Tian-tong Luo
Nanjing Agricultural University
Department of Zoology
College of Life Sciences
Jian-long Zhu
Nanjing Agricultural University
Department of Zoology
College of Life Sciences
Trond Reitan
University of Oslo
Centre for Ecological and
Evolutionary Synthesis
Department of Biology
trond.reitan@ibv.uio.no
Gabriel Yedid*
Nanjing Agricultural University
Department of Zoology
College of Life Sciences
gyedid02@gmail.com
Keywords
Coevolution, digital evolution,
evolutionary stasis, Red Queen, temporal
intransitivity
Alteration of (Frequency-
Dependent) Fitness in Time-Shift
Experiments Reveals Cryptic
Coevolution and Uncoordinated
Stasis in a Virtual Jurassic Park
Abstract Among the major unresolved questions in ecosystem
evolution are whether coevolving multispecies communities are
dominated more by biotic or by abiotic factors, and whether
evolutionary stasis affects performance as well as ecological profile;
these issues remain difficult to address experimentally. Digital
evolution, a computer-based instantiation of Darwinian evolution in
which short self-replicating computer programs compete, mutate, and
evolve, is an excellent platform for investigating such topics in a
rigorous experimental manner. We evolved model communities with
ecological interdependence among community members, which were
subjected to two principal types of mass extinction: a pulse extinction
that killed randomly, and a selective press extinction involving an
alteration of the abiotic environment to which the communities had
to adapt. These treatments were applied at two different strengths
(Strong and Weak), along with unperturbed Control experiments. We
performed several kinds of competition experiments using simplified
versions of these communities to see whether long-term stability that
was implied previously by ecological and phylogenetic metrics was
also reflected in performance, namely, whether fitness was static over
long periods of time. Results from Control and Weak treatment
communities revealed almost completely transitive evolution, while
Strong treatment communities showed higher incidences of temporal
intransitivity, with pre-treatment ecotypes often able to displace some
of their post-recovery successors. However, pre-treatment carryovers
more often had lower fitness in mixed communities than in their own
fully native conditions. Replacement and invasion experiments pitting
single ecotypes against pre-treatment reference communities showed
that many of the invading ecotypes could measurably alter the
fitnesses of one or more residents, usually with depressive effects,
and that the strength of these effects increased over time even in the
most stable communities. However, invaders taken from Strong
treatment communities often had little or no effect on resident
performance. While we detected periods of time when the fitness of a
particular evolving ecotype remained static, this stasis was not
permanent and never affected an entire community at once. Our
results lend support to the fitness-deterioration interpretation of the
Red Queen hypothesis, and highlight community context dependence
in determining fitness, the shaping of communities by both biotic
factors and abiotic forcing, and the illusory nature of evolutionary
stasis. Our results also demonstrate the potential of digital evolution
studies to illuminate many aspects of evolution in interacting
multispecies communities.
* Corresponding author.
© 2020 Massachusetts Institute of Technology.
Published under a Creative Commons Attribution
4.0 International (CC BY 4.0) license.
Artificial Life 26: 196–216 (2020) https://doi.org/10.1162/artl_a_00305
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
1 Introduction
1.1 Who Holds Court in Biological Communities, the Queen or the Jester?
The stability of biological communities made up of multiple interacting species may be considered
from both strictly ecological [39, 42, 48, 50] and evolutionary [17, 27, 31, 52] perspectives; namely,
whether the same factors that lead to ecological stability will also produce resistance to evolutionary
change, or whether these ecological interactions instead drive evolutionary change. There are two
major primary contrasting views of multispecies community evolution: the Red Queen [36, 52] and
the Court Jester [4]. In the former, the “environment” is defined largely by biotic interactions among
community members, which are the primary drivers of evolution. Any change in the strength or type
of these interactions stemming from evolutionary change in one interactor (individuals, species, etc.)
must then be matched by changes in others, with the corollary that any evolutionary advance in one
interactor will result in a loss of fitness in others, which must then evolve as quickly as possible
simply in order to maintain fitness. Taken to its logical extreme, the concept suggests that these
interactions might drive evolution indefinitely even if the external, abiotic environment could be held
completely constant.
At the other end of the continuum, the Court Jester view posits that evolution is governed by
chance survival of organisms in inconstant, unpredictable external environments, which may result
in massive redirections of the course of evolution. Although first proposed by a paleontologist [55],
the Red Queen has come to be favored by neontological biologists, particularly with regard to the
evolution of sexual reproduction and host-parasite coevolution (e.g., [6, 18, 40]), while the Court
Jester is favored by geoscience workers who see evolution of life on Earth as inseparable from major
geological and astronomical events in Earth history, to which life can only respond passively (e.g.,
[35]). A third view, dubbed “ecological locking” [41], also posits a web of biotic interactions among
community members, but these interactions come to impede evolution by generating community-
wide stabilizing selection, rather than driving evolution through continual directional selection. Evo-
lution may then only occur when that web of stabilizing interactions is destroyed in Court Jester-type
events. Clearly, real evolution contains elements of all these views [19, 20, 22, 24, 25, 46].
1.2 Looking at Going Somewhere While Running in Place
According to the Red Queen hypothesis, biotic interactions among organisms mean that they should
always be pushing each other towards ever greater states of adaptedness. Evolution may appear to
be standing still by some measures, but may actually be cryptically continuing [13]. Hence, if com-
munities were truly stable both ecologically and evolutionarily, and if communities from different
points in time could be competed repeatedly against each other, we could reasonably expect result-
ing mixed communities in which each member that occupied a particular ecological niche would
come 50% of the time from the earlier community and 50% of the time from the later community.
Types taken from the later time would be able to easily “slot in” to the earlier community (and vice
versa), without disrupting the performance of the rest of the community. While first proposed as
thought experiments several decades ago [52], such experiments remain in the realm of science fic-
tion, at least for vertebrate megafauna (e.g., [16]). However, they are easily achieved with microbial
experimental evolution, where samples of a microbial population evolving over many thousands of
generations may be kept frozen and revived as needed [14, 33]. An in silico alternative is digital
evolution, which offers similar capabilities at much greater speed and gives the experimenter a
greater degree of environmental control.
In previous work [38], we examined the tendency of cross-feeding communities of digital organ-
isms generated by the Avida digital evolution platform [44] to show stability with and without mass
extinction, as measured using metrics incorporating information from ecology and phylogeny. We
determined that 75% of communities generated under these conditions were ecologically unstable
even without mass extinction. However, about 25% of the replicate populations in our data set
showed some evidence for stability using eco-phenotypic metrics. It remains to be seen, however,
Artificial Life Volume 26, Number 2
197
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
whether these apparently stable communities are also stable in terms of performance, that is, whether
they would satisfy the aforementioned criteria.
Here, we use digital evolution to perform exactly such experiments, using simplified versions of
the communities used in previous work [38, 59, 62] to look for evidence of performance stasis in com-
munities diversified into multiple coexisting ecotypes. We examine whether or not such stability
exists at the whole-community or individual-niche level, whether there is any occurrence of temporal
intransitivity (i.e., where an earlier-evolved type displaces a later-evolved one), and also whether mass
extinctions can alter these kinds of evolutionary and population dynamics.
2 Methods
2.1 Experimental Platform
We used the digital evolution platform Avida [44] as our experimental platform, allowing for a com-
plete record of the course of evolution and precise manipulation of environmental parameters. An
Avida world consists of a grid of digital organisms (hereafter referred to as Avidians), each of which
consists of a genome of instructions written in an assembly-like, Turing-complete computer language,
along with a simple virtual computer chip that executes those instructions. Avida has been detailed at
length elsewhere [44], as has our experimental setup [38, 59, 62], but some details of our implemen-
tation deserve mention. First, the ecosystem has a limited amount of space, introducing an element of
drift as new organisms displace the old. Second, the environment features multiple depletable re-
sources, linked to certain computations, where a low concentration of a resource reduces the benefit
gained by performing an associated computation. This feature mimics density-dependent competition
for resources, favoring organisms that most efficiently consume particular resources or target under-
utilized ones. Resources are globally available to all organisms, with no spatial structure. Only a limited
number of resources are supplied exogenously, while the remainder can only be generated as by-products
by organisms when they successfully complete certain associated computational functions (these inter-
actions are discussed at greater length in Supplement A, and illustrated in Supplementary Figure S1,
which can be found in the online supplementary materials for this article at www.mitpressjournals.
org/doi/suppl/10.1162/artl_a_00305). This introduces ecological interdependence into the population
dynamics, which is a feature of real communities. Configuration files for performing the experiments
described here are available at doi:10.5061/dryad.67506q4.
We used two different versions of Avida for the work described here. The base communities
were generated using v. 2.4.4, while all competition trials and fitness assays were conducted with
v. 2.12. The source code for each version is available from https://www.dropbox.com/home/
Luo_etal_ALIFE_SourceCodes.
2.2 Experimental Methodology
2.2.1 Mass Extinction Treatments
We have previously used Avida [38, 59–62] to investigate the effects of two general types of mass
extinction—press and pulse—on recovery dynamics and community composition. A pulse extinction
is an instantaneous mass culling of individuals from the population, with survivors selected at random
and providing no opportunity for adaptation; by contrast, a press extinction involves a period of altered
environmental conditions—in this case, greatly reduced inflows of basal resources—that persists long
enough to allow an adaptive response in the population. Each of these two treatment types was applied
at two different strengths, Strong and Weak, along with a Control treatment in which evolution continued
uninterrupted. Details of the treatment regimens have been described at length in the aforementioned
publications, and are summarized in Supplement A in the online supplementary materials.
2.2.2 Generation of Ecologically Stable Communities
The base communities are those used in previous work [38, 59, 62]. Avida communities typically
contain many hundreds of genotypes and dozens of Avidians with distinct functional combinations.
