Transcranial Direct Current Stimulation Modulates
Connectivity of Left Dorsolateral Prefrontal
Cortex with Distributed Cortical Networks
Kamin Kim1, Matthew S. Sherwood2, Lindsey K. McIntire3,
R. Andy McKinley4, and Charan Ranganath1
Abstract
■ Studies have shown that transcranial direct current stimula-
tion increases neuronal excitability of the targeted region and
general connectivity of relevant functional networks. However,
relatively little is understood of how the stimulation affects the
connectivity relationship of the target with regions across the
network structure of the brain. Here, we investigated the effects
of transcranial direct current stimulation on the functional
connectivity of the targeted region using resting-state fMRI
scans of the human brain. Anodal direct current stimulation
was applied to the left dorsolateral prefrontal cortex (lDLPFC;
cathode on the right bicep), which belongs to the frontoparietal
control network (FPCN) and is commonly targeted for neuro-
modulation of various cognitive functions including short-term
memory, long-term memory, and cognitive control. lDLPFC’s
connectivity characteristics were quantified as graph theory
measures, from the resting-state fMRI scans obtained prior to
and following the stimulation. Critically, we tested pre- to post-
stimulation changes of the lDLPFC connectivity metrics following
an active versus sham stimulation. We found that the stimulation
had two distinct effects on the connectivity of lDLPFC: for
Brodmann’s area (BA) 9, it increased the functional connectivity
between BA 9 and other nodes within the FPCN; for BA 46, net
connectivity strength was not altered within FPCN, but connec-
tivity distribution across networks (participation coefficient) was
decreased. These findings provide insights that the behavioral
changes as the functional consequences of stimulation may
come about because of the increased role of lDLPFC in the
FPCN. ■
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
Transcranial direct current stimulation (tDCS) is widely used
as a neuromodulation tool for clinical and research purposes
(for reviews, see Dedoncker, Baeken, De Raedt, &
Vanderhasselt, 2021; Filmer, Mattingley, & Dux, 2020; Razza
et al., 2020; Galli, Vadillo, Sirota, Feurra, & Medvedeva, 2019;
Osório & Brunoni, 2019; Bennabi & Haffen, 2018; Lucchiari,
Sala, & Vanutelli, 2018; Strobach & Antonenko, 2017;
Dedoncker, Brunoni, Baeken, & Vanderhasselt, 2016a,
2016b; Jantz, Katz, & Reuter-Lorenz, 2016; Kim, Ekstrom, &
Tandon, 2016; Nitsche & Paulus, 2011; Paulus, 2011;
Nitsche et al., 2008; Wagner, Valero-Cabre, & Pascual-Leone,
2007; Gandiga, Hummel, & Cohen, 2006). For example, sev-
eral tDCS studies have targeted left dorsolateral prefrontal
cortex (lDLPFC) to modulate long-term memory (Mizrak
et al., 2018; Leshikar et al., 2017; Sandrini et al., 2014, 2016;
Javadi, Cheng, & Walsh, 2012; Javadi & Walsh, 2012;
Zwissler et al., 2014), working memory (Talsma,
Broekhuizen, Huisman, & Slagter, 2018; Talsma, Kroese, &
Slagter, 2017; Hill, Fitzgerald, & Hoy, 2016; Mancuso, Ilieva,
Hamilton, & Farah, 2016; Trumbo et al., 2016; Carvalho
et al., 2015; Andrews, Hoy, Enticott, Daskalakis, &
1University of California, Davis, 2Science and Space, KBR Inc.,
Beavercreek, OH, 3Infoscitex, Beavercreek, OH, 4U.S. Air
Force, Dayton, OH
© 2021 Massachusetts Institute of Technology
Fitzgerald, 2011; Keeser et al., 2011; Fregni et al., 2005;
Marshall, Mölle, Siebner, & Born, 2005), cognitive training
effects (Au et al., 2016; Martin, Liu, Alonzo, Green, & Loo,
2014), and attention and cognitive control (London &
Slagter, 2021; McIntire, McKinley, Nelson, & Goodyear,
2017; Nejati, Salehinejad, Nitsche, Najian, & Javadi, 2020;
Nelson, McKinley, Golob, Warm, & Parasuraman, 2014;
McKinley et al., 2013; Gladwin, den Uyl, Fregni, & Wiers,
2012). Prefrontal cortex is thought to contribute to these
functions by using information about behavioral goals to
modulate activity in posterior cortical areas (Miller &
Cohen, 2001; Tomita, Ohbayashi, Nakahara, Hasegawa, &
Miyashita, 1999). Accordingly, to the extent that stimulating
lDLPFC produces behavioral effects, we would expect that
stimulation should modulate potential interactions between
lDLPFC and other cortical areas. However, our under-
standing is still sparse on how the tDCS might influence
the interregional dynamics of lDLPFC.
We investigated whether tDCS applied to lDLPFC can
modulate network-level functional connectivity between
lDLPFC and other regions of the brain. We used the tem-
poral correlation of BOLD signal fluctuations as the mea-
sure of functional connectivity between brain regions (i.e.,
resting-state connectivity; Cole, Ito, Bassett, & Schultz,
2016; Cole, Bassett, Power, Braver, & Petersen, 2014;
Vincent, Kahn, Snyder, Raichle, & Buckner, 2008; Fox &
Journal of Cognitive Neuroscience 33:7, pp. 1381–1395
https://doi.org/10.1162/jocn_a_01725
Raichle, 2007; Damoiseaux et al., 2006; De Luca,
Beckmann, De Stefano, Matthews, & Smith, 2006;
Vincent et al., 2006). The spatial structure of these corre-
lations suggests that the brain is organized in multiple
semimodular “small-world” networks characterized by
high within-network connections and sparse between-
network connections (Ashourvan, Telesford, Verstynen,
Vettel, & Bassett, 2019; Power et al., 2011; Bullmore &
Sporns, 2009; Kaiser, Martin, Andras, & Young, 2007;
Sporns & Zwi, 2004), and we aim to investigate how stim-
ulation modulates the target region’s connectivity within
the network structure. Prior studies have shown that
tDCS applied to lDLPFC influences general connectivity
strength of the target or neighboring networks. In
Keeser et al. (2011), participants went through two exper-
imental sessions, one with active stimulation and the other
with sham stimulation, and each stimulation session was
preceded and followed by fMRI scanning. Authors showed
that the stimulation induced significant increases in
lDLPFC activation and significant changes of connectivity
within networks that included brain regions that are close
to lDLPFC. However, the connectivity changes specific to
lDLPFC or between lDLPFC and the networks were not di-
rectly tested in this study. In another study, connectivity
analyses using lDLPFC as the seed region showed that
the stimulation increased connectivity between lDLPFC
and bilateral parietal regions inferior parietal lobule
[IPL], superior parietal lobule [SPL]); however, this study
focused on pairwise functional connectivity relationships
and did not take the network structure into consideration
(Mondino et al., 2020).
We investigated how lDLPFC stimulation modulates the
way in which the target region interacts with other parts of
the brain network structure. Given that lDLPFC has been
identified as a node in the “frontoparietal control network
(FPCN)” (Schaefer et al., 2018), we considered the possi-
bility that tDCS might increase the relative importance of
lDLPFC within the FPCN compared to its importance
across other networks. An alternative possibility is that
stimulation might increase the breadth of lDLPFC connec-
tivity across different networks beyond FPCN. To test
these predictions, we conducted a combined stimulation
and imaging experiment, in which different groups of par-
ticipants underwent an active or sham stimulation.
Functional connectivity between regions was estimated
using resting-state fMRI scans acquired prior to and follow-
ing the stimulation, and used to compute graph theory
metrics that characterize a node’s connectivity in the brain
network structure. Finally, we examined how stimulation
altered the network relationships of lDLPFC.
METHODS
Participants
A total of 60 healthy participants completed this study
(Day 1 study session; see Procedure section for details).
Participants were active duty Air Force military members
recruited from Wright-Patterson Air Force Base. Inclusion
criteria for this study included the absence of neurological
or psychological disorders, head injury (concussion his-
tory, traumatic brain injury), recent trauma or hospitaliza-
tion, impairment of vision, hearing or motor control,
dependency on alcohol, caffeine, or nicotine, and any
current medication that may affect cognitive functions.
Of the 60 participants who completed the session, nine
participants were excluded from analysis because of ex-
cessive motion during fMRI scanning (> 3 mm) and a to-
tal number of 51 participants were included in the final
analyses (13 female, 38 male). Participants were randomly
assigned to the sham, 1 mA, or 2 mA active stimulation
group (Nsham = 22, Nactive = 29 [N1mA = 13, N2mA =
16]). All data were collected at Dayton Children’s
Hospital, and all experimental procedures were approved
by the Air Force Research Laboratory Institutional Review
Board at Wright-Patterson Air Force Base and were fully
described to the participants before they consented to
participate in the study.
Procedure
This study was conducted as part of a larger multiday
testing study. This study focused on noncumulative,
immediate effects of stimulation using data from Day 1
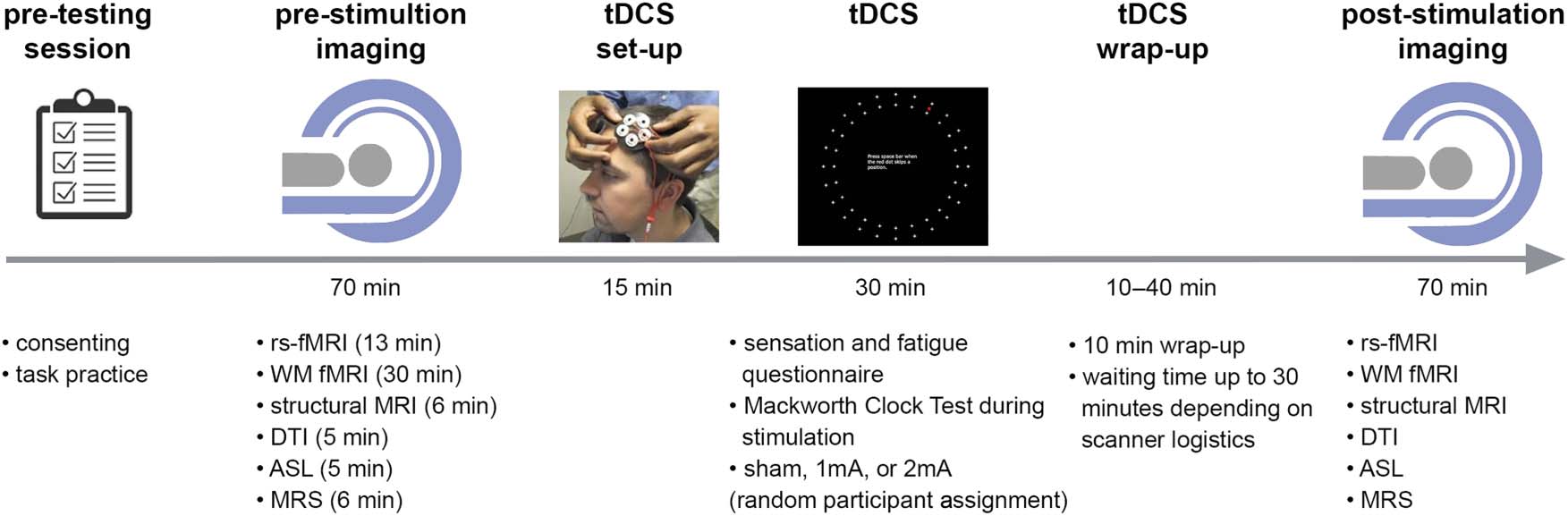
resting-state fMRI. Figure 1 depicts the procedure of the
pretesting and Day 1 sessions. One or 2 days prior to the
actual testing, participants came in for the consenting pro-
cedure and task practice. They were given a 5-min-long
practice training on the Mackworth Clock test (McKinley,
2018; McIntire et al., 2017) that was to be used during stim-
ulation. On each testing day, participants went through a
stimulation session and two MRI sessions: one preceding
and one following the stimulation session. An imaging ses-
sion included one block of resting-state fMRI, three blocks
of task fMRI (n-back task), structural MRI, diffusion tensor
imaging, magnetic resonance spectroscopy (MRS), and
arterial spin labeling (ASL). During resting-state fMRI scan-
ning, participants were instructed to keep their eyes open
and to focus on a fixation dot at the center of the screen.
