Approaching the Bad and Avoiding the Good: Lateral
Prefrontal Cortical Asymmetry Distinguishes
between Action and Valence
Elliot T. Berkman and Matthew D. Lieberman
Abstrait
■ Goal pursuit in humans sometimes involves approaching un-
pleasant and avoiding pleasant stimuli, such as when a dieter
chooses to eat vegetables (although he does not like them) dans-
stead of doughnuts (which he greatly prefers). Previous neuro-
science investigations have established a left–right prefrontal
asymmetry between approaching pleasant and avoiding unpleas-
ant stimuli, but these investigations typically do not untangle the
roles of action motivation (approach vs. avoidance) and stimulus
valence (pleasant vs. unpleasant) in this asymmetry. En plus,
studies on asymmetry have been conducted almost exclusively
using electroencephalography and have been difficult to replicate
using functional magnetic resonance imaging (IRMf). The present
fMRI study uses a novel goal pursuit task that separates action
motivation from stimulus valence and a region-of-interest analy-
sis approach to address these limitations. Results suggest that
prefrontal asymmetry is associated with action motivation and
not with stimulus valence. Spécifiquement, there was increased left
(vs. droite) activation in dorsolateral prefrontal cortex during ap-
proach (vs. avoidance) actions regardless of the stimulus valence,
but no such effect was observed for pleasant compared to unpleas-
ant stimuli. This asymmetry effect during approach–avoidance ac-
tion motivations occurred in the dorsolateral but not orbito-frontal
aspects of prefrontal cortex. Aussi, individual differences in
approach–avoidance motivation moderated the effect such that
increasing trait approach motivation was associated with greater
left-sided asymmetry during approach actions (regardless of the
stimulus valence). Ensemble, these results support the notion
that prefrontal asymmetry is associated with action motivation
regardless of stimulus valence and, as such, might be linked with
goal pursuit processes more broadly. ■
INTRODUCTION
Goal-relevant stimuli in the environment typically can
be categorized into goal-consistent stimuli that should be
approached or goal-inconsistent stimuli that should be
avoided. Once this categorization takes place, the impetus
to move toward (or away from) a stimulus, as well as the
action itself, is known as approach (or avoidance) motiva-
tion. This triage process is relatively straightforward for
most animals: move toward positively valenced stimuli
and move away from negatively valenced stimuli. For hu-
mans, goal pursuit is not always as easy. Although we share
with other animals the same general tendencies to ap-
proach positive and avoid negative stimuli, one of the de-
fining qualities of human goal pursuit is that we have the
capacity to approach negative and avoid positive stimuli if
that behavior serves our ultimate goal. Par exemple, pendant
dieting, someone with a sweet tooth who generally dislikes
vegetables might, nonetheless, have a motive to avoid
candy and instead snack on broccoli. Previous work has ex-
amined the prefrontal cortical asymmetry associated with
approaching positive and avoiding negative stimuli (par exemple.,
Sutton & Davidson, 1997). Cependant, with the exception
Université de Californie, Les anges
of studies on anger (discussed below), most of these stud-
ies have not shed light on whether this asymmetry is driven
by stimulus valence (c'est à dire., positive vs. negative) or action
motivation (approach vs. avoidance). The present study sep-
arates the roles of stimulus valence and action motivation
in hemispheric asymmetry for the first time using fMRI.
Neuroscience investigations that did not separate ac-
tion from valence have repeatedly observed left–right pre-
frontal cortical asymmetry between approaching pleasant
and avoiding unpleasant stimuli. Research conducted pre-
dominantly using electroencephalography (EEG) a
found that approach–pleasant is associated with relatively
greater activity in left lateral prefrontal cortex (PFC; relative
À droite), and avoid–unpleasant is associated with rela-
tively greater activity in right lateral PFC (compared to
gauche) (Sutton & Davidson, 1997; Davidson, Ekman, Saron,
& Senulis, 1990). Lesion studies in rats (Robinson, 1979)
and humans (Starkstein et al., 1991) have also supported
the link between left PFC and approach–pleasant behav-
iors. Other EEG studies replicate this finding but also do
not distinguish between stimulus valence and action moti-
vation. Par exemple, lateral frontal asymmetry (measured
by EEG) has been observed in response to smelling a
pleasant vanilla odor compared to a neutral odor (Kline,
Blackhart, Woodward, Williams, & Schwartz, 2000), et
© 2009 Massachusetts Institute of Technology
Journal des neurosciences cognitives 22:9, pp. 1970–1979
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
2
je
9
e
–
1
p
9
d
7
F
0
/
1
2
9
2
3
/
9
9
8
/
1
1
9
9
o
7
c
0
n
/
1
2
0
7
0
7
9
0
4
2
6
1
7
3
1
/
7
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
9
e
.
s
t
2
o
1
n
3
1
0
7
8
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
/
.
t
.
.
F
o
n
1
8
M.
un
oui
2
0
2
1
in two studies comparing monetary rewards compared to
pertes (Miller & Tomarken, 2001; Sobotka, Davidson, &
Senulis, 1992). In both of these examples, the valence of
the stimulus (pleasant/unpleasant) covaried completely
with the behavioral tendency (approach/avoid).
Several fMRI studies have replicated the basic finding of
hemispheric asymmetry being associated with approach–
pleasant versus avoid–unpleasant, but as in the EEG litera-
ture, these studies did not disentangle action from valence.
In the two fMRI studies that explicitly examined asymme-
try by directly comparing activation in the right and left
hemispheres, participants were either passively shown
pleasant and unpleasant words (Herrington et al., 2005)
or pictures (Canli, Desmond, Zhao, Glover, & Gabriela,
1998). These studies suggest that asymmetry during pleas-
ant stimuli is associated with increases in left dorsolateral
(Herrington et al., 2005) or throughout the left PFC (Canli
et coll., 1998), and one of the two found unpleasant stimuli
to be associated with activation in right PFC (Canli et al.,
1998). Another fMRI study examined the relation be-
tween hemispheric asymmetry and promotion/prevention
regulatory focus (Eddington, Dolcos, Cabeza, Krishnan,
& Strauman, 2007), a construct distinct from approach/
avoidance but, nonetheless, related (Cunningham, Raye,
& Johnson, 2005; Higgins, 1997). The authors found in-
creases in left orbito-frontal PFC when participants were
primed with positively valenced adjectives associated with
their “ideal self” (promotion regulatory focus) compared
to those associated with their “ought self ” (prevention
regulatory focus). Ensemble, these findings are broadly
consistent with the EEG literature but still cannot identify
whether the asymmetry is driven by action tendency or
stimulus valence, because in each study, pleasant and un-
pleasant stimuli were likely to have activated approach
and avoidance tendencies, respectivement.
