also holds true for smaller-scale spatial relationships. But
there may be other explanations for this finding. Hippo-
campal engagement here could merely reflect associative
binding between a landmark and a treasure chest,
although we believe this is unlikely given that the same
binding was involved for treasure that was near and far-
ther away and yet it was possible to differentiate the two
from activity within the hippocampus. Another consider-
ation is the nature of the treasure chest. Although each
trial contained a unique landmark, the treasure chest was
the same across trials, although it had a different conno-
tation in each case, and participants knew this and clearly
performed well at learning orienting relevance. Never-
theless, it could be argued that the distance effects we
noted were related to the amount of pattern separation
(Leutgeb, Leutgeb, Moser, & Moser, 2007) that was re-
quired, with near and farther away treasure chests being
perhaps more easily distinguishable. However, this can-
not explain the main MVPA finding where it was possible
to decode whether landmarks were relevant or not for
orienting. It is worth noting that, although the MVPA
effect sizes were small, it was nevertheless possible to
achieve significant above-chance classification. The mag-
nitude of classifier accuracy depends on various elements
of an experimental paradigm (as described by Chadwick
et al., 2012). Here the requirement for whole-brain cov-
erage precluded the use of high-resolution scanning,
which may have yielded a greater degree of classifiable
information.
We also found the angular gyrus was responsive to the
orienting relevance of landmarks. This brain region has
been posited to play a role in a broad array of cognitive
functions (Seghier, 2013), from reading, comprehension,
and number processing (Arsalidou & Taylor, 2011;
Houdé, Rossi, Lubin, & Joliot, 2010; Price & Mechelli,
2005) to social cognition (Mar, 2011). As such, the angu-
lar gyrus response in this study to landmarks with rel-
evance for orienting could be interpreted in several
ways. Knowledge of a landmark’s relevance for orienting
required linking this semantic concept to its visual attri-
butes, and this type of semantic processing has been con-
sistently attributed to angular gyrus (Binder, Desai,
Graves, & Conant, 2009; Vandenberghe, Price, Wise,
Josephs, & Frackowiak, 1996), particularly for concrete
rather than abstract concepts ( Wang, Conder, Blitzer, &
Shinkareva, 2010). Angular gyrus is also implicated in
shifting attention to stimuli with a particularly salient
value or meaning (Studer, Cen, & Walsh, 2014; Taylor,
Muggleton, Kalla, Walsh, & Eimer, 2011; Gottlieb,
2007), which the relevant landmarks could certainly be
said to have possessed. A third interpretation is that an-
gular gyrus was activated because of its role in conflict
resolution between inputs (Nee, Wager, & Jonides,
2007; Fan, Flombaum, McCandliss, Thomas, & Posner,
2003), perhaps the task of resolving the conflicting per-
manence and relevance information engaged angular
gyrus. This final interpretation is more problematic given
that the level of “conflict” between the landmarks’ prop-
erties would not necessarily be greater among the rele-
vant than nonrelevant landmarks; each landmark had
the same two binary conflicting attributes. That said, it
is difficult to conclusively distinguish whether the angular
gyrus response to landmarks with relevance for orienta-
tion reflected generalized semantic processing, more
specific direction of visuospatial attention, or the resolu-
tion of conflicting landmark properties. In truth, it may
be some combination of the three.
Interindividual Differences in
Landmark Knowledge
Responses in RSC also related to how well people had
learned about landmark permanence. Better learning
about permanence was associated with more differentia-
ble response patterns in RSC. Furthermore, the better
participants had learned which landmarks were perma-
nent, the more their RSC interacted with AThal while
viewing those landmarks.
Previous studies have demonstrated that activity in
RSC is related to a person’s ability to acquire new spatial
information and to navigate (Auger et al., 2012, 2017;
Auger & Maguire, 2013). In each case, variation in these
general spatial abilities was also found to be associated
with specific differences in processing landmark perma-
nence, both behaviorally and in fMRI responses in RSC.
This study adds to this growing body of evidence that per-
formance in some spatial tasks may be directly linked to
RSC permanence representations both of highly familiar
everyday items (Auger & Maguire, 2013; Auger et al.,
2012) as well as while learning new information about pre-
viously unfamiliar landmarks (Auger et al., 2015, 2017).
