The Phenomenology of Error Processing: The Dorsal
ACC Response to Stop-signal Errors Tracks Reports
of Negative Affect
Robert P. Spunt1, Matthew D. Lieberman1, Jessica R. Cohen1,2,
and Naomi I. Eisenberger1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
Abstract
■ A reliable observation in neuroimaging studies of cognitive
control is the response of dorsal ACC (dACC) to events that de-
mand increased cognitive control (e.g., response conflicts and
performance errors). This observation is apparently at odds with
a comparably reliable association of the dACC with the subjec-
tive experience of negative affective states such as pain, fear,
and anxiety. Whereas “affective” associates of the dACC are based
on studies that explicitly manipulate and/or measure the subjec-
tive experience of negative affect, the “cognitive” associates of
dACC are based on studies using tasks designed to manipulate
the demand for cognitive control, such as the Stroop, flanker,
and stop-signal tasks. Critically, extant neuroimaging research
has not systematically considered the extent to which these
cognitive tasks induce negative affective experiences and, if
so, to what extent negative affect can account for any variance
in the dACC response during task performance. While under-
going fMRI, participants in this study performed a stop-signal
task while regularly reporting their experience of performance
on several dimensions. We observed that within-subject var-
iability in the dACC response to stop-signal errors tracked
changes in subjective frustration throughout task performance.
This association remained when controlling for within-subject
variability in subjective reports of cognitive engagement and
several performance-related variables indexing task difficulty.
These results fit with existing models characterizing the dACC
as a hub for monitoring ongoing behavior and motivating adjust-
ments when necessary and further emphasize that such a function
may be linked to the subjective experience of negative affect. ■
INTRODUCTION
By the late 1990s, one of the most reliable observations
in cognitive neuroscience was the response of the dorsal
region of the ACC (dACC; the region of ACC superior and
posterior to the genu of the corpus callosum) to the per-
formance of cognitively engaging tasks, especially to events
that demand increases in cognitive control (Duncan &
Owen, 2000; Paus, Koski, & Caramanos, 1998). Even ear-
lier, electrophysiological investigations had identified
an event-related brain potential robustly associated with
performance errors assumed to be generated by the
dACC (Gehring, Goss, Coles, Meyer, & Donchin, 1993;
Falkenstein, Hohnsbein, Hoormann, & Blanke, 1991).
The reliability of these observations paved the way for a
still predominant account of the dACC as an information-
processing hub that operates to detect and/or signal the
need for adjustments in cognitive control (Ridderinkhof,
Ullsperger, Crone, & Nieuwenhuis, 2004; Yeung, Botvinick,
& Cohen, 2004; Holroyd & Coles, 2002; Botvinick,
Braver, Barch, Carter, & Cohen, 2001), and early on, this
“cognitive” account of dACC was categorically distinguished
1University of California, Los Angeles, 2University of California,
Berkeley
from an “emotional” characterization of the rostral sub-
division of ACC (rACC; the region of ACC anterior and
inferior to the genu of the corpus callosum; Bush, Luu, &
Posner, 2000; Devinsky, Morrell, & Vogt, 1995).
Although the evidence for an information-processing
account of dACC is compelling, there is arguably an
equally compelling body of evidence associating the
dACC with the subjective experience of negative affective
states such as pain, fear, and anxiety (for reviews, see Etkin,
Egner, & Kalisch, 2011; Shackman et al., 2011; Vogt, 2005;
Eisenberger & Lieberman, 2004). As early as the 1950s,
cingulotomies—a surgical procedure that lesions a por-
tion of the dACC but not the rACC—were shown to
be successful in treating medically intractable pain (e.g.,
Foltz & White, 1962) and anxiety disorders (e.g., Whitty,
Duffield, & Tow, 1952). Following cingulotomy for pain,
patients reported that they could still detect the presence
of pain but that it was no longer distressing (Foltz & White,
1962), and this observation accords with more recent
neuroimaging evidence linking the distress or “felt un-
pleasantness” of physical pain to the dACC (Rainville,
Duncan, Price, Carrier, & Bushnell, 1997). In addition,
the dACC has been consistently linked to feelings of dis-
tress following social rejection (e.g., Eisenberger, in press;
Eisenberger, Lieberman, & Williams, 2003) and negative
© 2012 Massachusetts Institute of Technology
Journal of Cognitive Neuroscience 24:8, pp. 1753–1765
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
affect inductions designed to produce fear, anger, or
disgust (Etkin et al., 2011; Shackman et al., 2011; Kober
et al., 2008).
Taken together, the evidence is compelling that the
dACC is linked to the subjective experience of negative
affective states. Yet tasks specifically designed to manip-
ulate cognitive control demands, such as the Stroop,
flanker, and stop-signal, reliably produce activation in
the dACC, especially on trials featuring performance
errors. Critically, it is unknown to what extent these
tasks induce experiences of negative affect, and if so,
whether these experiences explain any part of the dACC
response to the task. In other words, we still have not
answered the question posed by Rainville (2002): “How
does it feel when ACC is activated during a cognitive
task?” (p. 198).
There is indirect evidence to suggest that dACC activa-
tion during cognitive performance, in particular to perfor-
mance errors, may be associated with negative affective
experiences. The majority of this evidence comes from
work examining factors that influence the magnitude of
the error-related negativity (ERN; for a recent review, see
Weinberg, Riesel, & Hajcak, 2012), an event-related brain
potential reliably associated with error commission and
source-localized to the dACC (Debener et al., 2005). Early
on, Luu, Collins, and Tucker (2000) showed that during
performance of a flanker task, the magnitude of the ERN
was associated with individual differences in state and trait
negative affect. Other work has shown that the ERN is
associated with increased autonomic (Hajcak, McDonald,
& Simons, 2003) and startle (Hajcak & Foti, 2008) response
to errors. Moreover, individuals diagnosed with psycho-
pathologies where negative affect is a core feature ex-
hibit an enhanced dACC response to performance errors
(Weinberg, Olvet & Hajcak, 2010; Olvet, & Hajcak, 2008;
Chiu & Deldin, 2007; Fitzgerald et al., 2005; Gehring,
Himle, & Nisenson, 2000). In addition to studies demon-
strating a link between the ERN and negative affect, studies
using fMRI have observed that that the amplitude of the
dACC response during cognitive performance is associ-
ated with variation in autonomic arousal (Critchley, Tang,
Glaser, Butterworth, & Dolan, 2005) and the self-reported
desire to avoid a task (McGuire & Botvinick, 2010). On the
basis of findings such as these, Botvinick (2007) proposed
that the types of events that reliably activate the dACC—
namely, response conflicts and performance errors—may
be intrinsically aversive and that their aversiveness may
be pivotal to motivating adjustments in cognitive control.
The studies just reviewed demonstrate a link between
numerous correlates of negative affective experiences
(e.g., skin conductance, startle response, anxiety disorder)
and the dACC response to cognitive performance. How-
ever, it remains unknown to what extent dACC activation
during cognitive performance is associated with the sub-
jective experience of negative affect in response to such
performance. This study was designed to answer this
question. To do so, we had participants undergo fMRI
while performing a standard version of the stop-signal task
(SST; Logan, 1994), which is commonly used to investigate
cognitive control and which reliably activates the dACC
(e.g., Cohen et al., 2010). In addition, we assessed natural
variability in the subjective experience of performing the
task to test the hypothesis that within-subject variability
in the dACC response to task performance would be
associated with within-subject variability in self-reported
negative affect.