One possible way to disentangle valence and action is to
study an affective–motivational state in which the two are
naturally in opposition—approach–pleasant or avoid–
unpleasant. The emotion anger is believed to be one such
state because it possesses both an unpleasant valence
and an approach motive (Carver & Harmon-Jones, 2009;
Harmon-Jones & Allen, 1998). Harmon-Jones et al. (2002)
and Harmon-Jones and Sigelman (2001) have capitalized
on this fact and examined the effect of both state and
trait anger on hemispheric asymmetry. Their studies have
revealed increased left lateral prefrontal asymmetry in re-
sponse to an anger induction such as an insult. Supporting
this work, another study found that decreased memory
for anger following an experimental reduction of activity
in left (but not right) lateral PFC using repetitive transcranial
stimulation magnétique (SMTr) (van Honk & Schutter, 2006).
Ensemble, these studies suggest that asymmetry is associ-
ated with action motivation rather than valence.
Although these studies provide one way to separate
action and valence, the extent to which the results might
generalize to goal pursuit is unclear. Approaching during
anger is different than approaching an unpleasant stimulus
during goal pursuit. In the context of a goal, individuals
presumably engage in self-regulation in order to approach
unpleasant stimuli that they would otherwise avoid (par exemple.,
someone who wants to lose weight but doesnʼt like to
exercise might nonetheless go to the gym every day). Dans
contraste, the tendency to approach during anger seems
to be intrinsic to the experience of anger and does not
require overriding a natural tendency to avoid. Plus loin,
anger appears to be unique as a mental state that is nega-
tively valenced and also approach-motivated, and there is
no parallel affective state that is positively valenced and also
avoidance-motivated. Ainsi, it is important to determine
if this is unique to anger or generalizes to action versus
valence more broadly.
A second caveat to the broad conclusion that lateral pre-
frontal asymmetry is associated with approach–avoidance
motivation is that, although several studies have replicated
the link between approach motivation and increased left
prefrontal cortical activation, many studies fail to repli-
cate the finding that avoidance motivation is associated
with increased right prefrontal activation (Amodio, Master,
Yee, & Taylor, 2008; Hewig, Hagemann, Seifert, Naumann,
& Bartussek, 2006; Pizzagalli, Sherwood, Henriques,
& Davidson, 2005; Jackson et al., 2003; Coan, Allen, &
Harmon-Jones, 2001; Henriques & Davidson, 2000; Kline
et coll., 2000). Researchers have speculated that this in-
consistency might arise because of the theoretical com-
plexity of the avoidance construct (Coan & Allen, 2004),
or because approach is not necessarily the opposite of
avoidance (Amodio et al., 2008) et, instead, relies upon
distinct psychological processes that might not be elic-
ited by the experimental paradigms used to investigate
frontal asymmetry. Another explanation is that EEG is not
ideally suited to measuring differences in activation be-
tween hemispheres because it relies on comparisons
in voltage between two sites that are often nonindepen-
bosse (Allen, Coan, & Nazarian, 2004). If this were the case,
convergent evidence from a different neuroimaging mo-
dality such as fMRI would help to clarify the inconsistent
findings regarding right lateral prefrontal asymmetry and
avoidance.
The key limitation in the current literature on asym-
metry is the confounding of action and valence. It is dif-
ficult to disentangle action and valence because they are
most often confounded in the real world—people typi-
cally do approach pleasant things and avoid unpleasant
things, and it is typically easy to do so—but they are not
always confounded. During goal pursuit, action motivation
and stimulus valence are sometimes in opposition. En effet,
one reason why goals are challenging and we often fail at
them is because success frequently depends on actions
that are in opposition to our natural response to a stimulus,
such as when a dieter approaches healthy (but unpleasant)
foods or a smoker avoids a cigarette. For this reason, un
task that models this type of goal pursuit situation (c'est à dire.,
approaching unpleasant and avoiding pleasant stimuli) est
ideally suited to disentangle action from valence.
Berkman and Lieberman
1971
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
2
je
9
e
–
1
p
9
d
7
F
0
/
1
2
9
2
3
/
9
9
8
/
1
1
9
9
o
7
c
0
n
/
1
2
0
7
0
7
9
0
4
2
6
1
7
3
1
/
7
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
9
e
.
s
t
2
o
1
n
3
1
0
7
8
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
.
.
F
t
/
.
o
n
1
8
M.
un
oui
2
0
2
1
The present study employs a novel task that models goal
pursuit in order to cross action motivation (approach/
avoidance) with stimulus valence (pleasant/unpleasant)
and directly compare the role of these two factors in pre-
frontal cortical asymmetry. In the Nochmani paradigm
(Berkman, Lieberman, & Gable, 2009), participants first
read a fake National Geographic-style article about a newly
discovered tribe of people, the Nochmani. From the article,
participants learn that the Nochmani are similar to West-
erners in their enjoyment of sweets and their distaste for
fungi, but are dissimilar in their fondness for eating insects
and disgust when eating meats. In the subsequent response
time task, Western participants respond whether to “eat” or
“not eat” various foods from the perspective of the Noch-
mani (Chiffre 1). In this way, the action to “eat” (approche)
or “not eat” (avoidance) can be crossed with the valence of
the delicious (pleasant) or disgusting (unpleasant) nourriture
stimuli into an orthogonal design.
We used fMRI to measure neural responses to trials in
each of these conditions. We analyzed the data by generat-
ing a priori ROIs in left and right dorsolateral and orbito-
frontal cortices and directly comparing left to right activation
Chiffre 1. The Nochmani task. (UN) Trials are drawn from one of four
experimental conditions defined by the 2 (action motivation: approach/
avoidance) × 2 (valence: pleasant/unpleasant) within-subjects factorial
conception. (B) Two-second trials are presented in blocks of 10 that are
composed predominantly (80%) of trials from one cell.
in each of these regions during the approach, avoidance,
pleasant, and unpleasant conditions relative to baseline.
Activation in these ROIs was also correlated with a trait
measure of approach and avoidance motivation. Based
on the neuroimaging literature discussed above, we ex-
pected asymmetry to be associated with action motivation
rather than stimulus valence. There have been few attempts
to identify the source of the asymmetry within PFC. The only
EEG study to use a formal localization procedure identified
three foci in left dorsolateral prefrontal cortex (DLPFC) que
were linked to reward preference (Pizzagalli et al., 2005), et
several other EEG studies have identified DLPFC more
broadly using informal localization procedures (par exemple., scalp
topography) (Amodio et al., 2008; Harmon-Jones & Allen,
1998; Sutton & Davidson, 1997). The fMRI studies identified
either dorsolateral (Herrington et al., 2005) or orbito-frontal
(Eddington et al., 2007) PFC, although the Eddington et al.
(2007) study used a manipulation of promotion–prevention
focus instead of approach–avoidance. Based on these stud-
ies, we predicted that the asymmetry would be localized only
to DLPFC, and included orbito-frontal cortex as a compari-
son region. We also predicted that trait approach–avoidance
motivation would moderate asymmetry such that higher
levels of approach and avoidance would be associated with
increased left- and right-sided asymmetry, respectivement.
