The Surprising Creativity of
Digital Evolution: A Collection of
Anecdotes from the Evolutionary
Computation and Artificial Life
Research Communities
Abstract Evolution provides a creative fount of complex and subtle
adaptations that often surprise the scientists who discover them.
However, the creativity of evolution is not limited to the natural
world: Artificial organisms evolving in computational environments
have also elicited surprise and wonder from the researchers studying
them. The process of evolution is an algorithmic process that
transcends the substrate in which it occurs. Indeed, many researchers
in the field of digital evolution can provide examples of how their
evolving algorithms and organisms have creatively subverted their
expectations or intentions, exposed unrecognized bugs in their code,
produced unexpectedly adaptations, or engaged in behaviors and
outcomes, uncannily convergent with ones found in nature. Such
stories routinely reveal surprise and creativity by evolution in these
digital worlds, but they rarely fit into the standard scientific narrative.
Instead they are often treated as mere obstacles to be overcome,
rather than results that warrant study in their own right. Bugs are
fixed, experiments are refocused, and one-off surprises are collapsed
into a single data point. The stories themselves are traded among
researchers through oral tradition, but that mode of information
transmission is inefficient and prone to error and outright loss.
Moreover, the fact that these stories tend to be shared only among
practitioners means that many natural scientists do not realize how
interesting and lifelike digital organisms are and how natural their
evolution can be. To our knowledge, no collection of such anecdotes
has been published before. This article is the crowd-sourced product
of researchers in the fields of artificial life and evolutionary
computation who have provided first-hand accounts of such cases.
It thus serves as a written, fact-checked collection of scientifically
important and even entertaining stories. In doing so we also present
here substantial evidence that the existence and importance of
evolutionary surprises extends beyond the natural world, and may
indeed be a universal property of all complex evolving systems.
Joel Lehman*
Uber AI Labs
lehman.154@gmail.com
Jeff Clune*
OpenAI
jclune@gmail.com
Dusan Misevic*
Université de Paris
INSERM U1284
Center for Research and
Interdisciplinarity
dule@alife.org
Christoph Adami, Michigan State University.
Lee Altenberg, Univeristy of Hawaiʼi at Mānoa.
Julie Beaulieu, Université Laval.
Peter J. Bentley, University College London.
Samuel Bernard, INRIA, Institut Camille Jordan,
CNRS.
Guillaume Beslon, Université de Lyon, INRIA,
CNRS.
David M. Bryson, Michigan State University.
Nick Cheney, University of Vermont.
Patryk Chrabaszcz, University of Freiburg.
Antoine Cully, Imperial College London.
Stephane Doncieux, Sorbonne Universités,
UPMC Univ Paris 06, CNRS, Institute of
Intelligent Systems and Robotics (ISIR).
Fred C. Dyer, Michigan State University.
Kai Olav Ellefsen, University of Oslo.
Robert Feldt, Chalmers University of Technology.
Stephan Fischer, INRA.
Stephanie Forrest, Arizona State University.
Antoine Fŕenoy, Université Grenoble Alpes.
Christian Gagńe, Université Laval.
Leni Le Goff, Sorbonne Universités, UPMC Univ
Paris 06, CNRS, Institute of Intelligent Systems
and Robotics (ISIR).
Laura M. Grabowski, State University of New
York at Potsdam.
Babak Hodjat, Cognizant Technology Solutions.
Frank Hutter, University of Vermont.
Laurent Keller, Department of Fundamental
Microbiology, University of Lausanne.
Carole Knibbe, Université de Lyon, INRIA,
CNRS.
Peter Krcah, Charles University Prague.
Richard E. Lenski, Michigan State University.
Hod Lipson, Columbia University.
Robert MacCurdy, University of Colorado.
Carlos Maestre, Sorbonne Universités, UPMC
Univ Paris 06, CNRS, Institute of Intelligent
Systems and Robotics (ISIR).
Risto Miikkulainen, University of Texas at
Austin.
* Organizing, lead, and contact authors, who contributed equally. Jeff Cluneʼs work done while at Uber AI Labs.
© 2020 Massachusetts Institute of Technology.
Published under a Creative Commons Attribution
4.0 International (CC BY 4.0) license.
Artificial Life 26: 274–306 (2020) https://doi.org/10.1162/artl_a_00319
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
Sara Mitri, Department of Fundamental
Microbiology, University of Lausanne.
David E. Moriarty, Apple Inc.
Jean-Baptiste Mouret, INRIA Nancy Grand—
Est.
Anh Nguyen, Auburn University.
Charles Ofria, Michigan State University.
Marc Parizeau, Université Laval.
David Parsons, Michigan State University.
Robert T. Pennock, Michigan State University.
William F. Punch, Michigan State University.
Thomas S. Ray, University of Oklahoma.
Marc Schoenauer, INRIA, Université Paris-
Saclay.
Eric Schulte, GrammaTech.
Karl Sims
Kenneth O. Stanley, Uber AI Labs and
University of Central Florida.
François Taddei, Center for Research and
Interdisciplinarity, INSERM U1284, Université
de Paris.
Danesh Tarapore, University of Southampton.
Simon Thibault, Université Laval.
Richard Watson, University of Southampton.
Westley Weimer, University of Virginia.
Jason Yosinski, Uber AI Labs.
Keywords
Surprise, creativity, digital evolution,
experimental evolution, genetic algorithms,
evolutionary computation
1 Introduction
Evolution provides countless examples of creative, surprising, and amazingly complex solutions to
lifeʼs challenges. Some flowers act as acoustic beacons to attract echo-locating bats [121], extremophile
microbes repair their DNA to thrive in the presence of extreme radiation [89], bombardier beetles repel
predators with explosive chemical reactions [131], and parasites reprogram host brains, inducing
suicide for the parasiteʼs own gain [78]. Many more examples abound, covering the full range of
biological systems [24, 49, 88]. Even seasoned field biologists are still surprised by the new adaptations
they discover, which they express in popular press accounts of their work [77, 100, 111], but only more
rarely in official academic publications, for example, in [41] but not in [76, 142].
Thus, the process of biological evolution is extremely creative [9, 29], at least in the sense that it
produces surprising and complex solutions that would be deemed creative if produced by a human.
But the creativity of evolution need not be constrained to the organic world. Independently of its
physical medium, evolution can happen wherever replication, variation, and selection intersect
[23, 27]. Thus, evolution can be instantiated digitally [25, 75], as a computer program, either to
study evolution experimentally or to solve engineering problems through directed digital breeding.
Similarly to biological evolution, digital evolution experiments often produce strange, surprising,
and creative results. Indeed, evolution often reveals that researchers actually asked for something
far different from what they thought they were asking for—not so different from those stories in
which a genie satisfies the letter of a request in an unanticipated way. Sometimes evolution reveals
hidden bugs in code or displays surprising convergence with biology. Other times, evolution simply
surprises and delights by producing clever solutions that investigators did not consider or had
thought impossible.
Artificial Life Volume 26, Number 2
275
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
While some such unexpected results have been published [9, 55, 90, 109], most have not, and
they have not previously been presented together, as they are here. One obstacle to their dissemi-
nation is that such unexpected results often result from evolution thwarting a researcherʼs intentions:
by exploiting a bug in the code, by optimizing an uninteresting feature, or by failing to answer the
intended research question. That is, such behavior is often viewed as a frustrating distraction, rather
than a phenomenon of scientific interest. Additionally, surprise is subjective and thus fits poorly with
the objective language and narrative expected in scientific publications. As a result, most anecdotes
have been spread only through word of mouth, providing laughs and discussion in research groups,
at conferences, and as comic relief during talks. But such communications fail to inform the field as
a whole in a lasting and stable way.
Importantly, these stories of digital evolution “outsmarting” the researchers who study it provide
more than an exercise in humility; they also provide insight and constructive knowledge for practi-
tioners, because they show the pervasiveness of such obstacles and how, when necessary, they can
be overcome. Furthermore, these cases demonstrate that robust digital models of evolution do not
blindly reflect the desires and biases of their creators, but instead they have depth sufficient to yield
unexpected results and new insights. Additionally, these cases may be of interest to researchers in
evolutionary biology as well as animal and plant breeding, because of their compelling parallels to the
creativity of biological evolution.
For these reasons, this article draws attention to surprise and creativity in algorithmic evolution,
aiming to document, organize, and disseminate information that, until now, has been passed down
through oral tradition, which is prone to error and outright loss. To compile this archive, the orga-
nizers of this article sent out a call for anecdotes to digital evolution mailing lists and succeeded in
reaching both new and established researchers in the field. We then selected from 90 submissions to
produce this “greatest hits” collection of anecdotes, and all co-authors of each selected submission
were included as co-authors on the article. Before presenting these stories, the next section provides
background information useful for those outside the fields of digital evolution and evolutionary
computation.
2 Background
2.1 Evolution and Creativity
Intuitively, evolutionʼs creativity is evident from observing lifeʼs vast and complex diversity. This
sentiment is well captured by Darwinʼs famous concluding thoughts in On the Origin of Species, where
surveying the myriad co-inhabitants of a single tangled bank leads to grand reflections on the “end-
less forms most beautiful” that were produced by evolution [22, p. 307]. Varieties of life diverge
wildly along axes of complexity, organization, habitat, metabolism, and reproduction, spanning
from single-celled prokaryotes to quadrillion-celled whales [147]. Since lifeʼs origin, biodiversity
has expanded as evolution has conquered the sea, land, and air, inventing countless adaptations
along the way [147].
The functional abilities granted by such adaptations greatly exceed the capabilities of current hu-
man engineering, which has yet to produce robots capable of robust self-reproduction or of auton-
omous exploration in the real world, or that demonstrate human-level intelligence. It would thus be
parochial to deny attributing creativity to the evolutionary process, if human invention of such ar-
tefacts would garner the same label. Admittedly, “creativity” is a semantically rich word that can take
on many different meanings. Thus to avoid a semantic and philosophical quagmire, while acknowl-
edging that other definitions and opinions exist, we here adopt the “standard definition” [116]: Cre-
ativity requires inventing something both original (i.e., novel) and effective (i.e., functional). Many of
evolutionʼs inventions clearly meet this benchmark.
The root of natural evolutionʼs creativity, in this standard sense of the term, is the sieve of re-
production. This sieve can be satisfied in many different ways, and as a result, evolution has pro-
duced a cornucopia of divergent outcomes [29, 147]. For example, nature has invented many
276
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
different ways to siphon the energy necessary for lifeʼs operation from inorganic sources (e.g., from
the sun, iron, or ammonia), and it has created many different wing structures for flight among in-
sects, birds, mammals, and ancient reptiles. Evolutionʼs creative potential has also been boot-
strapped from ecological interactions; the founding of one niche often opens others, for example,
through predation, symbiosis, parasitism, or scavenging. Although evolution lacks the foresight and
intentionality of human creativity, structures evolved for one functionality are often opportunistically
adapted for other purposes, a phenomenon known as exaptation [54]. For example, a leading theory
is that feathers first evolved in dinosaurs for temperature regulation [71] and were later exapted for
flight in birds. Even in the absence of direct foresight, studies of evolvability suggest that genomic
architecture itself can become biased toward increasing creative potential [62, 65, 66].
One component of evolution is the selective pressures that adapt a species to better fit its en-
vironment, which often results in creativity within that species. That is, meeting evolutionary chal-
lenges requires inventing effective solutions, such as better protection from predators or from
natural elements like wind or radiation. Beyond creativity within species, there are also evolutionary
forces that promote creative divergence, that is, that lead to the accumulation of novel traits or niches.
One such force is negative frequency-dependent selection [35]; this dynamic occurs when some
traits are adaptive only when rare, which promotes the evolution of organisms that demonstrate
different traits. Another divergent force is adaptive radiation [118], which occurs when access to
new opportunities allows an organism to rapidly diversify into a range of new species, as when a
new modality such as flight is discovered. In this way, evolution is driven toward effectiveness (be-
ing well adapted and functional) and toward originality both through the optimizing force of natural
selection and through divergent forces, thereby producing artefacts that meet both criteria of the
standard definition of creativity.
One aim of this article is to highlight that such creativity is not limited to the biological medium,
but is also a common feature of digital evolution. We continue by briefly reviewing digital evolution.
2.2 Digital Evolution
Inspired by biological evolution, researchers in the field of digital evolution study evolutionary pro-
cesses embodied in digital substrates. The general idea is that there exist abstract principles under-
lying biological evolution that are independent of the physical medium [23], and that these principles
can be effectively implemented and studied within computers [84]. As noted by Daniel Dennett,
“evolution will occur whenever and wherever three conditions are met: replication, variation (mu-
tation), and differential fitness (competition)” [26, pp. E83–E92]; no particular molecule (e.g., DNA
or RNA) or substrate (e.g., specific physical embodiment) is required.
In nature, heredity is enabled through replicating genetic molecules, and variation is realized
through mechanisms like copy errors and genetic recombination. Selection in biological evolution
results from how survival and reproduction are a logical requirement for an organismʼs genetic ma-
terial to persist. The insight behind digital evolution is that processes fulfilling these roles of repli-
cation, variation, and selection can be implemented in a computer, resulting in an evolutionary algorithm
(EA) [25]. For example, replication can be instantiated simply by copying a data structure (i.e., a
digital genome) in memory, and variation can be introduced by randomly perturbing elements within
such a data structure. Selection in an EA can be implemented in many ways, but the two most
common are digital analogues of artificial and natural selection in biological evolution. Because
the similarities and differences between these kinds of selection pressure are important for under-
standing many of the digital evolution outcomes, we next describe them in greater detail.
Artificial selection in biological evolution is exemplified by horse breeders who actively decide
which horses to breed together, hoping to enhance certain traits, for example, by breeding together
the fastest ones, or the smallest ones. In this mode, selection reflects human goals. Similarly, in
digital evolution a researcher can implement a fitness function as an automated criterion for selection.
A fitness function is a metric describing which phenotypes are preferred over others, reflecting the
researcherʼs goal for what should be evolved. For example, if applying an EA to design a stable gait
Artificial Life Volume 26, Number 2
277
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
for a legged robot, an intuitive fitness function might be to measure how far a controlled robot walks
before it falls. Selection in this EA would breed together those robot controllers that traveled far-
thest, in hopes that their offspring might travel even farther. This mode of selection is most com-
mon in engineering applications, where digital evolution is employed to achieve a desired outcome.
The other common mode of digital selection implements natural selection as it occurs in nature,
where evolution is open-ended. The main difference is that in this mode there is no specific target
outcome, and no explicit fitness function. Instead, digital organisms compete for limited resources,
which could be artificial nutrients, CPU cycles needed to replicate their code, or digital storage space
in which to write their genomes [105, 113]. Given variation within the population, some organisms
will survive long enough to reproduce and propagate their genetic material, while others will disap-
pear, which enables evolution to occur naturally. Typically, digital evolution systems and experiments
of this sort do not serve a direct engineering purpose, but are instead used as a tool to study prin-
ciples of life and evolution in an easier setting than natural biology; that is, they provide artificial life
model systems for use in experimental evolution [75]. Note that the field of digital evolution over-
laps with the study of EAs and of artificial life, but is not synonymous with it. For the purposes of
this article, we define digital evolution as evolutionary processes in which the algorithm of evolution
and the genetic material evolved are instantiated digitally. As a result, digital evolution does not in-
clude “wet” artificial life, which seeks alternative physical substrates for life [7]. Digital evolution does
encompass paradigms like virtual creatures evolving completely in silico [126]. It also includes the
fields of evolvable hardware and evolutionary robotics, where evolved digital programs or controllers
are deployed on real-world devices [13].