METHODS
Participants
Twenty-one participants were recruited from either the
University of California, Los Angeles (UCLA) subject pool
or the surrounding community, provided informed con-
sent according to the procedures of the UCLA Institutional
Review Board, and received monetary compensation for
participating. Participants were screened to be safe for
the MR environment, not diagnosed with a mental illness,
and not taking psychotropic medications. Six participants
were excluded from the final image analysis because of
technical issues (1; head coil malfunction), abnormal
brain (1; cavernous malformation in the right frontal lobe),
poor SST performance (2; defined a priori as meeting at
least one of the following criteria: response rate on the
Go task of less than 90%; more than 10% incorrect trials
on the Go task; percentage of inhibition on the Stop task
less than 25% or greater than 75%; or a stop-signal RT
[SSRT] of less than 80 msec; Cohen et al., 2010), or a lack
of an emotional response to the task (2; defined a priori
as not meeting the criterion, described further below, for
classifying a participant as demonstrating an emotional
response to the SST). These exclusion procedures left
15 participants (nine women, mean age = 21.87 years,
age range = 19–28 years) for the analyses.
Experimental Design and Procedure
Participants performed four runs of the SST (Logan, 1994),
which features frequent “Go” trials, which require a
speeded response, and less frequent “Stop” trials, which
require the response to be withheld (Figure 1A). The Go
task was a two-choice RT task with spatially compatible
stimulus–response mappings: participants used their right
hand to press a left button with their index finger if an
arrow pointed leftward and a right button with their mid-
dle finger if an arrow pointed rightward. Before each trial, a
warning signal (open circle) appeared on the monitor for
500 msec (visual angle subtended 4.9° × 4.9°). An arrow
then appeared in the center of the circle until either the
participant responded or 1000 msec elapsed.
On 25% of trials, a tone was sounded at a variable delay
(the stop-signal delay [SSD]) after the onset of the arrow
stimulus; this served as a signal for participants to inhibit
1754
Journal of Cognitive Neuroscience
Volume 24, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
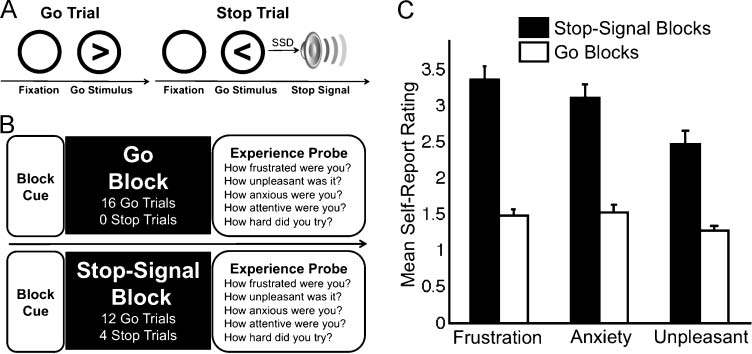
Figure 1. The SST. (A)
Schematic of go and stop trials.
(B) Schematic of Go and Stop-
signal blocks. (C) Mean self-
reports of each dimension of
negative affect for Stop-signal
and Go blocks in the behavioral
pilot study (n = 39).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
their manual response. If participants failed to inhibit,
the trial proceeded as if it were a Go trial. If participants
successfully inhibited, the arrow remained on the screen
for 1000 msec. Error rates were controlled using an adap-
tive tracking procedure (1 up, 1 down staircase) wherein
the SSD was adjusted throughout performance of the
task to produce approximately 50% successful inhibition
for each participant. Two independent staircase functions
with a step size of 50 msec were used, with starting de-
lays on the first run of 250 and 350 msec. For Runs 2–4,
the final SSD of each staircase on the previous run was
used as the starting values.
The first run was administered immediately before
functional scanning during an anatomical scan; the
purpose of this run was to ensure participants under-
stood the task. This run contained 128 trials (96 Go,
32 Stop) performed without interruption. For every
eight trials, there were four left and four right arrows
and two stop trials (one per ladder). Order of arrow
direction and Stop trial ladder were randomized. The
position of the stop-signals was pseudorandomized
with the constraint that the stop-signal could not occur
on three or more contiguous trials. A blank screen was
presented for a fixed interval of 500 msec between
trials (ISI).
The remaining three runs were during fMRI acquisi-
tion (Figure 1B). These runs were identical to the first
run, except for the following changes: (a) each run was
divided into eight blocks of 16 trials, and for two of these
blocks, stop-signals were removed; (b) before each
block, participants were presented with a screen inform-
ing them whether the upcoming block contained stop-
signals; (c) after each block, participants completed an
experience probe (described below); and (d) the ISI was
jittered between 500 and 4000 msec (mean = 1000 msec,
sampled from an exponential distribution).
At each probe, participants reported their experience
of the preceding block on three dimensions of nega-
tive affect: frustration (How frustrated were you?), anxi-
ety (How anxious were you?), and felt unpleasantness
(How unpleasant was it?). In a pilot study of 39 UCLA
undergraduates, we found that, compared with go-only
blocks, stop-signal blocks significantly increased re-
ports of negative affect on these three dimensions (Fig-
ure 1C). In addition to these dimensions of negative
affect, we also assessed two dimensions of cognitive en-
gagement at each probe: attention (How attentive were
you?) and effort (How hard did you try?). Participants
responded using a 7-point Likert scale (1 = not at all,
3 = slightly, 5 = quite a bit, 7 = extremely). To use
the scale, participants used three buttons with their
index, middle, and ring fingers: The first button de-
creased the response value, the second button increased
the response value, and the third button committed
the response value. The starting value was always an-
chored at 1. Participants were given 20 sec to complete
each probe.
The MATLAB (The MathWorks, Inc., Natick, MA) Psycho-
physics Toolbox (Brainard, 1997) version 7.4 was used to
present the stimuli to participants and to record their re-
sponses. Participants viewed the task through LCD goggles
and responded using an MR-compatible button box.
Image Acquisition
Imaging data were acquired using a Siemens Trio 3.0-T MRI
scanner at the UCLA Ahmanson-Lovelace Brainmapping
Center. For each participant, we acquired 828 functional
T2*-weighted EPI volumes (slice thickness = 4 mm,
34 slices, repetition time [TR] = 2000 msec, echo time
[TE] = 30 msec, flip angle = 90°, matrix = 64 × 64, field
of view = 192 mm) divided evenly across three runs.
We also acquired a T2-weighted matched-bandwidth
anatomical scan (same parameters as EPIs, except for
TR = 5000 msec, TE = 34 msec, flip angle = 90°, matrix =
128 × 128) and a T1-weighted magnetization-prepared
rapid acquisition gradient-echo anatomical scan (slice
Spunt et al.
1755
thickness = 1 mm, 176 slices, TR = 2530 msec, TE =
3.31 msec, flip angle = 7°, matrix = 256 × 256, field of
view = 256 mm).