Participants were taken out from the scanner after the first
(prestimulation) imaging session and guided into a behav-
ioral testing room for a stimulation session. A scanning
session was about 70 min long, and the average interval
between the start of the prestimulation and poststimula-
tion scanning sessions was 158 min. Poststimulation
resting-state fMRI was conducted within 1 hr from the
stimulation, and this interval is well within the duration
that our stimulation regime shows behavioral effects
for (McIntire, McKinley, Goodyear, & Nelson, 2014).
Nevertheless, one may argue that tDCS effects might wear
out before the scanning and mere epiphenomenal effects
of stimulation (i.e., thinking about the tingling sensation
during the stimulation session) could result in false-
positive findings. In order to address such possibility,
1382
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
Figure 1. Experiment procedure. tDCS setup figure is reused with permission (McIntire et al., 2014).1
we obtained participants’ rating on the arousal level (fa-
tigued vs. energized) and four types of sensation that are
often associated with stimulation (see under Stimulation
section below for details).
meant unbearable. The protocol was to turn off the stim-
ulation in the case one reports a sensation of 7 or higher
on any category, but there was no incident. Fatigue was
rated on a 7-point scale, with 1 feeling most fatigued
and 7 feeling energized.
Stimulation
Transcranial DC stimulation was delivered with a MagStim
DC stimulator (magstim.com, Whiteland). Instead of the
standard wet sponge electrodes, custom, composite elec-
trodes were used as anode and cathode (Sherwood,
Madaris, Mullenger, & McKinley, 2018; McIntire et al.,
2014, 2017; McKinley et al., 2013; Park, Hong, Kim, Suh,
& Im, 2011). As in the prior studies that used composite
electrodes, a composite electrode consisted of five
Na/NaCl EEG electrodes that were arranged in a circular
pattern with an inner diameter of 1.6 cm. EEG electrodes
in the array were separated by 0.1 cm from the neighbor-
ing ones (measured from outer edge to outer edge). The
anode was placed on the lDLPFC (approximately F3 in
International 10–20 EEG system), and the cathode was
placed on the contralateral (right) bicep. Electrical field
modeling using the Finite Element Method has shown
that this approach delivers the current evenly distributed
among the five EEG electrodes and, when used with a 2 mA
current intensity, delivers stimulation with an estimated
current density of 0.199 mA/cm2 to the target area
(McIntire et al., 2017; McKinley et al., 2013; see Figure 2
in McKinley et al., 2013, for the Finite Element Method
model of estimated current distribution). Conductive gel
was applied on the electrodes to ensure current conduc-
tion, and electrodes were secured to the target area using
medical bandages. A constant current of 1 mA or 2 mA
was delivered for 30 min in the active stimulation group
and for 30 sec in the sham stimulation group. Electrodes
remained in place for the full 30 min in both groups.
One minute into the stimulation, participants completed
a sensation and fatigue questionnaire. On this question-
naire, participants rated the intensity of four types of sensa-
tion (heat, itching, pain, and discomfort) on a 10-point
scale, where 0 corresponded to feeling nothing and 10
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
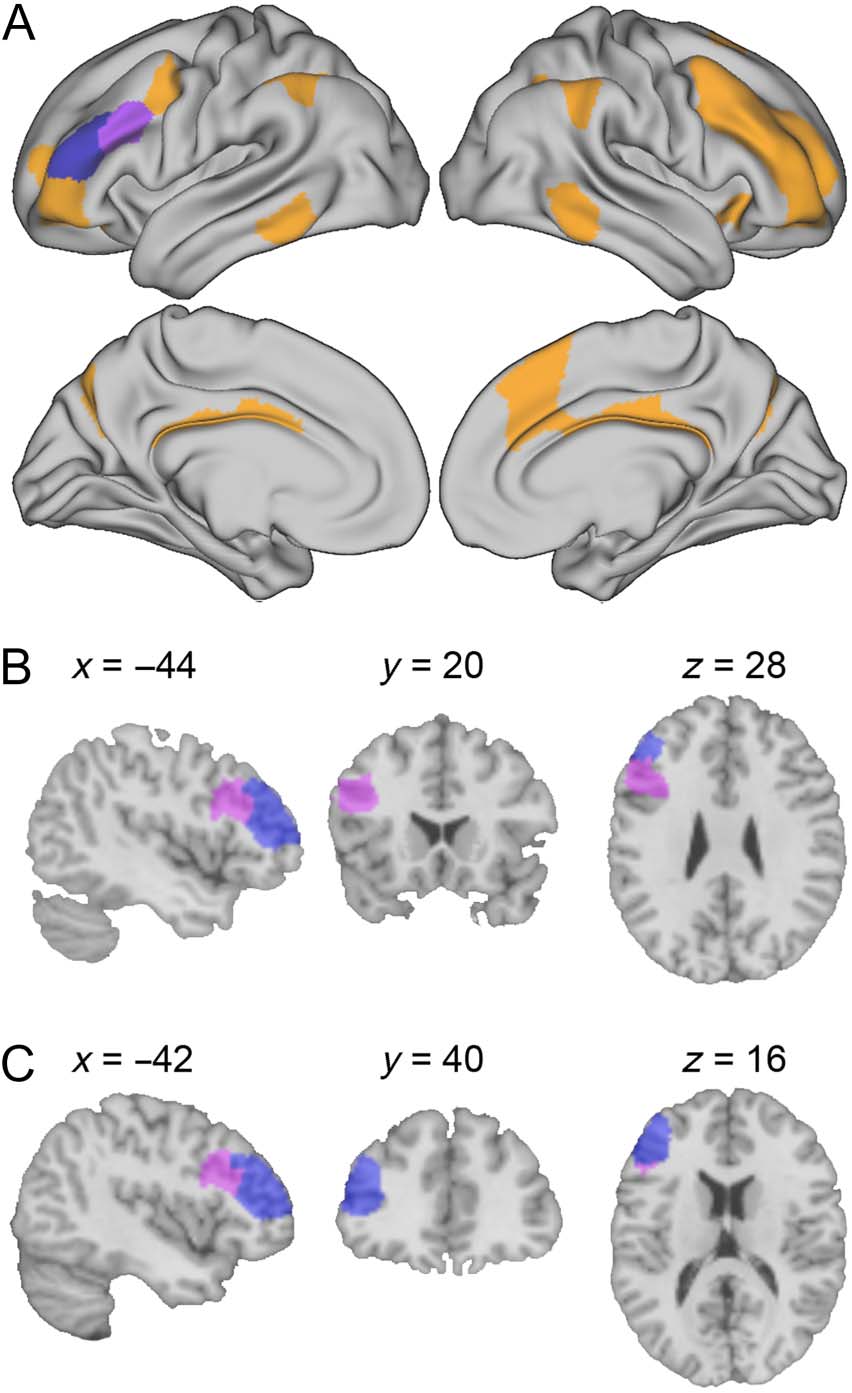
Figure 2. Stimulation target ROIs belong to the FPCN. (A) FPCN
is depicted in color: violet = parcels corresponding to BA 9
(LH_Cont_PFCl_4 in Schaefer atlas); blue = parcels corresponding to
BA 46 (labeled as LH_Cont_PFCl_3 the in Schaefer atlas); orange =
other parcels that belong to FPCN. (B–C) Left DLPFC ROIs are depicted
in volumetric views. Sections at the center of BA 9 (B, violet) and BA 46
(C, blue). Coordinates are in MNI space.
Kim et al.
1383
For the remaining duration of the 30-min stimulation,
participants completed a vigilance task (an adapted ver-
sion of Mackworth Clock Test; McIntire et al., 2017).
This was done as neuronal modulatory effects via electrical
stimulation are largely influenced by the neuronal state
before and during the stimulation (Silvanto, Muggleton,
& Walsh, 2008). Specifically for tDCS, prior research indi-
cated that anodal stimulation applied while the target
region is activated by task engagement has increased effec-
tiveness (Filmer, Lyons, Mattingley, & Dux, 2017; Ruf,
Fallgatter, & Plewnia, 2017; Andrews et al., 2011). Left
DLPFC, our stimulation target region, is highly engaged
in attentional/cognitive control functions such as vigilance
(Kim, Kim, & Im, 2017; Nelson et al., 2014; Langner &
Eickhoff, 2013) and working memory ( Jansma, Ramsey,
de Zwart, van Gelderen, & Duyn, 2007; Ranganath,
Johnson, & D’Esposito, 2003; D’Esposito, Postle, Ballard,
& Lease, 1999; D’Esposito et al., 1998; Barch et al., 1997;
Braver et al., 1997).
translation and three rotation parameters, and their first-
order derivatives), BOLD time series from subject-specific
white matter (top 5 PCA parameters) and CSF (top 5 PCA
parameters), and a binary vector that marked outlier time
points identified by ART (www.nitrc.org/projects/artifact
_detect). ART outliers were defined as images with a
framewise displacement greater than 0.5 mm from the
previous image and images with a global mean intensity
more than 2 SDs away from the mean image intensity of
all scans. These thresholds are slightly more conservative
than the default thresholds in the CONN toolbox
(0.9 mm, 5 standard deviations), which are considered
to be “intermediate” thresholds (97th percentile). On av-
erage, approximately 17% of scans were outliers (ranges:
2%–53%). The resulting BOLD time series were band-pass
filtered (0.009 Hz < F < 0.08 Hz) and averaged within
each ROI. Fisher-transformed correlations between the
resulting filtered time series of ROI pairs were used as
the final estimates of connectivity between ROI pairs.
Resting-State fMRI Data Acquisition
MRI images were collected using a 3T GE Discovery
scanner with a 24-channel head/neck coil at Dayton
Children’s Medical Center. T1-weighted structural images
were acquired using a spoiled gradient recalled acquisi-
tion sequence (flip angle = 16°, FoV = 25.6 cm, image
dimension: 256 × 256 × 164, voxel size = 1 × 1 ×
1 mm). Resting-state functional images were acquired using
gradient EPI sequences (TR = 2000 msec, TE = 20 msec,
flip angle = 90°, FoV = 24 cm, image dimension = 64 ×
64 × 41, slice thickness = 3 mm, voxel size = 3.75 × 3.75 ×
3.5 mm, top–down interleaved). The functional image
acquisition included four initial dummy scans to ensure
signal stabilization, and the scanning lasted 12 min and
14 sec. Dummy scans were discarded, and a total number
of 363 volumes were further processed for analyses.
Resting-State fMRI Data Processing
Preprocessing and data analyses were performed using
SPM12 (www.fil.ion.ucl.ac.uk/spm) and the CONN tool-
box ( Whitfield-Gabrieli & Nieto-Castanon, 2012).
