The Role of Action–Effect Contingency on Sensory
Attenuation in the Absence of Movement
Nathan Han1
, Bradley N. Jack2, Gethin Hughes3, and Thomas J. Whitford1
Abstract
■ Stimuli that have been generated by a person’s own willed
motor actions generally elicit a suppressed electrophysiologi-
cal, as well as phenomenological, response compared with
identical stimuli that have been externally generated. This
well-studied phenomenon, known as sensory attenuation,
has mostly been studied by comparing ERPs evoked by
self-initiated and externally generated sounds. However, most
studies have assumed a uniform action–effect contingency, in
which a motor action leads to a resulting sensation 100% of
the time. In this study, we investigated the effect of manipu-
lating the probability of action–effect contingencies on the
sensory attenuation effect. In Experiment 1, participants
watched a moving, marked tickertape while EEG was recorded.
In the full-contingency (FC) condition, participants chose
whether to press a button by a certain mark on the tickertape.
If a button press had not occurred by the mark, a sound would
be played a second later 100% of the time. If the button was
pressed before the mark, the sound was not played. In the
no-contingency (NC) condition, participants observed the same
tickertape; in contrast, however, if participants did not press
the button by the mark, a sound would occur only 50% of the
time (NC-inaction). Furthermore, in the NC condition, if a par-
ticipant pressed the button before the mark, a sound would
also play 50% of the time (NC-action). In Experiment 2, the
design was identical, except that a willed action (as opposed
to a willed inaction) triggered the sound in the FC condition.
The results were consistent across the two experiments:
Although there were no differences in N1 amplitude between
conditions, the amplitude of the Tb and P2 components were
smaller in the FC condition compared with the NC-inaction
condition, and the amplitude of the P2 component was also
smaller in the FC condition compared with the NC-action con-
dition. The results suggest that the effect of contingency on
electrophysiological indices of sensory attenuation may be
indexed primarily by the Tb and P2 components, rather than
the N1 component which is most commonly studied. ■
INTRODUCTION
Stimuli that have been generated by a person’s own willed
motor actions generally elicit a suppressed electrophysio-
logical, as well as phenomenological, response compared
with physically identical stimuli that have been externally
generated (Hughes, Desantis, & Waszak, 2013). This
well-studied phenomenon, known as sensory attenuation,
has mostly been studied by comparing ERPs evoked by
self-initiated and externally generated sounds (Pinheiro,
Schwartze, Gutiérrez-Domínguez, & Kotz, 2020; Pinheiro,
Schwartze, Gutierrez, & Kotz, 2019; Horváth, 2015;
Gentsch & Schütz-Bosbach, 2011; Ford, Roach, & Mathalon,
2010; Baess, Widmann, Roye, Schröger, & Jacobsen,
2009; Aliu, Houde, & Nagarajan, 2008; Martikainen,
Kaneko, & Hari, 2005; Schafer & Marcus, 1973). A large
body of literature shows that certain components of the
auditory-evoked potential are reduced in amplitude when
participants hear sounds initiated by their own motor
actions (e.g., overt speech, button press elicited tones),
1UNSW Sydney, 2Australian National University, Canberra,
Australia, 3University of Essex, Colchester, United Kingdom
© 2022 Massachusetts Institute of Technology
compared with when they passively listen to the same
sounds. This effect has been most commonly observed
with the N1 component ( Jack et al., 2021; Harrison et al.,
2021; Klaffehn, Baess, Kunde, & Pfister, 2019; Pinheiro
et al., 2019; Elijah, Le Pelley, & Whitford, 2018; Neszmélyi
& Horváth, 2017; Mifsud et al., 2016; Oestreich et al., 2016;
van Elk, Salomon, Kannape, & Blanke, 2014; Bäß, Jacobsen,
& Schröger, 2008) but has also been identified with the
Tb (SanMiguel, Widmann, Bendixen, Trujillo-Barreto, &
Schroger, 2013; Saupe, Widmann, Trujillo-Barreto, &
Schröger, 2013) and P2 components (Horváth & Burgyán,
2013; Knolle, Schröger, Baess, & Kotz, 2012).
Within the sensory attenuation literature, a topic that
has remained relatively unexplored is the role of stimulus
contingency (Horváth, 2015). Here, we operationalize
contingency as the probability of a stimulus (in this case,
a sound) occurring because of a willed action (or willed
inaction). Although there have been some attempts to
investigate contingency in the sensory attenuation litera-
ture, most previous studies have operationalized contin-
gency in terms of temporal contingency; specifically, most
previous studies have manipulated contingency by varying
the temporal delay between the motor action and sound.
Journal of Cognitive Neuroscience 34:8, pp. 1488–1499
https://doi.org/10.1162/jocn_a_01867
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
For example, Bäß et al. (2008) investigated the effect of
tone frequency and onset predictability on N1 suppression.
In their motor–auditory and auditory-only conditions, fre-
quency and onset predictability were manipulated in a 2 ×
2 design. Sounds could be (a) predictable in both fre-
quency (a 1000-Hz sound) and onset, in which the sound
immediately followed the button press; (b) unpredictable
in frequency (ranging from 400 to 1990 Hz) but predict-
able in onset; (c) predictable in frequency but unpredict-
able in onset, in which a random delay of 500–1000 msec
was imposed between the action and effect; and (d)
unpredictable in both frequency and sound onset. They
found that N1 suppression occurred regardless of the pre-
dictability of the frequency and onset of the sound.
Pinheiro et al. (2019) conducted a study where various
delays were inserted between the action and sound within
action–effect contingencies as to induce temporal predic-
tion errors. In 30% of trials, sounds that followed the but-
ton press were presented with a delay of either 50, 100, or
250 msec. They found that N1 suppression still occurred
with delays of up to 100 msec between the action and
the sound.
Another method of manipulating contingency is by
changing the probability of a sound resulting from an
action. Typical self-stimulation experiments often have
action–effect contingencies of 100%, meaning that actions
(e.g., button presses) lead to sounds in 100% of trials
(Horváth, 2015). Action–effect contingencies—especially
in ecological settings—do not always occur in the uniform
manner that they do in the typical laboratory setting. For
example, Knolle, Schröger, and Kotz (2013) tested
whether N1 suppression effects were larger when self-
generated sounds were correctly predicted rather than
when they were not. Self- and externally generated sounds
were categorized as frequent standard-pitched sounds (70%
probability) or infrequent deviant-pitched sounds (30%
probability). Although N1 suppression occurred for both
standard and deviant sounds, the size of the N1 suppres-
sion effect was smaller for deviant sounds compared with
the standard sounds, indicating that predictions for
self-generated sounds contained specificity. In another
study, SanMiguel, Widmann, et al. (2013) recorded EEG
from human participants while they pressed a button that
either consistently (88%), inconsistently (50%), or never
(0%) initiated a sound. During conditions where button
presses consistently produced a sound (88%), EEG
responses were also obtained when sounds were omitted
in 12% of trials. Their results showed that during such
omitted trials, EEG responses showed a pattern of activity
that shared brain sources and time course with exogenous
responses to trials where there were actual stimuli. Based
on these results, the authors argued that the brain acti-
vates a template of its response to predicted stimuli
before sensory input arrived at our senses.
Ecologically, people sometimes incorrectly predict the
outcome of an action (e.g., when trying to guess the out-
come of a button press on a slots machine). Essentially,
manipulation of action–effect contingency is a method
of modifying participants’ confidence in their predic-
tions regarding the sensory consequences of their
actions. In essence, both forms of operationalization—by
which we mean both manipulating of temporal onset versus
the manipulating the probability of sounds occurring—
perform the function of reducing the participant’s confi-
dence that the expected sensory event will occur.
In this study, we manipulated action–effect contingency
by varying the probability that actions (or inaction) would
result in a tone being played. In these two experiments,
participants observed a short animation and, on each trial,
were required to decide whether to press a button (the
space bar of the keyboard). This decision determined
whether a sound would subsequently be presented after
a significant delay. The two experiments were based on
the experimental paradigm used in Han, Jack, Hughes,
Elijah, and Whitford (2021), which demonstrated that
willed inaction might also result in sensory attenuation.