One persistent misconception of digital evolution is that, because it is instantiated in a compu-
tational substrate, it lacks relevance to the study of biological evolution. Yet both philosophical ar-
guments [23, 26, 27, 82, 109] and high-profile publications [2, 16, 18–21, 52, 59, 60, 83, 85, 86, 92,
141, 146, 148] suggest that digital evolution can be a useful tool to aid and complement the study of
biological evolution. Indeed, these evolving systems can be seen as real instances of evolution, rather
than mere simulation of evolution [110].
2.3 Surprise from Algorithms and Simulations
At first, it may seem counterintuitive that a class of algorithms can consistently surprise the re-
searchers who wrote them. Here we define surprise broadly as observing an outcome that signifi-
cantly differs from expectations, whether those expectations arise from intuitions, predictions from
past experiences, or theoretical models. Note that such surprise is a feature of the experimenterʼs
subjective experience. Because an algorithm is a formal list of unambiguous instructions that execute in a
prescribed order, it would seem sufficient to examine any algorithmʼs description to predict the full
range of its possible outcomes, undermining any affordance for surprise. However, a well-known
result in theoretical computer science is that, for many computer programs, the outcome of a pro-
gram cannot be predicted without actually running it [137]. Indeed, within the field of complex sys-
tems it is well known that simple programs can yield complex and surprising results when executed
[43, 74]; such results relate to the broader concept of emergence [6, 58], wherein a phenomenon re-
sulting from lower-level parts is best understood at a more abstract level of description (e.g., that a
copper atom exists at the tip of a statueʼs nose is better understood at the level of history and politics
than at the level of physics [28]).
This basic fact can be counterintuitive at first. Interactions with modern software, which is ex-
plicitly designed to be predictable, may understandably prime us with the expectation that innovation
and surprise cannot be captured by a computer algorithm. However, if surprising innovations are a
hallmark of biological evolution, then the default expectation ought to be that computer models that
instantiate fundamental aspects of the evolutionary process would naturally manifest similarly crea-
tive output. While we offer no formal proof in this article of digital evolutionʼs ability to generate
surprise, the diversity of anecdotes presented next highlights how common and widespread such
surprising results are in practice. It is important to note here that a facet of human psychology, called
278
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
hindsight bias, often obscures appreciating the subjective surprise of another person [115]. In other
words, humans often overestimate how predictable an event was after the fact. For many of the
anecdotes that follow, a post hoc understanding of the result is possible, which may lead the reader
to discount its surprisingness. While mediating this kind of cognitive bias is challenging, we mention
it here in hopes that readers might grant the original experimenters leeway for their inability to an-
ticipate what perhaps is easily recognized in retrospect. In other words, we believe that experi-
menters are in general well situated to objectively report on their subjective experience of surprise.
3 Routine Creative Surprise in Digital Evolution
The next subsections present 32 curated anecdotes representing the work of over 50 researchers. In
reviewing the anecdotes, we found that they roughly clustered into four representative categories:
misspecified fitness functions, in which digital evolution reveals the divergence between what an exper-
imenter is asking of evolution and what they think they are asking; unintended debugging, in which digital
evolution reveals and exploits previously unknown software or hardware bugs; exceeded experimenter
expectations, in which digital evolution discovers solutions that go beyond what an experimenter
thought evolution would produce; and convergence with biology, in which digital evolution discovers
solutions surprisingly convergent with those found in nature, despite vast divergence in medium
and conditions.
3.1 Misspecified Fitness Functions
When applying digital evolution to solve practical problems, the most common approach is for an
experimenter to choose a fitness function that reflects the desired objective of search. Such fitness
functions are often simple quantitative measures that seem to intuitively capture the critical features
of a successful outcome. These measures are a linchpin of EAs, as they serve as funnels to direct
search: Breeding is biased toward individuals with a high fitness score, in hopes that they will lead to
further fitness improvements, ultimately to culminate in the desired outcome.
This approach resembles the process of animal breeding and relies on the same evolutionary
principles for its success. However, as the following anecdotes illustrate, well-intentioned quantita-
tive measures are often maximized in counterintuitive ways. That is, experimenters often overesti-
mate how accurately their quantitative measure reflects the underlying qualitative success they have in
mind. This mistake is known as confusing the map with the territory (the metric is the map, whereas
what the experimenter intends is the actual territory [64]).
Exacerbating the issue, it is often functionally simpler for evolution to exploit loopholes in the quan-
titative measure than it is to achieve the actual desired outcome. Just as well-intentioned metrics in
human society can become corrupted by direct pressure to optimize them (known as Campbellʼs law
[14] or Goodhartʼs law [53]), digital evolution often acts to fulfill the letter of the law (i.e., the fitness
function) while ignoring its spirit. We often ascribe creativity to lawyers who find subtle legal loop-
holes, and digital evolution is often frustratingly adept at finding similar exploits.
In this section we describe many instances of this phenomenon, but the list is far from exhaus-
tive: Encountering the divergence between what we intended to select and what we actually selected
for is likely the most common way digital evolution surprises its practitioners.
3.1.1 Why Walk When You Can Somersault?
In a seminal work from 1994, Karl Sims evolved 3D virtual creatures that could discover walking,
swimming, and jumping behaviors in simulated physical environments. The creaturesʼ bodies were
made of connected blocks, and their “brains” were simple computational neural networks that gen-
erated varying torque at their joints based on perceptions from their limbs, enabling realistic-looking
motion. The morphology and control systems were evolved simultaneously, allowing a wide range of
possible bodies and locomotion strategies. Indeed, these virtual creatures remain among the most
iconic products of digital evolution [126, 127].
Artificial Life Volume 26, Number 2
279
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
Figure 1. Exploiting potential energy to locomote. Evolution discovers that it is simpler to design tall creatures that fall
strategically than it is to discover active locomotion strategies. The left photograph shows the creature at the start of a
trial, and the right photograph shows snapshots of the figure over time falling and somersaulting to preserve forward
momentum.
However, when Sims initially attempted to evolve locomotion behaviors, things did not go
smoothly. In a simulated land environment with gravity and friction, a creatureʼs fitness was mea-
sured as its average ground velocity during its lifetime of ten simulated seconds. Instead of inventing
clever limbs or snakelike motions that could push them along (as was hoped for ), the creatures
evolved to become tall and rigid. When simulated, they would fall over, harnessing their initial po-
tential energy to achieve high velocity. Some even performed somersaults to extend their horizontal
velocity (Figure 1). A video of this behavior can be seen here: https://goo.gl/pnYbVh. To prevent
this exploit, it was necessary to allocate time at the beginning of each simulation to relax the potential
energy inherent in the creatureʼs initial stance before motion was rewarded.
Building on Simsʼ work, but using a different simulation platform, Krcah [70] bred creatures to
jump as high above the ground as possible. In the first set of experiments, each organismʼs fitness
was calculated as the maximum elevation reached by the center of gravity of the creature during the
test. This setup resulted in creatures around 15 cm tall that jumped about 7 cm off the ground.
However, it occasionally also resulted in creatures that achieved high fitness values by simply having
a tall, static tower for a body, reaching high elevation without any movement. In an attempt to
correct this loophole, the next set of experiments calculated fitness as the furthest distance from
the ground to the block that was originally closest to the ground, over the course of the simulation.
When examining the quantitative output of the experiment, to the scientistʼs surprise, some evolved
Figure 2. Exploiting potential energy to pole-vault. Evolution discovers that it is simpler to produce creatures that fall and
invert than it is to craft a mechanism to actively jump.
280
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
individuals were extremely tall and also scored a nearly tenfold-improvement on their jumps! How-
ever, observing the creaturesʼ behaviors directly revealed that evolution had discovered another
cheat: somersaulting without jumping at all. The evolved body consisted of a few large blocks rem-
iniscent of a “head” supported by a long thin vertical pole (see Figure 2).
At the start of the simulation, the individual “kicks” the foot of its pole off the ground, and
begins falling head first, somersaulting its foot (originally the “lowest point” from which the jumping
score is calculated) away from the ground. Doing so created a large gap between the ground and the
“lowest point,” thus securing a high fitness score without having to learn the intended skill of jump-
ing. A video of the behavior can be seen here: https://goo.gl/BRyyjZ.
3.1.2 Creative Program Repair
In automated program repair, a computer program is designed to automatically fix other, buggy, computer
programs. A user writes a suite of tests that validate correct behavior, and the repair algorithmʼs goal
is to patch the buggy program so that it can pass all of the tests. One such algorithm is GenProg
[45], which applies digital evolution to evolve code (called genetic programming [68]). GenProgʼs evo-
lution is driven by a simple fitness function: the number of test cases a genetic program passes. That
is, the more tests an evolved program passes, the more offspring it is likely to have.
While GenProg is often successful, sometimes strange behavior results because human-written
test cases are written with human coders in mind. In practice, evolution often uncovers clever loop-
holes in human-written tests, sometimes achieving optimal fitness in unforeseen ways. For example,
when MIT Lincoln Lab evaluated GenProg on a buggy sorting program, researchers created tests
that measured whether the numbers output by the sorting algorithm were in sorted order. However,
rather than actually repairing the program (which sometimes failed to correctly sort), GenProg
found an easier solution: It entirely short-circuited the buggy program, having it always return an
empty list, exploiting the technicality that an empty list was scored as not being out of order [145].
In other experiments, the fitness function rewarded minimizing the difference between what the
program generated and the ideal target output, which was stored in text files. After several gener-
ations of evolution, suddenly and strangely, many perfectly fit solutions appeared, seemingly out of
nowhere. Upon manual inspection, these highly fit programs still were clearly broken. It turned out
that one of the individuals had deleted all of the target files when it was run! With these files missing,
because of how the test function was written, it awarded perfect fitness scores to the rogue candi-
date and to all of its peers [122]. In another project, to avoid runaway computation, the fitness func-
tion explicitly limited a programʼs CPU usage. In response, GenProg produced programs that slept
forever, which did not count toward CPU usage limits, as there were no computations actually per-
formed [145]. In all cases, updating the fitness function or disallowing certain program behaviors
eventually outwitted evolutionʼs creative mischief and resulted in debugged, improved programs.
3.1.3 Why Learn When You Can Exploit an Unintended Regularity?
One common trick that digital evolution often learns is to exploit subtle unintended patterns in data,
that is, ones that an experimenter may create without realizing they have provided an escape hatch
for evolution to latch onto, obviating the need to confront the intended challenge of the task. For
example, in a recent experiment, Ellefsen, Mouret, and Clune [33] investigated the problem of cat-
astrophic forgetting in neural networks: that learning a new task can destroy previous knowledge.
One element of the experiment was that neural connections could change during an agentʼs lifetime
through neuromodulatory learning [129]. The evolution of learning was promoted by presenting
objects several times and providing a reward or punishment for eating them (e.g., apple = edible,
mushroom = poisonous). The edibility of each object was randomized each generation, to force the
agents to learn these associations within their life instead of allowing evolution to hardcode the
knowledge.
The researchers were surprised to find that high-performing neural networks evolved that con-
tained nearly no connections or internal neurons: Even most of the sensory input was ignored. The
Artificial Life Volume 26, Number 2
281
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
networks seemed to learn associations without even receiving the necessary stimuli, as if a blind
person could identify poisonous mushrooms by color. A closer analysis revealed the secret to their
strange performance: Rather than actually learning which objects are poisonous, the networks
learned to exploit a pattern in how objects were presented. The problem was that food and poison
items were always presented in an alternating fashion: food, then poison, then food, then poison,
repeatedly. Evolution discovered networks that learn to simply reverse their most recent reward, so
they could alternate eating and not eating indefinitely, and correctly ignoring entirely what food
item was presented. In this way, evolution circumvented the intended research question, rather
than shedding light on it. The problem was easily solved by randomizing the order in which items
were presented.
3.1.4 Learning to Play Dumb on the Test
Like the previous one, this anecdote involves exploiting patterns inadvertently provided by researchers.
In research focused on understanding how organisms evolve to cope with high-mutation-rate en-
vironments [146], Ofria sought to disentangle the beneficial effects of performing tasks (which
would allow an organism to execute its code faster and thus replicate faster ) from evolved robust-
ness to the harmful effect of mutations. To do so, he tried to disable mutations that improved an
organismʼs replication rate (i.e., its fitness). He configured the system to pause every time a mutation
occurred, and then measured the mutantʼs replication rate in an isolated test environment. If the
mutant replicated faster than its parent, then the system eliminated the mutant; otherwise, the mu-
tant would remain in the population. He thus expected that replication rates could no longer im-
prove, thereby allowing him to study the effect of mutational robustness more directly. However,
while replication rates at first remained constant, they later unexpectedly started again rising. After a
period of surprise and confusion, Ofria discovered that he was not changing the inputs provided to
the organisms in the isolated test environment. The organisms had evolved to recognize those in-
puts and halt their replication. Not only did they not reveal their improved replication rates, but they
appeared to not replicate at all, in effect “playing dead” when presented with what amounted to a
predator.
Ofria then took the logical step of altering the test environment to match the same random dis-
tribution of inputs as would be experienced in the normal (non-isolated) environment. While this
patch improved the situation, it did not stop the digital organisms from continuing to improve their
replication rates. Instead they made use of randomness to probabilistically perform the tasks that
accelerated their replication. For example, if they did a task half of the time, they would have a 50%
chance of slipping through the test environment; then, in the actual environment, half of the organ-
isms would survive and subsequently replicate faster. In the end, Ofria eventually found a successful
fix, by tracking organismsʼ replication rates along their lineage, and eliminating any organism (in real
time) that would have otherwise out-replicated its ancestors.
3.1.5 Absurdly Thick Lenses, Impossible Superposition, and Geological Disarray
Optimization algorithms have often been applied to design lenses for optical systems (e.g., tele-
scopes, cameras, microscopes). Two families of solutions that were identified as likely being optimal
in an article using an optimization algorithm not based on evolution [107] were easily outperformed,
by a factor of two, by a solution discovered though digital evolution by Gagné et al. [50]. However,
the evolved solution, while respecting the formal specifications of the problem, was not realistic:
One lens in the evolved system was over 20 meters thick.
In a similarly underconstrained problem, William Punch collaborated with physicists, applying
digital evolution to find lower-energy configurations of carbon. The physicists had a well-vetted
energy model for between-carbon forces, which supplied the fitness function for evolutionary
search. The motivation was to find a novel low-energy buckyball-like structure. While the algorithm
produced very low-energy results, the physicists were irritated because the algorithm had found a
superposition of all the carbon atoms onto the same point in space. “Why did your genetic algorithm
282
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
violate the laws of physics?” they asked. “Why did your physics model not catch that edge condi-
tion?” was the teamʼs response. The physicists patched the model to prevent superposition, and
evolution was performed on the improved model. The result was qualitatively similar: great low-
energy results that violated another physical law, revealing another edge case in the simulator. At
that point, the physicists ceased the collaboration.