Functional images were slice-time corrected for differ-
ences in slice acquisition times, realigned, normalized,
spatially smoothed (FWHM = 8 mm), and segmented
using a local-global parcellation atlas (Schaefer et al.,
2018; the atlas provides parcellations of 10 different levels
of granularity ranging from 100 to 1000 parcellations,
and we used the atlas with 200 parcels).
Data were denoised using the component-based noise
correction (CompCor; Behzadi, Restom, Liau, & Liu,
2007) method and scrubbing as implemented in the
CONN toolbox. Specifically, the following time series
were used as temporal covariates in the first-level analysis
and removed from the BOLD functional data using linear
regression: 12 time series of the estimated motion (three
Graph Measures
Graph theory measures that characterize a node’s connec-
tivity strength (node degree) and connectivity distribution
(participation coefficient) were computed using the
resting-state connectivity measures and an independent,
predefined network structure. For valid testing of the stim-
ulation effects on graph measures, it was crucial to obtain
graph measures based on a common, objective network
structure across individuals and scanning sessions. To
achieve this, the following steps were taken: 1) We
acquired stable solutions of modular/network structures
from our resting-state connectivity data using a consensus
clustering approach (see below for details), and 2) we com-
puted graph measures by assessing the outcome connec-
tions against a network structure of our ROIs that was
predefined using resting-state functional connectivity from
a large sample (n = 1489; Schaefer et al., 2018). The
Schaeffer atlas assigns ROIs into networks of two different
granularity—seven and 17 networks—and we used the
seven-network solution that includes visual, somato-motor,
dorsal attention, ventral attention, limbic, frontoparietal
control, and default networks.
The functional connectivity network structure from
each individual and session was obtained using a consen-
sus clustering approach (Lancichinetti & Fortunato,
2012), which allows for obtaining stable community solu-
tions when the community detection method is not deter-
ministic. A consensus matrix was obtained as following.
First, an adjacency matrix of ROI-by-ROI connectivity
strength (Fisher’s Z ) was obtained for each participant
and session (see Resting-state fMRI Data Processing
section). The adjacency matrices were entered into a
clustering algorithm (community_louvain) implemented
in BrainConnectivityToolbox (sites.google.com/site
/bctnent; Rubinov & Sporns, 2011), and this procedure
was repeated 1000 times. For each ROI pair, the ratio that
1384
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
the two ROIs were partitioned into the same community
out of the 1000 solutions (i.e., “consensus” rate) was com-
puted, and the consensus rates were thresholded at 0.035
(consensus rates smaller than the threshold were set to
zero). The outcome ROI-by-ROI matrix of thresholded
consensus rate was used as an input to the next iteration.
This procedure (clustering an ROI-by-ROI matrix 1000
times, obtaining a consensus matrix, and thresholding
the consensus matrix) was repeated iteratively until the
resulting matrix reached a complete consensus (all ROI
pairs would have a consensus rate of 0 or 1). We consid-
ered an “edge” (i.e., connection) to exist between ROIs if
the consensus rate was 1 between the pair of ROIs.
Using the predefined network memberships of ROIs and
the consensus matrix solutions that are described above,
we estimated graph measures that quantify a node’s con-
nectivity strength with given networks (node degree) and
how well-distributed across different networks a node’s
connectivity is (participation coefficient). Connectivity
strength of an ROI (i.e., a network node), node degree,
was estimated as total number of the node’s edges divided
by the number of all possible edges. For example,
Brodmann’s area (BA) 9’s node degree within FPCN was
computed as the number of edges between BA 9 and other
nodes within the FPCN divided by the number of possible
edges between BA 9 and all FPCN nodes (i.e., the denom-
inator is N-1 in a network with N nodes). Participation co-
efficient was computed using the participation_coef_sign
function in BrainConnectivityToolbox.
Statistical Analysis
Statistical analyses were conducted using R statistical tool-
box 3.4.3. (www.r-project.org/) and custom MATLAB
(www.mathworks.com) codes. One-way between-group
ANOVA was used to test whether participant groups
(sham vs. active stimulation) differed in their age, scan-
ning time intervals, or the sensation and fatigue ratings.
A general linear mixed-effects model (GLMM) and
permutation-based nonparametric tests were used to test
for stimulation effects on node degree and participation
coefficient of targets. Graph theory measures (i.e., node
degree) were entered as the predicted variable, and stim-
ulation group (sham/active), scanning session (pre-/poststim-
ulation), and a Group × Session interaction were entered
as predictor variables, with the subject variable included
in the model as a random intercept. To determine
whether the variability of graph measures is explained
by the stimulation, the statistical significance of the inter-
action term (Sham/Active × Pre-/Poststimulation interac-
tion) was critically tested using permutation tests. A
permutation distribution of interaction coefficients was
acquired by running the regression model 1000 times
after shuffling group membership (active/sham) and
session (pre-/poststimulation) labels. Specifically, each
participant was randomly reassigned to one of the groups
(sham/active) while keeping the group size the same
(Nsham = 22, Nactive = 29). Session labels (pre-/poststimula-
tion) were flipped in randomly selected participants.
Shuffling variable labels in this manner guaranteed that
the within-subject variability was kept intact. The interac-
tion coefficient obtained from unshuffled, real data was
tested against the permutation distribution. The sign of
the interaction term is irrelevant to the interpretation of
results here; therefore, coefficients smaller than the 5th
or greater than the 95th percentile of the distribution
were considered statistically significant.
RESULTS
Demographic, Scanning, and Poststimulation
Questionnaire Data
Stimulation groups were comparable in age and the inter-
vals between the scanning sessions prior to and following
the stimulation (Table 1; all p > .1). On average, both
groups reported ratings below 2 (from 0 being nothing
and 10 being unbearable) on all four types of sensation,
and importantly, the rating did not differ between groups
for any type (all p > .1). In addition to sensation, partici-
pants also reported how fatigued or energized they felt on
a 7-point scale (with 1 being most fatigued and 7 being
energized ). The fatigue level that the sham and active
stimulation groups reported were identical (sham group =
4.09, active group = 4.10; p > .1). Therefore, it can be
assured that effects of the stimulation were not driven
by participants’ awareness of the stimulation condition.
When the active stimulation group was further broken
into 1- and 2-mA stimulation intensity groups, the three
groups (sham, active-1 mA, active-2 mA) did not differ in
any metrics reported here (Table 2).
ROIs and Functional Categories of the lDLPFC
Among the 200 parcels defined in the Schaefer atlas, ones
that were centered in BA 9 or BA 46 (MacDonald, Cohen,
Stenger, & Carter, 2000) were identified as lDLPFC. This
approach identified two neighboring parcels in the left
prefrontal cortex—a posterior dorsal one corresponding
to BA 9 (violet in Figure 2; Figure 2B for volumetric views)
and an anterior ventral one mapping to BA 46 (blue in
Figure 2; Figure 2C for volumetric views). Both of these
ROIs have been identified as nodes in the FPCN
(Schaefer et al., 2018; Figure 2A). In addition, to test the
specificity of stimulation effects, we used the rDLPFC as a
control region. Three parcels were identified as rDLPFC
using the same approach as for the lDLPFC described
above, and all three nodes belonged to the FPCN.
Active Stimulation Increased Connectivity of BA 9
within FPCN
We first examined how the stimulation influenced the
within-FPCN connectivity strength of lDLPFC ROIs. To
Kim et al.
1385
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Comparisons of Sham and Active Simulation Groups
Stimulation Group
Group Difference
demographic
scanning time
sensation
fatigue
sex
age
interval
itching
pain
heat
discomfort
Sham (n = 22)
Active (n = 29)
Female = 5
28.1 (5.22)
Female = 8
28.2 (5.85)
159.1 (16.86)
158.9 (15.54)
1.73 (1.70)
0.36 (0.85)
0.23 (0.43)
1.05 (1.43)
4.09 (1.02)
1.38 (1.21)
0.59 (0.91)
0.55 (0.91)
0.69 (0.97)
4.10 (1.35)
F
NA
0.009
0.003
0.733
0.796
2.387
1.112
0.001
p
NA
.92
.96
.40
.38
.13
.30
.97
Demographics, time interval between scanning sessions prior to and following the stimulation, and stimulation questionnaire scores in participant
groups that received sham and active stimulation. The sham and active stimulation group did not differ in age, scanning time, and sensation reports.
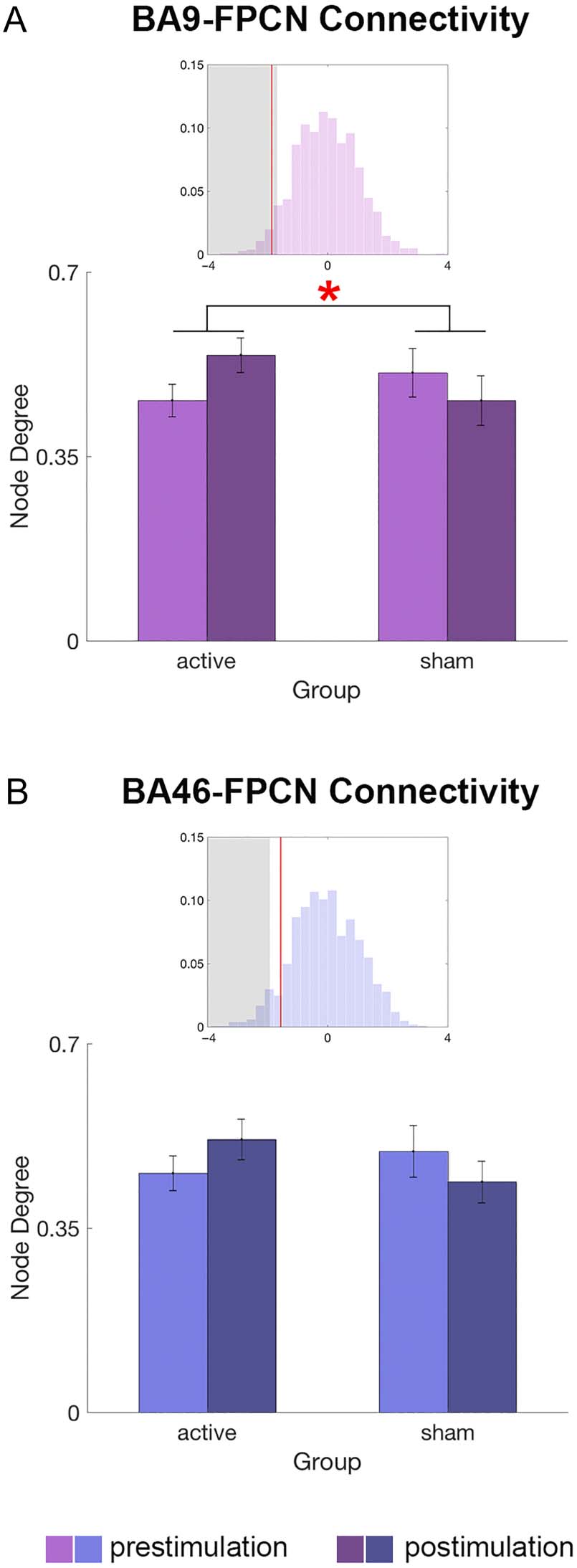
test this, GLMM was conducted on lDLPFC nodes’ within-
FPCN connectivity strength with the stimulation group
(active/sham), scanning session (pre-/poststimulation),
and an interaction term as predictor variables (see
Statistical Analysis section for details of the regression
model). GLMM permutation tests revealed that within-
FPCN connectivity strength was significantly increased
by stimulation for BA 9 (|z| = 1.90; Figure 3A), but not
for BA 46 (|z| = 1.39; Figure 3B) or any of the rDLPFC
control ROIs (all |z| < 0.7). We additionally investigated
whether the BA 9-FPCN connectivity change was stimula-
tion intensity-dependent. Specifically, we tested the
effects of the stimulation intensity (Intensity × Session
interaction) on BA 9-FPCN connectivity using GLMM per-
mutation tests. The analysis revealed no significant differ-
ence in the stimulation-driven BA 9-FPCN connectivity
changes between the 1- and 2-mA stimulation groups
(interaction term |z| = 0.11).
