A Double Dissociation between Anterior and Posterior
Superior Temporal Gyrus for Processing Audiovisual
Speech Demonstrated by Electrocorticography
Muge Ozker1,2, Inga M. Schepers3, John F. Magnotti2, Daniel Yoshor2,
and Michael S. Beauchamp2
D
o
w
n
je
o
un
d
e
d
F
r
o
m
Abstrait
■ Human speech can be comprehended using only auditory
information from the talker’s voice. Cependant, comprehension
is improved if the talker’s face is visible, especially if the audi-
tory information is degraded as occurs in noisy environments or
with hearing loss. We explored the neural substrates of audio-
visual speech perception using electrocorticography, direct re-
cording of neural activity using electrodes implanted on the
cortical surface. We observed a double dissociation in the re-
sponses to audiovisual speech with clear and noisy auditory
component within the superior temporal gyrus (STG), a region
long known to be important for speech perception. Anterior
STG showed greater neural activity to audiovisual speech with
clear auditory component, whereas posterior STG showed
similar or greater neural activity to audiovisual speech in which
the speech was replaced with speech-like noise. A distinct border
between the two response patterns was observed, demarcated by
a landmark corresponding to the posterior margin of Heschl’s
gyrus. To further investigate the computational roles of both re-
gions, we considered Bayesian models of multisensory integra-
tion, which predict that combining the independent sources of
information available from different modalities should reduce
variability in the neural responses. We tested this prediction by
measuring the variability of the neural responses to single audio-
visual words. Posterior STG showed smaller variability than
anterior STG during presentation of audiovisual speech with
noisy auditory component. Taken together, these results suggest
that posterior STG but not anterior STG is important for multi-
sensory integration of noisy auditory and visual speech. ■
INTRODUCTION
Human speech perception is multisensory, combining
auditory information from the talker’s voice with visual
information from the talker’s face. Visual speech informa-
tion is particularly important in noisy environments in
which the auditory speech is difficult to comprehend
(Ross, Saint-Amour, Leavitt, Javitt, & Foxe, 2007; Bernstein,
Auer, & Takayanagi, 2004; Sumby & Pollack, 1954).
Although visual speech can substantially improve the per-
ception of noisy auditory speech, little is known about the
neural mechanisms underlying this perceptual benefit.
Speech varies on a timescale of milliseconds, requiring
the brain to accurately integrate auditory and visual speech
with high temporal fidelity. Cependant, the most popular
technique for measuring human brain activity, BOLD fMRI,
is an indirect measure of neural activity with a temporal
resolution on the order of seconds, making it difficult to
accurately measure rapidly changing neural responses to
speech. To overcome this limitation, we recorded from
the brains of participants implanted with electrodes for the
treatment of epilepsy. This technique, known as electro-
1University of Texas Graduate School of Biomedical Sciences at
Houston, 2Baylor College of Medicine, 3University of Oldenburg
corticography, allows for the direct measurement of activ-
ity in small populations of neurons with millisecond
precision. We measured activity in electrodes implanted
over the superior temporal gyrus (STG), a key brain area
for speech perception (Mesgarani, Cheung, Johnson, &
Chang, 2014; Binder et al., 2000), as participants were pre-
sented with audiovisual speech with either clear or noisy
auditory or visual components.
The STG is functionally heterogeneous. Regions of ante-
rior STG lateral to Heschl’s gyrus are traditionally classified
as unisensory auditory association cortex (Rauschecker,
2015). In contrast, regions of posterior STG and STS are
known to be multisensory, responding to both auditory
and visual stimuli including faces and voices, letters and
voices, and recordings and videos of objects (Reale et al.,
2007; Miller & D'Esposito, 2005; Beauchamp, Lee, Argall, &
Martine, 2004; van Atteveldt, Formisano, Goebel, & Blomert,
2004; Foxe et al., 2002; Calvert, Campbell, & Brammer,
2000).
On the basis of this distinction, we hypothesized that
anterior and posterior regions of STG should differ in
their electrocorticographic response to clear and noisy
audiovisual speech. We expected that auditory asso-
ciation areas in anterior STG should respond strongly
to speech with a clear auditory component but show
© 2017 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 3.0 Unported (CC PAR 3.0) Licence.
Journal des neurosciences cognitives 29:6, pp. 1044–1060
est ce que je:10.1162/jocn_a_01110
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
un
/
_
j
0
o
1
c
1
n
1
0
_
un
p
_
d
0
1
b
1
oui
1
g
0
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
F
/
t
.
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
a reduced response to the reduced information avail-
able in speech with noisy auditory component. Multi-
sensory areas in posterior STG should be able to use
the clear visual speech information to compensate for
the noisy auditory speech, resulting in similar responses
to speech with clear and noisy auditory components.
A related set of predictions comes from theoretical
models of Bayesian integration. In these models, sensoriel
noise and the resulting neural variability is independent
in each modality. Combining the modalities through
multisensory integration results in a decreased neural
variability (and improved perceptual accuracy) relative
to unisensory stimulation (Fetsch, Pouget, DeAngelis, &
Angelaki, 2012; Knill & Pouget, 2004). Bayesian models
predict that unisensory areas, such as those in anterior
STG, should have greatly increased variability as the
sensory noise in their preferred modality increases. Multi-
sensory areas, like those in posterior STG, should be less
influenced by the addition of auditory noise, resulting in
similar variability for speech with clear and noisy auditory
components.
was replaced with noise that matched the spectrotemporal
power distribution of the original auditory speech. Le
total power of this speech-specific noise was equated to
the total power of the original auditory speech (Schepers,
Schneider, Hipp, Ange, & Senkowski, 2013). This process
generated speech-like noise.
To create speech stimuli with a noisy visual component,
the visual component of the speech stimulus was blurred
using a 2-D Gaussian low-pass filter (MATLAB function
fspecial, filter size = 30 pixels in each direction). Each vid-
eo frame (image size = 200 × 200 pixels) was filtered sep-
arately using 2-D correlation (MATLAB function imfilter).
Values outside the bounds of the images were assumed
to equal the nearest image border. These filter settings
resulted in highly blurred videos.
Thirty-two to 56 repetitions of each condition were
presented in random sequence. Each 5.4-sec trial con-
sisted of a single 1.4-sec video clip followed by an ISI
de 4 sec during which a fixation cross on a gray screen
was presented. Participants pressed a mouse button to
report which word was presented.
MÉTHODES
Participant Information
All experimental procedures were approved by the insti-
tutional review board of Baylor College of Medicine. Five
human participants with refractory epilepsy (3 femmes,
âge moyen = 31 années) were implanted with subdural
electrodes guided by clinical requirements. Following
surgery, participants were tested while resting comfort-
ably in their hospital bed in the epilepsy monitoring unit.
Stimuli, Experimental Design, and Task
Visual stimuli were presented on an LCD monitor posi-
tioned at 57-cm distance from the participant, and audi-
tory stimuli were played through loudspeakers positioned
next to the participant’s bed. Two video clips of a female
talker pronouncing the single syllable words “rain” and
“rock” with clear auditory and visual components (AV)
were selected from the Hoosier Audiovisual Multitalker
Database (Sheffert, Lachs, & Hernandez, 1996). The dura-
tion of each video clip was 1.4 sec, and the duration of the
auditory stimulus was 520 msec for “rain” and 580 msec
for “rock.” The auditory word onsets were 410 msec for
“rain” and 450 msec for “rock” after the video onset.
The face of the talker subtended approximately 15° hori-
zontally and 15° vertically.
Speech stimuli were consisted of four conditions:
Speech with clear auditory and visual components (AV),
clear visual but noisy auditory components (AnV), clear
auditory but noisy visual components (AVn), and finally
noisy auditory and noisy visual components (AnVn).
Electrode Localization and Recording
Before surgery, T1-weighted structural MRI scans were
used to create cortical surface models (Figure 1A) avec
FreeSurfer (Dale, Fischl, & Séréno, 1999; Fischl, Séréno,
& Dale, 1999) and visualized using SUMA (Argall, Saad, &
Beauchamp, 2006). Participants underwent a whole-
head CT after the electrode implantation surgery. Le
postsurgical CT scan and presurgical MR scan were
aligned using AFNI (Cox, 1996), and all electrode posi-
tions were marked manually on the structural MR im-
ages. Electrode positions were then projected to the
nearest node on the cortical surface model using the
AFNI program SurfaceMetrics. Resulting electrode posi-
tions on the cortical surface model were confirmed by
comparing them with the photographs taken during the
implantation surgery.
A 128-channel Cerebus amplifier (Blackrock Micro-
systèmes, Salt Lake City, UT) was used to record from
subdural electrodes (Ad-Tech Corporation, Racine, WI)
that consisted of platinum alloy discs embedded in a
flexible silicon sheet. Electrodes had an exposed surface
diameter of 2.3 mm and were located on strips or grids
with interelectrode distances of 10 mm. An inactive intra-
cranial electrode implanted facing the skull was used as a
reference for recording. Signals were amplified, filtered
(low-pass: 500 Hz, Butterworth filter with order 4; haut-
pass: 0.3 Hz, Butterworth filter with order 1) and digi-
tized at 2 kHz.
Electrophysiological Data Analysis
To create speech stimuli with a noisy auditory com-
ponent, the auditory component of the speech stimulus
Data were analyzed in MATLAB 8.5.0 (MathWorks, Inc.
Natick, MA) using the FieldTrip toolbox (Oostenveld,
Ozker et al.
1045
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
un
/
_
j
0
o
1
c
1
n
1
0
_
un
p
_
d
0
1
b
1
oui
1
g
0
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
F
/
t
.
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Fries, Maris, & Schoffelen, 2011). To remove common
artifacts, the average signal across all electrodes was sub-
tracted from each individual electrode’s signal (common
average referencing). The continuous data stream was
epoched into trials. Line noise at 60, 120, 180 Hz was re-
moved, and the data were transformed to time–frequency
space using the multitaper method (three Slepian tapers,
frequency window from 10 à 200 Hz, frequency steps of
2 Hz, time steps of 10 msec, temporal smoothing of
200 msec, frequency smoothing of ±10 Hz).
Our primary measure of neural activity was the broad-
band response in the high-gamma frequency band, rang-
ing from 70 à 110 Hz. This frequency range is thought
to reflect the frequency of action potentials in nearby
neurons ( Jacques et al., 2016; Ray & Maunsell, 2011;
Nir et al., 2007; Mukamel et al., 2005). For each trial,
the high-gamma response was measured in a window
depuis 0 à 500 msec following auditory stimulus onset
(reflecting the ∼500 msec duration of the auditory stim-
ulus) and converted to percent signal change measure by
comparing the high-gamma response to a within-trial base-
line window encompassing −500 to −100 msec before
auditory stimulus onset. Par exemple, un 100% changement de signal
on one trial would mean the power in the high-gamma
band doubled from the pre-stimulus to the post-stimulus
interval. For each electrode, the mean percent signal
change in the high-gamma band across all trials of a
given condition was calculated (m).
Our second analysis focused on neural variability across
repeated presentations of identical stimuli. One obvious
measure of variability is variance (defined as the square
of the standard deviation across all observations). Comment-
jamais, the variance of neural responses is known to increase
with increasing response amplitude (Ma, Beck, Latham,
& Pouget, 2006; Tolhurst, Movshon, & Dean, 1983), et
our initial analysis demonstrated differences in response
amplitude between speech with clear and noisy auditory
components (Tableau 1). To search for variability differences
without the confound of these amplitude differences, nous
used a different measure of variability known as the co-
efficient of variation (CV), which normalizes across ampli-
tude differences by dividing the standard deviation of
the response across trials by the mean response amplitude
(CV = σ/μ; Churchland et al., 2010; Gur, Beylin, & Snodderly,
1997). The CV assumes that variance covaries linearly with
amplitude. We tested this assumption by calculating the
Pearson correlation between the mean and variance of
the high-gamma response across all anterior and posterior
STG electrodes and found it to be reasonable for the
four different stimulus conditions (AV: r = .96, p = 10−16;
AnV: r = .86, p = 10−8; AVn: r = .97, p = 10−16; AnVn: r =
.91, p = 10−11). Although CV has the advantage of ac-
counting for the known correlation between amplitude
and variance, it has the disadvantage that it becomes
undefined as response amplitude approaches zero. Pour
this reason, response amplitudes of less than 15% étaient
excluded from the CV analysis, affecting 3 de 16 anterior
electrodes in Figure 3 et 8 de 216 condition-electrode
pairs in Table 2 and Table 7.