Performance and Experience Analysis
MATLAB was used to analyze performance and experience
data. For SST performance, we computed Go percent
accuracy and RT, standard deviation of Go RT, percentage
of responding on Go trials, percentage of successful inhibi-
tion, and SSRT. Only correct Go trials were included in the
Go RT calculation. SSRT was calculated according to the
race model of stopping (Logan & Cowan, 1984). The race
model assumes that the Go and Stop processes occur in
parallel and are stochastically independent, although when
using the tracking method, this assumption is not critical
(Band, Van der Molen, & Logan, 2003). To calculate SSRT,
we first arranged all correct Go RTs in an assumption-free
distribution in ascending order. Next, we determined the
proportion of failed inhibition. The RT corresponding to
that proportion was computed: the quantile RT (i.e., if
failed inhibition was .55, the RT corresponding to 55% of
the area under the RT distribution curve was the quantile
RT). SSRT was calculated as the difference between the
quantile RT and the average SSD.
For SST experience, we computed mean ratings for the
five experience dimensions for both Stop-signal and Go
blocks (Figure 1B) and used paired-sample t tests to test
differences across the two block types. For Stop-signal
blocks, we computed within-subject correlations among
time courses for the five measured experience dimen-
sions, the number of failed inhibitions per block, and the
average SSD per block.
We used a liberal criterion for classifying participants as
demonstrating an emotional response to the SST. Namely,
participants were considered responders only when their
mean emotion rating was greater than 1.25 during the
Stop-signal blocks (scale range, 1–7). Using this criterion,
15 participants were classified as demonstrating a frus-
tration response (meanresp = 2.42, meannon-resp = 1.19),
13 as demonstrating an anxious response (meanresp =
2.17, meannon-resp = 1.06), and 11 as demonstrating an un-
pleasant response (meanresp = 1.91, meannon-resp = 1.05).
These relatively high exclusion rates on the basis of a lack
of an emotional response were not anticipated based on
the pilot study. Using this same criterion, 97% of the par-
ticipants in the pilot study reported feeling anxiety and
frustration, whereas 82% reported feeling unpleasantness.
We note two explanations for the attenuated emotional
response to the SST in the imaging sample relative to the
pilot sample. The first regards the well-documented fact
that undergoing an MRI is itself emotionally distressing
(Quirk, Letendre, Ciottone, & Lingley, 1989). Given this,
it is plausible that some participants in the imaging sample
may have been at or near ceiling in their baseline levels of
negative affect. The second regards a major procedural dif-
ference across the pilot and imaging samples. As described
above, participants in the imaging sample performed a
practice run of the SST in which self-reports were not col-
lected; participants in the pilot sample did not perform
this practice run. Hence, it is possible that the emotional
response to the task partially habituated during this period.
Image Analysis
Functional data were analyzed using SPM (SPM8, Well-
come Department of Cognitive Neurology, London,
United Kingdom) implemented in MATLAB. Within each
run, image volumes were slice time corrected; realigned
to correct for head motion; normalized into Montreal
Neurological Institute (MNI) space (resampled at 3 ×
3 × 3 mm) using the default SPM segmentation routine;
smoothed with an 8-mm Gaussian kernel, FWHM; de-
trended using the linear model of the global signal
method (Macey, Macey, Kumar, & Harper, 2004); and
standardized so that the mean of each voxelʼs time series
was 100. Following this preprocessing, image volumes
from all runs were concatenated to produce a single
consecutive time series. This procedure of detrending
and standardizing the time course of each run separately
before their concatenation allows us to capitalize on
between-run variability in the emotional response in
the statistical analysis and follows procedures used in pre-
vious work (e.g., Zaki, Weber, Bolger, & Ochsner, 2009).
For each subject, we began by estimating three models
(henceforth referred to as the “Base Models”), one for each
of the three dimensions of negative affect (frustration,
anxiety, felt unpleasantness). In each of these models, we
separately modeled the three events that occur most
frequently during performance of the SST: accurate Go
responses (Go-success), successful inhibitions for Stop
trials (Stop-success), and failed inhibitions for Stop trials
(Stop-failures). The onset of each event was defined as
the onset of the Go (arrow) stimulus. Regressors were
created by convolving a delta function at each event onset
with a canonical (double-gamma) hemodynamic response
function. In addition to the regressors modeling the aver-
age (time-invariant) response, each model also included
one of the three dimensions of negative affect as a block-
wise parametric modulator of the response to the three
events of interest occurring with each block. These param-
etric regressors were created by modulating the amplitude
of the delta functions at each event by the demeaned
values of the parameter (e.g., self-reported frustration)
for each trial. Within these models, our primary goals were
to estimate both the parametric effect of negative affect on
each of the three event types, as well as the difference
in parametric effects across the event types. These param-
etric contrasts allowed us to determine regions where
the relationship between negative affect and brain activity
differs among the three event types.
To further specify the effect of frustration on the dACC
response to Stop-failure trials, we estimated one additional
model (referred to as the “Control Model”). This model
1756
Journal of Cognitive Neuroscience
Volume 24, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
was identical to the Base Model for frustration, except it
included four additional parametric modulators of the re-
sponse to Stop-failures. These were (1) trial-wise variability
in the latency of the erroneous manual response, (2) trial-
wise variability in the SSD, (3) block-wise variability in the
total number of errors occurring within each block, and
(4) block-wise variability in subjective cognitive engagement.
The latter parameter was computed by averaging the time
courses of self-reported attentiveness and effort. The latter
two measures were averaged into a composite because
their time courses were highly intercorrelated (r = .55). As
in the Base Models, all parameter regressors were created
by modulating the amplitude of the delta functions at each
event by the parameterʼs demeaned values.
All models included two additional regressors coding
for the position of the three runs in the concatenated time
series, and the time series at each voxel was high-pass
filtered to 1/128 Hz. Serial autocorrelations were modeled
as an AR(1) process. For each model, linear contrasts were
performed for comparisons of interest, and contrast im-
ages were then subjected to one-sample t tests for group
analysis. Our primary analyses were conducted within a
dACC ROI (Figure 2A). We defined this ROI to capture
the putatively cognitive, supracallosal region of the ante-
rior midcingulate (Bush et al., 2000). To produce this
ROI, we took the following steps: (a) we created a binary
ACC mask using the automated anatomical labeling atlas
(Tzourio-Mazoyer et al., 2002), (b) trimmed the mask in
the posterior-anterior plane (0 ≤ y ≤ 36), and (c) overlaid
the mask on the average of the participantsʼ normalized
anatomical images and edited it to minimize white matter
coverage and maximize gray matter coverage.
Analyses within the dACC ROI were performed using a
cluster-level family-wise error (FWE) rate of .05, with
clusters defined using a voxel-wise threshold of p < .005
(uncorrected). We followed all ROI analyses with whole-
brain analyses using a cluster-level FWE rate of .05 and a
cluster-defining threshold of .001 (uncorrected). For visual
presentation, t-statistic maps were overlaid on the average
of the participantsʼ normalized anatomical images.
RESULTS
Performance on and Experience of the SST
Table 1 summarizes all performance and experience data.
Performance variables were consistent with past research
using the SST (Cohen et al., 2010; Congdon et al., 2010).
