The Prospective Sleeping Brain: Age-Related Differences in
Episodic Future Thinking and Frontal Sleep Spindles
Paul Fillmore, Chenlu Gao, Jose Diaz, and Michael K. Scullin
Abstract
■ Sleep spindles are a physiological marker of off-line memory
consolidation. In young adults, sleep spindles are preferentially
responsive to encoded information that is tagged as having fu-
ture relevance. Older adults, on the other hand, show reduced
capacity for future simulation and alterations in sleep physiology.
Healthy young adults (n = 38) and older adults (n = 28) com-
pleted an adaptation night, followed by two in-laboratory poly-
somnography nights, in which they mentally simulated future
events or remembered past events, recorded via written descrip-
tions. We quantified the degree of future/past thinking using
linguistic analysis of time orientation. In young adults, greater
future thinking was linked to greater spindle density, even when
controlling for gender, age, and word count (rp = .370, p =
.028). The opposite was true for older adults, such that greater
future thinking was associated with reduced spindle density
(rp = −.431, p = .031). These patterns were selective to future
thinking (not observed for past thinking). The collective findings
implicate an impaired interaction between future relevance tag-
ging and sleep physiology as a mechanism by which aging com-
promises sleep-dependent cognitive processing. ■
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
2
8
7
1
9
2
1
2
3
9
/
/
j
o
c
n
_
a
_
0
1
7
1
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
The classic view of sleep is that it functions to restore the
body following daytime use. Indeed, sleep restores gly-
cogen levels, clears metabolites, and downregulates syn-
aptic weights. When sleep is lost, it must be recovered,
otherwise daily functioning becomes sluggish, the im-
mune system is more easily compromised, and risk for
diseases/disorders increases (Luyster, Strollo, Zee, &
Walsh, 2012). Slow wave activity (delta power) is typically
used to index sleep homeostasis or, in other words, the
degree to which sleep is needed to restore the brain and
body in response to the previous day (Borbély, 1982).
Though few scholars debate that sleep serves restor-
ative functions, the discovery that memories are reacti-
vated and consolidated during sleep challenges the
notion that sleep’s role is simply to homeostatically regu-
late the brain (Rasch & Born, 2013). Memory functioning,
after all, is a future-oriented ability. It serves little func-
tional value to veridically store all experiences from the
past; instead, memory systems evolved to aid survival in
the present and guide future behaviors (Schacter et al.,
2012).
One model of memory consolidation is that encoded
information must be tagged as future relevant to undergo
consolidation during sleep (Stickgold & Walker, 2013).
Broadly speaking, such tagging may occur based on ex-
pected future rewards, emotionality, and saliency of en-
coded experiences. Selective or preferential consolidation
Baylor University
© 2021 Massachusetts Institute of Technology
has been reported for memories for future actions (Scullin
et al., 2019; Barner, Seibold, Born, & Diekelmann, 2017), for
highly rewarded memories (Michon, Sun, Kim, Ciliberti, &
Kloosterman, 2019; Oudiette, Antony, Creery, & Paller, 2013;
Fischer, & Born, 2009), for “remember” but not “forget”
items in directed forgetting lists (Scullin, Fairley, Decker, &
Bliwise, 2017; Saletin, Goldstein, & Walker, 2011), and when
individuals are explicitly instructed to expect a later retrieval
test (Bennion, Payne, & Kensinger, 2016; Van Dongen,
Thielen, Takashima, Barth, & Fernández, 2012; Wilhelm
et al., 2011; for alternative findings, see Cordi & Rasch,
2021; Reverberi, Kohn, & Fernández, 2020; Wamsley,
Hamilton, Graveline, Manceor, & Parr, 2016). A sleep-based
hippocampal–neocortical dialogue is hypothesized to un-
derlie consolidation of future-relevant memories, with this
dialogue being indexed in humans using scalp EEG mea-
sures of spindle density (Studte, Bridger, & Mecklinger,
2017; Saletin et al., 2011), slow wave activity (Oudiette
et al., 2013), or both ( Wilhelm et al., 2011).
An important but unresolved question is whether and
how aging compromises sleep-dependent memory consol-
idation (Spencer, Gouw, & Ivry, 2007). Memory consolida-
tion is presumed to be a building block of overall cognitive
functioning, with cognitive functioning known to decline
progressively across the adult life span to the point that in-
dependent living becomes disrupted (e.g., early stages of
Alzheimer disease and related dementias). The preponder-
ance of evidence points to memory consolidation being re-
duced or absent even in healthy older adults (Leong, Lo, &
Chee, 2021; Jones, Mackay, Mantua, Schultz, & Spencer,
2018; Gui et al., 2017). However, researchers are still
Journal of Cognitive Neuroscience 33:7, pp. 1287–1294
https://doi.org/10.1162/jocn_a_01716
determining the psychological and neurobiological mecha-
nisms by which memory consolidation declines with aging
(Scullin & Gao, 2018; Mander, Winer, & Walker, 2017). For
example, memory consolidation may decline with aging be-
cause of changes in the neuroanatomy of cognitive systems,
alterations in spindle and slow wave physiology, changes in
encoding quality, or some combination of these cognitive–
sleep–neuroanatomical mechanisms (Muehlroth, Rasch, &
Werkle-Bergner, 2020; see also Huan, Liu, Lei, & Yu, 2020;
Alger, Kensinger, & Payne, 2018). Building on this “combi-
nation” view, we hypothesized that there are changes in
the aging brain’s ability to tag encoded information as fu-
ture relevant in a manner that effectively triggers sleep
physiological responses. We investigated this possibility
by examining sleep physiology markers in response to a
hippocampus-dependent intensive future-thinking task
(Addis, Wong, & Schacter, 2007).
METHODS
Participants
Sixty-six healthy participants spent three nights in the
sleep laboratory. The sample included 38 young adults
(ages 18–29 years, Mage = 20.37 ± 1.67 years, 57.9%
women, 57.9% white) and 28 older adults (ages 50–84 years,
Mage = 65.50 ± 9.07 years, 53.6% women, 82.1% white).