Auger and Maguire
709
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
RSC does not act in isolation and is connected to a
wide range of other cortical and subcortical brain regions
(Sugar, Witter, van Strien, & Cappaert, 2011; Greicius,
Supekar, Menon, & Dougherty, 2009; Vann et al., 2009;
van Groen & Wyss, 1990, 1992, 2003). This is particularly
true of AThal, where RSC shares dense reciprocal con-
nectivity with a number of different nuclei ( Wright,
Erichsen, Vann, O’Mara, & Aggleton, 2010; van Groen
& Wyss, 2003). These connections are not just structural.
The two regions influence one another’s processing of
space and together help support navigation ( Jankowski
et al., 2013; Clark, Bassett, Wang, & Taube, 2010), espe-
cially when it requires the use of environmental land-
marks ( Yoder et al., 2011). However, despite the large
amount of evidence of mutual interaction between RSC
and AThal in rodents, there are far fewer examples of
similar communication in humans, only evidence of sim-
ilar directional coding within both regions (Shine et al.,
2016). RSC and anterodorsal parts of the thalamus were
previously shown to be more active in good compared
with poor navigators when viewing permanent landmarks
(Auger et al., 2012). The current study builds upon this
finding using a more explicit measure of people’s under-
standing of landmark permanence. This revealed that,
rather than simply being coactive, RSC actually drove
activity in AThal, and as this directional coupling increased,
the better participants had learned which landmarks were
permanent.
RSC and AThal both contain neurons that display tun-
ing to the direction an animal’s head is facing ( Vann
et al., 2009; Taube, 1995; Chen et al., 1994). It is there-
fore interesting that we also observed increased RSC in-
teraction with a third region known to contain head
direction cells, the subiculum, albeit at a lower statistical
threshold (Taube et al., 1990). Therefore, these three re-
gions, which have consistently been shown to be densely
interconnected and functionally related in rodents, were
here, in humans, interacting in accordance with the ex-
tent of a person’s knowledge about permanent land-
marks. That said, it could be argued that, given the
survey-type paradigm we used here, head direction cells
would be unlikely to contribute to performance on this
task. Consequently, some other mechanism that is not
centered on head direction cells might underpin the per-
manence response. One way to examine this in the
future would be to directly compare permanent land-
mark representations in small- and large-scale space. If
the magnitude of RSC permanence responses is similar,
this might suggest that head direction cells are not of
prime relevance in this case.
It is also interesting to note that the subiculum is
directly implicated along with RSC in the spread of pa-
thology in Alzheimer’s dementia (George et al., 2014).
This provides further evidence that the disorientation
usually present in the early stages of the disease could
be a consequence of aberrant processing in RSC, one
of the first regions to show pathological changes in this
condition (Tu et al., 2015; Pengas et al., 2012; Pengas,
Hodges, Watson, & Nestor, 2010). The profound disorien-
tation commonly associated with lesions that involve the
RSC (Vann et al., 2009; Maguire, 2001) could also be ex-
plained by the failure to identify permanent features, with
deleterious effects on environmental representations.
Overall, therefore, we conclude that the coding of per-
manent landmarks in RSC may drive processing in regions
like AThal and subiculum, with possible implications for
the efficacy of functions such as spatial navigation.
Acknowledgments
E. A. M. is funded by a Wellcome Principal Research Fellowship
(101759/Z/13/Z) and by a Wellcome Centre Award (203147/Z/
16/Z). S. D. A. was funded by UCLH/UCL, who received a propor-
tion of funding from the Department of Health’s NIHR Biomedical
Research Centres funding scheme. We thank the Imaging Support
and Cognitive Interface teams for technical assistance.
Reprint requests should be sent to Eleanor A. Maguire, Wellcome
Centre for Human Neuroimaging, Institute of Neurology, Uni-
versity College London, 12 Queen Square, London WC1N
3BG, United Kingdom, or via e-mail: e.maguire@ucl.ac.uk.
REFERENCES
Aggleton, J. P. (2010). Understanding retrosplenial amnesia:
Insights from animal studies. Neuropsychologia, 48,
2328–2338.