Connectivity between lDLPFC and Regions Outside
FPCN: Stimulation May Bias BA 46 Connectivity to
Be Less Distributed across Networks
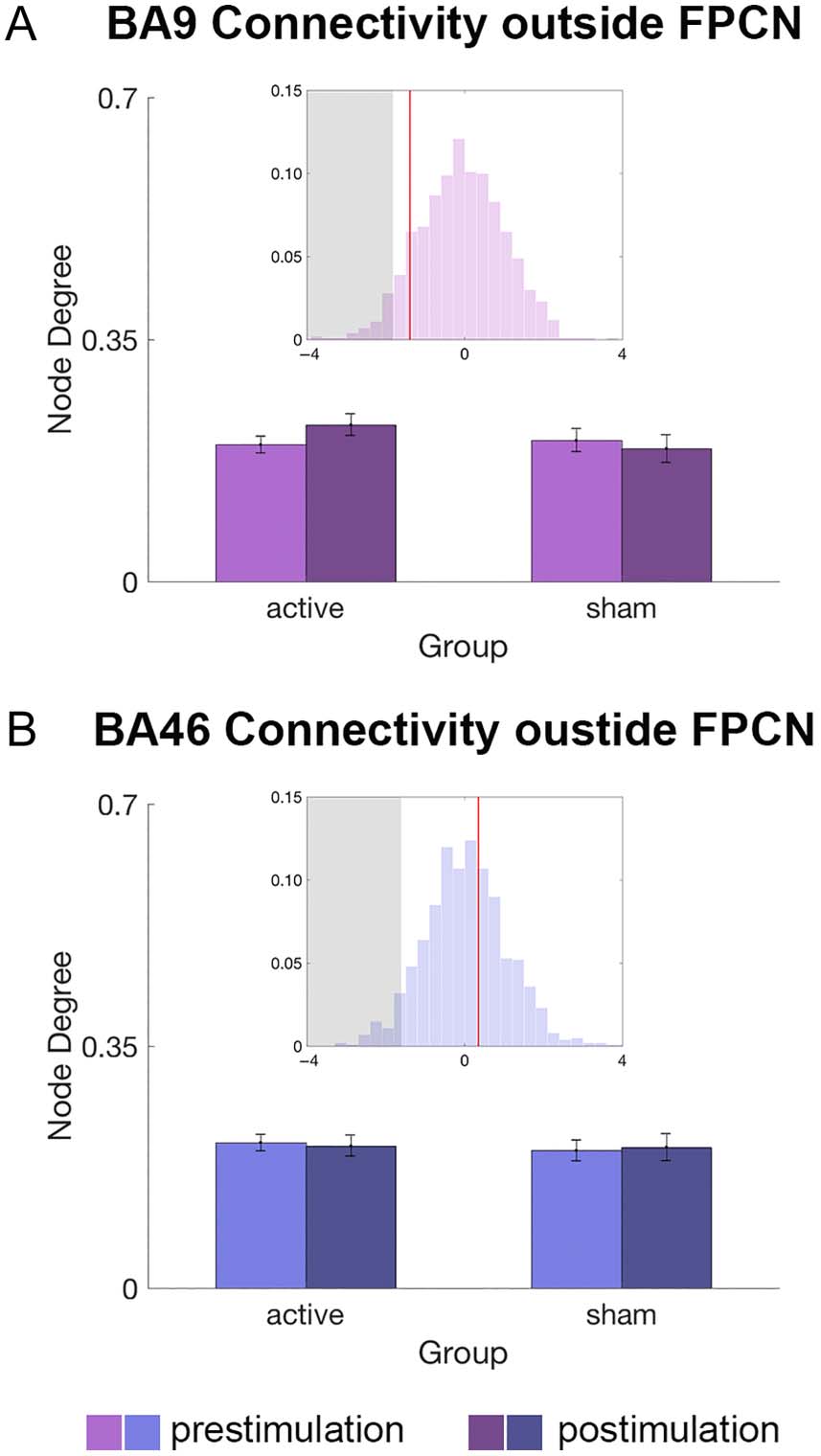
Next, we assessed stimulation effects on the stimulation
target’s connectivity characteristics outside FPCN. We
approached the outside-network connectivity via two dif-
ferent measures: the connectivity strength and the distri-
bution of connections outside FPCN.
First, outside-FPCN connectivity strength of an lDLPFC
ROI was quantified as node degree that only took nodes
that are not part of FPCN into consideration. For example,
BA 9’s outside-FPCN connectivity strength was computed
Table 2. Comparisons of Sham and Two Active Stimulation Groups
Stimulation Group
Group Difference
Sham (n = 22)
Active - 1mA (n = 13)
Active - 2mA (n = 16)
demographic
sex
age
F = 5
28.1 (5.2)
F = 4
28.8 (6.2)
F = 4
27.8 (5.7)
scanning time
interval
159.1 (16.86)
158.9 (17.39)
158.9 (14.45)
sensation
itching
pain
heat
1.73 (1.70)
0.36 (0.85)
0.23 (0.43)
discomfort
1.05 (1.43)
fatigue
4.09 (1.02)
1.31 (1.18)
0.77 (1.17)
0.54 (0.97)
0.85 (1.21)
3.69 (1.32)
Stimulation groups did not differ in age, scanning time, and sensation reports.
1.44 (1.26)
0.44 (0.63)
0.56 (0.59)
0.56 (0.73)
4.44 (1.31)
F
NA
0.108
0.001
0.388
0.906
1.173
0.756
1.396
p
NA
.9
.99
.68
.41
.32
.48
.26
1386
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
as the number of edges between BA 9 and non-FPCN
nodes divided by possible number of edges between
BA 9 and all non-FPCN nodes (i.e., denominator was the
total number of non-FPCN nodes). GLMM permutation
tests revealed that the connectivity strength of the
lDLPFC ROIs (BA 9, BA 46) with nodes outside FPCN was
not significantly altered by the stimulation (BA 9: |z| =
1.23, Figure 4A; BA 46 |z| = 0.29, Figure 4B). Similarly,
stimulation did not alter rDLPFC nodes’ connectivity with
nodes outside FPCN (all |z| < 0.8). In summary, the stim-
ulation did not alter the net strength of the connectivity
between lDLPFC ROIs and brain regions outside FPCN.
In our next analyses, we assessed the effects of tDCS on
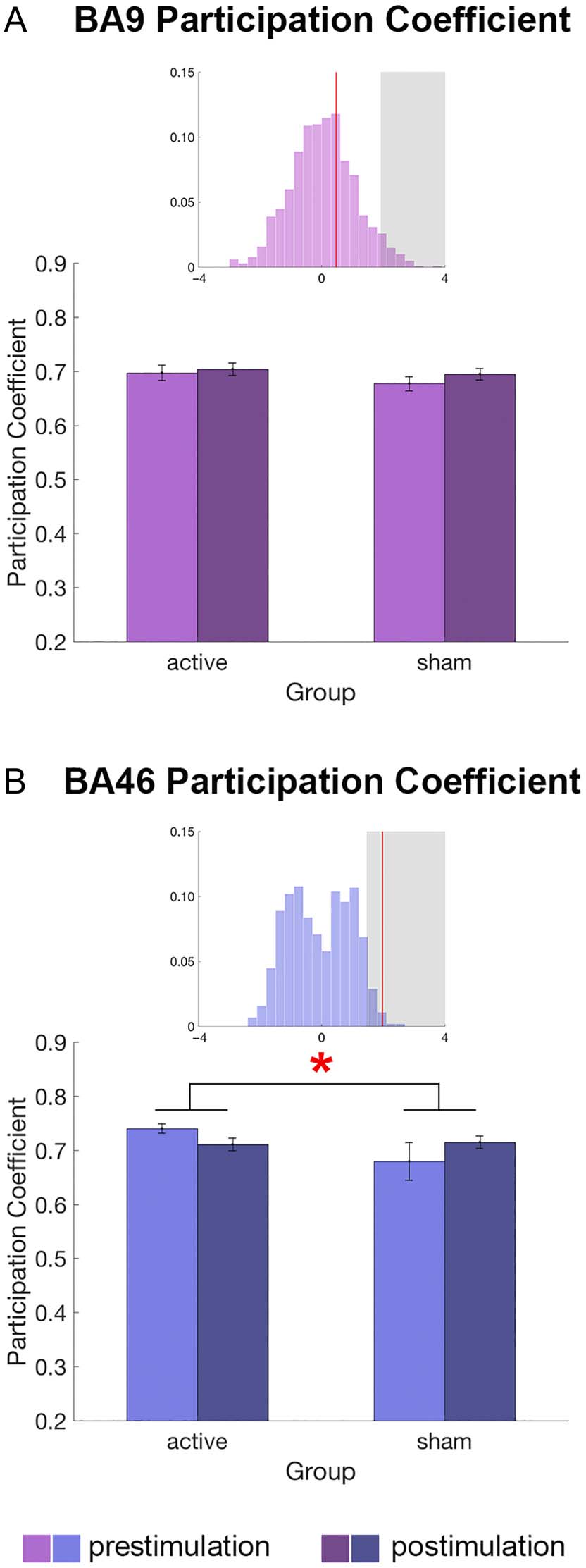
the participation coefficient, the measure that character-
izes how well-distributed a node’s connections are
(Meunier, Lambiotte, & Bullmore, 2010; Bullmore &
Sporns, 2009; Guimerà, Sales-Pardo, & Amaral, 2007;
Guimerà & Amaral, 2005) across the seven networks that
were predefined in the atlas (Schaefer et al., 2018). GLMM
regression revealed that the participation coefficient of
BA 46 was reduced by lDLPFC stimulation relative to sham
(|z| = 1.98; Figure 5B), whereas stimulation did not
significantly affect participation coefficient values in BA 9
(|z| = 0.37; Figure 5A) or the rDLPFC (|z| < 0.2). We
also tested whether the modulation of BA 46 participa-
tion coefficient was stimulation intensity-dependent,
and found no difference in the stimulation effects be-
tween the two intensity groups (|z| = 0.25).
Close examination of the data indicated potential
between-group differences in the baseline (prestimula-
tion) BA 46 participation coefficient values (Figure 5B,
light blue), and we further inspected this result.
Consistent with this impression, we found that prestimu-
lation BA 46 participation coefficient values were signifi-
cantly higher for participants in the active stimulation
condition relative to participants in the sham conditions
(|z| = 1.82). To address whether the interaction effect
was merely a consequence of prestimulation group differ-
ence, we conducted follow-up analyses on subgroups of
sham and active condition participants that were closely
matched in the prestimulation participation coefficient
values. Participation coefficient-matched groups were
identified as follows: First, we identified the range of the
BA 46 participation coefficients that overlapped between
the active and sham stimulation groups, and excluded
participants that fell outside the range. This eliminated a
few participants from each stimulation group, and yielded
20 and 27 participants for the sham and active stimulation
groups, respectively. Next, for the participation coeffi-
cient value of each participant in the sham stimulation
condition, the closest participation coefficient value was
identified from active stimulation participants. The search
for a matching pair was conducted through the sham
participants’ participation coefficient values in a random
order. This matching process was repeated 1000 times
with a new random order each time. The goodness of
matching was assessed as the mean difference of the
Kim et al.