Anatomical Classification and Electrode Selection
The STG was segmented on each participant’s cortical
surface model. The posterior margin of the most medial
portion of the transverse temporal gyrus of Heschl was
used as a landmark to separate the STG into anterior
and posterior portions (the A–P boundary). All of the
STG anterior to this point (extending to the temporal
pole) was classified as anterior STG. All of the STG pos-
terior to this point was classified as posterior STG.
The cortical surface atlases supplied with FreeSurfer
were used to automate ROI creation. The entire seg-
mented STG was obtained from the Destrieux atlas (droite
hemisphere STG atlas value = 152, left hemisphere = 78;
Destrieux, Fischl, Dale, & Halgren, 2010) and the anterior
and posterior boundaries of the posterior STG were ob-
tained from the Desikan-Killiany atlas (RH = 44, LH = 79;
Desikan et al., 2006).
A total of 527 intracranial electrodes were recorded
depuis. Of these, 55 were located on the STG. These were
examined for stimulus-related activity, defined as sig-
nificant high-gamma responses to audiovisual speech
compared with prestimulus baseline ( p < 10−3, equiva-
lent to ∼40% increase in stimulus power from baseline).
A total of 27 electrodes met both anatomical and func-
tional criteria and were selected for further analysis. To
simplify future meta-analyses and statistical comparisons
between experiments, we do not report p values as in-
equalities but instead report actual values (rounded to the
nearest order of magnitude for p values less than .001).
Response Timing Measurements
For each electrode, we calculated the response onset,
time to peak, and duration of the high gamma signal.
To calculate the response onset, we found the first time
point after the auditory speech onset at which the high-
gamma signal deviated three standard deviations from
baseline. To calculate the time to peak, we measured the
time after the auditory speech onset at which the signal
reached its maximum value. We also calculated the dura-
tion of the response curves. As a measure of response
duration, we used FWHM, which was calculated by finding
the width of the response curve at where the response is
at 50% of the peak amplitude. We calculated the response
onset, time to peak, and response duration for each trial
and then averaged across trials for each electrode.
Linear Mixed Effects Modeling
We used the lme4 package (Bates, Mächler, Bolker,
& Walker, 2014) available for the R statistical language
1046
Journal of Cognitive Neuroscience
Volume 29, Number 6
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
(R Core Team, 2015) to perform a linear mixed effect
(LME) analysis of the relationship between the neural
response and both fixed and random factors that may
influence the response. For the main LME analyses
(Tables 1–5), the fixed factors were the location of each
electrode (Anterior or Posterior), the presence or ab-
sence of auditory noise (Clear A or Noisy A), and the
presence or absence of visual noise Clear V of Noisy V.
The random factors were the mean response of each
electrode across all conditions and the stimulus exem-
plar. The use of stimulus exemplar as a random factor
accounts for differences in response to individual stimuli
and allows for inference beyond the levels of the factors
tested in the particular experiment (i.e., generalization to
other stimuli).
For each fixed factor, the LME analysis produced an
estimated effect in units of the dependent variable and
a standard error relative to a baseline condition (equiva-
lent to beta weights in linear regression). For the main
LME analyses, the baseline condition was always the re-
sponse to AV speech in anterior electrodes. The full re-
sults of all LME analyses and the baseline condition for
each analysis are shown in the tables and table legends.
Additional Experiment: Varying Levels of
Auditory Noise
In an additional control experiment, we recorded re-
sponses to audiovisual speech with varying levels of audi-
tory noise. Similar to the main experiment, for each
auditory word, noise that matched the spectrotemporal
power distribution of the auditory speech was generated,
then noise and the original auditory speech were added
together with different weights while keeping the total
power constant (Schepers et al., 2013). We parametrically
increased the amount of auditory noise in 11 steps from
0% to 100% in 10% increments. Forty-two to 44 repeti-
tions were presented for each noise level. The partici-
pant’s task was to discriminate between four different
words: Rain, Rock, Neck, and Mouth.
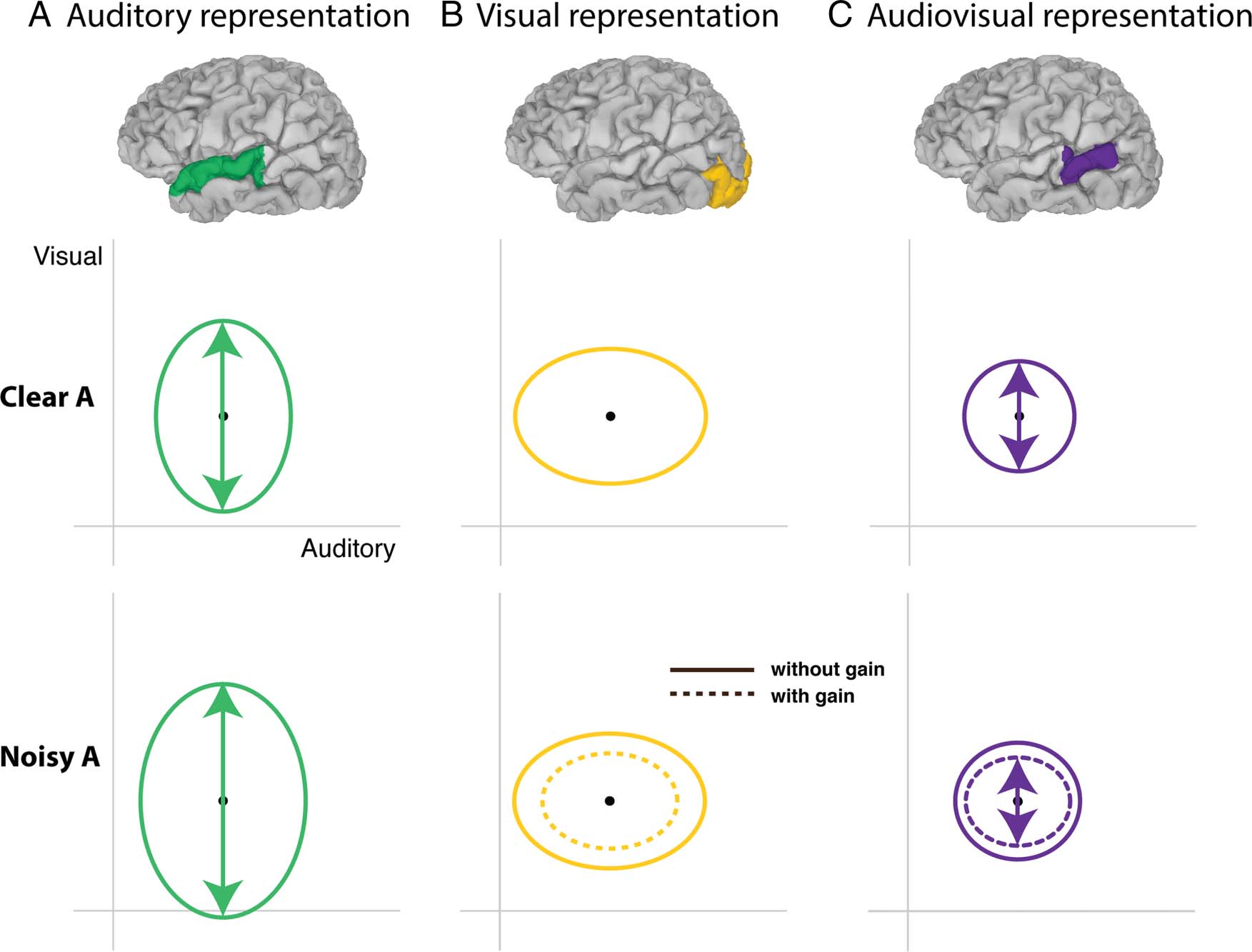
Model Creation
A simple Bayesian model was constructed to aid in interpre-
tation of the data (Figure 6) using a recently developed
model of human multisensory speech perception (Magnotti
& Beauchamp, 2017). Briefly, the high-dimensional neuro-
nal response vector is conceptualized as a point in 2-D
space. In this space, the x axis represents auditory feature
information and the y axis represents visual feature infor-
mation. Speech tokens are located at a fixed point in this
space (shown in Figure 6 as the black dot at the center of
each ellipse). For each presentation of an audiovisual
speech stimulus, the brain encodes the auditory and visual
information with noise. Over many trials, we characterize
the distribution of the encoded speech stimulus as an
ellipse. The axes of the ellipse correspond to the relative
precision of the representation along each axis. Modalities
are encoded separately, but through extensive experience
with audiovisual speech, encoding a unisensory speech
stimulus provides some information about the other
modality. Although the results are robust across a range
of parameters, for demonstration purposes, we assume
that the variability of the preferred to non-preferred
modality for audiovisual speech with a clear auditory
component is 2:1 (shown in Figure 6 as the asymmetry
of the ellipses in the auditory and visual representations).
The integrated representation is formed according to
Bayes rule, which combines the two modalities into a
single representation that has smaller variance than
−1 +
either of the component modalities: (cid:1)AV = ((cid:1)A
−1)−1 (Ma, Zhou, Ross, Foxe, & Parra, 2009). For audio-
(cid:1)V
visual speech with noisy auditory component, we assume
that the variability in the auditory representation increases
by 150% while keeping the relative variability at the
same ratio of 2:1 (shown in Figure 6 as larger ellipse).
We model the visual representation of speech with noisy
auditory component as being either identical to the rep-
resentation of speech with a clear auditory component
or with a gain term that reduces variability by 50% (with
the relative variability remaining at 2:1). The multisensory
representation is calculated in the same fashion with and
without gain.
RESULTS
Across participants, a total of 27 speech-responsive elec-
trodes were identified on the STG. Using the posterior
border of Heschl’s gyrus as an anatomical landmark, 16
of these electrodes were located over anterior STG and
11 electrodes were located over posterior STG (Figure 1A).
We hypothesized that the presence of noise in the speech
stimulus (Figure 1B–E) might differentially affect responses
in anterior and posterior electrodes. To test this hypothesis,
we used the response amplitude in the gamma band as the
dependent measure and fit a LME model with electrode
location (Anterior vs. Posterior), the presence or absence
of auditory noise in the stimulus (Clear A vs. Noisy A),
and the presence or absence of visual noise in the stimu-
lus (Clear V vs. Noisy V) as fixed factors. To account for
overall differences in response amplitude across elec-
trodes and stimulus exemplars, these were added to the
model as random factors.
Amplitude of the Responses to Clear and
Noisy Speech
As shown in Table 1, there were three significant effects
in the LME model. There was a small but significant effect
of electrode location ( p = .01) driven by a smaller overall
response in posterior electrodes (Anterior vs. Posterior:
136 ± 27% vs. 101 ± 24%, mean signal change from base-
line averaged across all stimulus conditions ± SEM ) and
Ozker et al.
