The Neural Representation of Events Is Dominated by
Elements that Are Most Reliably Present
Konstantinos Bromis1*, Petar P. Raykov1*, Leah Wickens1,
Warrick Roseboom1,2, and Chris M. Bird1
Abstract
■ An episodic memory is specific to an event that occurred at a
particular time and place. However, the elements that constitute
the event—the location, the people present, and their actions
and goals—might be shared with numerous other similar
events. Does the brain preferentially represent certain elements
of a remembered event? If so, which elements dominate its neu-
ral representation: those that are shared across similar events, or
the novel elements that define a specific event? We addressed
these questions by using a novel experimental paradigm com-
bined with fMRI. Multiple events were created involving conver-
sations between two individuals using the format of a television
chat show. Chat show “hosts” occurred repeatedly across multi-
ple events, whereas the “guests” were unique to only one event.
Before learning the conversations, participants were scanned
while viewing images or names of the (famous) individuals to
be used in the study to obtain person-specific activity patterns.
After learning all the conversations over a week, participants
were scanned for a second time while they recalled each event
multiple times. We found that during recall, person-specific
activity patterns within the posterior midline network were rein-
stated for the hosts of the shows but not the guests, and that
reinstatement of the hosts was significantly stronger than the
reinstatement of the guests. These findings demonstrate that
it is the more generic, familiar, and predictable elements of an
event that dominate its neural representation compared with
the more idiosyncratic, event-defining, elements. ■
INTRODUCTION
Our lives progress through a series of unique events.
When we remember these events, we reactivate their neu-
ral representations. Although the events themselves are
unique, defined by the combination of what happened
where and when, many will share common elements, such
as the same people or location. It is not known how such
repeated elements are activated during memory recall. Do
the predictable and repeated elements of an event domi-
nate its neural representation? If so, this might provide a
structure to retrieve less predictable, more idiosyncratic
elements. Alternatively, are the unique elements that dis-
tinguish similar events represented more robustly? The
current study aimed to address these questions.
When humans experience or recall an event, an
event-specific pattern of brain activity—as measured using
fMRI—is elicited within regions of the cortex (Raykov,
Keidel, Oakhill, & Bird, 2021; Chen et al., 2017; Oedekoven,
Keidel, Berens, & Bird, 2017; Lee & Kuhl, 2016; Bird,
Keidel, Ing, Horner, & Burgess, 2015; St-Laurent, Abdi,
& Buchsbaum, 2015; Kuhl & Chun, 2014). These
effects are observed most frequently in the brain’s poste-
rior medial network (Cooper & Ritchey, 2020; Ranganath
1School of Psychology, University of Sussex, 2School of Engi-
neering and Informatics, University of Sussex
*Co-first authorship.
& Ritchey, 2012), and although they reflect the neural rep-
resentation of an event, it is unclear what drives them.
Under the view that episodic memory is holistic in nature
(Tulving, 1983), it might be expected that all elements that
comprise an event contribute equally. Some recent evi-
dence speaks against that because the spatial context that
an event takes place in has been shown to be a major fac-
tor in determining its neural representation (Robin,
Buchsbaum, & Moscovitch, 2018). In this study, we
wanted to broadly compare nonspatial elements that
were repeated, familiar, and more predictable with those
that were event-unique, unfamiliar, and less expected.
Elements that are repeatedly encountered across events
inevitably become more familiar to us and also more pre-
dictable. Any viewer of the TV show “Friends” will become
familiar with the six main characters and would expect
them to feature in any new episode. Predictive coding
accounts of perception argue that we generate predictions
that serve to “explain away” variance in incoming sensory
information (Clark, 2013). Thus, during perception, the
activity of neuronal populations that code the predictable
features of an event are suppressed, whereas those that
code the unexpected or novel features are enhanced
(Sohoglu & Davis, 2020; Aitchison & Lengyel, 2017;
Friston, 2005). Importantly, the enhanced elements of
our perceptual experience are also encoded better into
memory, consistent with the view that “prediction errors”
© 2022 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Journal of Cognitive Neuroscience 34:3, pp. 517–531
https://doi.org/10.1162/jocn_a_01802
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
drive new learning (Quent, Henson, & Greve, 2021; Niv &
Schoenbaum, 2008). Given that the brain appears to prior-
itize unfamiliar and unexpected information, we might
expect that when we recall an event, it is the less predict-
able elements that dominate our representation of the
event. For example, if we recall a specific episode of
“Friends” that involved the unexpected arrival of one of
the sisters of one of the main characters, our representa-
tion of this individual might be more robust than the
others, as her presence is a key identifying element of this
particular episode.
Memory recall has also been argued to play a role in
updating our internal model, by the “off-line” generation
of fictive prediction errors (Barron, Auksztulewicz, &
Friston, 2020; see also Hinton, Dayan, Frey, & Neal,
1995). More generally, memory recall has long been
thought to involve reinstating both the processes and rep-
resentations that were active during encoding (Nyberg,
Habib, McIntosh, & Tulving, 2000; Morris, Bransford, &
Franks, 1977). It is therefore plausible that memory
recall—similarly to memory encoding—might be biased
toward reinstating representations of the more unex-
pected elements of events (see Wittkuhn, Chien, Hall-
McMaster, & Schuck,2021).
However, it has been argued that recent repeated expe-
riences are most useful in predicting future experiences
because they are more likely to be encountered again
(Anderson & Milson, 1989). By contrast, idiosyncratic
experiences are poor for making generalizations about
the future (Sherman & Turk-Browne, 2020). Behaviorally,
it is well established that prior knowledge exerts a strong
influence over what aspects of an event are recalled
(Popov & Reder, 2020; Smith, Hasinski, & Sederberg,
2013; Poppenk, Köhler, & Moscovitch, 2010; Brewer &
Treyens, 1981). Accordingly, we may expect that it is those
elements of an event that are more reliably present that
will dominate its neural representation.
More generally, it is well established that repeated expo-
sure to the same material is beneficial for memory ( Van
Strien, Hagenbeek, Stam, Rombouts, & Barkhof, 2005;
Glenberg, Smith, & Green, 1977). However, the situation
is less clear-cut when the same items are repeatedly
encoded in different contexts. Here, although the items
may become more familiar and are better recognized,
their associations with the contexts that they were experi-
enced in can weaken (Sievers, Bird, & Renoult, 2019; Yassa
& Reagh, 2013).
Although repetition can result in better memory, it is
also the case that more novel and distinctive items are also
better remembered (Hunt, 1995). This memory advantage
for more distinct items affects both recollection- and
familiarity-based recognition judgments (Kishiyama &
Yonelinas, 2003). Furthermore, novelty can act at the level
of a stimulus or whether a particular stimulus is novel
within a specific context (Ranganath & Rainer, 2003).
Taken together, there is substantial evidence that two
broad factors can determine the degree to which elements
of an event are more memorable. On the one hand, there
are items that are reliably present and predictable—often
as a result as having been frequently encountered in the
same context. On the other hand, there are items that
are more idiosyncratic and unexpected—perhaps only
occurring once in a context and therefore uniquely identi-
fying an event. These factors have different effects on
memory in different situations, indeed there are situations
where memory is superior both for highly unexpected and
highly expected events (Quent, Henson, & Greve, 2021).
Our focus in this study is how these factors affect the neu-
ral representation of complex naturalistic events. Specifi-
cally, if a complex memory can be accurately recalled, is
it those elements that are most reliably present that dom-
inate the neural representation or is it the less familiar
events that are uniquely associated with an event?
We first recorded fMRI patterns of activity elicited when
participants viewed images or read names of famous celeb-
rities. Following this, participants learned nine fictional
conversations between two celebrities set within a televi-
sion “chat show” format. The repeated elements were the
hosts of the shows (three in total), whereas the unique ele-
ments were the guests (nine in total). After they had learned
all of the conversations, participants then repeatedly
recalled them in a second fMRI session. By using the
person-specific patterns of activity from Session 1, we
were able to examine whether the hosts or the guests
were more robustly reinstated when participants recalled
the conversations. All analyses were performed on ROIs
from the posterior midline (PM) network identified in
a previous study of event memory (Robin, 2018).
METHODS
Participants
Thirty-one (21 women, 10 men) participants took part
in the experiment. Participants were aged 19–30 years
(mean = 24 years, SD = 3.54 years) and did not have a his-
tory of any psychiatric or neurological disorders. All partic-
ipants were right-handed and were fluent English
speakers. Four participants were excluded from any anal-
yses: two did not complete the experiment, one because
of excessive movement in the scanner, and one for failing
to recall three of the guests during the memory screening
test. Therefore, data from 27 participants were analyzed.
Note three participants had corrupted audio recording
and were not included in reports of the memory screening
test. This sample size is consistent with previous studies
examining fMRI pattern effects about stories or person
decoding (Raykov, Keidel, Oakhill, & Bird, 2020; di Oleggio
Castello, Halchenko, Guntupalli, Gors, & Gobbini, 2017;
Zadbood, Chen, Leong, Norman, & Hasson, 2017).
Informed consent was obtained from all participants
before the experiment, and they were reimbursed £40
for their time. This project was approved by the Brighton
and Sussex Medical School Research Governance and
Ethics Committee.
518
Journal of Cognitive Neuroscience
Volume 34, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
We also collected follow-up data from a group of sepa-
rate participants who completed a similar task online. This
group comprised 37 participants (20 women, 17 men)
with a mean age of 24.61 years (±4.94 years). We excluded
one participant who failed to learn the conversations after
five learning sessions. Therefore, 36 participants from the
online study were included in the analyses. Participants
received payment through online recruitment platform
Prolific (https://www.prolific.co/) for each of the learning
sessions. The project was approved by the University of
Sussex Cross School Research Ethics Committee.
Stimuli
We ran a pilot study to select 12 celebrities who would be
familiar to our sample (see https://osf.io/zpcv3/). Forty-
eight pictures of 12 (six men, six women) famous individ-
uals were used in Session 1. Four different pictures
sourced from Google Images were used for each
famous individual. The pictures only showed the famous
individual and were converted to grayscale using Adobe
Photoshop CC19. Additionally, five pictures of nonfamous
individuals were used in the experiment.
Nine short fictitious conversations were written by the
research team (see https://osf.io/zpcv3/). These were
learned after Session 1 and recalled in the scanner in
Session 2. The conversations were relatively short (162.5 ±
16.1 words on average) and took the form of chat show
conversations using a question-and-answer structure.
The conversations involved two people, a guest, and a
host, taken from the 12 famous celebrities. On average,
a similar number of words were “spoken” by the hosts
(mean = 80.7 words) and the guests (mean = 81.7).