Alexander, A. S., & Nitz, D. A. (2017). Spatially periodic
activation patterns of retrosplenial cortex encode route
sub-spaces and distance traveled. Current Biology, 27,
1551–1560.e4.
Arsalidou, M., & Taylor, M. J. (2011). Is 2 + 2 = 4? Meta-analyses
of brain areas needed for numbers and calculations.
Neuroimage, 54, 2382–2393.
Auger, S. D., & Maguire, E. A. (2013). Assessing the mechanism
of response in the retrosplenial cortex of good and poor
navigators. Cortex, 49, 2904–2913.
Auger, S. D., Mullally, S. L., & Maguire, E. A. (2012).
Retrosplenial cortex codes for permanent landmarks. PLoS
One, 7, e43620.
Auger, S. D., Zeidman, P., & Maguire, E. A. (2015). A central role
for the retrosplenial cortex in de novo environmental
learning. eLife, 4, e09031.
Auger, S. D., Zeidman, P., & Maguire, E. A. (2017). Efficacy of
navigation may be influenced by retrosplenial cortex-
mediated learning of landmark stability. Neuropsychologia,
104, 102–112.
Baumann, O., Chan, E., & Mattingley, J. B. (2010). Dissociable
neural circuits for encoding and retrieval of object locations
during active navigation in humans. Neuroimage, 49,
2816–2825.
Baumann, O., Chan, E., & Mattingley, J. B. (2012). Distinct
neural networks underlie encoding of categorical versus
coordinate spatial relations during active navigation.
Neuroimage, 60, 1630–1637.
Baumann, O., & Mattingley, J. B. (2010). Medial parietal cortex
encodes perceived heading direction in humans. Journal of
Neuroscience, 30, 12897–12901.
Baumann, O., & Mattingley, J. B. (2013). Dissociable
representations of environmental size and complexity in the
human hippocampus. Journal of Neuroscience, 33,
10526–10533.
710
Journal of Cognitive Neuroscience
Volume 30, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Bicanski, A., & Burgess, N. (2016). Environmental anchoring of
head direction in a computational model of retrosplenial
cortex. Journal of Neuroscience, 36, 11601–11618.
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and meta-
analysis of 120 functional neuroimaging studies. Cerebral
Cortex, 19, 2767–2796.
Boccia, M., Guariglia, C, Sabatini, U., & Nemmi, F. (2016).
Navigating toward a novel environment from a route or
survey perspective: Neural correlates and context-dependent
connectivity. Brain Structure and Function, 221, 2005–2021.
Boccia, M., Nemmi, F., & Guariglia, C. (2014). Neuropsychology
of environmental navigation in humans: Review and meta-
analysis of fMRI studies in healthy participants.
Neuropsychology Review, 24, 236–251.
Bonnici, H. M., Kumaran, D., Chadwick, M. J., Weiskopf, N., Hassabis,
D., & Maguire, E. A. (2012). Decoding representations of scenes
in the medial temporal lobes. Hippocampus, 22, 1143–1153.
Burnett, G., Smith, D., & May, A. (2001). Supporting the
navigation task: Characteristics of “good” landmarks. In
Contemporary Ergonomics 2001: Proceedings of the Annual
Conference of the Ergonomics Society (pp. 441–446).
London: Taylor and Francis.
Byrne, P., Becker, S., & Burgess, N. (2007). Remembering the
past and imagining the future: A neural model of spatial
memory and imagery. Psychological Review, 114, 340–375.
Chadwick, M. J., Bonnici, H. M., & Maguire, E. A. (2012).
Decoding information in the human hippocampus: A user’s
guide. Neuropsychologia, 50, 3107–3121.
Chang, C.-C., & Lin, C.-J. (2011). LIBSVM: A library for support
vector machines. ACM Transactions on Intelligent Systems
and Technology, 2, 1–39.
Chen, L. L., Lin, L. H., Green, E. J., Barnes, C. A., & McNaughton,
B. L. (1994). Head-direction cells in the rat posterior cortex.
I. Anatomical distribution and behavioral modulation.
Experimental Brain Research, 101, 8–23.