Importantly, we observed that during Stop-signal blocks,
compared with Go blocks, individuals reported experienc-
ing more frustration, t14 = 5.87, p < .001 (Mdiff = 1.07,
SDdiff = 0.70); anxiety, t14 = 2.81, p = .01 (Mdiff = .48,
SDdiff = .67); and felt unpleasantness, t14 = 4.86, p <
.001 (Mdiff = 0.49, SDdiff = .39). This replicates results from
the behavioral pilot study (Figure 1C) and confirms that
participants subjectively experienced negative emotions
in response to performing the SST during the scanning
session. Participants also reported being marginally more
attentive, t14 = 1.95, p = .07 (Mdiff = 0.31, SDdiff = .62),
and exerting marginally more effort, t14 = 2.08, p = .06
(Mdiff = 0.29, SDdiff = .54), during Stop-signal compared
with Go blocks.
Table 2 summarizes all within-subject correlations among
the five measured dimensions of experience and relevant
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
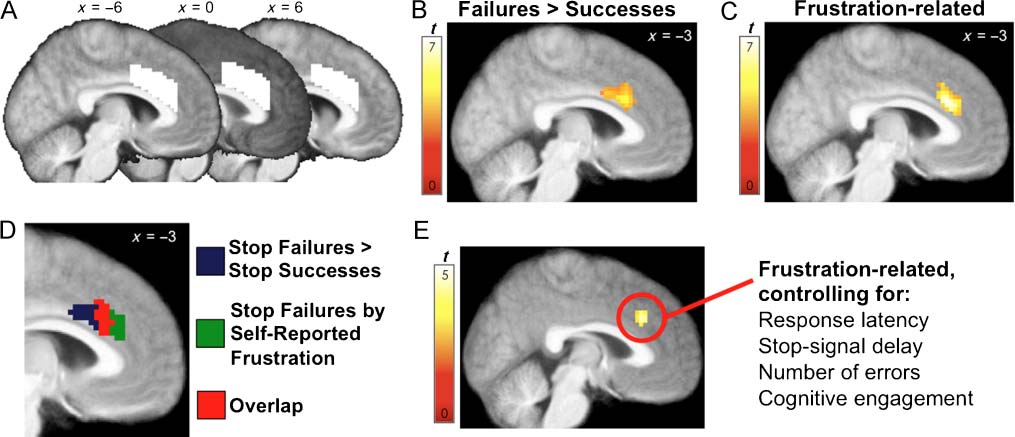
Figure 2. Results observed within dACC ROI analyses. All results are cluster-level corrected using a FWE rate of .05. (A) The dACC mask used
in ROI analyses. (B) The region of dACC emerging in the contrast Stop-failures > Stop-successes. (C) The region of dACC where within-subject
variability in the magnitude of the response to Stop-failures was associated with within-subject variability in self-reported frustration. (D) Overlap
of the regions of dACC observed in B and C. (E) The region of dACC whose response to Stop-failures was associated with self-reported frustration
in the Control Model.
Spunt et al.
1757
Table 1. Descriptive Statistics of SST Performance and
Experience for All Participants Included in the Analysis
(n = 15)
task performance. The only association observed among
experience and performance was between frustration and
block-by-block variation in the number of Stop-failures,
rmean = .33, t14 = 3.73, p = .002.
Mean
SD
Min
Max
99.70
0.57
97.92 100.00
581.60 109.16 370.40 746.18
Neural Correlates of the Experience of the SST
Variable
Go accuracy
Median Go RT
SD Go RT
133.17
28.65
91.20 175.11
Percentage of Go responses
98.01
2.30
91.67 100.00
Percentage of inhibition
53.10
6.42
41.67
64.58
SSRT
Frustration
Stop-signal
Go
Anxiety
Stop-signal
Go
Unpleasant
Stop-signal
Go
Attention
Stop-signal
Go
Effort
Stop-signal
Go
168.87
27.20 128.00 225.30
2.42
1.36
2.02
1.53
1.69
1.20
5.51
5.19
5.59
5.30
0.71
0.47
0.85
0.53
0.57
0.39
0.76
1.01
0.57
0.89
1.50
1.00
1.00
1.00
1.00
1.00
4.50
3.33
4.56
3.33
3.89
2.83
4.00
2.67
3.00
2.50
6.83
6.50
6.46
6.75
performance variables that demonstrated intrasubject vari-
ation. Correlations among the dimensions of experience
were moderate to low, with attentiveness and effort show-
ing the strongest relationship. This indicates that these
dimensions tap unique aspects of the phenomenology of
As described in the Methods, we conducted all primary
analyses within a dACC ROI (Figure 2A). All whole-brain
results all listed in Table 4. Our analysis began with con-
firming the presence of a dACC response to Stop-failures
in the contrast Stop-failures > Stop-successes. This re-
vealed a cluster (Figure 2B; listed in Table 3) in a region
of the dACC comparable with error-related activity iden-
tified in meta-analyses (e.g., Ridderinkhof et al., 2004).
Next, we used parametric modulation analyses to test
if within-subject variability in the subjective experience
of any of the three dimensions of negative affect were
associated with the amplitude of the dACC response to
any of three basic events that occur during the task: Go-
successes, Stop-successes, and Stop-failures. Reports of
frustration did not significantly explain any variability in
the dACC response to either Go- or Stop-successes. How-
ever, frustration did significantly explain variability in the
response to Stop-failures, both in the ROI and whole-brain
analyses (Figure 2C; listed in Tables 3 and 4). As shown in
Figure 2D, this cluster demonstrates considerable overlap
with the region of dACC observed in the contrast Stop-
failure > Stop-success, although it is notable that there
is some clear differentiation, with the average error effect
located more caudally than the parametric effect of frus-
tration. To determine how specific this effect was to error
trials, we directly contrasted the parametric effect of frus-
tration on Stop-failures to the same effect estimated for
both Go-successes and Stop-successes. For both contrasts,
we observed a cluster in dACC overlapping with the clus-
ter depicted in Figure 1C (listed in Table 3). This dem-
onstrates that the link between the dACC response and
self-reported frustration is selectively driven by the response
to error trials.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
f
t
Table 2. Average Within-subject Time Course Correlations among the Dimensions of Experience, Number of Errors per Block, and
Average SSD per Block, for All Participants Included in the Analysis (n = 15)
Frustration
Anxiety
Unpleasant
Attention
Effort
Frustration
Anxiety
Unpleasant
Attention
Effort
Errors
1
–
–
–
–
–
0.25*
1
–
–
–
–
0.08
0.33*
1
–
–
–
0.09
0.14
0.22*
1
–
–
0.06
0.15
0.10
0.55*
1
–
Errors
0.33*
0.01
0.13
0.02
−0.08
1
SSD
0.07
0.02
−0.09
−0.15
−0.13
0.15*
*Significantly different from zero at p < .05. 1758 Journal of Cognitive Neuroscience Volume 24, Number 8 u s e r o n 1 7 M a y 2 0 2 1 Next, we examined whether variability in felt unpleasant- ness and/or anxiety explained variance in the dACC re- sponse to the SST. For both affective dimensions and for the three event types, no clusters were observed that survived correction in either the whole-brain or dACC ROI analyses. Given the relatively low number of partici- pants showing evidence of experiencing unpleasantness (n = 11) and anxiety (n = 13) in response to the task, it is notable that two clusters do emerge when interrogating the dACC ROI using the cluster-defining threshold of p < .005. Namely, we observed associations between reports of felt unpleasantness and the dACC response to Stop-failures (peak: x = 0, y = 24, z = 33; t10 = 5.77, voxel extent = 11) and between reports of anxiety and the dACC response to Go-successful events (during stop-signal blocks; peak: x = 3, y = 18, z = 