Alpha Oscillations during Incidental Encoding Predict
Subsequent Memory for New “Foil” Information
Donne la vie. Vogelsang1, Matthias Gruber2,3, Zara M. Bergström4,
Charan Ranganath2, and Jon S. Simons1
Abstrait
■ People can employ adaptive strategies to increase the like-
lihood that previously encoded information will be successfully
retrieved. One such strategy is to constrain retrieval toward rel-
evant information by reimplementing the neurocognitive pro-
cesses that were engaged during encoding. Using EEG, nous
examined the temporal dynamics with which constraining re-
trieval toward semantic versus nonsemantic information affects
the processing of new “foil” information encountered during a
memory test. Time–frequency analysis of EEG data acquired
during an initial study phase revealed that semantic compared
with nonsemantic processing was associated with alpha de-
creases in a left frontal electrode cluster from around 600 msec
after stimulus onset. Successful encoding of semantic versus
nonsemantic foils during a subsequent memory test was related
to decreases in alpha oscillatory activity in the same left frontal
electrode cluster, which emerged relatively late in the trial at
around 1000–1600 msec after stimulus onset. Across partici-
pants, left frontal alpha power elicited by semantic processing
during the study phase correlated significantly with left frontal
alpha power associated with semantic foil encoding during the
memory test. En outre, larger left frontal alpha power de-
creases elicited by semantic foil encoding during the memory
test predicted better subsequent semantic foil recognition in
an additional surprise foil memory test, although this effect did
not reach significance. These findings indicate that constraining
retrieval toward semantic information involves reimplementing
semantic encoding operations that are mediated by alpha oscil-
lations and that such reimplementation occurs at a late stage
of memory retrieval, perhaps reflecting additional monitoring
processes. ■
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
un
_
0
1
2
3
4
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
Memory retrieval often requires goal-directed control pro-
cesses to optimize retrieval success. One possible strategy
people use to facilitate memory retrieval is to reimplement
the neurocognitive processes that were involved during
encoding (Rugg, Johnson, Parc, & Uncapher, 2008). Ce
idea has been manifested in the Transfer Appropriate
Processing Framework, which states that the likelihood
of retrieval success is dependent on the overlap between
encoding and retrieval operations (Roediger, Weldon, &
Challis, 1989; Morris, Bransford, & Franks, 1977). Accord-
franchement, the most effective encoding strategy depends on
the specific conditions at retrieval, and conversely, what
constitutes an optimal retrieval strategy depends on the
conditions under which the information was encoded
(Rugg et al., 2008). In line with this principle, previous
behavioral studies have shown that recognition memory
is enhanced when mental operations at encoding are re-
capitulated during a subsequent memory test (Dewhurst
& Brandt, 2007; Morris et al., 1977).
Jacoby, Shimizu, Daniels, and Rhodes (2005) developed
a behavioral “memory for foils” paradigm that enabled
1University of Cambridge, 2University of California at Davis,
3Cardiff University, 4University of Kent
such encoding–retrieval overlap to be investigated more
directly. In an initial study phase (Phase 1 of the para-
digm), participants studied nouns in two separate blocks,
one of which involved a semantic task (pleasant/unpleas-
ant judgment) and the other involved a nonsemantic task
(letter judgment). In a subsequent recognition memory
test (Phase 2), studied and nonstudied words were inter-
mixed, and participants undertook blocks in which they
judged whether they had previously encountered the
words in the pleasantness judgment task or whether they
were new (the memory test for semantically encoded
mots), or judged whether they had previously seen the
words in the letter judgment task or whether they were
nouveau (the memory test for nonsemantically encoded words).
Of special interest were the new words (so called “foils”)
in the semantic and nonsemantic memory test blocks. Le
semantic and nonsemantic foils were subsequently mixed
together with completely new words in a final foil recog-
nition test (Phase 3) in which participants were again in-
structed to make an old/new judgment, this time about
whether the words had been encountered at any time dur-
ing the experiment or were completely novel. Jacoby et al.
found that the “foil” words were differentially memorable
depending on the type of test in which they had been orig-
inally encountered: Recognition memory was significantly
© 2018 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 unported (CC PAR 4.0) Licence.
Journal des neurosciences cognitives 30:5, pp. 667–679
est ce que je:10.1162/jocn_a_01234
higher for semantic compared with nonsemantic foils.
Because semantic encoding typically leads to more accu-
rate memory compared with nonsemantic encoding, ce
“foil effect” implies that participants strategically orient
their retrieval toward a semantic processing mode when
attempting to retrieve semantic encoded information and
a nonsemantic processing mode when retrieving non-
semantic information, resulting in better incidental encod-
ing of semantic compared with nonsemantic foils. Jacoby
and colleagues interpreted this foil finding in light of the
transfer appropriate processing principle by emphasiz-
ing the importance of the overlap in study–test opera-
tions for optimizing retrieval success (see also Zawadzka,
Hanczakowski, & Wilding, 2017; Gray & Gallo, 2015; Kantner
& Lindsay, 2013; Alban & Kelley, 2012; Halamish, Goldsmith,
& Jacoby, 2012; Danckert, MacLeod, & Fernandes, 2011;
Marsh et al., 2009).
Recently, we collected fMRI data in a “memory for
foils” paradigm and applied subsequent memory analysis
(also known as “difference due to memory” or “DM effect”)
to study the neural mechanisms underlying the enhanced
encoding of foils in a semantic compared with nonsemantic
recognition test. The results revealed that the left inferior
frontal gyrus (LIFG) exhibited significantly greater sub-
sequent memory effects for semantic compared with non-
semantic foils (Vogelsang, Bonnici, Bergström, Ranganath,
& Simons, 2016). A conjunction analysis revealed signifi-
cant overlap in activity between semantic processing in
the initial study phase and semantic foil encoding during
the first memory retrieval test in the LIFG; cependant, ce
overlap in activation was not observed for the nonsemantic
condition. The LIFG has previously been associated with
semantic processing and semantic encoding strategies
across many studies (Kim, 2011; Fletcher, Stephenson,
Carpenter, Donovan, & Bullmorel, 2003; Poldrack et al.,
1999; Wagner et al., 1998). Together with the behavioral
result that semantic foils were recognized more accurately
than nonsemantic foils on the final surprise foil recogni-
tion test, these neuroimaging data support the hypothesis
that directing retrieval toward new semantic versus non-
semantic information leads to the recruitment of distinct
neural mechanisms that are predictive of subsequent
mémoire (Vogelsang et al., 2016).
A key element of the foil paradigm is the proposal that
retrieval is strategically oriented toward the relevant pro-
cessing mode to facilitate memory search before informa-
tion is retrieved. It has been hypothesized that there are
two ways that control can be exerted over retrieval: “front-
end” and “back-end” processes. Front-end control processes
are engaged shortly after a memory cue is encountered to
guide retrieval attempts ( Jacoby, Shimizu, Velanova, &
Rhodes, 2005). Alternativement, strategic control processes
can also be recruited later on in the trial when retrieval
attempts have failed or have produced ambiguous infor-
mation and additional monitoring or verification is re-
demandé, which has been referred to as back-end control
(Halamish et al., 2012) or “late correction strategy” (Jacoby,
Kelley, & McElree, 1999). Front-end control processes are
similar to the concept of retrieval orientation, which refers
to the type of processing participants engage in when they
are prompted with a retrieval cue to increase the likelihood
of retrieval success (Rugg & Wilding, 2000). Cependant,
when such early control processes result in retrieved in-
formation that is vague or ambiguous, participants may
engage back-end control processes, correcting their strat-
egy, to increase the likelihood of recollecting accurately
more contextual details about the encoded event ( Jacoby
et coll., 1999). Previous fMRI research was unable to distin-
guish “front-end” and “back-end” accounts ( Vogelsang
et coll., 2016) because the low temporal resolution of the
BOLD precludes investigation of at which stage of retrieval
(early vs. late) LIFG activity is reinstated for semantic com-
pared with nonsemantic foils. Donc, in the current
étude, we recorded EEG oscillations during performance
of the “memory for foils” paradigm. The fine-grained tem-
poral resolution of neural oscillations can provide more
information with regard to the question of “when” the
neural activity associated with initial encoding operations
during a study phase reoccur during the incidental encod-
ing of foils in a subsequent recognition test.
Neural oscillations and their relationship with memory
functions have gained considerable interest in recent years
(Fell & Axmacher, 2011). In the memory encoding litera-
ture, there is evidence that a decrease in alpha power
might be related to semantic processing (Hanslmayr &
Staudigl, 2014; Zion-Golumbic, Kutas, & Bentin, 2010;
Hanslmayr, Spitzer, & Bäuml, 2009; Bastiaansen, van der
Linden, ter Keurs, Dijkstra, & Hagoort, 2005; for a review,
see Klimesch, 1999). Par exemple, Hanslmayr and col-
leagues (2009) contrasted deep semantic encoding with
shallow nonsemantic encoding and found power de-
creases in alpha (and beta) frequency bands that were
related to successful semantic encoding. Fellner, Bäuml,
and Hanslmayr (2013) showed that alpha and beta de-
creases predicted subsequent memory in a semantic
condition, but not in a nonsemantic but still highly effi-
cient encoding condition (in this case, a survival process-
ing task), thereby suggesting that alpha decreases are
likely a reflection of semantic processing in particular,
rather than of efficient encoding strategies in general.
En outre, alpha decreases have been observed over
left frontal electrodes in tasks requiring high semantic pro-
cessing demands (Hanslmayr & Staudigl, 2014; Klimesch,
1999) but have also been associated with subsequent
memory effects (Klimesch, Doppelmayr, Schimke, &
Ripper, 1997), consistent with the idea that the left prefron-
tal cortex is important for successful encoding (Vogelsang
et coll., 2016; Wagner et al., 1998).
The main aim of the present experiment was to inves-
tigate the temporal dynamics of EEG oscillations asso-
ciated with encoding of new “foil” words during a memory
retrieval test. We focused our analysis on alpha EEG fre-
quencies (8–10 Hz) because previous research has shown
that alpha plays a role in both semantic processing (Bakker,
668
Journal des neurosciences cognitives
Volume 30, Nombre 5
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
un
_
0
1
2
3
4
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Takashima, van Hell, Janzen, & McQueen, 2015) and sub-
sequent memory effects (Hanslmayr et al., 2009). We first
examined alpha power associated with semantic versus
nonsemantic processing during the initial study phase.