1047
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
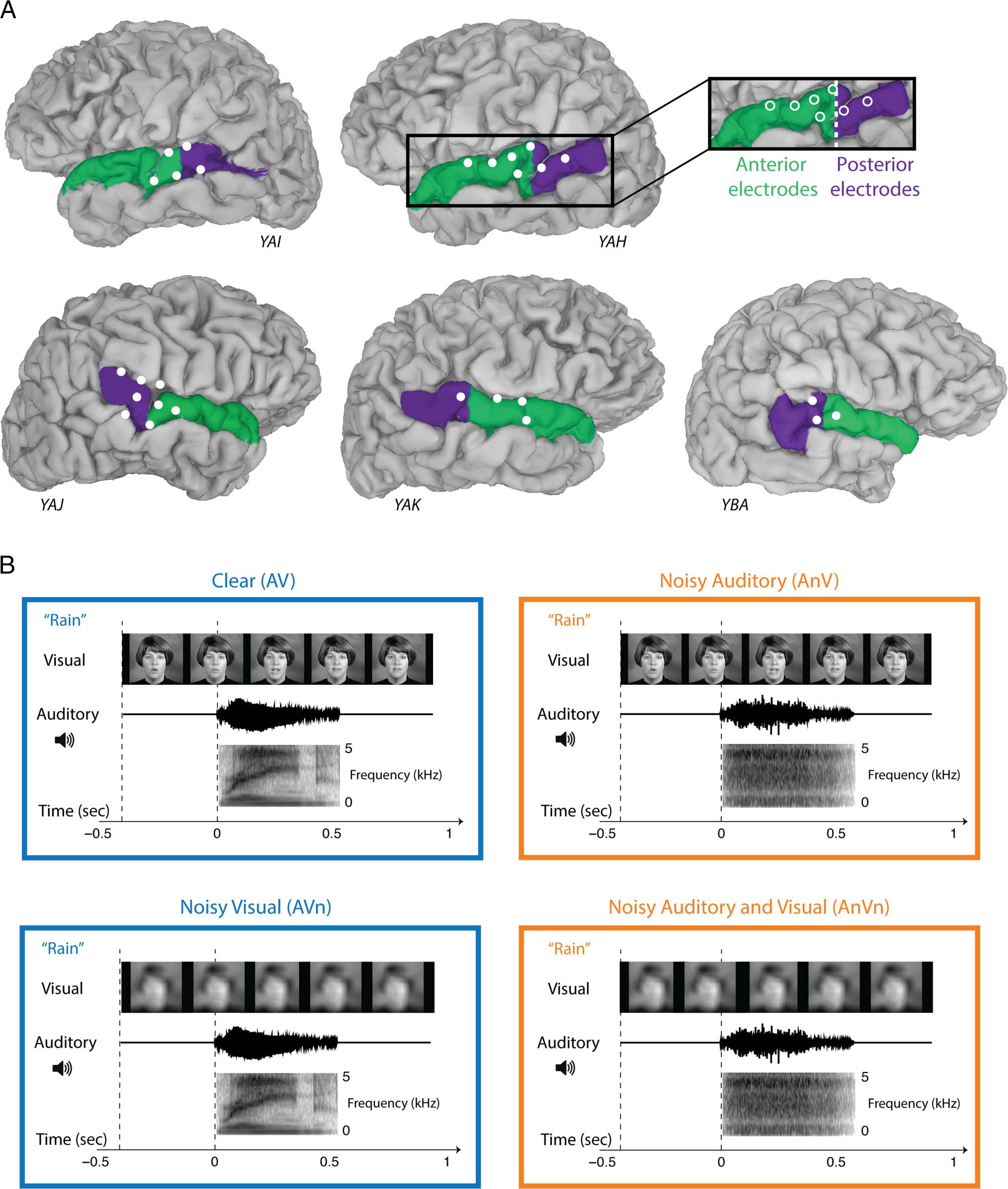
Figure 1. Electrode locations and audiovisual speech stimuli. (A) Cortical surface models of the brains of five participants (with anonymized subject
ID). White circles show the location of implanted electrodes with a significant response to speech stimuli in the left hemisphere (top row) and
right hemisphere (bottom row). In each hemisphere, the STG was parcellated into anterior (green) and posterior (purple) portions, demarcated by
the posterior-most portion of Heschl’s gyrus. (B) Clear audiovisual speech (AV) consisted of a movie of a talker pronouncing the word “rain” or
“rock.” Visual stimulus (top row) shows sample frames from the video. Auditory stimulus is shown as sound pressure level (middle row) and
spectrogram (bottom row). Black vertical dashed lines indicate visual and auditory stimulus onsets. For noisy auditory speech (AnV), the auditory
component was replaced with speech-specific noise of equal power to the original auditory speech. For noisy visual speech (AVn), the visual
component was blurred using a low-pass Gaussian filter. For noisy auditory and noisy visual speech (AnVn), the auditory component was replaced
with speech-specific noise and the visual component was blurred.
1048
Journal of Cognitive Neuroscience
Volume 29, Number 6
Table 1. Linear Mixed-effects Model of the Response Amplitude
Fixed Effects
Baseline
Auditory noise (An)
Posterior location × An
Posterior location
Visual noise ( Vn)
An × Vn
Posterior location × Vn
Posterior location × An × Vn
Estimate
183.1
−109.6
140.6
−101
21.6
−13.3
−8.9
3.6
SE
24.8
13.5
21.2
38.7
13.5
19.1
21.2
29.9
df
33.7
188
188
34.2
188
188
188
188
t
7.4
−8.1
6.6
−2.6
1.6
−0.7
−0.4
0.1
p
−8
10
10−13
10−10
.01
.11
.49
.67
.91
Results of an LME model of the response amplitude. The fixed effects were the location of each electrode (Anterior vs. Posterior), the presence or
absence of auditory noise (An) in the stimulus and the presence or absence of visual noise ( Vn) in the stimulus. Electrodes and stimulus exemplar
were included in the model as random factors. For each effect, the model estimates (in units of percent signal change) for that factor are shown
relative to baseline, the response in anterior electrodes to clear audiovisual speech (AV stimulus condition). The “SE” column shows the standard
error of the estimate. The degrees of freedom (“df”), t value, and p value derived from the model were calculated according to the Satterthwaite
approximation, as provided by the lmerTest package (Kuznetsova, Brockhoff, & Christensen, 2015). The baseline is shown first; all other effects are
ranked by absolute t value. Significant effects are shown in bold. The significance of the baseline fixed effect is grayed-out because it was prespe-
cified: only electrodes with significant amplitudes were included in the analysis.
two larger effects: the main effect of auditory noise ( p =
10−14) and the interaction between auditory noise and
the location of the electrode ( p = 10−10). Speech with
clear auditory components evoked a larger response than
speech with noisy auditory components (Clear A, consist-
ing of the average of the AV and AVn conditions, 151 ±
27% vs. Noisy A, consisting of the average of the AnV and
AnVn conditions, 93 ± 14%, mean ± SEM across elec-
trodes) driving the main effect of auditory noise. However,
the response patterns were very different in anterior
and posterior electrodes, leading to the significant inter-
action in the LME model (Figure 2A). Speech with clear
auditory components evoked a larger response than
speech with noisy auditory component in anterior elec-
trodes (Clear A vs. Noisy A: 194 ± 39% vs. 78 ± 16%,
mean ± SEM across electrodes) but speech with clear
auditory components evoked a smaller response than
speech with noisy auditory component in posterior elec-
trodes (88 ± 23% vs. 115 ± 25%).
To determine if the interaction between electrode loca-
tion and the response to auditory noise was consistent,
we plotted the amplitude of the response to Clear A
versus Noisy A for all electrodes using one symbol per
electrode (Figure 2B). All of the anterior electrodes lay
above the line of equality, indicating uniformly larger re-
sponses for Clear A, and all of the posterior electrodes lay
on or below the line of equality, indicating similar re-
sponses for Clear A and Noisy A.
To examine the interaction between location and audi-
tory noise in a single participant, we examined two elec-
trodes: an anterior electrode located just anterior to the
A–P boundary and an adjacent electrode located 10 mm
more posterior, just across the anterior–posterior bound-
ary (Figure 2C and D). In the anterior electrode, the re-
sponse to Clear A speech was much larger than the
response to Noisy A speech (Clear A vs. Noisy A: 461 ±
35% vs. 273 ± 21%, mean across trials ± SEM; unpaired
t test across trials: t(147) = 4.6, p = 10−6), whereas in
the adjacent posterior electrode, the response to Clear A
speech was similar to the response to Noisy A speech
(Clear A vs. Noisy A: 313 ± 21% vs. 349 ± 18%, t(147) =
1.3, p = .2). Hence, two electrodes located on either side
of the anterior–posterior boundary showed very different
patterns of responses to Clear A and Noisy A speech.
To examine the effect of anatomical location on the
response to Clear A and Noisy A speech in more detail,
we calculated each electrode’s location in a reference
frame defined by the STG (Figure 2E) and the difference
in the electrode’s response amplitude to Clear A and
Noisy A speech (Clear A − Noisy A). First, we examined
electrodes sorted by their medial-to-lateral position on
the STG and observed no discernible pattern (Figure 2F).
Second, we examined electrodes sorted by their anterior-
to-posterior position on the STG (Figure 2G). Anterior elec-
trodes showed uniformly positive values for Clear A – Noisy
A (Clear A) whereas posterior electrodes showed zero or
negative values for Clear A – Noisy A. However, we did not
observe a gradient of responses between more anterior and
more posterior electrodes, suggesting a sharp transition
across the A–P boundary rather than a gradual shift
in response properties along the entire extent of the STG.
To quantify this observation, we tested two simple models.
In the discrete model, there was a sharp transition between
response properties on either side of the A–P boundary; in
the continuous model, there was a gradual change
in response properties across the entire extent of the STG.
For the discrete model, we fit the amplitude versus
location points with two constants ( y = b; horizontal lines
with a fixed mean and zero slope, one mean for the ante-
rior electrodes and one for the posterior electrodes;
Ozker et al.
1049
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
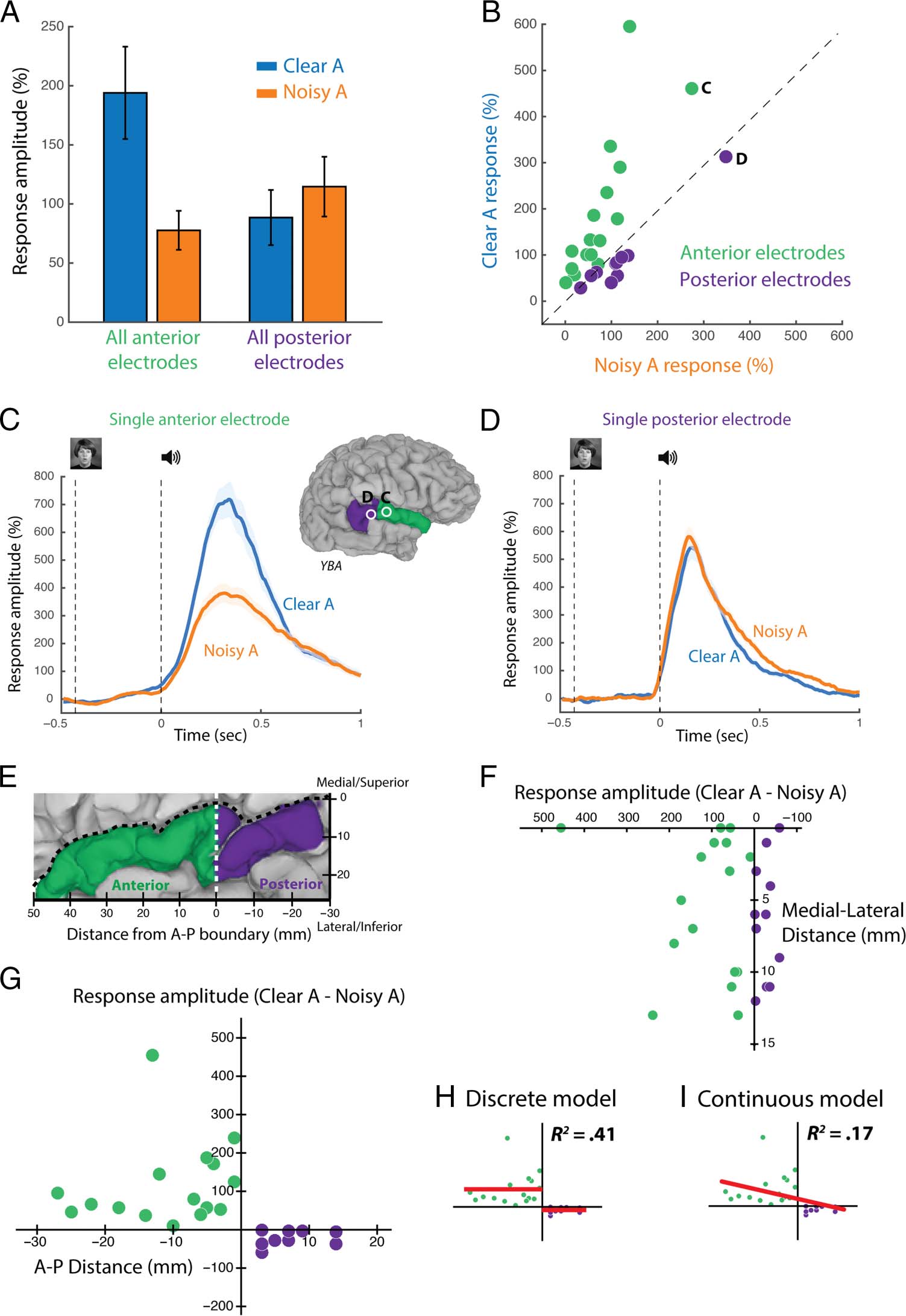
Figure 2. Response amplitudes.