The topic of each conversation was unique (e.g., giving
to charity, social media; see https://osf.io/zpcv3/ for the
complete transcripts of all conversations). Each conversa-
tion was associated with a particular day (Monday,
Wednesday, or Friday) and week ( Week 1, 2, or 3). Partic-
ipants were required to learn when each conversation
took place, as this was how memory would be cued in
the scanner. Each of the three hosts was associated with
three conversations that occurred on a specific day of
the week (Host 1, Mondays; Host 2, Wednesdays; and
Host 3, Fridays; see Figure 1). The guests were only asso-
ciated with one of the nine conversations. A pilot experi-
ment with an independent group of participants ensured
that there were not substantial differences between the
conversations in how memorable and interesting they
were (see https://osf.io/zpcv3/). A picture was created
using Photoshop to illustrate the context of each conver-
sation and to make them appear more plausible. The
picture presented the host and the guest sitting in a TV
studio (see Figure 1C). Three background pictures of TV
studios were used overall, which were consistent for the
three hosts (e.g., Studio 1 for Mondays, Studio 2 for
Wednesdays). Three counterbalancing lists were created
where the identity of hosts and guests was varied across
participants.
Procedure
fMRI Study
Prescanning. Before taking part in the fMRI experiment,
participants completed a short online questionnaire
establishing their knowledge of the famous individuals
used in the experiment (see https://osf.io/zpcv3/). Partici-
pants rated the following from 1–5: how familiar they are
with the person, how well they can imagine them, how
much they know about their career and/or personal life,
and whether they like the person. Participants were
selected if they responded to be familiar (e.g., responded
above 3) with the 12 celebrities used in the experiment.
Participants were also encouraged to learn more about
these celebrities before the experiment. Participants took
part in two fMRI sessions spaced approximately 7 days
apart (see Figure 1).
Scanning Session 1
In Session 1, images and names of 12 famous individuals
were presented in a blocked fMRI design. There were five
runs in total. Each run contained 24 blocks of trials, com-
prising one image and one text block for each of the 12
celebrities. In an image block, participants saw the four dif-
ferent images of the same individual (e.g., Daniel Craig).
Each of the four pictures were presented twice within a
single block, resulting in eight images per block (see
Figure 1). The presentation order within a block was ran-
domized. Each picture was presented for 800 msec with a
200-msec gap, during which a white fixation cross was
shown. Each block lasted 8 sec. In a text block, participants
saw multiple presentations of the name of 1 of the 12
famous individuals. Each text presentation was 800 msec,
and there was a 200-msec ISI. To increase engagement
with each stimulus, the font color and text position for
each name presentation was varied. Each image and text
name were presented on a black background and presen-
tation of identities across blocks was randomized.
In between picture and text blocks, participants per-
formed an odd–even number judgment task. This served
as an active baseline task. Here, participants saw a
sequence of four numbers randomly selected from the
range 1–98. Each number was presented for 1.8 sec and
was followed by a white fixation cross for 700 msec. There-
fore, a block of the odd–even task lasted 10 sec. The odd–
even task was followed by a red fixation cross presented
for 400 msec signaling the upcoming identity block. Par-
ticipants performed an odd-ball detection task on the
image blocks. In each run, there was an additional image
block that included a picture of an unfamiliar individual
embedded in the sequence of pictures of a famous indi-
vidual. These odd-ball blocks could appear at any point in
Bromis et al.
519
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
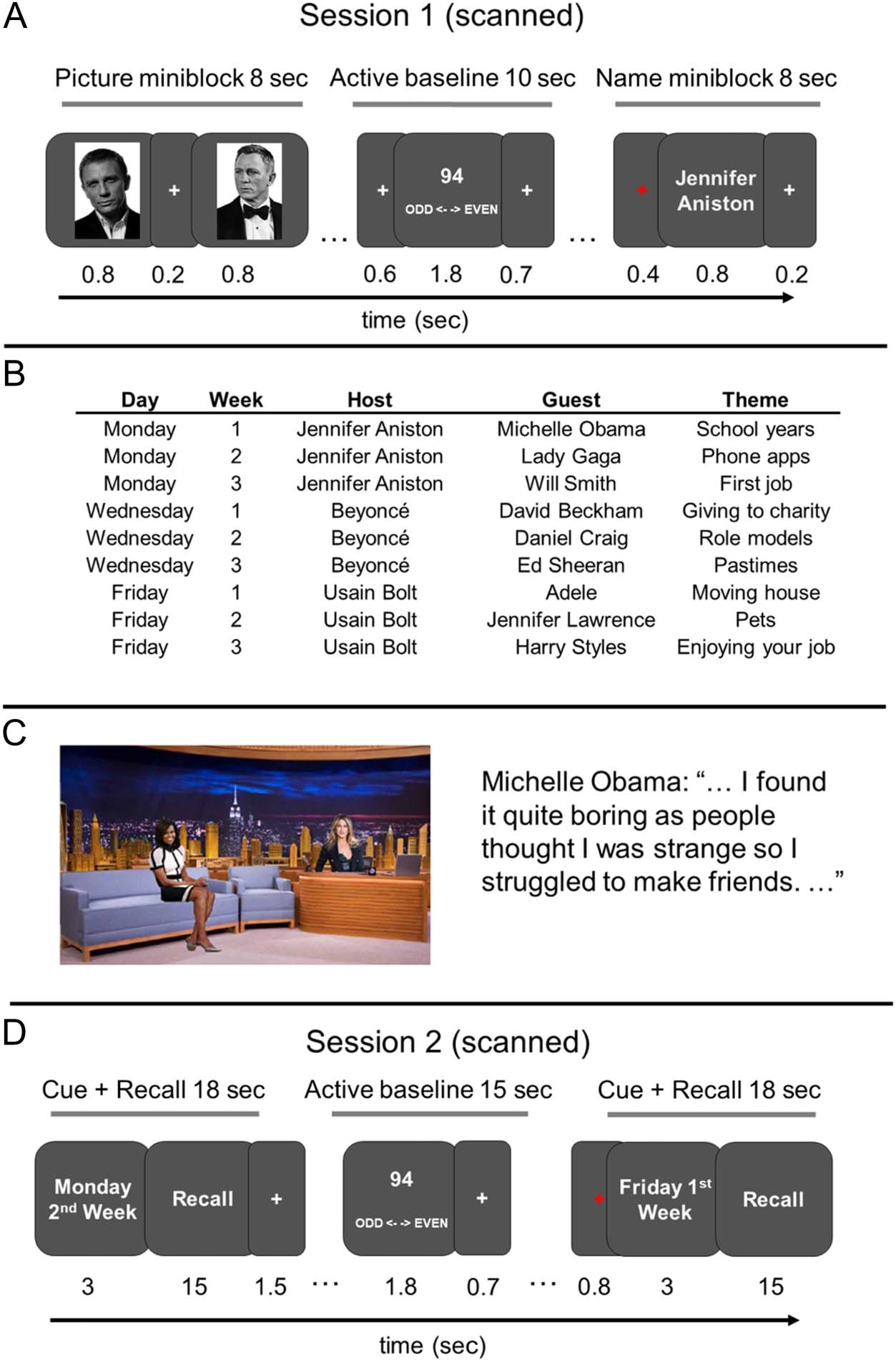
Figure 1. Study design.
Participants took part in two
fMRI sessions spaced a week
apart. (A) In Session 1,
participants viewed pictures and
names of famous individuals in
a blocked design (block length =
8 sec). There were 12 unique
identities, and within each run,
participants saw one picture and
one name block for each
identity. There were five runs in
total. (B) After Session 1,
participants were provided with
nine brief conversations to study
at their own leisure. Each
conversation included a host
and a guest discussing a
particular topic and was
associated with day and week.
The hosts were common to
three conversations, whereas
the guests were always unique.
(C) An image of the host and
guest in a studio was provided to
help participants visualize the
conversations. Conversations
were shown in the format of a
script (see https://osf.io/zpcv3/).
(D) In Session 2, participants
recalled the conversations
following a date cue. Each
conversation occurred once in
each run, and there were six runs
in total.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
the run apart from the first image block and were not
included in the main analyses. Identities were presented
in a randomized order.
Learning Phase
After completing scanning Session 1, participants were
provided with a list with the nine conversations involving
the 12 celebrities (see Figure 1). Participants were asked
to learn each conversation and were encouraged to visual-
ize each one, using the images provided. Participants
were instructed to learn the conversations well and were
told that their memory for the conversations would be
tested before being allowed to take part in the second
scanning session. They were given 4–6 days to learn the
conversations.
Following this, participants underwent a screening
test to ensure they had learned the conversations
before proceeding to the second scanning session. The
test was carried out at least 1 day before the scanning ses-
sion. Participants freely recalled all of the nine conversa-
tions in a random order, cued by the week and the day.
The experimenter provided feedback on parts of the con-
versations that the participants had failed to recall, and
all participants were asked to review the conversations
again before scanning.
Immediately before the second fMRI session, partici-
pants also provided subjective ratings on their memory
for the conversations. They rated on a scale of 0–100
how vividly and confidently they could remember the
conversations. They also rated how engaging they found
the conversations.
520
Journal of Cognitive Neuroscience
Volume 34, Number 3
Scanning Session 2
In Session 2, while in the scanner, participants were asked
to recollect the nine conversations they had previously
learned. There were six runs in total, and within each
run, participants recollected all nine of the conversations.
Participants were presented with the time information
(Monday first week) to cue their memory for the specific
conversation. The cue was presented for 3 sec and was
followed by a 15-sec recollection period. Participants were
asked to remember the conversation in as much detail
as possible for the 15 sec. Participants were provided with
the option to press a button if they failed to recollect the
conversation during that particular memory trial (e.g.,
because of mind-wandering). All such events were
removed from the main analyses. In between memory
blocks, participants performed the odd–even number
judgment task (described above) for 15 sec, which served
as active baseline. A red fixation cross followed the
number judgment task and signaled the upcoming recall
block (see Figure 1). Within each run, conversation
memory trials were randomized.
Online Study
In a follow-up online study, we asked a separate group of
participants to learn the nine conversations used in the
fMRI experiment and provide additional subjective ratings
about the conversations.
Similarly to the fMRI experiment, we initially pre-
screened participants to be familiar with the 12 celebrities
used in the experiment. Participants who were not familiar
with any of the 12 celebrities were not included in the
main experiment and were not included in the further
learning sessions.
Participants completed five learning sessions over a
week, each session released on a separate day. During
the learning sessions, participants read each of the nine
conversations at their leisure. The event cue (e.g., Monday
first) and the photoshopped studio picture showing
the host and the guest were presented to participants as
they read the conversations during each session. After
completing the reading session, participants answered
multiple-choice questions about the conversations. The
multiple-choice questions differed on each learning
session and aimed to help participants learn the conversa-
tions. The questions concerned the identities of the hosts
and guests, what they were talking about, and details from
the conversations. After the fifth learning session, partici-
pants provided subjective ratings on how confidently and
vividly they could remember each of the conversations.