In both experiments, there were two conditions: the
full-contingency (FC) condition and the no-contingency
(NC) condition. In the FC condition, sounds always
followed a willed inaction (in Experiment 1) or always
followed the button press (in Experiment 2). In contrast,
sounds followed willed inaction (Experiment 1) or a
button press (Experiment 2) only 50% of the time in
the NC condition. The definition of contingency—the
probability of an event occurring as a result of an action
minus the probability of an event occurring in the
absence of that action—was taken from Elsner and
Hommel (2004).
Like most previous studies in the sensory attenuation
literature, we analyzed the N1 and P2 components of the
ERP. We also included the Tb component, which is
believed to reflect activity of the secondary auditory cortex
(Rihs et al., 2013; Tonnquist-Uhlen, Ponton, Eggermont,
Kwong, & Don, 2003; Gallinat et al., 2002; Wolpaw &
Penry, 1975) and which has been found to be dependent
on the extent to which participants had agency over the
generation of auditory stimuli (Han et al., 2021). We ana-
lyzed all ERPs time-locked onto the sound. Previous
research using the traditional self-stimulation paradigm
has demonstrated suppression across the N1, Tb, and P2
components in the active condition relative to the passive
condition (SanMiguel, Todd, & Schröger, 2013; Knolle
et al., 2012). However, given that our studies use a differ-
ent methodology compared with the traditional
self-stimulation paradigm (Schafer & Marcus, 1973), we
do not expect to find any evidence of N1 suppression in
this study. This hypothesis was based on the results of
Han et al. (2021), which used a similar experimental
methodology. Consequently, effects of contingency were
expected to be mediated by the Tb and P2 components.
We therefore hypothesized that Tb amplitudes would be
smaller and P2 amplitudes to be larger when probabilities
of sound occurrence were 100% relative to when proba-
bilities were set at 50%.
Han et al.
1489
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
METHODS
Experiment 1
Participants
Forty undergraduate students from the University of New
South Wales (Sydney, Australia) participated in the study
in exchange for course credit (n = 40). All participants
gave written informed consent before the experiment.
Two participants were removed from analysis because
of insufficient number of artifact-free epochs (as
described in the EEG Recording and Analysis section),
leaving a final sample of n = 38 participants (mean
age = 21 years, SD = 7.5 years, 16 women). Study sam-
ple size was based on prior research by Han et al. (2021),
which used a similar experimental paradigm. The study
was approved by the Human Research Ethics Advisory
Panel (Psychology) at the University of New South Wales.
Stimuli, Materials, and Procedure
The audio stimulus was a sinusoid tone of frequency of 1000
Hz, 100-msec duration, with a 5-msec linear rise/fall time.
Audio stimuli were sent to participants through Sennheiser
HD 210 headphones. Audio input/output was controlled
by a specially written MATLAB script and was delivered
via an AudioFile Stimulus Processor (Cambridge
Research Systems). Participants made responses by press-
ing the space bar of a low-latency keyboard (DuckyShine 4,
1000 Hz report rate). Visual stimuli were displayed on a
high-performance 24-in. monitor (BenQ XL2420T).
During each trial, participants observed a visual anima-
tion, which was adapted from the paradigm employed by
Whitford et al. (2017) and Jack et al. (2019) and identical to
that employed by Han et al. (2021). The animation lasted
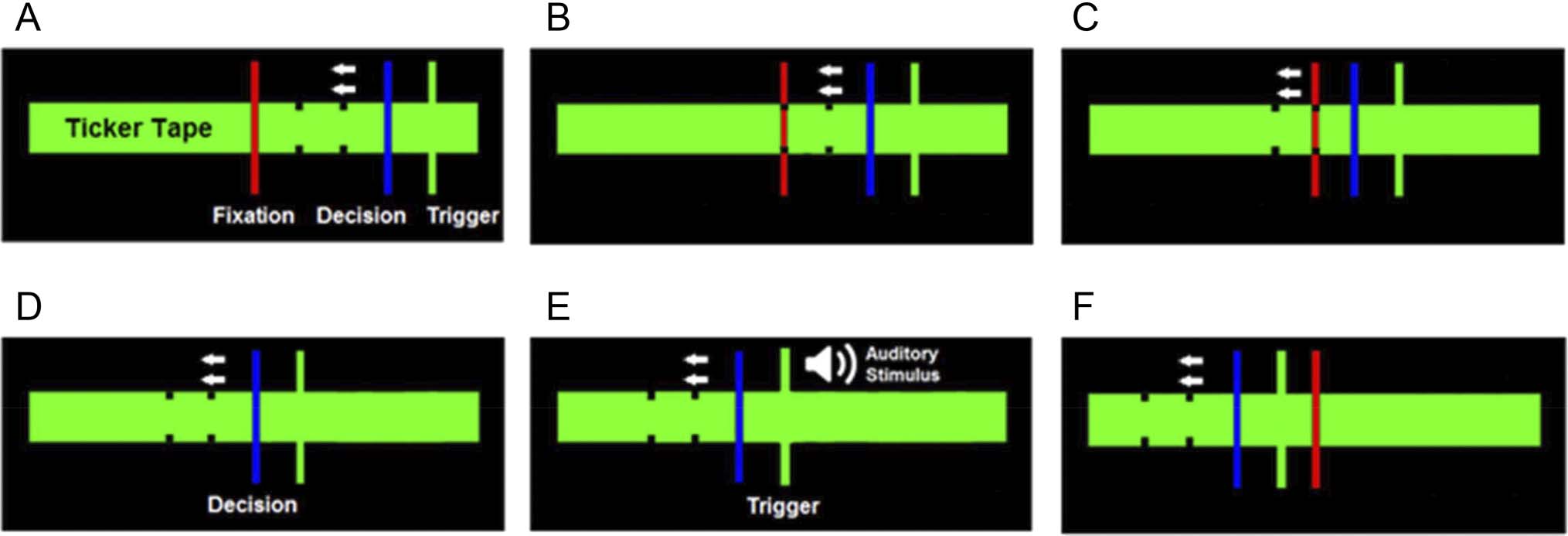
for about 6 sec. A schematic of the animation is presented
in Figure 1. The animation consisted of a central red fixa-
tion line that sat in the middle of a green horizontal bar,
which is referred to as the ticker tape. Participants were
instructed to keep their eyes fixated on the fixation line
during the trial. There was also a blue decision line and
a green trigger line located on the right side of the ticker
tape. The trigger line was initially positioned on the far
right-hand side of the ticker tape; the decision line sat to
the left of the trigger line (Figure 1A).
Upon commencement of the trial, after a 1-sec delay,
both the decision line and the trigger line started to move
toward the fixation line at a constant rate of 6.5°/sec (visual
angle). The decision line intersected the fixation line after
approximately 3 sec. The trigger line intersected the fixa-
tion line after approximately 4 sec; at this point, the audi-
tory stimulus was presented (depending on the trial, as
described below). The lines continued to move for
another 1 sec, before the trial was completed.
There were two conditions in the experiment: the FC
condition and the NC condition. In the FC condition, par-
ticipants had the option of pressing the space bar on the
keyboard before the decision line intersected the fixation
line (hereon referred to as the decision time). Partici-
pants were told that if they did not press the button by
the decision time (Figure 1B)—that is, if they performed
Figure 1. A schematic of the experimental protocol. Participants were instructed to fixate their eyes on the central red fixation line (A). After a 1-sec
delay, the blue decision line and the green trigger line moved slowly toward the central red fixation line at a rate of 6.5°/sec (visual angle; B–C).
Participants were told that they had the option of pressing the space bar of the keyboard by the decision time (D). In the FC condition in Experiment 1,
if the participant did not press the space bar before the decision line overlapped with the fixation line, this would cause the audio stimulus to be
played at trigger time (E). If the participant pressed the space bar during this time frame, the audio stimulus would not play at the trigger time. In the
FC condition in Experiment 2, this contingency was reversed, such that if participants did not press the space bar before the decision time, this would
inhibit the audio stimulus from being played at the trigger time. Vice versa, if participants pressed the space bar during this time frame, this would
cause the audio stimulus to be played at the trigger time. In the NC condition in both Experiments 1 and 2, the probabilities were set so that audio
onset would only follow button presses 50% of the time. Vice versa, audio onset would also occur to inaction 50% of the time. Participants were not
told that the probability in the NC condition was 50% but were told that sounds “may or may not be played.” The lines continued to move for another
1 sec, before the animation concluded and the trial concluded, and the trial was completed (F).