(A) The response to speech
with clear auditory component
(Clear A, combination of AV
and AVn stimulus conditions)
and noisy auditory component
(Noisy A, combination of AnV
and AnVn conditions) collapsed
across electrodes (error bars
show SEM ). The response
amplitude is the mean percent
change in high-gamma power
(70–110 Hz) in the 0–500 msec
time window relative to
prestimulus baseline (−500 to
−100 msec). (B) The response
to Clear A versus Noisy A speech
for each individual electrode,
with each anterior electrode
shown as a green circle and
each posterior electrode shown
as a purple circle. The black
dashed line represents the line
of equality. Electrodes shown
in C and D are labeled.
(C) High-gamma response
to Clear A speech (blue trace)
and Noisy A speech (orange
trace) for a single anterior
electrode (labeled “C” in inset
brain). Shaded regions indicate
the SEM across trials. Black
vertical dashed lines indicate
visual and auditory stimulus
onsets, respectively. (D) High-
gamma response to Clear A
and Noisy A speech in a single
posterior electrode (labeled “D”
in inset brain). (E) Coordinate
system for STG measurements.
y Axis indicates distance from
medial/superior border of STG
(black dashed line); x axis
shows distance from the A–P
boundary (white dashed line).
(F) The response amplitude
to Clear A speech minus the
response amplitude to Noisy A
speech as a function of
distance from the medial/
superior border, one symbol
per electrode (anterior
electrodes in green, posterior
electrodes in purple). (G) The
response amplitude to Clear A
minus Noisy A speech as a function of distance from the A–P boundary. (H) Discrete model: Constant values were fit separately to the anterior and
posterior electrode data in G ( y = a and y = b), and the correlation with the data was calculated. (I) Continuous model: A linear model with two
parameters was fit to both anterior and posterior electrodes ( y = mx + b).
Figure 2H). For the continuous model, we fit the ampli-
tude versus location points with a single line ( y = mx +
b; Figure 2I). Both models fit the data using an equal num-
ber of parameters (2). The two models were compared
using R2 as a measure of the explained variance and Akaike
Information Criterion (AIC) as a measure of likelihood. The
discrete model fit the amplitude versus location points
much better than the continuous model (R2 = .41 vs.
.17), and the AIC revealed that the discrete model was
more than 100 times more likely to explain the observed
data (e(AIC continuous − AIC discrete)/2 = 102).
To allow easier comparison of the A–P boundary with
the functional neuroimaging literature, we converted each
participant’s brain into standard space and measured
1050
Journal of Cognitive Neuroscience
Volume 29, Number 6
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the coordinates of each electrode. The average location
in standard space of the Heschl’s gyrus landmark, the
boundary between the anterior and posterior STG ROIs,
was y = −27 ± 2 (mean across participants ± SD). The
mean position in standard space of all anterior electrodes
was (x = ±66, y = −18, z = 6), whereas for posterior
electrodes the mean position was (x = ±67, y = −34,
z = 12).
Variability of the Responses to Clear and
Noisy Speech
Theoretical models predict that combining the informa-
tion available about speech content from the auditory
and visual modalities should reduce neural variability
(Fetsch et al., 2012; Knill & Pouget, 2004); see discussion
and Figure 6 for more details. We hypothesized that the
presence of noise in the speech stimulus might differen-
tially affect the response variability in anterior and poste-
rior electrodes. To test this hypothesis, we fit the same
LME model used to examine response amplitude, except
that response variability (CV) was used as the dependent
measure. As shown in Table 2, there were three signifi-
cant effects in the LME model, including an effect of elec-
trode location ( p = .02) driven by a larger overall
response variability in posterior electrodes than in ante-
rior electrodes (Anterior vs. Posterior: 0.85 ± 24% vs.
0.99 ± 0.1, mean CV averaged across all stimulus condi-
tions ± SEM ). The other two effects showed a larger
effect size: the main effect of auditory noise ( p = 10−6)
and the interaction between auditory noise and the loca-
tion of the electrode ( p = 10−8).
Speech with Noisy A resulted in larger response variabil-
ity than speech with Clear A (Clear A vs. Noisy A: 0.89 ±
0.06 vs. 0.93 ± 0.06, mean ± SEM across electrodes) driv-
ing the main effect of auditory noise in the model. How-
ever, the response patterns were very different in anterior
and posterior electrodes, leading to the significant inter-
action (Figure 3A). Speech with noisy auditory compo-
nent resulted in a larger response variability than speech
with clear auditory component in anterior electrodes
(Clear A vs. Noisy A: 0.73 ± 0.05 vs. 0.96 ± 0.1, mean ±
SEM across electrodes) but speech with a noisy auditory
component resulted in a smaller response variability
than speech with a clear auditory component in posterior
electrodes (Clear A vs. Noisy A: 1.1 ± 0.1 vs. 0.9 ± 0.1).
To determine if the interaction between electrode
location and the response variability for auditory noise
was consistent, we plotted the variability of the response
to Clear A versus Noisy A for all electrodes using one
symbol per electrode (Figure 3B). Most of the anterior
electrodes lay below the line of equality, indicating larger
variability for Noisy A, whereas most of the posterior
electrodes lay above the line of equality, indicating smaller
variability for noisy A.
To demonstrate the effect at the single electrode level,
we examined the interaction between location and audi-
tory noise in a single participant, we examined two elec-
trodes: an anterior electrode and a posterior electrode
(Figure 3C and D). Figure 3C shows the normalized re-
sponses for a single anterior electrode for single trials
of speech with clear and noisy auditory components. In
this anterior electrode, there was variability across trials
in both conditions, but the variability was much greater
for speech with a noisy auditory component than for
speech with a clear auditory component (Clear A vs.
Noisy A: 1.1 vs. 1.7, unpaired t test across normalized trial
amplitudes: t(221) = 5.4, p = 10−7). In a posterior elec-
trode from the same participant (Figure 3B), the opposite
pattern was observed: The variability was much greater for
speech with a clear auditory component than for speech
with a noisy auditory component (Clear A vs. Noisy A: 1.4 vs.
0.9, t(221) = 5, p = 10−6). Hence, two electrodes located
on either side of the anterior–posterior boundary showed
very different patterns of response variability.
To examine the effect of anatomical location on var-
iability, we calculated the difference in each elec-
trode’s variability to Clear A and Noisy A speech (CV for
Table 2. Linear Mixed-effects Model of the Response Variability
Fixed Effects
Baseline
Posterior location × An
Auditory noise (An)
Posterior location
Posterior location × Vn
Posterior location × An × Vn
An × Vn
Visual noise ( Vn)
Estimate
0.76
−0.59
0.31
0.35
−0.13
0.15
0.03
0.01
SE
0.1
0.1
0.07
0.14
0.1
0.15
0.09
0.06
df
29.8
179.9
180.4
39.8
179.5
179.5
179.6
179.5
t
8
−5.7
4.6
2.5
−1.3
1
0.3
0.1
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
p
10−8
10−7
10−5
.02
.2
.31
.77
.89
Results of an LME model of the response variability measure as CV. The baseline for the model was the response in anterior electrodes to clear
audiovisual speech (AV stimulus condition). Baseline is shown first; all other effects are ranked by absolute t value. Significant effects are shown in bold.
Ozker et al.
1051
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
f
.
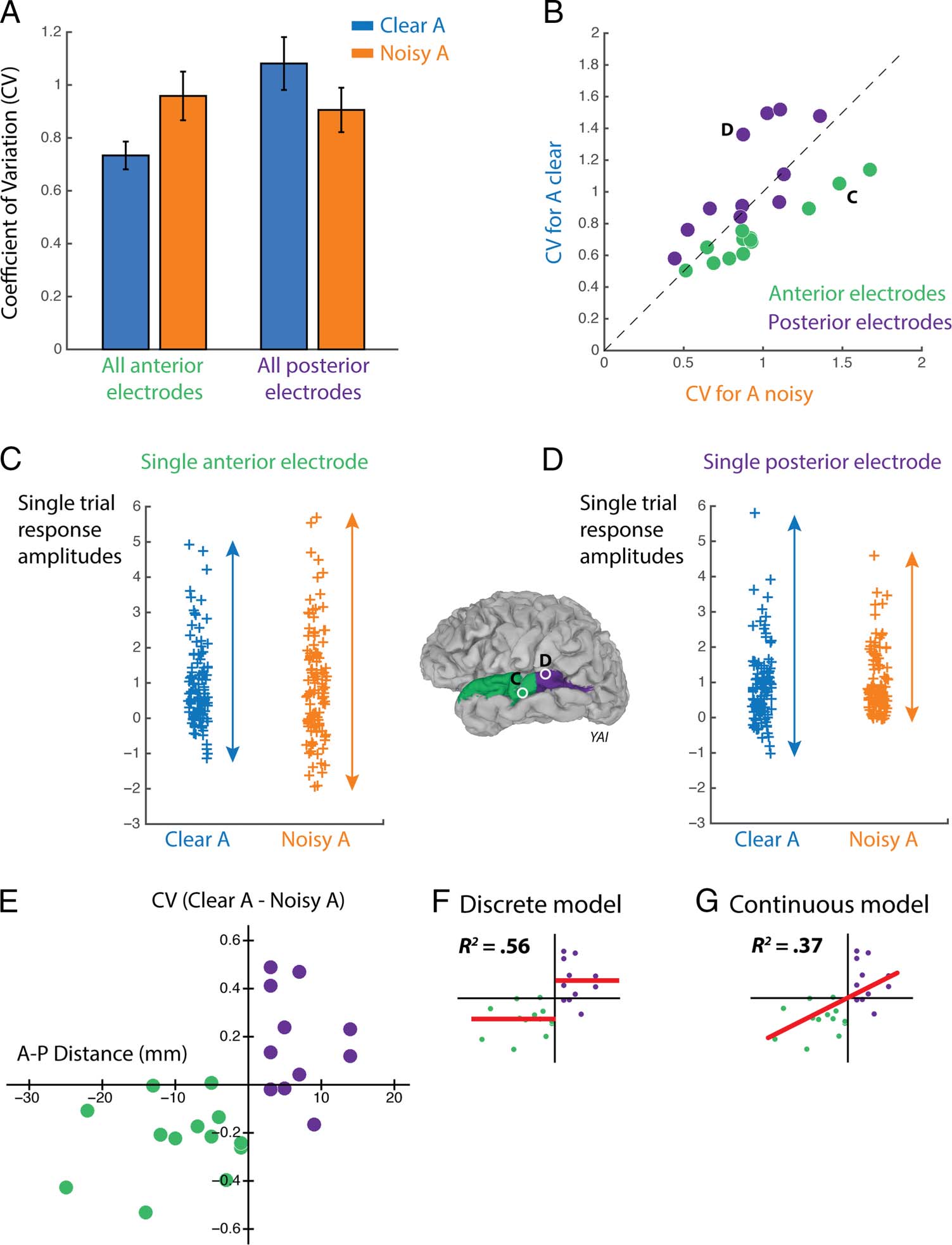
Figure 3. Response variability.