Participants also rated how confidently they could remem-
ber the host and the guest for each conversation and how
important they perceived each one was for the conversa-
tion. Additionally, participants were asked to guess who
the host and the guest on a hypothetical conversation
happening on Week 4 (Monday, Wednesday, and Friday)
would be. This was an open-ended question, and partici-
pants were instructed to make a guess or write they do
not know.
MRI Acquisition
All images were acquired on a 3-T Siemens Prisma scanner
with a 32-channel head coil. To minimize head movement,
soft cushions were inserted into the head coil. Functional
images were acquired with a gradient-echo EPI sequence
with multiband acceleration factor of 8 with the following
parameters (repetition time = 0.8 sec, echo time =
33.1 msec, 52° flip angle, field of view = 208 × 180 mm,
72 slices with sliced thickness of 2 mm and isotropic
2 mm voxels). Two SpinEcho fieldmap runs with reversed
phase-encode blips in both anterior to posterior and poste-
rior to anterior were acquired with the same parameters as
the functional images. Separate field maps were acquired
for Sessions 1 and 2. A high-resolution T1-weighted image
was acquired with 3-D MPRAGE sequence (repetition
time = 2.4 sec, echo time = 2.14 sec, 8° flip angle, field
of view = 224 × 224mm and 0.8 mm isotropic voxels).
Image Preprocessing
SPM 12 ( Wellcome Department of Imaging Neuroscience)
was used to preprocess all images, except for the field
maps. For each session, we first spatially realigned the
functional images to the mean session image. Session-
specific field maps were estimated with command line
functions from FSL (Smith et al., 2004) and were applied
to the motion-corrected data to correct for image distor-
tions (Hutton et al., 2002). The anatomical image was
segmented into gray, white, and cerebrospinal fluid using
tissue probability maps, and was coregistered to the mean
functional image. The segmented images were used to
estimate deformation fields, which were applied to the
functional data to transform them to Montreal Neurologi-
cal Institute space. A 3-mm smoothing FWHM Gaussian
kernel was applied to the data as recommended by
previous work, showing that a small amount of smoothing
can improve sensitivity of multivoxel pattern analyses
(Hendriks, Daniels, Pegado, & Op de Beeck, 2017;
Gardumi et al., 2016).
Data Analysis
Behavioral Data
In both scanning sessions, participants completed an odd–
even number judgment task that acted as an active base-
line. We analyzed accuracy and RTs from both sessions to
ensure participants were paying attention throughout the
main tasks.
We carried out a post hoc analysis of data from the
screening test before the second scanning session. This
analysis used the well-established procedure for scoring
Bromis et al.
521
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
performance on tests of prose recall (e.g., Wechsler,
1945). The script for each conversation was divided
into discrete “idea units,” and a point was allocated for
successfully recalling each unit. For each conversation,
we scored units spoken by the host and the guest sepa-
rately. Examples of the full script divided into units and
participants’ recalled conversations are available online
(https://osf.io/zpcv3/).
During the second scanning session, participants could
press a button to indicate that they could not retrieve the
conversation on this trial. These trials were not included
into further analyses. Participants provided subjective
ratings on a scale of 0–100 on how vividly and confidently
they could recall the conversations. Participants also rated
how familiar and engaging they found the conversations.
These ratings were averaged across participants separately
for each conversation.
In the online study, a separate group of participants pro-
vided subjective confidence and vividness ratings about
the same conversations. Participants also provided subjec-
tive ratings in their confidence to remember the host and
the guest for each conversation and their importance.
MRI Data
We note that data from the last run of Session 2 from 3 out
of the 27 participants were lost because of a technical
issue. Therefore, these participants had only five runs
rather than six from Session 2. MRI data were analyzed
with SPM 12, the CosMoMVPA toolbox (Oosterhof,
Connolly, & Haxby, 2016), and custom scripts written in
MATLAB ( Version 2017b, The MathWorks, Inc.). All analy-
ses were conducted on Montreal Neurological Institute
normalized images. The BrainNet Viewer toolbox was
used for visualizing the ROIs (see Figure 5A), and
bspmview toolbox was used for visualizing the whole-
brain parcellations reported in supplementary materials.
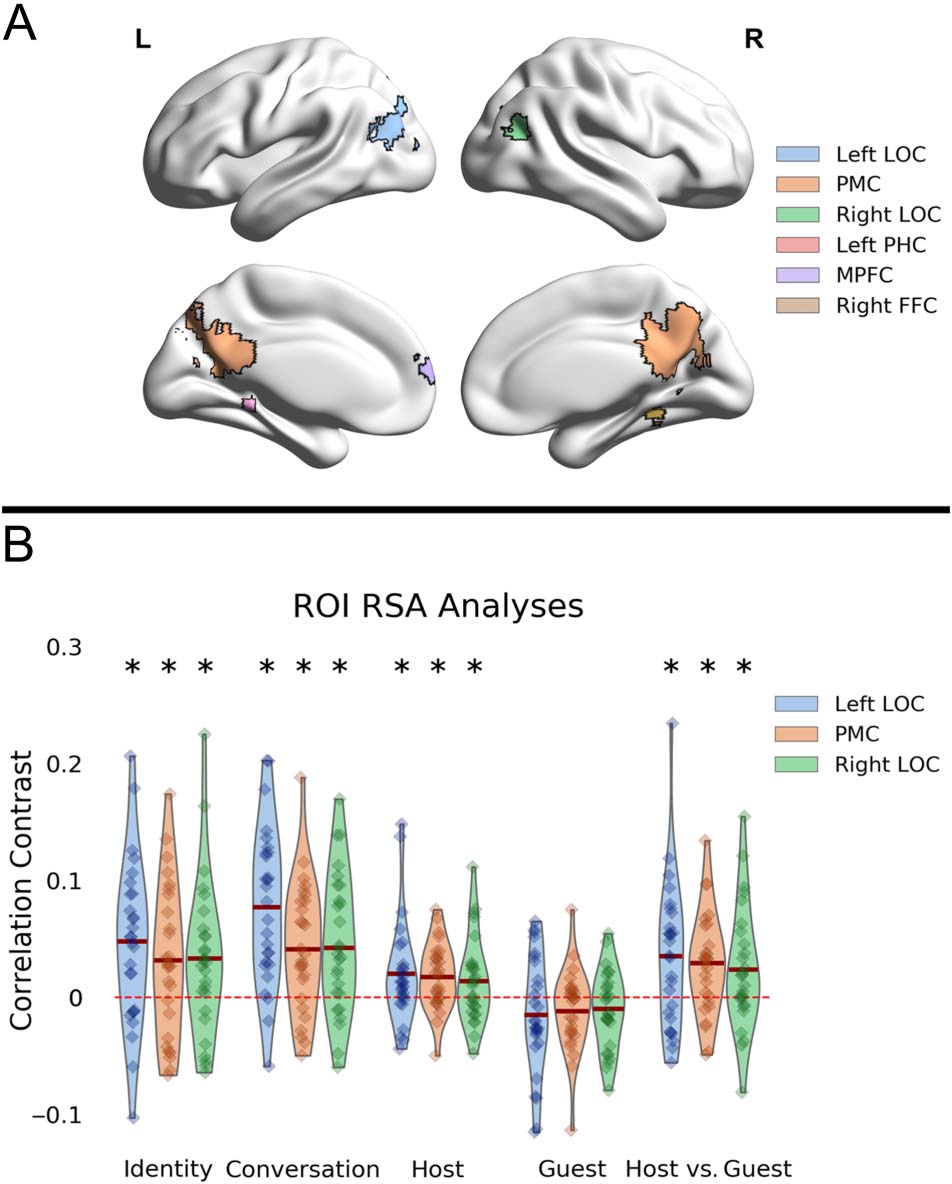
ROI Definition
We carried out our analyses on regions associated with the
PM network (see Introduction). The ROIs were taken from
a previous study of multielement event recall by Robin
et al. (2018). We used their multifeature ROI, which is a
set of regions comprising voxels that could classify
between different aspects of events—locations, people,
and objects (see Figure 5A). This comprises five different
regions: posterior medial cortex (PMC), dorsal medial pFC
(MPFC), left and right superior lateral occipital cortex
(LOC) extending to the angular gyrus in the lateral parietal
cortex (referred to as angular gyrus by Robin and col-
leagues), and left and right parahippocampal gyrus.
General Linear Models
To estimate activation patterns for later use in the repre-
sentational similarity analyses (RSAs), we used general
linear models (GLMs). In each run from Session 1, a
separate regressor for each block was included, such
that picture and text blocks were modeled separately
(i.e., 24 regressors per run). All trial regressors were
entered in a single first-level model as the least squares
all method described in Mumford, Turner, Ashby, and
Poldrack (2012). The patterns (t maps) from picture
and text blocks for the same identity within each run
were then averaged. Therefore, from all five runs, there
were 60 patterns (12 per run). After averaging the pic-
ture and text blocks, this resulted in five patterns for
each of the 12 famous individuals included in the exper-
iment. In Session 2, each retrieval trial block was modeled
with a separate regressor (nine per run) in a least squares
all approach. This resulted in 54 patterns for the nine con-
versations used (six per conversation). Separate regres-
sors of no interest for the six motion parameters, a session
constant term, and a high pass filter with cutoff of 1/128 Hz
were included in all GLMs.
RSA
We used the GLM estimated t maps as inputs to our RSAs.
A series of RSAs were carried out to examine whether
repeated features are more strongly represented during
recollection. Contrast matrices for each analysis are shown
in Figure 4. All similarity matrices were estimated using
Pearson correlation, and all correlation values were Fisher
transformed before computing further contrasts. Group-
level one-sample t test against zero was used to examine
the significance of the RSA contrasts (α = .05).
Before investigating whether the hosts and/or the
guests were represented in the ROIs during memory
retrieval, we first established which ROIs distinguished
the identities of the celebrities as well as the conversa-
tions. If an ROI could not discriminate between the celeb-
rities when presented in isolation, then it would not make
sense to seek evidence for identity-specific reactivation of
these patterns during memory retrieval. Similarly, if an ROI
could not discriminate between the conversations them-
selves during retrieval, then it would not make sense to
look for reactivation of the identities of the people taking
part in the conversations (see Supplementary Figure 5,
available online at https://osf.io/zpcv3/). Therefore, the
ROIs that we report showing a significant effect of “host”
or “guest” reinstatement had to not only pass the signifi-
cance threshold for these specific analyses but also show
significant effects in two additional independent analyses
(of “identity” and “conversation,” see below).