A final related example comes from an application of evolutionary algorithms to a problem in
geophysics. Marc Schoenauer relates attempting to infer underground geological composition by
analyzing echoes of controlled explosions [90]. The fitness function was a standard criterion used
in geology, based on properties of how waves align. To the experimentersʼ delight, evolution pro-
duced geological layouts with very high scores. However, an expert examining the underground lay-
outs selected by evolution declared that they “cannot be thought as a solution by anyone having
even the smallest experience in seismic data” [90, p. 645], as they described chaotic and unnatural
piles of polyhedral rocks.
These examples highlight how fitness functions often do not include implicit knowledge held by
experts, thus allowing for solutions that experts consider so ridiculous, undesirable, or unexpected
that they did not think to exclude or penalize such solutions when originally designing the fitness
function. Although failing to provide the desired type of solution, the surprising and unacceptable
results can catalyze thought and discussion that ultimately leads to more explicit understanding of
problems.
3.2 Unintended Debugging
Another manifestation of digital evolutionʼs creative freedom from human preconceptions about
what form a solution should take is that search will often learn how to exploit bugs in simulations
or hardware. One common approach in digital evolution is to start with a simulation of a physical
problem, so that evolution can initially be run completely in software. The benefit is that physics
simulations often run much faster than real time, thereby making more generations of evolution
feasible, which can allow studies that would be infeasible in the physical world. However, physics
simulations rarely mimic the real world exactly, meaning that subtle differences remain. As a result,
edge cases, bugs, or minor flaws in the implemented laws of physics are sometimes amplified and
exploited by evolution. The effect is the evolution of surprising solutions that achieve high fitness
scores by physically unrealistic or otherwise undesirable means. Because the researcher is often un-
aware of the bugs, these exploits almost by definition surprise; often, they are also entertaining.
While often frustrating to the experimenter, the benefit of such unintended debugging is to bring to
light latent faults in simulation or hardware that would otherwise remain liabilities. In effect, evolu-
tionʼs exploits can enable efficient debugging of the simulations, and thus can actually advance the
research program.
3.2.1 Evolving Virtual Creatures Reveal Imperfectly Simulated Physics
In further virtual-creature experiments [126, 127], Karl Simsʼ attempt to evolve swimming strategies
resulted in new ways for evolution to cheat. The physics simulator first used a simple Euler method
for numerical integration, which worked well for typical motion. However, with faster motion in-
tegration errors could accumulate, and some creatures learned to exploit that bug by quickly twitching
small body parts. In effect, they could obtain “free energy,” to propel them at unrealistic speeds
through the water. Similarly, when evolving jumping abilities, the creatures found a bug in the code
for collision detection and response. If the creatures hit themselves by contacting corners of two of
their body parts together in a certain way, an error was triggered that popped them airborne like
impossibly-strong grasshoppers. After such exploits were patched, the creatures eventually evolved
many other interesting methods of locomotion—ones that did not violate the laws of physics.
Later extensions of Simsʼ work encountered similar difficulties, as in Cheney et al.ʼs work evolv-
ing the morphology of soft robots [15]. One feature of their simulator was that it applied a heuristic
to estimate how coarsely it could simulate physics, to save on computation. The more cells that a
Artificial Life Volume 26, Number 2
283
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
Figure 3. Vibrating robots. Evolved behavior is shown in frames, where time is shown progressing from left to right. A
large time step enables the creatures to penetrate unrealistically through the ground plane, engaging the collision de-
tection system to create a repelling force, resulting in vibrations that propel the organism across the ground.
creature was composed of, the less stable the simulator estimated the creature to be, and it would
correspondingly simulate the world more granularly. In particular, the simulator decreased the time
delta separating subsequent simulation steps as the number of cells in the creature increased.
Creatures evolved to exploit this heuristic, paring down their body to only a few cells, resulting in a
large simulation time step. The large, less precise time step allowed the creatureʼs bottommost cells to
penetrate the ground between time steps without the collision being detected, which resulted in an up-
ward force from the physics engine to correct the unnatural state. This corrective force provided “free”
energy that enabled the creatures to vibrate and swiftly drift across the ground, producing a surprisingly
effective form of locomotion. (See Figure 3.) To achieve more realistic results, the system was patched.
Damping was increased when contacting the ground, the minimum creature size was raised, and the
time delta calculation was adjusted to reduce ground penetration. Evolution thus helped to surface un-
anticipated edge cases that were poorly handled by the physics simulator and experimental design.
3.2.2 Ticktacktoe Memory Bomb
In a graduate-level AI class at UT Austin in 1997 taught by Risto Miikkulainen, the capstone project
was a five-in-a-row ticktacktoe competition played on an infinitely large board. The students were
free to choose any technique they wanted, and most people submitted typical search-based solu-
tions. One of the entries, however, was a player based on the SANE neuroevolution approach
for playing Othello [96, 98]. As in previous work, the network received a board representation as
its input and indicated the desired move as its output. However, it had a clever mechanism for
encoding its desired move that allowed for a broad range of coordinate values (by using units with
an exponential activation function). A byproduct of this encoding was that it enabled the system to
request nonexistent moves very, very far away in the ticktacktoe board. Evolution discovered that
making such a move right away led to a lot of wins. The reason turned out to be that the other
players dynamically expanded the board representation to include the location of the far-away
move—and crashed because they ran out of memory, forfeiting the match.
3.2.3 Floating-Point Overflow Lands an Airplane
In 1997, Feldt applied digital evolution to simulations of mechanical systems to try to evolve mech-
anisms that safely, but rapidly, decelerate aircraft as they land on an aircraft carrier [38]. An incoming
aircraft attaches to a cable, and the system applies pressure on two drums attached to the cable. The
idea was to evolve the control software that would bring the aircraft to a smooth stop by dynamically
adapting the pressure. Feldt was expecting evolution to take many generations, given the difficulty of
the problem, but evolution almost immediately produced suspiciously nearly perfect solutions that were
very efficiently braking the aircraft, even when simulating heavy bomber aircraft coming in to land.
Given the perceived problem difficulty, and that no bugs in the evolutionary algorithm could be
found, the suspicion came to rest on the physics simulator. Indeed, evolution had discovered a loop-
hole in the force calculation for when the aircraftʼs hook attaches to the braking cable. By overflow-
ing the calculation, that is, exploiting the fact that numbers too large to store in memory roll over to
zero, the resulting force was sometimes estimated to be zero. This, in turn, would lead to a perfect
score, because of low mechanical stress on the aircraft, hook, cable, and pilot (because zero force
means very little deceleration, implying no damaging “g force” on the pilot). In this way, evolution
had discovered that creating enormous force would break the simulation, although clearly it was an
exceedingly poor solution to the actual problem. Interestingly, insights from this experiment led to
284
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
theories about using evolution in software testing (to find bugs and explore unusual behavior) and
engineering (to help refine knowledge about requirements) [38–40] that were later identified as im-
portant early works facilitating the field of “search-based software engineering” [56].
3.2.4 Why Walk Around the Wall When You Can Walk Over It?
The NeuroEvolving Robotic Operatives (NERO) video game applied digital evolution to enable
non-player characters to evolve in real time while the game is being played [130]. While the polished
version of the game that was released in 2005 portrays a world where order prevails, evolutionʼs
tendency to seek out and exploit loopholes led to some humorous and unrealistic behaviors during
development. For example, players of NERO are encouraged to place walls around their evolving
robots to help them learn to navigate around obstacles. However, evolution figured out how to do
something that should have been impossible: The robotic operatives consistently evolved a special
kind of “wiggle” that causes them to walk up the vertical walls, allowing them to ignore obstacles
entirely, and undermining the intent of the game. The NERO team had to plug this loophole, which
resulted from a little-known bug in the Torque game engine, after which the robots acquiesced in
the more physically realistic policy of walking around walls to get to the other side.
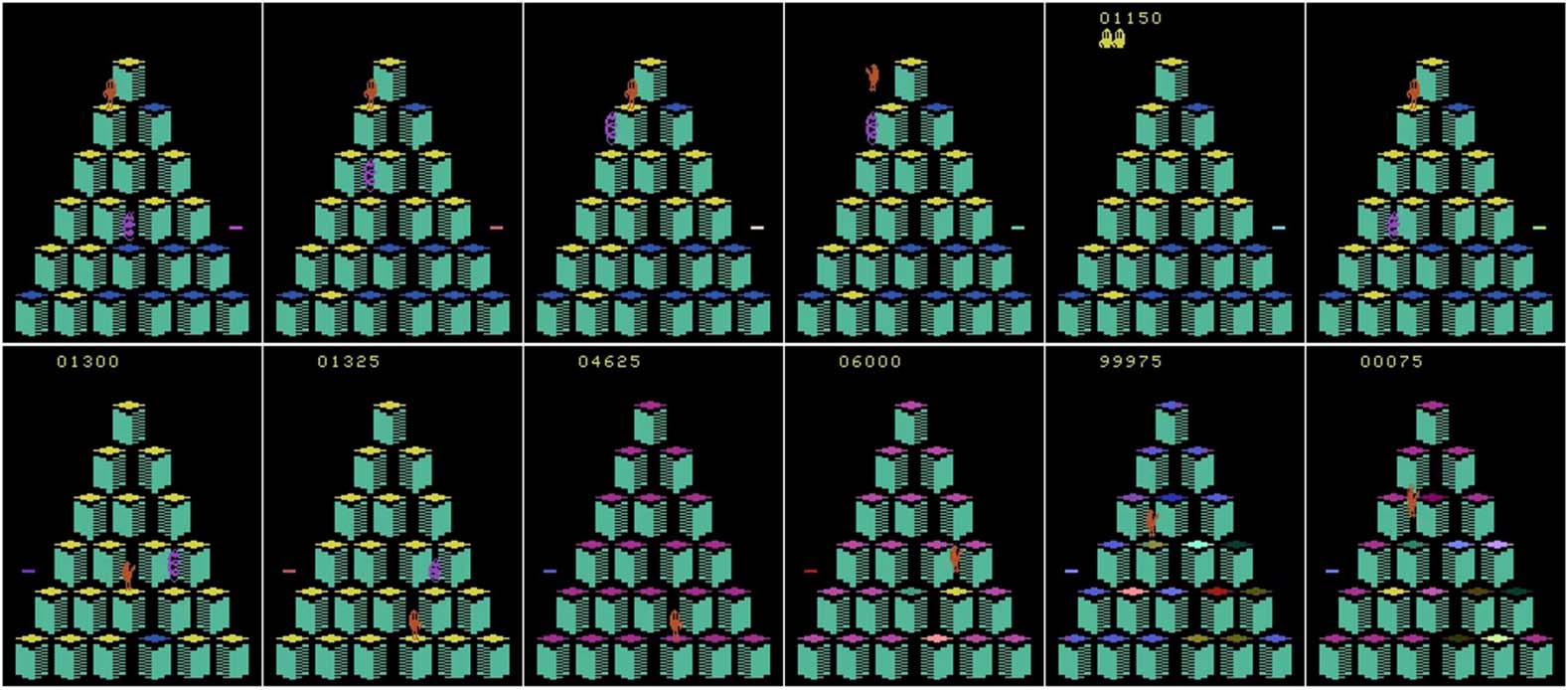
3.2.5 Exploiting a Bug in the Atari Game Q*bert
The next anecdote focuses on an evolutionary algorithm applied to a particular Atari game called
Q*bert. Atari games are a common benchmark in deep reinforcement learning [94, 95, 117, 132].
The challenge is to learn a policy that maps from raw pixels to actions at each time step, with the
objective of maximizing the game score. Typically, this policy is represented by a deep convolutional
neural network with many (often millions of) learned weight parameters. Relevant to this anecdote,
researchers at OpenAI [117], Uber [132], and the University of Freiburg [17] have recently shown
that evolutionary algorithms are a competitive approach to solving such games [94, 95].
In particular, the University of Freiburg team employed a simple version of a decades-old EA
called evolution strategies [114]. Interestingly, it learned to exploit two bugs in the Atari game
Q*bert. In the first case, which turned out to be a known bug, instead of completing the first level,
the agent baits an enemy to jump off the game platform with it, and scores the points for killing the
enemy; for some reason, the game engine does not count this suicide as a loss of life, and the agent is
able to repeat this process indefinitely (until the game cap of 100,000 frames). This pattern is shown
in Figure 4 (top) and in a video at https://goo.gl/2iZ5dJ.
In the second, more interesting and previously unknown, bug, the agent finds a sequence of
actions that completes the first level, but, for an unknown reason, this does not lead to the game
advancing to the next level; instead, all platforms start to blink and the agent is able to move around
seemingly aimlessly, but constantly gaining a huge number of points. The game counter was never
designed for such high scores and maxes out at 99,999. This exploit actually causes the game counter
to roll over many times (until the frame limit is reached) and seemingly could continue to do so
indefinitely; it improved the state-of-the-art high score from around 24,000 to nearly a million
points. Surprisingly, even the original developers of the game Q*bert (albeit a different version of
the game) were not aware of this bug, even after decades of continuous play [139]. This pattern is
shown in Figure 4 (bottom) and in a video at https://goo.gl/ViHRj2.