1490
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
a willed inaction—this would cause the audio stimulus to
be played at the exact moment that the trigger line inter-
sected the fixation line (hereon referred to as the trigger
time; Figure 1C). If they chose to press the space bar
before the decision time, this prevented the audio stimu-
lus from being played at the trigger time. The probabilities
were set so that inaction would always cause the audio
stimulus to be played at the trigger time; conversely,
pressing the button would always cause no sound to be
played at the trigger time. Participants were asked to press
the space bar on approximately half the trials while trying
not to conform to an obvious pattern of responses. At
the start of every trial, participants were reminded (by
means of instructions on the screen) as to what their
options were and what the consequences would be.
In the NC condition, participants were given instruc-
tions that were nearly identical to the instructions in the
FC condition. However, the probabilities were set so that
audio onset would only follow button presses 50% of the
time. Conversely, audio onset would also occur to inaction
50% of the time. Essentially, whether the participant did or
did not hear a sound during any given trial was random.
Participants were not told of the exact probabilities but
were instead told that sounds “may or may not” occur as
a result of their actions in the instructions.
The experiment consisted of five FC blocks and 10 NC
blocks, totaling 15 blocks for the whole experiment. For
ease of reference, we make the following distinction
between three different trial types:
(cid:129) FC trials in which participant inactions resulted in a
sound 100% of the time,
(cid:129) NC-action trials in which participant action resulted in
a sound 50% of the time, and
(cid:129) NC-inaction trials in which participant inaction resulted
in a sound 50% of the time. It is important to note that
NC-action and NC-inaction trials occurred within the
same block
There were twice the number of NC blocks compared
with FC blocks in the experiment. This was done to achieve
approximately equal number of usable epochs, given that
the sound was only presented on approximately 50% of
trials in the NC condition. We only analyzed trials in which
the auditory stimulus was played. Each block contained
24 trials. The order of the blocks alternated between the
FC and NC blocks so that there were two NC blocks for
every one FC block. Within the grouping of three blocks
(two NC blocks and one FC block), the order was counter-
balanced between participants. The starting block was also
counterbalanced between participants.
EEG Recording and Analysis
EEG was recorded with a BioSemi ActiveTwo system from
64 Ag/EgCl active electrodes (P1, FPz, FP2, AF7, AF3, AFz,
AF4, AF8, F7, F5,F3, F1, Fz, F2, F4, F6, F8, FT7, FC5, FC3,
FC1, FCz, FC2, FC4, FC6, FT8,T7, C5, C3, C1, Cz, C2, C4,
C6, T8, TP7, CP5, CP3, CP1, CPz, CP2, CP4,CP6, TP8, P9, P7,
P5, P3, P1, Pz, P2, P4, P6, P8, P10, PO7, PO3, POz,PO4, PO8,
O1, Oz, O2, Iz). A vertical EOG was recorded by placing an
electrode above and below the left eye; a horizontal EOG
was recorded by placing an electrode on the outer canthus
of each eye. Electrodes were also placed on each mastoid
and the nose. During data acquisition, the reference was
composed of sites CMS and DRL, and the sampling rate
was 2048 Hz.
For data analyses, we re-referenced the EEG data off-line
to the nose electrode, as is common in studies investigat-
ing the components of interest and necessary for extract-
ing the Tb component (SanMiguel, Widmann, et al., 2013;
Näätänen & Picton, 1987). Data were band-pass filtered
from 0.1 to 30 Hz using a half-amplitude phase shift-free
Butterworth filter and then notch-filtered (50 Hz) to
remove mains artifact. The filtered data were segmented
into 500-msec epochs, from −100 msec prestimulus to
400 msec poststimulus. Only trials in which the auditory
stimulus were played were analyzed. Epochs were
baseline-corrected to the mean voltage from −100 to
0 msec. We corrected the epochs for eye blinks and move-
ment artifacts using the technique described by Gratton,
Coles, and Donchin (1983) and Miller, Gration, and Yee
(1988). We excluded all epochs with signals exceeding
peak-to-peak amplitudes of 200 μV and had a maximal
allowed voltage step of 50 μV/msec. We analyzed
the amplitudes of the N1, Tb, and P2 components of the
auditory-evoked potential, which were calculated as the
average voltage within time windows (30 msec width),
the centers of which were defined using the collapsed
localizer approach (Luck & Gaspelin, 2017). The collapsed
localizer approach is a technique whereby one first
averages (or collapses) the ERP waveforms across all
conditions for all participants. The components of interest
(e.g., N1, Tb, P2) are identified on this collapsed wave-
form, and a time window is centered around these peaks,
which is then used for the statistical analysis of the original
(or uncollapsed) waveforms (Luck & Gaspelin, 2017).
For the N1, Tb, and P2 components, mean voltage in the
analysis window was submitted to paired samples t tests. All
paired samples t tests were analyzed two-tailed. For each
component, there were two contrasts of interest: (1) FC
versus NC-action and (2) FC versus NC-inaction. Electrodes
of interest for the N1 component were Fz, FCz, and Cz,
whereas the electrodes of interest for the P2 component
were FCz, Cz, and CPz. For the N1 and P2 components, elec-
trodes and analyses were chosen to be consistent with Han
et al. (2021), Whitford et al. (2017), and Jack et al. (2019).
Electrodes for the Tb component (T7 and T8) were based
on recommendations by Tonnquist-Uhlen et al. (2003) and
SanMiguel, Widmann, et al. (2013).
Experiment 1 Results
The summary results of Experiment 1 are illustrated in
Figure 4. In the FC condition, on average, participants
Han et al.
1491
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
opted for inaction, which led to a sound 61.3 (SD = 4.9)
times. After accounting for artifact exclusion, this led to an
average of 58.5 (SD = 7.1) epochs used for analysis.
In the NC condition, on average, there was a distribu-
tion of 60.8 sounds (SD = 6.9) because of inaction
and 62.4 sounds (SD = 6.7) because of action. After
accounting for artifact exclusion, this led to an average of
57.2 (SD = 7.4) epochs used for analysis in the NC-inaction
condition and an average of 59.8 (SD = 6.9) epochs used
for analysis in the NC-action condition.
