Transcutaneous Auricular Vagus Nerve Stimulation
Strengthens Semantic Representations of Foreign
Language Tone Words during Initial Stages of Learning
Ian Phillips1,2,3
, Regina C. Calloway1, Valerie P. Karuzis1, Nick B. Pandža1,4,
Polly O’Rourke1, and Stefanie E. Kuchinsky1,2
Abstract
■ Difficulty perceiving phonological contrasts in a second
language (L2) can impede initial L2 lexical learning. Such is
the case for English speakers learning tonal languages, like
Mandarin Chinese. Given the hypothesized role of reduced neu-
roplasticity in adulthood limiting L2 phonological perception,
the current study examined whether transcutaneous auricular
vagus nerve stimulation (taVNS), a relatively new neuromodula-
tory technique, can facilitate L2 lexical learning for English
speakers learning Mandarin Chinese over 2 days. Using
a double-blind design, one group of participants received
10 min of continuous priming taVNS before lexical training
and testing each day, a second group received 500 msec of
peristimulus (peristim) taVNS preceding each to-be-learned
item in the same tasks, and a third group received passive sham
stimulation. Results of the lexical recognition test administered
at the end of each day revealed evidence of learning for all
groups, but a higher likelihood of accuracy across days for the
peristim group and a greater improvement in response time be-
tween days for the priming group. Analyses of N400 ERP compo-
nents elicited during the same tasks indicate behavioral
advantages for both taVNS groups coincided with stronger
lexico-semantic encoding for target words. Comparison of these
findings to pupillometry results for the same study reported in
Pandža, N. B., Phillips, I., Karuzis, V. P., O’Rourke, P., and
Kuchinsky, S. E. (Neurostimulation and pupillometry: New
directions for learning and research in applied linguistics.
Annual Review of Applied Linguistics, 40, 56–77, 2020) suggest
that positive effects of priming taVNS (but not peristim taVNS)
on lexico-semantic encoding are related to sustained attentional
effort. ■
INTRODUCTION
Few individuals who begin learning a foreign or second
language (L2) as adults ultimately attain native-like percep-
tion of target language phonological categories (e.g., Díaz,
Mitterer, Broersma, & Sebastián-Gallés, 2012; Abrahamsson
& Hyltenstam, 2009; Long, 1990). This finding is often
attributed to early closure of a sensitive period for phono-
logical development, after which it becomes increasingly
difficult to perceive and produce new phonological
contrasts (e.g., Kuhl, 2010; Mattock, Molnar, Polka,
& Burnham, 2008). Enhanced neuroplasticity—the ability
of neural circuits in the brain to change in response to
experience—is thought to underlie the rapid learning
that characterizes sensitive periods (for a review, see
White, Hutka, Williams, & Moreno, 2013). As first lan-
guage (L1) phonemic categories become entrenched, a
diminished capacity for neural reorganization affects
1University of Maryland Applied Research Laboratory for
Intelligence & Security, College Park, MD, 2Walter Reed
National Military Medical Center, Audiology and Speech
Pathology Center, Bethesda, MD, 3The Geneva Foundation,
Bethesda, MD, 4University of Maryland Program in Second
Language Acquisition, College Park, MD
© 2021 Massachusetts Institute of Technology
the degree to which new categories are learned
( Werker & Hensch, 2015; Kuhl, 2004), which has conse-
quences across adult L2 learning domains, including the
lexicon. The ability to learn new L2 words is generally not
thought to be maturationally constrained (e.g., Hellman,
2011; but see Granena & Long, 2013, for other age-
related lexical constraints); however, difficulty perceiving
nonnative phonological contrasts impedes initial L2 lexi-
cal learning (Poltrock, Chen, Kwok, Cheung, & Nazzi,
2018; Cooper & Wang, 2012; Wong & Perrachione,
2007) and can have persistent negative effects on L2 lex-
ical processing (Pelzl, Lau, Guo, & DeKeyser, 2021; Ling
& Grüter, 2020; Pelzl, 2019; Sebastián-Gallés, Echeverría,
& Bosch, 2005). To test whether enhancing neuroplasticity
facilitates initial learning of L2 words featuring nonnative
phonological contrasts, we administered transcutaneous
auricular vagus nerve stimulation (taVNS)—a relatively
new approach to inducing neuroplasticity by electrically
stimulating peripheral afferent fibers of the vagus nerve—
to native speakers of English while they learned a set of
Mandarin pseudowords distinguished in part by lexical
tone—a phonological contrast notoriously difficult for
speakers of nontonal languages to master. In a previous
article, we reported novel findings that taVNS modulates
attentional effort and improves lexical learning in this
Journal of Cognitive Neuroscience 34:1, pp. 127–152
https://doi.org/10.1162/jocn_a_01783
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
paradigm (Pandža, Phillips, Karuzis, O’Rourke, &
Kuchinsky, 2020). In this study, we build on these find-
ings and report results for concurrently recorded
electrophysiological data that further illuminate the
mechanisms by which taVNS benefits L2 lexical learning.
Over half the world’s languages are tonal ( Yip, 2002),
using differences in pitch to distinguish word meaning, as
illustrated in the following common example from
Mandarin Chinese, which has four contrastive tones plus
a fifth, neutral tone ( Wang, Spence, Jongman, & Sereno,
1999): /ma/ means “mother” when produced with a high
flat tone (Tone 1), “hemp” when produced with a tone
rising from mid to high (Tone 2), “to scold” when pro-
duced with a tone falling from high to low (Tone 4),
and “horse” when produced with a tone that falls and
then rises (Tone 3). Studies of short-term vocabulary
training, focused on associating meanings with small sets
of highly controlled tone words, show naive learners can
achieve high levels of accuracy identifying tone words
heard in isolation over several training sessions
(Antoniou & Wong, 2016; Perrachione, Lee, Ha, &
Wong, 2011; Chandrasekaran, Sampath, & Wong, 2010).
Illustrating the important role of L2 phonetic perceptual
ability and lexical learning, short-term lexical tone per-
ception training has been shown to facilitate tone word
learning (Cooper & Wang, 2013; Ingvalson, Barr, &
Wong, 2013) similarly to benefits of musical experience
and ability in discriminating and identifying both linguis-
tic and nonlinguistic pitch categories (e.g., Poltrock et al.,
2018; Bowles, Chang, & Karuzis, 2016; Dittinger et al.,
2016; Wong & Perrachione, 2007).
During initial stages of L2 lexical learning, evidence of
lexico-semantic development following fairly limited
word exposure has been obtained using implicit mea-
sures, such as the N400 ERP component (e.g., Dittinger
et al., 2016; Pu, Holcomb, & Midgley, 2016). The N400 is
a negative-going deflection usually largest over centro-
parietal scalp regions that peaks around 400 msec follow-
ing word onset and is sensitive to a word’s expectancy or
plausibility (Nieuwland et al., 2020; Kutas & Federmeier,
2011; Kutas & Hillyard, 1980). The difference in N400
amplitude between unexpected and expected words
given a preceding context, dubbed the N400 effect, re-
flects ease of lexico-semantic processing. Items that match
an expectation set by the preceding context have reduced
N400 amplitude and are easier to access and/or integrate
than unexpected items (Lau, Phillips, & Poeppel, 2008).
During word learning, increases in N400 amplitude
(Borovsky, Elman, & Kutas, 2012; McLaughlin, Osterhout,
& Kim, 2004) and shifts from frontal to centro-parietal to-
pography with repeated word exposure are also thought
to reflect the development of semantic representations
(Dittinger et al., 2016; Rodríguez-Fornells, Cunillera,
Mestres-Missé, & de Diego-Balaguer, 2009). Frontal effects
early during learning may reflect increased working
memory or cognitive control required to access a new
item’s semantic representation (Elgort, Perfetti, Rickles,
& Stafura, 2015; Mestres-Missé, Rodriguez-Fornells, &
Münte, 2007; Rodríguez-Fornells, De Diego Balaguer, &
Münte, 2006) with demands lessening as items become
lexicalized.
In a study of the impact of musical training on initial L2
word learning, Dittinger et al. (2016) used the N400 to
measure lexico-semantic development over a single training
session in which French-speaking professional musicians
and nonmusicians learned L2 Thai words distinguished
by several nonnative phonological contrasts, including lex-
ical tone. Although musicians outperformed nonmusi-
cians in only two behavioral tests of word learning, N400
results indicated faster and better learning for musicians
across all tasks: Only musicians showed an increase in
N400 amplitude at centro-parietal electrodes during the
passive word learning task (indicating faster lexico-
semantic encoding) and a centro-parietal N400 effect in
the short-term matching task (larger amplitude for unex-
pected vs. expected words, indicating stronger semantic
encoding) and semantic relatedness task (larger ampli-
tude for unrelated vs. related words, indicating stronger
integration of target words in existing semantic networks).
These findings and those of other studies (e.g., Borovsky
et al., 2012; Perfetti, Wlotko, & Hart, 2005; McLaughlin
et al., 2004) illustrate the ability of the N400 to track initial
development of lexical knowledge, even before behavioral
changes occur, and support using the N400 to evaluate
effects of taVNS on initial lexico-semantic encoding.
Electrical stimulation of the vagus, the tenth cranial
nerve within the autonomic nervous system, has been ex-
plored over the past 30 years for treating neurological
and neuropsychiatric disorders, including refractory epi-
lepsy and depression. Findings from this research show
that direct vagus nerve stimulation administered via im-
planted devices (iVNS) can alter autonomic nervous sys-
tem activity (e.g., Desbeaumes Jodoin, Lespérance,
Nguyen, Fournier-Gosselin, & Richer, 2015) and improve
cognitive function, including learning and memory (e.g.,
Sun et al., 2017; Clark, Naritoku, Smith, Browning, &
Jensen, 1999; see Vonck et al., 2014, for a summary).
More recently, applying low voltage electrical stimulation
via taVNS to the skin of the external auditory canal, inner
tragus, or cymba conchae, which are innervated to varying
degrees by the auricular branch of the vagus nerve (Butt,
Albusoda, Farmer, & Aziz, 2020), has been shown to in-
crease activity in vagal brainstem projections in adult hu-
mans, including the nucleus of the solitary tract and the
locus coeruleus (LC; Yakunina, Kim, & Nam, 2017;
Frangos, Ellrich, & Komisaruk, 2015; Kraus et al., 2013)
and to confer similar cognitive benefits for healthy adults
as compared to iVNS (e.g., Jacobs, Riphagen, Razat,
Wiese, & Sack, 2015).
The exact mechanism by which VNS influences cogni-
tion is not fully understood, but evidence suggests in-
creased LC activity plays an important role. The LC is
the primary source of the neurotransmitter norepineph-
rine (NE) in cortex, modulating various cortical and
128
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
subcortical circuits involved in arousal, attention, sensory
processing, and memory formation (Berridge &
Waterhouse, 2003). The LC-NE system is thought to influ-
ence memory formation by regulating NE in the hippo-
campus and prefrontal cortex that supports long-term
potentiation (LTP; Vonck et al., 2014), and is thought
to optimize task performance by shifting between two
modes of neural activity that modulate responsivity of
task-relevant cortical circuits: a slow baseline, tonic pat-
tern of firing that shifts arousal state, and a more rapid,
task-evoked phasic burst of activity that facilitates task-
relevant responses and is maximal at moderate levels of
tonic LC activity (Aston-Jones & Cohen, 2005). The LC-NE
system also modulates basal forebrain cholinergic activity,
which is thought to play a critical role in regulating atten-
tional effort necessary to maintain performance in difficult
tasks by providing top–down support via the activation of
attentional systems and related executive functions
(Klinkenberg, Sambeth, & Blokland, 2011; Sarter, Gehring,
& Kozak, 2006).
Enhancing specific cognitive functions and learning via
VNS likely depends on a complex interplay of these neu-
romodulatory circuits (Hulsey, Shedd, Sarker, Kilgard, &
Hays, 2019; Hulsey et al., 2016) and the degree to which
each system contributes to plasticity and learning may
depend on VNS timing. It has been found that taVNS im-
proves associative memory when delivered continuously
during encoding and consolidation phases of an associa-
tion memory task ( Jacobs et al., 2015) and L2 letter–
sound mapping when paired with learning feedback
(Thakkar, Engelhart, Khodaparast, Abadzi, & Centanni,
2020). In a study of Mandarin lexical tone perception,
perceptual categorization was improved for tones that
were paired with taVNS during training but not for tones
that occurred in the same training task but were not
paired with taVNS (Llanos et al., 2020), which parallels
findings in animal models of iVNS affecting auditory pro-
cessing only when temporally coupled to stimuli
(Engineer, Engineer, Riley, Seale, & Kilgard, 2015).
The potential for taVNS to support L2 lexical learning
both by inducing more global, slow-changing effects in
tonic LC-NE activity and more rapid, transient effects in
LC-NE phasic activity motivated the comparison of two
taVNS timings in this study: delivering taVNS continuously
for 10 min before naive L2 Mandarin learners (L1 English)
completed tone categorization and lexical learning tasks
( priming taVNS ) or for 500 msec before each to-be-
learned item within the same tasks ( peristimulus [peri-
stim] taVNS ). We previously reported behavioral and
pupillometry results for this study in Pandža et al.
(2020) that showed better learning for priming and peri-
stim taVNS over a passive sham taVNS control group and
smaller task-evoked pupil responses for peristim taVNS,
reflecting reductions in the allocation of cognitive effort
as participants engaged in successful learning. These
findings provide some of the first evidence linking
taVNS-related learning improvements to changes in
cognitive effort; however, it has yet to be shown how
these differences in effort allocation relate to the develop-
ment of L2 lexical knowledge. This study takes a first step in
addressing this question by analyzing N400 ERP compo-
nents elicited during the lexical learning tasks reported
in Pandža et al. (2020) and interpreting them in the con-
text of the previous behavioral and pupillometry results.
With this analysis, we sought to answer two research
questions (RQs): RQ1. Does taVNS support initial devel-
opment of lexico-semantic representations for L2 words
as evidenced by N400 amplitude and topography? RQ2.
Do priming and peristim taVNS have differential effects
on initial L2 lexico-semantic development? For RQ1, we
hypothesized that both taVNS timings would lead to
more robust lexico-semantic encoding for novel L2 words
earlier in training, reflected in stronger centro-parietal
N400s earlier during learning and larger N400 effects in
later recognition testing. For RQ2, we hypothesized that
priming and peristim taVNS might have differential ef-
fects on N400 amplitude and topography given potential
timing-related differences in underlying neuromodula-
tory mechanisms, but we could not make specific predic-
tions because of the lack of literature characterizing VNS
timing effects on the N400 component.
METHODS
This study was approved by the University of Maryland’s
Institutional Review Board and the U.S. Department of
Navy Human Research Protection Program. Priming and
peristim taVNS were tested in separate experiments that
used identical materials and procedures except for as-
pects of taVNS delivery, described below in taVNS
Parameters. Participants completed either the priming
or the peristim experiment with active taVNS (priming
or peristim) compared to a separate passive taVNS
(sham) control group in each experiment. To directly
compare the effects of priming to peristim taVNS in this
study, all data were combined into a single analysis. The
method and results are reported for the overall study
with differences between the two experiments noted
when necessary.
Participants
Participants gave informed consent before enrolling in
this study and were paid for their time. Eighty-two partic-
ipants completed the study. All reported being right-
handed, native speakers of English, with normal or
corrected-to-normal vision, normal hearing, no previous
exposure to any tone languages, and no history of psy-
chological or neurological disorders (see Pandža et al.,
2020, for additional inclusion criteria). To balance
taVNS groups on nonlinguistic pitch contour identifica-
tion ability and musicianship, participants were assigned
to a taVNS group (priming/peristim vs. sham) based on
their overall accuracy on a shortened version (126 trials)
Phillips et al.
129
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. taVNS Group Descriptive Statistics
taVNS Group
N (female)
Age (SD)
PCID Score (SD)
NM
Priming
Peristim
Sham
12 (8)
13 (9)
20 (12)
22.7 (4.19)
21.7 (2.87)
22.1 (4.01)
0.67 (0.12)
0.68 (0.10)
0.65 (0.11)
2
4
4
OMSI Response Count
ML
5
5
10
AM
3
3
3
SA
2
1
3
NM = nonmusician; ML = music-loving nonmusician; AM = amateur musician; SA = serious amateur musician.
of a pitch contour identification task (PCID; Bowles et al.,
2016; Bent, Bradlow, & Wright, 2006) and their response
to one item from the Ollen Musical Sophistication Index
(OMSI; Zhang & Schubert, 2019; Ollen, 2006): What title
best describes you? 1 = Nonmusician, 2 = music-loving
nonmusician, 3 = amateur musician, 4 = serious am-
ateur musician, 5 = semiprofessional musician, 6 =
professional musician.
To link behavioral and electrophysiological indices of
learning, the present analysis includes only participants
with complete data sets for each measure and experi-
mental task (see Analysis Approach section). Forty-five
participants (n = 12 priming, n = 13 peristim, n = 20
sham) are included in the present analysis. These partic-
ipants (29 female) were 18–34 years old (M = 22.13,
SD = 3.70), and taVNS groups did not differ significantly
on mean PCID score (ANOVA: F(2, 42) = 0.34, p = .71),
PCID score variance (Levene’s test: F(2, 42) = 0.54, p = .59),
or OMSI score (Kruskal–Wallis rank sum test: χ2(2) = 0.77,
p = .68; see Table 1). Whereas all taVNS groups included
fewer participants than in Pandža et al. (2020; n = 17
priming, n = 17 peristim, n = 35 sham), the smaller sam-
ple sizes for the priming and peristim taVNS conditions in
the present analysis are similar to that of Llanos et al.