Cho, J. W., & Sharp, P. E. (2001). Head direction, place, and
movement correlates for cells in the rat retrosplenial cortex.
Behavioral Neuroscience, 115, 3–25.
Chrastil, E. R., Sherrill, K. R., Aselcioglu, I., Hasselmo, M. E., & Stern,
C. E. (2017). Individual differences in human path integration
abilities correlate with gray matter volume in retrosplenial cortex,
hippocampus, and medial prefrontal cortex. eNeuro, 4,
ENEURO.0346-16.2017.
Clark, B. J., Bassett, J. P., Wang, S. S., & Taube, J. S. (2010).
Impaired head direction cell representation in the
anterodorsal thalamus after lesions of the retrosplenial
cortex. Journal of Neuroscience, 30, 5289–5302.
Committeri, G., Galati, G., Paradis, A. L., Pizzamiglio, L., Berthoz,
A., & LeBihan, D. (2004). Reference frames for spatial
cognition: Different brain areas are involved in viewer-, object-,
and landmark-centered judgments about object location.
Journal of Cognitive Neuroscience, 16, 1517–1535.
Daunizeau, J., Stephan, K. E., & Friston, K. J. (2012). Stochastic
dynamic causal modelling of fMRI data: Should we care about
neural noise? Neuroimage, 62, 464–481.
Deichmann, R., Schwarzbauer, C., & Turner, R. (2004).
Optimisation of the 3D MDEFT sequence for anatomical
brain imaging: Technical implications at 1.5 and 3 T.
Neuroimage, 21, 757–767.
Duda, R. O., Hart, P. E., & Stork, D. G. (2001). Pattern
classification. New York: Wiley.
Duvernoy, H. (1999). The human brain: Surface, three-
dimensional sectional anatomy with MRI, and blood
supply. New York: Springer.
Epstein, R. A. (2008). Parahippocampal and retrosplenial
contributions to human spatial navigation. Trends in
Cognitive Sciences, 12, 388–396.
Epstein, R. A., & Vass, L. K. (2014). Neural systems for landmark-
based wayfinding in humans. Philosophical Transactions of
the Royal Society of London, Series B: Biological Sciences,
369, 20120533.
Fan, J., Flombaum, J. I., McCandliss, B. D., Thomas, K. M., &
Posner, M. I. (2003). Cognitive and brain consequences of
conflict. Neuroimage, 18, 42–57.
Friston, K. J., Buechel, C., Fink, G. R., Morris, J., Rolls, E. T., &
Dolan, R. J. (1997). Psychophysiological and modulatory
interactions in neuroimaging. Neuroimage, 6, 218–229.
Friston, K. J., Harrison, L., & Penny, W. D. (2003). Dynamic
causal modelling. Neuroimage, 19, 1273–1302.
Friston, K. J., Litvak, V., Oswal, A., Razi, A., Stephan, K. E., van
Wijk, B. C. M., et al. (2016). Bayesian model reduction and
empirical Bayes for group (DCM) studies. Neuroimage, 128,
413–431.
Galati, G., Pelle, G., Berthoz, A., & Committeri, G. (2010). Multiple
reference frames used by the human brain for spatial perception
and memory. Experimental Brain Research, 206, 109–120.
George, S., Rönnbäck, A., Gouras, G. K., Petit, G. H.,
Grueninger, F., Winblad, B., et al. (2014). Lesion of the
subiculum reduces the spread of amyloid beta pathology to
interconnected brain regions in a mouse model of
Alzheimer’s disease. Acta Neuropathologica
Communications, 2, 17.
Gottlieb, J. (2007). From thought to action: The parietal cortex
as a bridge between perception, action, and cognition.
Neuron, 53, 9–16.
Greicius, M. D., Supekar, K., Menon, V., & Dougherty, R. F.
(2009). Resting-state functional connectivity reflects
structural connectivity in the default mode network. Cerebral
Cortex, 19, 72–78.
Guyon, I., & Elisseeff, A. (2003). An introduction to variable and
feature selection. Journal of Machine Learning Research, 3,
1157–1182.
Haynes, J.-D., & Rees, G. (2006). Neuroimaging: Decoding
mental states from brain activity in humans. Nature Reviews
Neuroscience, 7, 523–534.