We then investigated whether alpha power differences
were again observed during successful encoding of seman-
tic versus nonsemantic foils in the first memory test, lequel
would support the hypothesis that the incidental encod-
ing of foils in a memory test involves the reimplemen-
tation of the neurocognitive processes that were involved
during initial encoding (Vogelsang et al., 2016; Bergström,
Vogelsang, Benoit, & Simons, 2015; Jacoby, Shimizu,
Daniels, et coll., 2005; Jacoby, Shimizu, Velanova, et coll., 2005).
Le plus important, the high temporal resolution of EEG
oscillations allowed us to examine whether alpha re-
instatement during foil encoding occurred early or late in
the trial, which we hypothesized would indicate that par-
ticipants used “front-end” or “back-end” control strategies,
respectivement. We also tested whether those individuals
who showed the largest alpha power differences during
semantic versus nonsemantic processing in the study
phase also showed the largest alpha power differences
during semantic encoding of foils in the retrieval test,
which would support the hypothesis that the alpha effects
during study and test were functionally related. Enfin, nous
tested the hypothesis that if alpha frequencies mediate
semantic foil encoding during the first recognition test,
then individuals who showed larger alpha differences for
successfully encoded foils during the first test should also
exhibit better foil recognition performance in the final foil
recognition test.
MÉTHODES
Participants
Fifty-three right-handed healthy English native speakers
with normal or corrected-to-normal vision participated
in this experiment. Written informed consent was ob-
tained before commencement of the experiment, et
all participants received £15 for their participation. Données
depuis 17 participants were excluded because they did not
produce enough trials of each type for the subsequent
memory analysis (see “Time–Frequency Analysis” section
for details). En outre, data from two participants were
excluded because of excessively noisy EEG data. The final
data set thus consisted of 34 participants (21 femmes, mean
age = 21.9 années, range 18–33 years). The study was
approved by the University of Cambridge psychology
research ethics committee.
Materials
The stimuli consisted of 552 nouns (par exemple., “book”) derived
from the MRC psycholinguistic database ( Wilson, 1988;
also used in Vogelsang et al., 2016). Le 552 words were
split into six lists that were matched for concreteness,
familiarity, Kucera–Francis frequency, word length, et
number of syllables, and we counterbalanced the as-
signment of lists to the experimental conditions across
participants.
Procedure
Participants were fitted with an EEG cap and were seated
in a sound- and light-attenuated room. Participants com-
pleted three phases: (1) a study phase (henceforth re-
ferred to as “Phase 1”), (2) an initial memory test
(henceforth referred to as “Phase 2”), et (3) a final sur-
prise memory test that assessed foil recognition (hence-
forth referred to as “Phase 3”). Throughout all phases,
participants provided their responses on a button box
with either their left or right hand (counterbalanced
across participants).
Phase 1 consisted of two separate incidental encoding
blocks during which participants were instructed to make
a simple judgment. In a semantic judgment study block,
participants made a pleasantness judgment (“Is this word
pleasant?»). In a nonsemantic study block, participants
made a letter judgment (“Is there a letter O or U in the
word?»). Each trial in the study phase started with a 500-msec
fixation cross followed by the stimulus that was presented
in the center of the screen for 2000 msec. Both the seman-
tic and nonsemantic judgment blocks consisted of 92 trials
chaque. The order of semantic and nonsemantic judgment
blocks was counterbalanced across participants. Partici-
pants were instructed to respond while the words were
on the screen.
In Phase 2, participants’ memory for the stimuli en-
countered during Phase 1 was assessed in an old/new
recognition test, which aimed to manipulate whether
participants oriented retrieval toward semantic or non-
semantic information. In the semantic test phase, 92 vieux
words from the semantic study phase were intermixed with
92 new words (semantic foils). In the nonsemantic test
phase, 92 old words from the nonsemantic study phase
were intermixed with 92 new words (nonsemantic foils).
For both blocks, participants were told in which specific
Phase 1 task any old words had been shown to encourage
them to engage different retrieval orientations for the two
blocks. The order of test block (semantic and nonseman-
tic) was counterbalanced across participants. Each test
trial began with a 500-msec fixation cross, suivi de
the presentation of the stimulus centrally on the screen
pour 2000 msec. Participants were instructed to provide
their response as to whether each word was old or new
while the stimulus was still on the screen.
In the final phase, a surprise old/new foil recognition
test (Phase 3) was administered in which participants
were asked to distinguish between the semantic and non-
semantic foils and completely new words. Participants
were instructed that they were “going to be presented
with a word that is either old or new. ‘Old’ in this case
means that you saw the word at some point earlier in
Vogelsang et al.
669
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
un
_
0
1
2
3
4
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
the experiment in any study or test phase. ‘New’ words
are words you have not seen at all in today’s experiment.”
This foil recognition test consisted of 368 mots (92 seman-
tic foils, 92 nonsemantic foils, et 184 completely new
mots, which were randomly intermixed). Each trial in
the final foil recognition test began with a 500-msec fixa-
tion cross followed by the stimulus presented centrally for
2000 msec.
EEG Recording and Preprocessing
EEG data were acquired during all phases of the experi-
ment and were recorded with an Electrical Geodesic
(Eugene, OR) Netamps 200 system with a 128-channel
HydroCel Geodesic Sensor Net (GSN 200; Tucker, 1993).
The recorded EEG data were referenced to Cz and were
filtered with a bandwidth of 0.01–70 Hz (250 Hz sampling
rate). The EEG data were analyzed in EEGLab 13 (Delorme
& Makeig, 2004). The continuous EEG data from the study
phase and first retrieval test were rereferenced to an aver-
age mastoid reference and high-pass filtered with a cutoff
de 0.5 Hz (two-way least-squares finite impulse response fil-
ter), and the continuous data were divided into epochs
ranging from −500 msec before cue onset until 2000 msec
thereafter. Artifact correction was applied using extended
info-max independent component analysis (Cloche &
Sejnowski, 1995, in Delorme & Makeig, 2004) using Runica
from the EEGLab toolbox, with default mode training pa-
rameters (Delorme & Makeig, 2004). Independent compo-
nents reflecting eye movements and other sources of noise
were identified by visual inspection of component scalp
topographies, time courses, and activation spectra and
were discarded from the data by back-projecting all but
these components to the data space. Trials that still con-
tained artifacts after running independent component anal-
ysis correction were removed after visual inspection,
resulting in only 5–10% of the trials being excluded.
Time–Frequency Analysis
Time–frequency analysis in EEGLab was applied using
Morlet wavelets (Percival & Walden, 1993) with six cycles
in a frequency range of 4–30 Hz, with steps of 1 Hz be-
tween each wavelet center frequency. These wavelets
were applied in a sliding window with 20-msec increments
in the 0–2000 msec interval. To minimize edge effects (dis-
tortions that occur at the edge of the time window of
analyse), we concatenated mirrored (c'est à dire., time inverted)
segments at the left and right edge of the original epochs.
We then performed the wavelet transform on these ex-
tended epochs and discarded the concatenated segments
from the final analysis (for detailed explanation of this
“reflection approach,” see Cohen, 2014; see Fell et al.,
2011, for example of a paper using this approach). Le
results reported here were not corrected for baseline in
the frequency domain because we were interested in oscil-
latory differences between the semantic and nonsemantic
conditions, and therefore, baseline correction is not nec-
essary (voir, for similar approaches, Gruber, Watrous,
Ekstrom, Ranganath, & Otten, 2013).
To identify the neural oscillations associated with se-
matic and nonsemantic processing, we first examined
the power spectra of epoched data from Phase 1. Pour
each of the 34 participants, EEG data during the study
phase were binned according to the type of processing
(semantic vs. nonsemantic). In this way, we could isolate
the EEG frequencies that were elicited by semantic and
nonsemantic processing to later examine whether these
frequencies were reinstated during the encoding of foils
in the first test phase (Phase 2). Mean trial numbers were
the following: semantic study mean = 90, range 46–92;
nonsemantic study mean = 92, range 88–92.
To analyze the neural oscillations during Phase 2, nous
binned the EEG data for each participant according to
condition (semantic vs. nonsemantic) and subsequent
mémoire (remembered vs. forgotten). Mean trial numbers
for each condition were as follows: semantic foils remem-
bered mean = 65, range 25–81; semantic foils forgotten
mean = 25, range 12–67; nonsemantic foils remembered
mean = 55, range 15–79; and nonsemantic foils forgot-
ten mean = 35, range 13–77.
Time–frequency analysis was conducted on EEG that
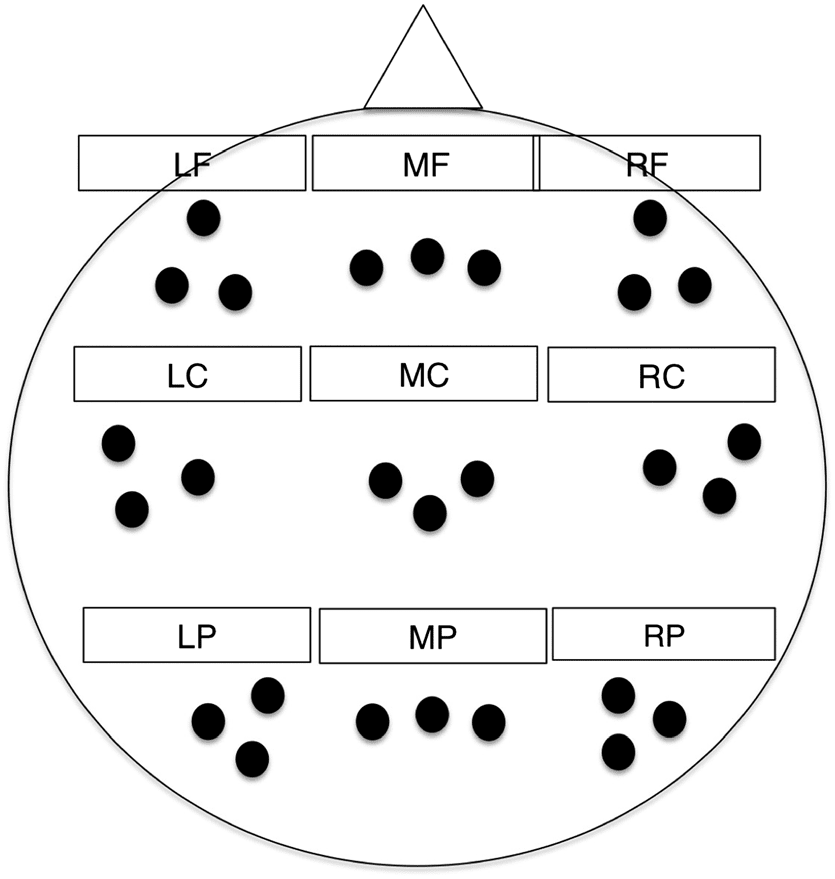
was averaged within nine electrode clusters (frontal vs.