(2020; 12 participants per group), which found peristim
taVNS effects on Mandarin tone categorization. Thus, the
final sample size in the present analysis was determined
a priori to be powerful enough to detect taVNS-related
differences of lexical learning.
Materials
The target items in this study were nine phonologically
plausible monosyllabic Mandarin pseudowords, consist-
ing of the syllables /ba/, /bi/, and /pi/, each produced with
Mandarin Tones 1 (flat), 2 (rising), and 4 (falling) by two
native speakers of Mandarin (one female, one male).
Tone 3 was not included in this study because of a creaky
quality often observed in Tone 3, which could lead to
easier identification from the other tone stimuli and to
avoid the added difficulty of learning to discriminate
between Tones 2 and 3, which are acoustically similar
and more easily confused by L2 and L1 speakers of
Mandarin (Hao, 2012; Shen & Lin, 1991). These items were
recorded along with stimuli created for a previous study
of Mandarin tone word learning (Bowles et al., 2016)
using the same recording procedure. Each auditory
recording was root-mean-squared normalized and pre-
sented at 70 dBA sound pressure level (SPL). The same
recordings of the Mandarin pseudowords were used
across tasks without any manipulations to stimulus
duration or other acoustic features. Each pseudoword
was paired with an English noun (COIN, GOWN, LENS,
MENU, OVEN, RAFT, SOFA, TRAY, VASE ), which appeared
only in written form across tasks. English pairings were
selected to control frequency (logSUBTLEX: 2.29–2.71;
Brysbaert & New, 2009), concreteness (4.61–5.00;
Brysbaert, Warriner, & Kuperman, 2014), word length
(four letters, one to two syllables), and animacy. Three
lists of Mandarin–English word pairings were created
such that across lists, each English word occurred once
with each tone (1, 2, 4) and once with each syllable (/ba/,
/bi/, /pi/). Each participant encountered only one list
across all tasks and sessions.
Procedures
Participants completed all tasks in a sound-attenuated,
dimly lit room, seated 65 cm from a 24-in. LCD computer
monitor. Tasks were delivered by E-Prime 2.0 software
(Psychology Software Tools, Inc., 2012) with auditory
Mandarin pseudowords presented via Neuvana
(Neuvana [previously Nervana], LLC) first generation
earbuds connected by a 3.5-mm audio plug to a Chronos
device, which was used to collect behavioral responses,
and by a 2.5-mm electrode plug to a Digitimer DS8R
Biphasic Constant Current Stimulator (Digitimer North
America, LLC), which generated the taVNS. The left
earbud had an embedded electrode with two contact
areas that made contact with the superior and inferior
walls of the outer ear canal when inserted into the ear.
The silicone tip fitting over this earbud was modified by
replacing the silicone located over the electrode contact
areas with 0.5–1 cm2 pieces of Axelgaard AG735 and/or
AG2550 hydrogel (Axelgaard Manufacturing Co., Ltd) as
a transmission medium to maintain consistent contact
between the electrode and outer ear canal wall. Once
the left earbud was inserted, the taVNS conducting
electrode made contact with the most lateral 1 cm of the
superior and inferior walls of the outer ear canal via the
hydrogel. Each participant was fit for earbud tip size, and
a new pair of earbuds was prepared each session. EEG was
acquired with an Electrical Geodesics Inc. 64-channel
130
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
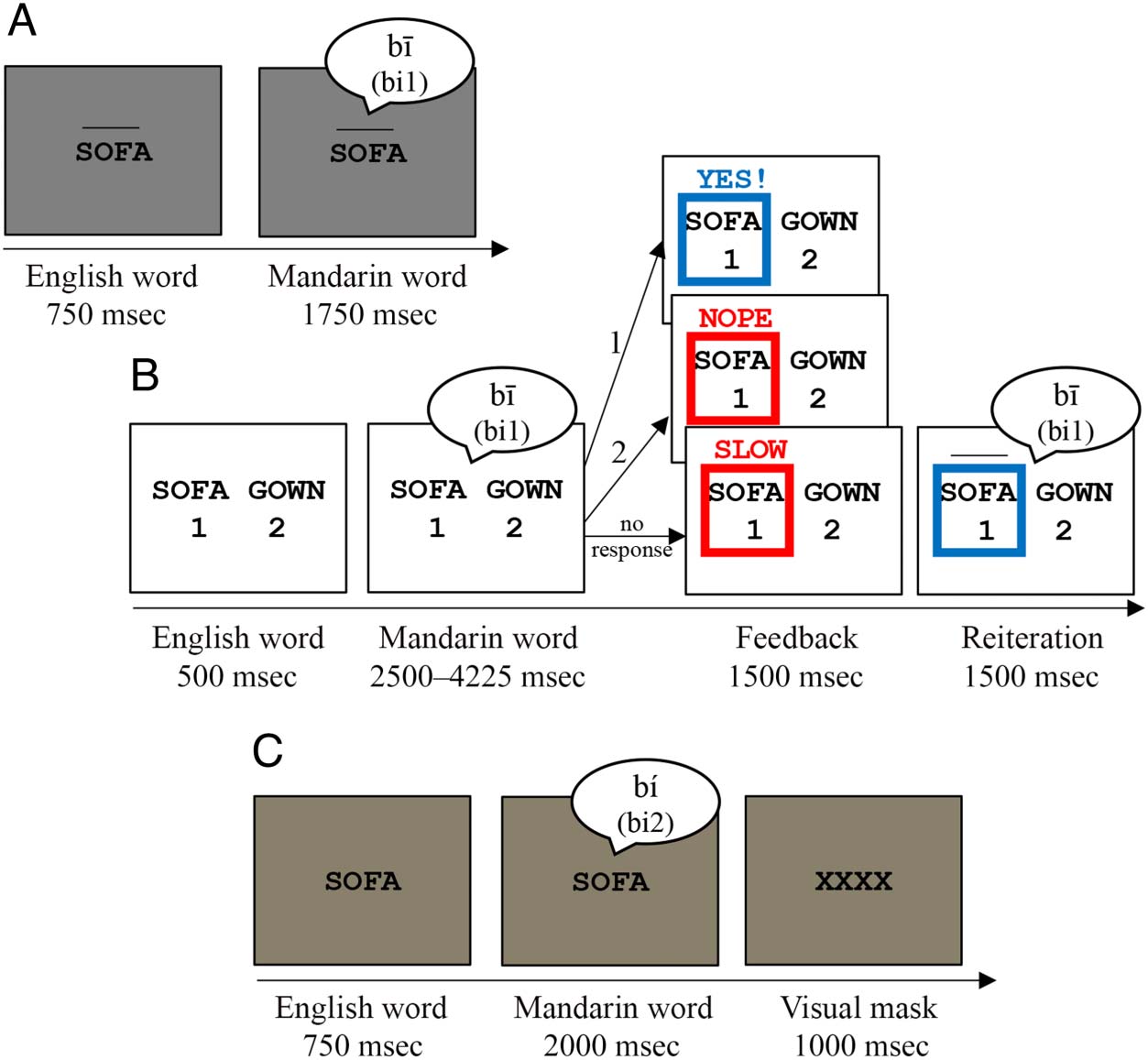
Figure 1. Sequence and
durations of events in each trial
for (A) the passive word
learning task, (B) active word
learning task, and (C) lexical
recognition test. For each task,
“Mandarin word” indicates the
time period when the Mandarin
pseudoword is auditorily
presented. For active word
learning (B) and lexical
recognition (C), participant
responses were recorded
during the “Mandarin word”
period of each trial.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
HydroCel Geodesic Sensor Net and an NA300 high-
impedance amplifier using Net Station software, Version
5.4.1.1 (r26882, Electrical Geodesics Inc.). Data were
sampled at 1000 Hz, referenced online to the vertex
(Cz), using a 400-Hz analog low-pass filter. Impedances
were kept below 50 KΩ where possible and otherwise
under 100 KΩ, as is customary with this system. EEG was
preprocessed off-line in MATLAB 2017b using EEGLab
Version 14.1.2 (Delorme & Makeig, 2004) and ERPLab
Toolbox Version 7.0.0 (Lopez-Calderon & Luck, 2014).
During a pretraining session, at least 1 day before train-
ing began, participants completed several tasks to estab-
lish study eligibility and collect individual difference
measures (including PCID and OMSI) and were assigned
a taVNS group number by a member of the research team
not involved in data collection or analysis. Training tasks
were programmed to deliver the appropriate taVNS
(priming/peristim/sham) based on participant number,
and this design ensured that both proctors and partici-
pants were blind to the taVNS condition (Pandža et al.,
2020). The two training sessions were completed on con-
secutive days or with 1 day in between. In each training
session, participants first completed a task to familiarize
them with the Mandarin tones appearing in the stimuli.
In this task, participants read descriptions of each tone
contour on the monitor and were shown corresponding
visual representations (flat, rising, or falling lines) and
listened to recorded examples of each tone produced
with the vowel
/a/ by a male native speaker of
Mandarin. No data were collected during this task. After
familiarization, participants completed phonological
categorization and discrimination pretests (Day 1 only),
a phonological training task, the passive and active word
learning tasks followed by the lexical recognition test
described below, and phonological categorization and
discrimination posttests (Day 2 only). The data presented
here are from a larger taVNS study, but only the tasks
relevant to word learning are detailed here because of
the scope of the present RQs (see Pandža et al., 2020,
for the full task sequence and details of the PCID,
OMSI, and tone familiarization tasks). Each session lasted
approximately 3 hr. Illustrations of each task are shown in
Figure 1.
Passive Word Learning
The passive word learning task comprised 90 trials and
lasted 7–8 min including instructions. Each trial began
with a 750-msec period in which one of the English
words appeared in the center of the monitor (40 pt. black
Courier New, gray background) with an image of its tonal
contour above the word, followed by a 1750-msec period
that began with the auditory presentation of the corre-
sponding Mandarin pseudoword while the English trans-
lation equivalent remained on screen. Visual depictions
of the tone contours were included based on results of
pilot testing, which indicated the task may have been
too difficult for participants to learn the tonal contrasts
when relying solely on auditory input. Each Mandarin
pseudoword was presented 10 times (5 times per
speaker), and trial order was pseudorandomized so that
no more than four consecutive trials had the same
Phillips et al.
131
syllable, tone, or speaker, and no consecutive trials had
the same Mandarin pseudoword. Participants were
instructed to try to memorize the meaning of each
Mandarin pseudoword and to focus on the center of
the screen and limit movements during the task. There
was no practice for this task. EEG was recorded continu-
ously during this task, and participants did not make
behavioral responses.
Active Word Learning
The active word learning task comprised 36 trials, lasting
3–4 min with instructions. Each trial began with a
500-msec period in which two of the English words
appeared side by side in the center of the monitor above
the numbers 1 and 2 (35 pt. black Courier New, white
background); followed by a 2500- to 4225-msec period
that began with the auditory presentation of one
Mandarin pseudoword while the English words remained
on screen and ended once participants pressed Button 1
or 2 to indicate the correct translation; then a 1500-msec
period in which a box appeared around the correct
response and a word (“YES!”, “NOPE,” or “SLOW”)
appeared above the English words to indicate perfor-
mance (blue if correct, red if incorrect); and a 1500-msec
period in which the Mandarin pseudoword was pre-
sented again with an image of its tone contour above
the correct English word. Each button was correct in
half of the trials. In each trial, the Mandarin translation
equivalents for the distractor and correct item differed
in one of three ways: same tone, different syllable; differ-
ent tone, same syllable; and different tone, different sylla-
ble. Each Mandarin pseudoword occurred with each
distractor type at least once, and there were three to five
occurrences of each tonal confusion pair (e.g., correct
word is Tone 1, distractor is Tone 2) and syllable confu-
sion pair (e.g., correct word is /ba/, distractor is /bi/). Each
Mandarin pseudoword was presented 4 times (2 times
per speaker), and trial order was pseudorandomized so
no more than four consecutive trials contained the same
syllable, tone, speaker, or distractor type, and no consec-
utive trials contained the same English words. Participants
were instructed to select the correct English equivalent
for each Mandarin pseudoword, and there was no prac-
tice for this task. Accuracy and RT were recorded for
each trial.
Lexical Recognition Test
The lexical recognition test included 216 trials, lasting
about 20 min with instructions. Each trial began with a
750-msec baseline period in which one of the English
words appeared in the center of the monitor (30 pt. black
Courier New, tan background); followed by a 2,000-msec
period in which one Mandarin pseudoword was auditorily
presented while the English word remained on screen
and participants pressed a button to indicate whether
stimuli were translation equivalents (match, Button 1)
or not (mismatch, Button 2); and a 1000-msec period
in which a visual mask (“XXXX”) replaced the English
word on screen. Each Mandarin pseudoword was pre-
sented 24 times, split over two testing blocks, and
occurred in an equal number of match and mismatch tri-
als. Trial order was pseudorandomized so that no more
than four consecutive trials contained the same syllable,
tone, speaker, or trial condition, and no consecutive trials
contained the same Mandarin pseudoword or English
word. Because of these requirements, the number of
times each auditory pseudoword was produced by each
speaker could not be perfectly balanced. Whereas some
pseudowords were spoken 12 times by each speaker,
others were spoken 16 times by one speaker and 8 times
by the other speaker. This varied across presentation
lists. The order of trial conditions was consistent within
each block, but the specific stimulus pair appearing in
each trial was randomized for each session. Participants
were instructed to indicate as quickly and accurately as
possible whether the English word and the Mandarin
pseudoword in each trial were translation equivalents
and to limit blinking to the mask portion of the trial.
There was no practice or feedback. Accuracy, RT, and
continuous EEG were recorded.
taVNS Parameters
The taVNS consisted of a biphasic square wave (50-μs
pulse width, 350-μs interphase dwell, 100% recovery
phase ratio) triggered at 300 Hz by a TTL pulse over a
BNC jack from a custom-programmed Arduino UNO
board controlled by E-Prime. All participants completed
a calibration and ramping procedure before each task
where taVNS was possible to determine their perceptual
threshold (see Pandža et al., 2020, for details). In the sub-
sequent tasks, priming and peristim participants received
taVNS at 0.2 mA below their perceptual threshold to
prevent them from feeling any sensation because of
taVNS that might unblind them to their taVNS condition
and affect their task performance. For the priming
experiment, taVNS was delivered to participants in the
priming group 3 times each session while they watched
a 10-min silent animated video used for resting-state fMRI
scans (Inscapes; Vanderwal, Kelly, Eilbott, Mayes, &
Castellanos, 2015). This task was administered about
9 min before passive word learning, 18 min before active
word learning, and immediately before the lexical recog-
nition test, but only participants in the priming group
received continuous taVNS during the video. In the peri-
stim experiment, the video task was not administered.
Instead, participants in the peristim group received
500-msec bursts of taVNS immediately preceding each
trial in both word learning tasks and the lexical recogni-
tion test. Participants in the sham taVNS group for each
experiment completed the same tasks but did not
receive taVNS outside of calibration and ramping.
132
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
During piloting, it was discovered that taVNS at higher
intensity levels occasionally produced an audible noise
artifact. To mask this noise, a 60 dB SPL pink noise mask
was played whenever taVNS was possible in the priming
experiment, and the pink noise mask overlaid with a
recording of the taVNS sound artifact was played in the
peristim experiment. More specifically, the priming
group and the sham group matched to the priming
group heard the same pink noise mask during the prim-
ing video, whereas the peristim group and the sham
group matched to the peristim group heard the same
pink noise mask overlaid with the taVNS sound artifact
for 500 msec prior to the onset of the Mandarin pseudo-
words in the word learning tasks and lexical recognition
test. All participants also heard their respective sound
masks during each taVNS calibration and ramping.
Analysis Approach
Preliminary analyses of active word learning and lexical
recognition accuracy and RT, following the procedure
outlined here, did not reveal major behavioral differences
between sham groups—the only significant difference
was a slowdown in RT to mismatch trials on Day 2 for
the peristim sham group but not the priming sham
group. Thus, data from the priming and peristim experi-
ments were combined into a single, three-taVNS-group
analysis, with one sham group containing all participants
who received sham taVNS across the priming and peri-
stim experiments. Accuracy was analyzed using logistic
mixed-effects models, and log RT and ERP mean ampli-
tude were analyzed using linear mixed-effects models,
in R Version 3.6.3 (R Core Team, 2020). The buildmer
package (v. 1.5; Voeten, 2020) was used to automatically
determine the best-fitting model for each measure by
first determining the maximal random- and fixed-effects
structures that allowed the model to converge, ordering
effects by the magnitude of their contribution to model
fit, and then removing terms in a backward stepwise pro-
cedure until the model contained only factors that signif-
icantly improved model fit. This method provides a more
objective and replicable way to fit exploratory mixed-
effects models. Improvements in fit between nested
models were assessed with likelihood ratio tests, and
p values reported for linear model fixed effects were cal-
culated using Satterthwaite’s approximation for degrees
of freedom. For significant fixed effects, model-predicted
values shown in the below tables were obtained using the
effects package (v. 4.1–4; Fox & Weisberg, 2018), which
weights factor levels by sample size, absorbs lower-order
terms for interactions, and averages over noninteracting
terms. In all models, factors were treatment coded and
PCID (centered, z-score transformed) and OMSI (cen-
tered on the mean) scores were included to model indi-
vidual differences known to impact tone word learning.
To interpret model fixed effects involving three-level fac-
tors, variables were releveled as necessary (and indicated
in the text) to obtain model estimates for all factor level
comparisons.