198
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
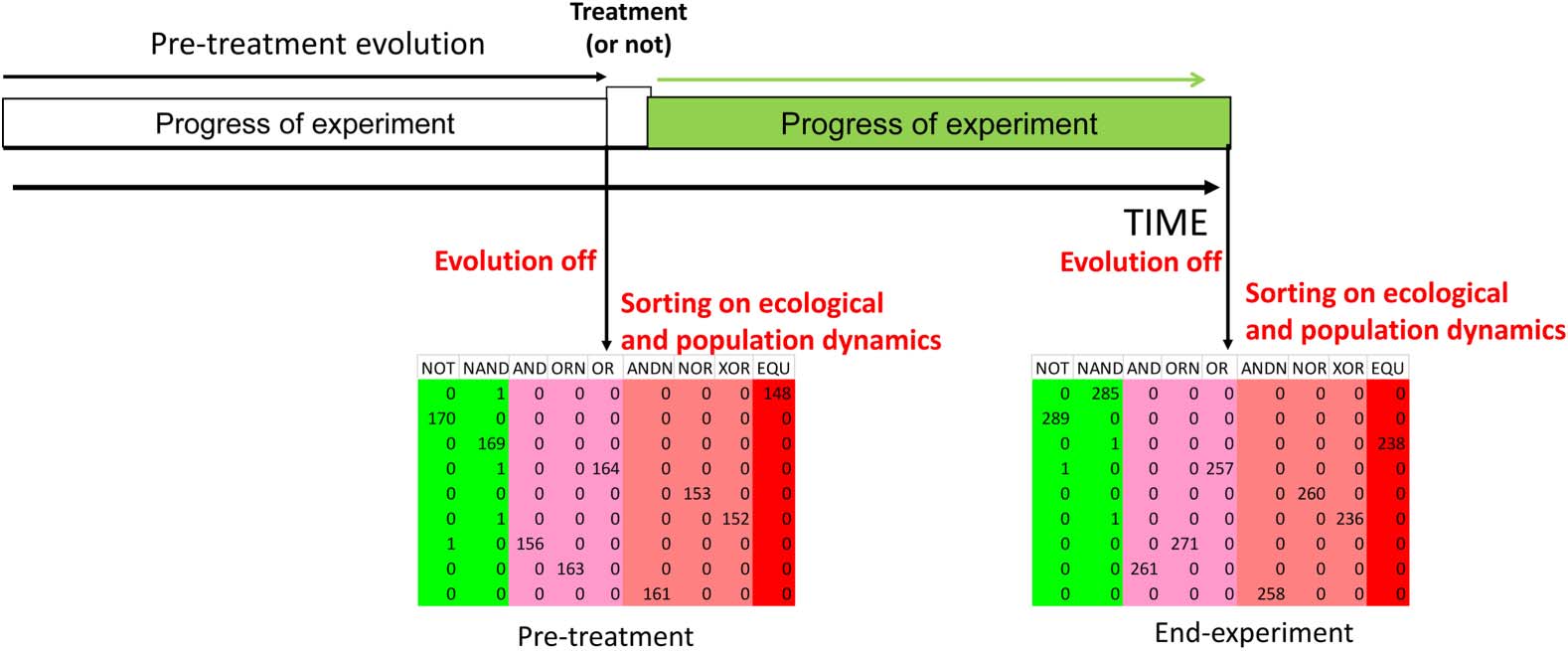
Figure 1. Schematic depiction of generation of ecologically stable communities. After an initial period of evolution and
diversification, all evolutionary forces are stopped, allowing communities containing thousands of genotypes to sort on
ecological and population dynamics to produce an ecologically stable community. Such communities may be produced
before or after a treatment entailing a major macroevolutionary event. Each row in the community represents a single
Avidian, while the number in each cell shows how many times the Avidian performs the computational function (column
headings) during the course of a single gestational cycle.
To enable tractable competition experiments, we generated ecologically stable communities accord-
ing to previously described methods [12, 15]. Briefly, Avida experiments were seeded and run as
described previously, but, at specific times, all mutation was turned off, allowing the extant geno-
types to sort on ecological and population dynamics for an additional 100,000 Avida updates. This
additional step resulted in a greatly reduced set of Avidians that could coexist with minimal ecolog-
ical overlap, representing the ecological “elites” of the population at the time evolution was halted
(Figure 1). Each unique Avidian with a particular ecological profile (computational functions per-
formed and corresponding resources used) is an ecotype. We generated such stable communities for
the following times:
100,000 updates (representing the immediate pre-treatment state)
Control: 105,000; 115,000; 130,000; 155,000; 180,000; 205,000 updates
Press treatments: 205,000 updates
Pulse treatments: 200,000 updates
2.2.3 Initial Assessment of Fitness in Stable Communities
We first used these ecologically stable communities to derive a measure of fitness for each compo-
nent ecotype. We did not use the native measure of fitness in Avida, which is more like a metabolic
rate. Instead, we opted for one more akin to what is used in real microbial evolution experiments, a
reproductive rate. For each pre-treatment community, we conducted fitness assays in which we
seeded an Avida population grid with a single individual of each stable ecotype at particular fixed
positions, with ample space around each individual to avoid early extinction of ecotypes. Each eco-
type was given a specific lineage label that would be carried by all descendants of that particular seed
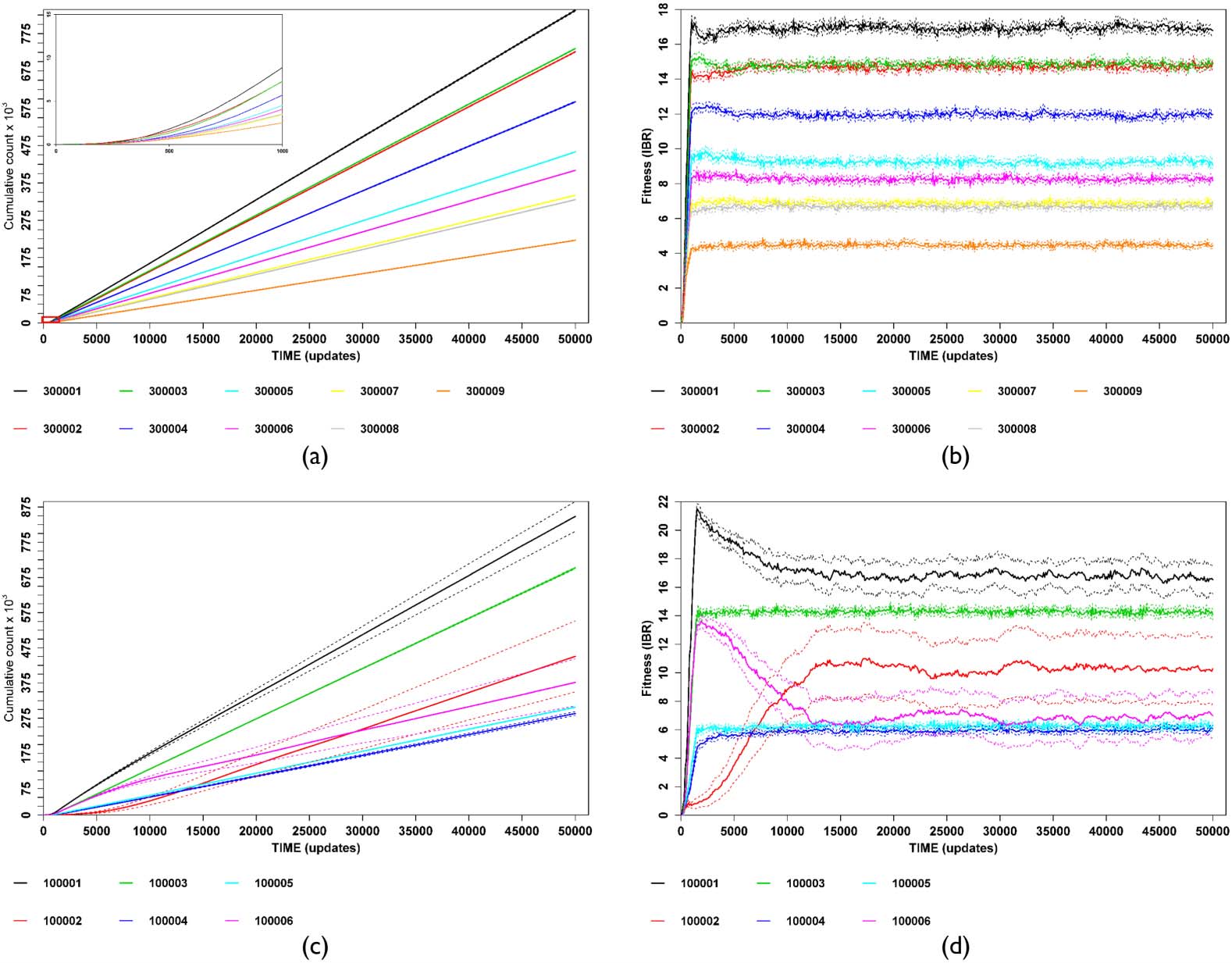
individual. For each assay, we recorded the total number of births (cumulative population count, or
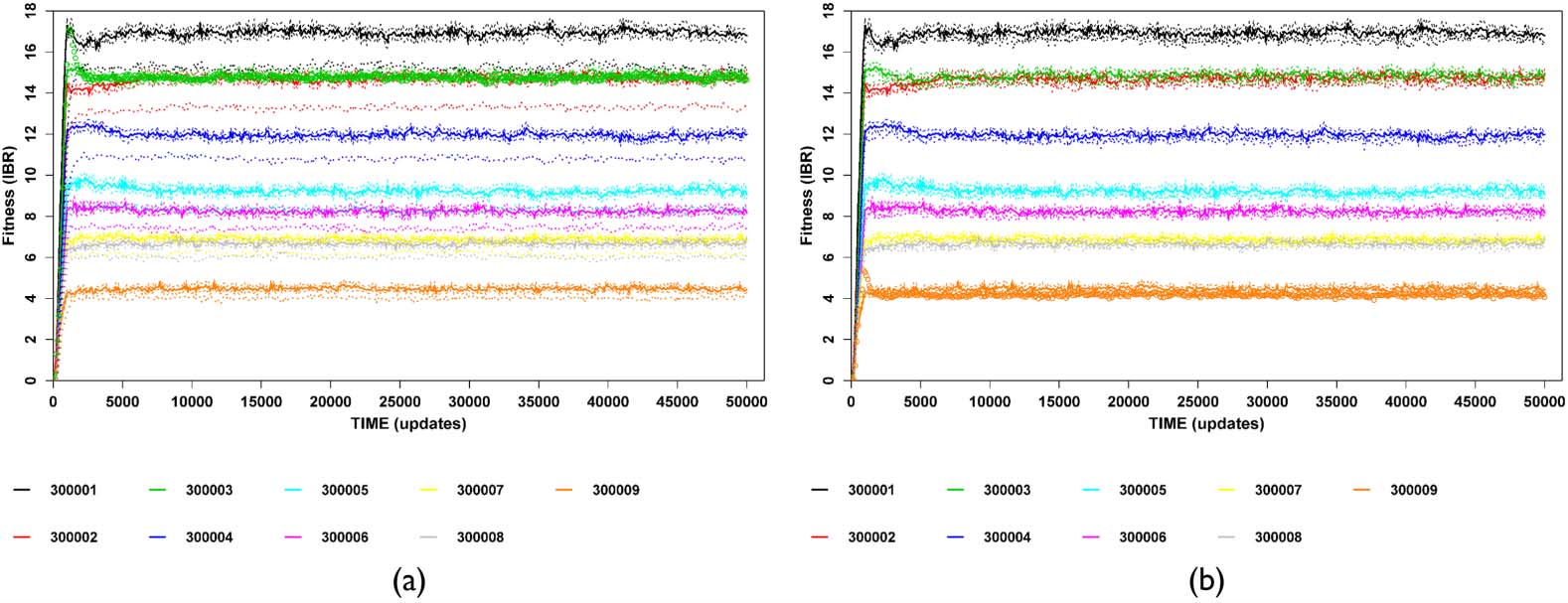
CPC [14]) of each labeled ecotype for 50,000 Avida updates (Figures 2(a), 2(c)). Each ecotypeʼs CPC
data were then converted into instantaneous birth rate (IBR, i.e., the first derivative of the CPC with
respect to time, see Figures 2(b), 2(d)), which was subsequently analyzed with breakpoint analysis
[64, 65] in order to determine the initial increase and equilibrium phases. The measure of fitness was
the average of all equilibrium-phase IBR values from the last breakpoint to the end of the data series.