central vs. posterior; left vs. middle vs. droite; voir la figure 1)
based on a previous study by Hsieh, Ekstrom, and Ranganath
(2011). These clusters included the following: left frontal
cluster (Channels 33, 24, 26; equivalent to F3, F7, AF7),
mid frontal cluster (Channels 19, 11, 4; equivalent to Fz,
F1, F2), right frontal cluster (Channels 124, 2, 122; equivalent
Chiffre 1. Locations of the electrode clusters, which included left
frontal (LF), mid frontal (MF), right frontal (RF), left central (LC), mid
central (MC), right central (RC), left posterior (LP), mid posterior (MP),
and right posterior (RP).
670
Journal des neurosciences cognitives
Volume 30, Nombre 5
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
un
_
0
1
2
3
4
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Tableau 1. Accuracy (Hits and FAs) and RT (for Hits and Correct Rejections) for Phase 2
Hits
FA
RT (msec)
Mean
SD
Mean
SD
Hits (Mean) Hits (SD)
Correct Rejections (Mean)
Correct Rejections (SD)
Semantic
Nonsemantic
0.88
0.50
0.07
0.15
0.13
0.26
0.11
0.13
890
981
113
138
938
982
117
161
to F4, F8, AF8), left central cluster (Channels 35, 36, 41;
equivalent to C5, C3, T7), mid central cluster (Channels
31, 55, 80; equivalent to Cz, C1, C2), right central cluster
(Channels 109, 104, 110; equivalent to C4, C6, T8), gauche
posterior cluster (Channels 52, 53, 60; equivalent to P3,
P1, PO3), mid posterior cluster (Channels 61, 62, 78;
equivalent to CP1, Pz, CP2), and right posterior cluster
(Channels 85, 86, 92; equivalent to P2, P4, PO4).
Statistical Analysis
Across-trial permutation tests were conducted to test for
significant effects in alpha power related to the semantic
versus nonsemantic processing in Phase 1 and successful
encoding of semantic versus nonsemantic foils in Phase 2.
For both phases, the 2000-msec epoch was split into time
windows of 200 msec each.
The permutation testing procedure consisted of three
steps. D'abord, for Phase 1, permutation testing was con-
ducted on the mean power alpha (8–10 Hz) frequencies
per condition for each time window and electrode cluster
(see Gruber et al., 2013, for similar procedure). We first
conducted two-tailed paired t tests on the relevant elec-
trode clusters comparing the two conditions. Deuxième, le
two conditions were then interchanged randomly for
each participant and each randomization run, so that
pseudoconditions were created in which systematic dif-
ferences between the conditions were eliminated. Pour
each permutation, we selected the largest negative and
positive t value across all nine electrode clusters. Ce
step was repeated 1000 times so that a null distribution
of largest negative and positive t values across all clusters
was created and the two tails of the null t distribution
were used as critical t values. Using an alpha level of .05
avec 1000 permutations, we used the 25th and 975th
values to represent the critical t values, and values below
or higher than these values were considered to be signifi-
cant effects. This stringent permutation method corrects
for multiple comparisons and was based on Blair and
Karniski (1993; for similar approaches, see Gruber et al.,
2013; Addante, Watrous, Yonelinas, Ekstrom, & Ranganath,
2011; Staudigl, Hanslmayr, & Bäuml, 2010; Hanslmayr
et coll., 2009).
For Phase 2, we conducted the permutation testing
only in those electrode clusters that showed a significant
alpha effect in Phase 1. Ainsi, the electrode clusters that
showed a significant effect in Phase 1 were taken as “elec-
trode clusters of interest” for the analysis for Phase 2
data, to examine alpha activity reimplementation during
encoding of foils in the same electrode clusters that
showed significant alpha effects in Phase 1. Pour tester notre
hypothesis that reimplementation of semantic processes
facilitate successful encoding of foils, we focused the
Phase 2 analysis on the interaction between condition
(semantic vs. nonsemantic) and subsequent memory in
Phase 2 (remembered vs. forgotten) by comparing the
difference between remembered and forgotten semantic
foils versus the difference between remembered and
forgotten nonsemantic foils. We also tested the simple
effects of subsequent memory for semantic versus non-
semantic conditions separately. The rest of the permuta-
tion procedure was the same as for the Phase 1 data.
RÉSULTATS
Behavioral Results
Recognition accuracy for Phase 2 was calculated using the
discrimination measure p(Hits) − p(False alarms [FA])
(Snodgrass & Corwin, 1988) and the results are presented
in Table 1. Recognition memory for semantic trials was
significantly more accurate compared with nonsemantic
trials (t(33)= 25.4, p < .001, 95% CI [0.47, 0.56], Cohen’s
Dz = 4.4). Furthermore, RTs were faster for old semantic
items compared with old nonsemantic items (t(33) =
4.39, p < .001, 95% CI [49, 134], Cohen’s Dz = 0.75). Foils
presented in the semantic condition were also endorsed
as new significantly more quickly than foils presented in
the nonsemantic condition (t(33) = 2.23, p = .033, 95%
CI [4, 84], Cohen’s Dz = 0.38).
The behavioral results of Phase 3 are presented in
Table 2. Note that we conducted the Phase 3 analysis on
accuracy scores (proportion correct) rather than Hits–FAs
because in the final foil recognition test completely new
Table 2. Accuracy (Proportion Correct) and RT for Phase 3
Accuracy
RT (msec)
Mean
0.72
0.61
0.77
SD
0.16
0.16
0.12
Mean
887
910
921
SD
122
118
127
Semantic foils
Nonsemantic foils
New items
Vogelsang et al.
671
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
a
_
0
1
2
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
a
_
0
1
2
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
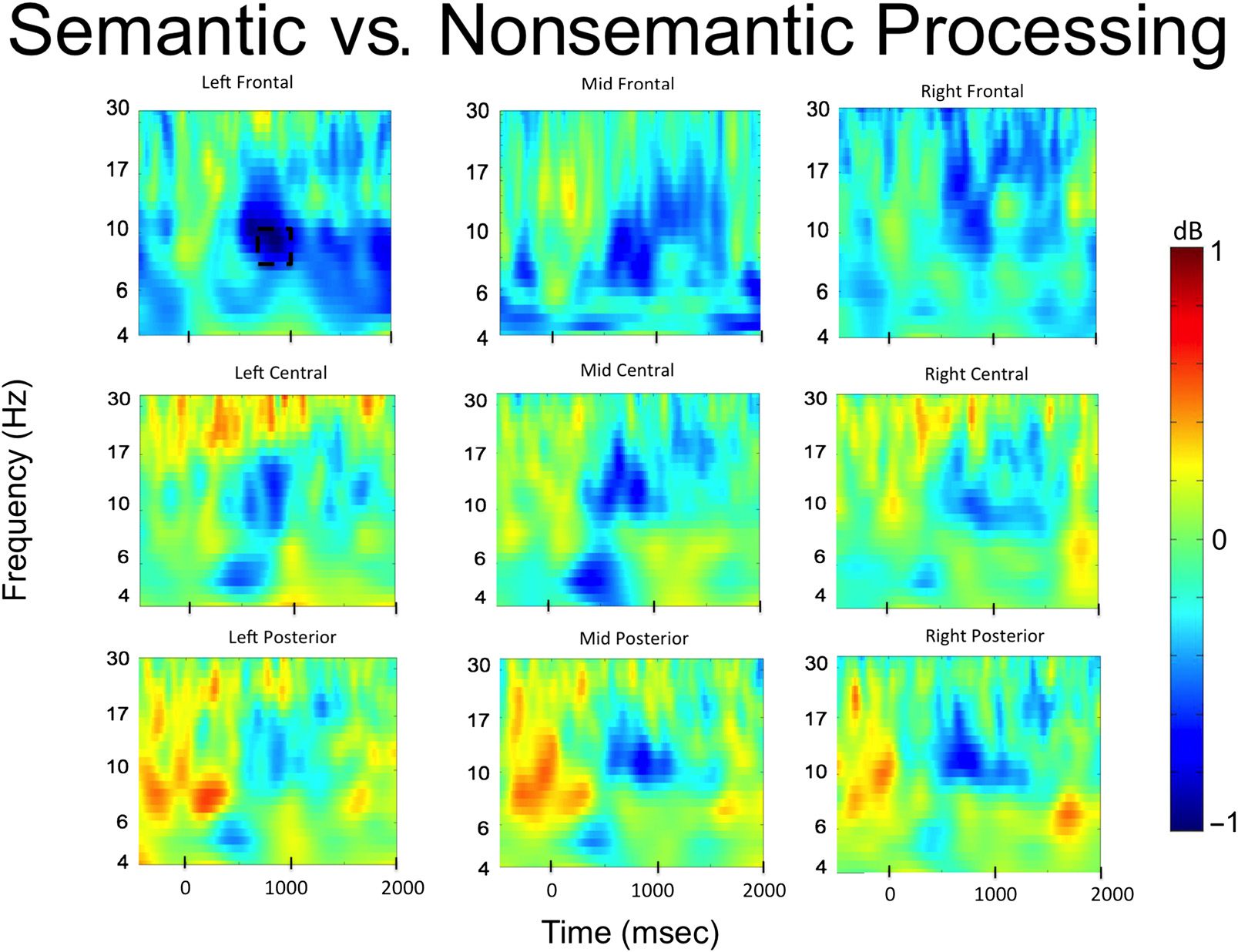
Figure 2. Time–frequency plots for semantic versus nonsemantic processing in the study phase. Significant decreases in alpha frequencies were
observed in left frontal and mid and right posterior electrode sites. The dotted boxes represent the significant alpha time window.
items were intermixed with semantic and nonsemantic
foils, so a proper Hits–FAs analysis cannot be conducted
here. In line with our main prediction, semantic foils were
recognized significantly more accurately than nonsemantic
foils (t(35) = 5.18, p < .001, 95% CI [0.066, 0.15], Cohen’s
Dz = 0.89) and significantly faster (t(33) = 3.42, p = .002,
95% CI [9, 37], Cohen’s Dz = 0.59). There was no signifi-
cant difference in RT between nonsemantic foils and new
items (t(33) = 1.5, p = .15); however, RT was faster for rec-
ognizing semantic foils compared with new items (t(33) =
4.03, p < .001, 95% CI [17, 52], Cohen’s Dz = 0.69). These
results replicate earlier findings of the “foil effect” obtained
in previous studies ( Vogelsang et al., 2016; Bergström
et al., 2015; Jacoby, Shimizu, Daniels, et al., 2005; Jacoby,
Shimizu, Velanova, et al., 2005).