Participants were recruited from the central Texas area
using outreach programs, fliers, and local news advertise-
ments for a larger study of sleep, cognition, and aging
(Scullin et al., 2019). Inclusion criteria were being 18 years
or older and scoring 24 or higher on the Mini-Mental
Status Examination (MYoung = 28.63 ± 1.26, MOlder =
27.96 ± 1.79; Folstein, Folstein, & McHugh, 1975).
Exclusion criteria were having a history of psychiatric or
neurological disorders, insomnia, or narcolepsy; taking
sleep-altering medications; showing an apnea–hypopnea
index of ≥30 on the adaptation night; or not completing
all three laboratory nights. Participants gave informed con-
sent, and the study was reviewed and approved by the
Baylor University institutional review board.
Sleep Measurement
Grass Comet XL Plus systems were used for overnight
polysomnography. Recordings were taken in a sound-
dampened sleep laboratory at Baylor University. The mon-
tage included EEG, recorded at 200 samples per second, at
positions Fp1, Fp2, F3, F4, Fz, C3, C4, Cz, P3, P4, Pz, O1,
and O2, using contralateral mastoids as reference points.
EOG (left and right), mentalis EMG, and measures of
breathing (i.e., nasal pressure, chest and abdomen move-
ments, and finger pulse oximetry) were also included. A
certified polysomnography technician scored sleep stages
in 30-sec epochs according to American Academy of Sleep
Medicine guidelines (Iber, Ancoli-Israel, Chesson, &
Quan, 2007).
Quantitative EEG Analysis
We utilized MATLAB (2019a; Mathworks, Inc.) to conduct
spectral analysis of the EEG. Trained research personnel
excluded epochs containing movement or other artifacts
(averaging 1.82% of all epochs). Following this, data were
re-referenced to the common average, band-pass filtered
utilizing a high-pass cutoff of 0.3 Hz and a low-pass cutoff
of 35 Hz, and down-sampled to the next power of two
(128 Hz), to allow for even spacing of points in subsequent
spectral analysis (Cooley & Tukey, 1965). For each stage of
sleep (N1, N2, N3, REM, and NREM), power was computed
using Welch’s method, using a symmetric 4-sec Hanning
window with 50% overlap, at a resolution of 0.25 Hz,
generating a spectral power density (μV2/Hz) at all scalp
channels. Next, data were corrected for aperiodic noise
components using the fitting oscillations and one-over
f method (Donoghue et al., 2020), which outputs log-
transformed values suitable for parametric analysis.
Finally, mean spectral power density was generated for
typical frequency bands: 0.5–1 Hz (slow oscillations),
1–4 Hz (delta), 4–8 Hz (theta), 8–12 Hz (alpha), 12–16 Hz
(sigma), and 16–32 Hz (beta). Because slow wave activity,
particularly in the frontal lobe, is known to change with
age (Muehlroth & Werkle-Bergner, 2020; Mander et al.,
2017), we were primarily interested in slow wave activity,
operationalized as slow oscillation and delta power aver-
aged across frontal sites (Fp1, Fp2, F3, F4, and Fz) during
NREM.
In a separate analysis pipeline, automated spindle de-
tection was conducted based on the algorithm described
by Wamsley et al. (2012), which performs time–frequency
analysis of the EEG using Morlet wavelets. Average-
referenced data were resampled to 100 Hz, and spindle
detection was conducted separately for each channel dur-
ing artifact-free epochs via identifying amplitude increases
(>4.5 times of mean signal) occurring in the canonical
spindle frequency band (10–16 Hz) and lasting between
0.3 and 3 sec. This procedure has been validated relative
to trained manual scoring and appears to outperform
other automated spindle detection tools ( Warby et al.,
2014). Spindle density was the number of spindles per
minute averaged across frontal sites during NREM.
Procedure
After completing an adaptation night, participants re-
turned to the laboratory for two experimental nights
(89.5% within 2 days, 100% within 1 week). On arrival, par-
ticipants completed questionnaires, had electrodes ap-
plied, and completed 30 min of cognitive and memory
tasks (data reported in Scullin et al., 2019). Afterward, par-
ticipants completed an episodic future-thinking task or a
past-thinking task using the Modified Future Crovitz Test
(MFCT; Crovitz & Schiffman, 1974). In the MFCT, partici-
pants are shown a common noun (e.g., apple) and asked
to generate and type in detail either a related past memory
1288
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
7
1
2
8
7
1
9
2
1
2
3
9
/
/
j
o
c
n
_
a
_
0
1
7
1
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
or simulate a novel future event. Future simulation in the
MFCT is a hippocampus-dependent ability (e.g., Addis
et al., 2007). On one night, participants completed only
future-thinking simulations, and on the other night, partic-
ipants completed only past thinking (night order counter-
balanced). There were 10 trials each night, with different
lists of nouns each night (order counterbalanced). On
each trial, participants viewed the cue word for 3.5 sec
and then typed their response for 60 sec. Participants
were in bed with lights out by 22:30 and with lights on
at 07:30 the next morning, giving approximately 9-hr time
in bed.
Linguistics Analysis
To index the intensity of future/past thinking, we subjected
participants’ typed responses on the MFCT to comput-
erized text analysis using Linguistic Inquiry and Word
Count Version 1.3.1 (LIWC, 2015; Pennebaker, Booth,
Boyd, & Francis, 2015). Using the Java programming
language, LIWC software compares digitized text against
a built-in dictionary to produce weighted estimates of
cognitive and psychological variables. Before analysis,
the data were preprocessed for spelling errors. Although
LIWC extracts dozens of linguistic markers, our focus
was on extractions of time orientation (future/past tense
dominance). All LIWC analyses were adjusted for total
word count.
Statistical Analysis
We first compared the age groups on LIWC time orienta-
tion, spindle density, and slow wave activity outcomes.