Prior to the analysis of accuracy and RT in the lexical
recognition test, the time until voicing in each sound file
was subtracted from the RT for corresponding trials be-
cause these portions of the Mandarin pseudowords did
not carry tone information, which was required to deter-
mine whether the items were a match or mismatch. This
was not done for the active word learning task because
the syllable-initial phoneme could distinguish the correct
item for some trials. For accuracy analyses in both tasks,
trials were excluded if the response was recorded within
60 msec of the adjusted onset of the Mandarin pseudo-
word (0.03% of trials for active word learning; 0.05% for
lexical recognition) and for RT analyses, trials that re-
ceived incorrect responses were also excluded (28.31%
of trials for active word learning; 27.34% for lexical
recognition).
Prior to analysis, EEG was resampled at 250 Hz, high-
pass filtered (0.1 Hz, Butterworth, second order) and
notch filtered at 60 Hz to remove line noise. Loose chan-
nels were interpolated from surrounding sites, epochs
were extracted from −200 to 1000 msec relative to the
onset of the Mandarin pseudoword in each trial, and oc-
ular artifacts were corrected based on an independent
components analysis (Luck, 2014). Individual epochs
were then low-pass filtered (30 Hz, Butterworth, second
order), channels were rereferenced to the average of all
sites, and epochs were baseline-corrected against the
prestimulus period. Epochs with deflections exceeding
±100 μV on any channel were excluded from analysis
(mean 9% for passive word learning; mean 6% for lexical
recognition). EEG data for participants who had greater
than 25% of epochs rejected due to artifacts in either
session were excluded from analyses. The 300–500 msec
following the onset of voicing in each trial was selected as
the analysis window based on the N400 literature. For the
passive word learning task, the dependent variable was
mean amplitude over the analysis window at each of nine
electrodes over lateral (left hemisphere: F3, C3, P3; right
hemisphere: F4, C4, P4) and midline sites (Fz, Cz, Pz)
covering frontal, central, and parietal regions (based on
the 10–10 system; Luu & Ferree, 2005) calculated by
participant and session. For the lexical recognition test,
the dependent variable was the difference in mean ampli-
tude between mismatch and match trials (mismatch
minus match) at each of the nine electrode sites calcu-
lated by participant and session.
The maximal fixed-effects structures for accuracy and
RT analyses included the three-way interaction between
taVNS group (GROUP: SHAM/PRIMING/PERISTIM), training ses-
sion (SESS: DAY 1/ DAY 2), and trial condition (COND:
TONE/SEGMENT/TONE + SEGMENT for active word learning;
COND: MATCH/ MISMATCH for lexical recognition) and all
lower-order terms, plus noninteracting terms for musi-
cianship (OMSI) and nonlinguistic pitch contour identifi-
cation task (PCID) ability. The maximal random-effects
Phillips et al.
133
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
structures included by-participant random intercepts and
slopes for the SESS × COND interaction and lower-order
terms plus by-item random intercepts and slopes for
the three-way GROUP × SESS × COND interaction and all
lower-order terms. The maximal fixed-effects structure
for ERP analyses included the four-way interaction
between GROUP, SESS, anterior–posterior electrode region
(REGION: FNT/CNT/ PAR [frontal/central/parietal]), and
electrode laterality (LAT: LEFT/ MIDLINE/ RIGHT) and all
lower-order terms, plus three-way and lower-order inter-
actions between the two electrode location factors and
OMSI and PCID; the maximal random-effects structure in-
cluded by-participant random intercepts and slopes for
the three-way SESS × REGION × LAT interaction and all
lower-order interactions. Treatment coding electrode
region and laterality factors provided direct comparisons
between all three factor levels (e.g., right vs. midline vs.
left) in the same model, and estimates for all comparisons
were obtained by releveling.
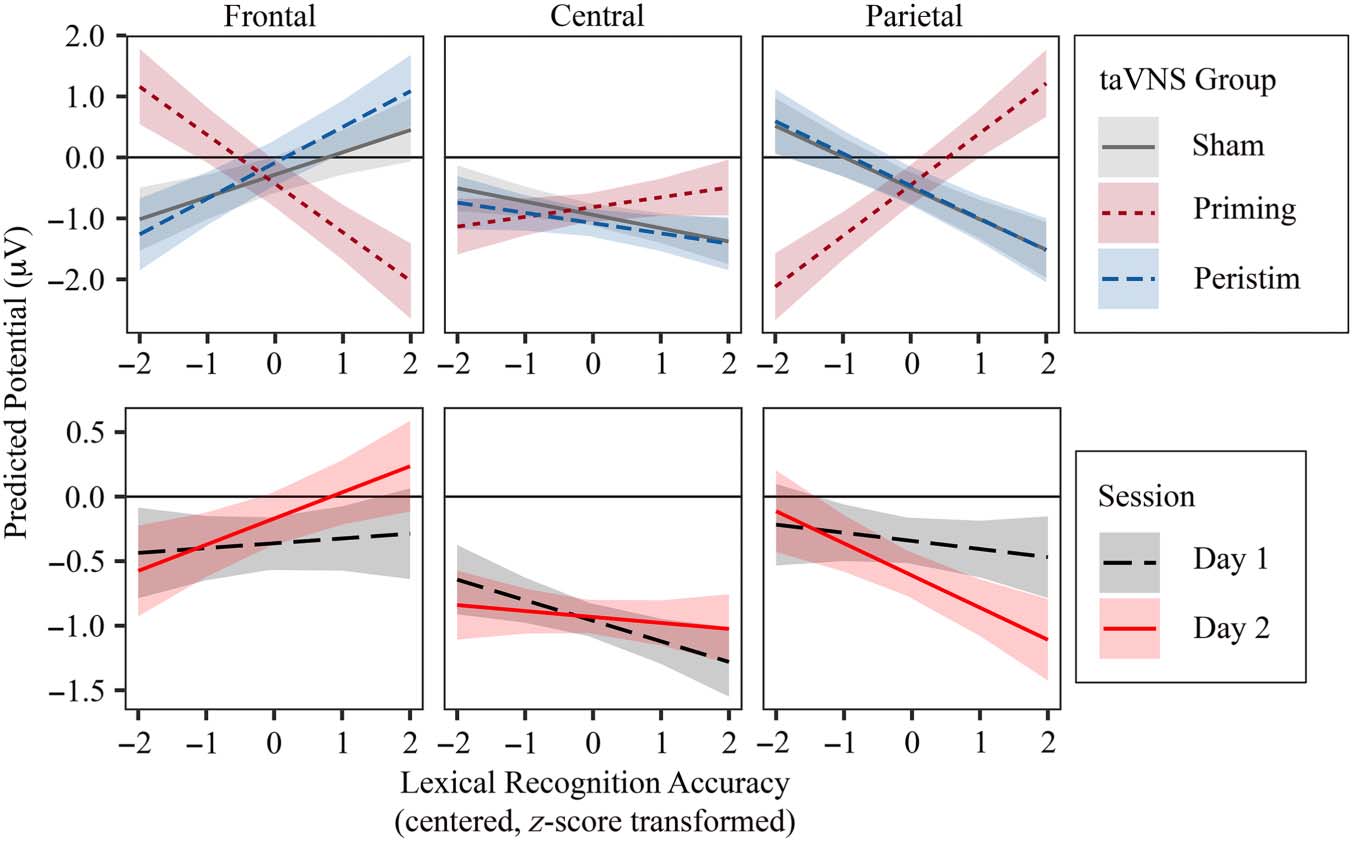
Following the main analyses, post hoc models were
run to more directly explore the relationship between be-
havioral outcomes in the lexical recognition test and dif-
ferences in semantic encoding reflected in passive word
learning N400 amplitude and topography. Lexical recog-
nition mean accuracy (ACC) and RT by participant and ses-
sion ( both centered, z score transformed across
sessions) were tested in two separate linear mixed-effects
models. In each model, the behavioral predictor was
added to the best-fitting passive word learning N400
model as a simple fixed effect and as interactions with
all other fixed effects. To determine whether the inclu-
sion of the behavioral predictor improved N400 model
fit, the maximal model underwent the same model fitting
procedure described above, with only model terms in-
volving behavioral predictors subject to removal.
RESULTS
Passive Word Learning Task
N400
By-participant N400 mean amplitude (300–500 msec) was
analyzed for the passive word learning task to determine
whether there were differences in N400 magnitude or to-
pography (by region and laterality) between taVNS
groups and sessions that would indicate differences in
the amount of exposure needed to encode novel tone
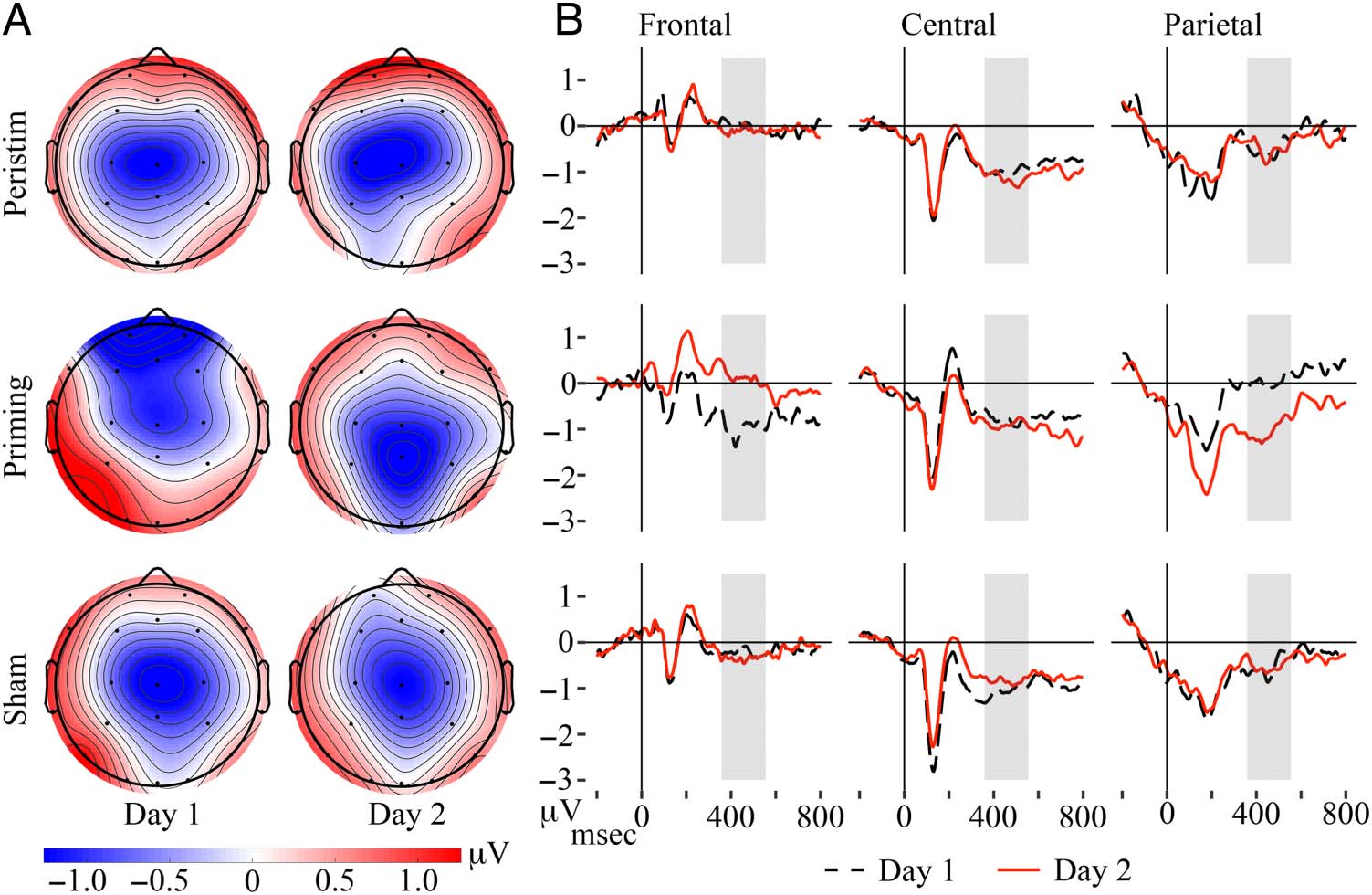
words in semantic memory. Topographic plots of mean
amplitude in Figure 2 reveal a prominent negativity cen-
tered over central or centro-parietal midline sites across
taVNS groups and sessions, with the exception of the
priming group on Day 1 where the negativity spans cen-
tral, frontal, and frontopolar sites.
The best-fitting model for these data (Table 2) retained
a significant REGION × SESS × GROUP interaction, indicating
differences in N400 anterior–posterior topography shifts
from Day 1 to 2 between taVNS groups. For the priming
group on Day 1, frontal sites were negative (b = −1.06,
SE = 0.36, p = .004) and not different from central (b =
−0.16, SE = 0.38, p = .67) and parietal sites (b = 0.63,
SE = 0.57, p = .27), although parietal sites were less neg-
ative than central sites (with PRIMING, CNT, DAY 1 reference
levels: REGION(PAR): b = 0.79, SE = 0.38, p = .039). For
priming on Day 2, frontal sites did not differ from zero
and central and parietal sites were more negative (with
PRIMING, FNT, DAY 2 reference levels: INTERCEPT: b = 0.03,
Figure 2. (A) Topographic
plots of by-participant N400
mean amplitude during the
analysis window in the passive
word learning task. (B)
By-participant ERPs averaged
over all sites in each region
comparing taVNS group and
session for the passive word
learning task. The onset of the
Mandarin pseudoword is at
0 msec; gray boxes indicate
the mean onset and offset for
the adjusted analysis window
across trials.
134
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 2. Best-Fitting Model for Passive Word Learning N400 Mean Amplitude
Fixed Effect
(INTERCEPT)
REGION(CNT)
REGION(PAR)
LAT(LEFT)
LAT(RIGHT)
SESS(DAY 2)
GROUP(SHAM)
GROUP(PERISTIM)
REGION(CNT) × LAT(LEFT)
REGION(PAR) × LAT(LEFT)
REGION(CNT) × LAT(RIGHT)
REGION(PAR) × LAT(RIGHT)
REGION(CNT) × SESS(DAY 2)
REGION(PAR) × SESS(DAY 2)
LAT(LEFT) × GROUP(SHAM)
LAT(RIGHT) × GROUP(SHAM)
LAT(LEFT) × GROUP(PERISTIM)
LAT(RIGHT) × GROUP(PERISTIM)
REGION(CNT) × GROUP(SHAM)
REGION(PAR) × GROUP(SHAM)
REGION(CNT) × GROUP(PERISTIM)
REGION(PAR) × GROUP(PERISTIM)
SESS(DAY 2) × GROUP(SHAM)
SESS(DAY 2) × GROUP(PERISTIM)
REGION(CNT) × SESS(DAY 2) × GROUP(SHAM)
REGION(PAR) × SESS(DAY 2) × GROUP(SHAM)
REGION(CNT) × SESS(DAY 2) × GROUP(PERISTIM)
REGION(PAR) × SESS(DAY 2) × GROUP(PERISTIM)
Random Effect
PARTICIPANT(INTERCEPT)
REGION(CNT)
REGION(PAR)
RESIDUAL
Est.
−1.06
−0.16
0.63
0.16
0.09
1.10
0.82
1.21
0.61
0.81
0.53
0.45
−1.20
−2.14
−0.10
−0.08
−0.58
−0.12
−1.04
−1.32
−1.23
−1.54
−1.22
−1.26
1.52
2.25
1.24
2.35
Variance
1.17
1.11
3.32
0.70
SE
0.36
0.38
0.57
0.17
0.17
0.20
0.44
0.49
0.18
0.18
0.18
0.18
0.28
0.28
0.18
0.18
0.19
0.19
0.46
0.71
0.50
0.78
0.25
0.27
0.35
0.35
0.39
0.39
SD
1.08
1.05
1.82
0.84
df
58.79
67.19
50.88
656.00
656.00
656.00
55.65
55.65
656.00
656.00
656.00
656.00
656.00
656.00
656.00
656.00
656.00
656.00
57.83
47.70
57.83
47.70
656.00
656.00
656.00
656.00
656.00
656.00
t value
p value
−2.99
−0.42
1.11
0.91
0.51
5.55
1.85
2.49
3.42
4.58
2.98
2.52
−4.28
−7.66
−0.56
−0.46
−3.00
−0.61
−2.27
−1.86
−2.45
−1.97
−4.87
−4.60
4.29
6.37
3.21
6.07
.004
**
.67
.27
.36
.61
< .001
***
.070
.016
< .001
< .001
.003
.012
< .001
< .001
.58
.65
.003
.54
.027
.069
.017
.055
< .001
< .001
< .001
< .001
.001
< .001
*
***
***
**
*
***
***
**
*
*
***
***
***
***
**
***
Correlation
−.76
−.93
.87
Reference levels: FNT, MID, PRIMING, DAY 1. Number of obs.: 810, participants: 45.
* p < .05.
** p < .01.
*** p < .001.
Phillips et al.
135
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 3. Model-Predicted Values for Passive Word Learning
N400 Amplitude (μV) by taVNS Group, Region, and Session
Table 4. Model-Predicted Values for Passive Word Learning
N400 Amplitude (μV) by Region and Laterality
taVNS Group
Peristim
Priming
Sham
Frontal
Central
Parietal
Frontal
Central
Parietal
Frontal
Central
Parietal
Day 1
0.00
−1.02
−0.49
−0.98
−0.76
0.07
−0.22
−1.04
−0.49
Day 2
−0.17
−1.14
−0.44
0.11
−0.86
−0.97
−0.34
−0.84
−0.50
SE = 0.36, p = .93; REGION(CNT): b = −1.35, SE = 0.38,
p < .001; REGION(PAR): b = −1.51, SE = 0.57, p = .011) with
no difference between central and parietal (with PRIMING,
CNT, DAY 2 reference levels: REGION(PAR): b = −0.15,
SE = 0.38, p = .69). Going from Day 1 to Day 2 for the
priming group, the frontal negativity disappeared (b =
1.10, SE = 0.20, p < .001), the negativity at central sites
did not change (with PRIMING, CNT, DAY 1 reference levels:
SESS(DAY 2): b = −0.10, SE = 0.20, p = .62), and parietal
sites became more negative (with PRIMING, PAR, DAY 1 refer-
ence levels: SESS(DAY 2): b = −1.04, SE = 0.20, p < .001).