MÉTHODES
Participants
Seventeen right-handed participants (9 men; ages = 19–
28 années, M = 23.4 ± 2.7) were recruited from the UCLA
community and paid $25 for their participation. Le
participants were prescreened to be nonvegetarians. All
participants provided written informed consent that was
approved by the UCLA Office for Protection of Research
Sujets.
Procedure
We generated an fMRI-compatible version of a validated
paradigm that was designed to separate action motivation
and valence (Berkman et al., 2009). Before entering the
scanner, participants read a realistic but fake article about
the Nochmani (see http://berkman.bol.ucla.edu/Nochma-
ni.pdf for download). Participants were told to read the arti-
cle carefully because, subsequently, in the scanner they
would be completing a memory task about the Nochmani. Al-
though the subjects were led to believe that the task involved
mémoire, success in the forthcoming task depended only
on participants remembering two unusual characteristics
of the Nochmani—that they enjoyed eating insects, et
were disgusted by eating meat. The Nochmani otherwise
share Western tastes in food as they enjoy eating desserts
and are disgusted by eating fungus-infested foods. Success
rates on the present study and a previous study (Berkman
1972
Journal des neurosciences cognitives
Volume 22, Nombre 9
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
2
je
9
e
–
1
p
9
d
7
F
0
/
1
2
9
2
3
/
9
9
8
/
1
1
9
9
o
7
c
0
n
/
1
2
0
7
0
7
9
0
4
2
6
1
7
3
1
/
7
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
9
e
.
s
t
2
o
1
n
3
1
0
7
8
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
.
.
.
t
F
/
o
n
1
8
M.
un
oui
2
0
2
1
et coll., 2009) confirm that participants are easily able to
retain this information for the duration of the task. Après
reading the article, participants completed a self-report
measure of trait approach and avoidance motivation (de-
scribed below).
Suivant, participants completed a computerized response
time task during fMRI acquisition. They were shown a se-
ries of trials that each displayed a single picture of a dessert,
meat, fungus, or insect. Participants were asked to respond
as quickly as possible via keypress whether Nochmani
would “eat” or “not eat” the food. Trials were arranged
into blocks that represented one of the four cells of the 2
(action motivation: approach/avoid) × 2 (valence: pleasant/
unpleasant) conception. Each block comprised 80% trials from
one cell (par exemple., approach–pleasant) et 20% “foil” trials from
cells with an opposite action (c'est à dire., avoid–pleasant or avoid–
unpleasant). These minority foil trials serve to prevent par-
ticipants from falling into a response set on a given block
(par exemple., simply responding “eat” to every trial without pay-
ing attention to the content of the image). The duration
of each trial was set to be sufficiently long (2 sec) so that
participants had enough time to consider each trial—
including the foil trials—and that these trials would not
act as “oddballs” as in a speeded detection task. The four
block types were evenly and randomly distributed through-
out the experiment.
The key blocks for unconfounding action and valence
are those in which the valence of the stimulus conflicts
with the direction of the action required for a correct re-
sponse. Spécifiquement, these trials occur when responding
to images of insects (approach–unpleasant) and meat
(avoid–pleasant).
Each trial lasted 2 sec, and participants could respond
at any point during that time. The images remained on the
screen for the duration of the trial following a response.
Trials occurred within 20 blocks of 10 trials each that began
avec 4 sec of instructions per block and 12 sec of resting
fixation between blocks. This yielded a total of 200 trials
spread across two 6-min functional runs.
Foam padding was placed around participantsʼ heads to
reduce motion. Stimuli were presented on LCD goggles,
and responses were recorded on a magnet-safe button
box placed in the right hand.
Materials
The pictures of food used in the task belonged to one of
four categories: fungi, desserts, meats, or insects. Là
étaient 40 pictures in each category for a total of 160 color
pictures. The four categories were pre-rated to be equiva-
lent on absolute ratings of valence (c'est à dire., the fungi and in-
sects are equally unpleasant, the desserts and meats were
equally pleasant, and all four have equal absolute values on
valence; Berkman et al., 2009). The images were standard-
ized on brightness, contraste, and fixed at a resolution of 500
par 375 to maintain a 4:3 width-to-height ratio.
Trait behavioral activation and inhibition was measured
using the Behavioral Inhibition/ Behavioral Activation
Scales (BIS/BAS; Carver & Blanc, 1994). The BAS has three
subscales: The BAS-Drive subscale measures persistent
pursuit of goals (par exemple., “I go out of my way to get things I
want”), α = .73; the BAS-Fun Seeking subscale measures
desire for new rewards (par exemple., “I crave excitement and new
sensations”), α = .78; and the BAS-Reward Responsive-
ness subscale relates to positive responses to reward
(par exemple., “When I get something I want, I feel excited and en-
ergized”), α = .68. The reliability for the overall BAS was
.84. The BIS scale is thought to be unidimensional, et
taps sensitivity to negative events (par exemple., “Criticism or scold-
ing hurts me quite a bit”), α = .82.
fMRI Data Acquisition and Analysis
Data were acquired on a Siemens Allegra 3-T scanner at the
UCLA Ahmanson-Lovelace Brainmapping Center. High-
resolution structural T2-weighted echo-planar images (spin-
echo; TR = 5000 msec; LE = 33 msec; matrix size 128 ×
128; 36 sagittal slices; FOV = 20 cm; 3 mm thick, skip
1 mm) were acquired coplanar with the functional scans.
Two functional scans lasting 6 min each were acquired
during the task (echo-planar T2*-weighted gradient-echo;
TR = 2000 msec; LE = 25 msec; flip angle = 90°; matrice
size 64 × 64; 34 tranches axiales; FOV = 20 cm; 3 mm thick; skip
1 mm). The imaging data were analyzed using SPM5 (Bien-
come Department of Cognitive Neurology, Institute for
Neurologie, Londres, ROYAUME-UNI). Images from each participant
were realigned to correct for head motion, normalized into
l'Institut neurologique de Montréal (MNI) standard stereo-
tactic space, and smoothed with an 8-mm Gaussian kernel,
full width at half maximum.
The design was modeled as a blocked 2 (action: approach/
avoidance) × 2 (valence: pleasant/unpleasant) factorial de-
sign with eight 12-sec fixation-cross periods per run form-
ing a baseline. Linear contrasts were computed to assess
the difference in neural activity during each condition com-
pared to baseline. For all analyses, individual-participant
contrasts were generated with fixed-effects models and
then grouped into a random-effects model for greater
generalizability.