1387
Figure 3. FPCN-connectivity changes of lDLPFC ROIs. Bar graphs
depict lDLPFC ROI’s connectivity with FPCN in each stimulation
group and session, and insets depict permutation test results for
the interaction effect (Stimulation Session × Group). Histogram:
permutation distribution, x-axis: coefficient size, y-axis: probability,
red line: regression coefficient, gray shade: < 5th percentile of the
permutation distribution. (A) BA 9-FPCN connectivity was significantly
increased by active stimulation. (B) BA 46-FPCN was not significantly
modulated by active stimulation.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
lDLPFC and networks of the brain. We used functional
connectivity measures computed from resting-state fMRI
scans acquired before and after the participants under-
went a stimulation session. Relative to sham stimulation,
active stimulation altered the functional connectivity of
lDLPFC in two ways. First, stimulation increased the func-
tional connectivity between BA 9 and other nodes within
the FPCN. Second, stimulation impacted the connectivity
of BA 46 across different networks. While the net connec-
tivity strength of BA 46 was not altered within FPCN or
outside FPCN, the connectivity distribution (participation
coefficient) of BA 46 was decreased by stimulation. In con-
trast to the effects of stimulation on lDLPFC, stimulation
had no significant effects on network-level functional con-
nectivity metrics in homologous rDLPFC regions that are
also part of the FPCN.
Our results build on prior findings showing that lDLPFC
stimulation impacts the connectivity of FPCN and the
functional connectivity of lDLPFC. Keeser et al. (2011) ex-
amined stimulation-induced changes in the functional
connectivity of resting-state networks. Using independent
component analysis on resting-state fMRI scans obtained
from 13 healthy participants, they identified four net-
works: default mode, left frontoparietal, right frontoparie-
tal, and self-referential networks. Next, authors used dual
regression method (Nickerson, Smith, Öngür, &
Beckmann, 2017; Beckmann, DeLuca, Devlin, & Smith,
2005) to identify subject-specific spatial maps for the net-
works, which were entered into the group-level statistical
tests to assess stimulation effects in a network (contrast:
(after real tDCS > baseline1) > (after sham tDCS > base-
line)). Analysis of the left frontoparietal network revealed
increased coactivation between regions within the superior
and inferior frontal gyri, the inferior parietal lobule, and
the posterior cingulate gyrus. Apart from the connectivity
changes, authors also found significant increase of local
activation in the anodal target region (BA 6). However,
the independent component analysis did not identify
the stimulation target as part of the left frontoparietal net-
work, and as a consequence, the dual regression analysis
did not assess the connectivity relationship between the
stimulation target and other regions in the network. In
this study, we capitalized on a network atlas that is prede-
fined from a large, independent sample (Schaefer et al.,
2018), and tested how stimulation impacts the stimula-
tion target’s network relationships with FPCN and other
networks.
Whereas Keeser et al. (2011) focused on the connectiv-
ity strength changes in different brain networks, other re-
search focused on pairwise connectivity between lDLPFC
and other brain regions. Using a seed-based connectivity
analysis, Mondino et al. (2020) showed that the lDLPFC
stimulation increased resting-state functional connectivity
between lDLPFC and bilateral parietal regions (IPL, SPL).
IPL and SPL belong to FPCN, and therefore these findings
are consistent with our finding that lDLPFC connectivity
within FPCN was increased by stimulation. In this study,
Figure 4. lDLPFC connectivity changes outside FPCN. Bar graphs
depict lDLPFC ROI’s connectivity with regions outside FPCN in each
stimulation group and session, and insets depict permutation test
results for the interaction effect (Stimulation Session × Group).
Histogram: permutation distribution, x- axis: coefficient size, y-axis:
probability, red line: regression coefficient, gray shade: < 5th percentile
of the permutation distribution. (A) BA 9 Connectivity with non-FPCN
regions was not significantly altered by active stimulation. (B) BA 46
Connectivity with non-FPCN regions was not significantly altered by
active stimulation.
participation coefficient, and the matching solution with
the smallest mean participation coefficient difference was
selected for further analysis (difference mean = 0.0258;
standard deviation = 0.0291). Permutation GLMM revealed
that 1) BA 46 participation coefficients were matched be-
tween the sham and the active stimulation groups (|z| =
1.26) in the selected subsample and that 2) stimulation
significantly reduced the BA 46 participation coefficients
(|z| = 1.68, significant interaction effects) in this subset
of participants.
DISCUSSION
The goal of this study was to delineate how tDCS applied
to the lDLPFC modulates interactions between the
1388
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
we further took the network structure into consideration,
and found that the lDLPFC stimulation might increase
the role of lDLPFC in the FPCN via modulating the con-
nectivity between lDLPFC and other regions within the
network.
Given that the participation coefficient reflects the dis-
tribution of connectivity across networks, it is somewhat
counterintuitive that stimulation significantly strength-
ened connectivity of BA 9 within FPCN but stimulation
did not significantly affect its participation coefficient.
We suspected that this might reflect a concomitant in-
crease of connectivity between BA 9 and other networks
in addition to the FPCN. The numerical increase of
outside-FPCN connectivity of BA 9 (Figure 4A) was consis-
tent with this idea, and when we further probed BA 9’s
connectivity with each network, tDCS numerically in-
creased connectivity between BA 9 and all other networks
with the exception of Salience Ventral Attention Network.
In other words, BA 9 obtained stronger affinity/functional
connectivity with a broad range of networks, which did
not significantly alter the relative distribution of its con-
nections across networks.
In contrast, there was a significant Group × Session inter-
action effect on the participation coefficient of BA 46. This
finding is interesting as it suggests that the stimulation may
not only modulate connectivity within the targeted network
but also modulate the target’s connectivity with other net-
works. Given that BA 46 connectivity differed between the
sham and active stimulation groups in the prestimulation
data (see interaction effects and prestimulation differences
in Results section), we performed a follow-up analysis on
subsamples of participants who were equated in the presti-
mulation participation coefficient values. Even after match-
ing prestimulation participation coefficient values, we
found a significant effect of stimulation on the BA 46 partic-
ipation coefficient, suggesting that our finding was not sim-
ply because of sampling error. The finding that the
connectivity distribution (participation coefficient) of BA
46 was decreased by stimulation suggests that BA 46 be-
came less likely to be interacting with multiple other
networks.
There are some limitations of this study. The study de-
sign included only sham and active stimulation groups
and did not have an active control condition. Although
the current design with a sham stimulation condition al-
lows for comparing effects of the stimulation to the ab-
sence of stimulation, we cannot conclude from this
study that lDLPFC is the only stimulation target that can
lead to the connectivity changes shown in our results.
Relatedly, one may argue that connectivity changes might
be mere epiphenomena of stimulation-related sensation,
or arousal. We ruled out this possibility via demonstrating
that there was no between-group difference in the sensa-
tion and arousal ratings. However, we acknowledge that
an optimal approach is to include an active control condi-
tion in the design and that studies following up on the pres-
ent findings should include an active control condition.
Kim et al.
1389
Figure 5. Changes of lDLPFC participation coefficient. (A)
Stimulation did not significantly alter participation coefficient of BA 9.
(B) Participation Coefficient of BA 46 was significantly reduced by
stimulation. The bar graph depicts participation coefficients in each
stimulation group and session, and insets depict permutation test
results for the interaction effect (Stimulation Session × Group).
Histogram: permutation distribution, x- axis: coefficient size, y-axis:
probability, red line: regression coefficient, gray shade: > 95th
percentile of the permutation distribution.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Another caveat is that the active stimulation group in-
cluded participants who received different stimulation in-
tensities (1 mA or 2 mA). We used these intensities as
lDLFPC stimulation delivered at 1 mA or 2 mA has been
shown to be effective for yielding behavioral effects in
prior research (e.g., for vigilance/attention, 1 mA: Nejati
et al., 2020; Nelson et al., 2014; Gladwin et al., 2012;
2 mA: McIntire et al., 2017; McKinley et al., 2013), but
we treated them together as the active stimulation group
in the analyses as we did not have strong a priori predic-
tions on the effects of stimulation intensity. Nevertheless,
to address the possibility that stimulation effects shown
in this study could have been driven by stimulation of
one intensity and not the other, we first assured that
there was no preexisting difference between 1 and 2 mA
groups (Table 2; demographic feature, scanning intervals,
and stimulation-related sensation reports). More impor-
tantly, we tested whether the two groups differed in any
of the stimulation effects using GLMM and permutation
tests, and found no between-group difference (see
Results section for details). Therefore, it is unlikely that
the connectivity changes observed in this study were pre-
dominantly driven by one stimulation intensity group.
With the present data, we are not able to directly relate
the observed network changes to cognitive performance
measures and, therefore, cannot draw conclusions on
what would be the functional benefit of the lDLPFC-
FPCN connectivity changes. Building on the present
results, future studies can investigate whether lDLPFC
stimulation improves cognition by modulating connectiv-
ity of BA 9 within the FPCN. Accumulated fMRI findings
have established that the lDLPFC plays essential roles in
various cognitive functions including cognitive control
(meta-analysis and reviews: Vanderhasselt, De Raedt, &
Baeken, 2009; Neumann, von Cramon, & Lohmann,
2008; Nee, Wager, & Jonides, 2007; Owen, McMillan,
Laird, & Bullmore, 2005), long-term memory processing
(meta-analysis and reviews: Blumenfeld & Ranganath,
2007; Fletcher & Henson, 2001; Nolde, Johnson, & Raye,
1998) and working memory maintenance (meta-analysis
and reviews: Wager & Smith, 2003). Therefore, on the
one hand, increasing neuronal excitability in the lDLPFC
via anodal tDCS might be sufficient for modulating these
task functions. On the other hand, it is notable that these
cognitive functions also engage other regions in the FPCN
(Lemire-Rodger et al., 2019; Roberts, Libby, Inhoff, &
Ranganath, 2018; Thakral, Wang, & Rugg, 2017; Spaniol
et al., 2009; Blumenfeld & Ranganath, 2006; Owen et al.,
2005; Wager & Smith, 2003; Dobbins, Foley, Schacter, &
Wagner, 2002; Pessoa, Gutierrez, Bandettini, & Ungerleider,
2002). This brings up the possibility that the tightened
lDLPFC-FPCN connectivity may contribute to behavioral
modulations associated with lDLPFC stimulations. Future
studies using a wider array of cognitive tasks should allow
for directly investigating this question.
The present results raise the question as to whether the
kinds of modulations seen following lDLPFC stimulation
would generalize to targeted stimulation of other nodes
of FPCN. For example, if tDCS is applied to a node of
FPCN other than lDLPFC, would the connectivity of that
node be modulated the same way as lDLPFC? Or is it a
unique characteristic of lDLPFC that its network relation-
ships are particularly malleable? Future research should
also address the importance of the duration and additivity
of modulation effects. Are the modulation effects on the
target connectivity long-lasting? If so, what does the decay
function look like? In addition to the duration of stimula-
tion effects, it would be informative to understand whether
the observed stimulation effects can be cumulative over
repeated stimulation. A prior transcranial magnetic
stimulation study showed that stimulation repeated over
five consecutive days (i.e., five stimulation sessions)
brought about memory enhancement that lasted for a
long term (e.g., 15 days; Wang & Voss, 2015). Some
behavioral and imaging studies have tested the effects of
repeated tDCS, but so far evidence is sparse for additive
effects. tDCS repeatedly applied with a 10-hr-long gap
between stimulation sessions did not have an additive be-
havioral benefit (McIntire, McKinley, Goodyear, McIntire,
& Nelson, 2020). tDCS applied repeatedly over three con-
secutive days significantly influenced brain perfusion
measured using arterial spin labeling (Sherwood et al.,
2018); however, we do not know whether these tDCS
effects additively increased across sessions in a dose-
dependent manner. Future research is needed to under-
stand whether the target connectivity modulation effects
can be sustained and/or enhanced via repeated applica-
tion of tDCS.