Artificial Life Volume 26, Number 2
199
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Illustration of conversion of cumulative population count (CPC) for each stable ecotype into instantaneous
birth rate (IBR), used here as the measure of fitness. Each colored trace represents a distinct ecotype. Values are av-
erages of ten subreplicates (thick traces) with 95% confidence intervals (thin traces). Panels (a) and (c) show the CPC,
while panels (b) and (d) show the IBR. (a, b) A specialist community with nine stable ecotypes. Inset in panel (a) shows
initial nonlinear growth corresponding to pre-plateau region of (b) (red rectangle in lower left-hand corner ). (c, d) A
more generalized community with six stable ecotypes, three of which (the black, red, and magenta traces) show sub-
stantial ecological overlap and frequency-dependent dynamics. Note the nonlinear CPC traces and the wider 95% CIs for
these ecotypes.
Given the cross-feeding environment (including self-feeding potential), as well as the ability of many
of the tested ecotypes to produce multiple resources, starting each trial with only a single individual
of each ecotype could conceivably result in positively frequency-dependent effects before equilibrium
was attained. Having reached this point, ecotypes with substantial niche overlap would then show
negative frequency dependence (Figures 2(b), 2(d)).
2.2.4 Time-Shift Competition Trials
We next used the ecologically stable communities to perform the following kinds of competition
trials:
a.
200
Full-community competitions. Each trial was seeded with a single individual of each ecotype
represented in each replicateʼs pre-treatment and end-treatment stable communities.
These trials were intended to test for the emergence of long-term community-level stasis
as described in the Introduction, that is, whether the final community would have one
source community completely eliminate the other, or would be a mixture of ecotypes
from each source community, with each component ecotype coming sometimes from
the earlier one and sometimes from the later one.
Artificial Life Volume 26, Number 2
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
b.
c.
d.
Replacement trials. For communities where there was a clear phenotypic correspondence
between the end-treatment and pre-treatment ecotypes, each trial was seeded by
replacing one of the pre-treatment ecotypes with its end-treatment eco-equivalent (a
replacer ), noting what effect (if any) this replacement had on the equilibrium IBRs of the
other Avidians in the pre-treatment community (termed residents). Each end-treatment
ecotype was tested in this way. These trials tested for two aspects of stasis at the level of
ecotype: whether the replacer had about the same fitness as the resident being replaced,
and the replacerʼs effects on the remaining residentsʼ fitnesses. Only 65/500 total
replicates (across all treatments) could be treated as replacements.
Invasion trials. For communities where the end-treatment community contained ecotypes
not present in the pre-treatment community, or vice versa, a single individual of an end-
treatment ecotype was allowed to invade against the whole pre-treatment community,
without a priori removal of any resident. Again, each end-treatment ecotype was
evaluated in this manner. We again noted how the invasion affected the fitnesses of the
other residents, as well as whether any residents were displaced by the invader. 435/500
total replicates were treated as invasions.
Inverse invasion trials. These were similar in nature to the invasion trials, except that they
featured a pre-treatment ecotype invading against the whole end-treatment community.
We performed these trials only for cases where a pre-treatment ecotype was present in
the final community from a full-competition experiment, or where invariance of resident
fitness was suggested by the results of a replacement or invasion trial as described above.
We performed 10 subreplicates of each trial, each of 50,000 updates in duration. The seed
Avidians for each trial were again always placed at particular fixed positions on the Avida population
grid, amply spaced so as to prevent early extinctions by overwrites.
2.3 Analytical Methodology
2.3.1 Determination of Fitness Effects
In order to assess the effects of replacements and invasions on the fitnesses of resident ecotypes, we
used a likelihood ratio test developed previously [38] to compare the average IBR (derived from the
equilibrium IBR time series data) of each ecotype in the absence and in the presence of the replacer/
invader. For each resident ecotype, we first determined the fold difference between the average IBR
with and without the replacer or invader. Each fold difference is interpreted as a coefficient of se-
lection, and was tested for statistical significance through use of a chi-square test to compare the log
likelihoods of a model assuming a difference between the averages of the with and without data
series versus a null model assuming no difference. We then determined a mean fold difference
for each replacer or invader against all (other ) residents. This way of assessing relative fitness is also
akin to the lag-load of [52].
2.3.2 Nomenclature and Terminology
In the results (Section 3), single ecotypes are named by the random seed used to generate their
community, followed by a two-digit number that indicates their time of origin and rank-abundance,
then by one or two letters that indicate which treatment generated them: PT for pre-treatment, C for
Control, SP for Strong Press, SU for Strong Pulse, WP for Weak Press, and WU for Weak Pulse.
Hence, 72000PT refers to the most common ecotype from replicate 7200ʼs pre-treatment commu-
nity, while 720091C refers to the most common ecotype from replicate 7200ʼs end-Control commu-
nity; similarly, 720093SP would be the third most common ecotype from replicate 7200ʼs end-Strong
press community. Identical numbers do not necessarily indicate ecologically equivalent types.
Artificial Life Volume 26, Number 2
201
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
Additionally, the term “intransitivity” should be understood to mean temporal intransitivity, where an
earlier-evolved type outperforms a later-evolved one.
3 Results
3.1 Full-Community Competitions
When pre-treatment communities were competed against end-Control communities, no evidence of
full-community intransitivity was found (summary results in Table 1; example in Figures 3(a), 3(b)).
Only 2/100 replicates (7200 and 8700) showed any evidence of intransitivity at the level of single
ecotypes, with a single pre-treatment ecotype surviving to the end of the trial at greater frequency
and higher fitness than the end-treatment eco-equivalent. Results for the two Weak treatments were
similar, with only 2/100 and 4/100 replicates for Weak Pulse (2400, 7200) and Weak Press (2400,
3100, 8700, 10000), respectively, showing any evidence of single-ecotype intransitivity. In all these
cases, however, the surviving pre-treatment ecotype was present at lower fitness in the mixed pop-
ulation than in its own fully native community.
The two Strong treatments showed markedly different results from Control and Weak treat-
ments. Although there were again no cases of full-community intransitivity, an order of magnitude
more replicates showed evidence of single-ecotype intransitivity. When communities that were not
ecologically complete before the treatment were removed, Strong Press still showed the greatest
number of replicates with single-ecotype intransitivity, followed by Strong Pulse. Even in these
cases, there was a strong tendency for the carryover ecotypes (pre-treatment ecotypes that were
present in the mixed community) to show lower fitness in the final mixed communities than in their
own fully native conditions (Table 2, Figures 3(c), 3(d)). It was much less common for carryovers to
have about the same fitness as in native conditions, and rare for their fitness to actually be higher
(Table 2). Many communities were ecologically incomplete because the difficult XOR and EQU func-
tions did not evolve, or, more often, failed to re-evolve after a strong mass extinction [60–62]. Even
in these last cases, carryovers—the pre-treatment XOR- and EQU-bearing ecotypes—tended to have
lower fitness even when they could just “slot in” to empty niches without competition (Table 2).
We next conducted a series of post hoc invasion experiments to determine the strength of the
intransitivity for ecotypes from the aforementioned Control competitions. Replicate 8700 showed
Table 1. Summary for full-community competitions. Data for means are across all 100 replicates for each treatment ± 2 s.e.
Mean no. of
ecotypes in
final community
Mean no. of
ecotypes from
pre-treatment
community
Mean no. of
ecotypes from
end-treatment
community
No. of replicates
where both communities
are ecologically
complete (/100)
Treatment
Control
5.97 ± 0.235
0.02 ± 0.028
5.95 ± 0.232
Strong Press
6.45 ± 0.238
1.62 ± 0.27
4.83 ± 0.283
Strong Pulse
6.12 ± 0.256
0.63 ± 0.179
5.49 ± 0.253
Weak Press
6.08 ± 0.221
0.06 ± 0.056
6.02 ± 0.222
Weak Pulse
5.99 ± 0.245
0.04 ± 0.056
5.95 ± 0.238
85
56
74
85
84
No. of replicates where
both communities are
ecologically complete
and at least one
pre-treatment
ecotype is present
in final community
2
40
25
4
2
202
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
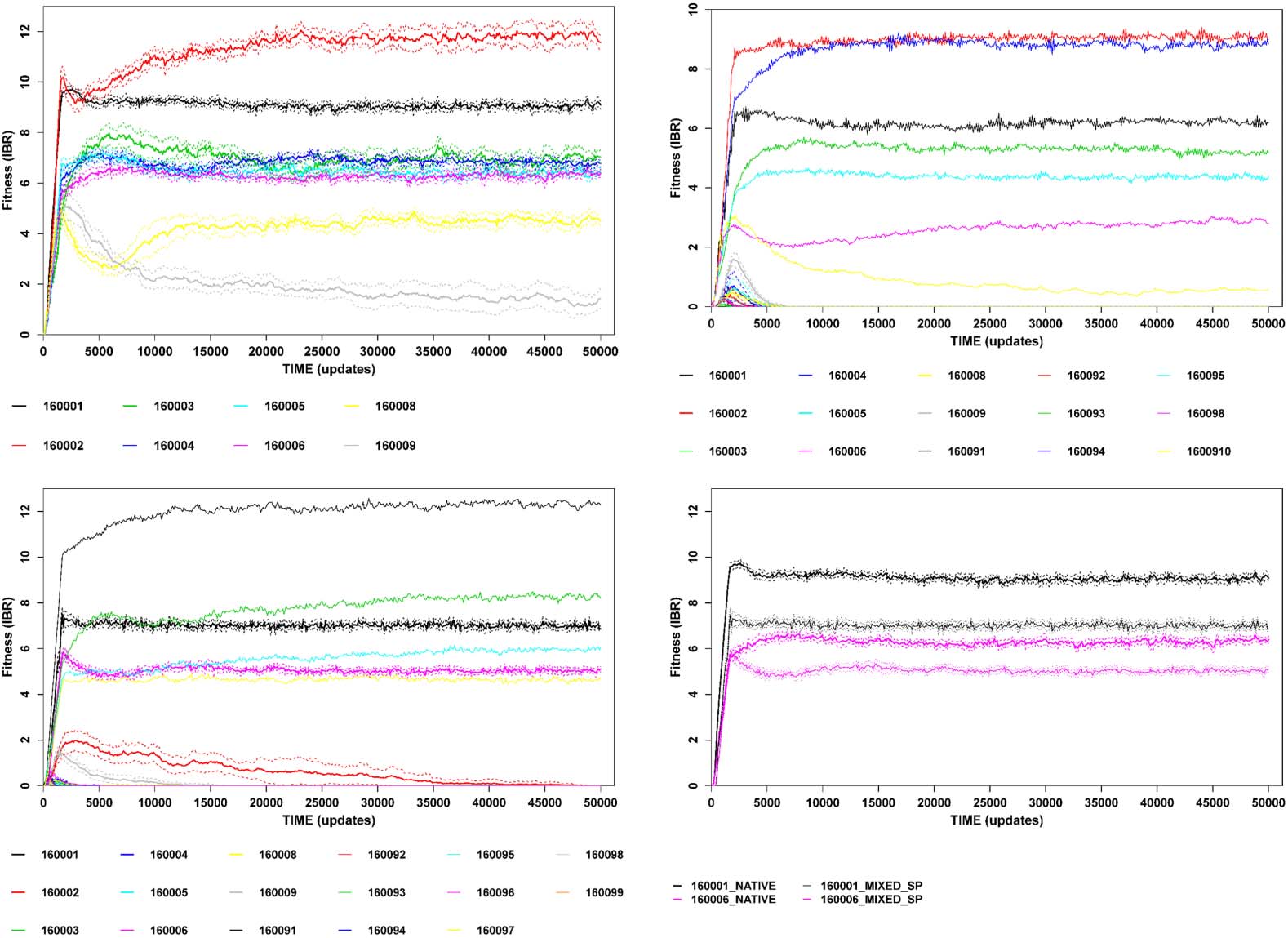
Figure 3. Example of partial evolutionary intransitivity in full-community competitions. All traces are means of ten sub-
replicates. Error series (where present) are 95% confidence intervals around means. (a) IBR plot for a community (rep-
licate 1600) with eight stable ecotypes. (b) Result of full-community competition for 1600PT (thick traces with errors)
versus 1600C (thin traces with no errors). All pre-treatment ecotypes are driven extinct rapidly. (c) Result of full-
community competition for 1600PT (thick traces with errors) versus 1600SP (thin traces with no errors). Eight eco-
types are present in the final mixed community, two of which are carryovers from the pre-treatment community
(note also long-term persistence of a third pre-treatment type before it goes extinct ). (d) Comparison of IBR for
carryover ecotypes in fully native conditions (thick traces with errors) versus mixed-community conditions (thin
traces with errors). The mixed-community fitnesses are well outside of the 95% CIs of the fully native values.