Next, we conducted regression analyses to determine
whether greater future thinking was associated with sleep
physiology in young and older adults. For quantitative
EEG analyses, we controlled for sex and chronological
age given the known decline in spindle density and slow
wave activity in male (relative to female) individuals and
with each advancing decade (Fernandez & Lüthi, 2020;
Wilckens, Ferrarelli, Walker, & Buysse, 2018). As a control,
we repeated the analyses with the degree of past thinking
(on past-thinking nights). Primary statistical analyses were
conducted in SPSS Version 27, and all tests were two-tailed
with alpha set to .05. For spatial visualization of our main
effects, we conducted individual-channel follow-up corre-
lation analyses in MATLAB, using a Bonferroni-corrected
alpha of .05/# channels ( p < .0038), with r values
Fisher-transformed to z values for ease of interpretation.
RESULTS
To assess the relationship between brain activity in aging
and future/past orientation, we conducted a series of
2 (Age) × 2 (Night) ANOVAs on the quantitative EEG
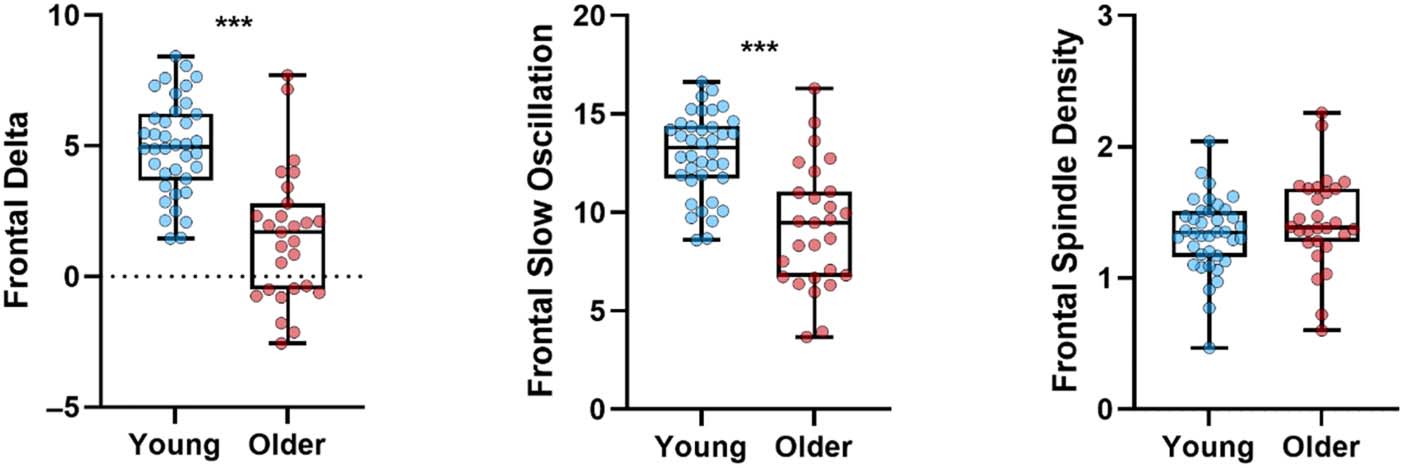
measures. Figure 1 illustrates significant age-related de-
cline in frontal delta power density, F(1, 63) = 39.46,
p < .001, ηp
2 = .385, frontal slow oscillation power density,
F(1, 63) = 28.40, p < .001, ηp
2 = .311, but not in frontal
spindle density, F(1, 62) = 0.14, p = .712, ηp
2 = .002.
There were no interactions with Night after adjusting for
age and sex covariates ( ps >.05). These patterns confirm
previous evidence that the aging brain produces less slow
wave activity (Mander et al., 2017) but diverge somewhat
from findings of fewer spindles in older individuals
(Fernandez & Lüthi, 2020).
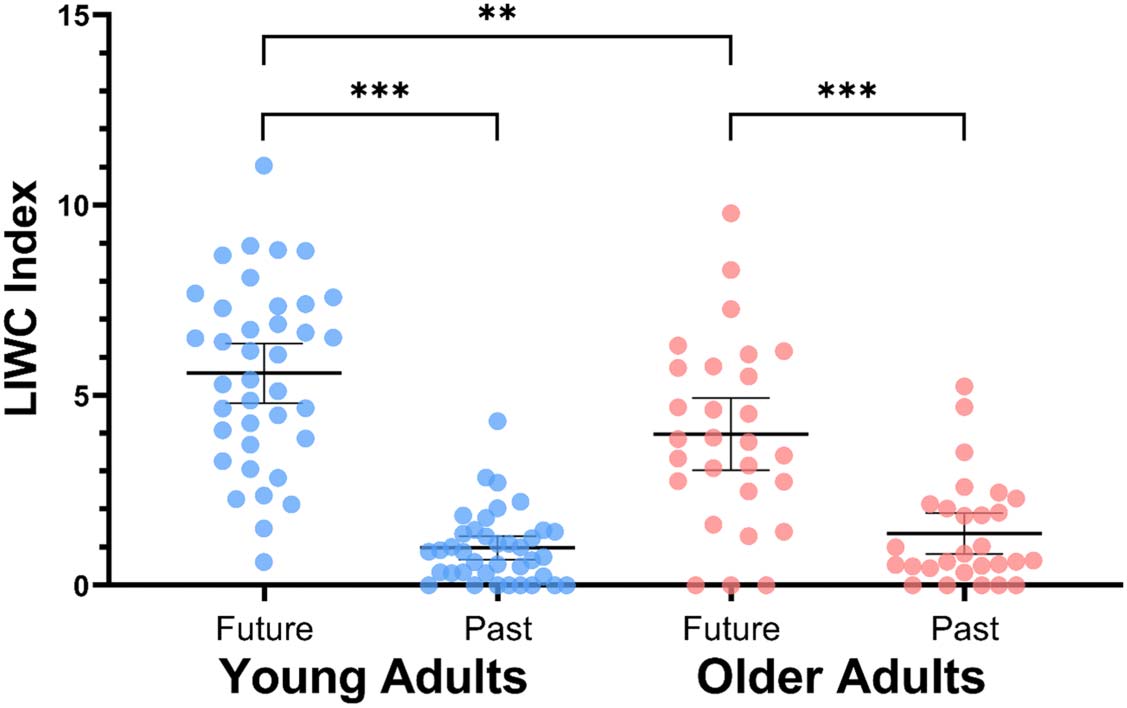
In terms of behavioral differences between age groups,
we also observed an age-related impairment in episodic
future thinking (Figure 2). After adjusting for total word
count, the 2 (Age) × 2 (Tense) ANCOVA showed a sig-
nificant interaction, F(1, 63) = 10.44, p = .002, ηp
2 =
.142. Young adults showed a greater future index than
older adults on the future-thinking night, F(1, 63) =
7.78, p = .007, ηp
2 = .110; this age difference in episodic
future thinking was observed without significant alter-
ations to past focus scores that night, F(1, 63) = 3.08,
p = .084, ηp
2 = .047, or alterations to future index scores
on the past-thinking night, F(1, 63) = 0.616, p = .435,
ηp
2 = .010.