Acknowledgments
We thank Casserly R. Mullenger and Aaron T. Madaris for their
assistance in data collection.
Reprint requests should be sent to Kamin Kim or Charan
Ranganath, Center for Neuroscience, University of California
Davis, 1544 Newton Cir., Davis, CA, or via e-mail: kmikim
@ucdavis.edu; cranganath@ucdavis.edu.
Author Contributions
Kamin Kim: Conceptualization; Formal analysis;
Investigation; Software; Visualization; Writing—Original
draft; Writing—Review & editing. Matthew S. Sherwood:
Conceptualization; Data curation; Investigation;
Methodology; Project administration; Writing—Review &
editing. Lindsey K. McIntire: Data curation; Investigation;
Project administration; Writing—Review & editing. Andy
R. McKinley: Conceptualization; Funding acquisition;
Resources; Supervision; Writing—Review & editing. Charan
Ranganath: Conceptualization; Funding acquisition;
Resources; Supervision; Writing—Original draft; Writing—
Review & editing.
1390
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Funding Information
This project was supported by the Air Force Research
Laboratory (AFRL) under the Human Interface and
Research Technology program (https://dx.doi.org/10
.13039/100000005), grant number: FA8650-14-D-6500;
and by a Vannevar Bush Faculty Fellowship (ONR grant
N00014-15-1-0033) to C.R. Any opinions, findings, and
conclusions or recommendations expressed in this mate-
rial are those of the authors and do not necessarily reflect
the views of the U.S. Department of Defense.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed
a persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/ W = .108, and W/ W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/ W = .102, and W/ W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance ex-
plicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender cita-
tion balance. The authors of this article report its propor-
tions of citations by gender category to be as follows:
M/M = .523, W/M = .233, M/ W = .116, and W/ W = .128.
Note
1. A stimulation session was preceded and followed by a scan-
ning session, and resting-state fMRI scans were acquired at the
beginning of each scanning session. Therefore, the interval be-
tween the prestimulation resting-state fMRI scanning and the
stimulation session (approximately 75 min) included some in-
scanner time, and stimulation setup time. During the in-scanner
time following resting-state fMRI scanning, participants engaged
in a working memory task for 30 min and then stayed task-free
for the remaining time in the scanner (∼22 min, structural MRI,
DTI, ASL, and MRS). Participants were instructed to relax and re-
main still during structural MRI, DTI, and MRS scanning. They
were further informed that they could close their eyes but needed
to remain awake. The fixation point remained on the screen; how-
ever, no other stimuli (auditory or visual) were provided. During
ASL scanning, participants were instructed to relax, clear their
mind and let their thoughts wander freely, and focus on the fixa-
tion dot. The interval between and the stimulation session and
the poststimulation resting-state fMRI scanning (approximately
10–40 min) included stimulation wrap-up and waiting time (be-
cause of scanner logistics, some participants had waiting time of
up to 30 min before the poststimulation scanning). During the
waiting time, participants stayed in the preparation area at the
testing site.
REFERENCES
Andrews, S. C., Hoy, K. E., Enticott, P. G., Daskalakis, Z. J., &
Fitzgerald, P. B. (2011). Improving working memory: The
effect of combining cognitive activity and anodal transcranial
direct current stimulation to the left dorsolateral prefrontal
cortex. Brain Stimulation, 4, 84–89. https://doi.org/10.1016/j
.brs.2010.06.004, PubMed: 21511208
Ashourvan, A., Telesford, Q. K., Verstynen, T., Vettel, J. M., &
Bassett, D. S. (2019). Multi-scale detection of hierarchical
community architecture in structural and functional brain
networks. PLoS One, 14, e0215520. https://doi.org/10.1371
/journal.pone.0215520, PubMed: 31071099
Au, J., Katz, B., Buschkuehl, M., Bunarjo, K., Senger, T., Zabel,
C., et al. (2016). Enhancing working memory training with
transcranial direct current stimulation. Journal of Cognitive
Neuroscience, 28, 1419–1432. https://doi.org/10.1162/jocn_a
_00979, PubMed: 27167403
Barch, D. M., Braver, T. S., Nystrom, L. E., Forman, S. D., Noll,
D. C., & Cohen, J. D. (1997). Dissociating working memory
from task difficulty in human prefrontal cortex.
Neuropsychologia, 10, 1373–1380. https://doi.org/10.1016
/S0028-3932(97)00072-9, PubMed: 9347483
Beckmann, C. F., DeLuca, M., Devlin, J. T., & Smith, S. M.
(2005). Investigations into resting-state connectivity using
independent component analysis. Philosophical Transactions
of the Royal Society of London, Series B: Biological Sciences,
360, 1001–1013. https://doi.org/10.1098/rstb.2005.1634,
PubMed: 16087444
Behzadi, Y., Restom, K., Liau, J., & Liu, T. T. (2007). A component
based noise correction method (CompCor) for BOLD and
perfusion based fMRI. Neuroimage, 37, 90–101. https://doi
.org/10.1016/j.neuroimage.2007.04.042, PubMed: 17560126
Bennabi, D., & Haffen, E. (2018). Transcranial direct current
stimulation (tDCS): A promising treatment for major
depressive disorder? Brain Sciences, 8, 81. https://doi.org/10
.3390/brainsci8050081, PubMed: 29734768
Blumenfeld, R. S., & Ranganath, C. (2006). Dorsolateral
prefrontal cortex promotes long-term memory formation
through its role in working memory organization. Journal of
Neuroscience, 26, 916–925. https://doi.org/10.1523
/JNEUROSCI.2353-05.2006, PubMed: 16421311
Blumenfeld, R. S., & Ranganath, C. (2007). Prefrontal cortex and
long-term memory encoding: An integrative review of
findings from neuropsychology and neuroimaging.
Neuroscientist, 13, 280–291. https://doi.org/10.1177
/1073858407299290, PubMed: 17519370
Braver, T. S., Cohen, J. D., Nystrom, L. E., Jonides, J., Smith,
E. E., & Noll, D. C. (1997). A parametric study of prefrontal
cortex involvement in human working memory.
Neuroimage, 5, 49–62. https://doi.org/10.1006/nimg.1996
.0247, PubMed: 9038284
Bullmore, E., & Sporns, O. (2009). Complex brain networks:
Graph theoretical analysis of structural and functional
systems. Nature Reviews Neuroscience, 10, 186–198. https://
doi.org/10.1038/nrn2575, PubMed: 19190637
Carvalho, S., Boggio, P. S., Gonçalves, Ó. F., Vigário, A. R., Faria,
M., Silva, S., et al. (2015). Transcranial direct current
stimulation based metaplasticity protocols in working
memory. Brain Stimulation, 8, 289–294. https://doi.org/10
.1016/j.brs.2014.11.011, PubMed: 25550147
Cole, M. W., Bassett, D. S., Power, J. D., Braver, T. S., &
Petersen, S. E. (2014). Intrinsic and task-evoked network
architectures of the human brain. Neuron, 83, 238–251.
https://doi.org/10.1016/j.neuron.2014.05.014, PubMed:
24991964
Cole, M. W., Ito, T., Bassett, D. S., & Schultz, D. H. (2016).
Activity flow over resting-state networks shapes cognitive task
activations. Nature Neuroscience, 19, 1718–1726. https://doi
.org/10.1038/nn.4406, PubMed: 27723746
D’Esposito, M., Aguirre, G. K., Zarahn, E., Ballard, D., Shin, R. K.,
& Lease, J. (1998). Functional MRI studies of spatial and
Kim et al.
1391
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
nonspatial working memory. Cognitive Brain Research, 7,
1–13. https://doi.org/10.1016/S0926-6410(98)00004-4,
PubMed: 9714705
D’Esposito, M., Postle, B. R., Ballard, D., & Lease, J. (1999).
Maintenance versus manipulation of information held in
working memory: An event-related fMRI study. Brain and
Cognition, 41, 66–86. https://doi.org/10.1006/brcg.1999.1096,
PubMed: 10536086
Damoiseaux, J. S., Rombouts, S. A. R. B., Barkhof, F., Scheltens,
P., Stam, C. J., Smith, S. M., et al. (2006). Consistent resting-
state networks across healthy subjects. Proceedings of the
National Academy of Sciences, U.S.A., 103, 13848–13853.
https://doi.org/10.1073/pnas.0601417103, PubMed: 16945915
De Luca, M., Beckmann, C. F., De Stefano, N., Matthews, P. M.,
& Smith, S. M. (2006). FMRI resting state networks define
distinct modes of long-distance interactions in the human
brain. Neuroimage, 29, 1359–1367. https://doi.org/10.1016/j
.neuroimage.2005.08.035, PubMed: 16260155
Dedoncker, J., Baeken, C., De Raedt, R., & Vanderhasselt, M.-A.
(2021). Combined transcranial direct current stimulation and
psychological interventions: State of the art and promising
perspectives for clinical psychology. Biological Psychology,
158, 107991. https://doi.org/10.1016/j.biopsycho.2020
.107991, PubMed: 33232800
Dedoncker, J., Brunoni, A. R., Baeken, C., & Vanderhasselt,
M.-A. (2016a). A systematic review and meta-analysis of the
effects of transcranial direct current stimulation (tDCS)
over the dorsolateral prefrontal cortex in healthy and
neuropsychiatric samples: Influence of stimulation
parameters. Brain Stimulation, 9, 501–517. https://doi.org/10
.1016/j.brs.2016.04.006, PubMed: 27160468
Dedoncker, J., Brunoni, A. R., Baeken, C., & Vanderhasselt,
M.-A. (2016b). The effect of the interval-between-sessions on
prefrontal transcranial direct current stimulation (tDCS) on
cognitive outcomes: A systematic review and meta-analysis.
Journal of Neural Transmission, 123, 1159–1172. https://doi
.org/10.1007/s00702-016-1558-x, PubMed: 27145765
Dobbins, I. G., Foley, H., Schacter, D. L., & Wagner, A. D.
(2002). Executive control during episodic retrieval: Multiple
prefrontal processes subserve source memory. Neuron,
35, 989–996. https://doi.org/10.1016/S0896-6273(02)00858-9,
PubMed: 12372291
Filmer, H. L., Lyons, M., Mattingley, J. B., & Dux, P. E. (2017).
Anodal tDCS applied during multitasking training leads to
transferable performance gains. Scientific Reports, 7, 1–11.
https://doi.org/10.1038/s41598-017-13075-y, PubMed:
29021526
Filmer, H. L., Mattingley, J. B., & Dux, P. E. (2020). Modulating
brain activity and behaviour with tDCS: Rumours of its
death have been greatly exaggerated. Cortex, 123, 141–151.
https://doi.org/10.1016/j.cortex.2019.10.006, PubMed:
31783223
Fletcher, P. C., & Henson, R. N. A. (2001). Frontal lobes and
human memory: Insights from functional neuroimaging.