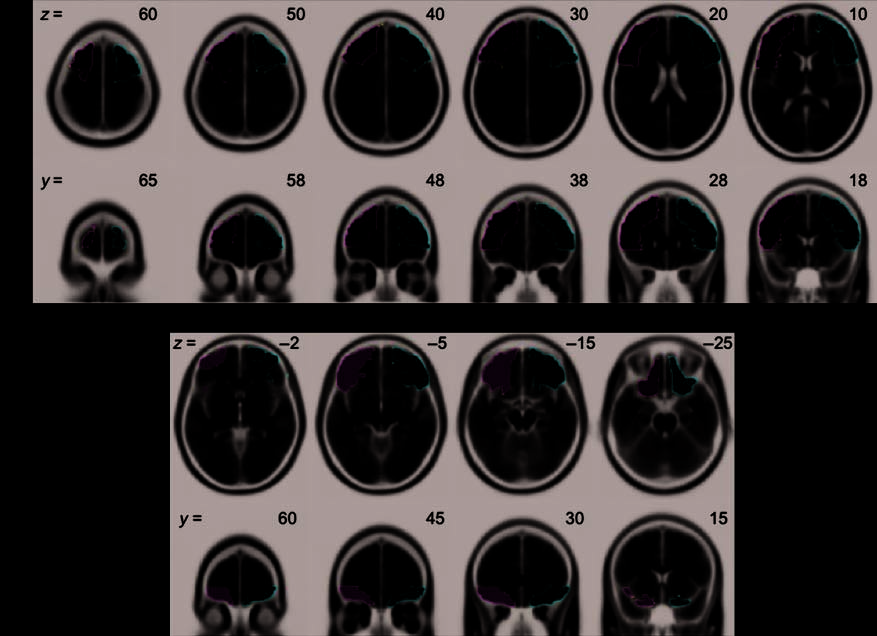
A priori ROIs were created for the dorsolateral and orbito-
frontal cortices, separately for left and right, using the Wake
Forest University Pickatlas Tool (Maldjian, Laurienti, Kraft,
& Burdette, 2003) based on the Automated Anatomical
Labeling atlas (AAL; Tzourio-Mazoyer et al., 2002). The dor-
solateral ROIs were a combination of AALʼs superior and
middle frontal gyri, pars opercularis, and pars triangularis,
all superior to and including the axial plane at MNI z = 2
(Figure 2A). The orbito-frontal ROIs were a combination
of AALʼs orbital subregions of the superior, middle, et
inferior frontal gyri, all inferior to and including the axial
plane at MNI z = 0 (Figure 2B). All voxels within each of
the four resulting ROIs were averaged to create a total of
four values per participant per condition.
Berkman and Lieberman
1973
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
2
je
9
e
–
1
p
9
d
7
F
0
/
1
2
9
2
3
/
9
9
8
/
1
1
9
9
o
7
c
0
n
/
1
2
0
7
0
7
9
0
4
2
6
1
7
3
1
/
7
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
9
e
.
s
t
2
o
1
n
3
1
0
7
8
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
.
/
t
.
.
F
o
n
1
8
M.
un
oui
2
0
2
1
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
2
je
9
e
–
1
p
9
d
7
F
0
/
1
2
9
2
3
/
9
9
8
/
1
1
9
9
o
7
c
0
n
/
1
2
0
7
0
7
9
0
4
2
6
1
7
3
1
/
7
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
9
e
.
s
t
2
o
1
n
3
1
0
7
8
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
.
F
t
.
.
/
o
n
1
8
M.
un
oui
2
0
2
1
Chiffre 2. The ROIs for the main analysis. Further details are provided in the text. (UN) The dorsolateral region (z ≥ 2) et (B) the orbito-frontal
region (z ≤ 0) defined using the AAL atlas.
The primary research questions regarding hemispheric
asymmetry were examined in the prefrontal ROIs. Following
convention in the EEG literature (par exemple., Harmon-Jones
et coll., 2002), an asymmetry score was calculated for
each participant for the dorsolateral and orbito-frontal
ROI. This score was then entered as the dependent mea-
sure in a within-subjects factorial ANOVA. Further analyses
examined whether trait-level approach and avoidance
motivation moderated activation in the ROIs across task
conditions. Because of the strong a priori hypotheses
about hemispheric asymmetry, and because our depen-
dent measure in the 2 × 2 ANOVA and regression analyses
was a single asymmetry score for each of the two regions
(instead of thousands of voxels), we used a traditional
p value of .05. All neuroimaging results are reported in
MNI coordinates.
RÉSULTATS
Behavioral Responses
As in previous work, participants achieved a high rate of
accuracy in all conditions (M = 97.4%) and no participant
was below 95% accuracy (Berkman et al., 2009). Because
of this high rate of accuracy, all trials were included in the
blockwise analysis.
Participants were significantly faster to approach [M =
753.6 msec, SD = 25.0 msec] than to avoid [M = 875.8 msec,
SD = 59.2 msec; t(16) = 5.31, p < .01]. There were no
differences in response time between the pleasant [M =
802.1 msec, SD = 43.7 msec] and unpleasant [M =
826.3 msec, SD = 127.8 msec; t(16) = 0.56, ns] valence
conditions. The Action × Valence interaction was not
significant [t(16) = 1.60, ns]. Because of the main effect
of action, mean response time for each block was entered
into the model as a covariate.
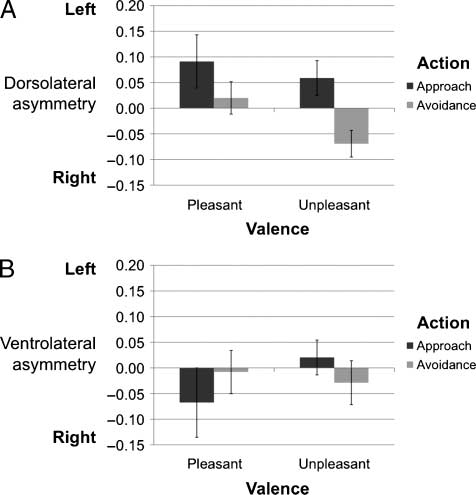
Main Effects of Task Condition
The main dependent measures were the left–right asym-
metry scores for the dorsolateral and orbito-frontal ROIs.
These scores were computed for both of the ROIs for each
of the four conditions relative to fixation baseline. For ex-
ample, the dorsolateral asymmetry score in the approach–
pleasant condition was computed as: (approach–pleasant
R DLPFC − baseline) − (approach–pleasant L DLPFC −
baseline). Once these scores were computed, 2 × 2 within-
subjects ANOVAs were run on the asymmetry scores sep-
arately for each ROI.
A main effect of action and no main effect of valence
would support the hypothesis that dorsolateral asymmetry
is driven by action motivation and not by stimulus valence.
In the dorsolateral ROIs, there was a main effect of ac-
tion [F(1, 16) = 7.07, p < .02], no main effect of valence
[F(1, 16) = 0.59, ns], and no interaction [F(1, 16) = 0.24,
1974
Journal of Cognitive Neuroscience
Volume 22, Number 9
asymmetry during avoidance relative to baseline (i.e.,
right-baseline > left-baseline). Neither comparison was sig-
nificant (both ps > .2), suggesting that the main effect of
action is driven by a combination of the increase in asym-
metry during approach and decrease in asymmetry during
avoidance.
Correlations with Trait Measures
of Approach–Avoidance
We demonstrated above that, on average, action motiva-
tion is associated with dorsolateral prefrontal asymmetry.
