Communicated by Patricia Churchland
The Brain Binds Entities and Events by
Multiregional Activation from Convergence Zones
Antonio R. Damasio
Department of Neurology,
Division of Behavioral Neurology and Cognitive Neuroscience,
University of Iowa College of Medicine, Iowa City, IA, Etats-Unis
The experience of reality, in both perception and recall, is spatially
and temporally coherent and “in-register.” Features are bound in enti-
liens, and entities are bound in events. The properties of these entities
and events, cependant, are represented in many different regions of the
brain that are widely separated. The degree of neural parcellation is
even greater when we consider that the perception of most entities and
events also requires a motor interaction on the part of the perceiver
(such as eye movements and hand movements) and often includes a
recordable modification of the perceiver’s somatic state. The question
of how the brain achieves integration starting with the bits and pieces
it has to work with, is the binding problem. Here we propose a new
solution for this problem, at the level of neural systems that integrate
functional regions of the telencephalon.
1 Introduction
Data from cognitive psychology, neurophysiology, and neuroanatomy in-
dicate unequivocally that the properties of objects and events that we
perceive through various sensory channels engage geographically sepa-
rate sensory regions of the brain (Posner 1980; Van Essen and Maunsell
1983; Damasio 1985; Livingstone and Hubel 1988). The need to “bind”
together the fragmentary representations of visual information has been
noted by Treisman and Gelade (19801, Crick (1984), et d'autres, but clearly
the problem is a much broader one and includes the need to integrate
both the sensory and motor components in both perception and recall,
at all scales and at all levels. This broader concept of binding is closer to
that of Sejnowski (1986).
The traditional and by now untenable solution to the binding problem
has been that the components provided by different sensory portals end
up being displayed together in so-called multimodal cortices, where the
most detailed and integrated representations of reality are achieved. Ce
intuitively reasonable view suggests that perception depends on a unidi-
rectional process which provides a gradual refinement of signal extraction
Neural Computation 1, 123-132 (1989) @ 1989 Massachusetts Institute of Technology
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
e
d
toi
n
e
c
o
un
r
t
je
c
e
–
p
d
/
je
F
/
/
/
/
/
1
1
1
2
3
8
1
1
8
0
7
n
e
c
o
1
9
8
9
1
1
1
2
3
p
d
.
.
.
.
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
124
Antonio R. Damasio
along a cascade aimed towards integrative cortices in anterior temporal
and anterior frontal regions. Some of the most influential accounts for the
neural basis of cognition in the post-war period as well as major discov-
eries of neurophysiology and neuroanatomy over the past two decades,
have seemed compatible with this view. After all, anatomical projections
do radiate from primary sensory cortices toward structures in the hip-
pocampus and prefrontal cortices via a multi-stage sequence (Pandya and
Kuypers 1969; Jones and Powell 1970; Nauta 1971; Van Hoesen 1982), et
the farther away neurons are from primary sensory cortices, the larger
their receptive fields become, and the less unimodal their responses are
(Desimone and Ungerleider 1989). Cependant, there are several lines of
evidence on the basis of which this traditional solution can be rejected.
2 Experimental Evidence
Evidence from Experimental Neuroanatomy: The notion that integration of
perceptual or recalled components depends on a single neural meeting
ground calls for the identification of a neuroanatomical site that would
receive projections from all neural regions involved in the processing of
entities and events as they occur in experience. Despite considerable ex-
ploration no such region has yet been found. The anterior temporal cor-
tices and the hippocampus do receive projections from multiple sensory
domaines, but not from motor regions (Van Hoesen 1982). The anterior frontal
cortices, the most frequently mentioned candidates for ultimate integra-
tion, are even less suited for that role. The sensory and motor streams
that reach them remain segregated in different regions (Goldman-Rakic
1988). Autrement dit, there seems to be no structural foundation to sup-
port the intuition that temporal and spatial integration occur at a single
site.