For the peristim and sham groups, N400 amplitude
was largest over central and parietal sites, with frontal
sites less negative and no difference between central
and parietal sites on both days (with PERISTIM, CNT, DAY 1
reference levels: INTERCEPT: b = −1.25, SE = 0.26, p <
.001; REGION(FNT): b = 1.39, SE = 0.36, p < .001; REGION
(PAR): b = 0.49, SE = 0.36, p = .18; with PERISTIM, CNT,
DAY 2 reference levels: INTERCEPT: b = −1.36, SE = 0.26,
p < .001; REGION(FNT): b = 1.35, SE = 0.36, p < .001;
REGION(PAR): b = 0.65, SE = 0.36, p = .076; with SHAM,
CNT, DAY 1 reference levels: INTERCEPT: b = −1.44, SE =
0.21, p < .001; REGION(FNT): b = 1.20, SE = 0.30, p <
.001; REGION(PAR): b = 0.51, SE = 0.30, p = .094; with
SHAM, CNT, DAY 2 reference levels: INTERCEPT: b = −1.24,
SE = 0.21, p < .001; REGION(FNT): b = 0.88, SE = 0.30,
p = .004; REGION(PAR): b = 0.30, SE = 0.30, p = .33).
Going from Day 1 to Day 2, there were no changes at
frontal, central, or parietal regions for peristim and sham
groups (with PERISTIM, FNT, DAY 1 reference levels: SESS
(DAY 2): b = −0.16, SE = 0.20, p = .39; with PERISTIM,
CNT, DAY 1 reference levels: SESS(DAY 2): b = −0.12, SE =
0.19, p = .54; with PERISTIM, PAR, DAY 1 reference levels:
SESS(DAY 2): b = 0.05, SE = 0.19, p = 0.80; with SHAM,
FNT, DAY 1 reference levels: SESS(DAY 2): b = −0.12,
Frontal
Central
Parietal
Left
−0.31
−0.76
−0.13
Midline
−0.25
−1.31
−0.88
Right
−0.24
−0.77
−0.42
SE = 0.15, p = .43; with SHAM, CNT, DAY 1 reference
levels: SESS(DAY 2): b = 0.2, SE = 0.15, p = .19; with
SHAM, PAR, DAY 1 reference levels: SESS(DAY 2): b = −0.01,
SE = 0.15, p = .95). Together, these effects reflect stable
centro-parietal N400 topography for sham and peristim
groups across days, whereas the priming group’s N400
shifted from fronto-central on Day 1 to centro-parietal on
Day 2. Model-predicted values are shown in Table 3.
Considering the remaining significant fixed effects, the
REGION × LAT interactions reflect a reduced amplitude
negativity for lateral versus midline central and parietal
sites, which together indicate a midline focus of the
centro-parietal N400 across taVNS groups and sessions
(REGION(CNT) × LAT(LEFT): b = 0.61, SE = 0.18, p <
.001; REGION(PAR) × LAT(LEFT): b = 0.81, SE = 0.18, p <
.001; REGION(CNT) × LAT(RIGHT): b = 0.53, SE = 0.18, p =
.003; REGION(PAR) × LAT(RIGHT): b = 0.45, SE = 0.18, p =
.012). Corresponding model predictions are shown in
Table 4. The nonsignificant GROUP × LAT interactions in-
dicate no difference in N400 laterality between priming
and sham groups (LAT(LEFT) × GROUP(SHAM): b = −0.10,
SE = 0.18, p = .58; LAT(RIGHT) × GROUP(SHAM): b = −0.08,
SE = 0.18, p = .65). However, the significant LAT(LEFT) ×
GROUP(PERISTIM) interaction reveals that the relative
difference in negativity between midline and left sites
observed for the priming and sham groups was
attenuated for the peristim group (b = −0.58, SE =
0.19, p = .003). This reflects a more left-lateralized
N400 distribution for the peristim group compared to
priming and sham (with SHAM, MIDLINE reference levels:
LAT(LEFT) × GROUP(PERISTIM): b = −0.48, SE = 0.17, p =
.005; LAT(RIGHT) × GROUP(PERISTIM): b = −0.04, SE =
0.17, p = .83). Corresponding model-predicted values
are shown in Table 5.
Table 5. Model-Predicted Values for Passive Word Learning
N400 Amplitude (μV) by taVNS Group and Laterality
Peristim
Priming
Sham
Left
−0.61
−0.28
−0.33
Midline
−0.66
−0.91
−0.86
Right
−0.36
−0.50
−0.53
136
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 6. Best-Fitting Model for Active Word Learning Accuracy
Fixed Effect
(INTERCEPT)
OMSI
PCID
SESS(DAY 2)
COND(TONE)
COND(SYLLABLE)
Est.
SE
z Value p Value
1.20
0.32
0.26
0.52
0.15
0.12
0.11
0.08
7.96 < .001
***
2.76
2.41
.006
**
.016
*
6.22 < .001
***
−0.65
−0.48
0.11 −5.89 < .001
0.13 −3.74 < .001
***
***
Random Effect
Variance SD
PARTICIPANT(INTERCEPT)
ITEM(INTERCEPT)
0.34
0.12
0.58
0.34
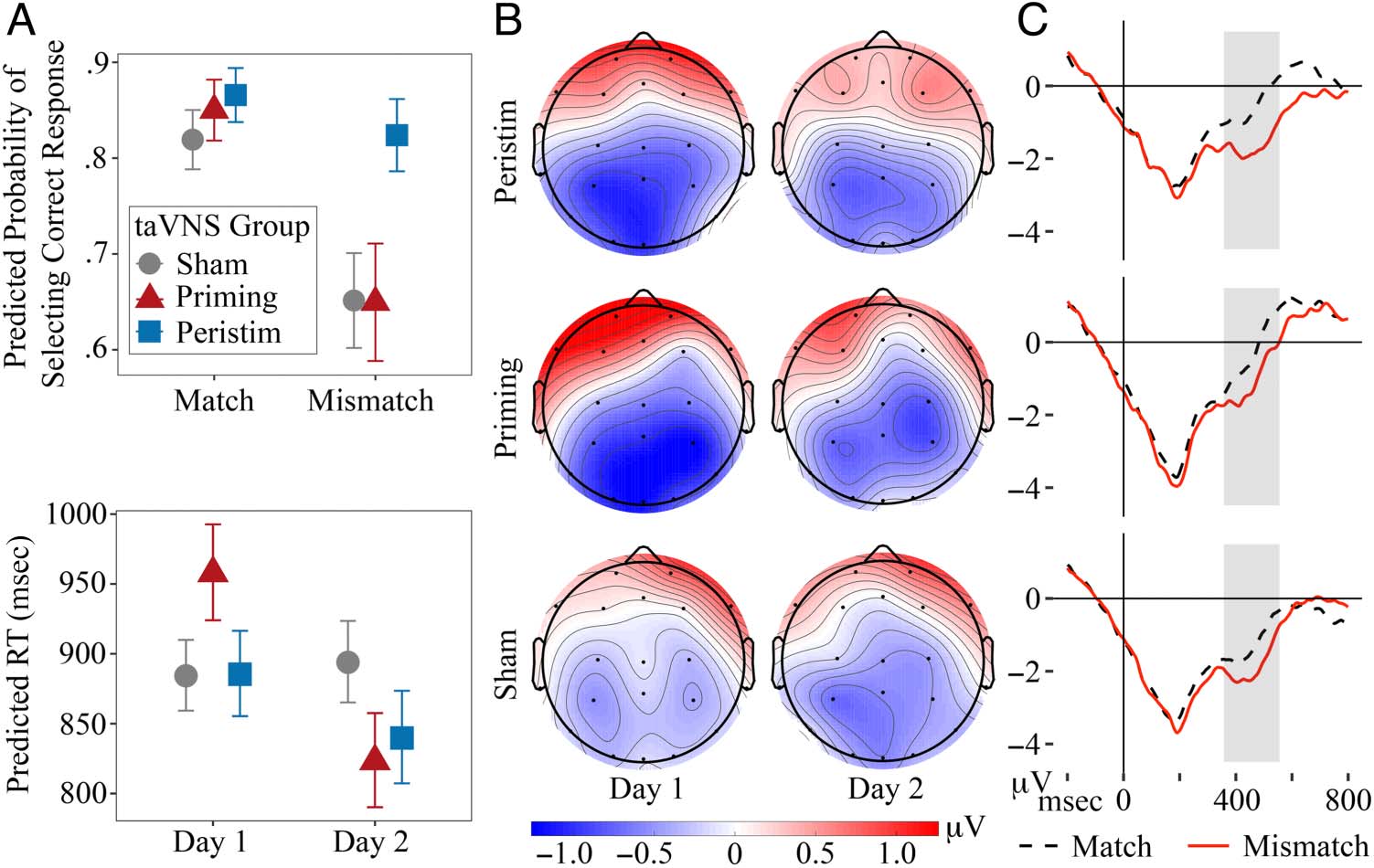
peristim (Day 1: M (SD) = 0.65 (0.17); Day 2: M (SD) =
0.76 (0.15)). The best-fitting model for these data
(Table 6) reveals that the likelihood of selecting the
correct response was higher on Day 2 than Day 1 across
taVNS groups (predicted = .79 vs. .69: b = 0.52, SE =
0.08, p < .001) and for individuals with higher PCID
and OMSI scores (PCID: b = 0.26, SE = 0.11, p = .016;
OMSI: b = 0.32, SE = 0.12, p = .006). Compared to the
likelihood of accuracy for trials where response items’
translation equivalents differed in both syllable and tone
(pred. = .81), accurate responses were less likely for tri-
als where response items differed only in tone (pred. =
.69; b = −0.65, SE = 0.11, p < .001) or syllable (pred. =
.73; b = −0.48, SE = 0.13, p < .001) with no difference
between these conditions (with TONE reference level:
COND(SYLLABLE): b = 0.17, SE = 0.13, p = .18).
Reference levels: DAY 1, TONE + SYLLABLE. Number of obs.: 3240, partic-
ipants: 45, items: 18.
RT
* p < .05.
** p < .01.
*** p < .001.
Active Word Learning Task
Accuracy
Active word learning task accuracy and RT were analyzed
to determine whether taVNS influenced behavior during
word learning. Across groups, mean accuracy improved
between training sessions: by 8% for the sham taVNS
group (Day 1: M (SD) = 0.66 (0.15); Day 2: M (SD) =
0.74 (0.13)), 9% for priming (Day 1: M (SD) = 0.71
(0.12); Day 2: M (SD) = 0.80 (0.12)), and 11% for
Table 7. Best-Fitting Model for Active Word Learning Log RT
Fixed Effect
(INTERCEPT)
SESS(DAY 2)
COND(TONE + SYLLABLE)
COND(SYLLABLE)
Estimate
7.06
−0.10
−0.04
−0.05
Random Effect
Variance
PARTICIPANT(INTERCEPT)
SESS(DAY 2)
ITEM(INTERCEPT)
RESIDUAL
0.03
0.03
0.00
0.07
SE
0.03
0.03
0.01
0.02
SD
0.18
0.17
0.04
0.26
Across groups, mean RT decreased from Day 1 to Day 2:
by 67 msec for the sham taVNS group (Day 1: M (SD) =
1185 (352); Day 2: M (SD) = 1118 (342)), 153 msec for
priming (Day 1: M (SD) = 1178 (387); Day 2: M (SD) =
1025 (324)), and 159 msec for peristim (Day 1: M (SD) =
1182 (339); Day 2: M (SD) = 1023 (335)). The best-fitting
model for RT (Table 7) reveals that, across taVNS groups,
accurate responses were given faster on Day 2 than Day 1
(pred. = 1126 msec vs. 1017 msec: b = −0.10, SE = 0.03,
p < .001) and, compared to trials were response item
translation equivalents differed only in tone (pred. =
1099 msec), accurate responses were given faster for tri-
als with response items that differed in syllable (pred. =
1042 msec: b = −0.05, SE = 0.02, p < .001) or syllable
t Value
p Value
232.43
−3.67
−2.58
−3.35
< .001
< .001
.010
< .001
***
***
**
***
df
61.17
43.92
1782.35
613.64
Correlation
−.43
Reference levels: DAY 1, TONE. Number of obs.: 2315, participants: 45, items: 18.
* p < .05.
** p < .01.
*** p < .001.
Phillips et al.
137
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 8. Best-Fitting Model for Lexical Recognition Accuracy
Fixed Effect
(INTERCEPT)
OMSI
SESS(DAY 2)
COND(MATCH)
GROUP(PRIMING)
GROUP(SHAM)
GROUP(PRIMING) × COND(MATCH)
GROUP(SHAM) × COND(MATCH)
SESS(DAY 2) × COND(MATCH)
Est.
1.03
0.40
1.03
0.34
−0.93
−0.92
0.80
0.57
−0.04
Random Effect
Variance
PARTICIPANT(INTERCEPT)
COND(MATCH)
SESS(DAY 2)
COND(MATCH) × SESS(DAY 2)
ITEM(INTERCEPT)
COND(MATCH)
SESS(DAY 2)
COND(MATCH) × SESS(DAY 2)
0.86
0.45
0.37
0.34
0.08
0.17
0.10
0.17
SE
0.25
0.11
0.13
0.22
0.34
0.29
0.28
0.24
0.16
SD
0.93
0.67
0.61
0.59
0.29
0.41
0.31
0.41
z Value
4.13
3.61
7.76
1.55
−2.74
−3.21
2.87
2.40
−0.23
−.68
.24
.38
−.28
−.23
.49
p Value
< .001
< .001
< .001
.12
.006
.001
.004
.017
.82
Correlation
−.02
−.37
.21
−.18
***
***
***
*
**
**
*
−.64
−.66
Reference levels: DAY 1, PERISTIM, MISMATCH. Number of obs.: 19429, participants: 45, items: 18.
* p < .05.
** p < .01.
*** p < .001.
and tone (pred. = 1060 msec: b = −0.04, SE = 0.01, p =
.010) with no difference between these conditions (with
TONE + SYLLABLE reference level: COND(SYLLABLE): b =
−0.02, SE = 0.02, p = .28).
Lexical Recognition Test
Accuracy
Accuracy and RT during the lexical recognition test were
analyzed to determine whether taVNS affected the level
of attainment for the Mandarin pseudowords at the end
of each session. Mean accuracy improved on Day 2 for all
groups: by 16% for the sham taVNS group (Day 1: M (SD) =
0.61 (0.14); Day 2: M (SD) = 0.77 (0.14)), 16% for priming
(Day 1: M (SD) = 0.67 (0.16); Day 2: M (SD) = 0.83 (0.15)),
and 10% for peristim (Day 1: M (SD) = 0.69 (0.16); Day 2:
M (SD) = 0.79 (0.14)). The best-fitting model for these
data (Table 8) reveals that the likelihood of being accurate
was higher on Day 2 than Day 1 for all groups (pred. = .86
vs. .69: b = 1.03, SE = 0.13, p < .001) and for participants
who reported higher musicianship (b = 0.40, SE = 0.11,
p < .001). The peristim group was equally likely to give
correct responses for match (pred. = .87) and mis-
match trials (pred. = .82; b = 0.34, SE = 0.22, p =
.12), but both priming and sham were less likely to
be accurate on mismatch trials (pred. priming = .65,
sham = .65) than match trials (pred. priming = .85,
sham = .82; with PRIMING, MATCH reference levels:
COND(MISMATCH): b = −1.14, SE = 0.23, p < .001; with
SHAM, MATCH reference levels: COND(MISMATCH): b =
−0.91, SE = 0.18, p < .001). The likelihood of accuracy
did not differ between taVNS groups for match trials
(with PERISTIM, MATCH reference levels: GROUP(PRIMING):
b = −0.13, SE = 0.29, p = 0.66; GROUP(SHAM): b =
−0.35, SE = 0.25, p = 0.15; with SHAM, MATCH reference
levels: GROUP(PRIMING): b = 0.27, SE = 0.28, p = .33).
However, the priming and sham groups were less likely
than the peristim group to give correct responses to
mismatch trials (GROUP(PRIMING): b = −0.93, SE = 0.34,
p = .006; GROUP(SHAM): b = −0.92, SE = 0.29, p =
.001), with no difference between these groups (with
138
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. (A) Model-predicted
values for accuracy likelihood
and RT in the lexical recognition
test, plotted with standard error
bars. (B) Topographic plots of
by-participant N400 effect
(mismatch–match) mean
amplitude during the adjusted
analysis window, comparing
taVNS group and session. (C)
By-participant ERPs averaged
over parietal sites (P3, Pz, P4)
and sessions comparing taVNS
group and trial condition for the
lexical recognition test. The
onset of the Mandarin
pseudoword is at 0 msec; gray
boxes indicate the mean onset
and offset for the adjusted
analysis window across trials.
SHAM, MISMATCH reference levels: GROUP(PRIMING): b =
−0.01, SE = 0.30, p = .98). Model-predicted values are
plotted in Figure 3.
(indicated by removal of the GROUP × COND term during
model selection because it did not improve model fit).
Model-predicted values are plotted in Figure 3.