Suivant, based on prior findings that individual differences
in approach–avoidance motivation are associated with
asymmetry as measured by EEG (Amodio, Shah, Sigelman,
Brazy, & Harmon-Jones, 2004; Harmon-Jones & Allen,
1998), we tested whether trait-level motivation would mod-
erate the main effect of action motivation on asymmetry
described above. Pour faire ça, we correlated asymmetry in
each ROI during each condition with scores on Carver
and Whiteʼs (1994) BAS and BIS. Par exemple, a positive
correlation between approach motivation and left–right
asymmetry during approach actions would suggest that,
even though there is a main effect of action on average,
individuals with higher approach motivation tend to show
greater left-lateralized asymmetry than those with lower
approach motivation during that condition.
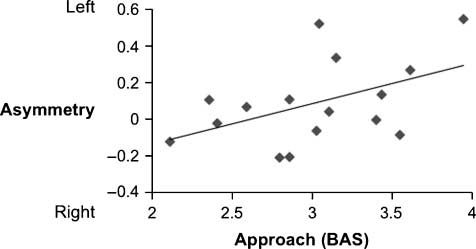
Left-sided asymmetry was correlated with the overall
BAS (controlling for BIS) during approach relative to base-
line [overall: r(16) = .52, p < .05; approach–pleasant:
r(16) = .37, ns; approach–unpleasant: r(16) = .29, ns].
The relationship between the overall BAS and approach
actions, controlling for BIS, is shown in Figure 4. Asymme-
try was not associated with BAS during avoidance actions
relative to baseline [r(16) = .29, ns], or with viewing pleas-
ant [r(16) = .36, ns] or unpleasant [r(16) = .35, ns] stimuli
compared to baseline. Because of the relatively small sam-
ple size, the nonsignificant correlations were not signifi-
cantly different from the significant association between
BAS and approach action motivation. These correlations
indicate that trait approach (but not avoidance) motivation
Figure 4. The correlation between left-sided asymmetry and trait
approach motivation (as measured by the BAS scale) during approach
actions [r(16) = .52, p < .05].
Berkman and Lieberman
1975
Figure 3. Asymmetry scores in (A) dorsolateral and (B) orbito-frontal
PFC. Each score was generated by calculating the average left–right
change from baseline during each condition defined by the 2 (action) ×
2 (valence) factorial design. The only significant effect is the main
effect of action in the dorsolateral region [F(1, 16) = 7.07, p < .02].
This effect was marginally significantly larger than the effect in the
orbito-frontal region [F(1, 16) = 3.83, p < .07]. Error bars represent
two standard errors.
ns; Figure 3A]. Specifically, there was greater relative left-
sided asymmetry in the dorsolateral ROI during approach
(M = 0.075, SD = 0.23) than avoidance (M = −0.025, SD =
0.23), but not during pleasant (M = 0.056, SD = 0.34) ver-
sus unpleasant (M = 0.005, SD = 0.17) stimuli. The null
interaction suggests that the relative left-sided asymmetry
during approach versus avoidance for the pleasant stimuli
(M = 0.071, SD = 0.35) was not different from the asym-
metry between approach and avoidance for unpleasant
stimuli (M = 0.128, SD = 0.24).
Next, we tested the same hypotheses in the orbito-
frontal region to examine the spatial specificity of the
asymmetry. If prefrontal asymmetry was localized only
to the dorsolateral region, we would expect to find no
effects of task condition on asymmetry. As expected,
there were no main effects in the orbito-frontal ROIs of
action [F(1, 16) = 0.01, ns] or valence [F(1, 16) = 0.06,
ns], and no Action × Valence interaction [F(1, 16) =
0.42, ns; Figure 3B]. The interaction between action
and region testing whether the effect of action on asym-
metry was greater in the dorsolateral than orbito-frontal re-
gion was marginally significant [F(1, 16) = 3.83, p < .07].
We then conducted a series of post hoc paired-samples
t tests to investigate whether the increased asymmetry
during approach compared to avoidance actions was being
driven by increased asymmetry during approach relative to
baseline (i.e., left-baseline > right-baseline) or decreased
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
2
je
9
e
–
1
p
9
d
7
F
0
/
1
2
9
2
3
/
9
9
8
/
1
1
9
9
o
7
c
0
n
/
1
2
0
7
0
7
9
0
4
2
6
1
7
3
1
/
7
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
9
e
.
s
t
2
o
1
n
3
1
0
7
8
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
/
.
.
.
F
t
o
n
1
8
M.
un
oui
2
0
2
1
moderates asymmetry specifically during approach actions,
but not during viewing of pleasant stimuli or avoidance
actes.
Based on this finding for the overall (c'est à dire., average) BAS,
we also examined the correlation of each of the BAS sub-
scales during approach baseline. The subscale correla-
tions revealed a similar pattern of results in terms of
direction and magnitude, although the only subscale that
yielded a significant correlation with asymmetry was the
Drive subscale during approach baseline [r(16) = .55,
p < .05].
Finally, we examined the relationship of trait avoidance
(as measured with the BIS scale) with asymmetry. A
negative correlation between avoidance and asymmetry
would indicate that higher levels of trait avoidance were
associated with greater right-sided asymmetry (because
asymmetry is always calculated as left–right). Asymmetry
was not significantly correlated with BIS (controlling for
overall BAS) in any of the four conditions individually,
nor with any of the marginal averages (e.g., approach
actions averaging across pleasant and unpleasant valences;
all ps > .1).
DISCUSSION
The present study is the first to use fMRI to disentangle the
contributions of action motivation and stimulus valence
in hemispheric asymmetry. Previous fMRI studies demon-
strated that positively and negatively valenced stimuli, que
naturally induce a tendency to approach or avoid, respecter-
tivement, are associated with asymmetry, but these studies
could not specify whether the asymmetry was linked to
action motivation, valence, or both. Our findings address
this limitation by demonstrating that asymmetry is asso-
ciated with action rather than valence, and specifically,
that approach motivation is associated with increased ac-
tivation in left relative to right PFC. En outre, the prior
work untangling action from valence has used EEG and
has yet to be replicated using an alternative neuroimaging
method such as fMRI. Using fMRI in combination with ROIs
that were guided by previous work allowed us to replicate
findings from the EEG literature to localize the asymme-
try to the dorsolateral and not the orbital aspects of PFC.
En plus, consistent with our expectations, trait motiva-
tion moderated the asymmetry effect such that individuals
with higher levels of trait approach motivation demon-
strated increased asymmetry during approach actions.
We used the novel Nochmani experimental paradigm
(Berkman et al., 2009) to completely cross two variables
that had previously been confounded—action motivation
(approach vs. avoidance) and stimulus valence (pleasant
vs. unpleasant). This allowed us to independently assess
the degree to which hemispheric asymmetry was linked
to action and valence. The data suggest that left-sided
asymmetry is associated with approach actions and not
viewing of pleasant stimuli per se. These results are con-
sistent with findings by Harmon-Jones and Sigelman (2001)
and Harmon-Jones and Allen (1997, 1998) that prefrontal
asymmetry is associated with state and trait anger, et ça
anger is an approach-related emotion (Carver & Harmon-
Jones, 2009). The fact that we observed increased left-
lateral asymmetry in response to nonanger unpleasant
émotions (par exemple., disgust) when they were coupled with an
approach response adds convergent evidence to this view.