3.2.6 Reenabling Disabled Appendages
In work by Ecarlat and colleagues [31], an EA called MAP-Elites [99] was applied to explore possible
interactions between a robot arm and a small box on a table. The goal was to accumulate a wide
variety of controllers, ones that would move the cube to as many different locations on the table as
possible. In the normal experimental setup, MAP-Elites was able to move the cube onto the table, to
grasp the cube, and even to launch it into a basket in front of the robot [31]. In a follow-up exper-
iment the robotʼs gripper was crippled, preventing it from opening and closing. The natural expec-
tation was that the robotic arm could then move the small box in only limited ways, that is, to push it
Artificial Life Volume 26, Number 2
285
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
Figure 4. Top: The agent (orange blob in the upper left part of the screen) learns to commit suicide to kill its enemy
(purple spring); because of the bug, the game does not count this as a loss of life. Bottom: The agent uses a bug in the
game: After solving the first level in a specific sequence, the game does not advance to the next level, but instead the
platforms start to blink and the agent gains a huge amount of points.
around clumsily, because it could no longer grasp the box. Surprisingly, MAP-Elites found ways to
hit the box with the gripper in just the right way to force the gripper open so that it gripped the box
firmly (Figure 5). Once holding the box, the gripper could move to a broad range of positions,
undermining the experimentersʼ intent by in effect reenabling the disabled gripper (video:
https://goo.gl/upTaiP).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
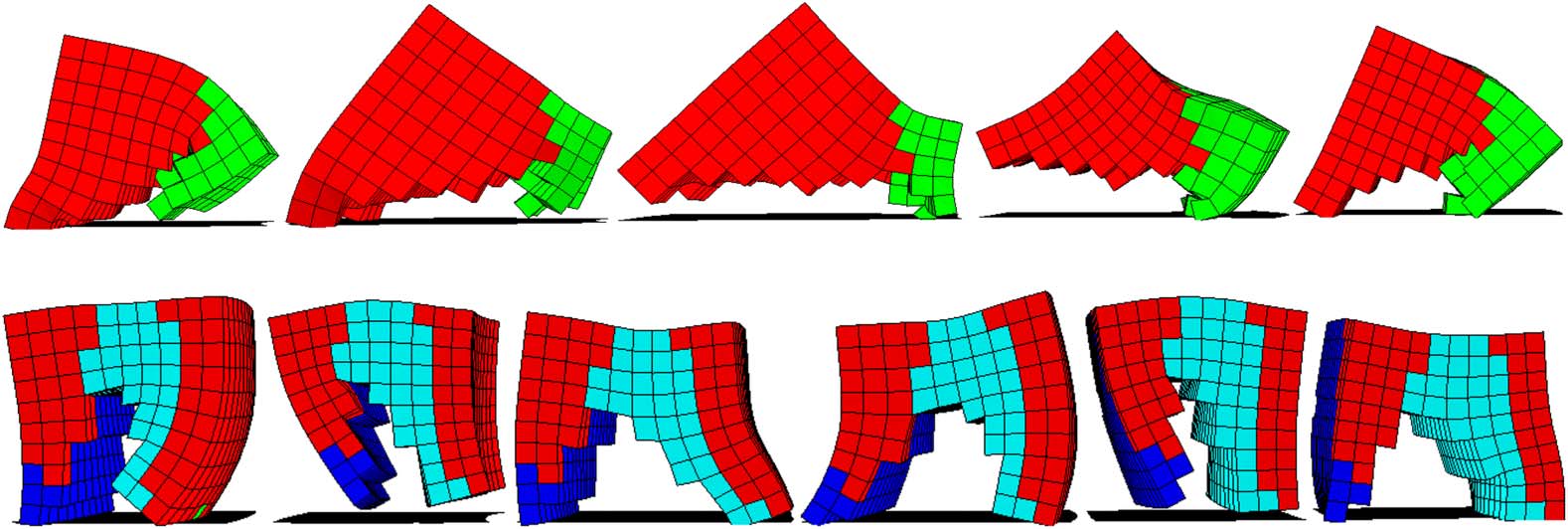
Figure 5. Snapshots of a forced-grasping trajectory. The robotic arm is in the initial position (a), with its gripper closed.
The arm pushes the small box (b) towards the armʼs base. The arm moves the gripper closer to its base (c), and ex-
ecutes a fast movement, sweeping across the table, forcing open its fingers, and grasping the small box. Finally, (d) the
arm moves its gripper to a position holding the small box.
286
Artificial Life Volume 26, Number 2
J. Lehman et al.
The Surprising Creativity of Digital Evolution
A similar result was noted by Moriarty and Miikkulainen in 1996 [97]. The researchers were
evolving neural networks to control a robot arm called OSCAR-6 [138] in a newly modified version
of the simulator. The goal was for the arm to reach a target point in midair; strangely, however, on
new experiments evolution took five times as long as it had previously. Observing the behavior of
the robot revealed a latent bug that arose when changing the simulator: The robot armʼs main motor
was completely disabled, meaning it could not directly turn towards targets that were far away from
its initial configuration. However, the arm still managed to complete the task: It slowly turned its
elbow away from the target, then quickly whipped it back—and the entire robot turned towards the
target from inertia. The movement sequence was repeated until the arm reached the target position.
It was not the solution that researchers were looking for, but one that revealed both a bug and an
unexpected way to solve the problem even under exceptional constraints.
3.3 Exceeding Experimenter Expectations
Another class of surprise is when evolution produces legitimate solutions that go beyond experimenter
expectations, rather than subverting experimenter intent or exploiting latent bugs.
3.3.1 Unexpected, Yet Valid, Solutions
Here we describe anecdotes in which evolution produces solutions that either were unconsidered or
thought impossible, or were more elegant or sophisticated than expected.
Unexpected Odometer
In an experiment evolving digital organisms to successfully navigate a connected trail of nutrients,
Grabowski et al. [55] encountered an unexpectedly elegant solution. Organisms were given the abil-
ity to sense whether there was nutrient underneath them and, if it was necessary to turn left or right,
to stay on the nutrient trail; but their sensors could not detect if they were at the end of the trail.
Organisms were rewarded for reaching more of the trail, and were penalized for stepping away from
the trail. Because it was impossible to directly sense where the trail ended, the best expected solution
was to correctly follow the trail one step past where it ended, which would incur a slight unavoidable
fitness penalty. However, in one run of evolution, the system achieved a perfect fitness score—an
analysis of the organism revealed that it had invented a step-counter, allowing it to stop precisely
after a fixed number of steps, exactly at the trailʼs end.
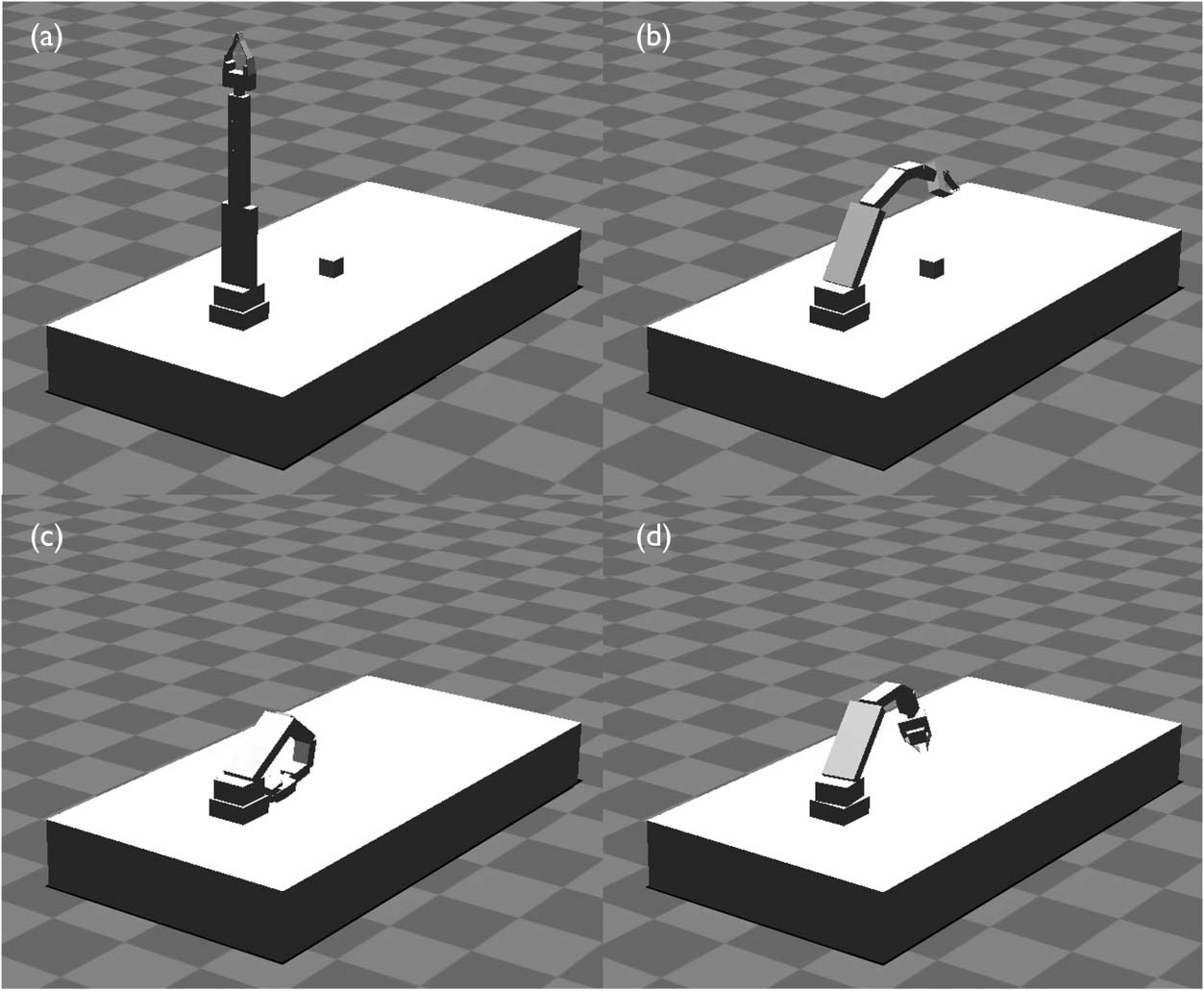
Elbow Walking
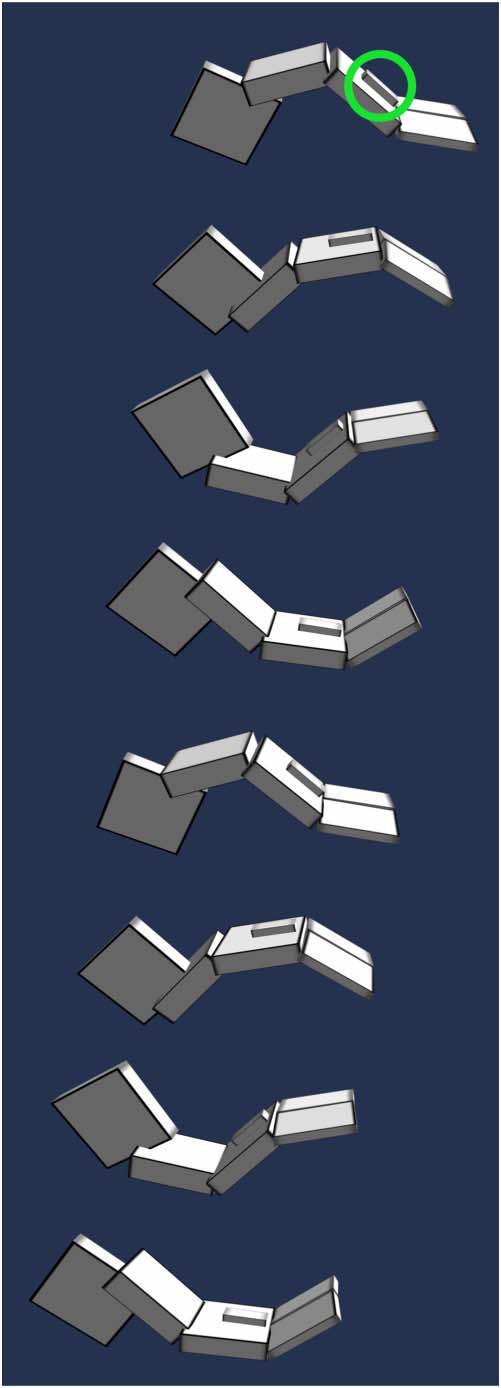
Cully et al. in 2015 [21] presented an algorithm that enables damaged robots to successfully adapt to
damage in under two minutes. The chosen robot had six legs, and evolutionʼs task was to discover
how to walk with broken legs or motors (Figure 6). To do so, ahead of the test, the researchers
coupled digital evolution with a robot simulator, to first learn a wide diversity of walking strategies.
Once damaged, the robot would then use the intuitions gained from simulated evolution to quickly
learn from test trials in the real world, zeroing in on a strategy that remained viable given the robotʼs
damage.
To evolve a large diversity of gaits, the team used the MAP-Elites evolutionary algorithm [99],
which simultaneously searches for the fittest organism over every combination of chosen dimen-
sions of variation (i.e., ways that phenotypes can vary). In this case, the six dimensions of variation
were the fraction of time that the foot of each leg touched the ground, a way to encourage learn-
ing diverse locomotion strategies. Thus, MAP-Elites searched for the fastest-moving gait possible
across every combination of how often each of the robotʼs six feet touched the ground. Naturally,
the team thought it impossible for evolution to solve the case where all six feet touch the ground
0% of the time, but to their surprise, it did. Scratching their heads, they viewed the video: It
showed a robot that flipped onto its back and successfully walked on its elbows, with its feet
in the air (Figure 7). A video with examples of the different gaits MAP-Elites found, including
Artificial Life Volume 26, Number 2
287
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
Figure 6. Six-legged robot. The robot uses the results of offline, simulated evolution to adapt quickly to a variety of damage
conditions, such as a broken leg. Each point on the colored floor represents a different type of gait, that is, a gait that uses
the robotʼs legs in different proportions. The assumption was that the cell in this map that required the robot to walk
without using any of its legs would be impossible to fill. But, to the researchersʼ surprise, evolution found a way.
this elbow walking gait (which is shown at the end starting at 1:49), can be viewed here: https://
goo.gl/9cwFtw.
Evolution of Unconventional Communication and Information Suppression
Mitri et al. [44, 93] applied digital evolution to groups of real and simulated robots, aiming to study
the evolution of communication. The small two-wheeled robots were equipped with blue lights,
which they could use as a simple channel of communication. Robots were rewarded for finding a
food source while avoiding poison, both of which were represented by large red lights distinguish-
able only in close proximity. Over many generations of selection, all the robots evolved to find the
food and avoid the poison, and under conditions that were expected to select for altruistic behavior,
they also evolved to communicate the location of the food, for example by lighting up after they had
reached it [44].
However, robots also solved the problem in surprising, unanticipated ways. In some cases,
when robots adapted to understand blue as a signal of food, competing robots evolved to signal
blue at poison instead, evoking parallels with dishonest signaling and aggressive mimicry in nature.
In other experiments that involved conditions selecting for competition between robots, authors
expected that the competitive robots simply would not communicate (i.e., not turn on their blue
light), because broadcasting the location of the food would potentially help competitors. But rath-
er than modifying how they signaled, some robots still lit up after finding food—but would then
Figure 7. Elbow-walking gait. The simulated robot, tasked with walking fast without touching any of its feet to the ground,
flips over and walks on its elbows. The red line shows the center of mass of the robot over time. Note that the robot
fulfills the task because the first few tenths of a second of the simulation are ignored, to focus on the gait in its limit cycle,
and not the robotʼs initial position.
288
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
literally hide the information from others by driving behind the food source (personal commu-
nication). Overall, a simple on-off light for communication revealed surprisingly rich evolutionary
potential.
Impossibly Compact Solutions
To test a distributed computation platform called EC-star [106], Babak Hodjat implemented a mul-
tiplexer problem [67], wherein the objective is to learn how to selectively forward an input signal.
Interestingly, the system had evolved solutions that involved too few rules to correctly perform the
task. Thinking that evolution had discovered an exploit, the impossibly small solution was tested
over all possible cases. The experimenters expected this test to reveal a bug in fitness calculation.
Surprisingly, all cases were validated perfectly, leaving the experimenters confused. Careful exami-
nation of the code provided the solution: The system had exploited the logic engineʼs rule evaluation
order to come up with a compressed solution. In other words, evolution opportunistically offloaded
some of its work into those implicit conditions.