Time–Frequency Results
Phase 1: Semantic versus Nonsemantic Processing
The time–frequency analysis of Phase 1 focused on a direct
comparison between all semantic and all nonsemantic trials.
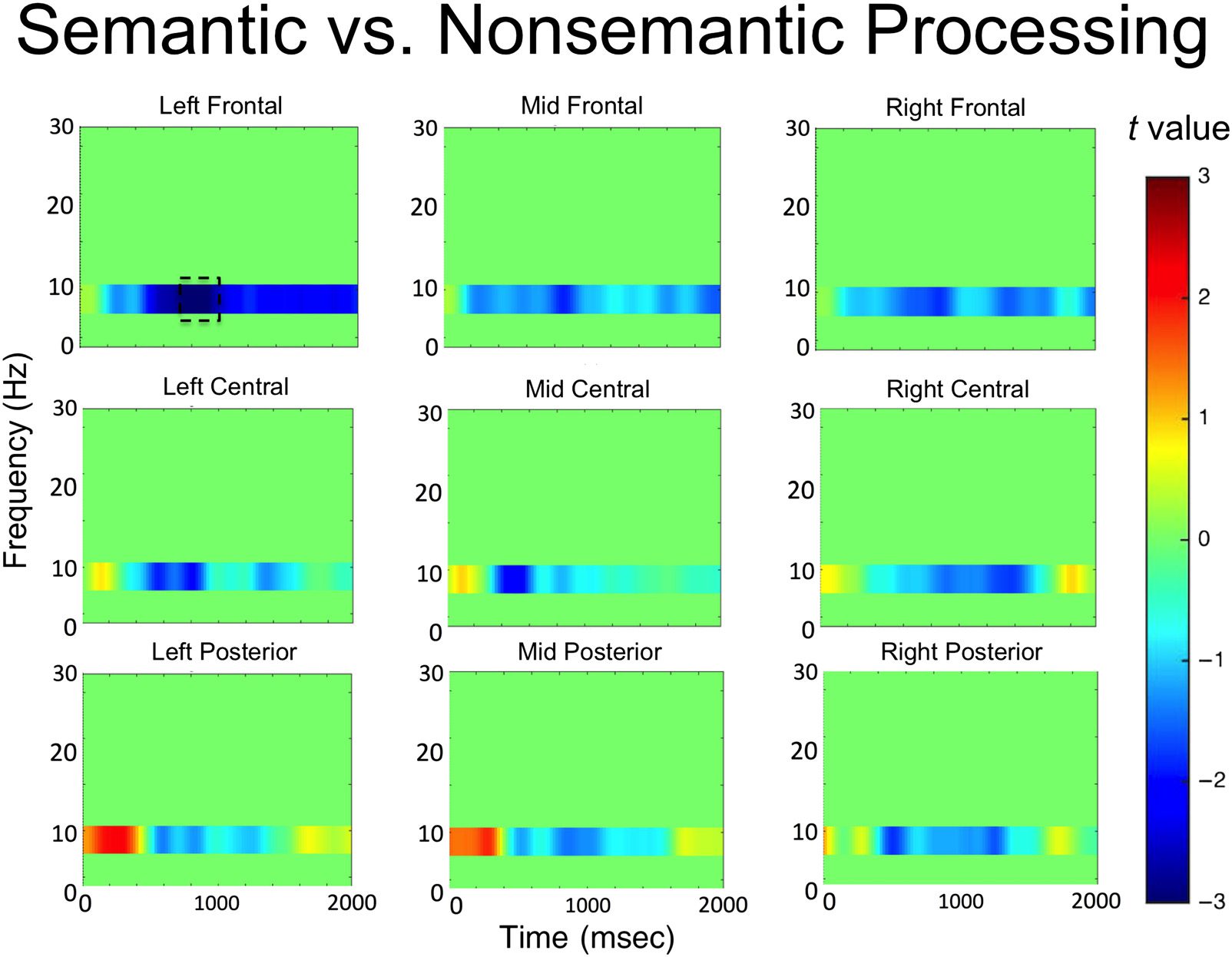
The results are presented in Figure 2. Significant decreases
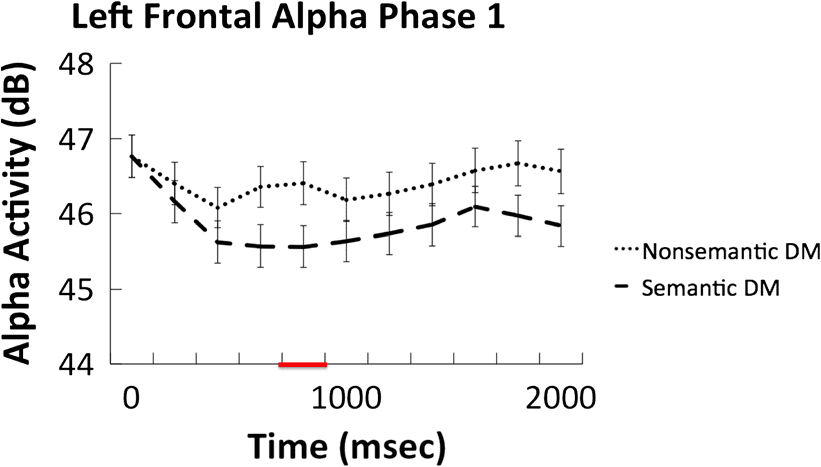
in alpha power were observed over the left frontal elec-
trode cluster between 800 and 1000 msec after word
onset (t(33) = −3.13, p = .004, 95% CI [−1.4, −0.3],
Cohen’s Dz = 0.54). No other significant effects were ob-
served in any of the other electrode clusters. The time
course of the alpha power changes in the left frontal elec-
trode cluster is presented in Figure 3, and the t values in
the alpha band across all electrode clusters is presented
in Figure 4.
Subsequent Memory Effect for Foils in Phase 2
The second part of the time–frequency analysis focused
on the temporal dynamics of subsequent memory effects
(DM effect) for foils during Phase 2 to investigate when
alpha activity was reinstated in a way that facilitated
Figure 3. Average alpha (8–10 Hz) frequency–time courses (in decibel)
for semantic and nonsemantic processing in the left frontal cluster.
Red line on x axis represents significant time window.
672
Journal of Cognitive Neuroscience
Volume 30, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
a
_
0
1
2
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. t values for semantic versus nonsemantic processing in all electrode clusters.
encoding of semantic foils. The time–frequency plot of
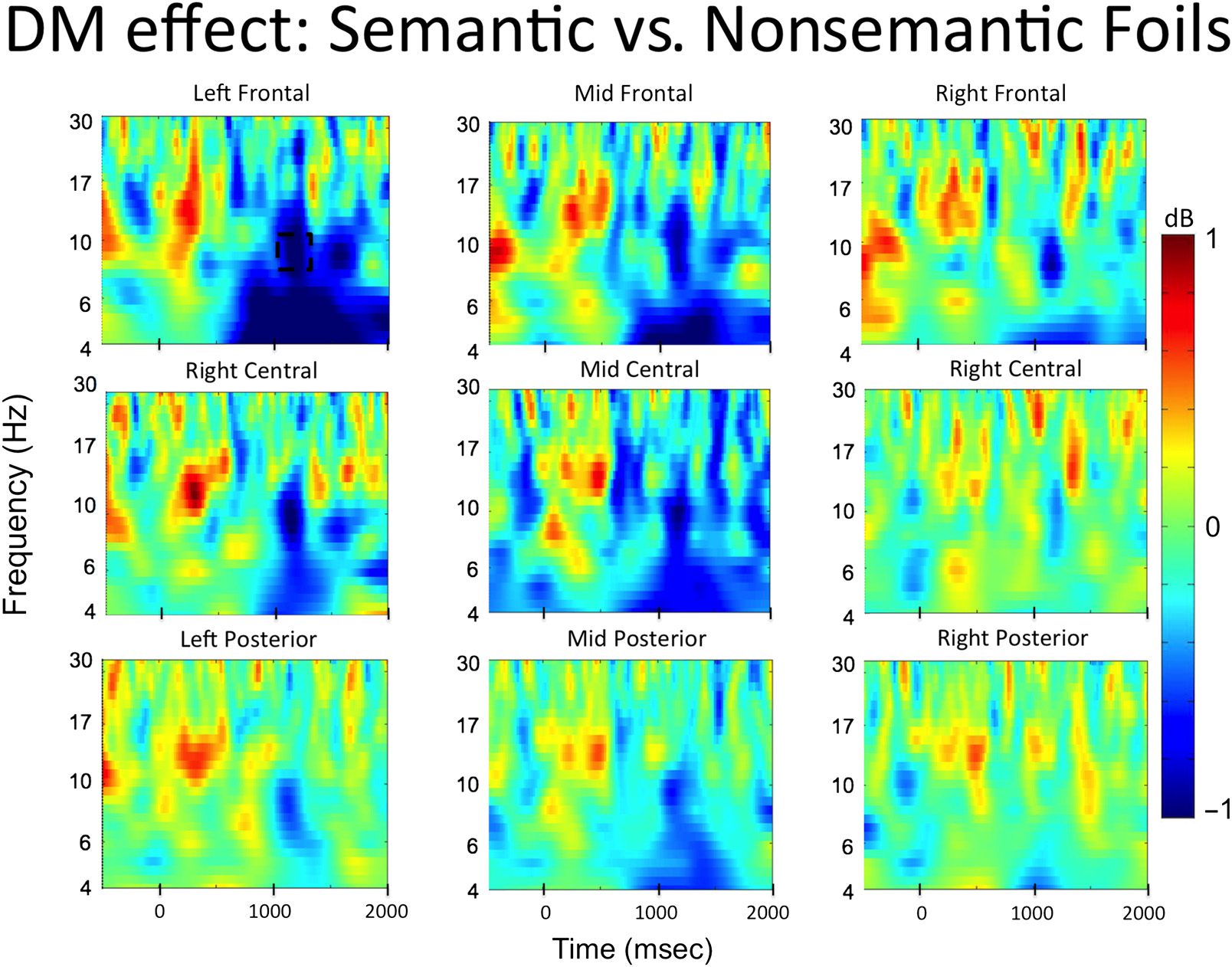
the interaction term (DM effect for semantic foils − DM
effect for nonsemantic foils) for all electrode clusters is
presented in Figure 5. Because significant alpha effects in
Phase 1 were only observed in the left frontal electrode
cluster, only this cluster was used to conduct the permuta-
tion analysis in Phase 2, which allowed us to directly test
the reimplementation hypothesis.