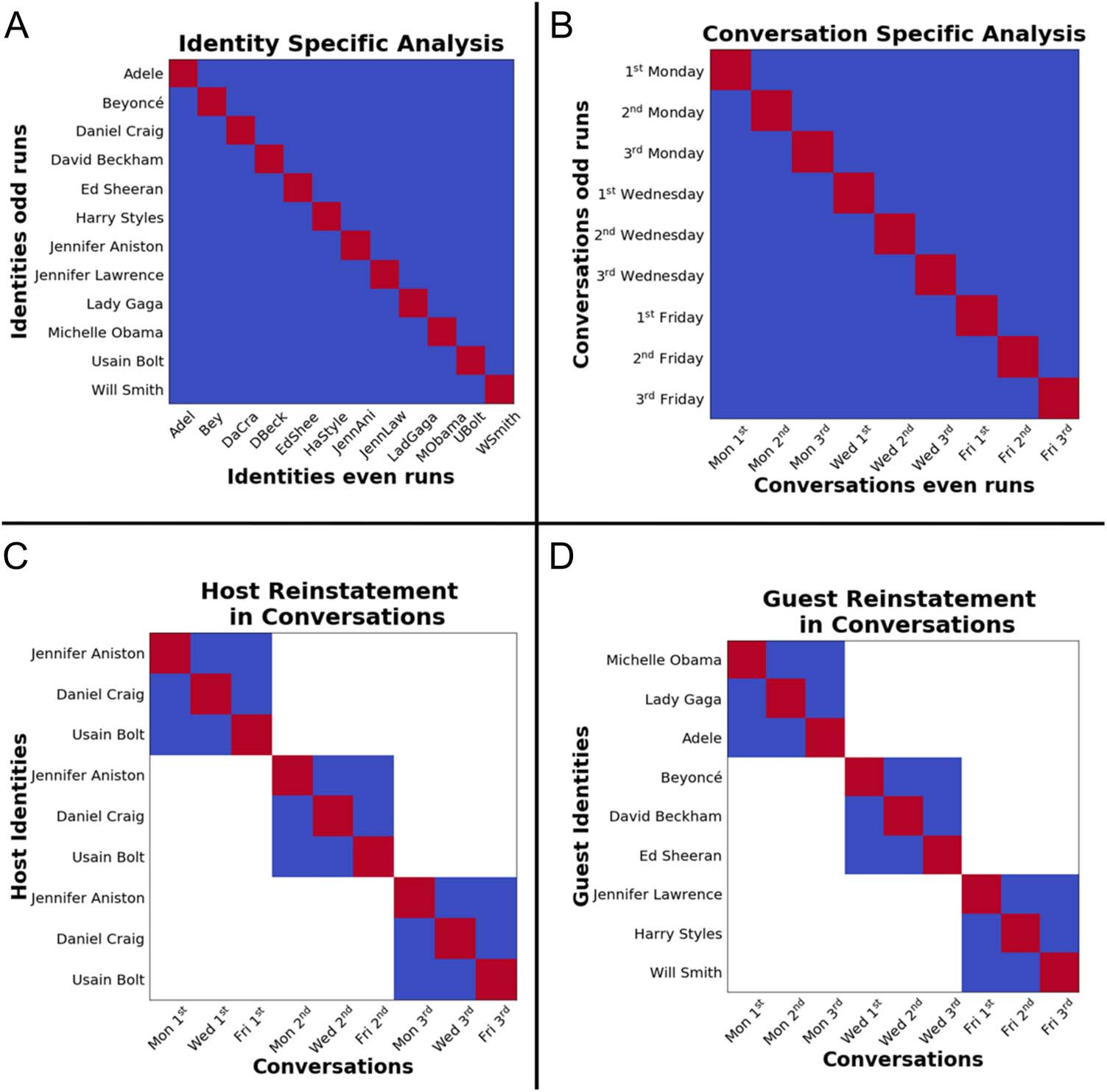
Identity. First, we examined which regions show reliable
identity-specific patterns (see Figure 4A). For this analysis,
only patterns from Session 1 were used. For each ROI, spa-
tial patterns of activity (t maps) for each identity were
extracted, vectorized, and used to construct an RSA matrix.
Patterns for the same identity in odd numbered runs and
even numbered runs were separately averaged. This
522
Journal of Cognitive Neuroscience
Volume 34, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
resulted in 12 identity patterns estimated from the odd
runs and 12 identity patterns estimated from the even runs.
The pairwise similarities between all of these patterns were
used to produce a 12 × 12 correlation matrix. The resulting
RSA matrix represents the neural similarity between the 12
identities. The diagonal values represent the matching
identities across runs, and the 132 off-diagonal values rep-
resent the similarity between nonmatching identities. To
examine which regions show reliable identity-specific
patterns, the mean average similarity between matching
identities versus the mean average similarity between
nonmatching identities was compared. ROIs that did
not show reliable identity-specific patterns were not
included into further analyses. Results from all ROIs are
available online at https://osf.io/zpcv3/.
Second, we examined whether the
Conversation.
remaining ROIs would exhibit reliable conversation-
specific effects. Using a similar logic, patterns for each
conversation across odd and even numbered runs were
separately averaged. Their similarity was then computed
using Pearson correlation, which resulted in a 9 × 9 similar-
ity matrix. Diagonal values represented similarity between
patterns of matching conversations, whereas off-diagonal
values represented values of mismatching conversations.
The mean matching similarity with the mean nonmatch-
ing similarity across conversations was compared. ROIs
that did not show reliable conversation-specific patterns
were also not included into further analyses.
In the host RSA, the patterns from Session 1 for
Hosts.
the host identities were averaged across all runs. Similarly,
all conversation patterns, from Session 2, were averaged
across all runs. The correlation between the host patterns
and the conversations was computed. The correlation
matrix was constructed such that the diagonal values rep-
resented the correlation between host identity (e.g.,
Jennifer Aniston pattern from Session 1) and the con-
versations with matching host. The off-diagonal values
represented correlation between host identity and mis-
matching conversations (e.g., where Jennifer Aniston was
not the host; see Figure 4C). Note that the host patterns
were repeated within the correlation matrix, and there-
fore, some off-diagonal values were not included into
the analyses. The contrast between matching host to con-
versation patterns and the mismatching patterns was
computed. This was done to examine whether host pat-
terns were reinstated during retrieval. ROIs that did not
show a significant host-specific reinstatement were not
removed from further analyses, as it was possible that they
would show a guest-specific reinstatement.
Guests. Afterward, the guest identity-specific patterns
from Session 1 were extracted and averaged them over
all runs. The similarity between guest patterns from Ses-
sion 1 and the conversation patterns from Session 2 was
computed. This resulted in a 9 × 9 correlation matrix
(see Figure 4D). The diagonal values represent similarity
between matching guest and conversation patterns (e.g.,
Michelle Obama Session 1 pattern and conversation where
Michelle Obama was the guest—Monday first). The
off-diagonal values that were used as a contrast repre-
sented the similarity between a mismatching guest and
conversation. To keep the number of contrast values sim-
ilar to the host analysis, described above, we focused on
the values representing mismatch between guest and con-
versation, coming from the same show. For instance, the
mismatch values for Michelle Obama who was a guest on
Monday first week were conversations where she was not
the guest, but still happened on Mondays and had the same
host (e.g., Monday second week and Monday third week).
To examine guest-specific reinstatement, the mean match-
ing guest to conversation similarity to the mean of the mis-
matching guest to conversation similarity was computed.
Simulations of Host and Guest RSAs
The RSAs for the hosts involve repeatedly correlating the
pattern of activity from Session 1 for each of the hosts with
three different conversations from Session 2. By contrast,
the RSAs for the guests involve unique pairwise correla-
tions between the pattern of activity for each guest and
their respective conversation. We wanted to check that
this procedure did not bias the analyses to find greater evi-
dence for reinstatement of either the hosts or the guests.
We therefore generated simulated patterns for the hosts
and guests and modeled the situation where both the host
and guest patterns were equally present in the pattern for
each conversation.
We first simulated 12 random patterns of the same
length as our PMC ROI. These patterns acted as the 12
celebrity patterns from Session 1. Three patterns were
taken to represent the hosts, and the other nine were
taken to represent the guests. We then simulated nine
conversation patterns that were linear combinations of
the host and guest patterns as well as noise (the correla-
tion between each conversation pattern and each of its
constituent “host” and “guest” pattern was assigned to
be 0.2). In the first simulation, we added white noise
to the simulated conversations patterns. This allowed us
to control the similarity between the simulated identity
patterns and the simulated conversation patterns. To
match our design, three of the simulated conversation pat-
terns were associated with the same host (but a unique
guest). We then ran our planned “host” and “guest” RSA
comparisons on the simulated data (as illustrated in
Figure 4). We ran these simulations 100,000 times each.
Next, we ran a further simulation using fMRI data to rule
out the possibility that correlated noise in the data would
bias the analyses to find greater evidence of reinstatement
for either the hosts or the guests. For each subject’s fMRI
data, we averaged the conversation patterns across the dif-
ferent runs of Session 2 and constructed a correlation
matrix between the different conversation patterns. We
Bromis et al.
523
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
then Fisher-transformed these correlation matrixes and
created an average correlation matrix across subjects that
represented the similarity between the nine conversations
participants were remembering during Session 2. We used
this average correlation matrix across conversations to add
correlated noise to the simulated conversations. Specifi-
cally, here, we modeled the conversations as combinations
of simulated host and guest patterns. Additionally, we
added simulated noise patterns that were drawn from a
multivariate Gaussian distribution with mean zero and cor-
relation matrix being equal to the empirical average corre-
lation matrix across conversations. We ran each simulation
100,000 times.
Hosts versus guests. To examine which regions show dif-
ferential reinstatement for host and guest identities during
retrieval of conversations, we performed a paired t test.
Specifically, we contrasted for each subject their host-
specific reinstatement effect with their guest-specific rein-
statement effect.
Time period analyses. We additionally ran post hoc
analyses to examine the time course of the host and guest
RSA effects. This examined whether any effects observed
in the main analyses evolve over time, and in particular,
whether the host effects are stronger during the initial
period of recall. To examine the time course of the host
and guest analyses, we ran three additional GLMs. We
modeled the first, middle, and last third of each of the con-
versations in separate GLMs. This allowed us to estimate
conversation patterns of brain activity that were specific
for the beginning, middle, and end of the 15-sec recall
period. We ran the host and guest analyses as described
previously, but separately for each of the three patterns.
We also ran the conversation-specific analysis (see
Figure 4B) that tested whether we could observe reliable
conversation-specific patterns.
Additional Analyses
Online (https://osf.io/zpcv3/), we report two additional
fMRI analyses. In the first, we ran exploratory RSAs to
investigate reinstatement of the hosts and guests during
the conversations within a whole-brain parcellation (see
Supplementary Figures 6 and 7 at https://osf.io/zpcv3/).