This offloading is similar to seminal work by Adrian Thompson in evolving real-world electronic
circuits [136]. In Thompsonʼs experiment, an EA evolved the connectivity of a reconfigurable field-
programmable gate area (FPGA) chip, with the aim of producing circuits that could distinguish
between a high-frequency and a lower-frequency square-wave signal. After 5,000 generations of
evolution, a perfect solution was found that could discriminate between the waveforms. This was
a hoped-for result, and not truly surprising in itself. However, upon investigation, the evolved cir-
cuits turned out to be extremely unconventional. The circuit had evolved to work only in the spe-
cific temperature conditions in the lab, and exploited manufacturing peculiarities of the particular
FPGA chip used for evolution. Furthermore, when attempting to analyze the solution, Thompson
disabled all circuit elements that were not part of the main powered circuit, assuming that discon-
nected elements would have no effect on behavior. However, he discovered that performance de-
graded after such pruning. Evolution had learned to leverage some type of subtle electromagnetic
coupling, something a human designer would not have considered (or likely have known how to
leverage).
The Fastest Route is Not Always a Straight Line
Richard Watson and Sevan Ficici evolved the behavior of physical robots. The simple robots they
built had two wheels, two motors, and two light sensors [143, 144]. This type of robot is well known
from Braitenbergʼs famous book Vehicles [12], which argued that connecting sensor inputs to motor
outputs in a particular way results in simple light-following behavior. For example, when the right
wheel is driven proportionally to how much light the left sensor detects and the left wheel is similarly
driven by the right sensor, the robot will move towards the light. In Watson and Ficiciʼs case the
weights of connections between the input from the light sensors and the two wheel speeds were
determined by evolution. The initial question was whether Braitenbergʼs original solution would ac-
tually be found [143, 144].
While the evolved robots successfully drove towards the light source, they often did so in unusual
and unintuitive ways. Some backed up into the light while facing the dark, which was certainly an
unexpected strategy. Others found the source by light-sensitive eccentric spinning, rather than
the Braitenberg-style movement (Figure 8). It turns out that such spinning can easily be fine-tuned,
by tightening or loosening the curvature, to produce effective light-seeking. After some analysis the
authors discovered that the portion of the genetic search space that results in spinning is extremely
large, while the classical Braitenberg solution requires delicate balance (e.g., the robot must execute
subtle turns, changing heading from slightly clockwise to slightly anticlockwise: Figure 8) and thus
occupies a relatively tiny part of genetic space. Further, despite its apparent inefficiency, spinning
remained functional even when driven at higher speeds, unlike the classical solution, which could
not adjust quickly enough when run at high motor speeds. Additionally, the spinning solution was
more robust to hardware differences between the individual robots, and was less likely to get stuck
Artificial Life Volume 26, Number 2
289
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
Figure 8. Light-seeking robot movement. The path of the hand-coded Braitenberg-style movement (left) and evolved
spinning movement (right ) when moving towards a light source.
in corners of the arena. Thus, evolution ultimately was able to discover a novel solution that was
more robust than what had initially been expected.
Evolving a Car without Trying
At first glance it may seem that interactive evolution [133] is unlikely to surprise anyone, because of
close and constant interactions with the user. And yet, in the case of Picbreeder [124], one such
surprise was career-altering. Picbreeder is a platform, similar in form to the classic blind-watchmaker
experiment [24], where the user can evolve new designs by choosing and recombining parents, with
mutations, through successive generations. The images are encoded by mathematical functions,
which are invisible to the user and may strongly constrain the direction and size of successive evo-
lutionary steps. The surprise snuck up on one of the platform co-authors, Stanley, while evolving
new images from one that resembled an alien face. As Stanley selected the parents, he suddenly
noticed that the eyes of the face had descended and now looked like the wheels of a car. To his
surprise, he was able to evolve a very different but visually interesting and familiar image of a car in
short order from there. This quick and initially unintended transition between recognizable but dis-
similar images was only the beginning of the story. The surprise inspired Stanley to conceive the
novelty search algorithm [80], which searches without an explicit objective (just as Stanley found
the car unintentionally), selecting instead the most different, novel outcomes at each evolution step.
Later formalized by Lehman and Stanley together, the now-popular algorithm owes its existence to
the unexpected evolution of a car from an alien face.
3.3.2 Impressive Evolved Art and Design
The anecdotes so far have focused on applications and insights related to computer science and
engineering. However, there is also a long tradition of applying digital evolution to art and design.
Here we detail two such examples. What is impressive and surprising about these stories is that the
290
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
outputs were not valued because they were decent attempts by computers to produce artistic cre-
ations, but were instead judged as valuable strictly on their own merits.
Evolving Tables and Tunes
In the 1990s digital evolution was often applied to optimization problems, but rarely to produce
novel and functional designs. Peter Bentley was interested in this challenge, but initial feedback from
professional designers was dismissive and discouraging: Such an approach is impossible, they said,
because computers cannot invent new designs. They argued that even something as simple as a table
could not be invented by evolution—how could it possibly find the right structure from a vast sea of
possibilities, and how would you specify a meaningful fitness function?
This challenge led Bentley to create the generic evolutionary design system [8], which provided evo-
lution with an expanding set of building blocks it could combine into complex configurations. Fit-
ness functions were developed that rewarded separate aspects of a functional design, such as: Is the
upper surface flat? Will it stand upright when supporting a mass on its upper surface? Is its mass
light enough to be portable?
The task put to the generic evolutionary design system was to evolve a table. From random initial
designs, there emerged multiple elegant designs, including a variety of different functional tables,
such as the classic four-legged type, one with a small but heavy base, and one with a flat base
and internal weight (the “washing machine principle”) (Figure 9). One of the evolved tables was
successfully built and has remained in functional use for nearly two decades.
In 1999 Bentley was approached by a group of musicians and developers who wanted to generate
novel music through digital evolution. Dance music was popular at the time, so the team aimed to
evolve novel dance tracks. They set different collections of number-one dance hits as targets, that is,
an evolving track would be scored higher the more it resembled the targets. The evolved results,
eight-bar music samples, were evaluated by a musician who selected the ones to be combined into an
overall piece, which was then professionally produced according to the evolved music score. The
results were surprisingly good: The evolved tracks incorporated complex drum rhythms with inter-
esting accompanying melodies and bass lines. Using bands such as The Prodigy as targets, digital
evolution was able to produce novel dance tracks with clear stylistic resemblance.
In 2000 the group formed a record label named J13 Records. A distribution contract was drawn
up and signed with Universal Music, stipulating that the true source of the music should not be
revealed (even to the distributors, because Universal Musicʼs CEO believed that no one would want
to buy computer-generated music). The companies produced several dance tracks together, some of
which were then taken by other music producers and remixed. Some of the music was successful in
dance clubs, with the clubgoers having no idea that key pieces of the tracks they were dancing to
were the products of digital evolution.
An Art Museum Accepts and Displays Evolved Art Produced by Innovation Engines
The innovation engine [101] is an algorithm that combines three keys ideas: (1) produce new innova-
tions (i.e., solutions) by elaborating upon already evolved ones, (2) simultaneously evolve the pop-
ulation toward many different objectives (instead of a single objective as in traditional digital
evolution), and (3) harness state-of-the-art deep neural networks to evaluate how interesting a
Figure 9. Three table designs evolved using the generic evolutionary design system [8].
Artificial Life Volume 26, Number 2
291
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
Figure 10. A selection of images evolved via an innovation engine. Underneath each image is the type of image that
evolution was challenged to generate.
new solution is. The approach successfully produced a large diversity of interesting images, many of
which are recognizable as familiar objects, both to deep neural networks and to human observers
(Figure 10). Interestingly, the images span a variety of aesthetic styles, and bear resemblance to ab-
stract “concept art” pieces (e.g., the two different images of prison cells, the beacon, and the folding
chairs in Figure 10). Furthermore, the digital genomes of these algorithmically produced images are
quantitatively similar to the elegant, compact genomes evolved by humans on the interactive evo-
lution website Picbreeder [124].
To test whether the images generated by the innovation engine could pass for quality art, the
authors submitted a selection of evolved images to a competitive art contest: the University of
Wyomingʼs 40th Annual Juried Student Exhibition. Surprisingly, not only was the innovation engine
piece among the 35.5% of submissions accepted, it was also among the 21.3% of submissions that
were given an award! The piece was hung on the museum walls alongside human-made art, without
visitors knowing it was evolved (Figure 11).
3.4 Convergence with Biology
Because it is inspired by biological evolution, digital evolution naturally shares with it the important
abstract principles of selection, variation, and heritability. However, there is no guarantee that digital
evolution will exhibit similar specific behaviors and outcomes to those found in nature, because the low-
level details are so divergent: mutation rates, genome sizes, how genotypes map to phenotypes,
population sizes, morphology, type of interactions, and environmental complexity. Interestingly,
however, this subsection demonstrates how in practice there often exists surprising convergence
between evolution in digital and biological media.
3.4.1 Evolution of Muscles and Bones
In further results from Cheney et al.ʼs virtual-creature system [15], evolution generated locomotion
strategies unexpectedly convergent with those of biological creatures, examples of which are shown
in Figure 12. The gait in the top figure is similar to the crawling of an inchworm, requiring evolution
to discover from scratch the benefit of complementary (opposing) muscle groups, similar to such
292
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
Figure 11. University of Wyoming art show. A collection of images evolved with innovation engines on display at the
University of Wyoming Art Museum. They have also been displayed in art exhibits in galleries, fairs, and conventions in
several countries around the world.
muscle pairs in humans (e.g., biceps and triceps), and also to place them in a functional way. The gait
in the bottom row highlights digital evolutionʼs use of a stiff bonelike material to support thinner
appendages, enabling them to be longer and skinnier without sacrificing their weight-bearing poten-
tial. The end product is a gait reminiscent of a horseʼs gallop.
3.4.2 Evolution of Parasitism
In 1990, Tom Ray developed his seminal artificial life system, Tierra [112], an early instance of evo-
lution by natural selection in a digital medium. Organisms in Tierra consist of self-replicating ma-
chine code, somewhat like computer viruses. However, unlike computer viruses, organisms in Tierra
live on virtual machines explicitly designed to enable evolution (e.g., the instruction set was designed
to be fault-tolerant and evolvable). Tierra manages a population of replicating programs, killing off
the oldest programs or those generating the most errors. Importantly, the operations (including
copying) are faulty, meaning that replication necessarily produces mutations. Rayʼs hope was that
Tierra would eventually create an interesting and alien tree of life in a computational universe,
Figure 12. A stop-motion view of a small sample of the evolved gaits from Cheney et al. [15], which produced surpris-
ingly effective and biologically reminiscent behaviors. Shown here are soft robots progressing from left to right across
the panel. Colors correspond to voxel types (with red and green denoting oppositely contracting muscle groups, and
dark and light blue representing stiff and soft support materials, respectively). In the top gait, notice how evolution
creates distinct regions of each muscle. It employs these opposing muscle groups to create an inchworm-like behavior.
In the bottom gait, the use of stiff (bonelike) support material allows evolution to create relatively long appendages
and produce a horselike galloping behavior. Videos of various soft-robot gaits, including these two, can be found at
Artificial Life Volume 26, Number 2
293
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
Figure 13. Parasites in Tierra. A self-replicator (green, left) has code that copies the genome from parent to offspring.
The parasite (blue, center ) lacks the genome replicating code, and executes that code in its neighbor, copying its genome
into its offspring (blue shell, right). The blue sphere represents the parasiteʼs CPU executing its neighborʼs code. Image
courtesy of Anti-Gravity Workshop.
but he expected to spend perhaps years tinkering before anything interesting would happen; surpris-
ingly, Tierra produced complex ecologies the very first time it ran without crashing [112].
What emerged was a series of competing adaptations between replicating organisms within the
computer, that is, an ongoing coevolutionary dynamic. The surprisingly large palette of emergent
behaviors included parasitism (Figure 13), immunity to parasitism, circumvention of immunity, hy-
perparasitism (Figure 14), obligate sociality, cheaters exploiting social cooperation, and primitive
forms of sexual recombination. All of these relied on digital templates, parts of code that provide
robust addressing for JMP and CALL, the machine instructions that enable subroutines and control
changes. By accessing templates not only in their own genomes, but in the genomes of others, Tierra
organisms unexpectedly exploited this feature to facilitate a variety of ecological interactions.
When two individuals have complementary templates, interaction occurs. Organisms that
evolved matching templates were able to execute code in neighboring organisms. They were selected
for, because by outsourcing computation they reduced the size of their genome, which made rep-
lication less costly. Such organisms effectively engaged in informational parasitism. Evolving matching
templates enabled exploitation, while non-complementary templates allowed individuals to escape
Figure 14. Hyperparasites in Tierra. A red hyperparasite (center ), has captured a CPU (blue sphere) from a parasite, and
is using it to replicate its genome into the shell on the right. The hyperparasite also has its own CPU (red sphere), which
it is using to replicate also into the shell at the left. Image courtesy of Anti-Gravity Workshop.
294
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
exploitation. Ray termed the underlying process bit-string races, echoing the idea of evolutionary and
ecological arms races in nature.
But the dynamics went further than bit-string races. Hyperparasites stole the CPUs of parasites,
exhibiting energy parasitism. Social cooperators executed some of their own code, and some of their
identical neighborʼs code, to their mutual advantage. Social cheaters stole CPUs as they passed, ex-
ploiting the implicit trust between social creatures. Echoing natural evolution, a diversity of social
and ecological interactions evolved in complex ways.
3.4.3 Digital Vestigial Organs
Virtual creatures evolved in the ERO system by Krcah [70] displayed a curious property: They sometimes
contained small body parts whose function was not immediately obvious, yet they seemed to be carefully
placed on the creatureʼs body. It was not clear what purpose, if any, such “decorations” served. See
Figure 15 for an example of a swimming creature with an ornamental “fin” on top of its back.
Analysis of the “fin” and its evolution demonstrated that its persistence was a consequence of a
specific limitation of the evolutionary algorithm: Mutation was implemented in such a way that body
parts were never entirely removed from any creature. The “fin” body part from Figure 15 had
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 15. Swimming creature with an atrophied body part. A body part that was functionally important to an ancestor of
the depicted creature became atrophied over evolutionary time. Mutations within this system did not allow deleting
parts entirely; as a result, evolution shrunk the part and tuned its placement to minimize its deleterious effect on swim-
ming speed. See https://youtu.be/JHOXzQeeUik?list=PL5278ezwmoxQODgYB0hWnC0-Ob09GZGe2 for full video.
Artificial Life Volume 26, Number 2
295
J. Lehman et al.
The Surprising Creativity of Digital Evolution
origins as a big randomly generated block added very early in the creatureʼs evolution. Because it
could not be later removed when it started to interfere with the movements of the creature, it was
instead quickly atrophied to the smallest allowed size and moved into the least obtrusive position, by
a series of mutations.
3.4.4 Whole-Genome Duplication in Avida
Avida is a rich and versatile platform for digital evolution, one that has been used to study many
fundamental evolutionary questions [1, 2, 16, 18, 20, 34, 52, 83, 85, 86, 92, 146]. During the sub-
mission process for an article on genetic architecture and sexual reproduction [92], reviewers pointed
out that some data was unexpectedly bimodal: Evolution had produced two types of populations
with distinct properties. After further investigation, the two types were found to be largely separable
by their genome size. One group had lengths similar to the ancestral genome (50 instructions), while
the other group had genomes about twice as long, suggestive of genome duplication events. Dupli-
cation mutations were known to be theoretically possible, but it was not obvious why or how such a
sharp change in genome length had evolved. Interestingly, the Avida organisms had found an un-
anticipated (and unintended) mechanism to duplicate their genomes.