RT
Across groups, mean RT decreased between Day 1 and
Day 2: by 6 msec for the sham taVNS group (Day 1: M
(SD) = 931 (313); Day 2: M (SD) = 925 (312)), 121 msec
for priming (Day 1: M (SD) = 990 (323); Day 2: M (SD) =
869 (293)), and 66 msec for peristim (Day 1: M (SD) =
951 (315); Day 2: M (SD) = 885 (288)). The best-fitting
model for these data (Table 9) reveals priming group RTs
sped up from Day 1 to Day 2 (b = −0.15, SE = 0.04, p <
.001) more so than RTs for peristim (b = 0.10, SE = 0.05,
p = .050) and sham (b = 0.16, SE = 0.05, p < .001). RTs
did not change between days for sham (pred. Day 1 =
884 msec, Day 2 = 894 msec; with SHAM, DAY 1 reference
levels: SESS(DAY 2): b = 0.02, SE = 0.03, p = .62) or peri-
stim (pred. Day 1 = 885 msec, Day 2 = 840 msec), and
RT changes did not differ between these groups (with
PERISTIM, DAY 1 reference levels: SESS(DAY 2): b = −0.05,
SE = 0.04, p = .17; GROUP(SHAM) × SESS(DAY 2): b = 0.06,
SE = 0.04, p = .15). There were no RT differences between
groups on Day 1 (GROUP(SHAM): b = −0.08, SE = 0.04, p =
.073; GROUP(PERISTIM): b = −0.08, SE = 0.05, p = .11; with
PERISTIM, DAY 1 reference levels: GROUP(SHAM): b = −0.001,
SE = 0.04, p = .98) or Day 2 (with PRIMING, DAY 2 reference
levels: GROUP(PERISTIM): b = 0.02, SE = 0.05, p = .71; GROUP
(SHAM): b = 0.08, SE = 0.05, p = .10; with PERISTIM, DAY 2
reference levels: GROUP(SHAM): b = 0.06, SE = 0.05, p =
.20). RTs were faster for match (pred. = 835 msec) than
mismatch trials (pred. = 934 msec; COND(MATCH): b =
−0.11, SE = 0.02, p < .001), but this did not differ be-
tween days (b = −0.01, SE = 0.02, p = .71) or groups
N400
Mean amplitude of by-participant N400 effects (mismatch–
match difference) was analyzed for the lexical recogni-
tion test to determine whether there were differences
between taVNS groups in the strength of semantic rep-
resentations for the Mandarin pseudowords at the end of
each session. Topographic plots of by-participant N400
effects averaged by taVNS condition and session are
shown in Figure 3 with time-series plots for the same
data averaged over parietal sites. These plots show broad
centro-parietal negativity on both days for all taVNS
groups, with comparatively larger amplitude for priming
and peristim.
The best-fitting model for these data (Table 10) reveals
a larger N400 effect amplitude at parietal sites for peri-
stim and priming compared to sham (with SHAM, PAR
reference levels: GROUP(PRIMING): b = −0.40, SE = 0.20,
p = .050; GROUP(PERISTIM): b = −0.43, SE = 0.20, p =
.030) but no difference between peristim and priming
(with PERISTIM, PAR reference levels: GROUP(PRIMING): b =
0.03, SE = 0.22, p = .88). N400 effect amplitude was not
different between groups at frontal sites (GROUP(PRIMING):
b = 0.25, SE = 0.20, p = .22; GROUP(PERISTIM): b = 0.32,
SE = 0.20, p = .11; with PERISTIM, ANT reference levels:
GROUP(PRIMING): b = −0.07, SE = 0.22, p = .76) or central
sites (with SHAM, CNT reference levels: GROUP(PRIMING): b =
−0.24, SE = 0.20, p = .25; GROUP(PERISTIM): b = −0.11,
SE = 0.20, p = .58; with PERISTIM, CNT reference levels:
GROUP(PRIMING): b = −0.13, SE = 0.22, p = .58). Central
and parietal sites were more negative than frontal sites for
Phillips et al.
139
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
***
***
***
*
***
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 9. Best-Fitting Model for Lexical Recognition Log RT
Fixed Effect
(INTERCEPT)
COND(MATCH)
SESS(DAY 2)
GROUP(PERISTIM)
GROUP(SHAM)
GROUP(PERISTIM) × SESS(DAY 2)
GROUP(SHAM) × SESS(DAY 2)
COND(MATCH) × SESS(DAY 2)
Estimate
6.92
−0.11
−0.15
−0.08
−0.08
0.10
0.16
−0.01
Random Effect
Variance
PARTICIPANT(INTERCEPT)
SESS(DAY 2)
COND(MATCH)
SESS(DAY 2) × COND(MATCH)
ITEM(INTERCEPT)
SESS(DAY 2)
COND(MATCH)
SESS(DAY 2) × COND(MATCH)
RESIDUAL
0.01
0.02
0.01
0.01
0.00
0.00
0.00
0.00
0.09
SE
0.04
0.02
0.04
0.05
0.04
0.05
0.05
0.02
SD
0.12
0.13
0.08
0.07
0.04
0.02
0.06
0.05
0.30
df
47.72
31.44
46.64
40.30
40.83
41.28
42.03
28.10
−.32
−.08
.18
−.08
−.47
−.28
t Value
p Value
192.08
−5.36
−4.02
−1.66
−1.84
2.02
3.63
−0.37
Correlation
−.25
−.17
−.44
−.12
< .001
< .001
< .001
.11
.073
.050
< .001
.71
−.48
.64
Reference levels: DAY 1, PRIMING, MISMATCH. Number of obs.: 14008, participants: 45, items: 18.
* p < .05.
** p < .01.
*** p < .001.
all groups (with SHAM, FNT reference levels: REGION(CNT):
b = −0.39, SE = 0.15, p = .012; REGION(PAR): b = −0.62,
SE = 0.15, p < .001; with PRIMING, FNT reference levels:
REGION(CNT): b = −0.87, SE = 0.20, p < .001; REGION(PAR):
b = −1.28, SE = 0.20, p < .001; with PERISTIM, FNT
reference levels: REGION(CNT): b = −0.82, SE = 0.19, p <
.001; REGION(PAR): b = −1.38, SE = 0.19, p < .001).
The difference between frontal and central sites did not
differ between groups (REGION(CNT) × GROUP(PERISTIM):
b = −0.43, SE = 0.25, p = .080; R E G I O N(C N T) ×
GROUP(PRIMING): b = −0.48, SE = 0.25, p = .054; with
PRIMING, FNT reference levels: REGION(CNT) × GROUP
(PERISTIM): b = 0.06, SE = 0.28, p = .84), but peristim
and priming showed larger differences between frontal
and parietal sites compared to sham (REGION(PAR) ×
GROUP(PERISTIM): b = −0.75, SE = 0.25, p = .002; REGION
(PAR) × GROUP(PRIMING): b = −0.65, SE = 0.25, p = .010)
with no difference between these groups (with PRIMING,
FNT reference levels: REGION(PAR) × GROUP(PERISTIM): b =
−0.10, SE = 0.28, p = .71). The SESS term was removed
from the model because it did not improve model fit,
indicating there was no difference in N400 effect topog-
raphy between days for any group. Model-predicted
values are given in Table 11.
Post Hoc Analyses Linking Behavioral Training
Outcomes to ERP Indices of Learning
Post hoc analyses including lexical recognition accuracy
and RT as predictors of N400 amplitude and topography
during passive word learning were conducted to deter-
mine whether there is a direct link between electrophys-
iological measures of semantic encoding during training
and behavioral training outcomes. The best-fitting model
testing inclusion of RT as a predictor for the passive word
learning N400 did not retain any fixed-effects involving
RT, indicating RT did not improve the fit of the passive
word learning N400 model without RT, reported above.
In contrast, the best-fitting model testing inclusion of
accuracy as a predictor for the passive word learning
N400 retained three-way REGION × GROUP × ACC and
REGION × SESS × ACC interactions and all lower-order terms
140
Journal of Cognitive Neuroscience
Volume 34, Number 1
Table 10. Best-Fitting Model for Lexical Recognition N400 Effect (Mismatch–Match) Mean Amplitude
Fixed Effect
(INTERCEPT)
REGION(CNT)
REGION(PAR)
GROUP(PERISTIM)
GROUP(PRIMING)
REGION(CNT) × GROUP(PERISTIM)
REGION(PAR) × GROUP(PERISTIM)
REGION(CNT) × GROUP(PRIMING)
REGION(PAR) × GROUP(PRIMING)
Est.
0.08
−0.39
−0.62
0.32
0.25
−0.43
−0.75
−0.48
−0.65
Random Effect
Variance
PARTICIPANT(INTERCEPT)
RESIDUAL
0.07
1.42
Reference levels: FNT, SHAM. Number of obs.: 810, participants: 45.
* p < .05.
** p < .01.
*** p < .001.
SE
0.12
0.15
0.15
0.20
0.20
0.25
0.25
0.25
0.25
SD
0.27
1.19
df
165.44
759.00
759.00
165.44
165.44
759.00
759.00
759.00
759.00
t Value
p Value
0.64
−2.51
−4.06
1.62
1.23
−1.75
−3.08
−1.93
−2.59
.52
.012
< .001
.10
.22
.080
.002
.054
.010
*
***
**
**
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
(Table 12, predicted values plotted in Figure 4). Removal of
the four-way REGION × GROUP × SESS × ACC term during
model fitting indicates that group differences in REGION ×
ACC interactions were consistent between days, but,
because of the retention of SESS in the model, estimates
for the REGION × GROUP × ACC interactions are given sepa-
rately for each day below.
For the peristim group, on both days, higher lexical
recognition accuracy predicted greater negativity over
parietal sites and reduced negativity over frontal sites
with the effect of ACC differing between frontal and
central/parietal sites but not between central and parietal
sites (with PERISTIM, FNT, DAY 1 reference levels: ACC: b =
0.51, SE = 0.24, p = .034; REGION(CNT) × ACC: b = −0.73,
SE = 0.27, p = .008; REGION(PAR) × ACC: b = −0.94, SE =
0.38, p = .015; with PERISTIM, FNT, DAY 2 reference levels:
ACC: b = 0.67, SE = 0.24, p = .005; REGION(CNT) × ACC: b =
−0.78, SE = 0.27, p = .005; REGION(PAR) × ACC: b = −1.29,
SE = 0.38, p < .001; with PERISTIM, PAR, DAY 1 reference
levels: ACC: b = −0.44, SE = 0.22, p = .046; REGION(CNT) ×
ACC: b = 0.21, SE = 0.27, p = .43; with PERISTIM, PAR, DAY 2
reference levels: ACC: b = −0.62, SE = 0.22, p = .005;
REGION(CNT) × ACC: b = 0.51, SE = 0.27, p = .056).
For the sham group, higher lexical recognition accuracy
predicted greater negativity over parietal sites on both
days and reduced negativity over frontal sites on Day 2
but not Day 1. On Day 1, the effect of ACC differed be-
tween frontal and parietal sites but not between central
and frontal or parietal sites whereas, on Day 2, the effect
of ACC differed between frontal and central/parietal sites
but not between central and parietal sites (with SHAM, FNT,
DAY 1 reference levels: ACC: b = 0.28, SE = 0.22, p = .20;
REGION(CNT) × ACC: b = −0.56, SE = 0.25, p = .025;
REGION(PAR) × ACC: b = −0.70, SE = 0.36, p = .055; with
SHAM, FNT, DAY 2 reference levels: ACC: b = 0.45, SE =
0.22, p = .043; REGION(CNT) × ACC: b = −0.61, SE = 0.25,
p = .015; REGION(PAR) × ACC: b = −1.05, SE = 0.36, p =
.004; with SHAM, PAR, DAY 1 reference levels: ACC: b =
−0.41, SE = 0.20, p = .039; REGION(CNT) × ACC: b = 0.14,
SE = 0.24, p = .56; with SHAM, PAR, DAY 2 reference levels:
ACC: b = −0.60, SE = 0.20, p = .003; REGION(CNT) × ACC:
b = 0.44, SE = 0.24, p = .070).
For the priming group, higher lexical recognition
accuracy predicted reduced negativity over parietal sites
and greater negativity over frontal sites with the effect of
ACC differing between frontal, central, and parietal sites on
Day 1 but only differing between frontal and central/parietal
but not between central and parietal sites on Day 2
Table 11. Model-Predicted Values for Lexical Recognition N400
Effect Amplitude (μV) by Region and taVNS Group
Peristim
Priming
Frontal
Central
Parietal
0.40
−0.42
−0.98
0.33
−0.54
−0.94
Sham
0.08
−0.31
−0.54
Phillips et al.
141
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 12. Best-Fitting Post Hoc Model of Passive Word Learning N400 Mean Amplitude Including Lexical Recognition Accuracy as a
Fixed-Effect Predictor
Fixed Effect
(INTERCEPT)
REGION(CNT)
REGION(PAR)
LAT(LEFT)
LAT(RIGHT)
SESS(DAY 2)
GROUP(SHAM)
GROUP(PERISTIM)
ACC
REGION(CNT) × LAT(LEFT)
REGION(PAR) × LAT(LEFT)
REGION(CNT) × LAT(RIGHT)
REGION(PAR) × LAT(RIGHT)
REGION(CNT) × SESS(DAY 2)
REGION(PAR) × SESS(DAY 2)
LAT(LEFT) × GROUP(SHAM)
LAT(RIGHT) × GROUP(SHAM)
LAT(LEFT) × GROUP(PERISTIM)
LAT(RIGHT) × GROUP(PERISTIM)
REGION(CNT) × GROUP(SHAM)
REGION(PAR) × GROUP(SHAM)
REGION(CNT) × GROUP(PERISTIM)
REGION(PAR) × GROUP(PERISTIM)
SESS(DAY 2) × GROUP(SHAM)
SESS(DAY 2) × GROUP(PERISTIM)
GROUP(PERISTIM) × ACC
GROUP(SHAM) × ACC
REGION(PAR) × ACC
REGION(CNT) × ACC
SESS(DAY 2) × ACC
REGION(CNT) × SESS(DAY 2) × GROUP(SHAM)
REGION(PAR) × SESS(DAY 2) × GROUP(SHAM)
REGION(CNT) × SESS(DAY 2) × GROUP(PERISTIM)
REGION(PAR) × SESS(DAY 2) × GROUP(PERISTIM)
REGION(CNT) × GROUP(SHAM) × ACC
REGION(PAR) × GROUP(SHAM) × ACC
REGION(CNT) × GROUP(PERISTIM) × ACC
Est.
−1.06
−0.16
0.63
0.16
0.09
1.10
0.82
1.21
−0.88
0.60
0.81
0.53
0.44
−1.20
−2.14
−0.10
−0.08
−0.58
−0.12
−1.04
−1.32
−1.23
−1.53
−1.22
−1.26
1.39
1.16
1.81
0.98
0.17
1.52
2.25
1.24
2.35
−1.54
−2.51
−1.71
SE
0.41
0.42
0.68
0.17
0.17
0.19
0.51
0.56
0.25
0.17
0.17
0.17
0.17
0.27
0.27
0.17
0.17
0.19
0.19
0.52
0.85
0.57
0.93
0.24
0.27
0.33
0.32
0.40
0.28
0.11
0.34
0.34
0.37
0.37
0.36
0.53
0.38
df
42.65
45.13
37.93
640.12
640.12
640.12
40.98
40.98
230.32
640.12
640.12
640.12
640.12
640.12
640.12
640.12
640.12
640.12
640.12
40.27
36.32
40.27
36.32
640.12
640.12
215.17
180.79
307.74
135.03
656.45
640.12
640.12
640.12
640.12
106.92
234.63
119.40
t Value
p Value
−2.62
−0.38
0.93
0.94
0.53
5.74
1.62
2.17
−3.55
3.54
4.74
3.08
2.60
−4.42
−7.91
−0.58
−0.47
−3.10
−0.63
−2.01
−1.56
−2.18
−1.66
−5.04
−4.76
4.15
3.61
4.49
3.46
1.55
4.44
6.58
3.31
6.27
−4.27
−4.72
−4.52
.012
*
.71
.36
.35
.60
< .001
***
.11
.036
< .001
< .001
< .001
.002
.009
< .001
< .001
.56
.64
.002
.53
.051
.13
.035
.11
< .001
< .001
< .001
< .001
< .001
< .001
.12
< .001
< .001
< .001
< .001
< .001
< .001
< .001
*
***
***
***
**
**
***
***
**
*
***
***
***
***
***
***
***
***
***
***
***
***
***
142
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 12. (continued )
Fixed Effect
REGION(PAR) × GROUP(PERISTIM) × ACC
REGION(CNT) × SESS(DAY 2) × ACC
REGION(PAR) × SESS(DAY 2) × ACC
Random Effect
PARTICIPANT(INTERCEPT)
REGION(CNT)
REGION(PAR)
RESIDUAL
Est.
−2.75
−0.05
−0.35
Variance
1.65
1.56
4.93
0.66
SE
0.55
0.15
0.15
SD
1.28
1.25
2.22
0.81
df
t Value
p Value
294.87
666.79
645.57
−5.04
−0.35
−2.32
< .001
***
.73
.021
*
Correlation
−.83
−.95
.90
Reference levels: FNT, MID, PRIMING, DAY 1. Number of obs.: 810, participants: 45.
* p < .05.
** p < .01.
*** p < .001.
(with PRIMING, FNT, DAY 1 reference levels: ACC: b =
−0.88, SE = 0.25, p < .001; REGION(CNT) × ACC: b =
0.98, SE = 0.28, p < .001; REGION(PAR) × ACC: b = 1.81,
SE = 0.40, p < .001; with PRIMING, FNT, DAY 2 reference
levels: ACC: b = −0.71, SE = 0.25, p = .004; REGION(CNT) ×
ACC: b = 0.93, SE = 0.28, p = .001; REGION(PAR) × ACC:
b = 1.46, SE = 0.40, p < .001; with PRIMING, PAR, DAY 1
reference levels: ACC: b = 0.93, SE = 0.23, p < .001;
REGION(CNT) × ACC: b = −0.82, SE = 0.28, p = .003; with
PRIMING, PAR, DAY 2 reference levels: ACC: b = 0.74, SE =
0.23, p = .001; REGION(CNT) × ACC: b = −0.52, SE = 0.28,
p = .061).