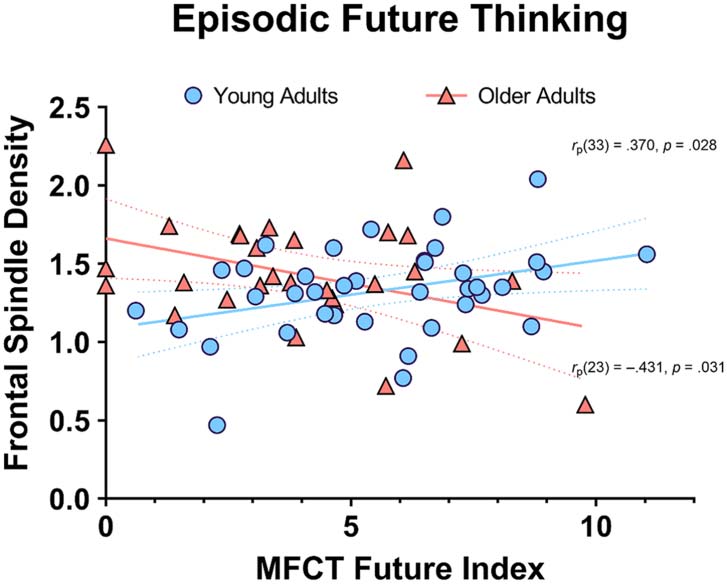
On the future-thinking night, there was an age-related
divergence in the relationship between LIWC future index
scores and subsequent spindle density. Figure 3 illustrates
that, after adjusting for covariates, greater future thinking
was associated with greater frontal spindle density in the
young adults, rp(33) = .370, 95% CI [.042, .626], p = .028;
interestingly, the opposite pattern emerged in the older
adults, rp(23) = −.431, 95% CI [−.706, −.043], p =
.031. Hierarchical regression analysis on frontal spindle
Figure 1. Age-related effects on
NREM slow wave activity and
spindle density, visualized on
the future-thinking night. Older
individuals had less average
power in both slow oscillation
and delta bands but were not
different from younger
individuals in mean spindle
density. Bars represent
minimum and maximum
values. ***p < .001.
Fillmore et al.
1289
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
8
7
1
9
2
1
2
3
9
/
/
j
o
c
n
_
a
_
0
1
7
1
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. LIWC analyses of time
orientation across age groups
during the future-thinking
night. Error bars represent
95% confidence intervals.
**p < .01, ***p < .001.
density provided additional support for this interaction:
After entering Age Group, Sex, Future Index, and Word
Count in Step 1, R2 = .110, F(4, 61) = 1.89, p = .123,
the Age Group × Future Index interaction in Step 2 ex-
plained considerable additional variance in frontal spindle
density, Δ R2 = .118, F(1, 60) = 9.17, p = .004. To visualize
the spatial extent of these effects, we replicated these anal-
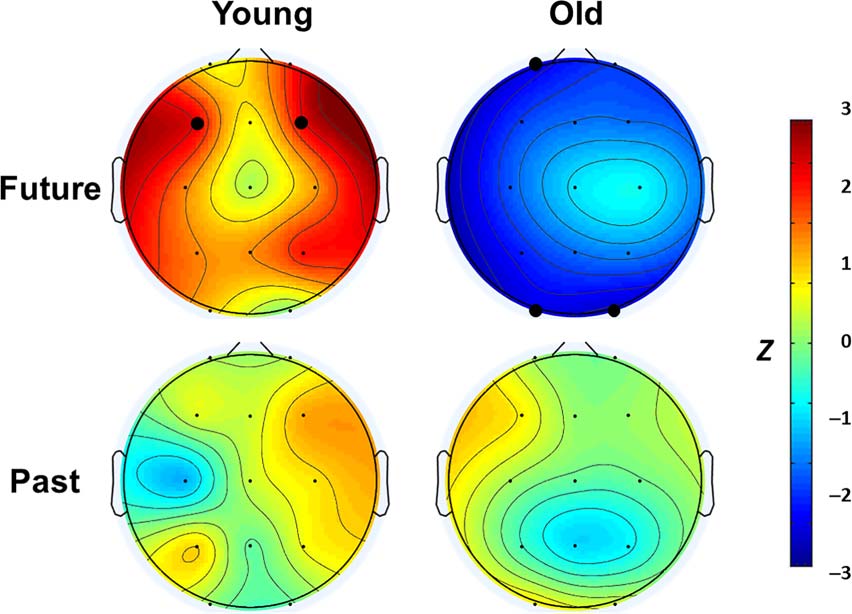
yses at the individual-channel level (Figure 4). We found
significant relationships at F3/ F4 for the young adults
and Fp1 in the older adults, in the same directions as
the main analysis. We also found similar effects at O1/O2
in the older adults. As expected, none of the relationships
between spindle density and past thinking on the past-
thinking night were significant.