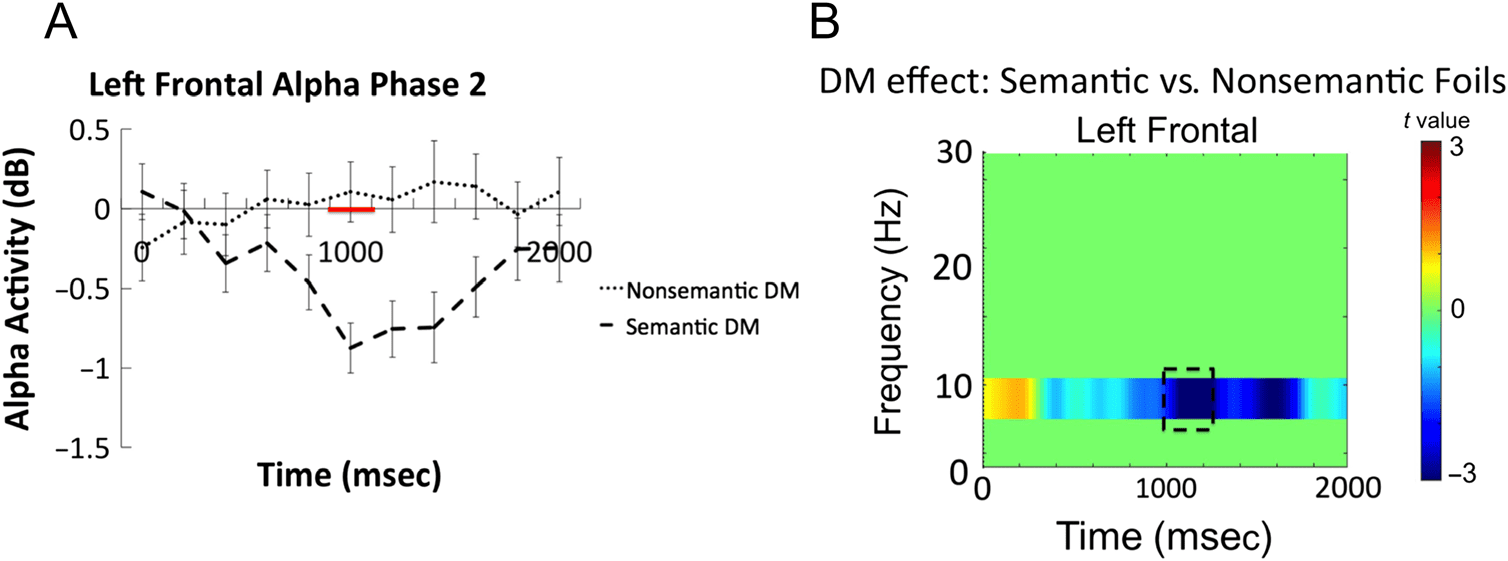
The left frontal electrode cluster showed a significant
interaction in the alpha band between 1000 and 1200 msec
after word onset (t(33) = −3.57, p = .001, 95% CI [−1.54,
−0.42], Cohen’s Dz = 0.61; see dashed box in Figure 5).
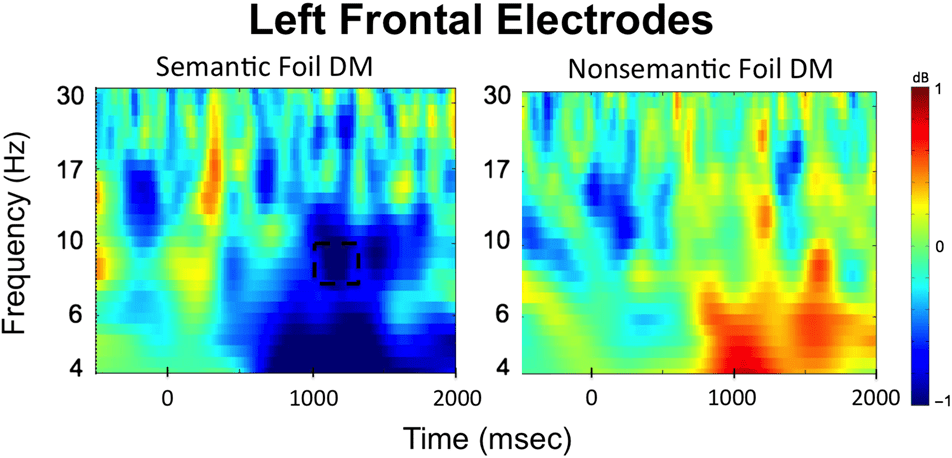
Time–frequency plots comparing EEG oscillations asso-
ciated with successful encoding of each type of foils sepa-
rately are presented in Figure 6, and the time courses for
alpha frequencies in the left frontal cluster for the seman-
tic and nonsemantic subsequent memory effect as well as
the t values for the left frontal electrode cluster are pre-
sented in Figure 7. These comparisons indicated that
the significant interaction arose because power differences
between remembered and forgotten items were observed
in the semantic but not in the nonsemantic condition. For
successful encoding of semantic foils, alpha in the 1000–
1200 msec time window (t(33) = −4.19, p = .0001, 95%
CI [−1.30, −0.45], Cohen’s Dz = 0.72) power was sig-
nificant depending on whether a word was later remem-
bered or forgotten. However, no significant differences
between remembered and forgotten words were observed
for nonsemantic foils (1000–1200 msec alpha: t(33) = 0.72,
p = .47, 95% CI [−0.19, 0.40], Cohen’s Dz = 0.12). Thus,
the subsequent memory effects observed here became
apparent over left frontal electrodes around 1000 msec after
stimulus presentation, which is at a relatively late stage in
the trial.
Relationship between Phase 1 and Phase 2
Alpha Frequencies
The third part of the analysis assessed the relationship
between alpha decreases in Phase 1 during semantic versus
nonsemantic processing and alpha decreases in Phase 2
during semantic (subsequently remembered vs. forgotten)
foil encoding to test whether these effects were func-
tionally related, which would support the hypothesis that
the neurocognitive processes engaged during initial en-
coding are reimplemented when attempting to retrieve
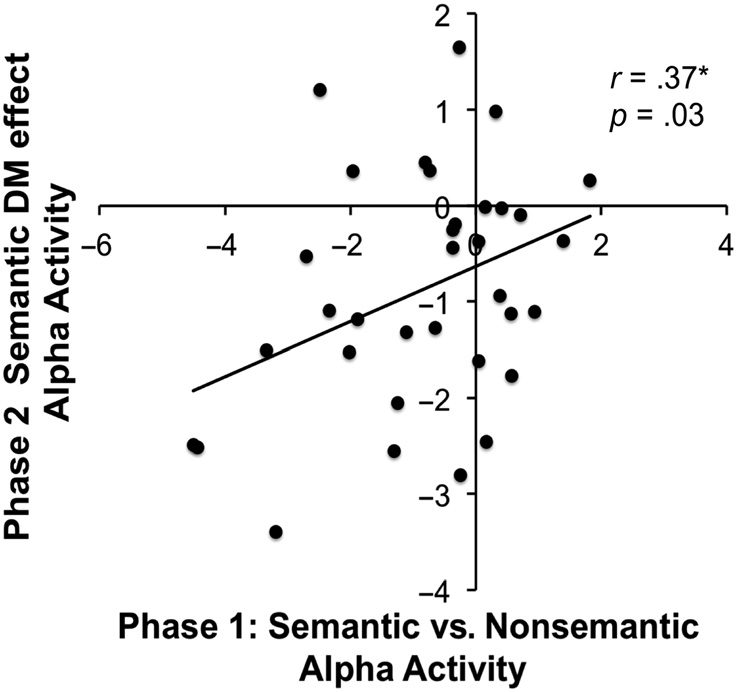
information. To examine the relationship between Phase 1
and Phase 2 alpha activity, we extracted the mean alpha
signal that showed a significant effect in the Phase 1 (800–
1000 msec) and Phase 2 (1000–1200 msec) time windows
for each participant and conducted an across-participant
Pearson correlation. A Pearson correlation is appropriate
here because the time–frequency data were normally
distributed.
There was a significant correlation (r = 0.37, p = .03;
two-tailed) between Phase 1 alpha activity associated
Vogelsang et al.