(A) Response variability to
speech with clear auditory
component (Clear A,
combination of AV and AVn
stimulus conditions) and noisy
auditory component (Noisy A,
combination of AnV and AnVn
conditions) collapsed across
electrodes (error bars show
SEM ). The response variability
was measured as the CV,
defined as the standard
deviation of the high-gamma
response divided by the mean
of the high-gamma response;
this measure accounts for the
differences in the mean
response between conditions
shown in Figure 2. (B) The
response variability to Clear A
versus Noisy A speech for each
individual electrode, with each
anterior electrode shown as a
green circle and each posterior
electrode shown as a purple
circle. The black dashed line
represents the line of equality.
Electrodes shown in C and D
are labeled. (C) High-gamma
response amplitudes to single
presentations of Clear A speech
(blue symbols) and Noisy A
speech (orange symbols) for a
single anterior electrode
(labeled “C” in inset brain),
normalized by the mean
response across trials (value of
one indicates a single trial
response equal to the mean
response across trials). Arrows
illustrate CV, a measure of
variability. (D) High-gamma
response amplitudes to single
presentations of speech for a
single posterior electrode
(labeled “D” in inset brain).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
Clear A − CV for Noisy A) and plotted it against that
electrode’s A–P location on the STG (Figure 3E). Paral-
leling the analysis performed on response amplitude,
discrete and continuous models were fit to the data
(Figure 3F and G). The discrete model fit the amplitude
versus location points much better than the continuous
model (R2 = .56 vs. .37) and the AIC revealed that the
discrete model was more likely to explain the observed
data (e(AIC continuous − AIC discrete)/2 = 74). Hence, the dif-
ference in response variability between electrodes is
more accurately described as arising from two groups
(Anterior and Posterior) with categorically different var-
iability rather than as a continuous change in variability
from anterior to posterior.
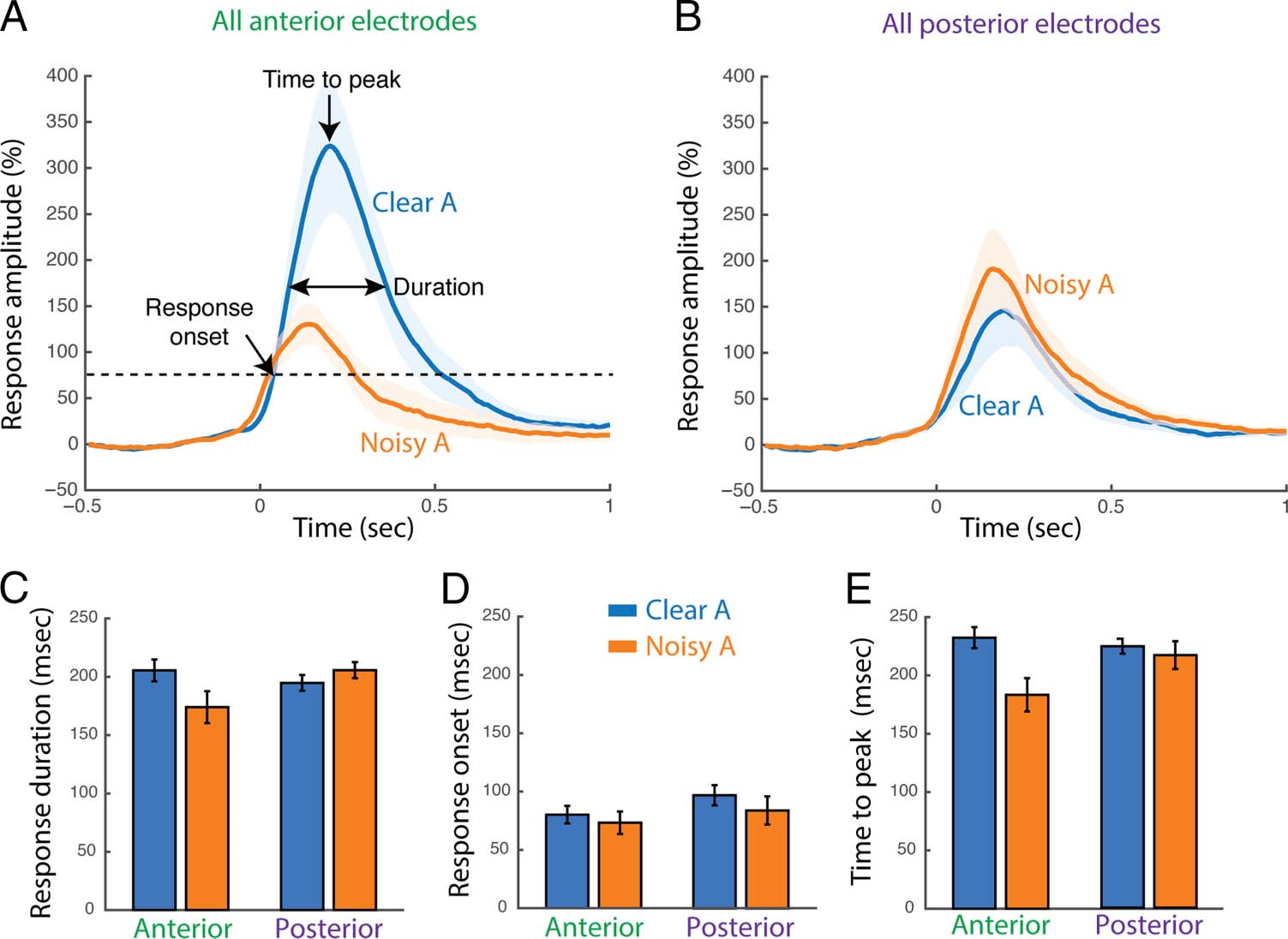
Timing of the Responses to Clear and Noisy Speech
The high temporal resolution of electrocorticography
allows for examination of the detailed timing of the neu-
ronal responses. Figure 4 (A and B) show the average re-
sponse of anterior and posterior electrodes to Clear A and
Noisy A speech. In anterior electrodes, the high-gamma
response to Clear A speech started at 77 msec after audi-
tory stimulus onset, reached half-maximum amplitude at
110 msec, peaked at 210 msec, and returned to the half-
maximum value at 290 msec, resulting in a total response
duration (measured as the FWHM) of 190 msec.
To determine the effects of auditory noise and elec-
trode location on the timing of the neuronal response,
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1052
Journal of Cognitive Neuroscience
Volume 29, Number 6
Figure 4. Response timing.
(A) High-gamma response
amplitudes to Clear A and Noisy
A speech averaged across all
anterior electrodes, shown as
percent signal change from
baseline relative to time from
auditory stimulus onset (error
bars show SEM ). Three
measures of the response were
calculated. Response onset time
is the first time point at which
the signal deviates three
standard deviations from
baseline. Time to peak is the
time point of maximal response
amplitude. Duration indicates
the time between the first and
last time points at which the
response is equal to half of its
maximum value (FWHM).
(B) High-gamma response
amplitudes to Clear A and Noisy
A speech averaged across all
posterior electrodes. (C) The
response duration for Clear A
versus Noisy A speech in
anterior electrodes (left) and
posterior electrodes (right).
Error bars show SEM. (D) The
response onset in anterior
and posterior electrodes.
(E) The time to peak in anterior
and posterior electrodes.
for each electrode we estimated response duration, onset
time, and time-to-peak and separately fit three LME
models with each temporal variable as the dependent
measure. For the LME model with response duration as
the dependent measure (Table 3 and Figure 4C) the only
significant effects were the main effect of auditory noise
( p = 10−5) and the interaction between auditory noise
and electrode location ( p = 10−5). These effects were
driven by an overall longer response duration for Clear
A speech than for Noisy A speech (Clear A vs. Noisy A:
194 ± 6 msec vs. 187 ± 9 msec, mean across electrodes ±
SEM), with anterior electrodes showing longer responses
for Clear A speech (Clear A vs. Noisy A: 205 ± 9 msec vs.
174 ± 14 msec) and posterior electrodes showing shorter
responses for Clear A speech (Clear A vs. Noisy A: 195 ±
7 msec vs. 206 ± 7 msec).
For the LME model with response onset as the depen-
dent measure, there were no significant main effects or
Table 3. Linear Mixed-effects Model of the Response Duration
Fixed Effects
Baseline
Posterior location × An
Auditory noise (An)
Posterior location
Posterior location × Vn
Posterior location × An × Vn
Visual noise ( Vn)
An × Vn
Estimate
206.2
48.6
−30.9
−15.1
8.9
−12.2
−1.4
−1.3
SE
9.6
10.9
7
15.1
10.9
15.5
7
9.9
df
41.4
189
189
41.4
189
189
189
189
t
21.4
4.4
−4.4
−1
0.8
−0.8
−0.2
−0.1
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
p
10−16
10−5
10−5
.32
.42
.43
.84
.89
Results of an LME model of the response duration. The baseline for the model was the response in anterior electrodes to clear audiovisual speech
(AV stimulus condition). Baseline is shown first; all other effects are ranked by absolute t value. Significant effects are shown in bold.
Ozker et al.
1053
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
Table 4. Linear Mixed-effects Model of the Response Onset
Fixed Effects
Estimate
SE
df
t
p
Baseline
81.5
9.2 27.6
8.8 10−9
Posterior location
17.6
13.6
41.3
1.3
Posterior location × An −9.1
9.8 187.9 −0.9
An × Vn
Auditory noise (An)
Visual noise ( Vn)
−7.1
−2.6
−2.6
8.8 187.9 −0.8
6.3 187.9 −0.4
6.3 187.9 −0.4
Posterior location ×
5
13.9 187.9
0.4
.2
.35
.42
.68
.68
.72
An × Vn
Posterior location × Vn −1.3
9.8 187.9 −0.1
.9
Results of an LME model of the response onset. The baseline for the
model was the response in anterior electrodes to clear audiovisual
speech (AV stimulus condition). Baseline is shown first; all other effects
are ranked by absolute t value. No factors were significant. Significant
effects are shown in bold.
interactions (Table 4 and Figure 4D). For the LME model
with time-to-peak as the dependent measure (Table 5
and Figure 4E), there was a significant main effect of
auditory noise ( p = 10−9) and an interaction between
auditory noise and electrode location ( p = 10−4 ) driven
by a longer time-to-peak for Clear A speech (Clear A vs.
Noisy A: 229 ± 6 msec vs. 197 ± 10 msec, mean across
electrodes ± SEM ), more so in anterior electrodes (Clear
A vs. Noisy A: 232 ± 9 msec vs. 183 ± 14 msec) than pos-
terior electrodes (Clear A vs. Noisy A: 224 ± 6 msec vs.
216 ± 12 msec).
Relationship between Neuronal Responses and
Perceptual Accuracy
Participants performed a task that required them to re-
spond to the identity of the word present in each trial.
Across participants, only AnVn trials consistently gen-
erated enough errors to compare correct and incorrect
trials (AV: 99 ± 3%, AVn: 98 ± 3%, AnV: 81 ± 20%, AnVn:
63 ± 15%; % correct, mean across participants ± SD). To
determine the relationship between neuronal response
amplitude and behavioral accuracy within AnVn trials,
an LME model was constructed with response amplitude
as the dependent measure, electrode location (Anterior vs.
Posterior), and behavioral accuracy (Correct vs. Incorrect)
as fixed factors and stimulus exemplar, participant, and
electrode (nested within participant) as random factors
(Table 6). In the LME model, the only significant effect
was an interaction between electrode location and behav-
ioral accuracy ( p = .01) driven by smaller amplitudes in
correct trials for anterior electrodes (Correct vs. Incorrect:
84 ± 15% vs. 93 ± 20%, mean gamma power signal change
relative to baseline across electrodes ± SEM) but larger
amplitudes in correct trials for posterior electrodes (Correct
vs. Incorrect: 122 ± 27% vs. 106 ± 26%). A similar model
with CV as the dependent measure did not show any sig-
nificant effects (Table 7).