The ACC × REGION interaction did not differ between
peristim and sham nor did the simple effect of ACC at any
region (with PERISTIM, FNT reference levels: GROUP(SHAM) ×
ACC: b = −0.22, SE = 0.31, p = .48; REGION(CNT) ×
GROUP(SHAM) × ACC: b = 0.17, SE = 0.35, p = .63;
REGION(PAR) × GROUP(SHAM) × ACC: b = 0.24, SE = 0.52,
p = .64; with PERISTIM, PAR reference levels: GROUP(SHAM) ×
ACC: b = 0.02, SE = 0.28, p = .94; REGION(CNT) × GROUP
(SHAM) × ACC: b = −0.07, SE = 0.34, p = .84; with
PERISTIM, CNT reference levels: GROUP(SHAM) × ACC: b =
−0.05, SE = 0.25, p = .84). In contrast, the simple effect
of ACC differed at frontal and parietal sites but not central
sites between priming and both peristim and sham (with
PRIMING, FNT reference levels: GROUP(PERISTIM) × ACC: b =
1.39, SE = 0.33, p < .001; GROUP(SHAM) × ACC: b = 1.16,
SE = 0.32, p < .001; with PRIMING, PAR reference levels:
GROUP(PERISTIM) × ACC: b = −1.36, SE = 0.30, p < .001;
GROUP(SHAM) × ACC: b = −1.34, SE = 0.29, p < .001; with
PRIMING, CNT reference levels: GROUP(PERISTIM) × ACC: b =
−0.33, SE = 0.27, p = .23; GROUP(SHAM) × ACC: b = −0.38,
Figure 4. Model-predicted
N400 mean amplitudes for the
effect of taVNS group (top row)
and session (bottom row) on
the interaction of lexical
recognition accuracy and
electrode region during passive
word learning, plotted with
standard error shading.
Phillips et al.
143
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
SE = 0.26, p = .14) driving the significant REGION × GROUP ×
ACC interactions (REGION(CNT) × GROUP(PERISTIM) × ACC:
b = −1.71, SE = 0.38, p < .001; R E G I O N(P A R) ×
GROUP(PERISTIM) × ACC: b = −2.75, SE = 0.55, p < .001;
REGION(CNT) × GROUP(SHAM) × ACC: b = −1.54, SE = 0.36,
p < .001; REGION(PAR) × GROUP(SHAM) × ACC: b = −2.51,
SE = 0.53, p < .001).
Across taVNS groups, the effect of ACC did not differ
between days at any region (with FNT, DAY 1 reference
levels: SESS(DAY 2) × ACC: b = 0.17, SE = 0.11, p = .12;
with CNT, DAY 1 reference levels: SESS(DAY 2) × ACC: b =
0.11, SE = 0.11, p = .29; with PAR, DAY 1 reference levels:
SESS(DAY 2) × ACC: b = −0.19, SE = 0.11, p = .081).
However, the REGION × SESS × ACC interaction reveals that
between-days trends in the effect of ACC differed between
parietal and frontal/central sites but not between frontal
and central sites (REGION(PAR) × SESS(DAY 2) × ACC: b =
−0.35, SE = 0.15, p = .021; REGION(CNT) × SESS(DAY 2) ×
ACC: b = −0.05, SE = 0.15, p = .73; with PAR, DAY 1 reference
levels: REGION(CNT) × SESS(DAY 2) × ACC: b = 0.30, SE =
0.15, p = .046). Over parietal sites, the trend was toward
higher lexical recognition accuracy predicting compara-
tively greater negativity on Day 2 than Day 1 (more nega-
tive slope on Day 2 compared to Day 1 in Figure 4) but
this trend was reversed over frontal and central sites.
DISCUSSION
The results of this study demonstrate that priming and
peristim taVNS enhance behavioral outcomes of initial
L2 lexical training for words partly distinguished by
Mandarin tone contrasts and that these improvements
are linked to stronger lexico-semantic encoding. All
groups showed evidence of learning, reflected in higher
likelihood of accuracy and improved processing speed
reflected in faster RTs on Day 2 across tasks, but pairing
taVNS with training led to greater behavioral gains in the
lexical recognition test than training alone in line with
findings reported for the larger data set (Pandža et al.,
2020). Differences in N400 effect amplitude between
taVNS groups in the lexical recognition test support the
RQ1 hypothesis that pairing taVNS with lexical training
results in more robust lexico-semantic representations.
The N400 results for the passive word learning task do
not clearly support the hypothesis that taVNS leads to fas-
ter semantic encoding during training, but they do pro-
vide some support for the RQ2 hypothesis that priming
and peristim taVNS benefit lexico-semantic development
in different ways. Behavioral and ERP results relevant to
each RQ are considered in turn below.
taVNS Improves Recognition of Newly Learned L2
Lexical Items
The priming group made accurate responses in the lexi-
cal recognition test faster on Day 2 than Day 1, reflecting
improved recognition speed or perhaps also participants’
confidence in their judgments, whereas accurate re-
sponse speed did not change for the other two taVNS
groups. The peristim group was highly and equally likely
to accurately identify matches and mismatches between
English words and Mandarin pseudowords, reflecting im-
proved recognition memory for word pairs, whereas the
priming and sham groups were less likely to correctly
identify mismatches than matches. Higher performance
on match trials in the priming and sham groups, on par
with the peristim group, likely reflects bias toward re-
sponding “match” that was observed for priming and
sham (and to a lesser extent, peristim) rather than learn-
ing per se, because each Mandarin pseudoword appeared
in the same number of match and mismatch trials.
Contrary to expectations, taVNS did not influence accu-
racy or RT for the active word learning task. One potential
explanation is that taVNS may have had a comparatively
weak effect on modulating the formation and strengthen-
ing of lexico-semantic representation during the passive
compared to active word learning task. In this case, prim-
ing and peristim would perform similarly to sham at the
start of active word learning (at least on Day 1), but im-
prove more rapidly during the task. Within-task differ-
ences in learning trajectory may not be captured in the
analysis of mean performance over the task. Another po-
tential explanation is that the active word learning task
(unlike the lexical recognition test) provided accuracy
feedback each trial, which could inform following re-
sponses. This may have allowed participants to perform
well without relying on memories formed in prior learn-
ing tasks, thereby attenuating any effects of taVNS on
prior learning. This attenuation combined with a low trial
number (n = 36) may have reduced the taVNS effect
size below a detectable level in these data. However, it
should be noted that identical accuracy and RT model
structures were selected, with very similar estimates,
when the same model fitting procedure was applied to
the full data set (n = 82 participants).
Aside from the null taVNS effect in active word learn-
ing, participants were slower to pick the correct response
when Mandarin translation equivalents for response
items differed only in lexical tone versus when they had
different syllables. Cue status (nonnative tone vs. native
syllable phonemes) may have contributed to this effect,
but differences in cue temporal features likely also played
a role. In two thirds (66.8%) of the trials with different
syllables, the difference occurred on the first phoneme.
The first phoneme also carried tone information in two
thirds of the stimuli (/ba/, /bi/), but tone direction contin-
ued to unfold over the second phoneme. Given that
speakers of nontonal languages are less sensitive to tone
direction (e.g., Chandrasekaran et al., 2010), participants
likely needed to hear more than the first phoneme to dif-
ferentiate items differing only in tone, while they more
often than not could distinguish items with different syl-
lables after hearing only the first phoneme. Accuracy
144
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
results also suggest that cue status played a comparatively
minor role in performance in this task. The likelihood
of responding accurately was higher when response item
translation equivalents differed in tone and syllable than
when they differed in tone or syllable alone, with no dif-
ference between these conditions. This pattern suggests
the number of cues (two vs. one) distinguishing re-
sponse items was more important than whether the
cue was contrastive in participants’ L1. If, instead, perfor-
mance was driven by cue status, accuracy differences
would be expected between trials where response item
translation equivalents differed only in tone or only in
syllable.
Lastly, this study replicated some previous findings in
the literature regarding individual differences in lexical
tone–word learning (e.g., Bowles et al., 2016; Dittinger
et al., 2016). Higher likelihood of accuracy on the active
word learning task was predicted by higher self-rated mu-
sicianship and ability to identify nonlinguistic pitch,
whereas higher accuracy on the lexical recognition test
was predicted by higher self-rated musicianship.
Notably, these factors did not affect RT or ERP results.
That these factors did not predict N400 amplitude or to-
pography in the passive word learning task or N400 effect
amplitude in the lexical recognition test is unexpected
because very similar tasks in Dittinger et al. (2016)
showed increased centro-parietal N400 amplitude in pas-
sive word learning and strong centro-parietal N400 effects
in lexical recognition for professional musicians. One
possible explanation for the difference is that Dittinger
et al. (2016) compared professional musicians to nonmu-
sicians, whereas there was more variation in musician-
ship and no professional musicians in this study.
taVNS Strengthens Lexico-Semantic Encoding
Beyond Training Alone
A strong centro-parietal N400 effect (mismatch–match)
was observed in the lexical recognition test on both days
for priming and peristim. The N400 effect was not statis-
tically different between priming and peristim, but was
larger over parietal sites for both groups on both days
compared to sham. Given N400 amplitude correlates
negatively with a word’s expectancy or predictability from
prior context (Lau, Holcomb, & Kuperberg, 2013; Van
Petten & Luka, 2012; Kutas & Federmeier, 2011; Lau
et al., 2008; Kutas & Hillyard, 1980), a larger N400 effect
at parietal sites indicates the English translation equiva-
lents primed Mandarin pseudowords to a greater degree
on both days for priming and peristim versus sham. This
difference in N400 effect strength largely parallels that in
Dittinger et al. (2016), where musicians showed a centro-
parietal N400 effect (mismatch–match) in both blocks of
a lexical recognition task very similar to that used in this
study, but nonmusicians showed the opposite effect over
fronto-central regions (larger N400 for match than mis-
match). The sham group in this study showed a centro-
parietal N400 effect (possibly indicating stimuli in this
study were easier to learn than those in Dittinger et al.,
2016), although the larger N400 effect at parietal sites
for priming and peristim suggests pairing either method
of taVNS delivery with lexical training provided an
advantage.
The N400 results in the passive word learning task sug-
gest taVNS did not reduce the number of exposures
needed to encode meaning of the Mandarin pseudo-
words. While the priming group showed a fronto-central
to centro-parietal N400 shift between days, perstim and
sham showed centro-parietal N400s on both days. The
shift observed for priming—consistent with previous
findings of frontal N400s during initial stages of word
learning and central-parietal N400s during later stages,
when learners are presumed to have developed stronger
semantic representations for learned words (Dittinger
et al., 2016; Rodríguez-Fornells et al., 2009)—may sug-
gest weaker semantic encoding of Mandarin pseudo-
words for the priming group on Day 1 of passive word
learning compared to peristim and sham but comparable
encoding strength by Day 2. There is some evidence that
iVNS delivered continuously at higher intensity levels
during learning and testing can negatively affect recogni-
tion memory for figures, although verbal memory was
unaffected and RT was improved in the same study
(Helmstaedter, Hoppe, & Elger, 2001). Thus, it is possi-
ble that priming taVNS could have impaired semantic en-
coding during passive word learning on Day 1. However,
any negative effects of priming taVNS in this study would
result from differences in total amount of current re-
ceived rather than current amplitude because taVNS am-
plitude was calibrated to the same perceptual threshold
for peristim and priming. It is also not clear why similar
levels of priming taVNS would be detrimental to encod-
ing 1 day but not the next. Thus, an alternative explana-
tion for the priming group’s fronto-central N400 on Day 1
of passive word learning that considers potential taVNS-
related changes in cognitive functions thought to contrib-
ute to frontal N400 topography during word learning is
given below.
The centro-parietal N400 on Day 1 for peristim and
sham suggests these individuals developed semantic rep-
resentations for target words with fewer word exposures
than were needed for participants in Dittinger et al.
(2016), where N400 amplitude was largest fronto-
centrally during Block 1 of a very similar passive word
learning task, only increasing at centro-parietal regions
during Block 2 for musicians. This again suggests stimuli
in this study were easier to learn, perhaps because of
items differing along only one nonnative phonological
contrast. Although we did not observe typical N400 topo-
graphic shifts between days that would indicate faster
lexico-semantic encoding for taVNS, the N400 for the
peristim group showed a left-lateralized distribution on
both days whereas N400s for priming and sham were fo-
cused over midline sites. Similarly, Dittinger et al. (2016)
Phillips et al.
145
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
showed an increased left-lateralized N400 amplitude for
learned words during Block 2 of passive word learning
for musicians, but not nonmusicians. Other studies inves-
tigating individual differences in word learning have also
shown left-lateralized N400 effects (e.g., Perfetti et al.,
2005). Within a single session of learning rare words,
Perfetti et al. (2005) found that, although more- and
less-skilled readers showed similar right-lateralized N400
effects for related versus unrelated words during a se-
mantic judgment task, only more-skilled readers also
showed a left-lateralized N400 effect. The left-lateralized
differences in this study, perhaps distinguishable by
some measure of skill, may indicate a greater integration
of new lexical items into an existing semantic network
that was facilitated by peristim taVNS. However, it should
be noted that post hoc analyses did not show a direct re-
lationship between these laterality differences and behav-
ioral performance during lexical recognition.
Recognition Accuracy (But Not RT) Predicts N400
Changes during Passive Word Learning
Post hoc modeling uncovered direct relationships be-
tween lexical recognition accuracy and N400 indices of
lexico-semantic encoding during passive word learning.
Across taVNS groups, for individuals with higher recogni-
tion accuracy compared to those with lower accuracy, pa-
rietal sites were numerically more negative whereas
frontal and central sites were numerically less negative
on Day 2 than Day 1. This reflects a parietal shift in the
passive word learning N400 between days that was
greater for individuals with higher recognition, which
aligns with previous observations of N400s elicited during
early word learning and between novel and known words
(e.g., Dittinger et al., 2016; Rodríguez-Fornells et al.,
2009). This finding suggests behavioral learning improve-
ments in this study are tied to changes in strength of
lexico-semantic representations of the Mandarin pseudo-
words, with individuals who showed better recognition of
learned words also showing increases in strength of
lexico-semantic representations during learning (via
shifts in N400 topography) whereas those with lower
recognition showing less change in N400 topography
between days.
Of greater interest to this study, post hoc modeling
also revealed differences between taVNS groups in the
link between lexical recognition accuracy and passive
word learning N400 topography. The same pattern was
observed for peristim and sham, with higher recognition
accuracy predicting increased negativity over parietal
sites and reduced negativity over frontal sites during
both days of passive word learning. This pattern aligns
with the literature linking stronger lexico-semantic repre-
sentations to centro-parietal N400s and weaker represen-
tations to fronto-central N400s and suggests that
behavioral learning improvements for the peristim and
sham groups are tied directly to the strength of the
Mandarin pseudoword lexico-semantic representations.
For peristim but not sham, there were other parallels
between ERP and behavioral measures of lexical learning
across days (higher accuracy for mismatch trials and
strong centro-parietal N400 effects in lexical recognition,
left-lateralized N400s in passive word learning) that may
suggest other potential links between stronger lexico-
semantic encoding and improved lexical recognition that
result from pairing peristim taVNS with lexical training. If
present, however, these links appear to be indirect as the
interaction between passive word learning N400 laterality
and lexical recognition accuracy was not retained in the
post hoc model, indicating that recognition accuracy
does not predict N400 laterality differences (or the rela-
tionship is nonlinear) in the present data.
Compared to peristim and sham, the priming group
showed a qualitatively different relationship between lex-
ical recognition accuracy and passive word learning N400
topography, with higher accuracy predicting greater neg-
ativity over frontal sites and reduced negativity over parie-
tal sites. This pattern does not align with an interpretation
of fronto-central N400 topography reflecting weaker
lexico-semantic representations, but it is perhaps consis-
tent with the idea that frontal negativity during initial lex-
ical learning may result from increased engagement of
certain cognitive functions. In this view, the fronto-
central focus of the passive word learning N400s elicited
for priming on Day 1 may more accurately reflect other
aspects of performance, potentially more effortful pro-
cessing related to cognitive control (Elgort et al., 2015;
Mestres-Missé et al., 2007; Rodriguez-Fornells et al.,
2006). Increased effort may be associated with executive
functions necessary to promote controlled semantic re-
trieval (Mestres-Missé et al., 2007), including interference
resolution, in which features of the nontarget language
are suppressed to promote processing of the target lan-
guage (Rodriguez-Fornells et al., 2006). This controlled
processing becomes more automatic as learned words
become more integrated into the lexicon (Bakker,
Takashima, van Hell, Janzen, & McQueen, 2015). In our
study, participants who received priming taVNS may
have had greater capacity to sustain attentional effort
that resulted in more controlled lexical access/retrieval
processes on Day 1, eliciting a frontal negativity. As
the Mandarin items became more integrated with more
training, processing would be expected to become more
automatic and the frontal distribution of the N400 would
be expected to lessen, consistent with the observed re-
sults on Day 2.