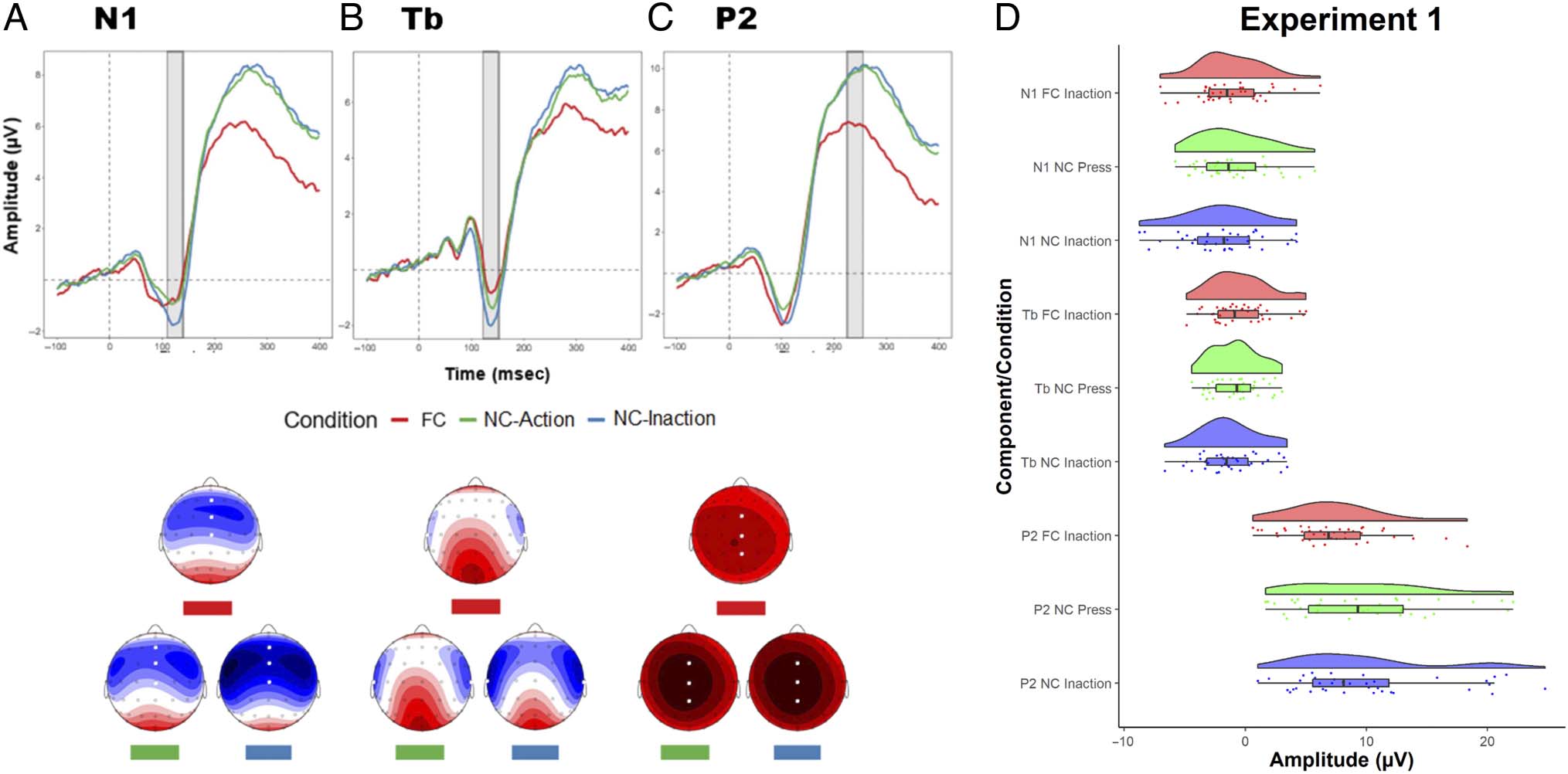
N1. Figure 2A shows the grand average N1 component
elicited in the FC and NC conditions. The time window
for the N1 analysis was 109.5–139.5 msec. The contrast
comparing FC versus NC-inaction did not reach signifi-
cance, t(37) = 1.769, p = .085, d = 0.29. Similarly, the
contrast comparing FC versus NC-action also did not
reach significance, t(37) = −.336, p = .739, d = 0.06.
These results indicate that the N1 amplitude did not differ
between the FC and NC conditions, regardless of whether
the sounds in the NC condition were associated with an
inaction or a button press.
t(37) = 1.162, p = .253, d = 0.19. The results indicate
that the Tb amplitude of the FC condition was sup-
pressed relative to the amplitude of the NC condition,
but only in the case where sounds in the NC condition
were elicited via inaction. However, Tb amplitude in
the FC condition did not differ from Tb amplitude in
the NC condition when sounds in the NC condition were
elicited by button press.
P2. Figure 2C shows the P2 component analysis elicited
in the FC and NC conditions. The time window for the P2
analysis was 224.7–254.7 msec. The contrast comparing FC
versus NC-inaction was significant, t(37) = −3.523, p =
.001, d = 0.57, as was the contrast comparing FC versus
NC-action, t(37) = −3.447, p = .001, d = 0.56. The results
indicate that the P2 amplitude of the FC condition was
strongly suppressed relative to the amplitude in the NC
condition both when sounds in the NC condition were
elicited by button press or when they were elicited via a
willed inaction.
Experiment 1 Discussion
Tb. Figure 2B shows the Tb component analysis elicited
in the FC and NC conditions. The time window for the Tb
analysis was 122.2–152.2 msec. The contrast comparing
FC versus NC-inaction reached significance, t(37) =
2.586, p = .014, d = 0.42, whereas the contrast com-
paring FC versus NC-action did not reach significance,
In Experiment 1, participants completed a variation of the
traditional self-stimulation task in which the sound was
not time-locked to a motor action. In the FC condition,
participants were instructed that choosing not to per-
form an action by the decision time would cause a
sound to be played at the subsequent trigger time. In
Figure 2. Experiment 1: Waveforms showing ERPs elicited by the FC condition and the NC-action and NC-inaction conditions in addition to
corresponding topographic mappings. White dots illustrate the electrodes used in the analysis. (A) The N1 component was measured at electrodes
Fz, FCz, and Cz, with time window 109.5–139.5 msec. (B) The Tb component was measured at electrodes T7 and T8, with time window
122.2–152.2 msec. (C) The P2 component was measured at electrodes FCz, Cz, and CPz, with time window 224.7–254.7 msec. (D) Raincloud graph
containing density plots and scatter plots of mean amplitudes for the N1, Tb, and P2 components for the FC and NC conditions.
1492
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
the NC condition, participants were given the same
choice, but the probability under which sounds occurred
was only 50% following a participant choice. That is, in the
NC condition, electing not to press a button (NC-inaction)
led to a sound on 50% of trials, whereas electing to press
the button (NC-action) also led to the sound being pre-
sented on 50% of trials. The results revealed that the
amplitude of the N1 components did not differ between
the FC and NC conditions. The amplitudes of the Tb con-
dition did differ, with Tb suppressed in the FC condition
relative to the NC-inaction condition, but not the
NC-action condition. The P2 component in the FC condi-
tion was strongly suppressed relative to both the
NC-inaction and NC-action conditions.
In Experiment 1, participants were instructed that the
performance of a willed inaction would elicit a sound in
the FC condition. They were also instructed that a willed
inaction “may or may not” cause a sound to be played in
the NC-inaction condition. The aim of Experiment 2 was
to investigate whether the key results of Experiment 1
(i.e., suppression of the Tb and P2 components in the
FC condition relative to the NC conditions) would be
replicated if the instructions were reversed, that is, if
participants were instructed that the performance of a
willed action (button press) would elicit a sound in
the FC condition. Furthermore, including this second
experiment might have helped to elucidate whether
there was any relation to the Tb suppression effect
being selective to the NC-inaction condition and the fact
that it was inaction that triggered sounds in the FC
condition.
Experiment 2
Participants
Forty-nine undergraduate students from the University of
New South Wales (Sydney, Australia) participated in the
study in exchange for course credit. The participant
samples of Experiments 1 and 2 were unique; none of
the participants in Experiment 2 had previously partici-
pated in Experiment 1. All participants gave written
informed consent before the experiment. Four partici-
pants were removed from analysis because of insufficient
number of artifact-free epochs (as described in the EEG
Recording and Analysis section), leaving a final sample of
n = 45 participants (mean age = 19 years, SD = 1.8 years,
24 women). Study sample size was based on Experiment 1
and on prior research by Han et al. (2021), which used a
similar experimental paradigm. The study was approved by
the Human Research Ethics Advisory Panel (Psychology)
at the University of New South Wales.
Stimuli, Materials, and Procedure
The stimuli and materials were identical to Experiment 1.
The only difference between the experiments was the
action–effect contingency in the FC condition. Specifi-
cally, in Experiment 1, the audio stimulus was played if
the participant opted not to press the button before the
decision time, and participants were informed of this
fact. In Experiment 2, this contingency was reversed:
The audio stimulus was played if and only if the partic-
ipant elected to press the button before the decision
time, and participants were informed of this fact. As in
Experiment 1, the audio stimulus was played at the trig-
ger time, which occurred 1 sec after the decision time.
There was no change to the instructions in the NC
condition.
EEG Recording and Analysis
The EEG recording and analysis were identical to Experi-
ment 1.
Experiment 2 Results
In the FC condition, on average, participants opted for a
button press, which led to a sound 62.4 (SD = 4.4) times.
After accounting for artifact exclusion, this led to an aver-
age of 60.3 (SD = 5.4) epochs used for analysis. In the NC
condition, on average, there was a distribution of 62.8
sounds (SD = 6.4) because of action and 57.3 sounds
(SD = 6.0) because of inaction. After accounting for arti-
fact exclusion, this led to an average of 60.3 (SD = 7.1)
epochs used for analysis in the NC-action condition and
an average of 54.8 (SD = 7.2) epochs used for analysis in
the NC-inaction condition.
N1. Figure 3A shows the N1 component analysis elicited
in the FC and NC conditions. The time window for the N1
analysis was 85.6–115.6 msec. The contrast comparing FC
versus NC-action did not reach significance, t(44) =
−1.102, p = .276, d = 0.16. Similarly, the contrast compar-
ing FC versus NC-inaction did not reach significance,
t(44) = −1.766, p = .084, d = 0.26.