Our finding that prefrontal hemispheric asymmetry is
tied to action motivation rather than stimulus valence fits
with current goal theory and our understanding of the
functions of PFC. In a hierarchical model of goal pursuit,
behavior can be determined by both proximal and distal
motives that vary in abstraction (Elliot, 2006; Elliot &
Church, 1997). Par exemple, in the Nochmani paradigm,
behavior can be motivated by either the proximal, lower-
level, hedonic value of the stimulus (c'est à dire., motivation to
approach pleasant and avoid unpleasant stimuli), or by
the more distal, higher-level, instrumental goal of success
(c'est à dire., motivation to respond correctly even if that some-
times requires approaching unpleasant and avoiding
pleasant stimuli). As with many real-world goals, acting
in line with the higher-level goal—even when it is in op-
position to the lower-level hedonic value—is the only way
to reliably attain success on the Nochmani task. Ainsi, un
novel and important feature of the Nochmani task is that
it contains conditions that require top–down control to
promote higher-level goals over lower-level tendencies.
The high rates of success by the participants in the pres-
ent investigation suggest that they were engaging control
by acting according to the higher-level goal and not the
lower-level stimulus-driven response. At a neuroanatomical
level, there is broad consensus that the main role of PFC is
to plan and coordinate precisely this kind of goal-directed
action that often requires control (Fuster, 2008; Miller &
Cohen, 2001). In contrast, the experience of pleasant and
unpleasant affective states is often associated with acti-
vation in subcortical structures such as the amygdala,
insula, anterior cingulate cortex, and basal ganglia (Phillips
et coll., 2004; Phan, Wager, Taylor, & Liberzon, 2002; Whalen
et coll., 2001; Damasio et al., 2000; Mayberg et al., 2000;
Morris, Ohman, & Dolan, 1998; Drevets et al., 1997; Georges,
Ketter, Parekh, Herscovitch, & Post, 1996). One meta-
analysis explicitly searched for hemispheric asymmetries
between pleasant and unpleasant emotions and found
no such asymmetries ( Wager, Phan, Liberzon, & Taylor,
2003). At least, in the current study, where attaining an ab-
stract goal was linked to action motivation, it makes sense
that action motivation, and not stimulus valence, is asso-
ciated with asymmetrical prefrontal activity. It is an open
question whether this interpretation would hold for other
findings in this literature.
The notion that there are hemispheric asymmetries in
approach compared to avoidance action motivation is
broadly consistent with prior work investigating processes
that can be characterized as predominantly approach or
avoidance. Par exemple, engaging in an avoidant emotion
1976
Journal des neurosciences cognitives
Volume 22, Nombre 9
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
2
je
9
e
–
1
p
9
d
7
F
0
/
1
2
9
2
3
/
9
9
8
/
1
1
9
9
o
7
c
0
n
/
1
2
0
7
0
7
9
0
4
2
6
1
7
3
1
/
7
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
9
e
.
s
t
2
o
1
n
3
1
0
7
8
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
F
.
.
/
t
.
o
n
1
8
M.
un
oui
2
0
2
1
regulation strategy (par exemple., by taking a distant or detached
perspective on emotional stimuli) has been shown to
produce activation in right lateral PFC (Kalisch et al.,
2005). Being distracted during a painful stimulus has also
been shown to increase activation in this region (Bantick
et coll., 2002). Enfin, Mitchell et al. (2007) found increases
in right dorsolateral PFC when participants attempt to
avoid thinking about white bears. D'autre part,
researchers have noted left lateral prefrontal activation dur-
ing approach-oriented tasks such as the planning and execu-
tion of tool use (Kroliczak & Frey, 2009; Frey, 2007, 2008),
action selection (Schluter, Krams, Rushworth, & Passing-
ham, 2001), and action planning (Bohlhalter et al., 2008). Al-
though suggestive, these studies often do not explicitly
compare left to right activation, Et ainsi, the current study
makes an important contribution by doing so. It is also
unclear whether action planning is intrinsically approach-
oriented, or whether the planning of avoidance actions
is different but simply not yet been investigated. Future
research can build upon these studies with the aim of clarify-
ing the action motivation of their tasks and directly assessing
hemispheric asymmetries.
Our use of a priori ROIs to directly compare activation in
left and right PFC during approach and avoidance actions
is novel, and allows for several interesting inferences. D'abord,
although functionally defined ROIs have been compared in
the past (par exemple., Herrington et al., 2005), the regions used in
the present article are relatively large for fMRI. The fact that
we were able to replicate findings from EEG, which has a
larger spatial extent than fMRI, suggests that using large
ROIs can be a useful tool to link across the two methods.
Deuxième, it is interesting to note that asymmetries emerge
in an ROI analysis although there have been no obvious
asymmetries in previous fMRI studies of action motivation
(par exemple., Wager et al., 2003). It might be the case that asym-
metries between approach- and avoidance-related action
motivations may be somewhat distributed across DLPFC,
or occur consistently at small number of sites that are each
below traditional significance thresholds. Besides pointing
to the general utility in comparing the results from ROI and
whole-brain analyses, this observation highlights the need
for future research to better understand the psychological
and neural components of approach-related motivation
and actions.
Enfin, we found that individuals with higher trait-level
approach motivation showed increased left-sided asymme-
try during approach actions. This correlation was specific
to approach actions and did not generalize to avoidance
actes, or to either valence condition averaging across ac-
tion motivation. Fait intéressant, trait-level avoidance motiva-
tion (as measured by Carver & Whiteʼs, 1994 BIS scale) était
not associated with asymmetry in any condition. The find-
ing that BIS scores were not associated with asymmetry
replicates other findings (par exemple., Amodio et al., 2008), et
is consistent with the notion that this scale is relevant to
conflict monitoring and detection more than avoidance
actions per se. Ainsi, the fact that the present study was de-
signed to maximize correct responses may contribute to
this null finding. En effet, Amodio et al. (2008) found that
during a go/no-go task, BIS correlated with one ERP signal
(the N2) during successful inhibition (“no-go” trials) mais
not successful execution (“go” trials), and with another
signal (the error-related negativity) during failed inhibition
but not during successful inhibition trials. Each of these
findings supports the role of BIS in conflict monitoring
and error detection rather than avoidance behaviors more
broadly.
Future research can build on the current study in a more
realistic goal pursuit context. Although one of the strengths
of the Nochmani paradigm is the external validity of the
stimuli, the task itself is meant to be a laboratory model
for real-world goal pursuits such as dieting or smoking
cessation. Based on the current results, it might be the case
that prefrontal asymmetry plays a role in coordinating goal-
directed approach and avoidance actions, and further, que
individual differences in approach–avoidance motivation
moderate asymmetry and are thus relevant to goal pur-
suit. We hope that the insights from the present study
will inform future work not only on the neuroanatomical
underpinnings of action motivation but also on the com-
plex goals to which those behaviors and motivations are
appliqué.