total intransitivity, as ecotype 870001PT always survived, while its end-treatment equivalent
870091C always went extinct. In replicate 7200, however, the intransitivity was partial, as ecotype
720001PT always survived, but its end-treatment equivalent 720092C did not always go extinct,
although the latter was always present at lower frequency and fitness, where it too survived in
the final mixed community.
The ecotypes that displayed intransitivity had little or no change in genome size (although there
had been a great deal of sequence evolution) and tended to be quite ecologically generalized, albeit
with low levels of functional expression (no function was performed more than ten times during a
single gestational cycle). An interesting observation about the type that showed complete intransi-
tivity was that the corresponding end-Control ecotype was actually missing one function, still present
in the pre-treatment version, which would have allowed it to be fully self-feeding. Instead, it was
rendered dependent on by-products from other community members in order to achieve its full
fitness, while the pre-treatment version was fully self-feeding.
3.2 Replacement Trials
In replacement trials, where a single pre-treatment ecotype was replaced by a clear future eco-
equivalent, most such replacements resulted in substantial depression of the pre-treatment residentsʼ
Artificial Life Volume 26, Number 2
203
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
Table 2. Pre-treatment carryovers in full-community competitions and their fitness in mixed versus fully native
conditions for Strong Press and Pulse extinctions.
Number of
pre-treatment
carryover ecotypes
(total |
mean ± 2 s.e.)
Number of carryovers
with lower fitness
than native conditions
(total | % of total
carryovers |
mean ± 2 s.e.)
Number of carryovers
with higher fitness
than native conditions
(total | % of total
carryovers |
mean ± 2 s.e.)
Number of carryovers
with approximately
same fitness as
native conditions
(total | % of total
carryovers | mean ± 2 s.e.)
Treatment
(No. of
replicates)
Both communities ecologically complete
Strong Press (40)
87 | 2.175 ± 0.319
69 | 79 | 1.725 ± 0.27
6 | 7 | 0.69 ± 0.114
12 | 14 | 0.3 ± 0.19
Strong Pulse (25)
38 | 1.52 ± 0.367
34 | 89 | 1.36 ± 0.303
1 | 3 | 0.04 ± 0.08
3 | 8 | 0.12 ± 0.13
One or both communities ecologically incomplete
Strong Press (35)
73 | 2.086 ± 0.397
54 | 74 | 1.54 ± 0.361
4 | 5 | 0.114 ± 0.109
15 | 21 | 0.428 ± 0.29
Strong Pulse (18)
26 | 1.444 ± 0.29
23 | 88 | 1.11 ± 0.454
1 | 4 | 0.056 ± 0.111
2 | 8 | 0.11 ± 0.15
fitnesses. Out of all replacement tests (65/500 total replicates across all treatments), only three (one
replacer each from 2/20 Control trials and three replacers from a single Strong Press trial) showed
any evidence of long-term stasis in a particular ecotypeʼs fitness, and not for all replacers. In only
these particular cases, the replacer had about the same fitness as the replaced resident, and negligible
effects on the remaining residents.
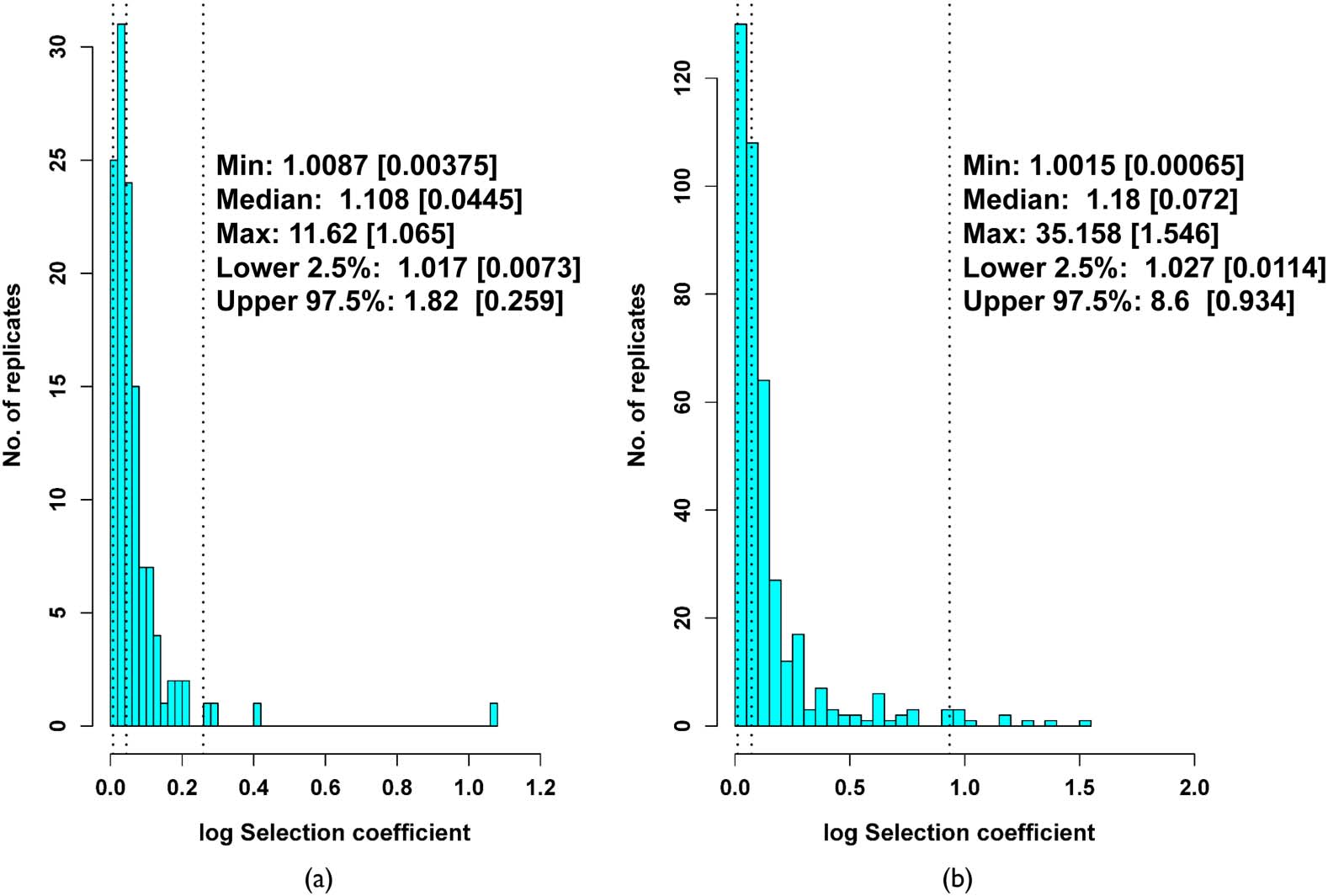
Across all replacement trials, the range of fitness effects as expressed by the IBR-based selection
coefficients (see Section 2) was extremely wide, as low as 0.87% (i.e., a less than 1% difference in
fitness) and as high as 1161% (Figure 4(a)). Within these extremes, the median change in the selec-
tion coefficient was a reduction of 10.8%, with 2.5 and 97.5 percentiles of 1.7% and 82%. Over the
entire length of the post-treatment period, ecotypes mostly did not remain static in their perfor-
mance, even if their ecological profiles remained unchanged; the replacements usually showed quite
substantial, measurable effects on the performance of the pre-treatment residents. Hence, long-term
evolution in these replacer communities was largely fully transitive. Over shorter time scales, how-
ever, either neutrality (as shown by inconsistent survival of particular ancestral ecotypes across assay
replicates) or intransitivity (consistent survival of particular ancestral ecotypes across assay replicates)
could still be observed (Supplement B in the online supplementary materials).