Tb. Figure 3B shows the Tb component analysis elic-
ited in the FC and NC conditions. The time window
for the Tb analysis was 133.9–163.9 msec. The contrast
comparing FC versus NC-action did not reach signifi-
cance, t(44) = 0.801, p = .427, d = 0.12. However,
the contrast comparing FC versus NC-inaction was statis-
tically significant, t(44) = 2.126, p = .039, d = 0.32, with
the NC-inaction condition showing a larger Tb amplitude
than the FC condition.
P2. Figure 3C shows the P2 component analysis elicited
in the FC and NC conditions. The time window for the P2
analysis was 175.9–205.9 msec. The contrast comparing FC
versus NC-action reached significance, t(44) = −2.208,
p = .032, d = 0.33, with the NC-action condition showing
a larger P2 amplitude than the FC condition. Similarly, the
contrast comparing FC versus NC-inaction also reached
Han et al.
1493
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Experiment 2: Waveforms showing ERPs elicited by the FC condition and the NC-action and NC-inaction conditions and the corresponding
topographic mappings. White dots illustrate the electrodes used in the analysis. (A) The N1 component was measured at electrodes Fz, FCz, and Cz,
with time window 85.6–115.6 msec. (B) The Tb component was measured at electrodes T7 and T8, with time window 133.9–163.9 msec. (C) The P2
component was measured at electrodes FCz, Cz, and CPz, with time window 175.9–205.9 msec. (D) Raincloud graph containing density plots and
scatter plots of mean amplitudes for the N1, Tb, and P2 components for the FC and NC conditions.
significance, t(44) = −3.305, p = .002, d = 0.49, with the
NC-inaction condition showing a larger P2 amplitude than
the FC condition.
GENERAL DISCUSSION
The results of both experiments are summarized in
Figure 4. In this study, participants were required to either
performed a willed inaction (in Experiment 1) or a willed
action (a button press, in Experiment 2) for a sound to be
played a second or more later. Participants’ electrophysio-
logical response to the sound was then measured with
EEG. The pattern of results was identical across the two
experiments. N1 amplitudes did not differ significantly
across the FC and NC conditions, regardless of whether
the audio stimulus was triggered by an inaction (NC-
inaction) or a button press (NC-action). The P2 compo-
nent was attenuated in the FC condition relative to both
the NC-inaction and NC-action conditions, in both exper-
iments. The results of the P2 component analyses suggest
Figure 4. Bar graphs of Experiments 1 and 2 illustrating mean amplitudes for the N1, Tb, and P2 components for the FC and NC conditions. Error
bars show the SEM. Asterisks represent levels of significance (*p < .05, **p < .01).
1494
Journal of Cognitive Neuroscience
Volume 34, Number 8
that effect of contingency on electrophysiological indices
of sensory attenuation may be indexed by the P2 compo-
nent. The Tb component was attenuated in the FC
condition relative to the NC-inaction condition in both
experiments. The results of the Tb analyses suggest that
the Tb component may index the increased sense of
agency associated with the 100% action–effect contin-
gency in the FC condition. Given that Tb amplitude has
been shown to be modulated by the degree to which par-
ticipants had agency over sound generation (Han et al.,
2021), this may explain why the Tb amplitudes in the
NC-action condition were driven closer toward the FC
condition in the current experiment. Even though in
actual fact, the probability of a tone resulting from a
button press was only 50% (and therefore had no actual
contingency, as inaction also resulted in a sound 50% of
the time), the existence of the button press meant that a
tenuous connection could be made between the action and
effect (similarly to the phenomenon of illusory control;
Harris & Osman, 2012; Dixon, 2000). However, it should
be noted that it is uncommon for evaluative (or
higher-order cognitive) processes—such as judgments of
agency—to influence ERP amplitude until after 300 msec
(Wascher & Beste, 2010). According to Synofzik, Vosgerau,
and Newen’s (2008) multifactorial account of agency, the
feeling of agency is the sense of agency someone experi-
ences when they perform a motor action that is followed
by a sensory event. This is what the literature typically
refers to when discussing agency within the context of
internal forward models (Synofzik et al., 2008). The judg-
ment of agency, on the other hand, requires an explicit
cognitive judgment of one’s agency and does not rely on
sensorimotor indicators. As such, given that there was no
explicit measure of agency in this study, it is not exactly
clear whether participants were experiencing feelings or
forming judgments of agency. In a previous study that
used a similar ticker tape design (which included two
experiments, the first of which used inaction to elicit
sounds; Han et al., 2021), it was argued that participants
formed judgments of agency rather than feelings of
agency because, in the first experiment, it was inaction
that led to a sound and, in the second experiment, sounds
were not time-locked to motor actions (with a gap of at
least 1 sec between action and effect). However, much like
the current study, in Han et al. (2021), there was no direct
measure of the sense of agency, leaving the possibility that
the ERP modulations reflect differences in the feelings of
agency rather than judgments of agency. There is evidence
that inaction can lead to temporal binding (Weller, Schwarz,
Kunde, & Pfister, 2020), a phenomenon that arguably
reflects feelings of agency as it occurs on a perceptual level.
We suggest that more research be done to investigate the
link between inaction–effects and sense of agency.
In both experiments, we found no difference in N1
amplitude between the FC and NC conditions, whether
triggered by inaction (NC-inaction) or a button press
(NC-action). Considering the different probabilities under
which sounds occurred, one might have expected differ-
ences in N1 amplitude between the FC and NC conditions.
Under the predictive coding account of perception, neu-
ronal responses to stimuli have been argued to reflect pre-
diction errors (Friston, 2005). In accordance with this
account, stimuli that are more predictable trigger smaller
neuronal responses than unpredictable stimuli because
they result in smaller prediction errors (SanMiguel,
Widmann, et al., 2013; Timm, SanMiguel, Saupe, &
Schröger, 2013; Bendixen, SanMiguel, & Schröger,
2012; Schafer, Amochaev, & Russell, 1981). For example,
Roth, Ford, Lewis, and Kopell (1976) investigated the
effects of probability on auditory processing by delivering
auditory stimuli in which a sequence of regularly occur-
ring 65-dB pip sounds were randomly interrupted by
white noise bursts. Half of the white noise bursts were
preceded by a warning tone (and were thus highly pred-
icable), and half were not (and were thus not predict-
able). They found that passively presented unpredictable
white noise bursts elicited larger N1 amplitudes than pas-
sively presented predictable noise bursts. Given the
results of Roth et al. (1976), one might expect that the
N1 amplitude generated in the NC conditions would be
larger than in the FC condition, considering that the occur-
rence of the sounds was less predictable in the NC condi-
tions. However, this discrepancy may be because the type
of predictability manipulated between the current study
and the one conducted by Roth et al. (1976) was different.
It has been demonstrated that sensory inputs are not only
predicted/mispredicted but can also be unpredicted (Hsu,
Hämäläinen, & Waszak, 2018; Hsu, Le Bars, Hamalainen, &
Waszak, 2015; Arnal & Giraud, 2012). Mispredicted stimuli
refer to instances where incoming stimuli are predicted
incorrectly; they are generally associated with larger pre-
diction errors because prediction errors involve a combi-
nation of incorrect prediction in addition to sensory input
that is not anticipated. On the other hand, unpredicted
stimuli refer to situations where sensory input is simply
not anticipated. Here, prediction errors tend to be smaller
because it only involves one type of prediction error: sen-
sory input that is not anticipated. In the current study, tim-
ing of auditory stimuli is always predictable whereas the
chance of stimuli being played is 50%, meaning that stim-
uli are unpredicted rather than mispredicted. However,
Roth et al.’s (1976) study involved stimuli that were mis-
predicted rather than unpredicted. This may explain why
prediction errors in the current study and that of Roth
et al. (1976) produced different effects on N1 amplitude.
Furthermore, although lower sound probability (and
hence higher prediction error when a sound does occur)
normally result in N1 amplitudes that are more negative
when compared with sounds with higher probabilities,
the long interstimilus intervals (ISIs) may have given par-
ticipants time to prepare cognitive resources to accommo-
date for whether the sound would play. For example, in a
study by Polich (1990) investigating the P3 component,
participants listened to a stream of 1000 Hz tones with
Han et al.