In sensitivity analyses that excluded participants with
Mini-Mental Status Examination scores below 27 (n = 7),
this age-based dissociation was maintained, with greater
future thinking being associated with greater frontal spindle
density in young adults, rp(30) = .380, 95% CI [.036, .643],
p = .032, but lower density in older adults, rp(19) =
−.487, 95% CI [−.759, −.070], p = .025. The connection
between future thinking and sleep physiology was selec-
tive to sleep spindles: No significant associations were
observed for slow wave activity, which is often used to
index sleep pressure/homeostasis (e.g., Borbély, 1982).
Furthermore, all correlations were specific to the future
condition night: No significant associations were observed
for the LIWC future index on the past-thinking night or the
LIWC past index in the young or older adults (all ps > .05).
Indeed, even when controlling for spindle activity on the
past-thinking night, there was still a trend toward associa-
tions between greater future thinking and spindle activity
on the future-thinking night: younger, rp(31) = .342, 95%
CI [−.001, .613], p = .051; older, rp(21) = −.385, 95% CI
[−.688, .032], p = .070. Therefore, the link between
future thinking and spindle activity seems night-specific
rather than reflecting generalized trait ability of spindle
density.
Figure 3. Greater future thinking was associated with higher frontal
spindle density in young adults and lower spindle density in older adults
(adjusted for age, sex, and word count). The solid lines represent lines
of best fit, whereas the dashed lines represent their 95% confidence
intervals.
Figure 4. Spatial specificity of future- and past-thinking effects. Individual-
channel analyses highlighted where future-thinking relationships with
spindle density were greatest for young and older adults; bolded dots
indicate corrected p < .05. Past-thinking effects were not significantly
related to spindle density at any channel in either group.
1290
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
8
7
1
9
2
1
2
3
9
/
/
j
o
c
n
_
a
_
0
1
7
1
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
GENERAL DISCUSSION
The current study demonstrated that spindle density is
sensitive to the intensity of recent future simulation.
Increased future simulation yielded higher spindling rates
in young adults, but lower spindling rates in older adults.
These patterns were robust when controlling for sex,
cognitive status, word count, and control night spindle
density. In this section, we consider these findings relative
to the theorized function of sleep spindles and comment
on why aging compromises spindle functioning.
Sleep spindles are theorized to represent off-line
memory consolidation processes (Fernandez & Lüthi,
2020; Zhang, Yetton, Whitehurst, Naji, & Mednick, 2020).
Thalamocortical sleep spindles mediate the coordinated
activity of cortical slow oscillations and hippocampal sharp
wave ripples (Poe, Walsh, & Bjorness, 2010; Latchoumane,
Ngo, Born, & Shin, 2017). In young adults, the coordinated
activity of spindles, slow oscillations, and sharp wave ripples
helps integrate recently encoded information into long-
term memory stores (Staresina et al., 2015). In older adults,
this coordinated activity is disrupted (Muehlroth et al.,
2019; Helfrich, Mander, Jagust, Knight, & Walker, 2018).
Of interest to the current work is evidence that spindling
activity is responsive to recent learning experiences (Gais,
Mölle, Helms, & Born, 2002), particularly intensive learning
of information with future relevance (Studte et al., 2017;
Saletin et al., 2011; Wilhelm et al., 2011). Our findings on
episodic future thinking are consistent with the hypothesis
that spindle activity underlies preferential processing of
future-relevant information. In other words, the sleeping
brain is prospective—at least in young adults—with its pro-
spective nature partially indexed by spindle activity.
In contrast to the findings in young adults, greater epi-
sodic future thinking did not lead to greater spindling in
older adults. If future thinking and spindling were simply
dissociated in older age, then that would fit well with
work that found sleep macro- and microarchitecture to
be less commonly linked to cognitive outcomes in older
adults than in children and young adults (Scullin &
Bliwise, 2015). However, to our surprise, spindling rates
in older adults were negatively associated with intensity
of recent future thinking. Though there is empirical pre-
cedent for this age by spindling dissociation (e.g., Fogel
et al., 2017), it remains poorly understood. We see three
potential interpretations.
First, there are known encoding deficits in older adults,
including deficiencies in simulating the future in detail
(Schacter et al., 2013). By this view, only very detailed
future simulations may pass a threshold by which spin-
dling rates are increased. Second, there are known alter-
ations to sleep physiology with aging, including increased
fragmentation and decreased structural and functional
integrity of the brain regions needed to generate and
coordinate spindle activity (Mander et al., 2017). By this
view, the future-thinking older brain may not be able
to trigger spindle bursting because of fragmented sleep,
cortical cell loss, or hippocampal cell loss (Fogel et al.,
2017). Third, there are changes in time perception with in-
creasing age (socioemotional selectivity theory; Carstensen,
2006). By this view, older adults are more likely to perceive
time as finite and therefore are motivated toward thinking
about the future in terms of regulating positive feelings
and well-being; by contrast, young people perceive time as
open-ended and are therefore more likely to think about
the future in terms of experiencing novelty and learning
new information. Socioemotional selectivity theory, there-
fore, offers a tantalizing explanation of the current findings:
Only specific qualities of future thinking (e.g., learning,
experiencing novelty) are sufficient to drive sleep-dependent
plasticity processes. Of course, some combination of the
above three views may best capture why aging compro-
mises future thinking–sleep physiology interactions.