Experiments in Avida typically start from a hand-coded ancestral organism, effectively a short
program that consists of a series of instructions capable of self-reproduction but nothing else. The
reproduction mechanism executes a loop that iteratively copies the genome instruction by instruc-
tion. The loop terminates when an “if” instruction detects that the last instruction in the genome has
been copied. The double-length organisms resulted from an unanticipated situation, which was trig-
gered when organisms had an odd number of instructions in their genome, and a mutation then
introduced a second “copy” instruction into the copy loop. Because the “if” condition was checked
only after every two “copy” instructions, the copying process could continue past the last instruction
in the genome, ultimately copying the whole genome again. In this way, through a particular detail of
the Avida reproduction mechanism, digital organisms managed to duplicate their entire genomes, as
sometimes also happens in biological evolution.
3.4.5 Evolving Complex Behavior May Involve Temporary Setbacks
In a pioneering study, Richard Lenski and colleagues used the Avida digital evolution platform to test
some of Darwinʼs hypotheses about the evolution of complex features [86]. In Avida, digital organ-
isms can perform a wide variety of computational functions, including copying themselves, by exe-
cuting instructions in their genome. The researchers were interested in the general processes by which
the evolutionary process produces complex features. The team specifically focused on whether and
how Avidians might evolve to perform the most complex logical function in the environment—EQU
—which requires comparing two 32-bit numbers and determining whether or not they are equal.
The experiment provided several surprises about the creative power of the evolutionary process.
The EQU function evolved in about half the replicate experimental populations, but each instance
was unpredictably different, using anywhere from 17 to 43 instructions. The most surprising out-
come was that the pathway that evolution followed was not always an upward climb to greater fit-
ness, nor even a path consisting of sideways, neutral steps. Instead, in several cases, mutations along
the line of descent to EQU were deleterious, some significantly so. In two cases, mutations reduced
fitness by half. Though highly deleterious themselves, these mutations produced a genetic state that
allowed a subsequent beneficial mutation to complete a sequence that could preform the EQU func-
tion. This result sheds light on how complex traits can evolve by traversing rugged fitness landscapes
that have fitness valleys that can be crossed to reach fitness peaks.
3.4.6 Drakeʼs Rule
The Aevol digital evolution model, which belongs to the so-called sequence-of-nucleotides formalism
[57], was originally developed by Carole Knibbe and Guillaume Beslon with the intent to study
the evolution of modularity in gene order. However, even if some preliminary results on gene order
296
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
were promising, none of them turned out statistically viable after deeper investigation, seemingly
indicating that the whole project was likely to fail. Then, one day in a corridor, Knibbe bumped into
Laurent Duret, a renowned bioinformatician. Knibbe related her disappointing Ph.D. advancement,
saying, “We have nothing interesting; the only clear signal is that genome size apparently scales with
mutation rates—both the coding part and the noncoding part, but thatʼs trivial, isnʼt it?” Laurent
disagreed, “The noncoding part also? But thatʼs a scoop!” It turned out that (i) without being de-
signed to do so, the Aevol model had spontaneously reproduced Drakeʼs rule, stating that the size of
microbial genomes scales inversely with their mutation rate [30], and (ii) no model had predicted a
scaling between the mutation rates and the noncoding size of a genome. Only the relation between
the size of the coding region of the genome and the mutation rates was theoretically expected, as a
result of the error threshold effect first identified by Eigen in his quasi-species model [32]. The
effect on the noncoding region could be observed in Aevol because the model included chromo-
somal rearrangements in addition to point mutations [63]. This random encounter opened a new
research direction that ultimately led to a more general mathematical model of genome evolution,
showing that indirect selection for robustness to large duplications and deletions strongly bounds
genome size [42].
3.4.7 Becoming Unbreakable Rather than Better
Many examples have described surprising ways that digital evolution optimizes its fitness function.
However, early experiments aiming to evolve computer programs (a technique called genetic program-
ming [68]) revealed another kind of surprise. As evolution progressed across generations, evolved ge-
netic programs kept becoming larger and larger, a phenomenon called bloat [73, 87, 125], which
eventually slowed the algorithm to a crawl because it took so long to run the huge evolved programs
to test their fitness. Upon closer examination, the evolved programs were found, surprisingly, to be full
of “junk code” that could be completely removed without changing the behavior of the program at all
(see also the rich literature on neutrality in digital and biological evolution [51, 61]).
The mystery was resolved when researchers discovered that bloat can have an evolutionary ad-
vantage by buffering against the disruptive effects of mutating and mating genetic programs [3]. In
early populations, creating a child program by mutating it, or replacing parts of one parent with those
from another, is, on average, very damaging to fitness. Once in a while, of course, the change is
beneficial, which fuels evolutionary adaptation. Additionally, some changes will by chance introduce
new branches of neutral junk code that have no effect on fitness [104]. These mutants are no better
at satisfying the fitness function, but their offspring are less likely to be harmed by mutation or mating;
that is because organisms with a higher percentage of neutral code have a lower chance of random
changes happening to critical code. The result is that bloated individuals are more likely to produce
unchanged offspring, which grants them an indirect evolutionary advantage because the average fit-
ness of unchanged offspring is higher than that of offspring with mutations that have affected func-
tioning code. Over generations, programs thus grow larger and become increasingly robust to the
effects of mutation and crossover.
The surprise is that evolution has its own “agenda” distinct from the programmerʼs. While the
programmer hopes to create an algorithm that discovers the fittest structures, evolution may instead
seek the structures whose fitnesses are least disturbed by reproduction. In some cases these two
agendas may come into conflict, as they do here in two distinct ways. First, bloat causes genetic
programming to consume increasing amounts of computation and memory over generations (due
to rapidly growing evolved programs) [72]. The more fundamental conflict is that the high fraction
of junk code in these huge evolved programs requires far more generations of evolution to adapt
and improve—because evolution learns to turn off the very thing that ultimately fuels its ability to
learn: random changes that alter behavior. A similar result has been observed in digital evolution,
where when evolution was given the ability to evolve its own mutation rate, with accepted wisdom
being that doing so would improve the results, evolution instead short-sightedly turned off muta-
tions entirely [18]! Just as with bloat, this behavior is beneficial in the short run because mutations
Artificial Life Volume 26, Number 2
297
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
tend to be harmful on average, but prevents adaptation over the long term, greatly hurting long-term
performance. Interestingly, this phenomenon goes away when evolution is not forced to focus on
short-term fitness improvement [81]. That natural evolution could favor robustness to change has
been known to biologists since the 1940s [119, 140]. But the idea that natural selection could favor
robustness over finding the fittest organisms was discovered only in the 1980s [123], and later, in
digital evolution, as selection for “conservative code” [3], neutral evolution, of mutational robustness
[10, 102], and “survival of the flattest” [123, 146].
3.4.8 Costly Genes Hiding from Natural Selection
Genes coding for cooperative behaviors—such as public good secretion or altruistic suicide—face
very specific selection pressures that have interested researchers for decades. Their existence may
seem counterintuitive, because they bring a benefit to the population at the expense of the individ-
uals bearing them. The Aevol system has recently been used to study cooperation [47, 91], by giving
individuals the ability to secrete a public good molecule, which benefits all digital organisms in the
neighborhood. However, the public good molecule is costly for an individual to produce, digitally
mirroring the challenges facing the evolution of cooperation in biological systems. In one experi-
ment, the researchers studied the dynamics behind the loss of such costly cooperative genes. To
evolve populations that would secrete the public good molecule, the researchers first lowered the
cost of the molecule. After public good secretion had evolved, the researchers continued evolution
with an increased cost to study if its production would cease. Interestingly, while most populations
quickly lost all their secretion genes when the cost was increased in the second stage of evolution,
some populations consistently did not, even when the second-stage experiments were repeated
many times (starting from the same population).
The genetic analysis of the populations in which individuals consistently continued cooperating
led to a surprising result. The secretion genes that survived the increase in cost were frequently
overlapping with crucial metabolic genes, meaning that they were physically encoded using the same
DNA base pairs as a metabolic gene, but using the opposite strand or another reading frame [48]. As
a result, it was challenging for mutations to alter secretion behavior without also destroying meta-
bolic genes. Costly secretion genes were effectively hiding behind directly beneficial metabolic ones.
There is anecdotal evidence of similar mechanisms reducing the evolutionary potential toward cheat-
ing behavior in microbes [46, 103], but overlapping genes had never been studied in this context. To
highlight how such results may often go unappreciated and unstudied, we note that when Frenoy, a
masterʼs student at the time, manually looked at the genomes that preserved secretion despite its
cost (to try to understand how they were different), he had not heard of gene overlap and thought
the result was likely an uninteresting artefact of the Aevol system. Only when presenting his results
during a lab meeting did his colleagues point him toward the existence of overlapping genes in na-
ture, and the fact that selection pressures on such genetic systems are not yet fully understood.
4 Discussion
A persistent misunderstanding is that digital evolution cannot meaningfully inform biological knowl-
edge because “it is only a simulation.” As a result, it is difficult to convince biologists, other scien-
tists, and the general public that these systems, like biological evolution, are complex, creative, and
surprising. Often such disagreements occur outside of published articles, in informal conversations
and responses to reviewers. During such discussions, it is common for researchers in digital evolu-
tion to relate anecdotes like those included in this paper as evidence that such algorithms indeed
unleash the creativity of the Darwinian process. However, such arguments lack teeth when rooted in
anecdotes perpetuated through oral tradition. Thus one motivation for this article was to collect and
validate the true stories from the original scientists and collect them for posterity.
Future work could move beyond collecting stories to directly study the prevalance of surprise
among digital evolution practitioners. For example, a survey could be conducted to measure how
298
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
often researchers experience surprise (note that, like the anecdotes listed here, such surveys would
depend on experimentersʼ self-reports of surprise). To go beyond self-reports is possible but likely
requires expensive interventions, such as measuring physiological signals as experimenters work and
correlating physiological signals with self-reports of surprise.
A separate motivation for collecting these anecdotes is to highlight pragmatic lessons for digital
evolution practitioners. Foremost, a practitioner should be more skeptical of their ability to correctly
specify robust fitness functions, and anticipate evolution iteratively, revealing such specification fail-
ures. This awareness is most important for safety-critical applications, and points to the need for
careful supervision when attempting to apply digital evolution in real-world systems [79]. Undesir-
able outcomes can result from subtle interactions between fitness functions and experimental
setups, suggesting that practitioners should adopt an adversarial mindset, looking for ways in which
an agent could exploit seemingly innocuous design decisions. Another lesson relates to training in
simulation, where evolution often exploits bugs to achieve high fitness: Practitioners should regu-
larly visualize their simulated solutions to test whether the proposed solutions are valid and reason-
able. A final lesson for practitioners (and beginning researchers) is to understand how the nature of
academic publishing can obscure the actual (often messy) process of human-driven research that cul-
minates in published articles. In other words, the phenomenon described in this article is common
knowledge to experienced digital evolution researchers, but is nowhere to be found in the academic
literature. We believe the main reasons are: (1) surprise is a subjective experience and many consider
such subjectivity outside the typical bounds of science, (2) the surprising phenomena were often
orthogonal (or a hindrance) to the research questions that researchers were pursuing, and (3) the
convention for academic articles (deviations from which are often punished by reviewers) is to re-
port the final successful experiments as if they were all that were performed; the effect is to distort
the human experience of applying digital evolution in practice (e.g., to a new domain). As a result,
practitioners should read articles with a critical outlook, and adjust expectations when adopting a
published technique in a new domain.
Further, the ubiquity of surprising and creative outcomes in digital evolution has implications for
other fields of artificial intelligence. For example, beyond their importance to digital evolution, the
many examples of “misspecified fitness functions” in this article connect to the broader field of
artificial intelligence safety: Many researchers therein are concerned with the potential for perverse
outcomes from optimizing reward functions that appear sensible on their surface [4, 11, 37, 134],
characterized in that community as problems such as avoiding negative side effects [4, 69], reward
hacking (also known as wire-heading) [4, 36], or more generally AI alignment [134]. The list com-
piled here provides additional concrete examples of how difficult it is to anticipate the optimal be-
havior created and encouraged by a particular incentive scheme. Additionally, the narratives from
practitioners highlight the iterative refinement of fitness functions often necessary to produce de-
sired results instead of surprising, unintended behaviors. Interestingly, more-seasoned researchers
develop better intuitions about how the creative process of evolution works, although even they
sometimes still observe comical results from initial explorations in new simulations or experiments.
Thus digital evolution may provide an interesting training ground for developing intuitions about
incentives and optimization, to better ground theories about how to craft safer reward functions
for AI agents.
Finally, there are interesting connections between surprising results in digital evolution and the
products of directed evolution in biology, wherein selection in an experimenter-controlled evolution-
ary process is manipulated with the hope of improving or adapting proteins or nucleic acids for prac-
tical purposes [5, 108]. Echoing our “misspecified fitness functions” section, the first rule of directed
evolution is “you get what you select for” [108, p. 992]. Selection for exactly the property you care
about in directed evolution is often difficult and time-consuming, motivating cheaper heuristics that
experimenters assume will lead to the desired outcome. However, the result is often something that
meets the heuristic but deviates from the ideal outcome in surprising ways [120, 149]. In a final ironic
twist, similar evolutionary arguments (applied to a higher level of biological organization) suggest that
current incentive systems in science similarly produce surprising (and undesirable) byproducts [128].
Artificial Life Volume 26, Number 2
299
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
5 Conclusion
The Surprising Creativity of Digital Evolution
Across a compendium of examples, we have reviewed many ways in which digital evolution pro-
duces surprising and creative solutions. We have also synthesized lessons from these examples, to
aid practitioners and to communicate implications for biology and artificial intelligence more broadly.
For every anecdote we included, there are likely others that have been already forgotten as re-
searchers retire and new ones are created. The diversity and abundance of these examples suggest
that surprise in digital evolution is common, rather than a rare exception, providing evidence that
evolution—whether biological or computational—is inherently creative, and should routinely be ex-
pected to surprise, delight, and even outwit us.
Acknowledgments
We thank Elizabeth Ostrowski for a suggestion in the Digital Evolution lab at Michigan State Uni-
versity over a decade ago that led to the idea for this article, and for suggestions of Avida anecdotes
to include. We also appreciate Tim Taylor, from whom we got the idea for crowd-sourcing the
writing of this article by an open call for material from the community in which those whose sub-
missions were accepted would become co-authors of the article; he successfully piloted that model
for an article we were a part of [135], and we adopted it here. Finally, we also thank all of those who
submitted anecdotes that we were not able to include.
Jeff Clune was supported by an NSF CAREER award (CAREER: 1453549).
References
1. Adami, C. (2006). Digital genetics: Unravelling the genetic basis of evolution. Nature Reviews Genetics, 7(2),
109–118.
2. Adami, C., Ofria, C., & Collier, T. (2000). Evolution of biological complexity. Proceedings of the National
Academy of Sciences of the USA, 97(9), 4463.