The outcomes of the full-community competitions strongly suggested that selection coefficients
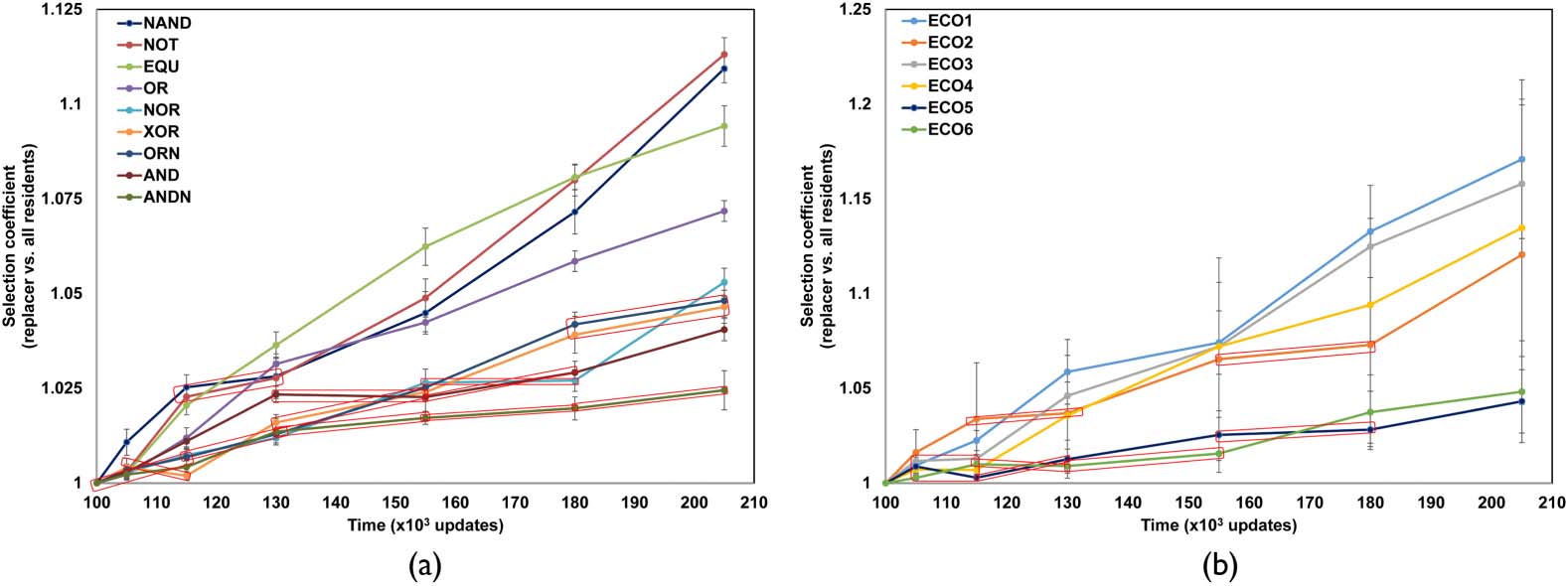
did not remain static over time. We investigated this in more detail by looking at seven case study
communities. Five of these showed both very high ecological and phylogenetic stability as deter-
mined previously [38] and were fairly generalized (6–7 stable ecotypes per community). The other
two were composed of pure specialists (9 stable ecotypes), and showed high ecological stability but
not phylogenetic stability. All chosen communities were also ecologically complete (i.e., all resources
were used). Replacement trials were done using the community states at 105,000; 115,000; 130,000;
155,000; and 180,000 updates as sources of replacers. The effects of replacements are best seen in
the pure specialist communities, where every end-experiment replacer had some measurable effect
on the residents (Figure 5(a), Figure 6). Over the duration of the post-treatment period, there was an
increase in performance for all ecotypes against the residents, but the strength of this increase was
not equal for all of them (Figure 6). Some of them (e.g., the ANDN ecotype) remained closer to
the pre-treatment level of performance, while the performance of others (the NOT, NAND, and
204
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
Figure 4. Distribution of selection coefficients (log-transformed) for invasion and replacement trials for end-treatment
communities versus pre-treatment communities. In both plots, the vertical dotted lines are (from left to right) the 2.5th
percentile, the median, and the 97.5th percentile of each distribution. Values in square brackets are log transformations
to aid interpretation of the x-axis. (a) Replacement trials. (b) Invasion trials.
EQU ecotypes) increased much more over the pre-treatment community (Figure 5(a)). Certain eco-
types (particularly ANDN, AND, OR, XOR) did not evolve much over certain time periods (Figure 5(a),
highlighted with red ovals), but others did during the periods of stasis for these types, and any ob-
served periods of stasis for any particular ecotype were not permanent. Further, certain replacers had
strongly negative effects on all other residents, reducing fitness well outside of the native range of
variation (Figure 6(a)), while others had weaker effects, reducing fitness only to around the lower
end of the native range (Figure 6(b)).
In contrast, the more generalized communities showed different behavior. While the overall trend
of increasing coefficients of selection was again evident, several ecotypes showed enormous varia-
tion around each estimate (Figure 5(b)). Unlike the pure specialist communities, certain replacers
tended to have extremely strong effects on one to three residents, and much weaker effects (if
any) on the others, sometimes to the extent of driving another resident extinct (even if that replacer
originated before the end of the experiment, as in Supplement B, in the online supplementary ma-
terials). Further, certain replacer ecotypes had only weak effects against all other residents over the
entire post-treatment period, acting largely like the residents they replaced. We also found some
cases where addition of the replacer actually increased the fitness of one or more residents while
decreasing the fitness of others; this was not observed in either of the specialist communities, where
fitness effects were always negative, even if the decrease was not significant.
3.3 Invasion Trials
The invasion trials, where a single end-experiment ecotype (a future invader, or FI) attempted to in-
vade against all of the pre-treatment residents, showed an even wider range of outcomes than did the
Artificial Life Volume 26, Number 2
205
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
Figure 5. Increase in coefficients of selection (see Section 2 for definition) over time for replacer ecotypes against pre-
treatment residents. In both plots, each trace represents the change in average selection coefficient for each replacer
ecotype (shown in legend) against all non-equivalent pre-treatment residents. Each value is the mean of all coefficients
across every resident for that ecotype. Errors are 95% confidence intervals. Red ovals indicate time periods where the
change in selection coefficient is strictly less than 1.00 percent. (a) A pure specialist community with nine stable ecotypes
where each Avidian specializes on one of the nine available resources and performs one of the nine standard Avida logic
functions (shown in legend). (b) A more generalized community with six stable ecotypes where each Avidian performs
two or three functions. Note the much larger errors associated with each estimate compared to panel (a).
replacement trials (Figure 4(b)). In these less ecologically stable communities, we observed a higher
incidence of invariance in resident performance (Table 3).
As with the replacements, residents usually showed a measurable reduction in fitness as a result
of the invasion (median 18%, 2.5 percentile 2.7%, 97.5 percentile 860%, Figure 4(b)). The most
common outcome was that a FI displaced at least one resident (although it was sometimes two
or more) and had strong depressive fitness effects on at least one other resident. However, there
was very wide among-resident variation in the strength of these effects; typically, a FI would have
strong effects on only one or two residents in a community, and much weaker or negligible effects
on others. This was true across all treatments, but was particularly noticeable for Strong Press and
Figure 6. Fitness assay plots for the specialist community (Figure 5(a)), showing differences in fitness change using dif-
ferent end-treatment replacer ecotypes. Each ecotype is represented by a different color. In both plots, the pre-treat-
ment residentsʼ native fitnesses (averaged over ten subreplicates) are shown by the thick traces with thin error series
(95% CIs), while their fitnesses in the presence of the replacer are shown by dotted traces. The fitness of the replacer is
shown as a trace of open circles, colored the same as the replaced resident. (a) Replacer is the end-control NAND eco-
type (colored bright green). The other residentsʼ equilibrium fitnesses are reduced by an average of ≈10.9%. (b) Replacer
is the end-control ANDN ecotype (colored orange). The other residentsʼ equilibrium fitnesses are reduced by an average
of ≈2.45%.
206
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
Table 3. Summary data for resident performance stasis in invasion trials.
Proportion of replicates
showing resident stasis
(total no. of replicates treated
as invasion trials)
Mean no. of invaders with
no significant effect on
residents (± 2 s.e.)
Proportion of invaders that
have no significant effects on
residents in the most
stable community
Control
Weak Pulse
Weak Press
Strong Pulse
Strong Press
4/80
7/79
4/81
26/96
41/99
1.25 ± 0.5
1.14 ± 0.286
1 ± 0.00
1.42 ± 0.297
2.024 ± 0.346
2/7
2/8
1/5
4/6
5/7
Strong Pulse (Table 3). We did not find any cases in which every tested invader failed to affect the
residentsʼ performance.
In a number of cases, the results of the invasion trials conflicted with those of the corresponding
full-community competitions, where particular FIs failed in every invasion attempt against the pre-
treatment community when tested individually, yet were present in the final community in the full
competition. We therefore performed a number of post hoc co-invasion trials, where one of these
failed FIs, or a replacer with a very weak fitness effect (1% or less), was allowed to invade accom-
panied by a second FI or replacer from the same community that was already determined separately
to have strong fitness effects on at least one member of the pre-treatment community. In every such
case, the double invasion succeeded, with both FIs or replacers establishing themselves and displa-
cing one or more residents.
3.4 Inverse Invasion Trials
The purpose of these trials was to confirm whether pre-treatment ecotypes that survived in the full-
community competitions could actually invade the end-treatment community. The results matched
the outcome of the full-community competitions almost perfectly, in that both series of experiments
ended with almost the same mixed community for each replicate. However, even here there were
two notable exceptions: Ecotypes 190004PT and 290004PT survived in the full-community com-
petitions, but always failed as single invaders of their Strong Press end-treatment communities.
4 Discussion
4.1 The Significance of Paleoecological Stasis, and the Need to See Through It
Paleontological examination of the fossil record has revealed the apparent stasis in morphology of a
number of single evolutionary lineages, and even whole paleocommunities, over significant spans of
geological time [26, 37]. It is unclear whether this stasis is more under abiotic or community-level
control, as theoretical and simulation studies allow for the emergence of evolutionary stasis depend-
ing on the strength and type of interactions among community members irrespective of abiotic con-
ditions [43, 52]. Further, it is unclear whether morphological stasis implies stasis in performance,
particularly in fitness, especially when the biotic context around the static lineages has changed
due to macroecological shifts, for example, survivors of a mass extinction existing among taxa that
diversified following the extinction. In this study, we used full-community competitions and time-
shift replacement and invasion trials to determine whether ecologically diversified communities of
Artificial Life Volume 26, Number 2
207
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
digital organisms that showed stasis based on ecological and phylogenetic data [38] were truly evo-
lutionarily static in terms of performance.
4.2 Ecological or Genealogical Stasis Does Not Mean Whole-Community
Stasis in Performance
Our primary interest here was to determine whether these digital communities showed performance
stasis in addition to the stasis implied from ecological and phylogenetic data. If this were in fact the
case, predictable consequences would follow (see Introduction). Our data show that for these digital
communities, these predictions were largely not fulfilled. In the absence of major ecological or pop-
ulation disruption, long-term evolution in this system appears to be almost completely transitive,
with later-evolved communities usually eliminating all members of an ancestral community, with
only rare intransitive exceptions. Even in cases of long-lasting ecological associations among com-
munity members (the appearance of coordinated stasis), performance does not remain constant over
long spans of time, though different communities appear to evolve at different rates (Figure 5 and
Supplement B in the online supplementary materials). This means that even when genealogical con-
tinuity between ecotypes can be established [38], the later versions are not truly the same as the
ancestral versions, despite niche persistence. While this conclusion holds for our Control and Weak
treatment communities, it is much less certain for the Strong treatment communities. Strong Press in
particular showed the highest amount of intransitive outcomes for the full-community competitions,
as well as the highest incidence of resident invariance in the invasion trials. Post-treatment ecotypes
were not necessarily superior to pre-treatment ones, as shown by the surprisingly high number of
outcomes where at least one pre-treatment ecotype was present in the mixed population rather than
a post-treatment one. These results show that a strong enough disruption, particularly one that rad-
ically reshapes a community at levels going all the way from ecological relationships to genomic
characteristics [38, 59, 60], can break the inherent transitivity of an evolving community. Post-treatment
ecotypes that evolve in these new ecological and genetic contexts, even if they successfully occupy a
particular niche and fit well with the rest of the post-treatment community, may in fact be less
effective than that nicheʼs previous occupants. This is normally difficult (if not impossible) to assess
when those previous occupants have been rendered completely extinct (as happened in the original
full experiment), but here we have effectively resurrected them and so the sensitivity of their post-
treatment successors may be demonstrated.