The main purpose of this analysis was to establish whether
any regions outside our predefined ROIs showed evidence
for reinstatement of the guests during recall of the conver-
sations. We therefore report results for the four main anal-
yses (see Figure 4) from 200 functionally defined regions
using a parcellation reported in Schaefer et al. (2018). We
also ran the four main analyses in a bilateral hippocampal
ROI (constructed using parcellations from Ritchey,
Montchal, Yonelinas, & Ranganath, 2015, and deposited
on https://neurovault.org/). Second, we used intersubject
pattern analysis to investigate whether the topics of the
conversations were represented in patterns of activity,
regardless of the identities of the hosts and guests. This
analysis addresses the issue of how narrative information
is represented in the brain (see Supplementary Figure 9
at https://osf.io/zpcv3/).
RESULTS
Behavioral Data
fMRI Study
Before taking part in the experiment, participants were
asked to rate how familiar they are with the 12 celebrities
and how easily they could imagine them (rating data were
missing from three participants). Participants, on average,
were familiar with the celebrities included in the experi-
ment (mean = 4.07, SD = 1.02, max = 5). One participant
initially reported relatively low familiarity with the celebri-
ties (mean = 2.66) and was asked to learn about each of
the celebrities by watching videos of them over the course
of a week.
In the first scanning session, participants were pre-
sented with an additional image block per run that
included a single picture of an unfamiliar person embed-
ded in a series of pictures of a famous celebrity. Partici-
pants, on average, identified 38% (SD = 19%) of these
oddball blocks. A coding error prevented us from record-
ing responses to the oddball blocks that occurred after
a block has finished, so this may be an underestimate of
the true percentage of oddball blocks detected. Between
picture and text blocks, participants performed an odd–
even number judgment task that served as an active base-
line and attention check. On average, participants made
the odd–even judgments accurately at 97.6% (SD = 0.02,
RT = 790 msec, SD = 101). Highly similar performance on
the odd–even judgment baseline task was also found
during the second scanning session (97%, SD = 0.03,
RT = 768 msec, SD = 105).
The memory screening test before the second scanning
session revealed that all of the participants had learnt the
nine conversations well. All participants correctly identi-
fied both the host and the guest for all conversations
(one participant failed to identify the guest for three of
the conversations and their data were excluded from the
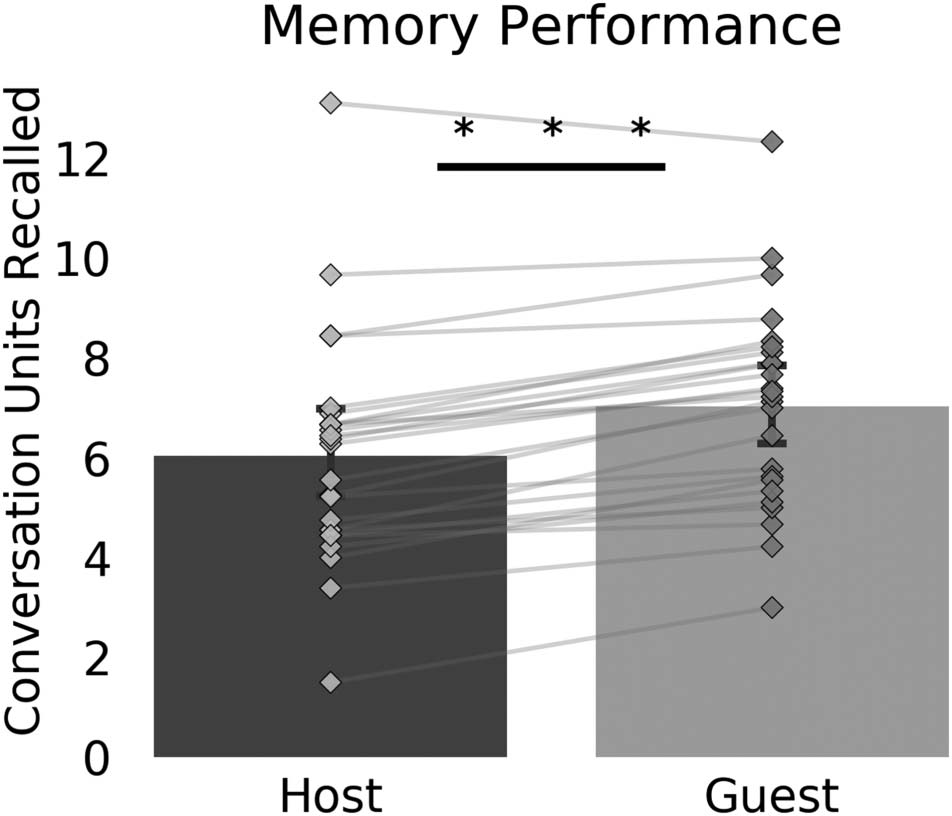
study). Participants identified 6.04 (SD = 2.29) idea units
spoken by the hosts and 7.03 (SD = 2.04) idea units spo-
ken by the guests, a difference that is highly significant,
t(24) = −8.18, p < .001 (see Figure 2). Thus, even though
the conversations were constructed so that the hosts and
guests spoke the same amount and both asked and
responded to questions, participants recalled more dia-
logue that was spoken by the guests than by the hosts.
Immediately before scanning, participants rated that
they could vividly and confidently remember the conver-
sations. Furthermore, they rated them as being familiar
and engaging (data available at https://osf.io/zpcv3/).
During the second scanning session, participants could
indicate if they did not recall a conversation in a specific
trial. Participants rarely reported that they could not recall
524
Journal of Cognitive Neuroscience
Volume 34, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
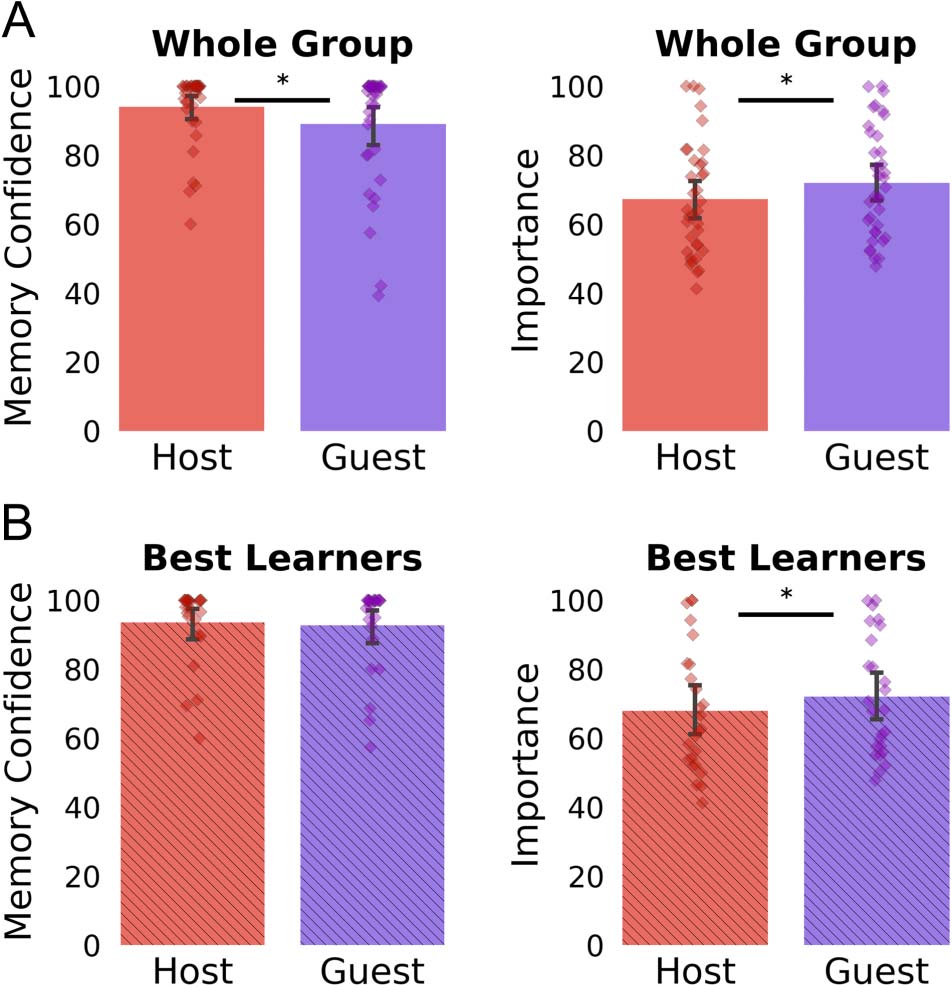
sample, we did not observe any significant differences in
host versus guest confidence (93.55 ± 11.15 vs. 92.69 ±
12.42), t(416) = 0.78, p = .43, but the guests were still per-
ceived to be more important than the hosts within each
conversation (72.03 ± 22.30 vs. 67.96 ± 23.38), t(416) =
−2.65, p = .008 (see Figure 3B). Last, when asked to guess
who the host and guests would be on a following week,
participants guessed 72.9% of the times that a following
week will involve the same hosts that were repeated in
the previous weekdays. For the guests, participants
responded that they did not know who the guest would
be for 73.83% of the future conversations and 25.2%
guessed that the guest on Week 4 would be a celebrity that
was not one of the previously seen celebrities. Only on 1%
of trials did one participant guess that a previously seen
guest would reappear on Week 4. Therefore, according
to this measure, the hosts were more “predictable” than
the guests.
Figure 2. Objective memory was higher for information said by the
guests in the fMRI study. Per subject average conversation units recalled
separately for hosts and guests were shown. On average, participants
remembered more of what the guest spoke.
fMRI Data
RSAs
the specific conversation on a given trial. On average,
participants had 1.71 (SD = 1.35) discarded trials out of
54, or 3.16%.
Online Study
A separate group of participants learned the same conver-
sations online and provided behavioral ratings about their
memory. Before completing the learning, sessions partic-
ipants were screened to be familiar with each of the 12
celebrities. Participants completed five learning sessions.
After the fifth session, participants answered multiple-
choice questions about the conversations and provided
subjective ratings about their memory.