3. Altenberg, L. (1994). The evolution of evolvability in genetic programming. In K. E. Kinnear (Ed.),
Advances in Genetic Programming, Vol. 1 (pp. 47–74). Cambridge, MA: MIT Press.
4. Amodei, D., Olah, C., Steinhardt, J., Christiano, P., Schulman, J., & Mané, D. (2016). Concrete problems
in AI safety. arXiv preprint arXiv:1606.06565.
5. Arnold, F. H. (1998). Design by directed evolution. Accounts of Chemical Research, 31(3), 125–131.
6. Bedau, M. A. (1997). Weak emergence. Noûs, 31, 375–399.
7. Bedau, M. A. (2003). Artificial life: Organization, adaptation and complexity from the bottom up. Trends in
Cognitive Sciences, 7(11), 505–512.
8. Bentley, P. J. (1996). Generic evolutionary design of solid objects using a genetic algorithm. Ph.D. thesis, The
University of Huddersfield.
9. Bentley, P. J. (1999). Is evolution creative? In P. J. Bentley & D. W. Corne (Eds.), Proceedings of the AISB,
Vol. 99 (pp. 28–34). Edinburgh: Society for the Study of Artificial Intelligence and Simulation of
Behaviour.
10. Bornberg-Bauer, E., & Chan, H. S. (1999). Modeling evolutionary landscapes: Mutational stability,
topology, and superfunnels in sequence space. Proceedings of the National Academy of Sciences of the USA, 96
(19), 10689–10694.
11. Bostrom, N. (2014). Superintelligence: Paths, dangers, strategies. Oxford, UK: Oxford University Press.
12. Braitenberg, V. (1986). Vehicles: Experiments in synthetic psychology. Cambridge, MA: MIT Press.
13. Bredeche, N., Haasdijk, E., & Eiben, A. (2009). On-line, on-board evolution of robot controllers. In P.
Collet, N. Monmarché, P. Legrand, M. Schoenauer, & E. Lutton (Eds.), International Conference on Artificial
Evolution EA2009 (pp. 110–121). Berlin, Heidelberg: Springer-Verlag.
14. Campbell, D. T. (1979). Assessing the impact of planned social change. Evaluation and Program Planning,
2(1), 67–90.
300
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
15. Cheney, N., MacCurdy, R., Clune, J., & Lipson, H. (2013). Unshackling evolution: Evolving soft robots
with multiple materials and a powerful generative encoding. In C. Blum (Ed.), Proceedings of the Genetic and
Evolutionary Computation Conference (pp. 167–174). New York: ACM.
16. Chow, S., Wilke, C., Ofria, C., Lenski, R., & Adami, C. (2004). Adaptive radiation from resource
competition in digital organisms. Science, 305(5680), 84.
17. Chrabaszcz, P., Loshchilov, I., & Hutter, F. (2018). Back to basics: Benchmarking canonical evolution
strategies for playing Atari. arXiv preprint arXiv:1802.08842.
18. Clune, J., Misevic, D., Ofria, C., Lenski, R., Elena, S., & Sanjuán, R. (2008). Natural selection fails to
optimize mutation rates for long-term adaptation on rugged fitness landscapes. PLoS Computational Biology,
4(9), e1000187.
19. Clune, J., Mouret, J.-B., & Lipson, H. (2013). The evolutionary origins of modularity. Proceedings of the Royal
Society B, 280(1755), 20122863.
20. Covert, A. W., Lenski, R. E., Wilke, C. O., & Ofria, C. (2013). Experiments on the role of deleterious
mutations as stepping stones in adaptive evolution. Proceedings of the National Academy of Sciences of the USA,
110(34), E3171–E3178.
21. Cully, A., Clune, J., Tarapore, D., & Mouret, J.-B. (2015). Robots that can adapt like animals. Nature,
521(7553), 503–507.
22. Darwin, C. (1859). On the origin of species. London: John Murray.
23. Dawkins, R. (1983). Universal Darwinism. In M. A. Bedau & C. E. Cleland (Eds.), The nature of life: Classical
and contemporary perspectives from philosophy and science. Cambridge, UK: Cambridge University Press.
24. Dawkins, R. (1986). The blind watchmaker: Why the evidence of evolution reveals a universe without design. New York:
W. W. Norton.
25. De Jong, K. A. (2006). Evolutionary computation: A unified approach. Cambridge, MA: MIT Press.
26. Dennett, D. C. (2002). The new replicators. In M. Pagel (Ed.), Encyclopedia of Evolution (pp. E83–E92).
Oxford, UK: Oxford University Press.
27. Dennett, D. C. (2014). Darwinʼs dangerous idea. New York: Simon and Schuster.
28. Deutsch, D. (1998). The fabric of reality. London: Penguin Books.
29. Dobzhansky, T. (1974). Chance and creativity in evolution. In F. J. Ayala & T. Dobzhansky (Eds.), Studies
in the philosophy of biology (pp. 307–338). Berlin: Springer.
30. Drake, J. W. (1991). A constant rate of spontaneous mutation in DNA-based microbes. Proceedings of the
National Academy of Sciences of the USA, 88(16), 7160–7164.
31. Ecarlat, P., Cully, A., Maestre, C., & Doncieux, S. (2015). Learning a high diversity of object
manipulations through an evolutionary-based babbling. In Proceedings of Learning Objects Affordances Workshop
at IROS 2016 (pp. 1–2). Hambourg: IROS.
32. Eigen, M. (1971). Self-organization of matter and the evolution of biological macromolecules.
Die Naturwissenschaften, 58(10), 465–523.
33. Ellefsen, K. O., Mouret, J.-B., & Clune, J. (2015). Neural modularity helps organisms evolve to learn new
skills without forgetting old skills. PLoS Computational Biology, 11(4), e1004128.
34. Elsberry, W. R., Grabowski, L. M., Ofria, C., & Pennock, R. T. (2009). Cockroaches, drunkards, and
climbers: Modeling the evolution of simple movement strategies using digital organisms. In Proceedings of
IEEE Symposium on Artificial Life (pp. 92–99). Piscataway, NJ: IEEE.
35. Endler, J. A., & Greenwood, J. J. D. (1988). Frequency-dependent predation, crypsis and aposematic
coloration. Philosophical Transactions of the Royal Society of London. B, Biological Sciences, 319(1196), 505–523.
36. Everitt, T., & Hutter, M. (2016). Avoiding wireheading with value reinforcement learning.
In B. Steunebrink, P. Wang, & B. Goertzel (Eds.), Artificial general intelligence (pp. 12–22).
Berlin: Springer.
37. Everitt, T., Lea, G., & Hutter, M. (2018). AGI safety literature review. arXiv preprint arXiv:1805.01109.
38. Feldt, R. (1998). Generating diverse software versions with genetic programming: An experimental study.
IEE Proceedings—Software Engineering, 145(6), 228–236.
Artificial Life Volume 26, Number 2
301
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
39. Feldt, R. (1999). Genetic programming as an explorative tool in early software development phases. In C.
Ryan & J. Buckley (Eds.), Proceedings of the 1st International Workshop on Soft Computing Applied to Software
Engineering (pp. 11–20). Limerick: Limerick University Press.
40. Feldt, R. (2002). Biomimetic software engineering techniques for dependability. Ph.D. thesis, Department of
Computer Engineering, Chalmers University of Technology, Gothenburg, Sweden.
41. Fire, A., Xu, S., Montgomery, M. K., Kostas, S. A., Driver, S. E., & Mello, C. C. (1998). Potent and specific
genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature, 391(6669), 806–811.
42. Fischer, S., Bernard, S., Beslon, G., & Knibbe, C. (2014). A model for genome size evolution. Bulletin of
Mathematical Biology, 76(9), 2249–2291.
43. Flake, G. W. (1998). The computational beauty of nature: Computer explorations of fractals, chaos, complex systems, and
adaptation. Cambridge, MA: MIT Press.
44. Floreano, D., Mitri, S., Magnenat, S., & Keller, L. (2007). Evolutionary conditions for the emergence of
communication in robots. Current Biology, 17(6), 514–519.
45. Forrest, S., Nguyen, T., Weimer, W., & Le Goues, C. (2009). A genetic programming approach to
automated software repair. In Proceedings of the Genetic and Evolutionary Computation Conference (pp. 947–954).
New York: ACM.
46. Foster, K. R., Shaulsky, G., Strassmann, J. E., Queller, D. C., & Thompson, C. R. (2004). Pleiotropy as a
mechanism to stabilize cooperation. Nature, 431(7009), 693–696.
47. Frénoy, A., Taddei, F., & Misevic, D. (2012). Robustness and evolvability of cooperation. In C. Adami,
D. M. Bryson, C. Ofria, & R. T. Pennock (Eds.), Artificial Life 13 Conference Proceedings (pp. 53–58).
Cambridge, MA: MIT Press.
48. Frénoy, A., Taddei, F., & Misevic, D. (2013). Genetic architecture promotes the evolution and
maintenance of cooperation. PLoS Computational Biology, 9(11), e1003339.
49. Futuyma, D. J. (2013). Natural selection and adaptation. In J. B. Losos, D. A. Baum, D. J. Futuyma, H. E.
Hoekstra, R. E. Lenski, A. J. Moore, C. L. Peichel, D. Schluter, & M. C. Whitlock (Eds.), The Princeton
Guide to Evolution (pp. 279–301). Princeton, NJ: Princeton University Press.
50. Gagné, C., Beaulieu, J., Parizeau, M., & Thibault, S. (2008). Human-competitive lens system design with
evolution strategies. Applied Soft Computing, 8(4), 1439–1452.
51. Galván-López, E., Poli, R., Kattan, A., OʼNeill, M., & Brabazon, A. (2011). Neutrality in evolutionary
algorithms… What do we know? Evolving Systems, 2(3), 145–163.
52. Goldsby, H. J., Dornhaus, A., Kerr, B., & Ofria, C. (2012). Task-switching costs promote the evolution
of division of labor and shifts in individuality. Proceedings of the National Academy of Sciences of the USA,
109(34), 13686–13691.
53. Goodhart, C. A. (1984). Problems of monetary management: The UK experience. Berlin: Springer.
54. Gould, S. J., & Vrba, E. S. (1982). Exaptation—a missing term in the science of form. Paleobiology, 8(01), 4–15.
55. Grabowski, L. M., Bryson, D. M., Dyer, F. C., Pennock, R. T., & Ofria, C. (2013). A case study of the de
novo evolution of a complex odometric behavior in digital organisms. PLoS One, 8(4), e60466.
56. Harman, M., Mansouri, S. A., & Zhang, Y. (2012). Search-based software engineering: Trends, techniques
and applications. ACM Computing Surveys, 45(1), 11:1–11:61.
57. Hindré, T., Knibbe, C., Beslon, G., & Schneider, D. (2012). New insights into bacterial adaptation
through in vivo and in silico experimental evolution. Nature Reviews Microbiology, 10(5), 352–365.
58. Holland, J. H. (2000). Emergence: From chaos to order. Oxford: Oxford University Press.
59. Kashtan, N., & Alon, U. (2005). Spontaneous evolution of modularity and network motifs. Proceedings of the
National Academy of Sciences of the USA, 102(39), 13773–13778.
60. Kashtan, N., Noor, E., & Alon, U. (2007). Varying environments can speed up evolution. Proceedings of the
National Academy of Sciences of the USA, 104(34), 13711.
61. Kimura, M. (1983). The neutral theory of molecular evolution. Cambridge: Cambridge University Press.
62. Kirschner, M., & Gerhart, J. (1998). Evolvability. Proceedings of the National Academy of Sciences of the USA, 95
(15), 8420.
302
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
63. Knibbe, C., Coulon, A., Mazet, O., Fayard, J.-M., & Beslon, G. (2007). A long-term evolutionary pressure
on the amount of non-coding DNA. Molecular Biology and Evolution, 24(10), 2344–2353.
64. Korzybski, A. (1958). Science and sanity: An introduction to non-Aristotelian systems and general semantics. Oxford,
UK: Institute of General Semantics.
65. Kounios, L., Clune, J., Kouvaris, K., Wagner, G. P., Pavlicev, M., Weinreich, D. M., & Watson, R. A.
(2016). Resolving the paradox of evolvability with learning theory: How evolution learns to improve
evolvability on rugged fitness landscapes. arXiv preprint arXiv:1612.05955.
66. Kouvaris, K., Clune, J., Kounios, L., Brede, M., & Watson, R. A. (2017). How evolution learns to
generalise: Using the principles of learning theory to understand the evolution of developmental
organisation. PLoS Computational Biology, 13, e1005358.
67. Koza, J. R. (1990). A hierarchical approach to learning the Boolean multiplexer function. In G. J. Rawlins
(Ed.), Foundations of genetic algorithms, Vol. 1 (pp. 171–192). Burlington, MA: Morgan Kaufmann.
68. Koza, J. R. (1992). Genetic programming: On the programming of computers by means of natural selection, Vol. 1.
Cambridge, MA: MIT Press.
69. Krakovna, V., Orseau, L., Martic, M., & Legg, S. (2018). Measuring and avoiding side effects using relative
reachability. arXiv preprint arXiv:1806.01186.
70. Krcah, P. (2008). Towards efficient evolutionary design of autonomous robots. In G. S. Hornby, L.
Sekanina, & P. C. Haddow (Eds.), Evolvable systems: From biology to hardware, Proceedings of the 8th International
Conference ICES (pp. 153–164). Berlin: Springer.
71. Kundrát, M. (2004). When did theropods become feathered?—evidence for pre-archaeopteryx feathery
appendages. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 302(4), 355–364.
72. Langdon, W. B. (2000). Quadratic bloat in genetic programming. In L. D. Whitley, D. E. Goldberg, E.
Cantú-Paz, & I. C. Parmee (Eds.), Proceedings of the 2nd Annual Conference on Genetic and Evolutionary
Computation (pp. 451–458). San Francisco: Morgan Kaufmann.
73. Langdon, W. B., & Poli, R. (1998). Fitness causes bloat. In P. K. Chawdhry & R. K. Pant (Eds.),
Soft Computing in Engineering Design and Manufacturing (pp. 13–22). Berlin: Springer.
74. Langton, C. G. (1990). Computation at the edge of chaos: phase transitions and emergent computation.
Physica D: Nonlinear Phenomena, 42(1–3), 12–37.
75. Langton, C. G. (1997). Artificial life: An overview. Cambridge, MA: MIT Press.
76. Last, K. S., Hobbs, L., Berge, J., Brierley, A. S., & Cottier, F. (2016). Moonlight drives ocean-scale mass
vertical migration of zooplankton during the Arctic winter. Current Biology, 26(2), 244–251.
77. Lau, N. C., & Bartel, D. P. (2003). Censors of the genome. Scientific American, 289(2), 34–41.
78. Lefevre, T., Adamo, S. A., Biron, D. G., Misse, D., Hughes, D., & Thomas, F. (2009). Invasion of the
body snatchers: The diversity and evolution of manipulative strategies in host–parasite interactions.
Advances in Parasitology, 68, 45–83.
79. Lehman, J. (2019). Evolutionary computation and AI safety: Research problems impeding routine and
safe real-world application of evolution. arXiv preprint arXiv:1906.10189.