Brain, 124, 849–881. https://doi.org/10.1093/brain/124.5.849,
PubMed: 11335690
Fox, M. D., & Raichle, M. E. (2007). Spontaneous fluctuations in
brain activity observed with functional magnetic resonance
imaging. Nature Reviews Neuroscience, 8, 700–711. https://
doi.org/10.1038/nrn2201, PubMed: 17704812
Fregni, F., Boggio, P. S., Nitsche, M., Bermpohl, F., Antal, A.,
Feredoes, E., et al. (2005). Anodal transcranial direct current
stimulation of prefrontal cortex enhances working memory.
Experimental Brain Research, 166, 23–30. https://doi.org/10
.1007/s00221-005-2334-6, PubMed: 15999258
episodic memory. Brain Stimulation, 12, 231–241.
https://doi.org/10.1016/j.brs.2018.11.008
Gandiga, P. C., Hummel, F. C., & Cohen, L. G. (2006).
Transcranial DC stimulation (tDCS): A tool for double-blind
sham-controlled clinical studies in brain stimulation. Clinical
Neurophysiology, 117, 845–850. https://doi.org/10.1016/j
.clinph.2005.12.003, PubMed: 16427357
Gladwin, T. E., den Uyl, T. E., Fregni, F. F., & Wiers, R. W.
(2012). Enhancement of selective attention by tDCS:
Interaction with interference in a Sternberg task.
Neuroscience Letters, 512, 33–37. https://doi.org/10.1016/j
.neulet.2012.01.056, PubMed: 22327036
Guimerà, R., & Amaral, L. A. N. (2005). Functional cartography
of complex metabolic networks. Nature, 433, 895–900.
https://doi.org/10.1038/nature03288, PubMed: 15729348
Guimerà, R., Sales-Pardo, M., & Amaral, L. A. N. (2007). Classes
of complex networks defined by role-to-role connectivity
profiles. Nature Physics, 3, 63–69. https://doi.org/10.1038
/nphys489, PubMed: 18618010
Hill, A. T., Fitzgerald, P. B., & Hoy, K. E. (2016). Effects of anodal
transcranial direct current stimulation on working memory: A
systematic review and meta-analysis of findings from healthy
and neuropsychiatric populations. Brain Stimulation, 9,
197–208. https://doi.org/10.1016/j.brs.2015.10.006, PubMed:
26597929
Jansma, J. M., Ramsey, N. F., de Zwart, J. A., van Gelderen, P., &
Duyn, J. H. (2007). fMRI study of effort and information
processing in a working memory task. Human Brain
Mapping, 28, 431–440. https://doi.org/10.1002/hbm.20297,
PubMed: 17133397
Jantz, T. K., Katz, B., & Reuter-Lorenz, P. A. (2016). Uncertainty
and promise: The effects of transcranial direct current
stimulation on working memory. Current Behavioral
Neuroscience Reports, 3, 109–121. https://doi.org/10.1007
/s40473-016-0071-8
Javadi, A. H., Cheng, P., & Walsh, V. (2012). Short duration
transcranial direct current stimulation (tDCS) modulates
verbal memory. Brain Stimulation, 5, 468–474. https://doi
.org/10.1016/j.brs.2011.08.003, PubMed: 21962975
Javadi, A. H., & Walsh, V. (2012). Transcranial direct current
stimulation (tDCS) of the left dorsolateral prefrontal cortex
modulates declarative memory. Brain Stimulation, 5,
231–241. https://doi.org/10.1016/j.brs.2011.06.007, PubMed:
21840287
Kaiser, M., Martin, R., Andras, P., & Young, M. P. (2007).
Simulation of robustness against lesions of cortical networks.
European Journal of Neuroscience, 25, 3185–3192. https://
doi.org/10.1111/j.1460-9568.2007.05574.x, PubMed: 17561832
Keeser, D., Padberg, F., Reisinger, E., Pogarell, O., Kirsch, V.,
Palm, U., et al. (2011). Prefrontal direct current
stimulation modulates resting EEG and event-related
potentials in healthy subjects: A standardized low
resolution tomography (sLORETA) study. Neuroimage,
55, 644–657. https://doi.org/10.1016/j.neuroimage.2010.12
.004, PubMed: 21146614
Kim, K., Ekstrom, A. D., & Tandon, N. (2016). A network
approach for modulating memory processes via direct and
indirect brain stimulation: Toward a causal approach for the
neural basis of memory. Neurobiology of Learning and
Memory, 134, 162–177. https://doi.org/10.1016/j.nlm.2016.04
.001, PubMed: 27066987
Kim, J.-H., Kim, D.-W., & Im, C.-H. (2017). Brain areas
responsible for vigilance: An EEG source imaging study.
Brain Topography, 30, 343–351. https://doi.org/10.1007
/s10548-016-0540-0, PubMed: 28054159
Galli, G., Vadillo, M. A., Sirota, M., Feurra, M., & Medvedeva,
A. (2019). A systematic review and meta-analysis of the
effects of transcranial direct current stimulation (tDCS) on
Lancichinetti, A., & Fortunato, S. (2012). Consensus clustering
in complex networks. Scientific Reports, 2, 336. https://doi
.org/10.1038/srep00336, PubMed: 22468223
1392
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Langner, R., & Eickhoff, S. B. (2013). Sustaining attention to
simple tasks: A meta-analytic review of the neural mechanisms
of vigilant attention. Psychological Bulletin, 139, 870–900.
https://doi.org/10.1037/a0030694, PubMed: 23163491
Lemire-Rodger, S., Lam, J., Viviano, J. D., Stevens, W. D.,
Spreng, R. N., & Turner, G. R. (2019). Inhibit, switch, and
update: A within-subject fMRI investigation of executive
control. Neuropsychologia, 132, 107134. https://doi.org/10
.1016/j.neuropsychologia.2019.107134, PubMed: 31299188
Leshikar, E. D., Leach, R. C., McCurdy, M. P., Trumbo, M. C.,
Sklenar, A. M., Frankenstein, A. N., et al. (2017). Transcranial
direct current stimulation of dorsolateral prefrontal cortex
during encoding improves recall but not recognition
memory. Neuropsychologia, 106, 390–397. https://doi.org/10
.1016/j.neuropsychologia.2017.10.022, PubMed: 29056368
London, R. E., & Slagter, H. A. (2021). No effect of transcranial
direct current stimulation over left dorsolateral prefrontal
cortex on temporal attention. Journal of Cognitive
Neuroscience, 33, 756–768. https://doi.org/10.1162/jocn_a
_01679, PubMed: 33464163
Lucchiari, C., Sala, P. M., & Vanutelli, M. E. (2018). Promoting
creativity through transcranial direct current stimulation
(tDCS). A critical review. Frontiers in Behavioral
Neuroscience, 12, 167. https://doi.org/10.3389/fnbeh.2018
.00167, PubMed: 30116184
MacDonald, A. W., Cohen, J. D., Stenger, V. A., & Carter, C. S.
(2000). Dissociating the role of the dorsolateral prefrontal
and anterior cingulate cortex in cognitive control. Science,
288, 1835–1838. https://doi.org/10.1126/science.288.5472
.1835, PubMed: 10846167
Mancuso, L. E., Ilieva, I. P., Hamilton, R. H., & Farah, M. J.
(2016). Does transcranial direct current stimulation improve
healthy working memory?: A meta-analytic review. Journal of
Cognitive Neuroscience, 28, 1063–1089. https://doi.org/10
.1162/jocn_a_00956, PubMed: 27054400
Marshall, L., Mölle, M., Siebner, H. R., & Born, J. (2005).
Bifrontal transcranial direct current stimulation slows
reaction time in a working memory task. BMC Neuroscience,
6, 23. https://doi.org/10.1186/1471-2202-6-23, PubMed:
15819988
Martin, D. M., Liu, R., Alonzo, A., Green, M., & Loo, C. K. (2014).
Use of transcranial direct current stimulation (tDCS) to
enhance cognitive training: Effect of timing of stimulation.
Experimental Brain Research, 232, 3345–3351. https://doi
.org/10.1007/s00221-014-4022-x, PubMed: 24992897
McIntire, L. K., McKinley, R.A., Goodyear, C., McIntire, J. P., &
Nelson, J. M. (2020). Cognitive performance after repeated
exposure to transcranial direct current stimulation (tDCS)
during sleep deprivation. In P. M. Arezes & R. L. Boring (Eds.),
Advances in safety management and human performance
(pp. 302–313). Springer International Publishing. https://doi
.org/10.1007/978-3-030-50946-0_41
McIntire, L. K., McKinley, R. A., Goodyear, C., & Nelson, J.
(2014). A comparison of the effects of transcranial direct
current stimulation and caffeine on vigilance and cognitive
performance during extended wakefulness. Brain
Stimulation, 7, 499–507. https://doi.org/10.1016/j.brs.2014.04
.008, PubMed: 25047826
McIntire, L. K., McKinley, R. A., Nelson, J. M., & Goodyear, C.
(2017). Transcranial direct current stimulation versus caffeine
as a fatigue countermeasure. Brain Stimulation, 10,
1070–1078. https://doi.org/10.1016/j.brs.2017.08.005,
PubMed: 28851554
McKinley, R. A. (2018). Chapter 139—Transcranial direct
current stimulation for fatigue and attentional disorders.
In E. S. Krames, P. H. Peckham, & A. R. Rezai (Eds.),
Neuromodulation (2nd ed., pp. 1637–1648). Academic
Press. https://doi.org/10.1016/B978-0-12-805353-9.00139-X
McKinley, R. A., McIntire, L., Bridges, N., Goodyear, C., Bangera,
N. B., & Weisend, M. P. (2013). Acceleration of image analyst
training with transcranial direct current stimulation.
Behavioral Neuroscience, 127, 936–946. https://doi.org/10
.1037/a0034975, PubMed: 24341718
Meunier, D., Lambiotte, R., & Bullmore, E. T. (2010). Modular
and hierarchically modular organization of brain networks.
Frontiers in Neuroscience, 4, 200. https://doi.org/10.3389
/fnins.2010.00200, PubMed: 21151783
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of
prefrontal cortex function. Annual Review of Neuroscience,
24, 167–202. https://doi.org/10.1146/annurev.neuro.24.1.167,
PubMed: 11283309
Mizrak, E., Kim, K., Roberts, B., Ragland, D. J., Carter, C., &
Ranganath, C. (2018). Impact of oscillatory tDCS targeting left
prefrontal cortex on source memory retrieval. Cognitive
Neuroscience, 9, 194–207. https://doi.org/10.1080/17588928
.2018.1512480, PubMed: 30124354
Mondino, M., Ghumman, S., Gane, C., Renauld, E., Whittingstall,
K., & Fecteau, S. (2020). Effects of transcranial stimulation with
direct and alternating current on resting-state functional
connectivity: An exploratory study simultaneously combining
stimulation and multiband functional magnetic resonance
imaging. Frontiers in Human Neuroscience, 13, 474. https://
doi.org/10.3389/fnhum.2019.00474, PubMed: 32116597
Nee, D. E., Wager, T. D., & Jonides, J. (2007). Interference
resolution: Insights from a meta-analysis of neuroimaging
tasks. Cognitive, Affective, & Behavioral Neuroscience, 7,
1–17. https://doi.org/10.3758/CABN.7.1.1, PubMed: 17598730
Nejati, V., Salehinejad, M. A., Nitsche, M. A., Najian, A., & Javadi,
A. H. (2020). Transcranial direct current stimulation improves
executive dysfunctions in ADHD: Implications for inhibitory
control, interference control, working memory, and cognitive
flexibility. Journal of Attention Disorders, 24, 1928–1943.
https://doi.org/10.1177/1087054717730611, PubMed:
28938852
Nelson, J. T., McKinley, R. A., Golob, E. J., Warm, J. S., &
Parasuraman, R. (2014). Enhancing vigilance in operators
with prefrontal cortex transcranial direct current stimulation
(tDCS). Neuroimage, 85, 909–917. https://doi.org/10.1016/j
.neuroimage.2012.11.061, PubMed: 23235272
Neumann, J., von Cramon, D. Y., & Lohmann, G. (2008). Model-
based clustering of meta-analytic functional imaging data.