Participants showed overall good learning of the conver-
sations after five sessions, with average accuracy being
95.38% (±5.72). Furthermore, participants had achieved
ceiling performance for the host (100% ± 0%) and near
ceiling performance for the guest (94.7% ± 9%) memory
questions. Participants rated that they could confidently
(75.75 ± 19.47) and vividly (75.03 ± 21.42) remember
the conversations. When examining all participants, we
observed that participants were more confident in their
memory for the hosts (94.01 ± 10.41) compared with
the guests (89.06 ± 16.92), t(603) = 4.33, p < .001. Inter-
estingly, participants rated the guests as playing a more
important role than the hosts in the conversations
(71.96 ± 21.77 vs. 67.24 ± 21.84), t(603) = −3.68, p <
.001 (see Figure 3A). To further investigate these effects
and to provide a better comparison to the participants
who took part in the main scanning experiment (who
identified both the host and the guest of every conversa-
tion), we analyzed data from participants who were able to
identify all of the hosts and the guests (25 in total). In this
Identity. We first examined which regions would show
reliable identity-specific patterns in Session 1 when the
celebrities were presented in isolation as both pictures
and names. See https://osf.io/zpcv3/ for results from all
ROIs. Reliable identity-specific decoding was observed
Figure 3. Similar average confidence and importance ratings for host
and guest in online study. (A) Memory confidence was slightly higher in
the whole group that completed the online study. This included people
who did not learn guests to a ceiling-level performance. (B) No memory
confidence difference was seen for hosts and guests in participants who
learned all conversations, similarly to the participants included in the
fMRI study. Nonetheless participants still rated the guest as playing a
more important role in the conversation.
Bromis et al.
525
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Contrast matrices demonstrating the different RSAs. Red indicates positive and blue indicates negative contrast weight. (A) Analysis of
Session 1 to examine which regions show reliable identity-specific patterns. (B) We examined Session 2 data to demonstrate regions that would show
reliable conversation-specific patterns. (C) We correlated patterns from Session 1 to Session 2 to examine host-specific reinstatement during the
conversations. (D) Examined guest reinstatement in the conversations.
in the left LOC, t(26) = 3.52, p = .001; PMC, t(26) = 2.47,
p = .02; right LOC, t(26) = 2.54, p = .01; and right fusi-
form cortex, t(26) = 3.62, p = .001 (see Figure 5B). The
other regions, left parahippocampal cortex, t(26) =
−0.59, p = .55, and MPFC, t(26) = 0.88, p = .38, did
not show significant identity-specific patterns. Therefore,
the regions taken forward to the next analysis were the
PMC, the left and right LOC, and the right fusiform cortex.
Conversations. We next examined which of the four
regions taken forward from the previous analyses would
show reliable conversation-specific patterns. See https://
osf.io/zpcv3/ for results from all four ROIs. The left LOC,
t(26) = 6.10, p < .001; PMC, t(26) = 4.04, p < .001; and
right LOC, t(26) = 4.06, p < .001, showed reliable
conversation-specific patterns (see Figure 5B). However,
the right fusiform cortex ROI did not show significantly
higher reliability for matching conversations, t(26) =
−0.28, p = .78. Therefore, the regions that were taken for-
ward for further analysis were the PMC and the left and
right LOC.
Simulations of host and guest RSAs. We ran simulations
to ensure that our analyses were not statistically biased to
find stronger effects for the host identities because the
hosts were repeated across three conversations. We simu-
lated conversation patterns that had an equal contribution
from both simulated host and simulated guest patterns.
The first simulation added white noise to the simulated
conversation patterns and ran the same analyses for the
hosts and guests as the ones ran in the main analyses.
The second simulation added noise that had the same cor-
relational structure as the conversation patterns in the fMRI
data (see Supplementary Figure 4 at https://osf.io/zpcv3/).
Importantly, we did not see any differences in the simu-
lated effects for host and guest analysis (simulation with
correlated noise: host mean = 0.202, SD = 0.005; guest
mean = 0.202, SD = 0.005), t(99999) = −0.58, p = .56.
Hosts. We next examined which ROIs would show host-
specific reinstatement. We compared the similarity
between brain patterns from the Session 1 to Session 2
conversations. We took the patterns of identities in
526
Journal of Cognitive Neuroscience
Volume 34, Number 3
Session 1 that would be hosts in Session 2 and computed
the similarity between host identities with the matching
conversations. The left LOC, t(26) = 2.34, p = .02; PMC,
t(26) = 3.27, p = .003; and right LOC, t(26) = 2.02, p = .05,
showed host-specific reinstatement during the conversa-
tions (see Figure 5B).
Guests. We next examined whether these regions would
also show guest-specific reinstatement. None of the ROIs
showed a reliable guest-specific reinstatement: left LOC:
t(26) = −1.56, p = .12; PMC: t(26) = −1.61, p = .12; right
LOC: t(26) = −1.62, p = .11. Surprisingly, these nonsignif-
icant effects are in the opposite direction to that which
would show evidence for reinstatement (see Figure 5B).
Hosts versus guests. We then performed a direct contrast
between the host-specific reinstatement and guest-specific
reinstatement effects. We observed higher host reinstate-
ment in the left LOC, t(26) = 2.90, p = .007; PMC, t(26) =
3.41, p = .002; and right LOC, t(26) = 2.46, p = .02.
In a post hoc analysis, we exam-
Time course analysis.
ined the time course of the host and guest effects reported
above. Specifically, we ran three separate GLMs to esti-
mate the first, second, and third part of the conversations
separately. This allowed us to estimate patterns for the
conversations for the beginning, middle, and end of the
recall period. We ran the host and guest analyses as
described above separately for each part of the recall
period. We further ran the conversation-specific analysis
in the three parts separately to ensure that we had enough
power to distinguish between the conversations when
modeling only a third of the duration (see Figure 6).
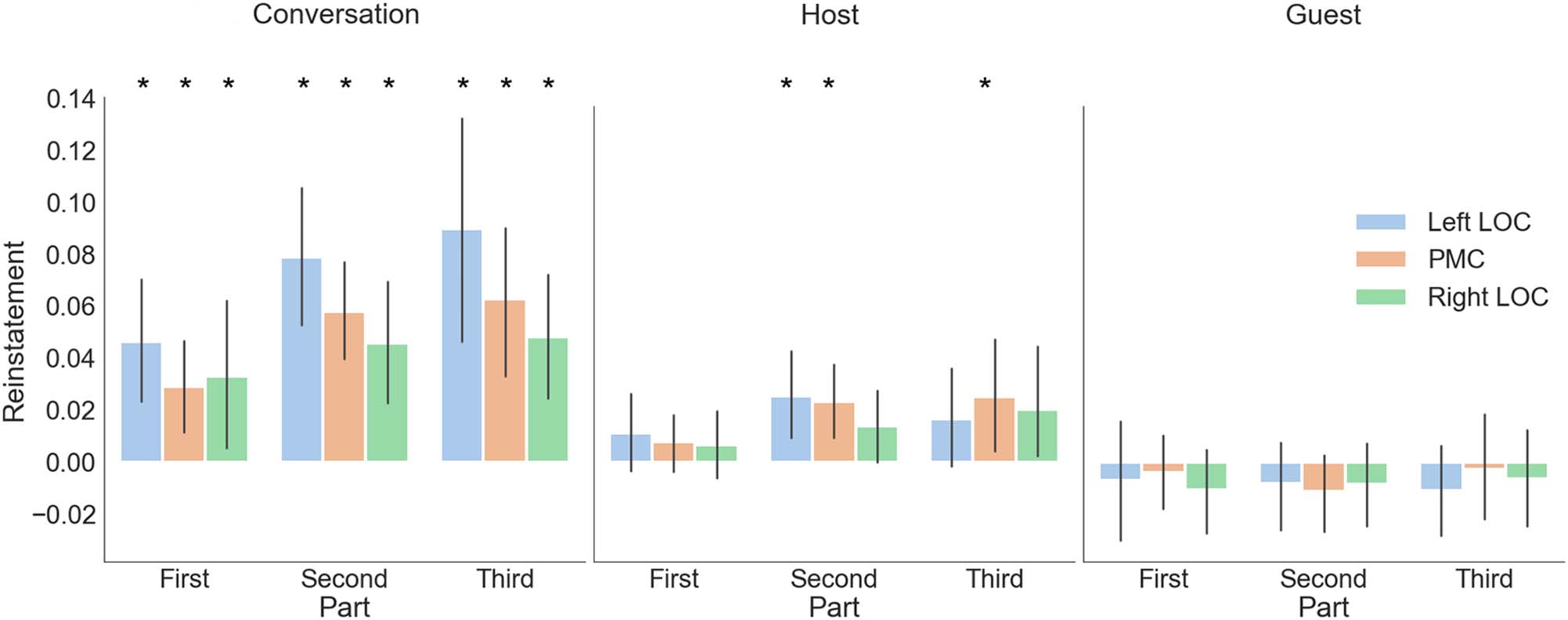
Throughout the beginning, middle, and end of the events
Figure 5. RSA comparisons within ROIs. (A) ROIs taken from Robin
et al. (2018): These ROIs were identified to equally represent multiple
features (location, people, and object) of events. The ROIs included
PMC, LOC, MPFC, and parahippocampal cortex (PHC). (B) RSA results
of ROIs showing reliable identity and conversation patterns, as well as
reliable host reinstatement effects. The x-axis represents the different
analyses (see Figure 3). The y-axis represents the correlation contrast
between the diagonal and off-diagonal values. The asterisk indicates
significance at p < .05. Each diamond represents the correlation
contrast value for a given subject in one of the three ROIs.
Figure 6. Reinstatement effects for host across time. Here, we show results for the conversation-specific host and guest analysis separately for the
first, second, and third part of the recall duration. Bar plot represents means, and error bar shows bootstrapped 95% confidence intervals. Asterisk
indicates significance at p < .05.
Bromis et al.
527
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
we observed conversation specific patterns. We observed
host reinstatement in the middle and end of recall periods,
but not during the beginning of events. In contrast, we
did not observe guest reinstatement during any point
of the recall period. We found that when we modeled
only the beginning of the conversations, we could reli-
ably detect conversation-specific patterns in all ROIs (left
LOC: t(26) = 3.98, p < .001; PMC: t(26) = 3.14, p = .004;
right LOC: t(26) = 2.25, p = .03), but could not observe
any reinstatement of the hosts or guests in any of the three
ROIs (all ps > .05). When modeling the middle part of the
recall, we again found conversation-specific patterns in all
ROIs: left LOC: t(26) = 5.47, p < .001; PMC: t(26) = 6.06,
p < .001; right ROC: t(26) = 3.86, p < .001. We also found
host reinstatement (left LOC: t(26) = 2.71, p = .01; PMC:
t(26) = 3.21, p = .003; right LOC: t(26) = 1.98, p = .058),
but not guest reinstatement (all ps > .05). When modeling
only the last 30% of the recall, we again found conversation-
specific patterns (left LOC: t(26) = 4.28, p < .001; PMC:
t(26) = 4.04, p < .001; right LOC: t(26) = 4.06, p < .001),
host reinstatement in PMC (left LOC: t(26) = 1.73, p = .09;
PMC: t(26) = 2.23, p = .03; right LOC: t(26) = 1.87, p =
.07), but no significant guest reinstatement (all ps > .05).