Houdé, O., Rossi, S., Lubin, A., & Joliot, M. (2010). Mapping
numerical processing, reading, and executive functions in the
developing brain: An fMRI meta-analysis of 52 studies
including 842 children. Developmental Science, 13, 876–885.
Hsu, C.-W., & Lin, C.-J. (2002). A comparison of methods for
multiclass support vector machines. IEEE Transactions on
Neural Networks, 13, 415–425.
Iaria, G., Chen, J.-K., Guariglia, C., Ptito, A., & Petrides, M.
(2007). Retrosplenial and hippocampal brain regions in
human navigation: Complementary functional contributions
to the formation and use of cognitive maps. European
Journal of Neuroscience, 25, 890–899.
Insausti, R., Juottonen, K., Soininen, H., Insausti, A. M.,
Partanen, K., Vainio, P., et al. (1998). MR volumetric analysis
of the human entorhinal, perirhinal, and temporopolar
cortices. American Journal of Neuroradiology, 19, 659–671.
Jacob, P.-Y., Casali, G., Spieser, L., Page, H., Overington, D., &
Jeffery, K. (2017). An independent, landmark-dominated
head-direction signal in dysgranular retrosplenial cortex.
Nature Neuroscience, 20, 173–175.
Jankowski, M. M., Ronnqvist, K. C., Tsanov, M., Vann, S. D.,
Wright, N. F., Erichsen, J. T., et al. (2013). The anterior thalamus
provides a subcortical circuit supporting memory and spatial
navigation. Frontiers in Systems Neuroscience, 7, 45.
Janzen, G., & van Turennout, M. (2004). Selective neural
representation of objects relevant for navigation. Nature
Neuroscience, 7, 673–677.
Konkle, T., & Oliva, A. (2012). A real-world size organization of
object responses in occipitotemporal cortex. Neuron, 74,
1114–1124.
Auger and Maguire
711
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Kriegeskorte, N., Goebel, R., & Bandettini, P. (2006).
Information-based functional brain mapping. Proceedings of
the National Academy of Sciences, U.S.A., 103, 3863–3868.
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S. F., & Baker,
C. I. (2009). Circular analysis in systems neuroscience: The
dangers of double dipping. Nature Neuroscience, 12,
535–540.
Leutgeb, J. K., Leutgeb, S., Moser, M.-B., & Moser, E. I. (2007).
Pattern separation in the dentate gyrus and CA3 of the
hippocampus. Science, 315, 961–966.
Lew, A. R. (2011). Looking beyond the boundaries: Time to put
landmarks back on the cognitive map? Psychological
Bulletin, 137, 484–507.
Lynch, K. (1960). The image of the city (Vol. 8). Cambridge, MA:
Technology Press.
Maguire, E. A. (2001). The retrosplenial contribution to human
navigation: A review of lesion and neuroimaging findings.
Scandinavian Journal of Psychology, 42, 225–238.
Mao, D., Kandler, S., McNaughton, B. L., & Bonin, V. (2017).
Sparse orthogonal population representation of spatial context
in the retrosplenial cortex. Nature Communications, 8, 243.
Mar, R. A. (2011). The neural bases of social cognition and story
comprehension. Annual Review of Psychology, 62, 103–134.
Marchette, S. A., Vass, L. K., Ryan, J., & Epstein, R. A. (2014).
Anchoring the neural compass: Coding of local spatial
reference frames in human medial parietal lobe. Nature
Neuroscience, 17, 1598–1606.
Marchette, S. A., Vass, L. K., Ryan, J., & Epstein, R. A. (2015).
Outside looking in: Landmark generalization in the human
navigational system. Journal of Neuroscience, 35,
14896–14908.
McLaren, D. G., Ries, M. L., Xu, G., & Johnson, S. C. (2012). A
generalized form of context-dependent psychophysiological
interactions (gPPI): A comparison to standard approaches.
Neuroimage, 61, 1277–1286.
Morgan, L. K., MacEvoy, S. P., Aguirre, G. K., & Epstein, R. A.
(2011). Distances between real-world locations are
represented in the human hippocampus. Journal of
Neuroscience, 31, 1238–1245.