24; t12 = 3.85, voxel extent = 15). Although preliminary, these results suggest that dimensions of nega- tive affect other than frustration may explain variance in the dACC response during performance of the SST. However, given that they did not survive correction for multiple com- parisons, we focused the remainder of our analyses on the dimension of frustration. Our final set of analyses tested the specificity of the ob- served association among self-reported frustration and the dACC response to errors. To do so, we estimated an additional model that was identical to the model in which the relationship between frustration and the re- sponse to Stop-failures was observed, except it included parametric regressors modeling the effect of four addi- tional variables on the response to Stop-failures: (1) trial- wise variability in the latency of the erroneous manual response, (2) trial-wise variability in the SSD, (3) block-wise variability in the total number of errors occurring within each block, and (4) block-wise variability in subjective cog- nitive engagement (average of self-reported attentiveness and effort). Within this model, we continued to observe a robust relationship between the dACC response to Stop- failures and subjective frustration (Figure 2E; listed in Table 3). When examining the four additional parametric effects, only one—response latency—explained signifi- cant variance in the response of the dACC to Stop-failures. This effect replicates previous work demonstrating that the dACC is sensitive to response latency (Grinband et al., 2011; McGuire & Botvinick, 2010). DISCUSSION Although the earliest work on dACC function, involving cingulotomies for pain, anxiety, and depression, all Table 3. Significant Clusters Identified in the Base and Control Models when Searching within the dACC ROI Tested Effect Contrast of Main Effects in the Base Model Stop-failures > Stop-successes
Parametric Effects in the Base Model
Frustration: Stop-failures
Frustration: Stop-successes
Frustration: Go-successes
Contrasts of Parametric Effects in the Base Model
Frustration: Stop-failures > Stop-successes
Frustration: Stop-failures > Go-successes
x
−12
−6
−6
0
Parametric Effects ( for Stop-failures Only) in the Control Model
Frustration
Response Latency
SSD
Number of errors
Subjective engagement
−6
−9
y
24
27
21
21
27
24
t
7.00
6.28
4.07
4.43
4.41
5.19
z
30
27
No significant clusters
No significant clusters
24
27
30
30
No significant clusters
No significant clusters
No significant clusters
k
104
125
50
93
24
119
All clusters were identified using a voxel-wise threshold of p < .005 (uncorrected) and are significant at a cluster-level FWE rate of .05. n = 15. x, y, and z refer to MNI coordinates of local maxima in the left–right, anterior–posterior, and inferior–superior dimensions, respectively. t refers to the t statistic at those coordinates. k refers to the cluster size (in voxels). Spunt et al. 1759 D o w n l o a d e d f r o m l l / / / / j t t f / i t . : / / h t t p : / D / o m w i n t o p a r d c e . d s f i r o l m v e h r c p h a d i i r r e . c c t . o m m / j e d o u c n o / c a n r a t r i t i c c l e e - p - d p d 2 f 4 / 8 2 4 1 / 7 8 5 / 3 1 1 7 9 5 4 3 4 / 1 1 0 7 1 7 o 8 c 1 n 7 _ 1 a / _ j 0 o 0 c 2 n 4 2 _ a p _ d 0 0 b 2 y 4 g 2 u . e p s t d o f n b 0 y 8 S M e I p T e m L i b b e r r a 2 r 0 2 i 3 e s / j t f / . u s e r o n 1 7 M a y 2 0 2 1 Table 4. Significant Clusters Identified in the Base and Control Models when Searching within the Whole Brain y 24 48 51 −57 15 −24 −90 −72 12 27 −36 −15 x −12 −12 −15 15 −3 6 −33 42 27 −6 −30 3 Anatomical Region Stop-failures > Stop-successes (Base Model)
dACC
Pregenual cingulate cortex
Dorsomedial pFC
Cerebellum
Pre-SMA
Midbrain
Stop-successes > Stop-failures (Base Model)
Middle occipital gyrus
Middle occipital gyrus
Putamen
L
L
L
R
L/R
L/R
L
R
R
Parametric Effect of Frustration on Stop-failures (Base Model)
Positive effect
dACC
Negative effect
Postcentral gyrus
SMA
L
L
L/R
Parametric Effect of Frustration on Stop-successes (Base Model)
No significant clusters
Parametric Effect of Frustration on Go-successes (Base Model)
No significant clusters
Contrasts of Parametric Effects of Frustration (Base Model)
−6
−30
30
−33
Stop-failures > Go-successes
dACC
Go-successes > Stop-failures
Postcentral gyrus
Stop-failures > Stop-successes
No significant clusters
Stop-successes > Stop-failures
Postcentral gyrus
Go-successes > Stop-successes
No significant clusters
Stop-successes > Go-successes
No significant clusters
L
L
L
z
30
6
27
−18
57
−12
15
−12
−6
27
57
57
24
57
t
7.00
5.93
4.77
5.43
5.41
5.09
7.42
7.46
6.27
6.28
9.31
5.32
k
43
76
–
92
59
45
346
513
45
67
210
58
4.50
5.69
52
124
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
−33
−30
54
5.16
86
1760
Journal of Cognitive Neuroscience
Volume 24, Number 8
Table 4. (continued )
Anatomical Region
x
y
z
t
k
Parametric Effects on Stop-failures Estimated in the Control Model
L
R
L
R
L
L
−33
−36
57
6.17
52
33
−36
12
−45
39
24
30
9
−9
45
6
−3
66
−9
9
6.17
5.56
5.96
5.14
5.13
170
97
33
38
36
Frustration
Positive effect
No significant clusters
Negative effect
Postcentral gyrus
Response latency
Positive effect
Anterior insula/inferior frontal gyrus
SMA
Superior temporal gyrus
Middle frontal gyrus
Negative effect
No significant clusters
SSD
No significant clusters
Number of errors
No significant clusters
Subjective engagement
No significant clusters
All clusters were identified using a voxel-wise threshold of p < .001 (uncorrected) and are significant at a cluster-level FWE rate of .05.
n = 15. L and R refer to left and right hemispheres. x, y, and z refer to MNI coordinates in the left–right, anterior–posterior, and inferior–superior
dimensions, respectively. t refers to the t score at those coordinates (local maxima). k refers to the cluster size (in voxels).
pointed to its role in the experience of negative affect,
contemporary theoretical accounts of the dACC tend to
focus exclusively on its role in cognitive control. The reli-
able association of the dACC with the exertion of cog-
nitive control is primarily based on the observation of
dACC activity during performance of tasks specifically
designed to manipulate cognitive control demands.