Muehlroth and Werkle-Bergner (2020) have outlined
an excellent list of key methodological concerns in under-
standing how sleep and aging interact, several of which are
worth mentioning here. First, there are many different
measures of sleep physiology that might seem similar
but show differing effects. For example, we have focused
on spindle density in the current work, but there may be
additional measures of interest (e.g., SO-spindle cou-
pling). Second, different spindle frequencies (fast vs.
slow) can differ in both topography and function and
these frequencies can shift over the lifespan, with fast
spindles becoming faster and slow spindles becoming
slower with increasing age. To minimize the number of
statistical tests, in our analyses, we used a single spindle
detector and did not differentiate between fast and slow
spindles. This approach could potentially cause a de-
creased number of spindles to be detected in older indi-
viduals. However, we saw no significant difference in
frontal spindle density between our young and older
groups, seeming to indicate that these age-related fre-
quency shifts did not lead to a noticeable loss in the num-
ber of detected spindles. Third, the neural generators of
sleep measures, such as spindles, are complex and best
detailed spatially with MRI and electrode setups that al-
low and source localization. Such approaches will be ad-
vantageous to advancing knowledge of sleep, aging, and
future tagging.
In conclusion, though sleep has historically been
viewed as a reactive, homeostatic process, it appears
now that the sleeping brain is more prospective than pre-
viously considered. Restoring glycogen levels, clearing
metabolites, and homeostatically regulating synaptic
weights are biological necessities, but equally important
is preparing the brain for probable future events
(Llewellyn & Hobson, 2015). Young adults’ sleeping brains
do just that. However, with increasing age, there is a dis-
connect between information tagging (as future relevant)
and physiological responses during sleep. Addressing
encoding–spindle interactions should illuminate why some
adults’ cognitive functioning is preserved into older age
whereas other older adults’ functioning declines.
Fillmore et al.
1291
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
8
7
1
9
2
1
2
3
9
/
/
j
o
c
n
_
a
_
0
1
7
1
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Acknowledgments
We are appreciative to Karl Szpunar for his comments on
methodology.
Reprint requests should be sent to Michael K. Scullin, Department
of Psychology and Neuroscience, Baylor University, One Bear
Place 97334, Waco, TX 76798, or via e-mail: Michael_Scullin@
Baylor.edu.
Author Contributions
Paul Fillmore: Formal analysis; Project administration;
Software; Supervision; Visualization; Writing—Review
& editing. Chenlu Gao: Data curation; Formal analysis;
Methodology; Validation; Visualization; Writing—Review
& editing. Jose Diaz: Formal analysis; Methodology;
Validation; Writing—Review & editing. Michael K. Scullin:
Conceptualization; Formal analysis; Funding acquisition;
Project administration; Supervision; Visualization; Writing—
Original draft; Writing—Review & editing.
Funding Information
This research was supported by the Sleep Research
Society Foundation and the National Science Foundation
(https://dx.doi.org/10.13039/100000173), grant numbers:
1920730, 1943323.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed a
persistent pattern of gender imbalance: Although the pro-
portions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/W = .108, and W/W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/W = .102, and W/W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explic-
itly when selecting which articles to cite and gives them
the opportunity to report their article’s gender citation
balance.
REFERENCES
Addis, D. R., Wong, A. T., & Schacter, D. L. (2007). Remembering
the past and imagining the future: common and distinct
neural substrates during event construction and elaboration.
Neuropsychologia, 45, 1363–1377. https://doi.org/10.1016/j
.neuropsychologia.2006.10.016, PubMed: 17126370
Alger, S. E., Kensinger, E. A., & Payne, J. D. (2018). Preferential
consolidation of emotionally salient information during a nap
is preserved in middle age. Neurobiology of Aging, 68, 34–47.
https://doi.org/10.1016/j.neurobiolaging.2018.03.030,
PubMed: 29704647
Barner, C., Seibold, M., Born, J., & Diekelmann, S. (2017).
Consolidation of prospective memory: Effects of sleep on
completed and reinstated intentions. Frontiers in Psychology,
7, 2025. https://doi.org/10.3389/fpsyg.2016.02025, PubMed:
28111558
Bennion, K. A., Payne, J. D., & Kensinger, E. A. (2016). The
impact of napping on memory for future-relevant stimuli:
Prioritization among multiple salience cues. Behavioral
Neuroscience, 130, 281–289. https://doi.org/10.1037
/bne0000142, PubMed: 27214500
Borbély, A. A. (1982). A two process model of sleep regulation.
Human Neurobiology, 1, 195–204. PubMed: 7185792
Boyer, P. (2009). What are memories for? Functions of recall
in cognition and culture. In P. Boyer & J. Wertsch (Eds.),
Memory in mind and culture (pp. 3–28). Cambridge, United
Kingdom: Cambridge University Press. https://doi.org/10
.1017/CBO9780511626999.002
Carstensen, L. L. (2006). The influence of a sense of time on
human development. Science, 312, 1913–1915. https://doi
.org/10.1126/science.1127488, PubMed: 16809530
Cooley, J. W., & Tukey, J. W. (1965). An algorithm for the
machine calculation of complex Fourier series. Mathematics
of Computation, 19, 297–301. https://doi.org/10.1090/S0025
-5718-1965-0178586-1
Cordi, M. J., & Rasch, B. (2021). How robust are sleep-mediated
memory benefits?. Current Opinion in Neurobiology, 67, 1–7.
https://doi.org/10.1016/j.conb.2020.06.002, PubMed: 32711356
Crovitz, H. F., & Schiffman, H. (1974). Frequency of episodic
memories as a function of their age. Bulletin of the
Psychonomic Society, 4, 517–518. https://doi.org/10.3758
/BF03334277
Donoghue, T., Haller, M., Peterson, E. J., Varma, P., Sebastian, P.,
Gao, R., et al. (2020). Parameterizing neural power spectra into
periodic and aperiodic components. Nature Neuroscience,
23, 1655–1665. https://doi.org/10.1038/s41593-020-00744-x,
PubMed: 33230329
Fernandez, L. M., & Lüthi, A. (2020). Sleep spindles: Mechanisms
and functions. Physiological Reviews, 100, 805–868. https://
doi.org/10.1152/physrev.00042.2018, PubMed: 31804897
Fischer, S., & Born, J. (2009). Anticipated reward enhances offline
learning during sleep. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 35, 1586–1593. https://
doi.org/10.1037/a0017256, PubMed: 19857029
Fogel, S., Vien, C., Karni, A., Benali, H., Carrier, J., & Doyon, J.