Potential Confound: Intelligibility
We observed very different neuronal responses to audio-
visual speech with noisy auditory component in anterior
compared with posterior electrodes, attributing this dif-
ference to the differential contributions of anterior and
posterior STG to multisensory integration. However, we
used only high levels of auditory noise in our audiovisual
speech stimuli. To determine how the level of auditory
noise influenced the effect, in one patient we presented
audiovisual speech with 11 different levels of auditory
noise and examined the neural response in two electrodes
located on either side of the anterior–posterior boundary
(Figure 5A). First, we examined how these data compared
with our previous results by collapsing the 11 different
levels of noise into just two categories “low noise” (0–
40% noise levels) and “high noise” (50–100% noise levels)
Table 5. Linear Mixed-effects Model of the Response Peak Time
Fixed Effects
Baseline
Auditory noise (An)
Posterior location × An
Posterior location
Posterior location × Vn
Visual noise ( Vn)
Posterior location × An × Vn
An × Vn
Estimate
234.3
−46.5
45.5
−12.5
8.7
−3.9
−8.4
−4.9
SE
10.4
7.4
11.5
15.8
11.5
7.4
16.3
10.4
df
36
187.9
187.9
41.6
187.9
187.9
187.9
187.9
t
22.6
−6.3
3.9
−0.8
0.8
−0.5
−0.5
−0.5
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
p
10−16
10−9
10−4
.44
.45
.6
.61
.64
Results of an LME model of the response peak time. The baseline for the model was the response in anterior electrodes to clear audiovisual speech
(AV stimulus condition). Baseline is shown first; all other effects are ranked by absolute t value. Significant effects are shown in bold.
1054
Journal of Cognitive Neuroscience
Volume 29, Number 6
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
Table 6. Linear Mixed-effects Model of the Effect of Accuracy on Response Amplitude
Fixed Effects
Baseline
Incorrect responses × Posterior location
Incorrect responses
Posterior location
Estimate
105.2
−25.6
11.3
19.6
SE
36.1
10.2
6.6
21.8
df
4.2
65.8
66.1
22.8
t
2.9
−2.5
1.7
0.9
p
.04
.01
.09
.38
Results of an LME model on the relationship between response amplitude and behavioral accuracy for auditory noise, visual noisy audiovisual speech
(AnVn stimulus condition). The fixed effects were the location of each electrode (Anterior vs. Posterior) and the behavioral accuracy of the par-
ticipant’s responses (Correct vs. Incorrect). Participants, electrodes nested in participants, and stimulus exemplar were included in the model as
random factors. The baseline for the model was the response in anterior electrodes for correct behavioral responses. Baseline is shown first; all other
effects are ranked by absolute t value. Significant effects are shown in bold.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
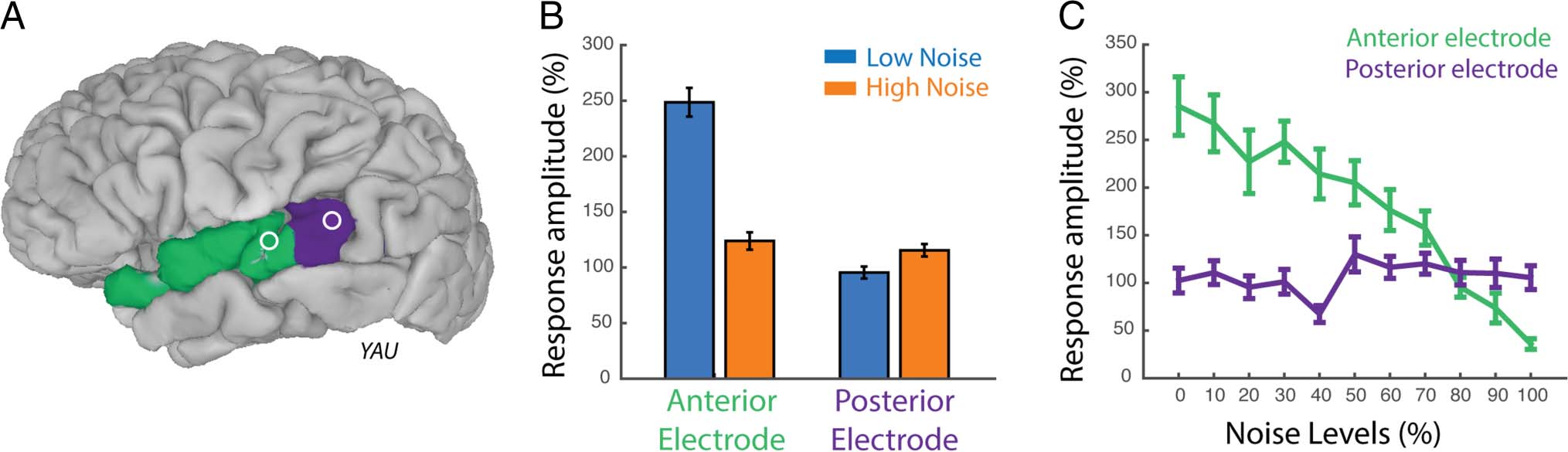
similar to our initial analysis of Clear A and Noisy A audio-
visual speech (Figure 5B). The responses were similar to
that observed with just two levels of noise (compare
Figure 5B and Figure 2A). An LME model fit to the data
across the different noise levels (Table 8) showed significant
effects of noise level ( p = 10−16), electrode location ( p =
10−16), and an interaction between noise level and location
( p = 10−16), driven by significantly greater response in an-
terior electrodes to low noise stimuli (Low vs. High: 248 ±
13% vs. 124 ± 8%, mean across trials ± SEM) and similar
responses in posterior electrodes to low and high noise
conditions (Low vs. High: 95 ± 5% vs. 115 ± 5%). Next,
we examined the response to each different level of audi-
tory noise. In the anterior electrode, increasing levels of
auditory noise led to smaller responses, whereas in the
posterior electrode, increasing levels of auditory noise
led to similar or slightly larger gamma band responses
(Figure 5C). We quantified this by fitting a line to the
anterior and posterior electrode responses at 11 different
auditory noise levels. The anterior electrode fit was sig-
nificant (R2 = .9, p = 10−6) with a negative slope (m =
−24), whereas the posterior electrode fit was not signif-
icant (R2 = .07, p = .4) with a slightly positive slope (m =
1.32).
The participant performed at a high level of accuracy
even in trials with a high level of auditory noise (zero
errors) demonstrating that the visual speech information
was able to compensate for the increased levels of auditory
noise.
DISCUSSION
We observed a double dissociation in the responses to
audiovisual speech with clear and noisy auditory compo-
nents for both amplitude and variability measures. In
anterior STG, the amplitude of the high-gamma response
was greater for speech with clear auditory components
than for speech with noisy auditory components, whereas
in posterior STG, responses were similar or slightly
greater for speech with noisy auditory component. In
anterior STG, the CV across single trials was greater for
speech with noisy auditory component, whereas in pos-
terior STG, it was greater for speech with clear auditory
components.
These data are best understood within the framework
of Bayes optimal models of multisensory integration
(Alais & Burr, 2004; Ernst & Banks, 2002) and speech per-
ception (Bejjanki, Clayards, Knill, & Aslin, 2011; Ma et al.,
2009). In these models, different sensory modalities are
posited to contain independent sources of environmen-
tal and sensory noise. Because of the independence of
noise sources across modality, Bayesian integration re-
sults in a multisensory representation that has smaller
variance than either of the unisensory variances (Fetsch
et al., 2012; Knill & Pouget, 2004).
Recently, a Bayesian model of causal inference in audio-
visual speech perception was proposed (Magnotti &
Beauchamp, 2017). Figure 6 shows an application of this
model to our data. We assume that anterior STG contains
Table 7. Linear Mixed-effects Model of the Effect of Accuracy on Response Variability
Fixed Effects
Baseline
Posterior location
Incorrect responses
Incorrect responses × Posterior location
Estimate
1
−0.13
0.02
0.01
SE
0.19
0.2
0.11
0.17
df
7
31.3
67.1
66.5
t
5.4
0.6
0.2
0.1
p
10−3
.53
.86
.95
Results of an LME model on the relationship between response variability (CV) and behavioral accuracy for noisy auditory and noisy visual speech
(AnVn stimulus condition). The baseline for the model was the response in anterior electrodes for correct behavioral responses. Baseline is shown
first; all other effects are ranked by absolute t value. No factors were significant. Significant effects are shown in bold.
Ozker et al.
1055
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 5. Response amplitude with varying levels of auditory noise. (A) The location of an anterior and a posterior electrode in a single participant.
(B) The response amplitude in the anterior electrode (left bars) and posterior electrode (right bars) to audiovisual speech with low levels of
auditory noise (Low Noise: 0% to 40%) and high levels of auditory noise (High Noise: 50% to 100%) averaged across trials (error bars show SEM ).
(C) Response amplitude for the anterior and posterior electrodes at each of 11 different auditory noise levels (0–100%) averaged across trials
(error bars show SEM ).
a unisensory representation of auditory speech, that ex-
trastriate visual areas contain a representation of visual
speech, and that posterior STG contains a representa-
tion of multisensory speech formed by integrating inputs
from unisensory auditory and visual areas (Bernstein &
Liebenthal, 2014; Nath & Beauchamp, 2011). The neural
implementation of Bayes optimal integration is thought
to rely on probabilistic population codes (Angelaki, Gu,
& DeAngelis, 2009; Ma et al., 2006) in which pools of
neurons encode individual stimuli in a probabilistic fash-
ion. These population codes are modeled as Gaussians
in which amplitude and variability are inversely related.
A smaller, more focal Gaussian indicates larger ampli-
tude and less variability in the population code, whereas
a larger Gaussian indicates smaller amplitude and more
variability.
For audiovisual speech with a clear auditory compo-
nent (Clear A), the neural population code in anterior
STG has a given amplitude and variability. When auditory
noise is added (Noisy A), the population code amplitude
decreases and the variability increases (Ma et al., 2006),
an accurate description of the response in anterior STG
for noisy compared with clear auditory speech.
For the visual representation in lateral extrastriate cor-
tex, the visual information is the same in the Clear A and
Noisy A conditions, predicting similar population codes
for both conditions (Figure 6B). For the multisensory
representation in posterior STG, the population code is
calculated as the optimal integration of the response in
auditory and visual representations. The visual informa-
tion serves to compensate for the increased auditory
noise in the Noisy A condition, so that the population
code for the integrated representation is only slightly
broader for Noisy A than Clear A speech, a match to
the observation that the amplitude and variability of the
response to Noisy A and Clear A speech are much more
similar in posterior STG than they are in anterior STG.
A close inspection of the data shows that, contrary to
Bayesian models, the response in posterior STG was
slightly more focal (30% greater amplitude and 16%
reduced variability) for Noisy A compared with Clear A
conditions. Although counterintuitive, this result is con-
sistent with evidence that visual cortex responds more
to noisy than clear audiovisual speech (Schepers, Yoshor,
& Beauchamp, 2015). This enhancement may be attribut-
able to top–down modulation from higher-level areas
Table 8. Linear Model of the Effect of Varying Auditory Noise Levels on Response Amplitude
Fixed Effects
Baseline
Posterior location
High auditory noise
High auditory noise × Posterior location
Estimate
248.5
−153.1
−124.8
144.8
SE
8.6
12.2
11.6
16.5
t
28.9
−12.6
−10.7
8.8
p
−16
10
10−16
10−16
10−16
Results of a linear model of the response amplitude for varying auditory noise levels in a single participant. Responses in individual trials were used as
samples. Electrode location (Anterior vs. Posterior) and noise level (Low vs. High) were used as factors. The baseline for the model was the response
in anterior electrodes to audiovisual speech with low auditory noise. Baseline is shown first; all other effects are ranked by absolute t value. Significant
effects are shown in bold. The significance of the baseline fixed effect is grayed-out because it was prespecified: Only electrodes responding to this
condition were included in the analysis.