1495
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
random presentations of a 2000-Hz target tone, which par-
ticipants had to identify via a finger tap. The chance of a
2000-Hz target tone appearing was either 20% or 80%,
depending on the condition. The results, as reported by
Polich (1990), demonstrated enhanced P3 amplitudes
for target sounds of 20% probability compared with target
sounds of 80% probability during low ISI conditions
(about 2–3 sec), but not during high ISI conditions (about
4–10 sec). The study by Polich (1990) can thus be adapted
to investigate the relationship between ISIs and probabil-
ity for the N1 component.
A central design feature of the current paradigm is that
the action (i.e., either a button press or a willed inaction)
was temporally dissociated from the outcome (i.e., the
sound). This design feature allowed us to explore the
electrophysiological index of sense of agency while
minimizing the potential confounds of motor-evoked
activity. Previous studies have explored the impact of
willed inactions on sense of agency using the measure
of temporal binding, which is a phenomenon closely
related to sensory attenuation, and found a temporal
binding effect for inactions, providing further evidence
that willed inactions can result in a sense of agency
( Weller et al., 2020). However, the aforementioned
design features of the current paradigm are—we
suggest—likely also partly responsible for our failure to
identify any modulation of the N1 component in the
experiments. Given this, we suggest that it would be
worthwhile studying the role of contingency in a more tra-
ditional self-stimulation paradigm in which the action is
time-locked to the outcome, and for which we would
expect to observe N1 suppression in the active condition
relative to the passive condition, based on the existing
literature. In such a hypothetical experiment, the active
condition would be the traditional motor–auditory
condition of the self-stimulation paradigm (Hughes et al.,
2013), where participants perform a motor action (such as
a button press) to elicit a sound that is time-locked to the
motor action. The passive condition would be the traditional
auditory condition of the self-stimulation paradigm, where
participants passively listen to an identical sound. It is
worth noting that this study did not include a passive
condition—willed inaction triggering a sound does not
count as a passive condition as normally defined.
Across both experiments, the P2 component was sup-
pressed in the FC condition relative to both NC conditions
(i.e., NC-inaction and NC-action). The functional signifi-
cance of the P2 component is less clear than that of the
N1 component (Crowley & Colrain, 2004). For example,
although the P2 component has shown results like that
of the N1 component in previous studies of sensory atten-
uation (i.e., suppression of the active condition relative to
the passive condition; Horváth & Burgyán, 2013; Knolle
et al., 2012), it has also demonstrated opposite results
(Pinheiro et al., 2019), although only when there was a
delay between the button press and the sound in their
active condition. These inconsistent results may be
attributed to the fact that factors underlying N1 and P2
suppression are likely caused by different factors (van
Elk et al., 2014; Knolle et al., 2012), even though they
have sometimes been seen as part of a single “N1–P2
complex” ( Jack et al., 2021; Timm, SanMiguel, Keil,
Schröger, & Schönwiesner, 2014; Crowley & Colrain, 2004).
In the current set of experiments, we observed P2 sup-
pression, a result in line with some other studies that have
investigated the P2 component in sensory attenuation
(Horváth & Burgyán, 2013; Knolle et al., 2012). Roth
et al. (1976) demonstrated that sounds with a higher prob-
ability elicited N1 components with smaller amplitudes
but P2 components with larger amplitudes. The results
in this study contradict this; sounds that were presented
in the FC condition (and hence had a 100% probability
of occurring after a button press) instead showed smaller
P2 amplitudes compared with lower-probability sounds in
the NC condition. However, as mentioned before, the
types of predictability involved between the current study
and that of Roth et al. (1976) were different. Therefore, we
suggest that more research is needed to elucidate the
nature of the relationship between stimulus probability
and P2 amplitude.
One limitation of the current series of experiments was
that—because of lack of time in the experimental
session—we were not able to include a passive condition
(i.e., a condition where participants passively listen to
auditory stimuli identical to those in the FC and NC condi-
tions), as is common in the traditional self-stimulation par-
adigm. As a result of this, we were unable to determine the
effect of manipulating action–outcome contingency
within the phenomenon of sensory attenuation. In the
future, it might be worthwhile conducting similar experi-
ments with a between-subjects design, with one group
completing an FC condition contrasting with a passive
condition and another group completing the NC condi-
tions contrasting with a passive condition. Furthermore,
participants were not asked to rate their sense of agency
in the current study. Therefore, the interpretation of the
Tb component for the NC-action condition being sugges-
tive of a residual sense of agency could be further investi-
gated in future studies by asking participants to judge the
degree to which they felt a sense of agency in the three
different conditions.
It is also worth noting that although the present studies
investigated action–outcome probability, the experiments
only used probability levels of 100% and 50%. There are
several issues related to this design that are worth noting.
First, this design does not assess probability on multiple
levels (e.g., 100%, 75%, 50%, and 25%), which may help
to address whether there is a direct correlation between
probability levels and auditory suppression or component
amplitude. Second, by setting the uncertain probability
at 50%, it does not allow for priors toward one outcome
to be formed, as the likely occurrence of sounds did not
lean toward tends to appear versus tends not to appear.
Without such a prior, prediction errors are less likely to
1496
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
be formed, meaning that the study was only assessing
unpredictions rather than mispredictions (Hsu et al.,
2015, 2018; Arnal & Giraud, 2012). Studies that have inves-
tigated the effect of probability of auditory processing
have generally used probabilities such as 10% versus 90%
or 20% versus 80% (Pereira et al., 2014; Pincze, Lakatos,
Rajkai, Ulbert, & Karmos, 2002; Polich, 1990). Using the
example of 10% versus 90%, when the probability of a
sound occurring is 10%, then participants generally expect
the sound not to occur; when the probability is 90%, the
general expectation is that the sound will mostly occur.
Future studies should therefore include more contingency
conditions that will examine a broader range of probabili-
ties as well as investigate the relationship between mis-
predictions versus unpredictions in sensory attenuation
(this was not done in the present set of studies because
of time constraints that would occur in a within-subject
design). Different aspects of contingency could also be
manipulated. One could manipulate not only how reliably
a button press results in a sound but also how reliably
sounds are associated with button presses. For example, a
button press might result in a sound 100% of the time, but
inaction could also result in a sound 50% of the time. This
would lead to a 75% contingency rate with strong
expectations of sound occurrence, but with the possibility
of reducing a participant’s sense of agency.
In conclusion, the results of the study suggest that dif-
ferences in action–effect contingency may be indexed by
the P2 and Tb components rather than the N1 component.
The results are consistent with the idea that the Tb compo-
nent may index judgments of agency, with participants in
the FC condition experiencing full agency over the sounds
as opposed to partial agency in the NC conditions. The
results of the N1 analyses provide further support for the
idea that N1 amplitude is not modulated by actions (or
willed inactions) that are not time-locked to sounds.
Reprint requests should be sent to Nathan Han, School of Psy-
chology, University of New South Wales, Sydney, NSW 2052,
Australia, or via e-mail: nathanthomas.han@unsw.edu.au.
Author Contributions
Nathan Han: Conceptualization; Data curation; Formal
analysis; Investigation; Methodology; Software; Visualiza-
tion; Writing—original draft; Writing—Review & editing.
Bradley N. Jack: Conceptualization; Formal analysis; Meth-
odology; Software; Writing—Review & editing. Gethin
Hughes: Writing—Review & editing. Thomas J. Whitford:
Conceptualization; Funding acquisition; Methodology;
Project administration; Resources; Supervision; Writing—
Review & editing.
Funding Information
Thomas J. Whitford: Ideas grant from the National Health
and Medical Research Council (NHMRC) of Australia, grant
number: APP2004067 Discovery Projects from the Austra-
lian Research Council, grant numbers: DP200103288 and
DP170103094: Nathan Han: Australian Government
Research Training Program Scholarship. Bradley Jack:
ARC DECRA, grant number: DE220100739.
Data Availability Statement
All raw EEG data and BrainVision Analyzer history tem-
plates are available on the Open Science Framework at
https://osf.io/p5f8n/. All enquiries about data processing
and analysis procedures can be made to the correspond-
ing author.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. The
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .42; W/M = −.42;
M/ W = .08; W/ W = −.08.