673
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
a
_
0
1
2
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Time–frequency plots from the left frontal cluster illustrating EEG oscillation differences associated with successful encoding (DM
effect) of semantic versus nonsemantic foils during the first memory test (Phase 2). The plot represents the difference of semantic foils later
remembered–forgotten versus nonsemantic foils later remembered–forgotten. Permutation testing conducted on the left frontal electrode cluster
(indicated by black squares) revealed a significant alpha decrease for subsequently remembered versus forgotten semantic versus nonsemantic
foils in the left frontal cluster. The dotted box represents the significant alpha time window that survived correction for multiple
comparisons.
with semantic versus nonsemantic processing and Phase 2
alpha activity associated with semantic subsequent memory
effect, indicating that those participants who showed a
large alpha effect in Phase 1 also showed an alpha effect
for semantic foil encoding in Phase 2. The results of the
correlation analysis are presented in Figure 8.
Relationship between Alpha Frequencies and
Subsequent Foil Recognition
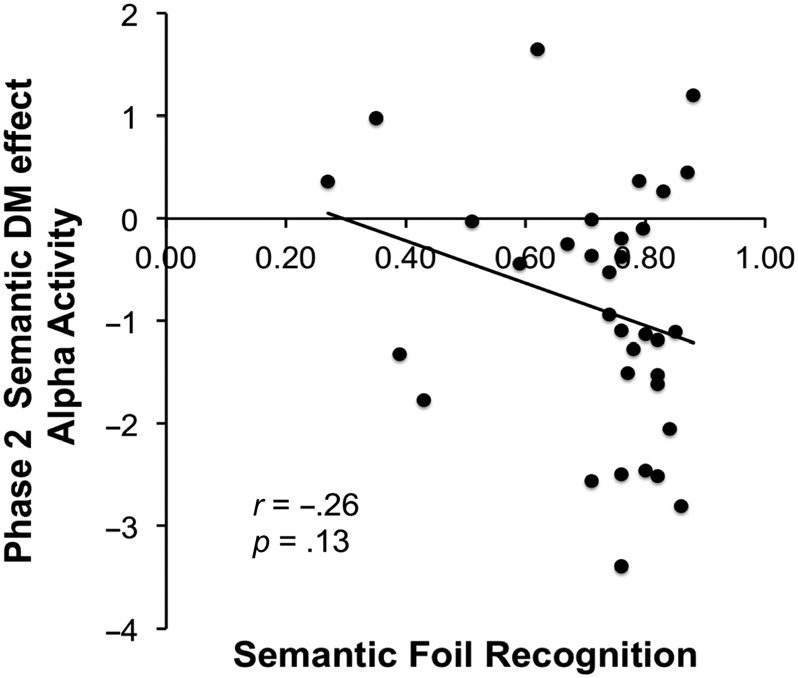
Finally, we examined whether individual differences in
Phase 2 alpha activity associated with semantic foil encod-
ing correlated with individual differences in behavioral se-
mantic foil recognition in Phase 3, which would provide
additional evidence that alpha power is functionally related
to semantic encoding success. We used the Phase 2 alpha
power from the 1000–1200 msec time window associated
with semantic foils that were later remembered versus
forgotten and correlated this with Phase 3 semantic foil rec-
ognition accuracy (proportion of correct responses). A
negative correlation was observed between Phase 2 alpha
power associated with later remembered semantic foils
and Phase 3 semantic foil recognition accuracy; however,
this correlation was not significant (r = −0.26, p = .13;
two-tailed). This correlation is displayed in Figure 9.
Figure 6. Time–frequency plots from the left frontal cluster
illustrating semantic and nonsemantic foil subsequent memory (DM)
EEG oscillation effects (remembered–forgotten). Successful encoding
of semantic foils was uniquely associated with a left frontal alpha
power decrease. The dotted box represents the significant alpha
time window.
DISCUSSION
The aim of the current experiment was to investigate the
neural oscillations involved in the successful encoding of
new “foil” information presented during a recognition test
674
Journal of Cognitive Neuroscience
Volume 30, Number 5
Figure 7. (A) Time courses of
left frontal alpha (average of
8–10 Hz) differences associated
with successful encoding of
semantic versus nonsemantic
foils. Red line on x axis
represents significant time
window that survived correction
for multiple comparisons.
(B) t Values for semantic
versus nonsemantic subsequent
memory effect in the left
frontal electrode cluster.
when participants engage a semantic versus nonsemantic
processing mode. We tested the hypothesis that attempt-
ing to retrieve information from memory involves the
reimplementation of the neurocognitive processes that
were engaged during initial encoding ( Vogelsang et al.,
2016; Bergström et al., 2015; Jacoby, Shimizu, Daniels,
et al., 2005; Jacoby, Shimizu, Velanova, et al., 2005). More
specifically, we used the excellent temporal resolution of
EEG to examine the temporal dynamics of the encoding
of foils to obtain a better understanding of when reimple-
mentation processes occur. It has previously been sug-
gested that the foil effect can be explained by source
constrained retrieval processes that reimplement encod-
ing processes in the early stage of a memory test trial to
guide memory search as a form of “front-end control”
(Kantner & Lindsay, 2013; Jacoby, Shimizu, Daniels, et al.,
2005), predicting that the neural activity associated with
such reimplementation should be apparent shortly after a
recognition cue is presented. An alternative, though not
mutually exclusive, proposal is that control processes
may be engaged at a later stage of processing, for example,
to monitor whether retrieved information is correct
(Halamish et al., 2012; Jacoby et al., 1999) or to elicit rec-
ollection if initial unconstrained retrieval attempts are
unsuccessful as a “late correction” or “back-end control”
strategy ( Jacoby et al., 1999).
Our behavioral findings replicated earlier studies in
demonstrating the typical foil effect: Semantic foils were
remembered significantly more accurately than non-
semantic foils, supporting the idea that participants imple-
mented a semantic processing mode during the semantic
memory test ( Vogelsang et al., 2016; Kantner & Lindsay,
2013; Alban & Kelley, 2012; Halamish et al., 2012; Danckert
et al., 2011; Marsh et al., 2009; Jacoby, Shimizu, Daniels,
et al., 2005; Jacoby, Shimizu, Velanova, et al., 2005).
Time–frequency analysis of EEG data collected during the
initial study phase revealed a power decrease in alpha
frequencies over left frontal electrodes between 800 and
1000 msec during the semantic as opposed to nonsemantic
task, consistent with prior literature highlighting a role for
alpha oscillations in semantic processing (e.g., Bastiaansen
et al., 2005; for a review, see Klimesch, 1999). Importantly,
the EEG data from the first recognition test revealed
decreases in alpha oscillatory activity in the left frontal elec-
trode cluster between 1000 and 1200 msec that predicted
subsequent recognition of semantic, but not nonsemantic,
Figure 8. Correlations between Phase 1 (800–1000 msec) and Phase 2
(1000–1200 msec) alpha activity.
Figure 9. Correlations between Phase 2 alpha related activity for
remembered versus forgotten semantic foils (1000–1200 msec) and
subsequent semantic foil recognition accuracy (proportion correct
responses).
Vogelsang et al.
675
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
a
_
0
1
2
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
foils during the final surprise recognition test. Thus, similar
oscillatory activity was associated with semantic processing
during initial study and during foil encoding while partici-
pants were trying to retrieve semantic information. Further-
more, individual differences in alpha activity during the
semantic study phase (Phase 1) were significantly cor-
related with individual differences in alpha activity for suc-
cessfully encoded foils during the semantic recognition test
(Phase 2), suggesting that the semantic neurocognitive
processes that were engaged during initial study were
reimplemented during the encoding of foils during the
Phase 2 recognition test. Finally, we tested whether alpha
decreases in phase semantic foil encoding during Phase 2
were negatively correlated with behavioral semantic foil
recognition during Phase 3; however, this analysis did not
reach significance, leaving the question open as to whether
alpha power is functionally related to semantic foil encod-
ing success.
Interestingly, alpha power decreases associated with
semantic foil encoding became apparent after the aver-
age time when participants provided their response, at
about 900 msec after stimulus presentation, suggesting
that alpha oscillations may reflect an implementation of
encoding operations at a relatively late processing stage,
contrary to what would be predicted if such reinstate-
ment was conducted as part of a front-end control strat-
egy (Gray & Gallo, 2015; Jacoby, Shimizu, Daniels, et al.,
2005; Jacoby, Shimizu, Velanova, et al., 2005). In a recent
fMRI study, we found that the LIFG was significantly
more active during successful encoding of semantic as
opposed to nonsemantic foils ( Vogelsang et al., 2016).
The LIFG has been widely associated with semantic pro-
cessing (Poldrack et al., 1999; Wagner et al., 1998), but
the low temporal resolution of fMRI precluded us from
determining whether LIFG activation reflected mentally
reenacting a semantic processing mode early or later in
the trial. The timing of the current EEG results suggest
that the left frontal alpha decreases, which we tentatively
interpret as possibly generated by the LIFG ( Vogelsang
et al., 2016), may reflect strategic processes that are en-
gaged during a later decision stage of retrieval, which
facilitates the incidental semantic encoding of foils.
The main underlying finding of the “foil effect” is that
“deep” semantic foils are recognized significantly better
than “shallow” nonsemantic foils despite the only differ-
ence between the foils being the type of information that
was oriented toward when they were first encountered.
There is nothing intrinsically “semantic” or “nonsemantic”
about the foils themselves. Instead, the instructions at
the start of the memory test determines which type of
encoding context (semantic vs. nonsemantic) participants
need to retrieve, appears in some manner to initiate
“deeper” retrieval for the semantic test and “shallower”
retrieval for the nonsemantic test ( Jacoby, Shimizu,
Velanova, et al., 2005), resulting in better incidental encod-
ing of semantic compared with nonsemantic foils. Further-
more, this encoding difference occurs spontaneously,
because participants are not explicitly asked to use differ-
ent retrieval strategies. The “foil effect” can be interpreted
as evidence for the source constrained retrieval hypothesis
as put forth by Jacoby, Shimizu, Daniels, et al. (2005) and
indicates that sought after information can be used to
guide information processing during recognition memory.
In this way, memory retrieval can be considered an encod-
ing event, determining whether information will be remem-
bered in the future (Dudukovic, Dubrow, & Wagner, 2009).