1056
Journal of Cognitive Neuroscience
Volume 29, Number 6
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 6. Bayesian model of audiovisual speech with auditory noise. (A) The model assumes that a neural representation of the auditory component
of audiovisual speech exists in anterior STG (top row: brain region colored green). The high-dimensional neural representation is projected onto
a 2-D space (middle and bottom rows) in which the x axis represents auditory feature information and the y axis represents visual feature
information. The stimulus representation is shown as an ellipse indicating the cross-trial variability in representation of an identical physical stimulus
due to sensory noise. For audiovisual speech with clear auditory component (Clear A) in anterior STG (green ellipse in middle row), there is
less variability along the auditory axis and more variability along the visual axis, indicated by the shape of the ellipse. For audiovisual speech with
noisy auditory component (Noisy A) in anterior STG (green ellipse in bottom row), there is greater variability along both axes due to the added
stimulus noise (see Methods for details). (B) The model assumes that a neural representation of the visual component of audiovisual speech exists in
lateral extrastriate visual cortex (top row: brain region colored yellow). In the visual representation, there is less variability along the visual axis and
more variability along the auditory axis, indicated by the shape of the ellipse. For audiovisual speech with noisy auditory component (Noisy A),
the visual component of the speech is identical, so the representation should be identical (yellow ellipse in bottom row). However, evidence from
Schepers et al. (2015) demonstrates that response in visual cortex to Noisy A speech is actually greater than to Clear A speech, suggesting an increase
in gain due to attentional modulation or other top–down factors. The representation with gain modulation is shown with the dashed yellow ellipse.
(C) The model assumes that a neural representation that integrates both auditory and visual components of audiovisual speech exists in posterior
STG (top row: brain region colored purple). Because of the principles of Bayesian integration, this representation has smaller variability than either
the auditory representation or the visual representation (compare size of purple ellipse in each row with green and yellow ellipses). Assuming gain
modulation, the integrated representation of Noisy A speech (dashed purple ellipse in bottom row) has smaller variability than the representation
of Clear A speech (purple ellipse in middle row).
that increase the gain in visual cortex, similar to atten-
tional modulation in which representations in visual cor-
tex are heightened and/or sharpened by spatial or
featural attention (Maunsell & Treue, 2006; Kastner,
Pinsk, De Weerd, Desimone, & Ungerleider, 1999). This
gain increase would be adaptive because it would
increase the likelihood of decoding speech from visual
cortex under conditions of low or no auditory infor-
mation, at the cost of additional deployment of atten-
tional and neural resources. We implemented this gain
modulation in our Bayesian model as reduced variance
in the visual representation for Noisy A compared with
Clear A speech. When this visual representation with re-
duced variance is integrated with the noisy auditory rep-
resentation, the resulting multisensory representation
becomes more focal for Noisy A than Clear A speech, a
fit to the observed increased amplitude and reduced vari-
ability for Noisy A compared with Clear A speech in pos-
terior STG.
Although the Bayesian model provides a conceptual
framework for understanding how multisensory inte-
gration could affect the amplitude and variance of neuro-
nal population responses, it is agnostic about the actual
stimulus features important for integration. We did not
Ozker et al.
1057
observe a main effect of visual noise (or an interaction
between visual noise and auditory noise) LME on ampli-
tude and variance (Tables 1 and 2). Most of the relevant
information provided by the visual signal during auditory–
visual speech perception is related to the timing of mouth
opening and closing relative to auditory speech. The blur-
ring procedure used to generate the noisy visual speech
may leave this timing information intact.
Our Bayesian model also does not make explicit pre-
dictions about the latency or duration of the neuronal re-
sponse. However, we observed the same pattern of
double dissociation between anterior and posterior STG
for response duration as in other response measures. At
the high levels of auditory noise used in our experiments,
the auditory representation contains little useful infor-
mation, so it would be adaptive for top–down modula-
tion to decrease both the amplitude and duration of
activity in the anterior STG for Noisy A speech. Interest-
ingly, the absolute duration of the response in posterior
STG during Noisy A speech was the same as the absolute
duration of the response in anterior STG during Clear A
speech (210 msec), raising the possibility that this is the
time frame of the selection process in which the com-
peting unisensory and multisensory representations are
selected for perception and action.
An interaction between electrode location and re-
sponse amplitude was also observed in an analysis of per-
ceptual accuracy (only speech with both noisy auditory
and noisy visual component generates enough errors
for this analysis). In anterior electrodes, responses were
larger for incorrect trials, whereas in posterior electrode
responses were larger for correct trials. This supports the
idea that posterior electrodes are particularly important
in the perception of noisy speech, with larger amplitude
indicating a more focal peak of activity in the population
code and less uncertainty about the presented stimulus.
Anterior versus Posterior
Anatomical Specialization
There was a strikingly sharp boundary between the ante-
rior and posterior response patterns, suggesting that
anterior and posterior STG are functionally distinct. We
divided STG at the posterior border of Heschl’s gyrus
(mean y = −27), a landmark that also has been used
in previous neuroimaging studies of speech processing
(Okada et al., 2010; Specht & Reul, 2003; Hickok & Poeppel,
2000). A functional division in STG near Heschl’s gyrus is
consistent with the division of the auditory system into
two processing streams, one of which runs anterior-ventral
from Heschl’s gyrus and one of which runs posterior-dorsal
(Pickles, 2015; Rauschecker, 2015). These two streams are
often characterized as specialized for processing “what” or
object identity features (anterior-ventral) and “where” or
object location features (posterior-dorsal) by analogy
with the different streams of visual processing (Mishkin
& Ungerleider, 1982). However, these labels do not
neatly map onto an anterior preference for clear speech
and a posterior preference for noisy speech (Leonard &
Chang, 2014) and may reflect preferences for different
rates of spectrotemporal modulation (Hullett, Hamilton,
Mesgarani, Schreiner, & Chang, 2016).
Although we are not aware of previous studies exam-
ining changes in the neural variability to Clear A and
Noisy A audiovisual speech, a number of neuroimaging
studies have reported A–P differences in the amplitude
of the neural response to Clear A and Noisy A audiovisual
speech. Stevenson and James (2009) presented clear au-
diovisual speech and audiovisual speech with noise
added to both modalities (noisy auditory + noisy visual),
contrasting both against a standard baseline condition con-
sisting of simple visual fixation. Anterior regions of STG/
STS showed greater responses to clear than noisy audiovi-
sual speech (Figure 5C and Table 3 in their paper, y = −20
compared with y = −18 in this study, mean across left and
right hemispheres). Their results slightly differ from ours
because they showed that posterior STG/STS (Figure 5D
and Table 1 in their paper, y = −37, compared with y =
−34 in this study) displays relatively weak responses
to moderately noisy speech. This could be explained by
their use of noisy auditory + noisy visual speech versus
our use of noisy auditory + clear visual speech: If poste-
rior regions respond to both auditory and visual speech
information, degraded visual information might be ex-
pected to reduce response amplitudes in posterior regions.
Consistent with these results, Lee and Noppeney (2011)
found that anterior STG/STS ( y = −16, their Table 2)
showed significant audiovisual interactions only for clear
speech, whereas posterior STG/STS (mean y = −36,
Table 2 in their paper) showed interactions for both clear
and noisy audiovisual speech.
Bishop and Miller (2009) reported greater responses
to clear versus noisy audiovisual speech in anterior re-
gions of STG (Table 1 in their paper, y = −13 mean
across left and right hemispheres), whereas McGettigan
and colleagues (2012) reported greater responses for
clear than noisy audiovisual speech in both anterior STG
( y = −12, Table 1 in their paper) and posterior STG
( y = −42).
Although most neuroimaging studies have reported
greater responses to clear than noisy audiovisual speech,
two studies have reported the opposite result of greater
responses to noisy speech in the STG (Callan et al., 2003;
Sekiyama, Kanno, Miura, & Sugita, 2003). However, the
interpretation of these studies is complex. Sekiyama
and colleagues tested clear and noisy speech consisting
of incongruent audiovisual speech (including McGurk
syllables) that are known to evoke responses in STS that
are both different from congruent syllables and vary
markedly from participant to participant (Erickson
et al., 2014; Nath & Beauchamp, 2012). Callan and col-
leagues performed an analysis in which they first sub-
tracted the response to auditory-only clear speech from
the response to audiovisual clear speech, then subtracted
1058
Journal of Cognitive Neuroscience
Volume 29, Number 6
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the response to auditory-only noisy speech from the
response to audiovisual noisy speech, and finally sub-
tracted the two differences. Without a direct comparison
between clear and noisy audiovisual speech, it is possible
that the reported preference for noisy audiovisual speech
was driven by the intermediate analysis step in which the
auditory-only response was subtracted from the audio-
visual response. For instance, even if clear and noisy
audiovisual speech evoked the exact same response, a
weak response to auditory-only noisy speech and a strong
response to auditory-only clear speech (a pattern observed
in a number of studies, see below) would result in the re-
ported greater response to noisy audiovisual speech.
The idea of an A–P double dissociation is also gen-
erally supported by the neuroimaging literature exam-
ining brain responses to clear and noisy auditory-only
speech, although the many differences in the stimulus
materials, task manipulations, and data analysis strategies
makes direct comparisons difficult. Obleser, Zimmermann,
Van Meter, and Rauschecker (2007) reported a double
dissociation, with posterior regions ( y = −26, Table 1 in
their paper) preferring noisy speech to clear speech,
whereas anterior regions ( y = −18) preferred clear
speech to noisy speech. A double dissociation was also
reported by Du, Buchsbaum, Grady, and Alain (2014):
Anterior regions of STG ( y = −15, Table S2 in their paper)
showed greater BOLD amplitude with less auditory noise,
whereas posterior regions ( y = −32) showed greater
BOLD amplitude with more auditory noise. Similarly,
Wild, Davis, and Johnsrude (2012) found that anterior re-
gions of STG ( y = −12, Table 1 in their paper) preferred
clear to noisy speech, whereas posterior regions ( y =
−30) preferred noisy speech to clear speech.
Single dissociations consistent with an anterior prefer-
ence for clear speech are also common in the literature.
Scott, Blank, Rosen, and Wise (2000) found that anterior
regions ( y = −12) showed greater response amplitudes
for clear speech, whereas posterior regions ( y = −38,
Figure 2A in their paper) showed similar response ampli-
tudes. Giraud and colleagues (2004) also reported greater
response amplitudes for clear than noisy speech in ante-
rior STG (Table 1 in their paper, y = −4 mean across left
and right hemispheres) but not posterior STG.
Reprint requests should be sent to Michael S. Beauchamp,
Department of Neurosurgery and Core for Advanced MRI,
Baylor College of Medicine, 1 Baylor Plaza, S104, Houston, TX
77030, or via e-mail: michael.beauchamp@bcm.edu.
REFERENCES
Alais, D., & Burr, D. (2004). The ventriloquist effect results from
near-optimal bimodal integration. Current Biology, 14, 257–262.
Angelaki, D. E., Gu, Y., & DeAngelis, G. C. (2009). Multisensory
integration: Psychophysics, neurophysiology, and computation.
Current Opinion in Neurobiology, 19, 452–458.
Argall, B. D., Saad, Z. S., & Beauchamp, M. S. (2006). Simplified
intersubject averaging on the cortical surface using SUMA.
Human Brain Mapping, 27, 14–27.
Bates, D., Mächler, M., Bolker, B., & Walker, S. (2014). Fitting
linear mixed-effects models using lme4. arXiv preprint
arXiv:1406.5823.
Beauchamp, M. S., Lee, K. E., Argall, B. D., & Martin, A. (2004).
Integration of auditory and visual information about objects
in superior temporal sulcus. Neuron, 41, 809–823.
Bejjanki, V. R., Clayards, M., Knill, D. C., & Aslin, R. N. (2011).
Cue integration in categorical tasks: Insights from audio-
visual speech perception. PLoS One, 6, e19812.
Bernstein, L. E., Auer, E. T., & Takayanagi, S. (2004). Auditory
speech detection in noise enhanced by lipreading. Speech
Communication, 44, 5–18.
Bernstein, L. E., & Liebenthal, E. (2014). Neural pathways for
visual speech perception. Frontiers in Neuroscience, 8, 386.