Finally, with regard to the issue of coordinated stasis, we failed to find any examples that showed
such for performance. Although our survey was hardly exhaustive, the most promising candidates that
showed high ecological and genealogical stability (Figure 5(b), Supplement B in the online supplemen-
tary materials) did not show any periods where performance remained static for fitness effects against
all resident ecotypes in a community. If even these best candidates failed to show coordinated stasis of
performance, it is unreasonable to expect it from other replicate populations that were demonstrably
unstable, as shown by shifting ecotype composition and division of labor [38]. Further, competitions
with sequentially occurring communities showed shifting bouts of short-term neutrality and longer-
term transitivity and intransitivity (Supplement B in the online supplementary materials). We conclude
that while such stasis can occur in our experimental system, it is impermanent and uncoordinated:
certain community members may remain static in performance for a particular time period, while
others evolve substantially over that same time, but the community as a whole is not static despite
outward appearances and does not represent an optimal fixed point [13, 45].
4.3 Fitness Is Highly Dependent on Community Context
Our results also demonstrate the extent to which an ecotypeʼs performance is affected by the other
community members around it. Several sources demonstrate this property for our digital commu-
nities. The first is the strong tendency for pre-treatment carryovers to have lower fitness in mixed
communities also containing post-treatment ecotypes, which obtained with both ecologically com-
plete and incomplete communities (Table 2). The second is the replacement and invasion trials,
208
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
where the great majority of fitness effects on residents by the invaders were negative (Table 2,
Figure 4), and which sometimes resulted in failure to invade by ecotypes that were successful in
the corresponding full-community trials, indicating that the presence of other ecotypes from their
native communities was necessary for their success. While the great majority of fitness effects
were negative, we did observe some cases where a replacer or invader increased the fitness of
one or more residents (while having negative or negligible effects on others). This was seen mostly
in communities where there was evidence for negative frequency dependence of interactions among
two or more residents with a substantial degree of niche overlap (e.g., Figure 2(d)). We see two
possible explanations for such results: (i) the replacer or invader affects one of those residents neg-
atively, alleviating frequency-dependent interactions, or (ii) the replacer or invader actually provides a
direct boost to the fitnesses of those positively affected residents through enhanced provisioning of
resources (relative to the replaced resident). However, we cannot currently distinguish between these
two possibilities.
While it is often politely acknowledged that a quantitative assessment of “fitness” (understood
here as a rate of reproduction) applies only under particular conditions (e.g., [8]), the contribution of
other community members to that determination often remains underappreciated. In our cross-
feeding communities, Avidians serve as resource transformers, and the effectiveness with which a
particular ecotype fulfills that role will play a large part in determining the response, and ultimately
the evolutionary characteristics, of other ecotypes that depend on those resources. Substituting an
ecotype with different such effectiveness (either by direct manipulation, or by evolutionary change)
will alter the responses, performance, and potentially the evolutionary trajectories of other ecotypes
in the community. Hence, in communities with these kinds of interactions, fitness (however mea-
sured) becomes an increasingly nebulous concept when every community memberʼs characteristics
depends on the performance and relative abundance of every other member.
4.4 The Queen and the Jester Rule Together
One of the long-standing issues in our exploration of evolution of cross-feeding digital communities
and how mass extinction and recovery affect them [38, 59–62] has been whether new and/or im-
proved ecotypes attain ecologically prominent positions through community drift effects [23, 37] or
Red Queen effects resulting from interactions among community members [18, 36]. Our data in-
dicate that in this system, it is the latter. The results of the full-community competitions and replace-
ment/invasion trials clearly show that without a major abiotic disruption, evolutionary transitivity
usually prevails. The more detailed trials (Figure 5 and Supplement B in the online supplementary
materials) demonstrate that the fitness effects of evolved ecotypes on the older pre-treatment res-
idents tend to increase over the length of the post-treatment period. Further, these effects are gen-
erally negative (though we did observe exceptions), measurably depressing the performance of at
least one resident. Our results lend support to the original, most general formulation of the Red
Queen hypothesis, which can apply to any system of interactors where evolutionary effects are me-
diated by ecological interactions [36, 55]: Evolutionary advance in one community member that in-
teracts ecologically with several others will result in deterioration of the fitnesses of those members,
which must continue evolving or go extinct.
Although all our trials were run under ecological conditions (with no new types allowed to
evolve), we argue that we still observed these predicted effects, though mostly indirectly. First,
we observed a preponderance of negative fitness effects, which generally increased in strength over
time. Second, we observed in the replacement and invasion trials the occasional secondary extinction
of pre-treatment residents whose ecotypes both did not go extinct and remained ecologically prom-
inent in the original evolutionary experiment, demonstrating that there was indeed evolutionary ad-
justment for these ecotypes during the post-treatment period (compare with [32]). Third, in the
sequential full-community competitions (Supplement B in the online supplementary materials),
the particular ecotypes that showed occasional intransitive effects sometimes changed between
the assayed time points, indicating that there was actual coevolution between ecotypes in these com-
Artificial Life Volume 26, Number 2
209
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
munities. These effects also help explain the results of the full-community competitions. In the re-
placement and invasion trials, each replacer or invader usually negatively affected at least one par-
ticular resident; hence, the older community is driven extinct not just because the later one is
composed of superior competitors for all available niches, but also because most members of that
later community may exert indirect negative effects on older ones through altered ecological inter-
actions. We draw these conclusions primarily from the Control experiments (where evolution con-
tinued uninterrupted), but they also apply to the Weak treatments, which produced very similar
results. However, the results of the Strong treatments seem to require a different explanation, given
the fairly frequent occurrence of mixed communities containing at least one pre-treatment ecotype
(Table 1), and the increased incidence of invasion trials where the invader had negligible effects on
performance of the remaining residents (especially in Strong Press), even if at least one resident was
eliminated (Table 3).
We believe that these results argue for a role of mixed Red Queen–Court Jester dynamics in
shaping the post-treatment communities [9, 19, 24, 25]. The pre-treatment communities are heavily
compromised or wiped out by massive abiotic forcing [38, 59, 62], allowing for redirection of evo-
lution and breaking of the prevailing dynamics that would otherwise yield a transitive outcome. The
communities that emerge in the aftermath of the Strong extinctions are made up of Avidians that
often have altered ecotype composition and functional properties, and whose ecological structure
may pass through a number of transitional stages during recovery [38]. Although we have not looked
directly at community web structure [30, 48] or ecological connectance [5] here, the greater inci-
dence of resident invariance when using post-Strong-treatment invaders suggests that those invaders
interact only weakly with the residents, implying that the post-treatment ecological structures from
which they are drawn differ substantially from those of the pre-treatment residents. This outcome
was already suggested by larger changes in the degree of a communityʼs division of labor after Strong
treatments [38]. Put another way, the lack of fitness effects on residents seen using these invaders is
paradoxically not because the invaders fit seamlessly into the remaining residentsʼ ecological network,
but because they are actually more ecologically disconnected from it. (This result should not be
overemphasized, though, as there are still plenty of post-Strong-treatment invaders that do produce
adverse effects on the residents.) We say this because the results from the Control and Weak ex-
periments (where resident invariance is much less common) show what happens when such a net-
work is invaded by a more evolved ecotype that matches the ecological profile of the replaced
resident, and that ought to fit into the network the same way. The change in post-treatment eco-
logical structure is also implied by the occasional persistence of pre-treatment ecotypes in the full-
community competitions, indicating that the structure of post-treatment communities is sometimes
less robust than the corresponding pre-treatment communityʼs.
4.5 Parallels with and Applications to Real Biological Systems
We have previously compared the broad qualitative similarities of the ecological and phylogenetic
aspects of our results to major extinction and recovery events from Earth history [38, 59, 61]. Our
present results are of general interest only if they are not system-specific. However, we argue that
these results too have broad qualitative parallels with diverse systems at different levels of real-world
biology and ecology.
1. “Softening up” of communities by strong invaders. Although uncommon among our
results, we still observed several incidents of replacers or invaders failing when attempting
invasion individually, but present in the mixed communities of full-community
competitions, and able to invade when accompanied by a co-invader with strong
depressive fitness effects on at least one resident. This result is broadly analogous to what
is expected under the invasion meltdown hypothesis [51], where invasive species with
strong ecosystem engineering capabilities can alter ecological dynamics in the recipient
community, rendering them vulnerable to further invasion by species that would not
210
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
otherwise succeed. While our experiments were replacements and invasions across time
rather than space, we are confident that similar dynamics would occur using spatially
separated, independently evolved communities that do not include the competing
residents as part of their own evolutionary history [57, 58].
2. Facilitation facilitates invasion. Related to the previous point, our cross-feeding
environmental setup only allows for facilitative resource conversion: Consumed resources
are transformed into by-products that then become resources themselves, but only
have positive effects on Avidians that consume them (Supplement A in the online
supplementary materials). However, these kinds of facilitative dynamics may also result in
increased susceptibility to invasion [3, 34, 54], which in the present context may also
contribute to the relative paucity of stable results for most of our digital communities [38].
Our model does not include direct negative interactions such as predation, parasitism, or
allelopathy, though such interactions are in principle possible with this digital system [10,
21, 63] and thus present intriguing possibilities for future work, particularly if inclusion of
them enhances ecological and evolutionary stability [41, 42, 50].
3. Microbial evolution population dynamics. The conventional paradigm that emerged for
interpreting the results of early microbial evolution experiments emphasized a periodic
selection process where successive, cumulative substitution of beneficial mutants in a
continuous lineage ought to produce a quite deterministic evolutionary process that agreed
well with theoretical predictions. Some experiments produced results highly inconsistent
with this expectation [1, 2], leading to an alternative interpretation of chemostat
populations as mixtures of genotypes that could possibly modify culture conditions and
alter the relative fitnesses of others [7]. This has been demonstrated directly even for serial
transfer experiments started from single clones in which different ecotypes emerge in both
mixed [11, 49, 53] and static cultures [47]. Here, we have shown that the fitness of any one
type is dependent on the characteristics of other types, and evolutionary advances in one
particular lineage can alter the fitnesses of coexisting lineages that remain invariant. Results
from our replacement and invasion trials involving extinction of resident ecotypes that
persisted throughout the source experiment are qualitatively comparable to those shown
previously for a two-ecotype microbial model system [32]. In this latter case, early clones
of a second ecotype failed in reciprocal invasion experiments against later-evolved clones
of the first ecotype, yet this second ecotype persisted across the original experiment,
demonstrating that it did evolve to avoid extinction in the intervening time. We believe
our results will apply to any evolving, closed-community system (see [56] for a critique of
the generality of the assumptions under which Red Queen dynamics will emerge) that
develops some degree of niche differentiation and/or capability for niche construction,
regardless of whether they are in chemostat-like or serial-transfer contexts.