Advances in experimental neuroanatomy have added a new element
to neuroanatomical reasoning about this problem: at every stage of the
chain of forward cortical projections, there exist prominent projections
back to the originating sites. De plus, the systems are just as rich in
multi-stage, reciprocating feedback projections as they are in feedforward
projections (Van Hoesen 1982; Van Essen 1985; Livingstone and Hubel
1987). The neuroanatomical networks revealed by these studies allow for
both forward convergence of some parallel processing streams, et pour
the flow of signaling back to points of origin. In the proposal we will
describe below, such networks operate as coherent phase-locked loops in
which patterns of neural activity in “higher” areas can trigger, enhance,
or suppress patterns of activity in “lower” domaines.
Evidence from Experimental Neuropsychology in Humans with Focal Cere-
bral Lesions: If temporal and frontal integrative cortices were the substrate
for the integration of neural activity on which binding depends, the bi-
lateral destruction of those cortices in humans should: (un) preclude the
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
e
d
toi
n
e
c
o
un
r
t
je
c
e
–
p
d
/
je
F
/
/
/
/
/
1
1
1
2
3
8
1
1
8
0
7
n
e
c
o
1
9
8
9
1
1
1
2
3
p
d
.
.
.
.
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The Brain Binds Entities and Events by Multiregional Activation
125
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
-Frontal
“integrative” cortices-
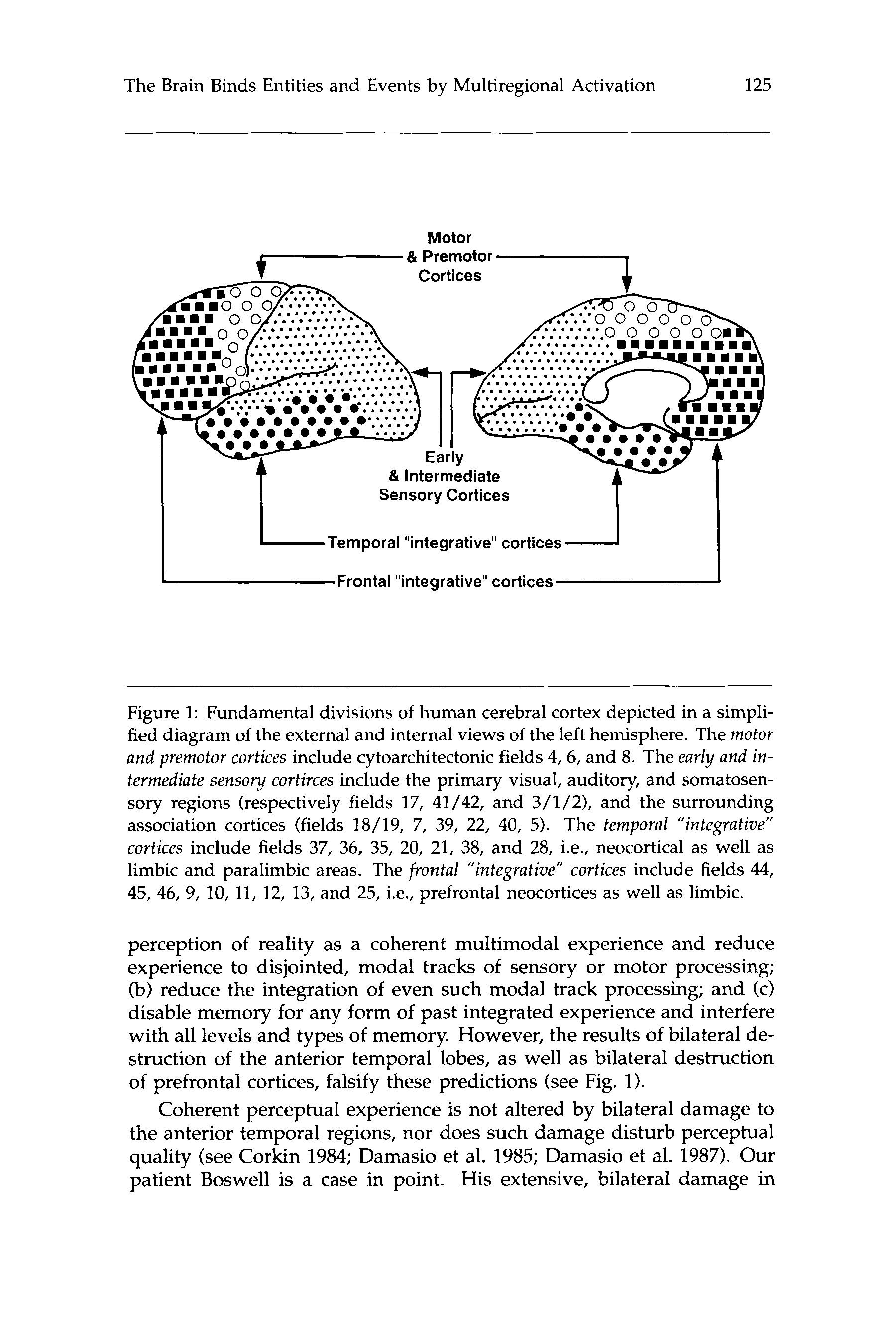
Chiffre 1: Fundamental divisions of human cerebral cortex depicted in a simpli-
fied diagram of the external and internal views of the left hemisphere. The motor
and premotor cortices include cytoarchitectonic fields 4,6, et 8. The early and in-
termediate sensory cortirces include the primary visual, auditory, and somatosen-
sory regions (respectively fields 17, 41 /42, et 3/1/2), and the surrounding
association cortices (fields 18/19, 7, 39, 22, 40, 5). The temporal “integrative”
cortices include fields 37, 36, 35, 20, 21, 38, et 28, c'est à dire., neocortical as well as
limbic and paralimbic areas. The frontal “integrative” cortices include fields 44,
45, 46, 9, 10, 11, 12, 13, et 25, c'est à dire., prefrontal neocortices as well as limbic.
perception of reality as a coherent multimodal experience and reduce
experience to disjointed, modal tracks of sensory or motor processing;
(b) reduce the integration of even such modal track processing; et (c)