Binder, J. R., Frost, J. A., Hammeke, T. A., Bellgowan, P. S.,
Springer, J. A., Kaufman, J. N., et al. (2000). Human temporal
lobe activation by speech and nonspeech sounds. Cerebral
Cortex, 10, 512–528.
Bishop, C. W., & Miller, L. M. (2009). A multisensory cortical
network for understanding speech in noise. Journal of
Cognitive Neuroscience, 21, 1790–1805.
Callan, D. E., Jones, J. A., Munhall, K., Callan, A. M., Kroos, C., &
Vatikiotis-Bateson, E. (2003). Neural processes underlying
perceptual enhancement by visual speech gestures.
NeuroReport, 14, 2213–2218.
Calvert, G. A., Campbell, R., & Brammer, M. J. (2000). Evidence
from functional magnetic resonance imaging of crossmodal
binding in the human heteromodal cortex. Current Biology,
10, 649–657.
Churchland, M. M., Yu, B. M., Cunningham, J. P., Sugrue, L. P.,
Cohen, M. R., Corrado, G. S., et al. (2010). Stimulus onset
quenches neural variability: A widespread cortical
phenomenon. Nature Neuroscience, 13, 369–378.
Cox, R. W. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers
in Biomedical Research, 29, 162–173.
Dale, A. M., Fischl, B., & Sereno, M. I. (1999). Cortical surface-
based analysis. I. Segmentation and surface reconstruction.
Neuroimage, 9, 179–194.
Desikan, R. S., Segonne, F., Fischl, B., Quinn, B. T., Dickerson,
B. C., Blacker, D., et al. (2006). An automated labeling
system for subdividing the human cerebral cortex on MRI
scans into gyral based regions of interest. Neuroimage,
31, 968–980.
Destrieux, C., Fischl, B., Dale, A., & Halgren, E. (2010).
Automatic parcellation of human cortical gyri and sulci using
standard anatomical nomenclature. Neuroimage, 53, 1–15.
Du, Y., Buchsbaum, B. R., Grady, C. L., & Alain, C. (2014).
Noise differentially impacts phoneme representations in the
auditory and speech motor systems. Proceedings of the
National Academy of Sciences, U.S.A., 111, 7126–7131.
Erickson, L. C., Zielinski, B. A., Zielinski, J. E., Liu, G.,
Turkeltaub, P. E., Leaver, A. M., et al. (2014). Distinct cortical
locations for integration of audiovisual speech and the
McGurk effect. Frontiers in Psychology, 5, 534.
Ernst, M. O., & Banks, M. S. (2002). Humans integrate visual
and haptic information in a statistically optimal fashion.
Nature, 415, 429–433.
Fetsch, C. R., Pouget, A., DeAngelis, G. C., & Angelaki, D. E. (2012).
Neural correlates of reliability-based cue weighting during
multisensory integration. Nature Neuroscience, 15, 146–154.
Fischl, B., Sereno, M. I., & Dale, A. M. (1999). Cortical surface-
based analysis. II: Inflation, flattening, and a surface-based
coordinate system. Neuroimage, 9, 195–207.
Foxe, J. J., Wylie, G. R., Martinez, A., Schroeder, C. E., Javitt,
D. C., Guilfoyle, D., et al. (2002). Auditory-somatosensory
multisensory processing in auditory association cortex: An
fMRI study. Journal of Neurophysiology, 88, 540–543.
Ozker et al.
1059
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Giraud, A. L., Kell, C., Thierfelder, C., Sterzer, P., Russ, M. O.,
Preibisch, C., et al. (2004). Contributions of sensory input,
auditory search and verbal comprehension to cortical activity
during speech processing. Cerebral Cortex, 14, 247–255.
Gur, M., Beylin, A., & Snodderly, D. M. (1997). Response
variability of neurons in primary visual cortex ( V1) of alert
monkeys. Journal of Neuroscience, 17, 2914–2920.
Hickok, G., & Poeppel, D. (2000). Towards a functional
neuroanatomy of speech perception. Trends in Cognitive
Sciences, 4, 131–138.
Hullett, P. W., Hamilton, L. S., Mesgarani, N., Schreiner, C. E., &
Chang, E. F. (2016). Human superior temporal gyrus
organization of spectrotemporal modulation tuning derived
from speech stimuli. Journal of Neuroscience, 36, 2014–2026.
Jacques, C., Witthoft, N., Weiner, K. S., Foster, B. L., Rangarajan,
V., Hermes, D., et al. (2016). Corresponding ECoG and fMRI
category-selective signals in human ventral temporal cortex.
Neuropsychologia, 83, 14–28.
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R., &
Ungerleider, L. G. (1999). Increased activity in human visual
cortex during directed attention in the absence of visual
stimulation. Neuron, 22, 751–761.
Knill, D. C., & Pouget, A. (2004). The Bayesian brain: The role of
uncertainty in neural coding and computation. Trends in
Neurosciences, 27, 712–719.
Kuznetsova, A., Brockhoff, P. B., & Christensen, R. H. B. (2015).
Package “lmerTest”. R package version, 2.0-29.
Lee, H., & Noppeney, U. (2011). Physical and perceptual factors
shape the neural mechanisms that integrate audiovisual
signals in speech comprehension. The Journal of
Neuroscience, 31, 11338–11350.
Leonard, M. K., & Chang, E. F. (2014). Dynamic speech
representations in the human temporal lobe. Trends in
Cognitive Sciences, 18, 472–479.
Ma, W. J., Beck, J. M., Latham, P. E., & Pouget, A. (2006).
Bayesian inference with probabilistic population codes.
Nature Neuroscience, 9, 1432–1438.
Ma, W. J., Zhou, X., Ross, L. A., Foxe, J. J., & Parra, L. C. (2009).
Lip-reading aids word recognition most in moderate noise: A
Bayesian explanation using high-dimensional feature space.
PLoS One, 4, e4638.
Nir, Y., Fisch, L., Mukamel, R., Gelbard-Sagiv, H., Arieli, A.,
Fried, I., et al. (2007). Coupling between neuronal firing rate,
gamma LFP, and BOLD fMRI is related to interneuronal
correlations. Current Biology, 17, 1275–1285.
Obleser, J., Zimmermann, J., Van Meter, J., & Rauschecker, J. P.
(2007). Multiple stages of auditory speech perception reflected
in event-related fMRI. Cerebral Cortex, 17, 2251–2257.
Okada, K., Rong, F., Venezia, J., Matchin, W., Hsieh, I. H., Saberi,
K., et al. (2010). Hierarchical organization of human auditory
cortex: Evidence from acoustic invariance in the response to
intelligible speech. Cerebral Cortex, 20, 2486–2495.
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J. M. (2011).
FieldTrip: Open source software for advanced analysis of MEG,
EEG, and invasive electrophysiological data. Computational
Intelligence and Neuroscience, 2011, 156869.
Pickles, J. O. (2015). Auditory pathways: Anatomy and
physiology. Handb Clin Neurol, 129, 3–25.
Rauschecker, J. P. (2015). Auditory and visual cortex of
primates: A comparison of two sensory systems. European
Journal of Neuroscience, 41, 579–585.
Ray, S., & Maunsell, J. H. (2011). Different origins of gamma
rhythm and high-gamma activity in macaque visual cortex.
PLoS Biology, 9, e1000610.
Reale, R. A., Calvert, G. A., Thesen, T., Jenison, R. L., Kawasaki, H.,
Oya, H., et al. (2007). Auditory-visual processing represented
in the human superior temporal gyrus. Neuroscience, 145,
162–184.
Ross, L. A., Saint-Amour, D., Leavitt, V. M., Javitt, D. C., & Foxe,
J. J. (2007). Do you see what I am saying? Exploring visual
enhancement of speech comprehension in noisy
environments. Cerebral Cortex, 17, 1147–1153.
Schepers, I. M., Schneider, T. R., Hipp, J. F., Engel, A. K., &
Senkowski, D. (2013). Noise alters beta-band activity in
superior temporal cortex during audiovisual speech
processing. Neuroimage, 70, 101–112.
Schepers, I. M., Yoshor, D., & Beauchamp, M. S. (2015).
Electrocorticography reveals enhanced visual cortex
responses to visual speech. Cerebral Cortex, 25, 4103–4110.
Scott, S. K., Blank, C. C., Rosen, S., & Wise, R. J. S. (2000).
Identification of a pathway for intelligible speech in the left
temporal lobe. Brain, 123, 2400–2406.
Magnotti, J. F., & Beauchamp, M. S. (2017). Causal inference
Sekiyama, K., Kanno, I., Miura, S., & Sugita, Y. (2003). Auditory-
explains perception of the McGurk effect and other incongruent
audiovisual speech. PLoS Computational Biology, 13, e1005229.
Maunsell, J. H. R., & Treue, S. (2006). Feature-based attention
in visual cortex. Trends in Neurosciences, 29, 317–322.
McGettigan, C., Faulkner, A., Altarelli, I., Obleser, J., Baverstock,
H., & Scott, S. K. (2012). Speech comprehension aided by
multiple modalities: Behavioural and neural interactions.
Neuropsychologia, 50, 762–776.
Mesgarani, N., Cheung, C., Johnson, K., & Chang, E. F. (2014).
Phonetic feature encoding in human superior temporal
gyrus. Science, 343, 1006–1010.
Miller, L. M., & D’Esposito, M. (2005). Perceptual fusion and
stimulus coincidence in the cross-modal integration of
speech. Journal of Neuroscience, 25, 5884–5893.
Mishkin, M., & Ungerleider, L. G. (1982). Contribution of striate
inputs to the visuospatial functions of parieto-preoccipital
cortex in monkeys. Behavioural Brain Research, 6, 57–77.
Mukamel, R., Gelbard, H., Arieli, A., Hasson, U., Fried, I., & Malach,
R. (2005). Coupling between neuronal firing, field potentials,
and fMRI in human auditory cortex. Science, 309, 951–954.
Nath, A. R., & Beauchamp, M. S. (2011). Dynamic changes in
superior temporal sulcus connectivity during perception of noisy
audiovisual speech. Journal of Neuroscience, 31, 1704–1714.
Nath, A. R., & Beauchamp, M. S. (2012). A neural basis
for interindividual differences in the McGurk effect, a
multisensory speech illusion. Neuroimage, 59, 781–787.
visual speech perception examined by fMRI and PET.
Neuroscience Research, 47, 277–287.
Sheffert, S. M., Lachs, L., & Hernandez, L. R. (1996). Research
on spoken language processing progress report no. 21
(pp. 578–583). Bloomington, IN: Speech Research Laboratory,
Indiana University.
Specht, K., & Reul, J. (2003). Functional segregation of the
temporal lobes into highly differentiated subsystems for
auditory perception: An auditory rapid event-related fMRI-task.
Neuroimage, 20, 1944–1954.
Stevenson, R. A., & James, T. W. (2009). Audiovisual integration
in human superior temporal sulcus: Inverse effectiveness and
the neural processing of speech and object recognition.
Neuroimage, 44, 1210–1223.
Sumby, W. H., & Pollack, I. (1954). Visual contribution to
speech intelligibility in noise. Journal of the Acoustical
Society of America, 26, 212–215.
Tolhurst, D. J., Movshon, J. A., & Dean, A. F. (1983). The
statistical reliability of signals in single neurons in cat and
monkey visual cortex. Vision Research, 23, 775–785.
van Atteveldt, N., Formisano, E., Goebel, R., & Blomert, L.
(2004). Integration of letters and speech sounds in the
human brain. Neuron, 43, 271–282.
Wild, C. J., Davis, M. H., & Johnsrude, I. S. (2012). Human
auditory cortex is sensitive to the perceived clarity of speech.
Neuroimage, 60, 1490–1502.
1060
Journal of Cognitive Neuroscience
Volume 29, Number 6
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
6
2
9
1
/
0
6
4
/
4
1
1
0
9
4
5
4
2
/
6
1
1
7
5
8
o
6
c
3
n
2
_
9
a
/
_
j
0
o
1
c
1
n
1
0
_
a
p
_
d
0
1
b
1
y
1
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1