(2017). Sleep spindles: A physiological marker of age-related
changes in gray matter in brain regions supporting motor
skill memory consolidation. Neurobiology of Aging, 49,
154–164. https://doi.org/10.1016/j.neurobiolaging.2016.10
.009, PubMed: 27815989
Folstein, M. F., Folstein, S. E., & McHugh, P. R. (1975). “Mini-
Mental State”: A practical method for grading the cognitive
state of patients for the clinician. Journal of Psychiatric
Research, 12, 189–198. https://doi.org/10.1016/0022-3956(75)
90026-6, PubMed: 1202204
Gais, S., Mölle, M., Helms, K., & Born, J. (2002). Learning-
dependent increases in sleep spindle density. Journal of
Neuroscience, 22, 6830–6834. https://doi.org/10.1523
/JNEUROSCI.22-15-06830.2002, PubMed: 12151563
Gui, W. J., Li, H. J., Guo, Y. H., Peng, P., Lei, X., & Yu, J.
(2017). Age-related differences in sleep-based memory
consolidation: A meta-analysis. Neuropsychologia, 97,
46–55. https://doi.org/10.1016/j.neuropsychologia.2017.02
.001, PubMed: 28161367
Helfrich, R. F., Mander, B. A., Jagust, W. J., Knight, R. T., &
Walker, M. P. (2018). Old brains come uncoupled in sleep:
Slow wave-spindle synchrony, brain atrophy, and forgetting.
Neuron, 97, 221–230. https://doi.org/10.1016/j.neuron.2017
.11.020, PubMed: 29249289
1292
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
8
7
1
9
2
1
2
3
9
/
/
j
o
c
n
_
a
_
0
1
7
1
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Huan, S. Y., Liu, K. P., Lei, X., & Yu, J. (2020). Age-related
emotional bias in associative memory consolidation: The
role of sleep. Neurobiology of Learning and Memory, 171,
107204. https://doi.org/10.1016/j.nlm.2020.107204, PubMed:
32145405
Iber, C., Ancoli-Israel, S., Chesson, A. L., & Quan, S. (2007). The
AASM manual for the scoring of sleep and associated events:
Rules, terminology and technical specifications. Westchester,
IL: American Academy of Sleep Medicine.
Jones, B. J., Mackay, A., Mantua, J., Schultz, K. S., & Spencer, R. M.
(2018). The role of sleep in emotional memory processing
in middle age. Neurobiology of Learning and Memory, 155,
208–215. https://doi.org/10.1016/j.nlm.2018.08.002, PubMed:
30081153
Latchoumane, C. F. V., Ngo, H. V. V., Born, J., & Shin, H. S.
(2017). Thalamic spindles promote memory formation
during sleep through triple phase-locking of cortical,
thalamic, and hippocampal rhythms. Neuron, 95, 424–435.
https://doi.org/10.1016/j.neuron.2017.06.025, PubMed:
28689981
Leong, R. L., Lo, J. C., & Chee, M. W. (2021). Sleep-dependent
prospective memory consolidation is impaired with aging.
Sleep, zsab069. https://doi.org/10.1093/sleep/zsab069,
PubMed: 33755184
Llewellyn, S., & Hobson, J. A. (2015). Not only … but also: REM
sleep creates and NREM Stage 2 instantiates landmark
junctions in cortical memory networks. Neurobiology of
Learning and Memory, 122, 69–87. https://doi.org/10.1016/j
.nlm.2015.04.005, PubMed: 25921620
Luyster, F. S., Strollo, P. J., Zee, P. C., & Walsh, J. K. (2012).
Sleep: A health imperative. Sleep, 35, 727–734. https://doi.org
/10.5665/sleep.1846, PubMed: 22654183
Mander, B. A., Winer, J. R., & Walker, M. P. (2017). Sleep and
human aging. Neuron, 94, 19–36. https://doi.org/10.1016/j
.neuron.2017.02.004, PubMed: 28384471
Michon, F., Sun, J. J., Kim, C. Y., Ciliberti, D., & Kloosterman, F.
(2019). Post-learning hippocampal replay selectively reinforces
spatial memory for highly rewarded locations. Current Biology,
29, 1436–1444. https://doi.org/10.1016/j.cub.2019.03.048,
PubMed: 31031113
Muehlroth, B. E., Rasch, B., & Werkle-Bergner, M. (2020).
Episodic memory consolidation during sleep in healthy
aging. Sleep Medicine Reviews, 52, 101304. https://doi.org/10
.1016/j.smrv.2020.101304, PubMed: 32278267
Muehlroth, B. E., & Werkle-Bergner, M. (2020). Understanding
the interplay of sleep and aging: Methodological challenges.
Psychophysiology, 57, e13523. https://doi.org/10.1111/psyp
.13523, PubMed: 31930523
Muehlroth, B. E., Sander, M. C., Fandakova, Y., Grandy, T. H.,
Rasch, B., Shing, Y. L., et al. (2019). Precise slow oscillation–
spindle coupling promotes memory consolidation in
younger and older adults. Scientific Reports, 9, 1–15.
https://doi.org/10.1038/s41598-018-36557-z, PubMed:
30760741
Oudiette, D., Antony, J. W., Creery, J. D., & Paller, K. A. (2013).
The role of memory reactivation during wakefulness and
sleep in determining which memories endure. Journal of
Neuroscience, 33, 6672–6678. https://doi.org/10.1523
/JNEUROSCI.5497-12.2013, PubMed: 23575863
Pennebaker, J. W., Booth, R. J., Boyd, R. L., & Francis, M. E.
(2015). Linguistic Inquiry and Word Count: LIWC2015.
Austin, TX: Pennebaker Conglomerates (www.LIWC.net).
Rasch, B., & Born, J. (2013). About sleep’s role in memory.