Remerciements
This research was supported by NIDA grants T90DA022768 and
F31DA024904 (E. T. B.) and NIMH grants R21MH071521 and
R01MH084116 (M.. D. L.).
Reprint requests should be sent to Elliot T. Berkman, UCLA Psy-
chology Department, 1285 Franz Hall, Box 951563, Les anges,
Californie 90095-1563, ou par e-mail: berkman@ucla.edu.
RÉFÉRENCES
Allen, J.. J., Coan, J.. UN., & Nazarian, M.. (2004). Issues and
assumptions on the road from raw signals to metrics of
frontal EEG asymmetry in emotion. Biological Psychology,
67, 183–218.
Amodio, D. M., Master, S. L., Yee, C. M., & Taylor, S. E. (2008).
Neurocognitive components of the behavioral inhibition
and activation systems: Implications for theories of
self-regulation. Psychophysiology, 45, 11–19.
Amodio, D. M., Shah, J.. Y., Sigelman, J., Brazy, P.. C., &
Harmon-Jones, E. (2004). Implicit regulatory focus
associated with asymmetrical frontal cortical activity.
Journal of Experimental Social Psychology, 40, 225–232.
Bantick, S. J., Sage, R.. G., Ploghaus, UN., Clare, S., Forgeron, S. M.,
& Tracey, je. (2002). Imaging how attention modulates
pain in humans using functional MRI. Cerveau, 125,
310–319.
Berkman, E. T., Lieberman, M.. D., & Gable, S. L. (2009). BIS,
BAS, and response conflict: Testing predictions of the
revised reinforcement sensitivity theory. Personality and
Individual Differences, 46, 586–591.
Bohlhalter, S., Hattori, N., Wheaton, L., Fridman, E., Shamim,
E. UN., Garraux, G., et autres. (2008). Gesture subtype-dependent
left lateralization of praxis planning: An event-related fMRI
étude. Cortex cérébral, 19, 1256–1262.
Berkman and Lieberman
1977
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
2
je
9
e
–
1
p
9
d
7
F
0
/
1
2
9
2
3
/
9
9
8
/
1
1
9
9
o
7
c
0
n
/
1
2
0
7
0
7
9
0
4
2
6
1
7
3
1
/
7
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
9
e
.
s
t
2
o
1
n
3
1
0
7
8
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
F
/
.
t
.
.
o
n
1
8
M.
un
oui
2
0
2
1
Canli, T., Desmond, J.. E., Zhao, Z., Glover, G., & Gabriela,
Harmon-Jones, E., & Sigelman, J.. (2001). State anger and
J.. D. (1998). Hemispheric asymmetry for emotional stimuli
detected with fMRI. NeuroReport, 9, 3233–3239.
Carver, C. S., & Harmon-Jones, E. (2009). Anger is an
approach-related affect: Evidence and implications.
Psychological Bulletin, 135, 183–204.
Carver, C. S., & Blanc, T. L. (1994). Behavioral inhibition,
behavioral activation, and affective responses to impending
reward and punishment: The BIS/BAS Scales. Journal de
Personality & Social Psychology, 67, 319–333.
Coan, J.. UN., & Allen, J.. J.. B. (2004). Frontal EEG asymmetry
as a moderator and mediator of emotion. Biological
Psychologie, 67, 7–49.
Coan, J.. UN., Allen, J.. J.. B., & Harmon-Jones, E. (2001). Voluntary
facial expression and hemispheric asymmetry over the
frontal cortex. Psychophysiology, 38, 912–925.
Cunningham, W. UN., Raye, C. L., & Johnson, M.. K. (2005).
Neural correlates of evaluation associated with promotion
and prevention regulatory focus. Cognitive, Affective, et
Behavioral Neuroscience, 5, 202–211.
Damasio, UN. R., Grabowski, T. J., Bechara, UN., Damasio, H.,
Ponto, L. L., Parvizi, J., et autres. (2000). Subcortical and
cortical brain activity during the feeling of self-generated
émotions. Neurosciences naturelles, 3, 1049–1056.
Davidson, R.. J., Ekman, P., Saron, C. D., & Senulis, J.. UN. (1990).
Approach/withdrawal and cerebral asymmetry: Emotional
expression and brain physiology: je. Journal of Personality
& Social Psychology, 58, 330–341.
Drevets, W. C., Prix, J.. L., Simpson, J.. R., Todd, R.. D., Reich, T.,
Vannier, M., et autres. (1997). Subgenual prefrontal cortex
abnormalities in mood disorders. Nature, 386, 824–827.
Eddington, K. M., Dolcos, F., Cabeza, R., Krishnan, K. R., &
Strauman, T. J.. (2007). Neural correlates of promotion
and prevention goal activation: An fMRI study using an
idiographic approach. Journal des neurosciences cognitives,
19, 1152–1162.
Elliot, UN. J.. (2006). The hierarchical model of approach–avoidance
motivation. Motivation & Émotion, 30, 111–116.
Elliot, UN. J., & Church, M.. UN. (1997). A hierarchical model of
approach and avoidance achievement motivation. Journal
of Personality & Social Psychology, 72, 218–232.
Frey, S. H. (2007). What puts the how in where? Tool use
and the divided visual streams hypothesis. Cortex, 43,
368–375.
Frey, S. H. (2008). Tool use, communicative gesture and
cerebral asymmetries in the modern human brain.
Philosophical Transactions of the Royal Society of
Londres, Série B, Sciences biologiques, 363, 1951–1957.
Fuster, J.. M.. (2008). The prefrontal cortex (4th ed.). Boston,
MA: Academic Press/Elsevier.
Georges, M.. S., Ketter, T. UN., Parekh, P.. JE., Herscovitch, P.,
& Post, R.. M.. (1996). Gender differences in regional
cerebral blood flow during transient self-induced sadness
or happiness. Biological Psychiatry, 40, 859–871.
Harmon-Jones, E., Abramson, L. Y., Sigelman, J., Bohlig, UN.,
Hogan, M.. E., & Harmon-Jones, C. (2002). Proneness to
hypomania/mania symptoms or depression symptoms and
asymmetrical frontal cortical responses to an anger-evoking
event. Journal of Personality & Social Psychology, 82,
610–618.
Harmon-Jones, E., & Allen, J.. J.. (1997). Behavioral activation
sensitivity and resting frontal EEG asymmetry: Covariation
of putative indicators related to risk for mood disorders.
Journal of Abnormal Psychology, 106, 159–163.
Harmon-Jones, E., & Allen, J.. J.. (1998). Anger and frontal
brain activity: EEG asymmetry consistent with approach
motivation despite negative affective valence. Journal de
Personality & Social Psychology, 74, 1310–1316.
prefrontal brain activity: Evidence that insult-related relative
left-prefrontal activation is associated with experienced
anger and aggression. Journal of Personality & Social
Psychologie, 80, 797–803.