We have shown that one such commonly used task, the
SST, although not designed to investigate negative af-
fect, nevertheless induces experiences of negative affect
which vary throughout task performance. Importantly,
this natural variability in the experience of negative affect—
namely, self-reports of frustration—explained a signifi-
cant amount of variability in the dACC response during
task performance. This strongly suggests that the asso-
ciation of dACC with the experience of negative affect
cannot be easily reduced to a mere “cognitive” account of
its function. Even in its response to a canonical “cognitive”
event—a simple performance error on a speeded response
inhibition task—the dACC appears to support an affective
function.
Relationship to Previous Studies
Although the cognitive account of dACC has been domi-
nant for more than 15 years, several reviews and meta-
analyses have highlighted the dACCʼs association with
both cognitive control and the experience of negative
affect (Weinberg et al., 2012; Etkin et al., 2011; Shackman
et al., 2011; Botvinick, 2007; Eisenberger & Lieberman,
2004). For instance, Shackman and colleagues (2011)
marshaled data from 192 neuroimaging studies of negative
affect, physical pain, and cognitive control and found that
all three categories reliably report activation in an over-
lapping region of the dACC. Although strongly suggestive,
the interpretation of meta-analytically defined overlap is
not unambiguous, as it is well accepted that the same
region can perform different functions in different contexts
Spunt et al.
1761
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
(Yarkoni, Poldrack, Nichols, Van Essen, & Wager, 2011;
Poldrack, 2006). For instance, from these meta-analyses,
it would be invalid to infer that during the performance
of a task designed to investigate variation in cognitive de-
mand, the presence of dACC activity indicates the presence
of a subjective experience of negative affect. Here, we pro-
vide the first demonstration of an overlap in the regions
associated with the objectively defined (error > nonerror)
and subjectively defined (self-reported frustration) aspects
of cognitive performance. Moreover, we have shown that
the association of self-reported frustration with the dACC
response to errors is independent of self-reported cogni-
tive engagement and several performance-related vari-
ables. To recapitulate Rainvilleʼs (2002) query, “How does
it feel when ACC is activated during a cognitive task?”
(p. 198). The results of this study provide the best evidence
yet that, at least in the response to performance errors,
ACC activation—in particular, the dorsal region typically
associated with signaling the need for adjustments in
cognitive control—feels like frustration.
Integrative Perspectives on dACC Function
These two correlates of dACC activity—the presence/absence
of error and the magnitude of subjective frustration—
invite consideration of integrative models of dACC function
which can accommodate its association with both cog-
nitive control and negative affect. Indeed, the dACC is
well positioned to support highly integrative functions,
which cut across traditional boundaries between cogni-
tion and emotion, as it is densely interconnected with
brain systems involved in affect, executive control, and
action (Shackman et al., 2011; Pessoa, 2008; Paus, 2001).
Eisenberger and Lieberman (2004) make analogy to an
alarm in describing the multiple correlates of dACC activity.
Any functional alarm system (e.g., smoke alarm) requires
two operations: (1) detecting discrepancies from a desired
set point (e.g., too many particles of smoke) and (2) gen-
erating a signal (e.g., alarm bell) capable of motivating
corrective action. The performance-defined contrast of
error and nonerror events may isolate the first operation
(i.e., detection), whereas the magnitude of self-reported
frustration in response to error events may isolate an
aspect of the second operation (i.e., signaling). The idea
that the dACC is involved in both the detection of prob-
lems in ongoing behavior and the motivation of adjust-
ments in future behavior is implied by other integrative
models of dACC function (Weinberg et al., 2012; Shackman
et al., 2011; Botvinick, 2007). For instance, Botvinick
(2007) proposed that the dACC may underlie a form of
avoidance learning in which cognitive demand (e.g., re-
sponse conflicts and errors) is registered as aversive to
motivate adjustments in future behavior. In support of this
proposed link between the aversiveness of cognitive de-
mand and the dACC, McGuire and Botvinick (2010; see
also Botvinick, Huffstetler, & McGuire, 2009) observed that
variability in self-reported desire to avoid a cognitively
demanding task tracked the dACC response to the task,
although in contrast to the present study this association
was not robust when controlling for block-wise variability
in response latency and error commission. In support of
the proposed link between the aversiveness of cognitive de-
mand and the adjustment of ensuing behavior, several be-
havioral studies have demonstrated that negative affect
during cognitive performance facilitates the deployment of
cognitive control (van Steenbergen, Booij, Band, Hommel,
& Does, 2012; van Steenbergen, Band, & Hommel, 2009,
2010) and informs future decisions regarding the avoidance
of cognitively demanding tasks (Kool, McGuire, Rosen, &
Botvinick, 2010). Hence, negative affective experiences
may not simply be an incidental byproduct of performing
cognitively demanding tasks; rather, they may be central
to the way in which individuals use information about
cognitive difficulties to adjust their future behavior.
Limitations
There are several limitations that merit discussion. The
first is that our results do not directly bear on computa-
tional perspectives on dACC function. This is because
functional (computational) and phenomenological (self-
reported phenomenal experience) perspectives on men-
tal processes sit squarely at different levels of analysis.
However, it is noteworthy that the relationship between
self-reported frustration and the dACC response to the
task was specific to error events. This provides neural evi-
dence for the face valid association of error commission
with the experience of frustration. More importantly, these
findings demonstrate that, in the response of the dACC to
errors, not all errors are created equal and that, whatever
attribute of an error the dACC computes, this attribute is
likely intimately linked with feelings of frustration. This
study suggests that research on the computational basis
of affective states, such as frustration, will likely be critical
for fully understanding the nature of the interface between
cognition and affect in the dACC.
The second limitation regards the nature of our mea-
surement of negative affect. Namely, although the resolu-
tion of our experience probes was at the level of individual
blocks (1 every 16 trials), subjective experience undoubt-
edly varied substantially at the level of individual trials.
Choosing to measure experience at the end of blocks of
trials was intended to provide a good balance between
the temporal resolution of our measurement of experience
and the amount of disruption this measurement would
have on task performance. Despite the block-level resolu-
tion of self-reports, we observed strong evidence for a trial-
specific association between dACC and frustration: This
association was observed for error trials only, and the
strength of this association was significantly stronger than
that observed for the other trial types occurring within
each block. This suggests that future research on the
neural bases of meta-cognitive experiences, such as frus-
tration, might benefit by directing participants to report
1762
Journal of Cognitive Neuroscience
Volume 24, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
experience only of error trials. This would enable an in-
crease in temporal resolution of negative affect measure-
ment while at the same time minimizing the number of
interruptions to task performance.
A related limitation concerns the absence of any non-
subjective measures of negative affect, such as measures
of autonomic activity. Although it is no doubt true that
such measures would have invited consideration of addi-
tional questions, they were not necessary for answering
the question of interest in this study: What is the relation-
ship between the subjective experience of negative affect
and dACC activity during cognitive performance? In fact,
it is already known from both neuroimaging and electro-
physiological work that the dACC response to cogni-
tive performance is associated with various measures
of autonomic activity (e.g., Critchley et al., 2005; Hajcak
et al., 2003; for a review, see Weinberg, et al., 2012).