DISCUSSION
Rich and detailed episodic memories comprise many
individual elements: Some of these elements are common
to several different memories, whereas others are unique to
one. Using a novel paradigm involving recall of com-
plex, naturalistic events (custom written “chat show” con-
versations), we found that the neural representation of
the events was dominated by the repeated and more pre-
dictable elements (the “hosts”) rather than the more idio-
syncratic, yet event-defining elements (“the guests”). These
effects were found in PMC as well as lateral parietal/occipital
cortex. Importantly, analyses of behavioral data showed that
participants actually had better memory for dialogue spo-
ken by the guests compared with the hosts and also rated
the guests as being more important to the conversations.
This shows that the fMRI effects we report are not driven
by better memory for the hosts compared with the guests.
Many studies have shown that episodic memories tend to
be recalled in a holistic manner, with the retrieval of one ele-
ment being dependent on retrieval of others ( Joensen,
Gaskell, & Horner, 2020; Ngo, Horner, Newcombe, &
Olson, 2019; Horner & Burgess, 2014). Furthermore,
retrieval of one element tends to result in reactivation of
the representation of other elements (Horner, Bisby, Bush,
Lin, & Burgess, 2015), suggesting that no particular element
dominates in the overall representation of the event. How-
ever, in these studies, the events are all unique combina-
tions of elements. Our results suggest that when we recall
memories of particular events, it is the repeated and pre-
dictable elements that serve to frame or scaffold the rep-
resentation of the memory. It has already been shown that
the spatial context of an episode can dominate the
representation of episodic memories (Robin et al., 2018),
but in the current study, we specifically chose nonspatial ele-
ments (well-known people) to be the repeated or novel ele-
ments. We therefore speculate that when we recall detailed
episodic memories, those elements that are reliably present
serve as a form of “context” within which to retrieve the less
predictable details that are specific to a particular occasion.
Our study focused on regions of the brain’s PM network,
and we used ROIs that were identified in a previous study of
multielement event recall (Robin et al., 2018). The regions
where we found that hosts were represented more strongly
than guests—the PMC and lateral occipital/parietal
cortex—have been shown to support event-specific pat-
terns of activity in many previous studies (Masís-Obando,
Norman, & Baldassano, 2021; Raykov et al., 2021; Reagh &
Ranganath, 2021; Bird, 2020; Chen et al., 2017; Oedekoven
et al., 2017; Bird et al., 2015; St-Laurent et al., 2015; Kuhl &
Chun, 2014). This brain network has been argued to pre-
dominantly represent contextual information in a broad
sense—not only spatial contexts but also temporal and
social contexts (Ranganath & Ritchey, 2012). Similarly, it
is associated with integrating multimodal information over
long timescales to build mental models of an overarching
situation ( Yeshurun, Nguyen, & Hasson, 2021; Hasson,
Chen, & Honey, 2015). Nevertheless, there is a large body
of evidence that these regions also support representa-
tions of known individuals and more basic semantic con-
cepts (Fairhall & Caramazza, 2013). Here, we show that
although these regions do support representations of
famous individuals, when two individuals occur together
in an event, it is the person who occurs most reliably
within that context who is represented most robustly.
The aim of this study was to contrast those elements that
are more generic, familiar, and reliably present with those
that are more idiosyncratic, unexpected, and event-
defining. Our behavioral results revealed that it was actually
the parts of the conversations spoken by the guests that
were more memorable. Moreover, a follow-up study found
that participants judged the guests to be more important to
the conversations compared with the hosts. Given this and
the research summarized in the Introduction, it was
unclear which elements would dominate the neural repre-
sentation of the conversations. We, therefore, feel that the
finding that the host representations were consistently
stronger than the guest representations is an important
step to understanding how recalled episodic memories
are represented by the brain. Our design does not allow
us to further tease apart which factors contribute to the
representations of the hosts being dominant. For example,
the number of repetitions of the hosts across the conversa-
tions is confounded with their predictability. Moreover, it is
possible that chat show “hosts” are associated with a qual-
itatively different structural or social representation.
Future work should aim to deconfound the effects of
number of repetitions and predictability as well as verify
that our results generalize to elements of an episode that
do not have strong preexisting contextual associations.
528
Journal of Cognitive Neuroscience
Volume 34, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Episodic memory recall is thought to comprise an initial
period of memory search and construction followed by a
period of memory elaboration (Addis, Wong, & Schacter,
2007; Conway, Pleydell-Pearce, & Whitecross, 2001). It is
possible that, in our study, the initial search period of epi-
sodic recall was dominated by the representations of the
hosts. This is because the hosts were reliably associated
with a particular day of the week across three separate con-
versations (e.g., all Fridays), whereas the guests were only
associated with one unique combination of day and week
(e.g., Monday Week 3). However, we found no evidence
that the representation of the hosts was preferentially
active at the beginning of the recall period or that the
activation of the representation of the guest became
stronger as the recall phase progressed. Furthermore, we
observed conversation-specific patterns of activity in all
ROIs included in the main analyses. Indeed, we also
observed conversation-specific patterns shared across
participants who learned the conversations with different
host and guests (see https://osf.io/zpcv3/). Taken together,
these results and our behavioral memory findings suggests
that participants were unlikely to be remembering only the
hosts during the whole retrieval period.
The guests in the conversations were all well-known
celebrities and key elements in each of the events. Never-
theless, the representational similarity between the guest-
related patterns of activity in Session 1 and recall of the
conversations in Session 2 was not reliably above chance
in any of the ROIs. Furthermore, an exploratory analysis
of 200 cortical regions and the hippocampus did not reveal
any locations where guest-related patterns of activity were
reliably activated when recalling the conversations (see
https://osf.io/zpcv3/). By modeling our analyses, we verified
that they were not biased toward detecting reactivation
effects in the hosts rather than the guests. It remains a pos-
sibility that brain regions supporting representations of the
guests during retrieval did not correspond to shape of the
parcellated brain regions used in the exploratory analysis.
It has been argued that a function of episodic memory is to
enable the prediction of future events (Barron et al., 2020;
Lu, Hasson, & Norman, 2020; Schacter, Benoit, & Szpunar,
2017; see also Sun, Advani, Spruston, Saxe, & Fitzgerald,
2021). Predictive processing accounts of perception and
cognition typically emphasize the role of unexpected infor-
mation in driving attention and new learning (Niv &
Schoenbaum, 2008; Friston, 2005). Since memory offers
an opportunity for “off-line” learning (Hinton et al., 1995),
we might expect that the less typical aspects of an event
would be overrepresented when the event is recalled.
However, we found the opposite effect—the repeated
and more predictable elements of the event—dominated
its neural representation.
Instead, our results are compatible with the view that
information that has been encountered more frequently
is likely to be most useful to us in the future (Anderson
& Milson, 1989). When recalling an event, activating robust
representations of the elements that are most reliably
present may help us to generalize our experiences to
new situations (Gershman, 2017). To reconcile our find-
ings with predictive coding accounts of cognition, it may
be that, during perception, prediction errors drive the
learning of any unexpected information, whereas memory
recall offers the opportunity to improve our internal
model of the world by selectively enhancing those ele-
ments that are likely to be encountered again and remov-
ing noisy or idiosyncratic components that are unlikely to
be repeated (Barron et al., 2020; see also Sun et al., 2021).
In summary, our results show that the elements of an
event are not represented equally in memory. Those
elements that are most reliably present across similar
events—being both more frequently encountered and
more predictable—are the ones that are most robustly acti-
vated when recalling these events. The brain may use these
elements to provide context and scaffold the representa-
tion of a remembered event within the PM network. This,
in turn, may facilitate the activation of the more idiosyn-
cratic elements that are unique to one specific occasion.
Acknowledgments
We thank Ediz Sohoglu and Sam Berens for helpful discussions
about the study. We would like to thank Jessica Robin for pro-
viding us with the multifeature ROI and Charlotte Sutherland
for helping to score the behavioral data.
Reprint requests should be sent to Petar P. Raykov, School of
Psychology, University of Sussex, Falmer BN1 9QH, UK, or via
e-mail: P.Raykov@sussex.ac.uk.
Author Contributions
Petar P. Raykov: Conceptualization; Formal analysis; Method-
ology; Project administration; Visualization; Writing—
Original draft; Writing—Review & editing. Konstantinos
Bromis: Conceptualization; Formal analysis; Methodol-
ogy; Project administration. Leah Wickens: Data curation;
Project administration. Warrick Roseboom: Conceptuali-
zation. Chris M. Bird: Conceptualization; Funding acquisi-
tion; Methodology; Project administration; Supervision;
Writing—Original draft; Writing—Review & editing.
Funding Information
This project has received funding from the European
Research Council (ERC) under the European Union’s
Horizon 2020 research and innovation programme
(https://dx.doi.org/10.13039/100010663), grant number:
819526 to C. M. B. P. P. R. was additionally supported by
an Economic and Social Research Council studentship
(https://dx.doi.org/10.13039/501100000269), grant num-
ber: ES/J500173/1 and fellowship: ES/ V012444/1.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions
Bromis et al.
529
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
of authorship teams (categorized by estimated gender
identification of first author/last author) publishing in
the Journal of Cognitive Neuroscience ( JoCN ) during
this period were M(an)/M = .407, W(oman)/M = .32,
M/W = .115, and W/W = .159, the comparable propor-
tions for the articles that these authorship teams cited
were M/M = .549, W/M = .257, M/W = .109, and W/W =
.085 (Postle and Fulvio, JoCN, 34:1, pp. 1–3). Conse-
quently, JoCN encourages all authors to consider gender
balance explicitly when selecting which articles to cite and
gives them the opportunity to report their article’s gender
citation balance.
REFERENCES
Addis, D. R., Wong, A. T., & Schacter, D. L. (2007). Remembering
the past and imagining the future: common and distinct
neural substrates during event construction and elaboration.
Neuropsychologia, 45, 1363–1377. https://doi.org/10.1016/j
.neuropsychologia.2006.10.016, PubMed: 17126370
Aitchison, L., & Lengyel, M. (2017). With or without you:
Predictive coding and Bayesian inference in the brain.