disable memory for any form of past integrated experience and interfere
with all levels and types of memory. Cependant, the results of bilateral de-
struction of the anterior temporal lobes, as well as bilateral destruction
of prefrontal cortices, falsify these predictions (see Fig. 1).
Coherent perceptual experience is not altered by bilateral damage to
the anterior temporal regions, nor does such damage disturb perceptual
qualité (see Corkin 1984; Damasio et al. 1985; Damasio et al. 1987). Notre
patient Boswell is a case in point. His extensive, bilateral damage in
/
e
d
toi
n
e
c
o
un
r
t
je
c
e
–
p
d
/
je
F
/
/
/
/
/
1
1
1
2
3
8
1
1
8
0
7
n
e
c
o
1
9
8
9
1
1
1
2
3
p
d
.
.
.
.
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
126
Antonio R. Damasio
anterior temporal cortices and hippocampus, disables his memory for
unique autobiographical events, but not his ability to perceive the world
around in fully integrated fashion and to recall and recognize the entities
and events that he encounters or participates in, at non-unique level.
His binding ability breaks down at the level of unique events, when the
integration of extremely complex combinatorial arrangement of entities
is required.
Bilateral lesions in prefrontal cortices, especially when restricted to
the orbitofrontal sector, are also compatible with normal perception and
even with normal memory for most entities and events except for those
that pertain to the domain of social knowledge (Eslinger and Damasio
1985; Damasio and Tranel 1988).
Enfin, it is damage to certain sectors of sensory association cortices
that can affect both the quality of some aspects of perception within the
modality served by those cortices, and recognition and recall. Depend-
ing on precisely which region of visual cortex is affected, lesions in early
visual association cortices can disrupt perception of shape, or color, ou
texture, or stereopsis, or spatial placement of the physical components
of a stimulus (Damasio 1985; Damasio et al. 1989). A patient may lose
the ability to perceive color and yet perceive shape, depth and motion
normally. More importantly, damage within some sectors of modal as-
sociation cortices can disturb recall and recognition of stimuli presented
through that modality, even when basic perceptual processing is not com-
promised. Par exemple, patients may become unable to recognize famil-
iar faces that they perceive flawlessly (bien que, intriguingly, they can
discriminate familiar from unfamiliar faces at covert level; Tranel and
Damasio 1985; 1988).