Human Brain Mapping, 29, 177–192. https://doi.org/10.1002
/hbm.20380, PubMed: 17390315
Nickerson, L. D., Smith, S. M., Öngür, D., & Beckmann, C. F.
(2017). Using dual regression to investigate network shape
and amplitude in functional connectivity analyses. Frontiers
in Neuroscience, 11, 115. https://doi.org/10.3389/fnins.2017
.00115, PubMed: 28348512
Nitsche, M. A., Cohen, L. G., Wassermann, E. M., Priori, A., Lang,
N., Antal, A., et al. (2008). Transcranial direct current
stimulation: State of the art 2008. Brain Stimulation, 1,
206–223. https://doi.org/10.1016/j.brs.2008.06.004, PubMed:
20633386
Nitsche, M. A., & Paulus, W. (2011). Transcranial direct current
stimulation—Update 2011. Restorative Neurology and
Neuroscience, 29, 463–492. https://doi.org/10.3233/RNN
-2011-0618, PubMed: 22085959
Nolde, S. F., Johnson, M. K., & Raye, C. L. (1998). The role of
prefrontal cortex during tests of episodic memory. Trends in
Cognitive Sciences, 2, 399–406. https://doi.org/10.1016/S1364
-6613(98)01233-9, PubMed: 21227255
Osório, A. A. C., & Brunoni, A. R. (2019). Transcranial direct
current stimulation in children with autism spectrum
disorder: A systematic scoping review. Developmental
Medicine & Child Neurology, 61, 298–304. https://doi.org/10
.1111/dmcn.14104, PubMed: 30451290
Kim et al.
1393
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Owen, A. M., McMillan, K. M., Laird, A. R., & Bullmore, E.
(2005). N-back working memory paradigm: A meta-analysis of
normative functional neuroimaging studies. Human Brain
Mapping, 25, 46–59. https://doi.org/10.1002/hbm.20131,
PubMed: 15846822
Park, J.-H., Hong, S. B., Kim, D.-W., Suh, M., & Im, C.-H. (2011).
A novel array-type transcranial direct current stimulation
(tDCS) system for accurate focusing on targeted brain areas.
IEEE Transactions on Magnetics, 47, 882–885. https://doi.org
/10.1109/TMAG.2010.2072987
Paulus, W. (2011). Transcranial electrical stimulation
(tES – tDCS; tRNS, tACS) methods. Neuropsychological
Rehabilitation, 21, 602–617. https://doi.org/10.1080
/09602011.2011.557292, PubMed: 21819181
Pessoa, L., Gutierrez, E., Bandettini, P. A., & Ungerleider, L. G.
(2002). Neural correlates of visual working memory: fMRI
amplitude predicts task performance. Neuron, 35, 975–987.
https://doi.org/10.1016/S0896-6273(02)00817-6, PubMed:
12372290
Power, J. D., Cohen, A. L., Nelson, S. M., Wig, G. S., Barnes,
K. A., Church, J. A., et al. (2011). Functional network
organization of the human brain. Neuron, 72, 665–678.
https://doi.org/10.1016/j.neuron.2011.09.006, PubMed:
22099467
Ranganath, C., Johnson, M. K., & D’Esposito, M. (2003).
Prefrontal activity associated with working memory and
episodic long-term memory. Neuropsychologia, 41, 378–389.
https://doi.org/10.1016/S0028-3932(02)00169-0, PubMed:
12457762
Razza, L. B., Palumbo, P., Moffa, A. H., Carvalho, A. F., Solmi,
M., Loo, C. K., et al. (2020). A systematic review and meta-
analysis on the effects of transcranial direct current
stimulation in depressive episodes. Depression and
Anxiety, 37, 594–608. https://doi.org/10.1002/da.23004,
PubMed: 32101631
Roberts, B. M., Libby, L. A., Inhoff, M. C., & Ranganath, C.
(2018). Brain activity related to working memory for
temporal order and object information. Behavioural Brain
Research, 354, 55–63. https://doi.org/10.1016/j.bbr.2017.05
.068, PubMed: 28602963
Rubinov, M., & Sporns, O. (2011). Weight-conserving
characterization of complex functional brain networks.
Neuroimage, 56, 2068–2079. https://doi.org/10.1016/j
.neuroimage.2011.03.069, PubMed: 21459148
Ruf, S. P., Fallgatter, A. J., & Plewnia, C. (2017). Augmentation of
working memory training by transcranial direct current
stimulation (tDCS). Scientific Reports, 7, 1–11. https://doi.org
/10.1038/s41598-017-01055-1, PubMed: 28432349
Sandrini, M., Brambilla, M., Manenti, R., Rosini, S., Cohen, L. G.,
& Cotelli, M. (2014). Noninvasive stimulation of prefrontal
cortex strengthens existing episodic memories and reduces
forgetting in the elderly. Frontiers in Aging Neuroscience, 6,
289. https://doi.org/10.3389/fnagi.2014.00289, PubMed:
25368577
Sandrini, M., Manenti, R., Brambilla, M., Cobelli, C., Cohen,
L. G., & Cotelli, M. (2016). Older adults get episodic
memory boosting from noninvasive stimulation of
prefrontal cortex during learning. Neurobiology of Aging,
39, 210–216. https://doi.org/10.1016/j.neurobiolaging.2015
.12.010, PubMed: 26923418
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N.,
Holmes, A. J., et al. (2018). Local-global parcellation of the
human cerebral cortex from intrinsic functional connectivity
MRI. Cerebral Cortex, 28, 3095–3114. https://doi.org/10.1093
/cercor/bhx179, PubMed: 28981612
Sherwood, M. S., Madaris, A. T., Mullenger, C. R., & McKinley,
R. A. (2018). Repetitive transcranial electrical stimulation
induces quantified changes in resting cerebral perfusion
measured from arterial spin labeling. Neural Plasticity, 2018,
5769861. https://doi.org/10.1155/2018/5769861, PubMed:
30254668
Silvanto, J., Muggleton, N., & Walsh, V. (2008). State-
dependency in brain stimulation studies of perception and
cognition. Trends in Cognitive Sciences, 12, 447–454.
https://doi.org/10.1016/j.tics.2008.09.004, PubMed:
18951833
Spaniol, J., Davidson, P. S. R., Kim, A. S. N., Han, H.,
Moscovitch, M., & Grady, C. L. (2009). Event-related fMRI
studies of episodic encoding and retrieval: Meta-analyses
using activation likelihood estimation. Neuropsychologia, 47,
1765–1779. https://doi.org/10.1016/j.neuropsychologia.2009
.02.028, PubMed: 19428409
Sporns, O., & Zwi, J. D. (2004). The small world of the cerebral
cortex. Neuroinformatics, 2, 145–162. https://doi.org/10
.1385/NI:2:2:145, PubMed: 15319512
Strobach, T., & Antonenko, D. (2017). tDCS-induced effects on
executive functioning and their cognitive mechanisms: A
review. Journal of Cognitive Enhancement, 1, 49–64. https://
doi.org/10.1007/s41465-016-0004-1
Talsma, L. J., Broekhuizen, J. A., Huisman, J., & Slagter, H. A.
(2018). No evidence that baseline prefrontal cortical
excitability (3T-MRS) predicts the effects of prefrontal tDCS
on WM performance. Frontiers in Neuroscience, 12, 481.
https://doi.org/10.3389/fnins.2018.00481, PubMed: 30065625
Talsma, L. J., Kroese, H. A., & Slagter, H. A. (2017). Boosting
cognition: Effects of multiple-session transcranial direct
current stimulation on working memory. Journal of
Cognitive Neuroscience, 29, 755–768. https://doi.org/10.1162
/jocn_a_01077, PubMed: 27897670
Thakral, P. P., Wang, T. H., & Rugg, M. D. (2017). Decoding the
content of recollection within the core recollection network
and beyond. Cortex, 91, 101–113. https://doi.org/10.1016/j
.cortex.2016.12.011, PubMed: 28077212
Tomita, H., Ohbayashi, M., Nakahara, K., Hasegawa, I., &
Miyashita, Y. (1999). Top–down signal from prefrontal cortex in
executive control of memory retrieval. Nature, 401, 699–703.
https://doi.org/10.1038/44372, PubMed: 10537108
Trumbo, M. C., Matzen, L. E., Coffman, B. A., Hunter, M. A.,
Jones, A. P., Robinson, C. S. H., et al. (2016). Enhanced
working memory performance via transcranial direct current
stimulation: The possibility of near and far transfer.
Neuropsychologia, 93, 85–96. https://doi.org/10.1016/j
.neuropsychologia.2016.10.011, PubMed: 27756695
Vanderhasselt, M.-A., De Raedt, R., & Baeken, C. (2009).
Dorsolateral prefrontal cortex and Stroop performance:
Tackling the lateralization. Psychonomic Bulletin & Review,
16, 609–612. https://doi.org/10.3758/PBR.16.3.609, PubMed:
19451392
Vincent, J. L., Kahn, I., Snyder, A. Z., Raichle, M. E., & Buckner,
R. L. (2008). Evidence for a frontoparietal control system
revealed by intrinsic functional connectivity. Journal of
Neurophysiology, 100, 3328–3342. https://doi.org/10.1152/jn
.90355.2008, PubMed: 18799601
Vincent, J. L., Snyder, A. Z., Fox, M. D., Shannon, B. J., Andrews,
J. R., Raichle, M. E., et al. (2006). Coherent spontaneous
activity identifies a hippocampal-parietal memory network.
Journal of Neurophysiology, 96, 3517–3531. https://doi.org
/10.1152/jn.00048.2006, PubMed: 16899645
Wager, T. D., & Smith, E. E. (2003). Neuroimaging studies of
working memory: A meta-analysis. Cognitive, Affective, &
Behavioral Neuroscience, 3, 255–274. https://doi.org/10
.3758/CABN.3.4.255, PubMed: 15040547
Wagner, T., Valero-Cabre, A., & Pascual-Leone, A. (2007).
Noninvasive human brain stimulation. Annual Review of
Biomedical Engineering, 9, 527–565. https://doi.org/10.1146
/annurev.bioeng.9.061206.133100, PubMed: 17444810
1394
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Wang, J. X., & Voss, J. L. (2015). Long-lasting enhancements of
memory and hippocampal-cortical functional connectivity
following multiple-day targeted noninvasive stimulation.
Hippocampus, 25, 877–883. https://doi.org/10.1002/hipo
.22416, PubMed: 17444810
Whitfield-Gabrieli, S., & Nieto-Castanon, A. (2012). Conn: A
functional connectivity toolbox for correlated and anticorrelated
brain networks. Brain Connectivity, 2, 125–141. https://doi
.org/10.1089/brain.2012.0073, PubMed: 22642651
Zwissler, B., Sperber, C., Aigeldinger, S., Schindler, S., Kissler,
J., & Plewnia, C. (2014). Shaping memory accuracy by left
prefrontal transcranial direct current stimulation. Journal of
Neuroscience, 34, 4022–4026. https://doi.org/10.1523
/JNEUROSCI.5407-13.2014, PubMed: 24623779
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
3
8
1
1
9
2
1
2
3
2
/
/
j
o
c
n
_
a
_
0
1
7
2
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Kim et al.
1395