For the priming group, the improvement in lexical rec-
ognition RT between days would also be consistent with
the above interpretation, although post hoc modeling in-
dicates any potential link between passive word learning
N400 topography and recognition RT is indirect. More
controlled processing could be expected to result in
slower RTs on Day 1 whereas more automatic processing
would result in faster RTs on Day 2. However, all terms
146
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
involving recognition RT were removed from the post hoc
passive word learning N400 model, indicating that lexical
recognition RT does not predict passive word learning N400
topography (or the relationship is nonlinear) in these
data. Attributing the priming group’s RT improvement
to a shift from more controlled to more automatic process-
ing (rather than purely reflecting semantic representation
strength) also makes sense in light of the apparent incon-
sistency between the lexical recognition RT improvement
from Day 1 to Day 2 and the strong centro-parietal N400
effect that was observed on both days in the same task.
The strong N400 effect on Day 1 suggests priming taVNS
strengthened semantic encoding for Mandarin pseudo-
words without parallel effects seen in explicit behavioral
responses. This mirrors effects seen in other studies of
L2 early word learning (e.g., McLaughlin et al., 2004) and
aligns with other studies of lexico-semantic processing
that find ERP but not behavioral differences (e.g.,
Barber, Otten, Kousta, & Vigliocco, 2013; Balass, Nelson,
& Perfetti, 2010).
Priming and Peristim taVNS May Differentially
Influence Initial L2 Lexical Learning
The behavioral learning advantages observed for priming
and peristim over sham and the striking similarities be-
tween N400 effects elicited for peristim and priming in
the lexical recognition test suggest both taVNS timings
enhanced lexico-semantic encoding to a similar degree.
However, the finding that priming taVNS improved RTs
but not accuracy, and peristim taVNS improved accuracy
but not RT, paired with distinct N400 patterns during pas-
sive word learning may suggest priming and peristim
taVNS support L2 lexical learning through different un-
derlying mechanisms. One way taVNS could potentially
lead to stronger semantic encoding is by enhancing
LTP during word learning tasks, thus impacting memory
formation for the Mandarin pseudowords. Although this
explanation has been suggested for memory improve-
ments following VNS administered during memory en-
coding and consolidation, after stimulus presentation
( Jacobs et al., 2015), priming and peristim taVNS could
potentially affect LTP because increased NE levels in
the basolateral amygdala have been observed to last
more than an hour after administering 30 sec of VNS in
animal models (Hassert, Miyashita, & Williams, 2004).
Stimuli in our training tasks were presented within a
few seconds of peristim taVNS and within 30 min of
priming taVNS. Another possibility is that, by modulating
LC-NE activity, taVNS helped participants to maintain an
optimal state of focused attention during the word
learning tasks, thus indirectly leading to better encoding
of target items.
The data collected in this study do not permit direct
examination of LC-NE activity or LTP. However, the re-
sults of the post hoc analyses contextualized with respect
to the passive word learning task pupillometry results
presented in Pandža et al. (2020) suggest that beneficial
effects of priming taVNS, but not peristim taVNS, may be
in part related to sustained attentional effort during learn-
ing. The potential links drawn below between the pres-
ent results and maintenance of attentional effort are
necessarily speculative as the multidimensional nature
of pupillary changes that characterize differences in effort
deployment in Pandža et al. (2020) does not easily permit
linking the present results to a unidimensional indicator
of sustained effort, but it is worth some discussion for its
potential value in guiding future research. Although N400
amplitude has been linked to concurrently recorded task-
evoked pupillary response amplitude previously (Kuipers
& Thierry, 2011), the design of this study does not permit
the same statistical comparisons.
During the passive word learning task, participants
were instructed to memorize the meaning of the
Mandarin pseudowords. Thus, stimulus-evoked changes
in pupil size were interpreted in Pandža et al. (2020) to
reflect the level of attention or effort that participants al-
located to memorizing Mandarin–English word pairs.
With this interpretation, the pupillometry results for pas-
sive word learning indicated the priming group main-
tained similar levels of sustained effort during trials on
Day 1 and 2, whereas peristim and sham groups showed
less sustained effort during trials on Day 2. Peristim and
sham had similar between-days changes in pupillary re-
sponses compared to priming, showing an earlier deploy-
ment of cognitive effort that was less sustained on Day 2
compared to Day 1. This between-days reduction in sus-
tained effort was significantly greater for peristim than
sham and priming, and greater for sham compared to
priming. This pattern of results suggests the lexical
learning advantage for the priming group in this study
is related to sustaining attentional effort during learning
whereas the advantage for the peristim group is not. The
post hoc analyses in this study lend further support to
this conclusion by revealing a direct link between recog-
nition accuracy and N400 amplitude during learning, with
a qualitatively distinct pattern observed for priming
where accuracy predicted a frontal negativity thought to
reflect potentially effortful cognitive control processes.
Explaining stronger lexico-semantic encoding and faster
lexical processing for the priming group as a result of
better sustained attentional effort because of taVNS is
straightforward given the role of attention in learning:
“…learning, particularly after a sensitive period, appears
to be a gated system, through which attention (via acetyl-
choline) can facilitate or restrict plasticity” ( White et al.,
2013, p. 12).
The explanation for stronger lexico-semantic encoding
and better learning for the peristim group seems unlikely
to involve sustained attentional effort because these par-
ticipants had the biggest drop off in effort between days
yet they were the most likely to respond correctly in the
lexical recognition test. These results would be expected,
however, if peristim taVNS enhanced memory formation
Phillips et al.
147
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
directly via its effects on LTP. The N400s elicited for peri-
stim and sham on Day 1 of passive word learning suggest
individuals in these groups had developed some level of
lexico-semantic representation for the Mandarin pseudo-
words during this task and the fact that there was no sig-
nificant change between days in any ERP or behavioral
measure for either group indicates that the semantic rep-
resentations that peristim and sham developed early dur-
ing training on Day 1 remained stable through Day 2.
Because there is no indication from the passive word
learning task pupillometry findings that the peristim
group exerted more effort than the sham group during
learning, the stronger lexico-semantic representations
developed by the peristim group evidenced in the lexical
recognition test results are presumably because of peri-
stim taVNS increasing learning efficiency, which is consis-
tent with the idea of taVNS benefiting memory encoding
more directly via its effects on LTP.
Limitations
As an initial exploration into potential taVNS effects,
taVNS was administered prior to (priming) and during
(peristim) the word learning tasks and the lexical recog-
nition test in this study. Thus, we cannot rule out the
possibility that taVNS effects on task engagement or
memory retrieval during the lexical recognition test con-
tributed to better performance without necessarily
influencing learning. Although lexical recognition test
ERP results indicate participants who received taVNS de-
veloped stronger lexico-semantic representations, sug-
gesting taVNS affected learning itself, future work
should tease apart the effects of taVNS administered dur-
ing training and testing phases to further support this
conclusion. As pointed out by one reviewer, the inclusion
of visual representations of the lexical tones in this study
may have potentially diminished taVNS effects. Future
work may gain further insight into the efficacy of taVNS
by studying its effects in more ecologically valid language
learning contexts. The design of this study also does not
provide evidence on whether taVNS facilitates integration
of lexico-semantic representations for newly learned L2
words into existing semantic networks or the longevity
of taVNS-related learning improvements. Dittinger et al.
(2016) found evidence from a semantic priming task that
musical experience facilitated the integration of target
item lexico-semantic representations into existing seman-
tic networks; however, this study did not include a simi-
lar semantic priming task and therefore does not provide
data necessary to support a similar claim for taVNS.
Similarly, Dittinger et al. found that the subset of profes-
sional musicians who repeated the matching and seman-
tic tasks 5 months after training showed higher accuracy
on the match task compared to the subset of nonmusi-
cians who completed the same posttest. (The advantage
for musicians in the semantic task, however, disappeared
at 5 months.) Although this study provides useful initial
evidence of the impact of taVNS on language learning,
future studies should assess semantic integration and
the longevity of taVNS effects. Lastly, it is possible that
the effects of taVNS on Mandarin pseudoword learning
are partly because of taVNS enhancing low-level auditory
processing of the novel phonological features distin-
guishing Mandarin pseudowords. Recent studies have
shown that pairing direct VNS with pure tones and
speech sounds can strengthen responses of primary au-
ditory cortex neurons to these stimuli in humans and an-
imal models (Engineer et al., 2011, 2015; De Ridder,
Vanneste, Engineer, & Kilgard, 2014). Although these
changes have been observed following several weeks of
iVNS, they were not found within a single day of taVNS-
paired training of Mandarin tones (Llanos et al., 2020),
and it is unclear whether changes could be expected
following just 2 days of training in this study.
Conclusion
Individuals lose sensitivity at an early age to phonological
features that do not distinguish word meaning in their
native language, which can negatively impact L2 lexical
learning and use. In this study, we found that combining
two timings of taVNS with lexical training markedly im-
proved native English speakers’ recognition ability for a
relatively small set of L2 pseudowords distinguished in
part by a particularly challenging nonnative phonological
contrast: Mandarin lexical tone. Our novel ERP findings
show that taVNS strengthened lexico-semantic encoding
of the target items, which likely contributed to behavioral
improvements although the links appear to be indirect.
Both priming and peristim taVNS strengthened lexico-
semantic representations to a similar degree, although
they likely accomplish this via different underlying mech-
anisms. These findings contribute to the limited literature
examining the effects of neuromodulatory interventions
on L2 lexical learning and suggest that taVNS holds prom-
ise as a safe and effective technique for supporting adult
L2 acquisition. Although these data are not able to eluci-
date the neurophysiological mechanisms driving taVNS-
enhanced learning, interpreting these novel findings in
the context of pupillometry results from the same study
(Pandža et al., 2020) suggests that stronger semantic en-
coding is in part facilitated by modulation of attentional
effort and thus the positive effects of taVNS likely have
more general applicability beyond the specific case of
L2 lexical learning.
Acknowledgments
We thank Eric Pelzl for his assistance with stimuli selection and
design; Matthew Turner and Sara McConnell for their assistance
with data collection; Meredith Hughes, Jason Struck, and Alison
Tseng for their assistance with blinding the data and assigning
groups; Jarrett Lee for assistance programming taVNS; and
Henk Haarmann and Greg Colflesh for contributions to earlier
portions of the overall project. This material is based upon work
148
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
supported by the Naval Information Warfare Center and
D e f e n s e A d v a n c e d R e s e a r c h P r o j e c t s A g e n c y u n d e r
Cooperative Agreement No. N66001–17-2-4009. The identifica-
tion of specific products or scientific instrumentation is consid-
ered an integral part of the scientific endeavor and does not
constitute endorsement or implied endorsement on the part
of the author, DoD, or any component agency. The views ex-
pressed in this article are those of the author and do not reflect
the official policy of the Department of Army/Navy/Air Force,
Department of Defense, or U.S. Government.
Reprint requests should be sent to Ian Phillips, Audiology and
Speech Pathology Center, Walter Reed National Military
Medical Center, Room 5400, 4954 North Palmer Road, America
Building, Bethesda, MD 20889, United States, or via e-mail:
iphillip@umd.edu.
Author Contributions
Ian Phillips: Data curation; Formal analysis; Investigation;
Methodology; Project administration; Visualization;
Writing—Original draft; Writing—Review & editing.
Regina C. Calloway: Data curation; Formal analysis;
Writing—Original draft; Writing—Review & editing.
Valerie P. Karuzis: Data curation; Formal analysis;
Investigation; Methodology; Project administration;
Software; Visualization; Writing—Original draft; Writing—
Review & editing. Nick B. Pandža: Data curation; Formal
analysis; Investigation; Methodology; Writing—Review &
editing. Polly O’Rourke: Conceptualization; Funding ac-
quisition; Methodology; Project administration;
Supervision; Writing—Review & editing. Stefanie E.
Kuchinsky: Conceptualization; Funding acquisition;
Methodology; Project administration; Supervision;
Writing—Review & editing.
Funding Information
Polly O'Rourke and Stephanie E. Kuchinsky, Defense
Advanced Research Projects Agency (https://dx.doi.org/10
.13039/100000185), grant number: N66001-17-2-4009.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed
a persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by estimated
gender identification of first author/last author) publish-
ing in the Journal of Cognitive Neuroscience ( JoCN )
during this period were M(an)/M = .408, W(oman)/M =
.335, M/ W = .108, and W/ W = .149, the comparable pro-
portions for the articles that these authorship teams cited
were M/M = .579, W/M = .243, M/ W = .102, and W/ W =
.076 (Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently,
JoCN encourages all authors to consider gender balance
explicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender citation
balance. The authors of this article report its proportions of
citations by gender category to be as follows: M/M = .462;
W/M = .262; M/ W = .108; W/ W = .169.
REFERENCES
Abrahamsson, N., & Hyltenstam, K. (2009). Age of onset and
nativelikeness in a second language: Listener perception
versus linguistic scrutiny. Language Learning, 59, 249–306.
https://doi.org/10.1111/j.1467-9922.2009.00507.x
Antoniou, M., & Wong, P. C. (2016). Varying irrelevant phonetic
features hinders learning of the feature being trained.
Journal of the Acoustical Society of America, 139, 271–278.
https://doi.org/10.1121/1.4939736, PubMed: 26827023
Aston-Jones, G., & Cohen, J. D. (2005). An integrative theory of
locus coeruleus-norepinephrine function: Adaptive gain and
optimal performance. Annual Review of Neuroscience, 28,
403–450. https://doi.org/10.1146/annurev.neuro.28.061604
.135709, PubMed: 16022602
Bakker, I., Takashima, A., van Hell, J. G., Janzen, G., &
McQueen, J. M. (2015). Tracking lexical consolidation with
ERPs: Lexical and semantic-priming effects on N400 and LPC
responses to newly-learned words. Neuropsychologia, 79,
33–41. https://doi.org/10.1016/j.neuropsychologia.2015.10
.020, PubMed: 26476370
Balass, M., Nelson, J. R., & Perfetti, C. A. (2010). Word learning:
An ERP investigation of word experience effects on
recognition and word processing. Contemporary
Educational Psychology, 35, 126–140. https://doi.org/10.1016
/j.cedpsych.2010.04.001, PubMed: 22399833
Barber, H. A., Otten, L. J., Kousta, S. T., & Vigliocco, G. (2013).
Concreteness in word processing: ERP and behavioral effects
in a lexical decision task. Brain and Language, 125, 47–53.
https://doi.org/10.1016/j.bandl.2013.01.005, PubMed:
23454073
Bent, T., Bradlow, A. R., & Wright, B. A. (2006). The influence of
linguistic experience on the cognitive processing of pitch in
speech and nonspeech sounds. Journal of Experimental
Psychology: Human Perception and Performance, 32,
97–103. https://doi.org/10.1037/0096-1523.32.1.97, PubMed:
16478329
Berridge, C. W., & Waterhouse, B. D. (2003). The locus
coeruleus–noradrenergic system: Modulation of behavioral
state and state-dependent cognitive processes. Brain
Research Reviews, 42, 33–84. https://doi.org/10.1016/S0165
-0173(03)00143-7, PubMed: 12668290
Borovsky, A., Elman, J. L., & Kutas, M. (2012). Once is enough:
N400 indexes semantic integration of novel word meanings
from a single exposure in context. Language Learning and
Development, 8, 278–302. https://doi.org/10.1080/15475441
.2011.614893, PubMed: 23125559
Bowles, A. R., Chang, C. B., & Karuzis, V. P. (2016). Pitch ability
as an aptitude for tone learning. Language Learning, 66,
774–808. https://doi.org/10.1111/lang.12159
Brysbaert, M., & New, B. (2009). Moving beyond Kučera and
Francis: A critical evaluation of current word frequency norms
and the introduction of a new and improved word frequency
measure for American English. Behavior Research Methods,
41, 977–990. https://doi.org/10.3758/BRM.41.4.977, PubMed:
19897807
Brysbaert, M., Warriner, A. B., & Kuperman, V. (2014). Concreteness
ratings for 40 thousand generally known English word lemmas.
Behavior Research Methods, 46, 904–911. https://doi.org/10
.3758/s13428-013-0403-5, PubMed: 24142837
Butt, M. F., Albusoda, A., Farmer, A. D., & Aziz, Q. (2020). The
anatomical basis for transcutaneous auricular vagus nerve
stimulation. Journal of Anatomy, 236, 588–611. https://doi
.org/10.1111/joa.13122, PubMed: 31742681
Phillips et al.
149
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Chandrasekaran, B., Sampath, P. D., & Wong, P. C. M. (2010).
Individual variability in cue-weighting and lexical tone
learning. Journal of the Acoustical Society of America,
128, 456–465. https://doi.org/10.1121/1.3445785, PubMed:
20649239
Clark, K. B., Naritoku, D. K., Smith, D. C., Browning, R. A., &
Jensen, R. A. (1999). Enhanced recognition memory
following vagus nerve stimulation in human subjects. Nature
Neuroscience, 2, 94–98. https://doi.org/10.1038/4600,
PubMed: 10195186
Cooper, A., & Wang, Y. (2012). The influence of linguistic and
musical experience on Cantonese word learning. Journal of
the Acoustical Society of America, 131, 4756–4769. https://
doi.org/10.1121/1.4714355, PubMed: 22712948
Cooper, A., & Wang, Y. (2013). Effects of tone training on
Cantonese tone–word learning. Journal of the Acoustical
Society of America, 134, EL133–EL139. https://doi.org/10
.1121/1.4812435, PubMed: 23927215
De Ridder, D., Vanneste, S., Engineer, N. D., & Kilgard, M. P.
(2014). Safety and efficacy of vagus nerve stimulation paired
with tones for the treatment of tinnitus: A case series.
Neuromodulation: Technology at the Neural Interface,
17, 170–179. https://doi.org/10.1111/ner.12127, PubMed:
24255953
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003
.10.009, PubMed: 15102499
Desbeaumes Jodoin, V., Lespérance, P., Nguyen, D. K.,
Fournier-Gosselin, M.-P., & Richer, F. (2015). Effects of vagus
nerve stimulation on pupillary function. International
Journal of Psychophysiology, 98, 455–459. https://doi.org/10
.1016/j.ijpsycho.2015.10.001, PubMed: 26437126
Díaz, B., Mitterer, H., Broersma, M., & Sebastián-Gallés, N.