4. Negative effects on performance of pathways by temporally displaced members. While we
have treated the Avidian communities as analogues for real natural communities, they may
also be seen as analogous to enzymes in a metabolic pathway, and the virtual metabolism
of resources as analogous to the modification of substrates by one enzyme, which are then
passed to the next one in the pathway. (The analogy here is not entirely accurate, because
the Avidians often use more than one resource and may consume their own by-products.)
It has recently become possible to produce reconstructed ancestral sequences of particular
enzymes, insert them into extant living cells, and analyze the physiological and
evolutionary responses of the unmodified remainder of the pathway [28, 29]. An inferred
ancestral variant of the essential gene tufB (which in bacteria encodes elongation factor
Tu), when inserted into a modern E. coli cell, was not immediately lethal, but did cause a
twofold decrease in fitness due to reduced protein dosage. This resembles the reduction in
fitness seen in multiple ecotypes (sometimes the whole community) in the presence of a
Artificial Life Volume 26, Number 2
211
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
temporally displaced replacer or invader. These qualitatively similar results suggest that
both our digital communities and real metabolic pathways are made up of mutually co-
adapted elements, and that the context as a whole suffers when one element is out of
evolutionary sync with the others. Our results also suggest another interesting avenue for
future work by allowing evolutionary responses [29] in the kinds of experiments we have
performed here, rather than just ecological-scale experiments where selection does not act
on either the replaced ecotype or the rest of the community. Better still would be
experiments where the invader is kept evolutionarily frozen while the rest of the
community is allowed to evolve.
5 Concluding Remarks
Our results show the potential for digital evolution to contribute to studying dynamics of multispe-
cies community interactions, combining them with aspects of macroevolution for a fuller under-
standing of how community composition and ecological and evolutionary responses are shaped
over time. While digital organisms and the communities they form lack many details of real-world
analogues, our results bear a number of similarities to what is known from real communities, par-
ticularly microbial ones, and suggest a number of promising avenues for future research. Here, we
have shown that “fitness” (measured as a reproductive rate as for real organisms) in cross-feeding
communities of digital organisms can depend heavily on community context. We have also shown
that evolution continues in a cryptic but discontinuous manner even when eco-phenotypic data
would imply stasis, and that major macroevolutionary events such as mass extinctions can produce
unforeseen redirections of prevailing evolutionary tendencies, leading to unexpected evolutionary
intransitivity when pre- and post-extinction communities meet. Remarkably, our results obtain with-
out any “sophisticated” biotic interactions, such as predation and parasitism; rather, they arise from
relatively simple facilitative resource dynamics. Further development of digital evolution platforms
(for example, to allow lineage-specific “freezing” of evolution), when combined with measurement
approaches developed for experimental evolution and community ecology, make digital evolution a
capable alternative to more traditional modeling and biological approaches for elucidating general
principles of evolution in interacting multispecies communities.
Although we have already highlighted commonalities between our results and previous real bio-
logical ones, perhaps the greatest implications of our results are for still-speculative areas of science.
Here, we have been able to resurrect organisms lost during mass extinctions and compete them
against post-extinction ones, often revealing the inadequacy of the latter. While still the domain
of science fiction novels and movies, advances in modern biotechnology could conceivably resurrect
extinct megafauna within the next century. While great caution must be exercised in extrapolating
results from a digital system like ours to the real world (or even a still-fictional extension of it), we
believe that our results have bearing on the wisdom of actually attempting this. Unless resurrected
species are kept in tightly contained artificial environments, our results suggest that, if released into
the modern natural world, such species might well have unforeseen negative interactions with mem-
bers of the modern biota, perhaps to the point of driving those modern organisms extinct. Our
results also imply that even if resurrected species succeed in establishing themselves in the modern
world, it is quite likely that, due to mismatched ecological interactions, their performance will be
reduced relative to what it was in their own fully native community prior to their initial extinction.
Acknowledgments
We thank three anonymous reviewers for comments and feedback that helped us improve the man-
uscript. This study was supported by research grants #31470435 from the National Natural Science
Foundation of China (http://www.nsfc.gov.cn) and #KYRC201301 from the Chinese Ministry of
Education (http://www.moe.edu.cn) to G.Y. The funders had no role in study design, data collec-
212
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
tion and analysis, decision to publish, or preparation of the manuscript. G.Y. gives special thanks to
Dr. Tim Cooper (School of Natural and Computational Sciences, Massey University, Palmerston
North, New Zealand) for suggesting the sequential full-community competition approach; to Dr.
Richard E. Lenski (Department of Microbiology and Molecular Genetics, Michigan State University,
East Lansing, Michigan, USA) and Dr. Josef Voglmeir (College of Food Science, Nanjing Agricul-
tural University, Nanjing, Jiangsu, China) for fruitful advice and discussions; and to Dr. Nils Christian
Stenseth (Centre for Ecological and Evolutionary Synthesis, University of Oslo, Oslo, Norway) for
providing the initial intellectual stimulus for this project. This manuscript grew out of a talk presented
at the celebration for Dr. Lenskiʼs 60th birthday at the 2016 BEACON Congress, held August 10–13,
2016 at Michigan State University.
References
1. Adams, J., & Oeller, P. W. (1986). Structure of evolving populations of Saccharomyces cerevisiae: Adaptive
changes are frequently associated with sequence alterations involving mobile elements belonging to the Ty
family. Proceedings of the National Academy of Sciences of the USA, 83(18), 7124–7127. http://doi.org/10.1073/
pnas.83.18.7124.
2. Adams, J., Paquin, C., Oeller, P. W., & Lee, L. W. (1985). Physiological characterization of adaptive clones
in evolving populations of the yeast, Saccharomyces cerevisiae. Genetics, 110, 175–185.
3. Altieri, A. H., van Wesenbeeck, B. K., Bertness, M. D., & Silliman, B. R. (2010). Facilitation cascade drives
positive relationship between native biodiversity and invasion success. Ecology, 91, 1269–1275.
4. Barnosky, A. D. (2001). Distinguishing the effects of the Red Queen and Court Jester on Miocene mammal
evolution in the northern Rocky Mountains. Journal of Vertebrate Paleontology, 21(1), 172–185. https://doi.
org/10.1671/0272-4634(2001)021[0172:DTEOTR]2.0.CO;2.
5. Beckerman, A. P., Petchey, O. L., & Warren, P. H. (2006). Foraging biology predicts food web complexity.
Proceedings of the National Academy of Sciences of the USA, 103(37), 13745–13749. https://doi.org/10.1073/
pnas.0603039103.
6. Bell, G. (1982). The masterpiece of nature: The evolution and genetics of sexuality. Cambridge, UK: Cambridge
University Press Archive.
7. Bell, G. (1997). Selection: The mechanism of evolution. New York: Chapman & Hall.
8. Bell, G. (2015). The evolution of life. Oxford, UK: Oxford University Press.
9. Benton, M. J. (2009). The Red Queen and the Court Jester: Species diversity and the role of biotic and
abiotic factors through time. Science, 323, 728–732.
10. Biswas, R., Ofria, C., Bryson, D. M., & Wagner, A. P. (2014). Causes vs benefits in the evolution of prey
grouping. In H. Sayama, J. Rieffel, S. Risi, R. Doursat, & H. Lipson (Eds.), ALIFE 2014: The Fourteenth
International Conference on the Synthesis and Simulation of Living Systems (pp. 641–648). Cambridge, MA: MIT
Press.
11. Blount, Z. D., Borland, C. Z., & Lenski, R. E. (2008). Historical contingency and the evolution of a key
innovation in an experimental population of Escherichia coli. Proceedings of the National Academy of Sciences of the
USA, 105, 7899–7906.
12. Chow, S. S., Wilke, C. O., Ofria, C., Lenski, R. E., & Adami, C. (2004). Adaptive radiation from resource
competition in digital organisms. Science, 305(5680), 84–86. http://doi.org/10.1126/science.1096307.
13. Cliff, D., & Miller, G. F. (1995). Tracking the Red Queen: Measurements of adaptive progress in co-
evolutionary simulations. In F. Morán, A. Moreno, J. J. Merelo, & P. Chacón (Eds.), Advances in artificial life:
Proceedings of the Third European Conference on Artificial Life (pp. 200–218). Berlin: Springer-Verlag.
14. Collins, S. (2011). Many possible worlds: Expanding the ecological scenarios in experimental evolution.
Evolutionary Biology, 38(1), 3–14. https://doi.org/10.1007/s11692-010-9106-3.
15. Cooper, T., & Ofria, C. (2002). Evolution of stable ecosystems in populations of digital organisms. In R. K.
Standish & M. A. Bedau (Eds.), Eighth International Conference on Artificial Life, December 9–13, Sydney,
New South Wales (pp. 227–232). Cambridge, MA: MIT Press.
16. Crichton, M. (1991). Jurassic Park. New York: Knopf.
Artificial Life Volume 26, Number 2
213
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
17. De Mazancourt, C., Johnson, E., & Barraclough, T. G. (2008). Biodiversity inhibits speciesʼ evolutionary
responses to changing environments. Ecology Letters, 11(4), 380–388. https://doi.org/10.1111/j.1461-
0248.2008.01152.x.
18. Decaestecker, E., & King, K. (2018). Red Queen dynamics. In B. D. Fath (Ed.), Encyclopedia of ecology Vol. 3
(2nd ed.) (pp. 185–192). Amsterdam, London, Cambridge, MA: Elsevier.
19. Ezard, T. H. G., Aze, T., Pearson, P. N., & Purvis, A. (2011). Interplay between changing climate and
speciesʼ ecology drives macroevolutionary dynamics. Science, 332(6027), 349–351. https://doi.org/
10.1126/science.1203060.
20. Ezard, T. H. G., & Purvis, A. (2016). Environmental changes define ecological limits to species richness
and reveal the mode of macroevolutionary competition. Ecology Letters, 19(8), 899–906. https://doi.org/
10.1111/ele.12626.
21. Goings, S., Clune, J., Ofria, C., & Pennock, R. T. (2004). Kin selection: The rise and fall of kin-cheaters. In
J. Pollack, M. Bedau, P. Husbands, T. Ikegami, & R. A. Watson (Eds.), Proceedings of the Ninth International
Conference on Artificial Life (pp. 303–308). Cambridge, MA: MIT Press.
22. Harmon, L. J., & Harrison, S. (2015). Species diversity is dynamic and unbounded at local and continental
scales. The American Naturalist, 185(5), 584–593. https://doi.org/10.1086/680859.