Why did the neural markers of semantic encoding of
foils occur so late? Because reinstating encoding opera-
tions is an effortful, self-initiated process (Alban & Kelley,
2012), it is possible that participants chose to engage
such a strategy to elicit recollection only if an initial un-
constrained retrieval attempt was unsuccessful. A related
account suggests that participants may reinstate encod-
ing operations to verify and possibly correct their initial
more automatic retrieval assessments, and such a moni-
toring strategy may contribute to enhanced encoding of
semantic foils together with earlier “front-end” control
processes (Halamish et al., 2012). According to Jacoby
et al. (1999), participants engage in such late correction
strategies primarily when the retrieved information is
vague or ambiguous (perhaps eliciting a sense of famil-
iarity without recollection of decisive contextual informa-
tion). The current oscillatory findings are consistent with
reinstatement of encoding operations occurring at a late
retrieval stage but do not rule out the possibility that
encoding operations were also reinstated to constrain
retrieval at the front-end without being reflected in our
EEG results (EEG oscillations of course only capture cer-
tain aspects of neural activity).
Our oscillatory findings are consistent with prior litera-
ture highlighting a role for alpha frequencies in successful
semantic encoding (Hanslmayr & Staudigl, 2014; Zion-
Golumbic et al., 2010; Hanslmayr et al., 2009) and semantic
processing (Long, Burke, & Kahana, 2014; Klimesch,
Doppelmayr, & Hanslmayr, 2006). In the oscillations litera-
ture, alpha frequencies have been linked with a wide variety
of cognitive functions ranging from inhibitory processes
during memory suppression (Park et al., 2014), to fine-
grained resolution of visual processing (Samaha & Postle,
2015), working memory (Myers, Stokes, Walther, & Nobre,
2014; Sauseng et al., 2009), and active inhibition of a not-
to-be applied rule (Buschman, Denovellis, Diogo, Bullock,
& Miller, 2012). One of the first studies that found a rela-
tionship between decreases in alpha and later memory
performance was conducted by Klimesch (1997), who ob-
served that decreases in alpha frequencies over parietal
electrodes during semantic encoding were positively cor-
related with later memory retrieval. Hanslmayr et al.
(2009) contrasted deep semantic encoding with shallow
nonsemantic encoding and found power decreases in
alpha (and beta) frequency bands that were related to suc-
cessful semantic encoding only, and Fellner et al. (2013)
showed that alpha likely reflects semantic processing spe-
cifically, rather than elaborative and efficient encoding
676
Journal of Cognitive Neuroscience
Volume 30, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
a
_
0
1
2
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
strategies in general. In our experiment, individual differ-
ences in alpha power decreases during the semantic rec-
ognition test for foils that were later remembered did not
correlate significantly with individual differences in seman-
tic foil recognition accuracy during the final surprise mem-
ory test. Therefore, we were not able to obtain additional
converging evidence that alpha power decreases reflect
successful semantic encoding.
Jacoby and colleagues ( Jacoby, Shimizu, Daniels, et al.,
2005; Jacoby, Shimizu, Velanova, et al., 2005) have hypoth-
esized that a possible explanation for the enhanced encod-
ing of semantic versus nonsemantic foils in the memory for
foils paradigm might lie in the Transfer Appropriate Pro-
cessing Framework and the related Encoding Specificity
Principle, both of which predict that retrieval success
depends on the amount of overlap between encoding and
retrieval processes (Roediger, 1990; Morris et al., 1977;
Tulving & Thomson, 1973). While attempting to retrieve
words that had either been semantically or nonseman-
tically encoded, participants may mentally reenact the orig-
inal study task, resulting in all recognition probes (both old
items and foils) being processed semantically during the
semantic test block and nonsemantically during the non-
semantic test block. Semantic retrieval attempts might
involve thinking about the meaning of a foil word (e.g.,
“do I think a strawberry is pleasant?”), whereas nonseman-
tic retrieval attempts might involve examining the letters
of the word in the hope that such a strategy will help to
decide whether the word is old or new. Such reenactment
may be a relatively late strategy that participants engage in
after an initial heuristic familiarity assessment and may
therefore be expressed in neural activity around the time or
even after participants have made their memory judgment.
The current time–frequency results in combination with
previous research indicate that neural oscillations are a use-
ful tool for studying the temporal dynamics of encoding–
retrieval overlap (Staresina et al., 2016; Waldhauser, Braun,
& Hanslmayr, 2016; Staudigl, Vollmar, Noachtar, & Hanslmayr,
2015; Jafarpour, Fuentemilla, Horner, Penny, & Duzel, 2014;
Staudigl & Hanslmayr, 2013; Waldhauser, Johansson, &
Hanslmayr, 2012). Burke et al. (2013), for example, found
that high gamma activity (44–100 Hz) during successful
encoding of information is also observed in similar brain
areas during the memory test phase in which previously
studied items need to be recalled. Waldhauser and col-
leagues (2012) observed that decreases in alpha/beta fre-
quencies during retrieval were associated with reactivation
of encoded target information, whereas increases in alpha/
beta power were associated with the inhibition of encoded
distracter information. Cortical reinstatement has also been
identified in an entrainment study in which participants
studied words presented on flickering backgrounds of
either 6 or 10 Hz (Wimber, Maaß, Staudigl, Richardson-
Klavehn, & Hanslmayr, 2012). EEG measurements during
successful retrieval of studied words exhibited 6- and 10-Hz
frequency oscillations similar to the background flicker rates
in which the words had been studied and the strength of
this reactivation was related to whether a word was remem-
bered or forgotten (Wimber et al., 2012). More work needs
to be done, however, to examine what mechanisms under-
lie the principle of encoding reimplementation and how
that facilitates retrieval. One prominent view is that, dur-
ing retrieval, a cue reactivates only a part of the encoded
memory and that activity of a fraction of the original pat-
tern triggers the reactivation of the entire trace (Rugg
et al., 2008). This “pattern completion” process has been
linked with the hippocampus, and a role for gamma
power increases and alpha power decreases has been
proposed as a neural mechanism underlying pattern com-
pletion (Staresina et al., 2016). However, there is also evi-
dence that suggests that alpha and beta frequency bands
in the cortex are important for content-specific processing
(Hanslmayr, Staresina, & Bowman, 2016), which is in line
with our current findings of alpha frequencies represent-
ing semantic processing.
To conclude, we investigated the neural oscillations in-
volved in the encoding of new “foil” information presented
during a retrieval test as a function of whether the test re-
quired participants to retrieve semantic versus nonseman-
tic information. Our findings show that semantic encoding
during retrieval attempts was associated with power de-
creases in left frontal alpha oscillations, which may origi-
nate from the LIFG ( Vogelsang et al., 2016). Consistent
with previous findings, our results support the view that
participants reimplement the distinct neurocognitive oper-
ations that were engaged during initial encoding, and we
extend previous research by identifying that the time
course of this reimplementation may be at a relatively late
processing stage. Thus, retrieving memories does not only
involve recollecting events from the past but may also
determine what type of information will be remembered
in the future.
Acknowledgments
This study was supported by a James S. McDonnell Foundation
Scholar Award to J. S. S. and was carried out within the University
of Cambridge Behavioural and Clinical Neuroscience Institute,
funded by a joint award from the Medical Research Council and
the Wellcome Trust.
Reprint requests should be sent to Jon S. Simons, Department of
Psychology, University of Cambridge, Downing Street, Cambridge
CB2 3EB, United Kingdom, or via e-mail: jss30@cam.ac.uk.
REFERENCES
Addante, R. J., Watrous, A. J., Yonelinas, A. P., Ekstrom, A. D.,
& Ranganath, C. (2011). Prestimulus theta activity predicts
correct source memory retrieval. Proceedings of the National
Academy of Sciences, U.S.A., 108, 10702–10707.
Alban, M. W., & Kelley, C. M. (2012). Variations in constrained
retrieval. Memory and Cognition, 40, 681–692.
Bakker, I., Takashima, A., van Hell, J. G., Janzen, G., & McQueen,
J. M. (2015). Changes in theta and beta oscillations as
signatures of novel word consolidation. Journal of Cognitive
Neuroscience, 27, 1286–1297.
Vogelsang et al.
677
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
a
_
0
1
2
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Bastiaansen, M. C. M., van der Linden, M., ter Keurs, M., Dijkstra,
T., & Hagoort, P. (2005). Theta responses are involved in
lexical–semantic retrieval during language processing. Journal
of Cognitive Neuroscience, 17, 530–541.
Bell, A. J., & Sejnowski, T. J. (1995). An information-maximization
approach to blind separation and blind deconvolution.
Neural Computation, 7, 1129–1159.
Bergström, Z. M., Vogelsang, D. A., Benoit, R. G., & Simons, J. S.
(2015). Reflections of oneself: Neurocognitive evidence for
dissociable forms of self-referential recollection. Cerebral
Cortex, 25, 2648–2657.
Blair, R. C., & Karniski, W. (1993). An alternative method for
significance testing of waveform difference potentials.
Psychophysiology, 30, 518–524.
Burke, J. F., Zaghloul, K. A., Jacobs, J., Williams, R. B., Sperling,
M. R., Sharan, A. D., et al. (2013). Synchronous and
asynchronous theta and gamma activity during episodic
memory formation. Journal of Neuroscience, 33, 292–304.
Buschman, T. J., Denovellis, E. L., Diogo, C., Bullock, D., &
Miller, E. K. (2012). Synchronous oscillatory neural ensembles
for rules in the prefrontal cortex. Neuron, 76, 838–846.
Cohen, M. X. (2014). Analyzing neural time series data: Theory
and practice. Cambridge, MA: MIT Press.
Danckert, S. L., MacLeod, C. M., & Fernandes, M. A. (2011).
Source-constrained retrieval influences the encoding of new
information. Memory and Cognition, 39, 1374–1386.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
Dewhurst, S. A., & Brandt, K. R. (2007). Reinstating effortful
encoding operations at test enhances episodic remembering.
Quarterly Journal of Experimental Psychology, 60, 543–550.
Dudukovic, N. M., Dubrow, S., & Wagner, A. D. (2009).
Attention during memory retrieval enhances future
remembering. Memory Cognition, 37, 953–961.
Fell, J., & Axmacher, N. (2011). The role of phase synchronization
in memory processes. Nature Reviews Neuroscience, 12,
105–118.
Fell, J., Ludowig, E., Staresina, B. P., Wagner, T., Kranz, T., Elger,
C. E., et al. (2011). Medial temporal theta/alpha power
enhancement precedes successful memory encoding: Evidence
based on intracranial EEG. Journal of Neuroscience, 31,
5392–5397.