The key point is that damage in a posterior and unimodal association
cortex can disrupt recall and recognition at virtually every level of the
binding chain, from the entity-categorical level to the event-unique level.
It can preclude the kind of integrated experience usually attributed to
the anterior cortices.
3 A New View on the Binding Problem
The evidence then indicates: (un) that substantial binding, relative to en-
tities or parts thereof, occurs in unimodal cortices and can support recall
and recognition at the level of categories; (b) that recall and recognition at
category level, are generally not impaired by damage confined to anterior
integrative cortices, c'est à dire., knowledge recalled at categoric levels depends
largely on posterior sensory cortices and interconnected motor cortices;
(c) that recall and recognition of knowledge at the level of unique enti-
ties or events, requires both anterior and posterior sensory cortices, c'est à dire.,
a more complex network is needed to map uniqueness; anterior integra-
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
e
d
toi
n
e
c
o
un
r
t
je
c
e
–
p
d
/
je
F
/
/
/
/
/
1
1
1
2
3
8
1
1
8
0
7
n
e
c
o
1
9
8
9
1
1
1
2
3
p
d
.
.
.
.
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The Brain Binds Entities and Events by Multiregional Activation
127
tive structures alone are not sufficient to record and reconstruct unique
connaissance.
The implication is that the early and intermediate posterior sensory
cortices contain fragmentary records of featural components which can
be reactivated, on the basis of appropriate combinatorial arrangements
(by fragmentary featural components we mean “parts of entities,” at a
multiplicity of scales, most notably at feature level, par exemple., color, mouvement,
texture, shape and parts thereof). They also contain records of the combi-
natorial arrangement of features that defined entities (”local” or ”entity”
binding), but do not contain records of the spatial and temporal relation-
ships assumed by varied entities within an event (“non-local” or “event
binding”). The latter records, the complex combinatorial codes needed
for event recall, are inscribed in anterior cortices. In this perspective the
posterior cortices contain the fragments with which any experience of en-
tities or events can potentially be re-enacted, but only contain the binding
mechanism to re-enact knowledge relative to entities. Posterior cortices
require binding mechanisms in anterior structures in order to guide the
pattern of multiregional activations necessary to reconstitute an event.
Thus posterior cortices contain both basic fragments and local binding
records and are essential for recreating any past experience. Anterior
cortices contain non-local or event-binding records and are only crucial
for reconstitution of contextually more complex events.
Perhaps the most important distinction between this perspective and
the traditional view, is that higher-order anterior cortices are seen as
repositories of combinatorial codes for inscriptions that lie elsewhere
and can be reconstructed elsewhere, rather than being the storage site
for the more refined ”multimodal” representations of experiences. Al-
though anterior cortices receive multimodal projections we conceptualize
the records they harbor as amodal.
If parts of the representation of an entity are distributed over distant
regions of the brain, then mechanisms must be available to bind together
the fragments. A proposal for a new solution to the binding problem
(Damasio 1989) is illustrated in figure 2 and presented in outline as fol-
lows:
1. The neural activity prompted by perceiving the various physical
properties of any entity, occurs in fragmented fashion and in geo-
graphically separate regions located in early sensory cortices and in
motor cortices. So-called ”integrative” cortices do not contain such
fragmentary inscriptions.
2. The integration of multiple aspects of external and internal reality in
perceptual or recalled experiences, depends on the phase-locked co-
activation of geographically separate sites of neural activity within
the above mentioned sensory and motor cortices, rather than on
a transfer and spatial integration of different representations to-
wards anterior higher-order cortices. Consciousness of those co-
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
e
d
toi
n
e
c
o
un
r
t
je
c
e
–
p
d
/
je
F
/
/
/
/
/
1
1
1
2
3
8
1
1
8
0
7
n
e
c
o
1
9
8
9
1
1
1
2
3
p
d
.
.
.
.
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
128
Antonio R. Damasio
activations depends on their being attended to, c'est à dire., on simultane-
ous enhancement of a pertinent set of activity against background
activité.