(2012). Individual differences in late bilinguals’ L2
phonological processes: From acoustic-phonetic analysis
to lexical access. Learning and Individual Differences,
22, 680–689. https://doi.org/10.1016/j.lindif.2012.05.005
Dittinger, E., Barbaroux, M., D’Imperio, M., Jäncke, L., Elmer, S.,
& Besson, M. (2016). Professional music training and novel
word learning: From faster semantic encoding to longer-
lasting word representations. Journal of Cognitive
Neuroscience, 28, 1584–1602. https://doi.org/10.1162/jocn_a
_00997, PubMed: 27315272
Brain Stimulation: Basic, Translational, and Clinical
Research in Neuromodulation, 8, 624–636. https://doi.org
/10.1016/j.brs.2014.11.018, PubMed: 25573069
Granena, G., & Long, M. H. (2013). Age of onset, length of
residence, language aptitude, and ultimate L2 attainment in
three linguistic domains. Second Language Research, 29,
311–343. https://doi.org/10.1177/0267658312461497
Hao, Y.-C. (2012). Second language acquisition of Mandarin
Chinese tones by tonal and non-tonal language speakers.
Journal of Phonetics, 40, 269–279. https://doi.org/10.1016/j
.wocn.2011.11.001
Hassert, D. L., Miyashita, T., & Williams, C. L. (2004). The effects
of peripheral vagal nerve stimulation at a memory-
modulating intensity on norepinephrine output in the
basolateral amygdala. Behavioral Neuroscience, 118,
79–88. https://doi.org/10.1037/0735-7044.118.1.79, PubMed:
14979784
Hellman, A. B. (2011). Vocabulary size and depth of word
knowledge in adult-onset second language acquisition.
International Journal of Applied Linguistics, 21, 162–182.
https://doi.org/10.1111/j.1473-4192.2010.00265.x
Helmstaedter, C., Hoppe, C., & Elger, C. E. (2001). Memory
alterations during acute high-intensity vagus nerve
stimulation. Epilepsy Research, 47, 37–42. https://doi.org/10
.1016/S0920-1211(01)00291-1, PubMed: 11673019
Hulsey, D. R., Hays, S. A., Khodaparast, N., Ruiz, A., Das, P.,
Rennaker, R. L., et al. (2016). Reorganization of motor cortex
by vagus nerve stimulation requires cholinergic innervation.
Brain Stimulation, 9, 174–181. https://doi.org/10.1016/j.brs
.2015.12.007, PubMed: 26822960
Hulsey, D. R., Shedd, C. M., Sarker, S. F., Kilgard, M. P., & Hays,
S. A. (2019). Norepinephrine and serotonin are required
for vagus nerve stimulation directed cortical plasticity.
Experimental Neurology, 320, 112975. https://doi.org/10
.1016/j.expneurol.2019.112975, PubMed: 31181199
Ingvalson, E. M., Barr, A. M., & Wong, P. C. (2013). Poorer
phonetic perceivers show greater benefit in phonetic-
phonological speech learning. Journal of Speech, Language,
and Hearing Research, 56, 1045–1050. https://doi.org/10
.1044/1092-4388(2012/12-0024), PubMed: 23275405
Jacobs, H. I., Riphagen, J. M., Razat, C. M., Wiese, S., & Sack, A. T.
(2015). Transcutaneous vagus nerve stimulation boosts
associative memory in older individuals. Neurobiology of
Aging, 36, 1860–1867. https://doi.org/10.1016/j.neurobiolaging
.2015.02.023, PubMed: 25805212
Elgort, I., Perfetti, C. A., Rickles, B., & Stafura, J. Z. (2015).
Klinkenberg, I., Sambeth, A., & Blokland, A. (2011).
Contextual learning of L2 word meanings: Second language
proficiency modulates behavioural and event-related brain
potential (ERP) indicators of learning. Language, Cognition
and Neuroscience, 30, 506–528. https://doi.org/10.1080
/23273798.2014.942673, PubMed: 25984550
Engineer, C. T., Engineer, N. D., Riley, J. R., Seale, J. D., &
Kilgard, M. P. (2015). Pairing speech sounds with vagus nerve
stimulation drives stimulus-specific cortical plasticity. Brain
Stimulation, 8, 637–644. https://doi.org/10.1016/j.brs.2015.01
.408, PubMed: 25732785
Engineer, N. D., Riley, J. R., Seale, J. D., Vrana, W. A.,
Shetake, J. A., Sudanagunta, S. P., et al. (2011). Reversing
pathological neural activity using targeted plasticity. Nature,
470, 101–104. https://doi.org/10.1038/nature09656, PubMed:
21228773
Fox, J., & Weisberg, S. (2018). Visualizing fit and lack of fit in
complex regression models with predictor effect plots and
partial residuals. Journal of Statistical Software, 87, 1–27.
https://doi.org/10.18637/jss.v087.i09
Frangos, E., Ellrich, J., & Komisaruk, B. R. (2015). Non-invasive
access to the vagus nerve central projections via electrical
stimulation of the external ear: fMRI evidence in humans.
Acetylcholine and attention. Behavioural Brain Research,
221, 430–442. https://doi.org/10.1016/j.bbr.2010.11.033,
PubMed: 21108972
Kraus, T., Kiess, O., Hösl, K., Terekhin, P., Kornhuber, J., &
Forster, C. (2013). CNS BOLD fMRI effects of sham-
controlled transcutaneous electrical nerve stimulation in the
left outer auditory canal—A pilot study. Brain Stimulation, 6,
798–804. https://doi.org/10.1016/j.brs.2013.01.011, PubMed:
23453934
Kuhl, P. K. (2004). Early language acquisition: Cracking the
speech code. Nature Reviews Neuroscience, 5, 831–843.
https://doi.org/10.1038/nrn1533, PubMed: 15496861
Kuhl, P. K. (2010). Brain mechanisms in early language
acquisition. Neuron, 67, 713–727. https://doi.org/10.1016/j
.neuron.2010.08.038, PubMed: 20826304
Kuipers, J. R., & Thierry, G. (2011). N400 amplitude reduction
correlates with an increase in pupil size. Frontiers in Human
Neuroscience, 5, 61. https://doi.org/10.3389/fnhum.2011
.00061, PubMed: 21747766
Kutas, M., & Federmeier, K. D. (2011). Thirty years and
counting: Finding meaning in the N400 component of the
event-related brain potential (ERP). Annual Review of
150
Journal of Cognitive Neuroscience
Volume 34, Number 1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Psychology, 62, 621–647. https://doi.org/10.1146/annurev
.psych.093008.131123, PubMed: 20809790
Kutas, M., & Hillyard, S. A. (1980). Reading senseless sentences:
Brain potentials reflect semantic incongruity. Science, 207,
203–205. https://doi.org/10.1126/science.7350657, PubMed:
7350657
Lau, E. F., Holcomb, P. J., & Kuperberg, G. R. (2013).
Dissociating N400 effects of prediction from association in
single word contexts. Journal of Cognitive Neuroscience, 25,
484–502. https://doi.org/10.1162/jocn_a_00328, PubMed:
23163410
Lau, E. F., Phillips, C., & Poeppel, D. (2008). A cortical network
for semantics: (De)constructing the N400. Nature Reviews
Neuroscience, 9, 920–933. https://doi.org/10.1038/nrn2532,
PubMed: 19020511
Ling, W., & Grüter, T. (2020). From sounds to words: The
relation between phonological and lexical processing of tone
in L2 Mandarin. Second Language Research. https://doi.org
/10.1177/0267658320941546
Llanos, F., McHaney, J. R., Schuerman, W. L., Yi, H. G., Leonard,
M. K., & Chandrasekaran, B. (2020). Non-invasive
peripheral nerve stimulation selectively enhances speech
category learning in adults. NPJ Science of Learning, 5, 1–11.
https://doi.org/10.1038/s41539-020-0070-0, PubMed:
32802406
Long, M. H. (1990). Maturational constraints on language
development. Studies in Second Language Acquisition, 12,
251–285. https://doi.org/10.1017/S0272263100009165
Lopez-Calderon, J., & Luck, S. J. (2014). ERPLAB: An open-
source toolbox for the analysis of event-related potentials.
Frontiers in Human Neuroscience, 8, 213. https://doi.org/10
.3389/fnhum.2014.00213, PubMed: 24782741
Luck, S. J. (2014). An introduction to the event-related
potential technique (2nd ed.). Cambridge, MA: MIT Press.
Luu, P., & Ferree, T. (2005). Determination of the HydroCel
Geodesic Sensor Nets’ average electrode positions and their
10–10 International equivalents (p. 11) [Technical Note].
Electrical Geodesics, Inc. https://www.egi.com/images
/HydroCelGSN_10-10.pdf.
Mattock, K., Molnar, M., Polka, L., & Burnham, D. (2008). The
developmental course of lexical tone perception in the first
year of life. Cognition, 106, 1367–1381. https://doi.org/10
.1016/j.cognition.2007.07.002, PubMed: 17707789
McLaughlin, J., Osterhout, L., & Kim, A. (2004). Neural
correlates of second-language word learning: Minimal
instruction produces rapid change. Nature Neuroscience, 7,
703–704. https://doi.org/10.1038/nn1264, PubMed: 15195094
Mestres-Missé, A., Rodriguez-Fornells, A., & Münte, T. F. (2007).
Watching the brain during meaning acquisition. Cerebral
Cortex, 17, 1858–1866. https://doi.org/10.1093/cercor/bhl094,
PubMed: 17056648
Nieuwland, M. S., Barr, D. J., Bartolozzi, F., Busch-Moreno, S.,
Darley, E., Donaldson, D. I., et al. (2020). Dissociable effects
of prediction and integration during language
comprehension: Evidence from a large-scale study using
brain potentials. Philosophical Transactions of the Royal
Society of London, Series B, Biological Sciences, 375,
20180522. https://doi.org/10.1098/rstb.2018.0522, PubMed:
31840593
Ollen, J. E. (2006). A criterion-related validity test of selected
indicators of musical sophistication using expert ratings
(Doctoral dissertation). Retrieved from https://www.ohiolink
.edu/etd/view.cgi?osu1161705351.
Pandža, N. B., Phillips, I., Karuzis, V. P., O’Rourke, P., &
Kuchinsky, S. E. (2020). Neurostimulation and pupillometry:
New directions for learning and research in applied
linguistics. Annual Review of Applied Linguistics, 40, 56–77.
https://doi.org/10.1017/S0267190520000069
Pelzl, E. (2019). What makes second language perception of
Mandarin tones hard? Chinese as a second language. Journal
of the Chinese Language Teachers Association, 54, 51–78.
https://doi.org/10.1075/csl.18009.pel
Pelzl, E., Lau, E. F., Guo, T., & DeKeyser, R. (2021). Even in the
best-case scenario L2 learners have persistent difficulty
perceiving and utilizing tones in Mandarin: Findings from
behavioral and event-related potentials experiments. Studies
in Second Language Acquisition, 43, 268–296. https://doi.org
/10.1017/S027226312000039X
Perfetti, C. A., Wlotko, E. W., & Hart, L. A. (2005). Word learning
and individual differences in word learning reflected in
event-related potentials. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 31,
1281–1292. https://doi.org/10.1037/0278-7393.31.6.1281,
PubMed: 16393047
Perrachione, T. K., Lee, J., Ha, L. Y. Y., & Wong, P. C. M. (2011).
Learning a novel phonological contrast depends on interactions
between individual differences and training paradigm design.
Journal of the Acoustical Society of America, 130, 461–472.
https://doi.org/10.1121/1.3593366, PubMed: 21786912
Poltrock, S., Chen, H., Kwok, C., Cheung, H., & Nazzi, T. (2018).
Adult learning of novel words in a non-native language:
Consonants, vowels, and tones. Frontiers in Psychology,
9, 1211. https://doi.org/10.3389/fpsyg.2018.01211, PubMed:
30087631
Psychology Software Tools, Inc. (2012). E-Prime ( Version 2.0).
[Computer software]. https://www.pstnet.com
Pu, H., Holcomb, P. J., & Midgley, K. J. (2016). Neural changes
underlying early stages of L2 vocabulary acquisition. Journal
of Neurolinguistics, 40, 55–65. https://doi.org/10.1016/j
.jneuroling.2016.05.002, PubMed: 28983152
R Core Team. (2020). R: A language and environment for
statistical computing. R Foundation for Statistical
Computing ( Version 3.6.3). R Foundation for Statistical
Computing. https://www.R-project.org/
Rodríguez-Fornells, A., Cunillera, T., Mestres-Missé, A., & de
Diego-Balaguer, R. (2009). Neurophysiological mechanisms
involved in language learning in adults. Philosophical
Transactions of the Royal Society of London, Series B:
Biological Sciences, 364. https://doi.org/10.1098/rstb.2009
.0130, PubMed: 19933142
Rodríguez-Fornells, A., De Diego Balaguer, R., & Münte, T. F.
(2006). Executive control in bilingual language processing.
Language Learning, 56, 133–190. https://doi.org/10.1111/j
.1467-9922.2006.00359.x
Sarter, M., Gehring, W. J., & Kozak, R. (2006). More attention
must be paid: The neurobiology of attentional effort. Brain
Research Reviews, 51, 145–160. https://doi.org/10.1016/j
.brainresrev.2005.11.002, PubMed: 16530842
Sebastián-Gallés, N., Echeverría, S., & Bosch, L. (2005). The
influence of initial exposure on lexical representation:
Comparing early and simultaneous bilinguals. Journal of
Memory and Language, 52, 240–255. https://doi.org/10.1016
/j.jml.2004.11.001
Shen, X. S., & Lin, M. (1991). A perceptual study of Mandarin
tones 2 and 3. Language & Speech, 34, 145–156. https://doi
.org/10.1177/002383099103400202
Sun, L., Peräkylä, J., Holm, K., Haapasalo, J., Lehtimäki, K.,
Ogawa, K. H., et al. (2017). Vagus nerve stimulation improves
working memory performance. Journal of Clinical and
Experimental Neuropsychology, 39, 954–964. https://doi.org
/10.1080/13803395.2017.1285869, PubMed: 28492363
Thakkar, V. J., Engelhart, A. S., Khodaparast, N., Abadzi, H., &
Centanni, T. M. (2020). Transcutaneous auricular vagus nerve
stimulation enhances learning of novel letter–sound
relationships in adults. Brain Stimulation, 13, 1813–1820.
https://doi.org/10.1016/j.brs.2020.10.012, PubMed: 33127581
Phillips et al.
151
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Van Petten, C., & Luka, B. J. (2012). Prediction during language
comprehension: Benefits, costs, and ERP components.
International Journal of Psychophysiology, 83, 176–190.
https://doi.org/10.1016/j.ijpsycho.2011.09.015, PubMed:
22019481
Vanderwal, T., Kelly, C., Eilbott, J., Mayes, L. C., & Castellanos,
F. X. (2015). Inscapes: A movie paradigm to improve
compliance in functional magnetic resonance imaging.
Neuroimage, 122, 222–232. https://doi.org/10.1016/j
.neuroimage.2015.07.069, PubMed: 26241683
Voeten, C. C. (2020). buildmer: Stepwise elimination and term
reordering for mixed-effects regression ( Version 1.5). https://
CRAN.R-project.org/package=buildmer
Vonck, K., Raedt, R., Naulaerts, J., De Vogelaere, F., Thiery, E.,
Van Roost, D., et al. (2014). Vagus nerve stimulation…25
years later! What do we know about the effects on cognition?
Neuroscience & Biobehavioral Reviews, 45, 63–71. https://
doi.org/10.1016/j.neubiorev.2014.05.005, PubMed: 24858008
Wang, Y., Spence, M. M., Jongman, A., & Sereno, J. A. (1999).
Training American listeners to perceive Mandarin tones.
Journal of the Acoustical Society of America, 106, 3649–3658.
https://doi.org/10.1121/1.428217, PubMed: 10615703
Werker, J. F., & Hensch, T. K. (2015). Critical periods in speech
perception: New directions. Annual Review of Psychology,
66, 173–196. https://doi.org/10.1146/annurev-psych-010814
-015104, PubMed: 25251488
White, E. J., Hutka, S. A., Williams, L. J., & Moreno, S. (2013).
Learning, neural plasticity and sensitive periods: Implications
for language acquisition, music training and transfer
across the lifespan. Frontiers in Systems Neuroscience,
7, 90. https://doi.org/10.3389/fnsys.2013.00090, PubMed:
24312022
Wong, P. C., & Perrachione, T. K. (2007). Learning pitch
patterns in lexical identification by native English-speaking
adults. Applied PsychoLinguistics, 28, 565–585. https://doi
.org/10.1017/S0142716407070312
Yakunina, N., Kim, S. S., & Nam, E.-C. (2017). Optimization of
transcutaneous vagus nerve stimulation using functional MRI.
Neuromodulation: Technology at the Neural Interface,
20, 290–300. https://doi.org/10.1111/ner.12541, PubMed:
27898202
Yip, M. (2002). Tone. Cambridge University Press. https://doi
.org/10.1017/CBO9781139164559
Zhang, J. D., & Schubert, E. (2019). A single item measure
for identifying musician and nonmusician categories
based on measures of musical sophistication. Music
Perception, 36, 457–467. https://doi.org/10.1525/mp.2019
.36.5.457
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
1
1
2
7
2
0
0
7
7
0
6
/
j
o
c
n
_
a
_
0
1
7
8
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
152
Journal of Cognitive Neuroscience
Volume 34, Number 1