80. Lehman, J., & Stanley, K. O. (2011). Abandoning objectives: Evolution through the search for novelty
alone. Evolutionary Computation, 19(2), 189–223. URL http://www.mitpressjournals.org/doi/pdf/10.1162/
EVCO_a_00025.
81. Lehman, J., & Stanley, K. O. (2011). Improving evolvability through novelty search and self-adaptation. In
Proceedings of the 2011 Congress on Evolutionary Computation (CEC 2011) (pp. 2693–2700). Washington, DC:
IEEE.
82. Lehman, J., & Stanley, K. O. (2014). Investigating biological assumptions through radical
reimplementation. Artificial Life, 21(1), 21–46.
83. Lenski, R., Ofria, C., Collier, T., & Adami, C. (1999). Genome complexity, robustness and genetic
interactions in digital organisms. Nature, 400(6745), 661–664.
84. Lenski, R. E. (1998). Get a life. Science, 280(5365), 849–850.
85. Lenski, R. E., Barrick, J. E., & Ofria, C. (2006). Balancing robustness and evolvability. PLoS Biology,
4(12), e428.
Artificial Life Volume 26, Number 2
303
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
86. Lenski, R. E., Ofria, C., Pennock, R. T., & Adami, C. (2003). The evolutionary origin of complex features.
Nature, 423(6936), 139–144.
87. Luke, S., & Panait, L. (2006). A comparison of bloat control methods for genetic programming.
Evolutionary Computation, 14(3), 309–344.
88. Madigan, M. T. (2000). Bacterial habitats in extreme environments. In J. Seckbach (Ed.), Journey to diverse
microbial worlds (pp. 61–72). Berlin: Springer.
89. Makarova, K. S., Aravind, L., Wolf, Y. I., Tatusov, R. L., Minton, K. W., Koonin, E. V., & Daly, M. J.
(2001). Genome of the extremely radiation-resistant bacterium Deinococcus radiodurans viewed from the
perspective of comparative genomics. Microbiology and Molecular Biology Reviews, 65(1), 44–79.
90. Mansanne, F., Carrere, F., Ehinger, A., & Schoenauer, M. (1999). Evolutionary algorithms as fitness
function debuggers. In Z. W. Rás & A. Skowron (Eds.), International Symposium on Methodologies for Intelligent
Systems (pp. 639–647). Berlin, Heidelberg: Springer.
91. Misevic, D., Frénoy, A., Parsons, D. P., & Taddei, F. (2012). Effect of public good properties on the
evolution of cooperation. In C. Adami, D. M. Bryson, C. Ofria, & R. T. Pennock (Eds.), Artificial Life 13
Conference Proceedings (pp. 218–225). Cambridge, MA: MIT Press.
92. Misevic, D., Ofria, C., & Lenski, R. E. (2006). Sexual reproduction reshapes the genetic architecture of
digital organisms. Proceedings of the Royal Society of London B: Biological Sciences, 273(1585), 457–464.
93. Mitri, S., Floreano, D., & Keller, L. (2009). The evolution of information suppression in communicating
robots with conflicting interests. Proceedings of the National Academy of Sciences of the USA, 106(37),
15786–15790.
94. Mnih, V., Badia, A. P., Mirza, M., Graves, A., Lillicrap, T., Harley, T., Silver, D., & Kavukcuoglu, K.
(2016). Asynchronous methods for deep reinforcement learning. In M. F. Balcan & K. Q. Weinberger
(Eds.), Proceedings of the 33rd International Conference on Machine Learning (pp. 1928–1937). New York: PMLR.
URL http://proceedings.mlr.press/v48/mniha16.html.
95. Mnih, V., Kavukcuoglu, K., Silver, D., Rusu, A. A., Veness, J., Bellemare, M. G., Graves, A., Riedmiller,
M., Fidjeland, A. K., Ostrovski, G., et al. (2015). Human-level control through deep reinforcement
learning. Nature, 518(7540), 529.
96. Moriarty, D. E., & Miikkulainen, R. (1995). Discovering complex Othello strategies through evolutionary
neural networks. Connection Science, 7(3), 195–209.
97. Moriarty, D. E., & Miikkulainen, R. (1996). Evolving obstacle avoidance behavior in a robot arm. In P. Maes,
M. J. Mataric, J.-A. Meyer, J. Pollack, & S. W. Wilson (Eds.), From animals to animats 4: Proceedings of the
Fourth International Conference on Simulation of Adaptive Behavior (pp. 468–475). Cambridge, MA: MIT Press.
98. Moriarty, D. E., & Miikkulainen, R. (1997). Forming neural networks through efficient and adaptive
co-evolution. Evolutionary Computation, 5(4), 373–399.
99. Mouret, J.-B., & Clune, J. (2015). Illuminating search spaces by mapping elites. arXiv preprint
arXiv:1504.04909 (pp. 1–15).
100. Nelson, B. (2013). University of Utah biologists surprised to discover “ultrafast recycling” at nerve
synapses. https://kuer.org/post/u-u-biologists-surprised-discover-ultrafast-recycling-nerve-synapses.
Accessed: 2016-07-28.
101. Nguyen, A., Yosinski, J., & Clune, J. (2016). Understanding innovation engines: Automated creativity and
improved stochastic optimization via deep learning. Evolutionary Computation, 24(3).
102. Nimwegen, E., Crutchfield, J. P., & Huynen, M. (1999). Neutral evolution of mutational robustness.
Proceedings of the National Academy of Sciences of the USA, 96(17), 9716–9720.
103. Nogueira, T., Rankin, D. J., Touchon, M., Taddei, F., Brown, S. P., & Rocha, E. P. (2009). Horizontal
gene transfer of the secretome drives the evolution of bacterial cooperation and virulence. Current Biology,
19(20), 1683–1691.
104. Nordin, P., Francone, F., & Banzhaf, W. (1996). Explicitly defined introns and destructive crossover in
genetic programming. In P. J. Angeline & K. E. Kinnear, Jr. (Eds.), Advances in genetic programming, Vol. 2
(pp. 111–134). Cambridge, MA: MIT Press.
105. Ofria, C., & Wilke, C. O. (2004). Avida: A software platform for research in computational evolutionary
biology. Artificial Life, 10(2), 191–229.
304
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
106. OʼReilly, U.-M., Wagy, M., & Hodjat, B. (2013). Ec-star: A massive-scale, hub and spoke, distributed
genetic programming system. In R. Riolo, E. Vladislavleva, M. Ritchie, & J. H. Moore (Eds.), Genetic
programming theory and practice X (pp. 73–85). Berlin: Springer.
107. OʼShea, D. C. (1991). Monochromatic quartet: A search for the global optimum. In G. N. Lawrence
(Ed.), International Lens Design Conference (pp. 548–554). Bellingham, WA: SPIE Press.
108. Peisajovich, S. G., & Tawfik, D. S. (2007). Protein engineers turned evolutionists. Nature Methods, 4(12),
991–994.
109. Pennock, R. T. (2000). Can Darwinian mechanisms make novel discoveries?: Learning from discoveries
made by evolving neural networks. Foundations of Science, 5(2), 225–238.
110. Pennock, R. T. (2007). Models, simulations, instantiations, and evidence: The case of digital evolution.
Journal of Experimental & Theoretical Artificial Intelligence, 19(1), 29–42.
111. Press Release. (2016). In Arctic winter, marine creatures migrate by the light of the moon. https://www.
eurekalert.org/pub_releases/2016-01/cp-iaw123015.php. Accessed: 2019-06-18.
112. Ray, T. (1992). Jʼai joué á Dieu et créé la vie dans mon ordinateur. Le Temps stratégique, 47, 68–81.
113. Ray, T. S. (1993). An evolutionary approach to synthetic biology: Zen and the art of creating life. Artificial
Life, 1(1–2), 179–209.
114. Rechenberg, I. (1973). Evolutionsstrategie–Optimierung technisher Systeme nach Prinzipien der biologischen Evolution.
Stuttgart, Germany: Frommann-Holzboog.
115. Roese, N. J., & Vohs, K. D. (2012). Hindsight bias. Perspectives on Psychological Science, 7(5), 411–426.
116. Runco, M. A., & Jaeger, G. J. (2012). The standard definition of creativity. Creativity Research Journal, 24(1),
92–96.
117. Salimans, T., Ho, J., Chen, X., & Sutskever, I. (2017). Evolution strategies as a scalable alternative to
reinforcement learning. arXiv preprint arXiv:1703.03864.
118. Schluter, D. (2000). The ecology of adaptive radiation. Oxford: Oxford University Press.
119. Schmalhausen, I. I. (1949). Factors of evolution: The theory of stabilizing selection. Chicago: University of Chicago
Press. 1986 edition.
120. Schmidt-Dannert, C., & Arnold, F. H. (1999). Directed evolution of industrial enzymes. Trends in
Biotechnology, 17(4), 135–136.
121. Schöner, M. G., Schöner, C. R., Simon, R., Grafe, T. U., Puechmaille, S. J., Ji, L. L., & Kerth, G. (2015).
Bats are acoustically attracted to mutualistic carnivorous plants. Current Biology, 25(14), 1911–1916.
122. Schulte, E., Forrest, S., & Weimer, W. (2010). Automated program repair through the evolution of
assembly code. In Proceedings of the IEEE/ACM International Conference on Automated Software Engineering
(pp. 313–316). New York: ACM.
123. Schuster, P., & Swetina, J. (1988). Stationary mutant distributions and evolutionary optimization. Bulletin of
Mathematical Biology, 50(6), 635–660.
124. Secretan, J., Beato, N., DʼAmbrosio, D. B., Rodriguez, A., Campbell, A., Folsom-Kovarik, J. T., &
Stanley, K. O. (2011). Picbreeder: A case study in collaborative evolutionary exploration of design space.
Evolutionary Computation, 19(3), 373–403.
125. Silva, S., & Costa, E. (2009). Dynamic limits for bloat control in genetic programming and a review of past
and current bloat theories. Genetic Programming and Evolvable Machines, 10(2), 141–179.
126. Sims, K. (1994). Evolving 3D morphology and behavior by competition. Artificial Life, 1(4), 353–372.
127. Sims, K. (1994). Evolving virtual creatures. In Proceedings of the 21st Annual Conference on Computer Graphics
and Interactive Techniques (pp. 15–22). New York: ACM.
128. Smaldino, P. E., & McElreath, R. (2016). The natural selection of bad science. Royal Society Open Science,
3(9), 160384.
129. Soltoggio, A., Bullinaria, J. A., Mattiussi, C., Durr, P., & Floreano, D. (2008). Evolutionary advantages of
neuromodulated plasticity in dynamic, reward-based scenarios. In S. Bullock, J. Noble, R. Watson, &
M. A. Bedau (Eds.), Proceedings of the 11th International Conference on Artificial Life (Alife XI) (pp. 569–576).
Cambridge, MA: MIT Press.
Artificial Life Volume 26, Number 2
305
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
J. Lehman et al.
The Surprising Creativity of Digital Evolution
130. Stanley, K. O., Bryant, B. D., & Miikkulainen, R. (2005). Real-time neuroevolution in the NERO video
game. IEEE Transactions on Evolutionary Computation, 9(6), 653–668.
131. Strahs, G. (1969). Biochemistry at 1000c: Explosive secretory discharge of bombardier beetles
(Brachinus). Science, 165(3888), 61–63.
132. Such, F. P., Madhavan, V., Conti, E., Lehman, J., Stanley, K. O., & Clune, J. (2017). Deep neuroevolution:
Genetic algorithms are a competitive alternative for training deep neural networks for reinforcement
learning. arXiv preprint arXiv:1712.06567.
133. Takagi, H. (2001). Interactive evolutionary computation: Fusion of the capacities of EC optimization and
human evaluation. Proceedings of the IEEE, 89(9), 1275–1296.
134. Taylor, J., Yudkowsky, E., LaVictoire, P., & Critch, A. (2016). Alignment for advanced machine learning
systems. Technical Report, Machine Intelligence Research Institute.
135. Taylor, T., Auerbach, J. E., Bongard, J., Clune, J., Hickinbotham, S., Ofria, C., Oka, M., Risi, S., Stanley,
K. O., & Yosinski, J. (2016). WebAL comes of age: A review of the first 21 years of artificial life on the
web. Artificial Life, 22(3), 364–407.
136. Thompson, A. (1996). An evolved circuit, intrinsic in silicon, entwined with physics. In T. Higuchi, M.
Iwata, & W. Liu (Eds.), International Conference on Evolvable Systems (pp. 390–405). Berlin, Heidelberg:
Springer-Verlag.
137. Turing, A. M. (1936). On computable numbers, with an application to the entscheidungsproblem.
American Journal of Mathematics, 58, 345–363.
138. van der Smagt, P. (1994). Simderella: A robot simulator for neuro-controller design. Neurocomputing, 6(2),
281–285.
139. Vincent, J. (2018). A video game-playing AI beat Q*bert in a way no oneʼs ever seen before. The Verge.
https://www.theverge.com/tldr/2018/2/28/17062338/ai-agent-atari-q-bert-cracked-bug-cheat.
140. Waddington, C. H. (1942). Canalization of development and the inheritance of acquired characters.
Nature, 150(3811), 563–565.
141. Wagner, G., Pavlicev, M., & Cheverud, J. M. (2007). The road to modularity. Nature Reviews Genetics, 8(12),
921–931.
142. Watanabe, S., Rost, B. R., Camacho-Pérez, M., Davis, M. W., Söhl-Kielczynski, B., Rosenmund, C., &
Jorgensen, E. M. (2013). Ultrafast endocytosis at mouse hippocampal synapses. Nature, 504(7479), 242.
143. Watson, R. A., Ficici, S. G., & Pollack, J. B. (2002). Embodied evolution: Distributing an evolutionary
algorithm in a population of robots. Robotics and Autonomous Systems, 39(1), 1–18.
144. Watson, R. A., Ficiei, S., & Pollack, J. B. (1999). Embodied evolution: Embodying an evolutionary
algorithm in a population of robots. In Proceedings of the 1999 Congress on Evolutionary Computation
(pp. 335–342). Piscataway, NJ: IEEE.
145. Weimer, W. (2013). Advances in automated program repair and a call to arms. In G. Ruhe & Y. Zhang
(Eds.), Proceedings of Search Based Software Engineering—5th International Symposium (pp. 1–3). Berlin,
Heidelberg: Springer-Verlag.
146. Wilke, C., Wang, J., Ofria, C., Lenski, R., & Adami, C. (2011). Evolution of digital organisms at high
mutation rates leads to survival of the flattest. Nature, 412(6844), 331–333.
147. Wilson, E. O. (1999). The diversity of life. New York: W. W. Norton.
148. Yedid, G., & Bell, G. (2002). Macroevolution simulated with autonomously replicating computer
programs. Nature, 420(6917), 810.
149. Zhao, H., & Arnold, F. H. (1997). Combinatorial protein design: Strategies for screening protein libraries.
Current Opinion in Structural Biology, 7(4), 480–485.
306
Artificial Life Volume 26, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
a
r
t
l
/
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
6
2
2
7
4
1
8
9
6
0
7
1
a
r
t
l
/
_
a
_
0
0
3
1
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3