Nee, D. E., Wager, T. D., & Jonides, J. (2007). Interference
resolution: Insights from a meta-analysis of neuroimaging
tasks. Cognitive, Affective, & Behavioral Neuroscience, 7,
1–17.
Norman, K. A., Polyn, S. M., Detre, G. J., & Haxby, J. V. (2006).
Beyond mind-reading: Multi-voxel pattern analysis of fMRI
data. Trends in Cognitive Sciences, 10, 424–430.
Patai, E. Z., Javadi, A.-H., Ozubko, J. D., O’Callaghan, A., Ji, S.,
Robin, J., et al. (preprint). Long-term consolidation switches
goal proximity coding from hippocampus to retrosplenial
cortex. bioRxiv, 167882.
Pengas, G., Hodges, J. R., Watson, P., & Nestor, P. J. (2010).
Focal posterior cingulate atrophy in incipient Alzheimer’s
disease. Neurobiology of Aging, 31, 25–33.
Pengas, G., Williams, G. B., Acosta-Cabronero, J., Ash, T. W. J.,
Hong, Y. T., Izquierdo-Garcia, D., et al. (2012). The
relationship of topographical memory performance to
regional neurodegeneration in Alzheimer’s disease. Frontiers
in Aging Neuroscience, 4, 17.
Price, C. J., & Mechelli, A. (2005). Reading and reading
disturbance. Current Opinion in Neurobiology, 15, 231–238.
Seghier, M. L. (2013). The angular gyrus: Multiple functions and
multiple subdivisions. The Neuroscientist, 19, 43–61.
Sharp, P. E., Blair, H. T., & Cho, J. (2001). The anatomical and
computational basis of the rat head-direction cell signal.
Trends in Neurosciences, 24, 289–294.
Shelton, A. L., & Gabrieli, J. D. (2002). Neural correlates of
encoding space from route and survey perspectives. Journal
of Neuroscience, 22, 2711–2717.
Sherrill, K. R., Erdem, U. M., Ross, R. S., Brown, T. I., Hasselmo,
M. E., & Stern, C. E. (2013). Hippocampus and retrosplenial
cortex combine path integration signals for successful
navigation. Journal of Neuroscience, 33, 19304–19313.
Shine, J. P., Valdés-Herrera, J. P., Hegarty, M., & Wolbers, T.
(2016). The human retrosplenial cortex and thalamus code
head direction in a global reference frame. Journal of
Neuroscience, 36, 6371–6381.
Siegel, A., & White, S. (1975). The development of spatial
representations of large-scale environments. In H. W. Reese
(Ed.), Advances in child development and behaviour.
New York: Academic Press.
Spiers, H. J., & Maguire, E. A. (2006). Thoughts, behaviour, and
brain dynamics during navigation in the real world.
Neuroimage, 31, 1826–1840.
Spiers, H. J., & Maguire, E. A. (2007). A navigational guidance
system in the human brain. Hippocampus, 17, 618–626.
Spreng, R. N., Mar, R. A., & Kim, A. S. (2009). The common
neural basis of autobiographical memory, prospection,
navigation, theory of mind, and the default mode: A
quantitative meta-analysis. Journal of Cognitive
Neuroscience, 21, 489–510.
Stephan, K. E., Penny, W. D., Daunizeau, J., Moran, R. J., &
Friston, K. J. (2009). Bayesian model selection for group
studies. Neuroimage, 46, 1004–1017.
Studer, B., Cen, D., & Walsh, V. (2014). The angular gyrus and
visuospatial attention in decision-making under risk.
Neuroimage, 103, 75–80.
Sugar, J., Witter, M. P., van Strien, N. M., & Cappaert, N. L. M.
(2011). The retrosplenial cortex: Intrinsic connectivity and
connections with the (para)hippocampal region in the
rat. An interactive connectome. Frontiers in
Neuroinformatics, 5, 7.
Sulpizio, V., Committeri, G., Lambrey, S., Berthoz, A., & Galati,
G. (2013). Selective role of lingual/parahippocampal gyrus
and retrosplenial complex in spatial memory across
viewpoint changes relative to the environmental reference
frame. Behavioural Brain Research, 242, 62–75.