REFERENCES
Aliu, S. O., Houde, J. F., & Nagarajan, S. S. (2008). Motor-
induced suppression of the auditory cortex. Journal of
Cognitive Neuroscience, 21, 791–802. https://doi.org/10.1162
/jocn.2009.21055, PubMed: 18593265
Arnal, L. H., & Giraud, A.-L. (2012). Cortical oscillations and
sensory predictions. Trends in Cognitive Sciences, 16,
390–398. https://doi.org/10.1016/j.tics.2012.05.003, PubMed:
22682813
Baess, P., Widmann, A., Roye, A., Schröger, E., & Jacobsen, T.
(2009). Attenuated human auditory middle latency response
and evoked 40-Hz response to self-initiated sounds.
European Journal of Neuroscience, 29, 1514–1521. https://
doi.org/10.1111/j.1460-9568.2009.06683.x, PubMed:
19323693
Bäß, P., Jacobsen, T., & Schröger, E. (2008). Suppression of the
auditory N1 event-related potential component with
unpredictable self-initiated tones: Evidence for internal
forward models with dynamic stimulation. International
Journal of Psychophysiology, 70, 137–143. https://doi.org/10
.1016/j.ijpsycho.2008.06.005, PubMed: 18627782
Bendixen, A., SanMiguel, I., & Schröger, E. (2012). Early
electrophysiological indicators for predictive processing in
audition: A review. International Journal of
Psychophysiology, 83, 120–131. https://doi.org/10.1016/j
.ijpsycho.2011.08.003, PubMed: 21867734
Han et al.
1497
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Crowley, K. E., & Colrain, I. M. (2004). A review of the evidence
for P2 being an independent component process: Age, sleep
and modality. Clinical Neurophysiology, 115, 732–744.
https://doi.org/10.1016/j.clinph.2003.11.021, PubMed:
15003751
Dixon, M. R. (2000). Manipulating the illusion of control:
Variations in gambling as a function of perceived control over
chance outcomes. The Psychological Record, 50, 705–719.
https://doi.org/10.1007/BF03395379
Elijah, R. B., Le Pelley, M. E., & Whitford, T. J. (2018). Act now,
play later: Temporal expectations regarding the onset of
self-initiated sensations can be modified with behavioral
training. Journal of Cognitive Neuroscience, 30, 1145–1156.
https://doi.org/10.1162/jocn_a_01269, PubMed: 29668396
Elsner, B., & Hommel, B. (2004). Contiguity and contingency in
action–effect learning. Psychological Research, 68, 138–154.
https://doi.org/10.1007/s00426-003-0151-8, PubMed:
14685854
Ford, J. M., Roach, B. J., & Mathalon, D. H. (2010). Assessing
corollary discharge in humans using noninvasive
neurophysiological methods. Nature Protocols, 5,
1160–1168. https://doi.org/10.1038/nprot.2010.67, PubMed:
20539291
Friston, K. (2005). A theory of cortical responses. Philosophical
Transactions of the Royal Society of London, Series B:
Biological Sciences, 360, 815–836. https://doi.org/10.1098
/rstb.2005.1622, PubMed: 15937014
Gallinat, J., Mulert, C., Bajbouj, M., Herrmann, W. M., Schunter,
J., Senkowski, D., et al. (2002). Frontal and temporal
dysfunction of auditory stimulus processing in schizophrenia.
Neuroimage, 17, 110–127. https://doi.org/10.1006/nimg.2002
.1213, PubMed: 12482071
Gentsch, A., & Schütz-Bosbach, S. (2011). I did it: Unconscious
expectation of sensory consequences modulates the
experience of self-agency and its functional signature.
Journal of Cognitive Neuroscience, 23, 3817–3828. https://
doi.org/10.1162/jocn_a_00012, PubMed: 21452945
Gratton, G., Coles, M. G., & Donchin, E. (1983). A new
method for off-line removal of ocular artifact.
Electroencephalography and Clinical Neurophysiology, 55,
468–484. https://doi.org/10.1016/0013-4694(83)90135-9,
PubMed: 6187540
Han, N., Jack, B. N., Hughes, G., Elijah, R. B., & Whitford, T. J.
(2021). Sensory attenuation in the absence of movement:
Differentiating motor action from sense of agency. Cortex,
141, 436–448. https://doi.org/10.1016/j.cortex.2021.04.010,
PubMed: 34146742
Harris, A. J. L., & Osman, M. (2012). The illusion of control: A
Bayesian perspective. Synthese, 189, 29–38. https://doi.org/10
.1007/s11229-012-0090-2
Harrison, A. W., Mannion, D. J., Jack, B. N., Griffiths, O.,
Hughes, G., & Whitford, T. J. (2021). Sensory attenuation is
modulated by the contrasting effects of predictability and
control. Neuroimage, 237, 118103. https://doi.org/10.1016/j
.neuroimage.2021.118103, PubMed: 33957233
Horváth, J. (2015). Action-related auditory ERP attenuation:
Paradigms and hypotheses. Brain Research, 1626, 54–65.
https://doi.org/10.1016/j.brainres.2015.03.038, PubMed:
25843932
Horváth, J., & Burgyán, A. (2013). No evidence for peripheral
mechanism attenuating auditory ERPs to self-induced tones.
Psychophysiology, 50, 563–569. https://doi.org/10.1111/psyp
.12041, PubMed: 23551061
Hsu, Y.-F., Hämäläinen, J. A., & Waszak, F. (2018). The
processing of mispredicted and unpredicted sensory inputs
interact differently with attention. Neuropsychologia, 111,
85–91. https://doi.org/10.1016/j.neuropsychologia.2018.01
.034, PubMed: 29407593
Hsu, Y.-F., Le Bars, S., Hamalainen, J. A., & Waszak, F. (2015).
Distinctive representation of mispredicted and unpredicted
prediction errors in human electroencephalography. Journal
of Neuroscience, 35, 14653–14660. https://doi.org/10.1523
/JNEUROSCI.2204-15.2015, PubMed: 26511253
Hughes, G., Desantis, A., & Waszak, F. (2013). Mechanisms
of intentional binding and sensory attenuation: The role of
temporal prediction, temporal control, identity prediction,
and motor prediction. Psychological Bulletin, 139, 133–151.
https://doi.org/10.1037/a0028566, PubMed: 22612280
Jack, B. N., Chilver, M. R., Vickery, R. M., Birznieks, I.,
Krstanoska-Blazeska, K., Whitford, T. J., et al. (2021).
Movement planning determines sensory suppression: An
event-related potential study. Journal of Cognitive
Neuroscience, 33, 2427–2439. https://doi.org/10.1162/jocn_a
_01747, PubMed: 34424986
Jack, B. N., Le Pelley, M. E., Han, N., Harris, A. W. F., Spencer,
K. M., & Whitford, T. J. (2019). Inner speech is accompanied
by a temporally-precise and content-specific corollary
discharge. Neuroimage, 198, 170–180. https://doi.org/10
.1016/j.neuroimage.2019.04.038, PubMed: 31002966
Klaffehn, A. L., Baess, P., Kunde, W., & Pfister, R. (2019).
Sensory attenuation prevails when controlling for temporal
predictability of self- and externally generated tones.
Neuropsychologia, 132, 107145. https://doi.org/10.1016/j
.neuropsychologia.2019.107145, PubMed: 31319119
Knolle, F., Schröger, E., Baess, P., & Kotz, S. A. (2012). The
cerebellum generates motor-to-auditory predictions: ERP
lesion evidence. Journal of Cognitive Neuroscience, 24,
698–706. https://doi.org/10.1162/jocn_a_00167, PubMed:
22098261
Knolle, F., Schröger, E., & Kotz, S. A. (2013). Prediction errors in
self- and externally-generated deviants. Biological
Psychology, 92, 410–416. https://doi.org/10.1016/j.biopsycho
.2012.11.017, PubMed: 23246535
Luck, S. J., & Gaspelin, N. (2017). How to get statistically
significant effects in any ERP experiment (and why you
shouldn’t). Psychophysiology, 54, 146–157. https://doi.org/10
.1111/psyp.12639, PubMed: 28000253
Martikainen, M. H., Kaneko, K., & Hari, R. (2005). Suppressed
responses to self-triggered sounds in the human auditory
cortex. Cerebral Cortex, 15, 299–302. https://doi.org/10.1093
/cercor/bhh131, PubMed: 15238430
Mifsud, N. G., Oestreich, L. K. L., Jack, B. N., Ford, J. M., Roach, B. J.,
Mathalon, D. H., et al. (2016). Self-initiated actions result in
suppressed auditory but amplified visual evoked components
in healthy participants. Psychophysiology, 53, 723–732. https://
doi.org/10.1111/psyp.12605, PubMed: 26751981
Miller, G. A., Gration, G., & Yee, C. M. (1988). Generalized
implementation of an eye movement correction procedure.