However, it remained unknown to what extent the dACC
is associated with self-reports of negative affect during
cognitive performance. Although self-reported subjec-
tive experience is only one correlate of negative affect,
it may be a particularly important correlate. Indeed, self-
report remains a gold standard method in the literature
on and clinical assessment of physical pain. As Coghill,
McHaffie, and Yen (2003) wrote in discussing the asso-
ciation of subjective reports of pain and the dACC re-
sponse, “pain is defined by the first-person experiential
perspective and must be diagnosed and treated with
significant consideration of the subjective report…[The]
subjective report will likely remain the single most reliable
index of the magnitude of pain” (p. 8542). In this spirit,
we would argue that in the measurement of any affective
response, such as the frustration felt in response to per-
forming a difficult task, self-report should be treated as
the gold standard method.
A final limitation regards the exclusive focus on the
emotion of frustration. As a group, the sample reported
very low amounts of anxiety and felt unpleasantness. Still,
exploratory analyses suggest that these two dimensions
may explain the dACC response during cognitive per-
formance. A fruitful direction for future research will be
to more systematically examine distinct dimensions of
negative affect as they relate to distinct aspects of cogni-
tive performance, such as the monitoring of information
processing for errors (captured by Go trials in Stop-signal
blocks) and the detection of errors (captured by Stop-
failure trials).
Conclusion
Nearly 75 years ago, Papez (1937) suggested that the
ACC “…is the seat of dynamic vigilance by which environ-
mental experiences are endowed with an emotional con-
sciousness” (p. 737). Intriguingly, Papez foreshadowed the
two faces of the dACC presented here: a cognitive module
for vigilantly monitoring the current state of the organism
and an affective alarm that endows events—such as simple
errors in the performance of a cognitive task—with an
emotional consciousness. One particularly influential view
(Bush et al., 2000) might suggest that Papezʼs comment
linked dACC with the cognitive module and rACC with
the affective module. This study underscores the possibility
that the dACC is central to both.
Acknowledgments
The authors thank Edward Kobayashi and John Mezzanotte for
their assistance with data collection; Russ Poldrack and Adam
Aron for helpful advice in the early stages of the project; and
Matthew Botvinick, Wouter Kool, and several anonymous review-
ers for comments on the manuscript. For generous support, the
authors also thank the Brain Mapping Medical Research Organi-
zation, Brain Mapping Support Foundation, Pierson-Lovelace
Foundation, The Ahmanson Foundation, William M. and Linda R.
Dietel Philanthropic Fund at the Northern Piedmont Community
Foundation, Tamkin Foundation, Jennifer Jones-Simon Founda-
tion, Capital Group Companies Charitable Foundation, Robson
Family, and Northstar Fund.
Reprint requests should be sent to Naomi I. Eisenberger, De-
partment of Psychology, 1285 Franz Hall, University of California,
Los Angeles, Los Angeles, CA 90095-1563, or via e-mail: neisenbe@
ucla.edu.
REFERENCES
Band, G. P. H., Van der Molen, M. W., & Logan, G. D. (2003).
Horse-race model simulations of the stop-signal procedure.
Acta Psychologica, 112, 105–142.
Botvinick, M., Braver, T., Barch, D., Carter, C., & Cohen, J.
(2001). Conflict monitoring and cognitive control.
Psychological Review, 108, 624.
Botvinick, M. M. (2007). Conflict monitoring and decision
making: Reconciling two perspectives on anterior cingulate
function. Cognitive, Affective, & Behavioral Neuroscience,
7, 356–366.
Botvinick, M. M., Huffstetler, S., & McGuire, J. T. (2009). Effort
discounting in human nucleus accumbens. Cognitive,
Affective, & Behavioral Neuroscience, 9, 16–27.
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 433–436.
Bush, G., Luu, P., & Posner, M. (2000). Cognitive and emotional
influences in anterior cingulate cortex. Trends in Cognitive
Sciences, 4, 215–222.
Chiu, P., & Deldin, P. (2007). Neural evidence for enhanced
error detection in major depressive disorder. American
Journal of Psychiatry, 164, 608–616.
Coghill, R. C., McHaffie, J. G., & Yen, Y.-F. (2003). Neural
correlates of interindividual differences in the subjective
experience of pain. Proceedings of the National Academy
of Sciences, U.S.A., 100, 8538–8542.
Cohen, J., Asarnow, R. F., Sabb, F. W., Bilder, R. M., Bookheimer,
S. Y., Knowlton, B. J., et al. (2010). Decoding developmental
differences and individual variability in response inhibition
through predictive analyses across individuals. Frontiers in
Human Neuroscience, 4, 1–12.
Congdon, E., Mumford, J. A., Cohen, J. R., Galvan, A., Aron,
A. R., Xue, G., et al. (2010). Engagement of large-scale
networks is related to individual differences in inhibitory
control. Neuroimage, 53, 653–663.
Critchley, H. D., Tang, J., Glaser, D., Butterworth, B., &
Dolan, R. J. (2005). Anterior cingulate activity during error
and autonomic response. Neuroimage, 27, 885–895.
Spunt et al.
1763
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Debener, S., Ullsperger, M., Siegel, M., Fiehler, K., von Cramon,
D. Y., & Engel, A. K. (2005). Trial-by-trial coupling of
concurrent electroencephalogram and functional magnetic
resonance imaging identifies the dynamics of performance
monitoring. Journal of Neuroscience, 25, 11730–11737.
Devinsky, O., Morrell, M., & Vogt, B. (1995). Contributions
of anterior cingulate cortex to behaviour. Brain, 118,
279–306.
Duncan, J., & Owen, A. M. (2000). Common regions of
the human frontal lobe recruited by diverse cognitive
demands. Trends in Neurosciences, 23, 475–483.
Eisenberger, N. I. (in press). The pain of social disconnection:
Examining the shared neural underpinnings of physical and
social pain. Nature Reviews Neuroscience.
Eisenberger, N. I., & Lieberman, M. D. (2004). Why rejection
hurts: A common neural alarm system for physical and
social pain. Trends in Cognitive Sciences, 8, 294–300.
Eisenberger, N. I., Lieberman, M. D., & Williams, K. (2003).
Does rejection hurt? An fMRI study of social exclusion.
Science (New York, NY), 302, 290–292.
Etkin, A., Egner, T., & Kalisch, R. (2011). Emotional processing
in anterior cingulate and medial prefrontal cortex. Trends
in Cognitive Sciences, 15, 85–93.
Falkenstein, M., Hohnsbein, J., Hoormann, J., & Blanke, L.
(1991). Effects of crossmodal divided attention on late ERP
components. II. Error processing in choice reaction tasks.
Electroencephalography and Clinical Neurophysiology,
78, 447–455.
Fitzgerald, K. D., Welsh, R. C., Gehring, W. J., Abelson, J. L., Himle,
J. A., Liberzon, I., et al. (2005). Error-related hyperactivity
of the anterior cingulate cortex in obsessive-compulsive
disorder. Biological Psychiatry, 57, 287–294.
Foltz, E., & White, L. (1962). Pain “relief ” by frontal
cingulumotomy. Journal of Neurosurgery, 19, 89–100.
Gehring, W., Goss, B., Coles, M., Meyer, D., & Donchin, E.
(1993). A neural system for error detection and
compensation. Psychological Science, 4, 385–390.