Taube, J. S. (1995). Head direction cells recorded in the anterior
thalamic nuclei of freely moving rats. Journal of
Neuroscience, 15, 70–86.
Taube, J. S., & Muller, R. U. (1998). Comparisons of head
direction cell activity in the postsubiculum and anterior
thalamus of freely moving rats. Hippocampus, 8, 87–108.
Taube, J. S., Muller, R. U., & Ranck, J. B. (1990). Head-direction
cells recorded from the postsubiculum in freely moving rats.
I. Description and quantitative analysis. Journal of
Neuroscience, 10, 420–435.
Taylor, P. C. J., Muggleton, N. G., Kalla, R., Walsh, V., & Eimer,
M. (2011). TMS of the right angular gyrus modulates priming
of pop-out in visual search: Combined TMS-ERP evidence.
Journal of Neurophysiology, 106, 3001–3009.
Troiani, V., Stigliani, A., Smith, M. E., & Epstein, R. A. (2014).
Multiple object properties drive scene-selective regions.
Cerebral Cortex, 24, 883–897.
Tu, S., Wong, S., Hodges, J. R., Irish, M., Piguet, O., &
Hornberger, M. (2015). Lost in spatial translation – A novel
tool to objectively assess spatial disorientation in Alzheimer’s
disease and frontotemporal dementia. Cortex, 67, 83–94.
van Groen, T., & Wyss, J. M. (1990). Connections of the
retrosplenial granular a cortex in the rat. Journal of
Comparative Neurology, 300, 593–606.
van Groen, T., & Wyss, J. M. (1992). Connections of the
retrosplenial dysgranular cortex in the rat. Journal of
Comparative Neurology, 315, 200–216.
van Groen, T., & Wyss, J. M. (2003). Connections of the
retrosplenial granular b cortex in the rat. Journal of
Comparative Neurology, 463, 249–263.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
712
Journal of Cognitive Neuroscience
Volume 30, Number 5
Vandenberghe, R., Price, C., Wise, R., Josephs, O., & Frackowiak,
R. S. J. (1996). Functional anatomy of a common semantic
system for words and pictures. Nature, 383, 254–256.
Vann, S. D., Aggleton, J. P., & Maguire, E. A. (2009). What does
the retrosplenial cortex do? Nature Reviews Neuroscience,
10, 792–802.
Vedder, L. C., Miller, A. M. P., Harrison, M. B., & Smith, D. M.
(2017). Retrosplenial cortical neurons encode navigational
cues, trajectories and reward locations during goal directed
navigation. Cerebral Cortex, 27, 3713–3723.
Wang, J., Conder, J. A., Blitzer, D. N., & Shinkareva, S. V. (2010).
Neural representation of abstract and concrete concepts: A
meta-analysis of neuroimaging studies. Human Brain
Mapping, 31, 1459–1468.
Weiskopf, N., Hutton, C., Josephs, O., & Deichmann, R. (2006).
Optimal EPI parameters for reduction of susceptibility-
induced BOLD sensitivity losses: A whole-brain analysis at 3 T
and 1.5 T. Neuroimage, 33, 493–504.
Wolbers, T., & Büchel, C. (2005). Dissociable retrosplenial and
hippocampal contributions to successful formation of survey
representations. Journal of Neuroscience, 25, 3333–3340.
Wolbers, T., Weiller, C., & Büchel, C. (2004). Neural
foundations of emerging route knowledge in complex spatial
environments. Cognitive Brain Research, 21, 401–411.
Wright, N. F., Erichsen, J. T., Vann, S. D., O’Mara, S. M., &
Aggleton, J. P. (2010). Parallel but separate inputs from limbic
cortices to the mammillary bodies and anterior thalamic
nuclei in the rat. Journal of Comparative Neurology, 518,
2334–2354.
Yoder, R. M., Clark, B. J., & Taube, J. S. (2011). Origins of
landmark encoding in the brain. Trends in Neurosciences,
34, 561–571.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
9
8
1
7
8
7
4
3
3
/
j
o
c
n
_
a
_
0
1
2
3
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Auger and Maguire
713