3. The patterns of neural activity that correspond to distinct physical
properties of entities are recorded in the same neural ensembles
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
e
d
toi
n
e
c
o
un
r
t
je
c
e
–
p
d
/
je
F
/
/
/
/
/
1
1
1
2
3
8
1
1
8
0
7
n
e
c
o
1
9
8
9
1
1
1
2
3
p
d
.
.
.
.
.
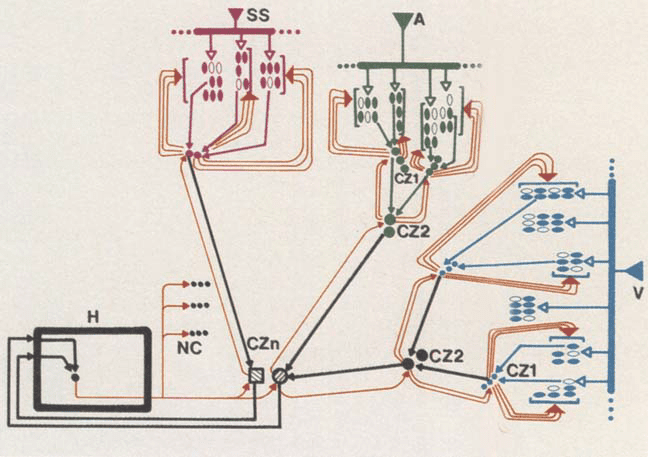
Chiffre 2: Simplified diagram of some aspects of the proposed neural architec-
ture. V, SS, and A depict early and intermediate sensory cortices in visual,
somatosensory, and auditory modalities. In each of those sensory sectors, sep-
arate functional regions are represented by open and filled dots. Note feed-
forward projections (black lines) from those regions toward several orders of
convergence zones (CZ1, CZ2, CZn), and note also feedback projections from
each CZ level toward originating regions (red lines). H depicts hippocampal
système, one of the structures where signals related to a large number of activ-
ity sites can converge. Note outputs of H toward last station of feedforward
convergence zones (CZn) and toward noncortical neural stations (Caroline du Nord) in basal
forebrain, brain stem, and neurotransmitter nuclei. Feedforward and feedback
pathways should not be seen as rigid channels. They are conceived as facili-
tated lines which become active when concurrent firing in early cortices or CZs
takes place. En outre, those pathways terminate over neuron ensembles, dans
distributed fashion, rather than on specific single neurons.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The Brain Binds Entities and Events by Multiregional Activation
129
in which they occur during perception, but the combinatorial ar-
rangements (binding codes) that describe their pertinent linkages
in entities and in events (their spatial and temporal coincidences),
are stored in separate neural ensembles called convergence zones.
4. Convergence zones trigger and synchronize neural activity patterns
corresponding to topographically organized fragment representa-
tions of physical structure, that were pertinently associated in ex-
expérience, on the basis of similarity, spatial placement, temporal se-
quence, or temporal coincidence, or combinations thereof. The trig-
gering and synchronization depends on feedback projections from
the convergence zone to multiple cortical regions where fragment
records can be activated.
5. Convergence zones are located throughout the telencephalon, à
multiple neural levels, in association cortices of different orders,
limbic cortices and subcortical limbic nuclei, and non-limbic sub-
cortical nuclei such as the basal ganglia.
6. The geographic location of convergence zones for different entities
varies among individuals but is not random. It is constrained by
the subject matter of the recorded material (its domain), and by
contextual complexity of events (the number of component entities
that interact in an event and the relations they adopt), and by the
anatomical design of the system. Convergence zones that bind fea-
tures into entities are located earlier in the processing streams, et
convergence zones that bind entities into progressively more com-
plex events are gradually placed more anteriorly in the processing
streams.
7. The representations inscribed in the above architecture, both those
that preserve topographic/topologic relationships and those that
code for combinatorial arrangements, are committed to populations
of neuron ensembles and their synapses, in distributed form.
8. The co-occurrence of activities in multiple sites that is necessary
for binding conjunctions, is achieved by recurrent feedback interac-
tion.
Ainsi, we propose that the processing does not proceed in a single
direction but rather through temporally coherent phase-locking amongst
multiple regions. Although the convergence zones that realize the more
encompassing integration are placed more anteriorly, it is activity in the
more posterior cortical regions that is more directly related to conscious
expérience.