Gehring, W., Himle, J., & Nisenson, L. (2000). Action-monitoring
dysfunction in obsessive-compulsive disorder. Psychological
Science, 11, 1–7.
Grinband, J., Savitskaya, J., Wager, T. D., Teichert, T., Ferrera,
V. P., & Hirsch, J. (2011). The dorsal medial frontal cortex
is sensitive to time on task, not response conflict or error
likelihood. Neuroimage, 57, 303–311.
Hajcak, G., & Foti, D. (2008). Errors are aversive: Defensive
motivation and the error-related negativity. Psychological
Science, 19, 103–108.
Hajcak, G., McDonald, N., & Simons, R. F. (2003). To err is
autonomic: Error-related brain potentials, ANS activity,
and post-error compensatory behavior. Psychophysiology,
40, 895–903.
Holroyd, C. B., & Coles, M. G. (2002). The neural basis of
human error processing: Reinforcement learning, dopamine,
and the error-related negativity. Psychological Review, 109,
679–709.
Kober, H., Barrett, L. F., Joseph, J., Bliss-Moreau, E., Lindquist, K.,
& Wager, T. D. (2008). Functional grouping and cortical-
subcortical interactions in emotion: A meta-analysis of
neuroimaging studies. Neuroimage, 42, 998–1031.
Kool, W., McGuire, J. T., Rosen, Z. B., & Botvinick, M. M.
(2010). Decision making and the avoidance of cognitive
demand. Journal of Experimental Psychology: General,
139, 665–682.
Logan, G. (1994). On the ability to inhibit thought and
action: A usersʼ guide to the stop signal paradigm.
In D. Dagenbach & T. Carr (Eds.), Inhibitory processes
in attention, memory, and language (pp. 189–240).
San Diego, CA: Academic Press.
Logan, G., & Cowan, W. (1984). On the ability to inhibit simple
and choice reaction time responses: A model and a method.
Journal of Experimental Psychology, 10, 276–291.
Luu, P., Collins, P., & Tucker, D. (2000). Mood, personality,
and self-monitoring: Negative affect and emotionality in
relation to frontal lobe mechanisms of error monitoring.
Journal of Experimental Psychology: General, 129,
43–60.
Macey, P., Macey, K., Kumar, R., & Harper, R. (2004).
A method for removal of global effects from fMRI time
series. Neuroimage, 22, 360–366.
McGuire, J. T., & Botvinick, M. M. (2010). Prefrontal cortex,
cognitive control, and the registration of decision costs.
Proceedings of the National Academy of Sciences, U.S.A.,
107, 7922–7926.
Olvet, D., & Hajcak, G. (2008). The error-related negativity
(ERN) and psychopathology: Toward an endophenotype.
Clinical Psychology Review, 28, 1343–1354.
Papez, J. (1937). A proposed mechanism of emotion.
Archives of Neurology & Psychiatry, 38, 725.
Paus, T. (2001). Primate anterior cingulate cortex: Where
motor control, drive and cognition interface. Nature
Reviews Neuroscience, 2, 417–424.
Paus, T., Koski, L., & Caramanos, Z. (1998). Regional differences
in the effects of task difficulty and motor output on blood
flow response in the human anterior cingulate cortex:
A review of 107 PET activation studies. NeuroReport, 9,
37–47.
Pessoa, L. (2008). On the relationship between emotion and
cognition. Nature Reviews Neuroscience, 9, 148–158.
Poldrack, R. (2006). Can cognitive processes be inferred
from neuroimaging data? Trends in Cognitive Sciences,
10, 59–63.
Quirk, M. E., Letendre, A. J., Ciottone, R. A., & Lingley,
J. F. (1989). Anxiety in patients undergoing MR imaging.
Radiology, 170, 463–466.
Rainville, P. (2002). Brain mechanisms of pain affect and
pain modulation. Current Opinion in Neurobiology, 12,
195–204.
Rainville, P., Duncan, G. H., Price, D. D., Carrier, B., &
Bushnell, M. C. (1997). Pain affect encoded in human
anterior cingulate but not somatosensory cortex. Science
(New York, NY), 277, 968–971.
Ridderinkhof, K. R., Ullsperger, M., Crone, E. A., &
Nieuwenhuis, S. (2004). The role of the medial frontal
cortex in cognitive control. Science (New York, NY),
306, 443–447.
Shackman, A. J., Salomons, T. V., Slagter, H. A., Fox, A. S.,
Winter, J. J., & Davidson, R. J. (2011). The integration
of negative affect, pain and cognitive control in the
cingulate cortex. Nature Reviews Neuroscience, 12,
154–167.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D.,
Crivello, F., Etard, O., Delcroix, N., et al. (2002).
Automated anatomical labeling of activations in
SPM using a macroscopic anatomical parcellation
of the MNI MRI single-subject brain. Neuroimage,
15, 273–289.
van Steenbergen, H., Band, G. P. H., & Hommel, B. (2009).
Reward counteracts conflict adaptation: Evidence for a role
of affect in executive control. Psychological Science, 20,
1473–1477.
van Steenbergen, H., Band, G. P. H., & Hommel, B. (2010).
In the mood for adaptation: How affect regulates conflict-
driven control. Psychological Science, 21, 1629–1634.
van Steenbergen, H., Booij, L., Band, G. P. H., Hommel, B.,
& Does, A. J. W. (2012). Affective regulation of
cognitive-control adjustments in remitted depressive
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1764
Journal of Cognitive Neuroscience
Volume 24, Number 8
patients after acute tryptophan depletion. Cognitive,
Affective, & Behavioral Neuroscience, 12, 280–286.
Vogt, B. A. (2005). Pain and emotion interactions in subregions
of the cingulate gyrus. Nature Reviews Neuroscience, 6,
533–544.
Weinberg, A., Olvet, D. M., & Hajcak, G. (2010). Increased
error-related brain activity in generalized anxiety disorder.
Biological Psychology, 85, 472–480.
Weinberg, A., Riesel, A., & Hajcak, G. (2012). Integrating
multiple perspectives on error-related brain activity:
The ERN as a neural indicator of trait defensive reactivity.
Motivation and Emotion, 36, 84–100.
Whitty, C., Duffield, J., & Tow, P. (1952). Anterior cingulectomy
in the treatment of mental disease. Lancet, 1, 475–481.
Yarkoni, T., Poldrack, R. A., Nichols, T. E., Van Essen, D. C.,
& Wager, T. D. (2011). Large-scale automated synthesis of
human functional neuroimaging data. Nature Methods, 8,
665–670.
Yeung, N., Botvinick, M., & Cohen, J. (2004). The neural basis
of error detection: Conflict monitoring and the error-related
negativity. Psychological Review, 111, 931–959.
Zaki, J., Weber, J., Bolger, N., & Ochsner, K. (2009). The neural
bases of empathic accuracy. Proceedings of the National
Academy of Sciences, 106, 11382–11387.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
4
/
8
2
4
1
/
7
8
5
/
3
1
1
7
9
5
4
3
4
/
1
1
0
7
1
7
o
8
c
1
n
7
_
1
a
/
_
j
0
o
0
c
2
n
4
2
_
a
p
_
d
0
0
b
2
y
4
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Spunt et al.
1765