Physiological Reviews, 93, 681–766. https://doi.org/10.1152
/physrev.00032.2012, PubMed: 23589831
Reverberi, S., Kohn, N., & Fernández, G. (2020). No evidence
for an effect of explicit relevance instruction on consolidation
of associative memories. Neuropsychologia, 143, 107491.
https://doi.org/10.1016/j.neuropsychologia.2020.107491,
PubMed: 32422141
Saletin, J. M., Goldstein, A. N., & Walker, M. P. (2011). The role
of sleep in directed forgetting and remembering of human
memories. Cerebral Cortex, 21, 2534–2541. https://doi.org/10
.1093/cercor/bhr034, PubMed: 21459838
Schacter, D. L., Addis, D. R., Hassabis, D., Martin, V. C., Spreng,
R. N., & Szpunar, K. K. (2012). The future of memory:
remembering, imagining, and the brain. Neuron, 76, 677–694.
https://doi.org/10.1016/j.neuron.2012.11.001, PubMed:
23177955
Schacter, D. L., Gaesser, B., & Addis, D. R. (2013). Remembering
the past and imagining the future in the elderly. Gerontology,
59, 143–151. https://doi.org/10.1159/000342198, PubMed:
22987157
Scullin, M. K., & Bliwise, D. L. (2015). Sleep, cognition, and
normal aging: Integrating a half century of multidisciplinary
research. Perspectives on Psychological Science, 10,
97–137. https://doi.org/10.1177/1745691614556680, PubMed:
25620997
Scullin, M. K., Fairley, J., Decker, M. J., & Bliwise, D. L. (2017).
The effects of an afternoon nap on episodic memory in
young and older adults. Sleep, 40, zsx035. https://doi.org/10
.1093/sleep/zsx035, PubMed: 28329381
Scullin, M. K., & Gao, C. (2018). Dynamic contributions of slow
wave sleep and REM sleep to cognitive longevity. Current
Sleep Medicine Reports, 4, 284–293. https://doi.org
/10.1007/s40675-018-0131-6, PubMed: 31737466
Scullin, M. K., Gao, C., Fillmore, P., Roberts, R. L., Pruett, N., &
Bliwise, D. L. (2019). Rapid eye movement sleep mediates
age-related decline in prospective memory consolidation.
Sleep, 42, zsz055. https://doi.org/10.1093/sleep/zsz055,
PubMed: 30860593
Spencer, R. M., Gouw, A. M., & Ivry, R. B. (2007). Age-related
decline of sleep-dependent consolidation. Learning &
Memory, 14, 480–484. https://doi.org/10.1101/lm.569407,
PubMed: 17622650
Staresina, B. P., Bergmann, T. O., Bonnefond, M., Van Der Meij,
R., Jensen, O., Deuker, L., et al. (2015). Hierarchical nesting
of slow oscillations, spindles and ripples in the human
hippocampus during sleep. Nature Neuroscience, 18,
1679–1686. https://doi.org/10.1038/nn.4119, PubMed:
26389842
Stickgold, R., & Walker, M. P. (2013). Sleep-dependent
memory triage: Evolving generalization through selective
processing. Nature Neuroscience, 16, 139–145. https://doi
.org/10.1038/nn.3303, PubMed: 23354387
Studte, S., Bridger, E., & Mecklinger, A. (2017). Sleep spindles
during a nap correlate with post sleep memory performance
for highly rewarded word-pairs. Brain and Language, 167,
28–35. https://doi.org/10.1016/j.bandl.2016.03.003, PubMed:
27129616
Van Dongen, E. V., Thielen, J. W., Takashima, A., Barth, M., &
Fernández, G. (2012). Sleep supports selective retention
of associative memories based on relevance for future
utilization. PLoS One, 7, e43426. https://doi.org/10.1371
/journal.pone.0043426, PubMed: 22916259
Wamsley, E. J., Hamilton, K., Graveline, Y., Manceor, S., &
Parr, E. (2016). Test expectation enhances memory
consolidation across both sleep and wake. PLoS One, 11,
e0165141. https://doi.org/10.1371/journal.pone.0165141,
PubMed: 27760193
Wamsley, E. J., Tucker, M. A., Shinn, A. K., Ono, K. E., McKinley,
S. K., Ely, A. V., et al. (2012). Reduced sleep spindles and
spindle coherence in schizophrenia: Mechanisms of impaired
memory consolidation? Biological Psychiatry, 71, 154–161.
https://doi.org/10.1016/j.biopsych.2011.08.008, PubMed:
21967958
Fillmore et al.
1293
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
8
7
1
9
2
1
2
3
9
/
/
j
o
c
n
_
a
_
0
1
7
1
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Warby, S. C., Wendt, S. L., Welinder, P., Munk, E. G., Carrillo, O.,
Wilhelm, I., Diekelmann, S., Molzow, I., Ayoub, A., Mölle, M.,
Sorensen, H. B., et al. (2014). Sleep-spindle detection:
Crowdsourcing and evaluating performance of experts, non-
experts and automated methods. Nature Methods, 11, 385–392.
https://doi.org/10.1038/nmeth.2855, PubMed: 24562424
Wilckens, K. A., Ferrarelli, F., Walker, M. P., & Buysse, D. J.
(2018). Slow-wave activity enhancement to improve
cognition. Trends in Neurosciences, 41, 470–482. https://doi
.org/10.1016/j.tins.2018.03.003, PubMed: 29628198
& Born, J. (2011). Sleep selectively enhances memory
expected to be of future relevance. Journal of
Neuroscience, 31, 1563–1569. https://doi.org/10
.1523/JNEUROSCI.3575-10.2011, PubMed: 21289163
Zhang, J., Yetton, B., Whitehurst, L. N., Naji, M., & Mednick, S. C.
(2020). The effect of zolpidem on memory consolidation over
a night of sleep. Sleep, 43, zsaa084. https://doi.org/10.1093
/sleep/zsaa084
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
8
7
1
9
2
1
2
3
9
/
/
j
o
c
n
_
a
_
0
1
7
1
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
1294
Journal of Cognitive Neuroscience
Volume 33, Number 7