By means of feedback, convergence zones repeatedly return process-
ing to earlier cortices where activity can proceed again towards the same
or other convergence zones. Integration takes place when activations oc-
cur within the same time window, in earlier cortices. There is no need
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
e
d
toi
n
e
c
o
un
r
t
je
c
e
–
p
d
/
je
F
/
/
/
/
/
1
1
1
2
3
8
1
1
8
0
7
n
e
c
o
1
9
8
9
1
1
1
2
3
p
d
.
.
.
.
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
130
Antonio R. Damasio
to postulate a “final” and single integration area. This model accommo-
dates the segregation of neural processing streams that neuroanatomical
and neurophysiological data continue to reveal so consistently, and is
compatible with the increase in receptive fields of neurons that occurs
in cerebral cortex, in the posterior-anterior direction. It accords with the
proposal that fewer and fewer neurons placed anteriorly in the system
are projected on by structures upstream and thus subtend a broader com-
pass of feed-forwarding regions. Broad receptive field neurons serve as
pivots for reciprocating feedback projections rather than as accumulators
of the knowledge inscribed at earlier levels. They are intermediaries in
a continuous process that centers on early cortices.
4 Conclusions
The problem of how the brain copes with the fragmentary representations
of information is central to our understanding of brain function. It is not
enough for the brain to analyze the world into its components parts:
the brain must bind together those parts that make whole entities and
events, both for recognition and recall. Consciousness must necessarily
be based on the mechanisms that perform the binding. The hypothesis
suggested here is that the binding occurs in multiple regions that are
linked together through activation zones; that these regions communi-
cate through feedback pathways to earlier stages of cortical processing
where the parts are represented; and that the neural correlates of con-
sciousness should be sought in the phase-locked signals that are used to
communicate between these activation zones.
Several questions are raised by this new view. Par exemple, what is
the precise nature of the feedback signals that provide temporally coher-
ent phase-locking among multiple regions? How large are the conver-
gence zones in different parts of the brain? How are the decisions made
to store an aspect of experience in a particular zone? There are several
possible approaches to test the hypothesis proposed here. One approach
is to develop new techniques for recording from many neurons simul-
taneously in communicating brain regions. Another relies on neuropsy-
chological experiments in neurological patients with small focal lesions in
key areas of putative networks dedicated to specific cognitive processes.
Enfin, modeling studies should illuminate the collective properties of
convergence zones and provide us with the intuition we need to sharpen
our questions.
Remerciements
Supported by NINCDS Grant PO1 NS19632.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
e
d
toi
n
e
c
o
un
r
t
je
c
e
–
p
d
/
je
F
/
/
/
/
/
1
1
1
2
3
8
1
1
8
0
7
n
e
c
o
1
9
8
9
1
1
1
2
3
p
d
.
.
.
.
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The Brain Binds Entities and Events by Multiregional Activation
131
Les références
Corkin, S. 1984. Lasting Consequences of Bilateral Medial Temporal Lobectomy:
Clinical Course and Experimental Findings in HM. Seminars in Neurology 4,
249-259.
Crick, F. 1984. Function of the Thalamic Reticular Complex: The Searchlight
Hypothesis. Proc. Natl. Acad. Sci. Etats-Unis 81, 4586-4590.
Damasio, UN. 1989. Multiregional Retroactivation: A Systems Level Model for
Some Neural Substrates of Cognition. Cognition, in press.
. 1985. Disorders of Complex Visual Processing. Dans: Principles of Be-
havioral Neurology, éd. M.M. Mesulam, Contemporary Neurology Series,
259-288. Philadelphia: F.A. Davis.
Damasio, UN., P.. Eslinger, H. Damasio, G.W. Van Hoesen, et S. Cornell. 1985.
Multimodal Amnesic Syndrome Following Bilateral Temporal and Basal
Forebrain Damage. Archives of Neurology 42, 252-259.
Damasio, UN., H. Damasio, D. Tranel, K. Welsh, and J. Brandt. 1987. Additional
Neural and Cognitive Evidence in Patient DRB. Society for Neuroscience 13,
1452.