Henriques, J.. B., & Davidson, R.. J.. (2000). Decreased
responsiveness to reward in depression. Cognition &
Émotion, 14, 711–724.
Herrington, J.. D., Mohanty, UN., Koven, N. S., Pêcheur, J.. E.,
Stewart, J.. L., Banich, M.. T., et autres. (2005). Emotion-modulated
performance and activity in left dorsolateral prefrontal
cortex. Émotion, 5, 200–207.
Hewig, J., Hagemann, D., Seifert, J., Naumann, E., & Bartussek,
D. (2006). The relation of cortical activity and BIS/BAS
on the trait level. Biological Psychology, 71, 42–53.
Higgins, E. T. (1997). Beyond pleasure and pain. Américain
Psychologist, 52, 1280–1300.
Jackson, D. C., Mueller, C. J., Dolski, JE., Dalton, K. M., Nitschke,
J.. B., Urry, H. L., et autres. (2003). Now you feel it, now you
donʼt: Frontal brain electrical asymmetry and individual
differences in emotion regulation. Sciences psychologiques,
14, 612–617.
Kalisch, R., Wiech, K., Critchley, H. D., Seymour, B., OʼDoherty,
J.. P., Oakley, D. UN., et autres. (2005). Anxiety reduction through
detachment: Subjective, physiological, and neural effects.
Journal des neurosciences cognitives, 17, 874–883.
Kline, J.. P., Blackhart, G. C., Woodward, K. M., Williams, S. R.,
& Schwartz, G. E. (2000). Anterior electroencephalographic
asymmetry changes in elderly women in response to a
pleasant and an unpleasant odor. Biological Psychology,
52, 241–250.
Kroliczak, G., & Frey, S. H. (2009). A common network in the
left cerebral hemisphere represents planning of tool use
pantomimes and familiar intransitive gestures at the hand-
independent level. Cortex cérébral. Epub ahead of print.
Maldjian, J.. UN., Laurienti, P.. J., Kraft, R.. UN., & Burdette, J.. H.
(2003). An automated method for neuroanatomic and
cytoarchitectonic atlas-based interrogation of fMRI data
sets. Neuroimage, 19, 1233–1239.
Mayberg, H. S., Brannan, S. K., Tekell, J.. L., Silva, J.. UN.,
Mahurin, R.. K., McGinnis, S., et autres. (2000). Regional metabolic
effects of fluoxetine in major depression: Serial changes
and relationship to clinical response. Biological Psychiatry,
48, 830–843.
Miller, UN., & Tomarken, UN. J.. (2001). Task-dependent changes
in frontal brain asymmetry: Effects of incentive cues,
outcome expectancies, and motor responses.
Psychophysiology, 38, 500–511.
Miller, E. K., & Cohen, J.. D. (2001). An integrative theory of
prefrontal cortex function. Revue annuelle des neurosciences,
24, 167–202.
Mitchell, J.. P., Heatherton, T. F., Kelley, W. M., Wyland, C. L.,
Wegner, D. M., & Neil Macrae, C. (2007). Separating
sustained from transient aspects of cognitive control
during thought suppression. Sciences psychologiques, 18,
292–297.
Morris, J.. S., Ohman, UN., & Dolan, R.. J.. (1998). Conscious and
unconscious emotional learning in the human amygdala.
Nature, 393, 467–470.
Phan, K. L., Wager, T., Taylor, S. F., & Liberzon, je. (2002).
Functional neuroanatomy of emotion: A meta-analysis of
emotion activation studies in PET and fMRI. Neuroimage,
16, 331–348.
Phillips, M.. L., Williams, L. M., Heining, M., Herba, C. M.,
Russell, T., Andrew, C., et autres. (2004). Differential neural
responses to overt and covert presentations of facial
expressions of fear and disgust. Neuroimage, 21,
1484–1496.
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
2
je
9
e
–
1
p
9
d
7
F
0
/
1
2
9
2
3
/
9
9
8
/
1
1
9
9
o
7
c
0
n
/
1
2
0
7
0
7
9
0
4
2
6
1
7
3
1
/
7
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
9
e
.
s
t
2
o
1
n
3
1
0
7
8
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
.
t
F
/
.
.
o
n
1
8
M.
un
oui
2
0
2
1
1978
Journal des neurosciences cognitives
Volume 22, Nombre 9
Pizzagalli, D. UN., Sherwood, R.. J., Henriques, J.. B., &
Davidson, R.. J.. (2005). Frontal brain asymmetry and
reward responsiveness: A source-localization study.
Sciences psychologiques, 16, 805–813.
Robinson, R.. G. (1979). Differential behavioral and biochemical
effects of right and left hemispheric cerebral infarction in
the rat. Science, 205, 707–710.
Schluter, N. D., Krams, M., Rushworth, M.. F., & Passingham,
R.. E. (2001). Cerebral dominance for action in the human
brain: The selection of actions. Neuropsychologie, 39,
105–113.
Sobotka, S. S., Davidson, R.. J., & Senulis, J.. UN. (1992).
Anterior brain electrical asymmetries in response to reward
and punishment. Electroencephalography and Clinical
Neurophysiologie, 83, 236–247.
Starkstein, S. E., Bryer, J.. B., Berthier, M.. L., Cohen, B., Prix,
T. R., & Robinson, R.. G. (1991). Depression after stroke:
The importance of cerebral hemisphere asymmetries.
Journal of Neuropsychiatry and Clinical Neurosciences,
3, 276–285.
Sutton, S. K., & Davidson, R.. J.. (1997). Prefrontal brain
asymmetry: A biological substrate of the behavioral
approach and inhibition systems. Sciences psychologiques,
8, 204–210.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D.,
Crivello, F., Etard, O., Delcroix, N., et autres. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject
brain. Neuroimage, 15, 273–289.
van Honk, J., & Schutter, D. J.. (2006). From affective valence
to motivational direction. Sciences psychologiques, 17,
963–965.
Wager, T. D., Phan, K. L., Liberzon, JE., & Taylor, S. F. (2003).
Valence, genre, and lateralization of functional brain
anatomy in emotion: A meta-analysis of findings from
neuroimaging. Neuroimage, 19, 513–531.
Whalen, P.. J., Shin, L. M., McInerney, S. C., Fischer, H.,
Wright, C. JE., & Rauch, S. L. (2001). A functional MRI study
of human amygdala responses to facial expressions of
fear versus anger. Émotion, 1, 70–83.
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
2
je
9
e
–
1
p
9
d
7
F
0
/
1
2
9
2
3
/
9
9
8
/
1
1
9
9
o
7
c
0
n
/
1
2
0
7
0
7
9
0
4
2
6
1
7
3
1
/
7
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
9
e
.
s
t
2
o
1
n
3
1
0
7
8
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
/
.
F
.
t
.
o
n
1
8
M.
un
oui
2
0
2
1
Berkman and Lieberman
1979