23. Hubbell, S. P. (2001). The unified neutral theory of biodiversity and biogeography. Princeton, NJ: Princeton
University Press.
24. Hull, P. (2015). Life in the aftermath of mass extinctions. Current Biology, 25, R941–R952. https://doi.org/
10.1016/j.cub.2015.08.053.
25. Hull, P. M., Norris, R. D., Bralower, T. J., & Schueth, J. D. (2011). A role for chance in marine recovery
from the end-Cretaceous extinction. Nature Geoscience, 4(12), 856–860. https://doi.org/10.1038/ngeo1302.
26. Ivany, L. C., Brett, C. E., Wall, H. L. B., Wall, P. D., & Handley, J. C. (2009). Relative taxonomic and
ecologic stability in Devonian marine faunas of New York State: A test of coordinated stasis. Paleobiology,
35(4), 499–524. https://doi.org/10.1666/0094-8373-35.4.499.
27. Johansson, J. (2007). Evolutionary responses to environmental changes: How does competition affect
adaptation? Evolution, 62, 421–435. https://doi.org/10.1111/j.1558-5646.2007.00301.x.
28. Kacar, B. (2016). Rolling the dice twice: Evolving reconstructed ancient proteins in extant organisms.
In G. Ramsey & C. Pence (Eds.) Chance in evolution (pp. 264–276). Chicago: University of
Chicago Press.
29. Kacar, B., Ge, X., Sanyal, S., & Gaucher, E. A. (2017). Experimental evolution of Escherichia coli harboring
an ancient translation protein. Journal of Molecular Evolution, 84(2–3), 69–84. https://doi.org/10.1007/
s00239-017-9781-0.
30. Kéfi, S., Berlow, E. L., Wieters, E. A., Navarrete, S. A., & Petchey, O. L. (2012). More than a meal:
Integrating non-feeding interactions into food webs. Ecology Letters, 15, 291–300.
31. Lawrence, D., Fiegna, F., Behrends, V., Bundy, J. G., Phillimore, A. B., Bell, T., & Barraclough, T. G.
(2012). Species interactions alter evolutionary responses to a novel environment. PLoS Biology, 10(5).
https://doi.org/10.1371/journal.pbio.1001330.
32. Le Gac, M., Plucain, J., Hindré, T., Lenski, R. E., & Schneider, D. (2012). Ecological and evolutionary
dynamics of coexisting lineages during a long-term experiment with Escherichia coli. Proceedings of
the National Academy of Sciences of the USA, 109(24), 9487–9492. https://doi.org/10.1073/pnas.1207091109.
33. Lenski, R. E. (2017). Experimental evolution and the dynamics of adaptation and genome evolution in
microbial populations. ISME Journal, 11(10), 2181–2194. https://doi.org/10.1038/ismej.2017.69.
34. Li, M., Wei, Z., Wang, J., Jousset, A., Friman, V. P., Xu, Y., Shen, Q., & Pommier, T. (2019). Facilitation
promotes invasions in plant-associated microbial communities. Ecology Letters, 22(1), 149–158. https://doi.
org/10.1111/ele.13177.
35. Lieberman, B. S., Miller III, W., & Eldredge, N. (2007). Paleontological patterns, macroecological dynamics
and the evolutionary process. Evolutionary Biology, 34, 28–48. https://doi.org/10.1007/s11692-007-9005-4.
36. Liow, L. H., Van Valen, L., & Stenseth, N. C. (2011). Red Queen: From populations to taxa and
communities. Trends in Ecology & Evolution, 26(7), 349–358. https://doi.org/10.1016/j.tree.2011.03.016.
214
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
37. Loreau, M., & de Mazancourt, C. (2008). Species synchrony and its drivers: Neutral and nonneutral
community dynamics in fluctuating environments. The American Naturalist, 172(2), E48–E66. http://doi.
org/10.1086/589746.
38. Luo, T.-T., Heier, L., Khan, Z. A., Hasan, F., Reitan, T., Yasseen III, A. S., Xie, Z.-X., Zhu, J.-L., & Yedid,
G. (2019). Examining community stability in the face of mass extinction in communities of digital
organisms. Artificial Life, 24, 250–276. https://doi.org/10.1162/artl_a_00272.
39. Monteiro, A. B., & Del Bianco Faria, L. (2017). Causal relationships between population stability and food-
web topology. Functional Ecology, 31(6), 1294–1300. https://doi.org/10.1111/1365-2435.12833.
40. Morran, L. T., Schmidt, O. G., Gelarden, I. A., Parrish, R. C., & Lively, C. M. (2011). Running with the Red
Queen: Host-parasite coevolution selects for biparental sex. Science, 333(6039), 216–218. https://doi.org/
10.1126/science.1206360.
41. Morris, P. J., Ivany, L. C., Schopf, K. M., & Brett, C. E. (1995). The challenge of paleoecological stasis:
Reassessing sources of evolutionary stability. Proceedings of the National Academy of Sciences of the USA, 92,
11269–11273.
42. Mougi, A., & Kondoh, M. (2012). Diversity of interaction types and ecological community stability. Science,
337(6092), 349–351. https://doi.org/10.1126/science.1220529.
43. Nordbotten, J. M., & Stenseth, N. C. (2016). Asymmetric ecological conditions favor Red Queen type of
continued evolution over stasis. Proceedings of the National Academy of Sciences of the USA, 113(7), 1847–1852.
https://doi.org/10.1073/pnas.1525395113.
44. Ofria, C., Bryson, D., & Wilke, C. (2009). Avida: A software platform for research in computational
evolutionary biology. In A. Adamatzky & M. Komosinski (Eds.), Artificial life models in software (2nd ed.)
(pp. 3–35). London: Springer.
45. Olson, R. S., Mirmomeni, M., Brom, T., Bruger, E., Hintze, A., Knoester, D. B., & Adami, C. (2013).
Evolved digital ecosystems: Dynamic steady state, not optimal fixed point. In P. Liò, O. Miglino, G.
Nicosia, S. Nolfi, & M. Pavone (Eds.), Advances in artificial life (ECAL 2013) (pp. 126–133). Cambridge,
MA: MIT Press.
46. Rabosky, D. L., & Hurlbert, A. H. (2015). Species richness at continental scales is dominated by ecological
limits. The American Naturalist, 185(5), 572–583. https://doi.org/10.1086/680850.
47. Rainey, P. B., & Travisano, M. (1998). Adaptive radiation in a heterogeneous environment. Nature, 394,
69–72.
48. Rooney, N., & McCann, K. S. (2012). Integrating food web diversity, structure and stability. Trends in
Ecology & Evolution, 27(1), 40–46. https://doi.org/10.1016/j.tree.2011.09.001.
49. Rozen, D. E., Schneider, D., & Lenski, R. E. (2005). Long-term experimental evolution in Escherichia coli.
XIII. Phylogenetic history of a balanced polymorphism. Journal of Molecular Evolution, 61(2), 171–180.
http://doi.org/10.1007/s00239-004-0322-2.
50. Sauve, A. M. C., Fontaine, C., & Thébault, E. (2013). Structure–stability relationships in networks
combining mutualistic and antagonistic interactions. Oikos, 123(3), 378–384.
51. Simberloff, D., & Von Holle, B. (1999). Positive interactions of nonindigenous species: Invasional
meltdown? Biological Invasions, 1, 21–32.
52. Stenseth, N. C., & Maynard Smith, J. (1984). Coevolution in ecosystems: Red Queen evolution or stasis?
Evolution, 38(4), 870–880. http://www.jstor.org/stable/2408397.
53. Turner, C. B., Blount, Z. D., & Lenski, R. E. (2015). Replaying evolution to test the cause of extinction of
one ecotype in an experimentally evolved population. PLoS ONE, 10, e0142050.
54. Traveset, A., & Richardson, D.M. (2014). Mutualistic interactions and biological invasions. Annual Review of
Ecology, Evolution, and Systematics, 45, 89–113.
55. Van Valen, L. M. (1973). A new evolutionary law. Evolutionary Theory, 1, 1–30.
56. Vermeij, G. J., & Roopnarine, P. D. (2013). Reining in the Red Queen: The dynamics of adaptation and
extinction reexamined. Paleobiology, 39(04), 560–575. https://doi.org/10.1666/13009.
57. Walker, B. L. (2012). The evolution of digital communities under limited resources. Ph.D. thesis, Department of
Computer Science and Engineering, Michigan State University.
Artificial Life Volume 26, Number 2
215
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
T.-T. Luo et al.
Alteration of (Frequency-Dependent) Fitness
58. Warren, J., Topping, C. J., & James, P. (2011). An evolutionary modelling approach to understanding the
factors behind plant invasiveness and community susceptibility to invasion. Journal of Evolutionary Biology,
24(10), 2099–2109. https://doi.org/10.1111/j.1420-9101.2011.02337.x.
59. Yedid, G., & Heier, L. (2012). Effects of random and selective mass extinction on community composition
in communities of digital organisms. In P. Pontarotti (Ed.), Evolutionary biology: Mechanisms and trends
(pp. 43–64). Berlin, Heidelberg: Springer. https://doi.org/10.1007/978-3-642-30425-5.
60. Yedid, G., Ofria, C. A., & Lenski, R. E. (2008). Historical and contingent factors affect re-evolution of a
complex feature lost during mass extinction in communities of digital organisms. Journal of Evolutionary
Biology, 21, 1335–1357. https://doi.org/10.1111/j.1420-9101.2008.01564.x.
61. Yedid, G., Ofria, C. A., & Lenski, R. E. (2009). Selective press extinctions, but not random pulse
extinctions, cause delayed ecological recovery in communities of digital organisms. The American Naturalist,
173(4), E139–E154. https://doi.org/10.1086/597228.
62. Yedid, G., Stredwick, J., Ofria, C., & Agapow, P. (2012). A comparison of the effects of random and
selective mass extinctions on erosion of evolutionary history in communities of digital organisms. PLoS
ONE, 7(5), 1–13. https://doi.org/10.1371/journal.pone.0037233.
63. Zaman, L., Meyer, J. R., Devangam, S., Bryson, D. M., Lenski, R. E., & Ofria, C. (2014). Coevolution
drives the emergence of complex traits and promotes evolvability. PLoS Biology, 12(12). https://doi.org/
10.1371/journal.pbio.1002023.
64. Zeileis, A., Kleiber, C., Walter, K., & Hornik, K. (2003). Testing and dating of structural changes in
practice. Computational Statistics and Data Analysis, 44(1–2), 109–123. http://doi.org/10.1016/S0167-9473
(03)00030-6.
65. Zeileis, A., Leisch, F., Hornik, K., & Kleiber, C. (2002). strucchange: An R package for testing for
structural change in linear regression models. Journal of Statistical Software, 7(2), 1–38. http://doi.org/
10.18637/jss.v007.i02.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
1
9
6
1
8
9
6
0
3
6
a
r
t
l
/
_
a
_
0
0
3
0
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
216
Artificial Life Volume 26, Number 2