Damasio, UN. and D. Tranel. 1988. Domain-specific Amnesia for Social Knowl-
bord. Société des neurosciences 14, 1289.
Damasio, A.R., H. Damasio, and D. Tranel. 1989. Impairments of Visual Recog-
nition as Clues to the Processes of Memory. Dans: Signal and Sense: Local and
Global Order in Perceptual Maps, éd.. G. Edelman, E. Gall, and M. Cowan,
Neuroscience Institute Monograph. Wiley and Sons.
Désimone, R.. and L. Ungerleider. 1989. Neural Mechanisms of Visual Process-
ing in Monkeys. Dans: Handbook of Neuropsychology, Disorders of Visual
Processing, éd. UN. Damasio, in press.
Eslinger, P.. and A. Damasio. 1985. Severe Disturbance of Higher Cognition
after Bilateral Frontal Lobe Ablation. Neurologie 35, 1731-1741.
Goldman-Rakic, P.S. 1988. Topography of Cognition: Parallel Distributed Net-
works in Primate Association Cortex. Dans: Annual Review of Neuroscience
11, Annual Reviews Inc., Palo Alto, Californie, 137-156.
Jones, E.G. and T.P.S. Powell. 1970. An Anatomical Study of Converging Sen-
sory Pathways within the Cerebral Cortex of the Monkey. Cerveau 93,793420.
Livingstone, M.. and D. Hubel. 1988. Segregation of Form, Color, Movement,
and Depth Anatomy, Physiology, and Perception. Science 240, 740-749.
. 1987. Connections between Layer 4B of Area 17 and Thick Cytochrome
Oxidase Stripes of Area 18 in the Squirrel Monkey. Journal of Neuroscience 7,
3371-3377.
Nauta, W.J.H. 1971. The Problem of the Frontal Lobe: A Reinterpretation. J.. Psy-
chiat. Res. 8, 167-187.
Pandya, D.N. and H.G.J.M. Kuypers. 1969. Cortico-cortical Connections in the
Rhesus Monkey. Brain Res. 13, 13-36.
Posner, M.I. 1980. Orienting of Attention. Quarterly Journal of Experimental
Psychologie 32, 3-25.
Sejnowski, T.J. 1986. Open Questions about Computation in Cerebral Cortex. Dans:
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
e
d
toi
n
e
c
o
un
r
t
je
c
e
–
p
d
/
je
F
/
/
/
/
/
1
1
1
2
3
8
1
1
8
0
7
n
e
c
o
1
9
8
9
1
1
1
2
3
p
d
.
.
.
.
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
132
Antonio R. Damasio
Parallel Distributed Processing, éd.. J.L. McClelland and D.E. Rummelhart,
372-389. Cambridge: MIT Press.
Tranel, D. et un. Damasio. 1988. Nonconscious Face Recognition in Patients
with Face Agnosia. Behavioral Brain Research 30, 235-249.
. 1985. Knowledge without Awareness: An Autonomic Index of Facial
Recognition by Prosopagnosics. Science 22821, 1453-1454.
Treisman, UN. and G. Gelade. 1980. A Feature-integration Theory of Attention.
Cognitive Neuropsychology 12, 97-1 36.
Van Essen, D.C. 1985. Functional Organization of Primate Visual Cortex. Dans:
Cortex cérébral, éd.. UN. Peters and E.G. Jones, 259-329. Plenum Publishing.
Van Essen, D.C. and J.H.R. Maunsell. 1983. Hierarchical Organization and
Functional Streams in the Visual Cortex. Trends in Neuroscience 6, 370-375.
Van Hoesen, G.W. 1982. The Primate Parahippocampal Gyrus: New Insights
Regarding its Cortical Connections. Trends in Neurosciences 5, 345-350.
Reçu 18 Novembre; accepted 25 Novembre 1988.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
e
d
toi
n
e
c
o
un
r
t
je
c
e
–
p
d
/
je
F
/
/
/
/
/
1
1
1
2
3
8
1
1
8
0
7
n
e
c
o
1
9
8
9
1
1
1
2
3
p
d
.
.
.
.
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3