Fellner, M.-C., Bäuml, K.-H., & Hanslmayr, S. (2013). Brain
oscillatory subsequent memory effects differ in power and
long-range synchronization between semantic and survival
processing. Neuroimage, 79, 361–370.
Fletcher, P. C., Stephenson, C. M., Carpenter, T. A., Donovan,
T., & Bullmorel, E. T. (2003). Regional brain activations
predicting subsequent memory success: An event-related
fMRI study of the influence of encoding tasks. Cortex, 39,
1009–1026.
Gray, S. J., & Gallo, D. A. (2015). Disregarding familiarity during
recollection attempts: Content-specific recapitulation as a
retrieval orientation strategy. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 41, 134–147.
Gruber, M. J., Watrous, A. J., Ekstrom, A. D., Ranganath, C., &
Otten, L. J. (2013). Expected reward modulates encoding-
related theta activity before an event. Neuroimage, 64, 68–74.
Halamish, V., Goldsmith, M., & Jacoby, L. L. (2012). Source
constrained recall: Front-end and back-end control of retrieval
quality. Journal of Experimental Psychology: Learning,
Memory, and Cognition, 38, 1–15.
Hanslmayr, S., Spitzer, B., & Bäuml, K.-H. (2009). Brain
oscillations dissociate between semantic and nonsemantic
encoding of episodic memories. Cerebral Cortex, 19,
1631–1640.
Hanslmayr, S., Staresina, B. P., & Bowman, H. (2016). Oscillations
and episodic memory: Addressing the synchronization/
desynchronization conundrum. Trends in Neurosciences,
39, 16–25.
Hanslmayr, S., & Staudigl, T. (2014). How brain oscillations
form memories—A processing based perspective on
oscillatory subsequent memory effects. Neuroimage, 85,
648–655.
Hsieh, L.-T., Ekstrom, A. D., & Ranganath, C. (2011). Neural
oscillations associated with item and temporal order
maintenance in working memory. Journal of Neuroscience,
31, 10803–10810.
Jacoby, L. L., Kelley, C. M., & McElree, B. D. (1999). The role
of cognitive control: Early selection vs. late correction. In
S. Chaiken & Y. Trope (Eds.), Dual-process theories in social
psychology (pp. 383–400). New York: Guilford Press.
Jacoby, L. L., Shimizu, Y., Daniels, K. A., & Rhodes, M. G. (2005).
Modes of cognitive control in recognition and source
memory: Depth of retrieval. Psychonomic Bulletin & Review,
12, 852–857.
Jacoby, L. L., Shimizu, Y., Velanova, K., & Rhodes, M. G. (2005).
Age differences in depth of retrieval: Memory for foils.
Journal of Memory and Language, 52, 493–504.
Jafarpour, A., Fuentemilla, L., Horner, A. J., Penny, W., & Duzel,
E. (2014). Replay of very early encoding representations
during recollection. Journal of Neuroscience, 34, 242–248.
Kantner, J., & Lindsay, D. S. (2013). Top–down constraint on
recognition memory. Memory and Cognition, 41, 465–479.
Kim, H. (2011). Neural activity that predicts subsequent
memory and forgetting: A meta-analysis of 74 fMRI studies.
Neuroimage, 54, 2446–2461.
Klimesch, W. (1997). EEG-alpha rhythms and memory
processes. International Journal of Psychophysiology, 26,
319–340.
Klimesch, W. (1999). EEG alpha and theta oscillations reflect
cognitive and memory performance: A review and analysis.
Brain Research Reviews, 29, 169–195.
Klimesch, W., Doppelmayr, M., & Hanslmayr, S. (2006). Upper
alpha ERD and absolute power: Their meaning for memory
performance. Progress in Brain Research, 159, 151–165.
Klimesch, W., Doppelmayr, M., Schimke, H., & Ripper, B.
(1997). Theta synchronization and alpha desynchronization
in a memory task. Psychophysiology, 34, 169–176.
Long, N. M., Burke, J. F., & Kahana, M. J. (2014). Subsequent
memory effect in intracranial and scalp EEG. Neuroimage,
84, 488–494.
Marsh, R. L., Meeks, J. T., Cook, G. I., Clark-Foos, A., Hicks, J. L.,
& Brewer, G. A. (2009). Retrieval constraints on the front
end create differences in recollection on a subsequent test.
Journal of Memory and Language, 61, 470–479.
Morris, C. D., Bransford, J. D., & Franks, J. J. (1977). Levels of
processing versus transfer appropriate processing. Journal
of Verbal Learning and Verbal Behavior, 16, 519–533.
Myers, N. E., Stokes, M. G., Walther, L., & Nobre, A. C. (2014).
Oscillatory brain state predicts variability in working memory.
Journal of Neuroscience, 34, 7735–7743.
Park, H., Lee, D. S., Kang, E., Kang, H., Hahm, J., Kim, J. S., et al.
(2014). Blocking of irrelevant memories by posterior alpha
activity boosts memory encoding. Human Brain Mapping,
35, 3972–3987.
Percival, D. B., & Walden, A. T. (1993). Spectral analysis for
physical applications: Multitaper and conventional
univariate techniques. Cambridge, UK: Cambridge
University Press.
Poldrack, R. A., Wagner, A. D., Prull, M. W., Desmond, J. E.,
Glover, G. H., & Gabrieli, J. D. (1999). Functional specialization
for semantic and phonological processing in the left inferior
prefrontal cortex. Neuroimage, 10, 15–35.
678
Journal of Cognitive Neuroscience
Volume 30, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
a
_
0
1
2
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Roediger, H. L., III. (1990). Implicit memory: Retention without
remembering. American Psychologist, 45, 1043–1056.
Roediger, H. L., III, Weldon, M. S., & Challis, B. H. (1989).
Explaining dissociations between implicit and explicit
measures of retention: A processing account. In H. L.
Roediger III & F. I. M. Craik (Eds.), Variations of memory
and consciousness: Essays in honor of Endel Tulving
(pp. 3–41). Hillsdale, NJ: Lawrence Erlbaum Associates.
Rugg, M. D., Johnson, J. D., Park, H., & Uncapher, M. R. (2008).
Encoding–retrieval overlap in human episodic memory: A
functional neuroimaging perspective. Progress in Brain
Research, 169, 339–352.
Rugg, M. D., & Wilding, E. L. (2000). Retrieval processing and
episodic memory. Trends in Cognitive Science, 4, 108–115.
Samaha, J., & Postle, B. R. (2015). The speed of alpha-band
oscillations predicts the temporal resolution of visual
perception. Current Biology, 25, 2985–2990.
Sauseng, P., Klimesch, W., Heise, K. F., Gruber, W. R., Holz, E.,
Karim, A. A., et al. (2009). Brain oscillatory substrates of
visual short-term memory capacity. Current Biology, 19,
1846–1852.
Snodgrass, J. G., & Corwin, J. (1988). Pragmatics of measuring
recognition memory: Applications to dementia and amnesia.
Journal of Experimental Psychology: General, 117, 34–50.
Staresina, B. P., Michelmann, S., Bonnefond, M., Jensen, O.,
Axmacher, N., & Fell, J. (2016). Hippocampal pattern
completion is linked to gamma power increases and alpha
power decreases during recollection. eLife, 5, e17397.
Staudigl, T., & Hanslmayr, S. (2013). Theta oscillations at
encoding mediate the context-dependent nature of human
episodic memory. Current Biology, 23, 1101–1106.
Staudigl, T., Hanslmayr, S., & Bäuml, K.-H. (2010). Theta
oscillations reflect the dynamics of interference in episodic
memory retrieval. Journal of Neuroscience, 30, 11356–11362.
Staudigl, T., Vollmar, C., Noachtar, S., & Hanslmayr, S. (2015).
Temporal-pattern similarity analysis reveals the beneficial
and detrimental effects of context reinstatement on human
memory. Journal of Neuroscience, 35, 5373–5384.
Tucker, D. M. (1993). Spatial sampling of head electrical fields:
The geodesic sensor net. Electroencephalography and
Clinical Neurophysiology, 87, 154–163.
Tulving, E., & Thomson, D. M. (1973). Encoding specificity and
retrieval processes in episodic memory. Psychological
Review, 80, 352–373.
Vogelsang, D. A., Bonnici, H. M., Bergström, Z. M., Ranganath,
C., & Simons, J. S. (2016). Goal-directed mechanisms that
constrain retrieval predict subsequent memory for new “foil”
information. Neuropsychologia, 89, 356–363.
Wagner, A. D., Schacter, D. L., Rotte, M., Koutstaal, W., Maril, A.,
Dale, A. M., et al. (1998). Building memories: Remembering
and forgetting of verbal experiences as predicted by brain
activity. Science, 281, 1188–1191.
Waldhauser, G. T., Braun, V., & Hanslmayr, S. (2016). Episodic
memory retrieval functionally relies on very rapid reactivation
of sensory information. Journal of Neuroscience, 36, 251–260.
Waldhauser, G. T., Johansson, M., & Hanslmayr, S. (2012).
Alpha/beta oscillations indicate inhibition of interfering visual
memories. Journal of Neuroscience, 32, 1953–1961.
Wilson, M. (1988). The MRC psycholinguistic database:
Machine-usable dictionary, version 2.00. Behavior Research
Methods, Instruments, and Computers, 20, 6–10.
Wimber, M., Maaß, A., Staudigl, T., Richardson-Klavehn, A.,
& Hanslmayr, S. (2012). Rapid memory reactivation
revealed by oscillatory entrainment. Current Biology, 22,
1482–1486.
Zawadzka, K., Hanczakowski, M., & Wilding, E. L. (2017). Late
consequences of early selection: When memory monitoring
backfires. Journal of Memory and Language, 92, 114–127.
Zion-Golumbic, E., Kutas, M., & Bentin, S. (2010). Neural
dynamics associated with semantic and episodic memory for
faces: Evidence from multiple frequency bands. Journal of
Cognitive Neuroscience, 22, 263–277.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
5
6
6
7
1
7
8
7
5
0
7
/
j
o
c
n
_
a
_
0
1
2
3
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Vogelsang et al.
679