Psychophysiology, 25, 241–243. https://doi.org/10.1111/j.1469
-8986.1988.tb00999.x
Näätänen, R., & Picton, T. (1987). The N1 wave of the human
electric and magnetic response to sound: A review and an
analysis of the component structure. Psychophysiology, 24,
375–425. https://doi.org/10.1111/j.1469-8986.1987.tb00311.x,
PubMed: 3615753
Neszmélyi, B., & Horváth, J. (2017). Consequences matter:
Self-induced tones are used as feedback to optimize
tone-eliciting actions. Psychophysiology, 54, 904–915. https://
doi.org/10.1111/psyp.12845, PubMed: 28240775
Oestreich, L. K. L., Mifsud, N. G., Ford, J. M., Roach, B. J.,
Mathalon, D. H., & Whitford, T. J. (2016). Cortical
suppression to delayed self-initiated auditory stimuli in
schizotypy: Neurophysiological evidence for a continuum
of psychosis. Clinical EEG and Neuroscience, 47, 3–10.
https://doi.org/10.1177/1550059415581708, PubMed:
25898988
1498
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Pereira, D. R., Cardoso, S., Ferreira-Santos, F., Fernandes, C.,
Cunha-Reis, C., Paiva, T. O., et al. (2014). Effects of
inter-stimulus interval (ISI) duration on the N1 and P2
components of the auditory event-related potential.
International Journal of Psychophysiology, 94, 311–318.
https://doi.org/10.1016/j.ijpsycho.2014.09.012, PubMed:
25304172
Pincze, Z., Lakatos, P., Rajkai, C., Ulbert, I., & Karmos, G. (2002).
Effect of deviant probability and interstimulus/interdeviant
interval on the auditory N1 and mismatch negativity in the cat
auditory cortex. Cognitive Brain Research, 13, 249–253.
https://doi.org/10.1016/S0926-6410(01)00105-7, PubMed:
11958968
Pinheiro, A. P., Schwartze, M., Gutierrez, F., & Kotz, S. A.
(2019). When temporal prediction errs: ERP responses to
delayed action-feedback onset. Neuropsychologia, 134,
107200. https://doi.org/10.1016/j.neuropsychologia.2019
.107200, PubMed: 31557484
Pinheiro, A. P., Schwartze, M., Gutiérrez-Domínguez, F., & Kotz,
S. A. (2020). Real and imagined sensory feedback have
comparable effects on action anticipation. Cortex, 130,
290–301. https://doi.org/10.1016/j.cortex.2020.04.030,
PubMed: 32698087
Polich, J. (1990). P300, probability, and interstimulus interval.
Psychophysiology, 27, 396–403. https://doi.org/10.1111/j.1469
-8986.1990.tb02333.x, PubMed: 2236442
Rihs, T. A., Tomescu, M. I., Britz, J., Rochas, V., Custo, A.,
Schneider, M., et al. (2013). Altered auditory processing in
frontal and left temporal cortex in 22q11.2 deletion syndrome:
A group at high genetic risk for schizophrenia. Psychiatry
Research: Neuroimaging, 212, 141–149. https://doi.org/10
.1016/j.pscychresns.2012.09.002, PubMed: 23137800
Roth, W. T., Ford, J. M., Lewis, S. J., & Kopell, B. S. (1976).
Effects of stimulus probability and task-relevance on event-
related potentials. Psychophysiology, 13, 311–317. https://doi
.org/10.1111/j.1469-8986.1976.tb03082.x, PubMed: 951472
SanMiguel, I., Todd, J., & Schröger, E. (2013). Sensory
suppression effects to self-initiated sounds reflect the
attenuation of the unspecific N1 component of the auditory
ERP. Psychophysiology, 50, 334–343. https://doi.org/10.1111
/psyp.12024, PubMed: 23351131
SanMiguel, I., Widmann, A., Bendixen, A., Trujillo-Barreto, N., &
Schroger, E. (2013). Hearing silences: Human auditory
processing relies on preactivation of sound-specific brain
activity patterns. Journal of Neuroscience, 33, 8633–8639.
https://doi.org/10.1523/JNEUROSCI.5821-12.2013, PubMed:
23678108
Saupe, K., Widmann, A., Trujillo-Barreto, N. J., & Schröger, E.
(2013). Sensorial suppression of self-generated sounds and
its dependence on attention. International Journal of
Psychophysiology, 90, 300–310. https://doi.org/10.1016/j
.ijpsycho.2013.09.006, PubMed: 24095710
Schafer, E., Amochaev, A., & Russell, M. J. (1981). Knowledge of
stimulus timing attenuates human evoked cortical potentials.
Electroencephalography and Clinical Neurophysiology,
52, 9–17. https://doi.org/10.1016/0013-4694(81)90183-8,
PubMed: 6166459
Schafer, E. W. P., & Marcus, M. M. (1973). Self-stimulation alters
human sensory brain responses. Science, 181, 175–177.
https://doi.org/10.1126/science.181.4095.175, PubMed:
4711735
Synofzik, M., Vosgerau, G., & Newen, A. (2008). I move,
therefore I am: A new theoretical framework to investigate
agency and ownership. Consciousness and Cognition, 17,
411–424. https://doi.org/10.1016/j.concog.2008.03.008,
PubMed: 18411059
Timm, J., SanMiguel, I., Keil, J., Schröger, E., & Schönwiesner,
M. (2014). Motor intention determines sensory attenuation of
brain responses to self-initiated sounds. Journal of Cognitive
Neuroscience, 26, 1481–1489. https://doi.org/10.1162/jocn_a
_00552, PubMed: 24392902
Timm, J., SanMiguel, I., Saupe, K., & Schröger, E. (2013).
The N1-suppression effect for self-initiated sounds is
independent of attention. BMC Neuroscience, 14, 2.
https://doi.org/10.1186/1471-2202-14-2, PubMed: 23281832
Tonnquist-Uhlen, I., Ponton, C. W., Eggermont, J. J., Kwong, B.,
& Don, M. (2003). Maturation of human central auditory
system activity: The T-complex. Clinical Neurophysiology,
114, 685–701. https://doi.org/10.1016/s1388-2457(03)00005-1,
PubMed: 12686277
van Elk, M., Salomon, R., Kannape, O., & Blanke, O. (2014).
Suppression of the N1 auditory evoked potential for sounds
generated by the upper and lower limbs. Biological
Psychology, 102, 108–117. https://doi.org/10.1016/j.biopsycho
.2014.06.007, PubMed: 25019590
Wascher, E., & Beste, C. (2010). Tuning perceptual
competition. Journal of Neurophysiology, 103, 1057–1065.
https://doi.org/10.1152/jn.00376.2009, PubMed: 20032246
Weller, L., Schwarz, K. A., Kunde, W., & Pfister, R. (2020).
Something from nothing: Agency for deliberate nonactions.
Cognition, 196, 104136. https://doi.org/10.1016/j.cognition
.2019.104136, PubMed: 31760322
Whitford, T. J., Jack, B. N., Pearson, D., Griffiths, O., Luque, D.,
Harris, A. W., et al. (2017). Neurophysiological evidence of
efference copies to inner speech. eLife, 6, e28197. https://doi
.org/10.7554/eLife.28197, PubMed: 29199947
Wolpaw, J. R., & Penry, J. K. (1975). A temporal component of
the auditory evoked response. Electroencephalography and
Clinical Neurophysiology, 39, 609–620. https://doi.org/10
.1016/0013-4694(75)90073-5
Han et al.
1499
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
4
8
8
2
0
3
3
1
9
4
/
/
j
o
c
n
_
a
_
0
1
8
6
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3