Current Opinion in Neurobiology, 46, 219–227. https://doi
.org/10.1016/j.conb.2017.08.010, PubMed: 28942084
Anderson, J. R., & Milson, R. (1989). Human memory: An
adaptive perspective. Psychological Review, 96, 703–719.
https://doi.org/10.1037/0033-295X.96.4.703
Barron, H. C., Auksztulewicz, R., & Friston, K. (2020). Prediction
and memory: A predictive coding account. Progress in
Neurobiology, 192, 101821. https://doi.org/10.1016/j
.pneurobio.2020.101821, PubMed: 32446883
Bird, C. M. (2020). How do we remember events? Current
Opinion in Behavioral Sciences, 32, 120–125. https://doi.org
/10.1016/j.cobeha.2020.01.020
Bird, C. M., Keidel, J. L., Ing, X. L. P., Horner, X. A. J., & Burgess,
N. (2015). Consolidation of complex events via reinstatement
in posterior cingulate cortex. Journal of Neuroscience, 35,
14426–14434. https://doi.org/10.1523/JNEUROSCI.1774-15
.2015, PubMed: 26511235
Brewer, W. F., & Treyens, J. C. (1981). Role of schemata in
memory for places. Cognitive Psychology, 13, 207–230.
https://doi.org/10.1016/0010-0285(81)90008-6
Chen, J., Leong, Y. C., Honey, C. J., Yong, C. H., Norman, K. A., &
Hasson, U. (2017). Shared memories reveal shared structure in
neural activity across individuals. Nature Neuroscience, 20,
115–125. https://doi.org/10.1038/nn.4450, PubMed: 27918531
Clark, A. (2013). Whatever next? Predictive brains, situated
agents, and the future of cognitive science. Behavioral
and Brain Sciences, 36, 181–204. https://doi.org/10.1017
/S0140525X12000477, PubMed: 23663408
Conway, M. A., Pleydell-Pearce, C. W., & Whitecross, S. E.
(2001). The neuroanatomy of autobiographical memory: A
slow cortical potential study of autobiographical memory
retrieval. Journal of Memory and Language, 45, 493–524.
https://doi.org/10.1006/jmla.2001.2781
Fairhall, S. L., & Caramazza, A. (2013). Brain regions that
represent amodal conceptual knowledge. Journal of
Neuroscience, 33, 10552–10558. https://doi.org/10.1523
/JNEUROSCI.0051-13.2013, PubMed: 23785167
Friston, K. (2005). A theory of cortical responses. Philosophical
Transactions of the Royal Society of London, Series B:
Biological Sciences, 360, 815–836. https://doi.org/10.1098
/rstb.2005.1622, PubMed: 15937014
Gardumi, A., Ivanov, D., Hausfeld, L., Valente, G., Formisano, E.,
& Uludağ, K. (2016). The effect of spatial resolution on
decoding accuracy in fMRI multivariate pattern analysis.
Neuroimage, 132, 32–42. https://doi.org/10.1016/j
.neuroimage.2016.02.033, PubMed: 26899782
Gershman, S. J. (2017). Predicting the past, remembering
the future. Current Opinion in Behavioral Sciences, 17,
7–13. https://doi.org/10.1016/j.cobeha.2017.05.025,
PubMed: 28920071
Glenberg, A., Smith, S. M., & Green, C. (1977). Type I rehersal:
Maintenance and more. Journal of Memory and Language,
16, 339–352. https://doi.org/10.1016/S0022-5371(77)80055-8
Hasson, U., Chen, J., & Honey, C. J. (2015). Hierarchical process
memory: Memory as an integral component of information
processing. Trends in Cognitive Sciences, 19, 304–313.
https://doi.org/10.1016/j.tics.2015.04.006, PubMed: 25980649
Hendriks, M. H. A., Daniels, N., Pegado, F., & Op de Beeck, H. P.
(2017). The effect of spatial smoothing on representational
similarity in a simple motor paradigm. Frontiers in Neurology,
8, 222. https://doi.org/10.3389/fneur.2017.00222, PubMed:
28611726
Hinton, G. E., Dayan, P., Frey, B. J., & Neal, R. M. (1995). The
“wake–sleep” algorithm for unsupervised neural networks.
Science, 268, 1158–1161. https://doi.org/10.1126/science
.7761831, PubMed: 7761831
Horner, A. J., Bisby, J. A., Bush, D., Lin, W.-J., & Burgess, N. (2015).
Evidence for holistic episodic recollection via hippocampal
pattern completion. Nature Communications, 6, 1–11.
https://doi.org/10.1038/ncomms8462, PubMed: 26136141
Horner, A. J., & Burgess, N. (2014). Pattern completion in
multielement event engrams. Current Biology, 24, 988–992.
https://doi.org/10.1016/j.cub.2014.03.012, PubMed: 24746796
Hunt, R. R. (1995). The subtlety of distinctiveness: What von
Restorff really did. Psychonomic Bulletin & Review, 2, 105–112.
https://doi.org/10.3758/BF03214414, PubMed: 24203592
Hutton, C., Bork, A., Josephs, O., Deichmann, R., Ashburner, J.,
& Turner, R. (2002). Image distortion correction in fMRI: A
quantitative evaluation. Neuroimage, 16, 217–240. https://doi
.org/10.1006/nimg.2001.1054, PubMed: 11969330
Joensen, B. H., Gaskell, M. G., & Horner, A. J. (2020). United we
fall: All-or-none forgetting of complex episodic events.
Journal of Experimental Psychology: General, 149, 230.
https://doi.org/10.1037/xge0000648, PubMed: 31305093
Kishiyama, M. M., & Yonelinas, A. P. (2003). Novelty effects on
recollection and familiarity in recognition memory. Memory
& Cognition, 31, 1045–1051. https://doi.org/10.3758
/BF03196125, PubMed: 14704019
Kuhl, B. A., & Chun, M. M. (2014). Successful remembering
elicits event-specific activity patterns in lateral parietal cortex.
Journal of Neuroscience, 34, 8051–8060. https://doi.org/10
.1523/JNEUROSCI.4328-13.2014, PubMed: 24899726
Cooper, R. A., & Ritchey, M. (2020). Progression from feature-
Lee, H., & Kuhl, B. A. (2016). Reconstructing perceived and
specific brain activity to hippocampal binding during episodic
encoding. Journal of Neuroscience, 40, 1701–1709. https://doi
.org/10.1523/JNEUROSCI.1971-19.2019, PubMed: 31826947
di Oleggio Castello, M. V., Halchenko, Y. O., Guntupalli, J. S., Gors,
J. D., & Gobbini, M. I. (2017). The neural representation of
personally familiar and unfamiliar faces in the distributed
system for face perception. Scientific Reports, 7, 12237. https://
doi.org/10.1038/s41598-017-12559-1, PubMed: 28947835
retrieved faces from activity patterns in lateral parietal cortex.
Journal of Neuroscience, 36, 6069–6082. https://doi.org/10
.1523/JNEUROSCI.4286-15.2016, PubMed: 27251627
Lu, Q., Hasson, U., & Norman, K. A. (2020). Learning to use
episodic memory for event prediction. BioRxiv.
Masís-Obando, R., Norman, K. A., & Baldassano, C. (2021).
Schema representations in distinct brain networks support
narrative memory during encoding and retrieval. BioRxiv.
530
Journal of Cognitive Neuroscience
Volume 34, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Morris, C. D., Bransford, J. D., & Franks, J. J. (1977). Levels of
processing versus transfer appropriate processing. Journal of
Verbal Learning and Verbal Behavior, 16, 519–533. https://
doi.org/10.1016/S0022-5371(77)80016-9
Mumford, J. A., Turner, B. O., Ashby, F. G., & Poldrack, R. A.
(2012). Deconvolving BOLD activation in event-related
designs for multivoxel pattern classification analyses.
Neuroimage, 59, 2636–2643. https://doi.org/10.1016/j
.neuroimage.2011.08.076, PubMed: 21924359
Ngo, C. T., Horner, A. J., Newcombe, N. S., & Olson, I. R.
(2019). Development of holistic episodic recollection.
Psychological Science, 30, 1696–1706. https://doi.org/10.1177
/0956797619879441, PubMed: 31672085
Niv, Y., & Schoenbaum, G. (2008). Dialogues on prediction
errors. Trends in Cognitive Sciences, 12, 265–272. https://doi
.org/10.1016/j.tics.2008.03.006, PubMed: 18567531
Nyberg, L., Habib, R., McIntosh, A. R., & Tulving, E. (2000).
Reactivation of encoding-related brain activity during
memory retrieval. Proceedings of the National Academy of
Sciences, U.S.A., 97, 11120–11124. https://doi.org/10.1073
/pnas.97.20.11120, PubMed: 11005878
Oedekoven, C. S. H., Keidel, J. L., Berens, S. C., & Bird, C. M. (2017).
Reinstatement of memory representations for lifelike events
over the course of a week. Scientific Reports, 7, 14305. https://
doi.org/10.1038/s41598-017-13938-4, PubMed: 29084981
Oosterhof, N. N., Connolly, A. C., & Haxby, J. V. (2016).
CoSMoMVPA: Multi-modal multivariate pattern analysis of
neuroimaging data in Matlab/GNU octave. Frontiers in
Neuroinformatics, 10, 27. https://doi.org/10.3389/fninf.2016
.00027, PubMed: 27499741
Popov, V., & Reder, L. M. (2020). Frequency effects on memory:
A resource-limited theory. Psychological Review, 127, 1–46.
https://doi.org/10.1037/rev0000161, PubMed: 31524424
Poppenk, J., Köhler, S., & Moscovitch, M. (2010). Revisiting the
novelty effect: When familiarity, not novelty, enhances
memory. Journal of Experimental Psychology: Learning,
Memory, and Cognition, 36, 1321–1330. https://doi.org/10
.1037/a0019900, PubMed: 20804299
Quent, J. A., Henson, R. N., & Greve, A. (2021). A predictive
account of how novelty influences declarative memory.
Neurobiology of Learning and Memory, 179, 107382. https://
doi.org/10.1016/j.nlm.2021.107382, PubMed: 33476747
Ranganath, C., & Rainer, G. (2003). Neural mechanisms for
detecting and remembering novel events. Nature Reviews
Neuroscience, 4, 193–202. https://doi.org/10.1038/nrn1052,
PubMed: 12612632
Ranganath, C., & Ritchey, M. (2012). Two cortical systems for
memory-guided behaviour. Nature Reviews Neuroscience, 13,
713–726. https://doi.org/10.1038/nrn3338, PubMed: 22992647
Raykov, P. P., Keidel, J. L., Oakhill, J., & Bird, C. M. (2020). The
brain regions supporting schema-related processing of people’s
identities. Cognitive Neuropsychology, 37, 8–24. https://doi.org
/10.1080/02643294.2019.1685958, PubMed: 31710265
Raykov, P. P., Keidel, J. L., Oakhill, J., & Bird, C. M. (2021).
Activation of person knowledge in medial prefrontal cortex
during the encoding of new lifelike events. Cerebral Cortex.
https://doi.org/10.1093/cercor/bhab027, PubMed: 33866362
Reagh, Z. M., & Ranganath, C. (2021). A cortico-hippocampal
scaffold for representing and recalling lifelike events. BioRxiv.
Ritchey, M., Montchal, M. E., Yonelinas, A. P., & Ranganath, C.
(2015). Delay-dependent contributions of medial temporal
lobe regions to episodic memory retrieval. eLife, 4, e05025.
https://doi.org/10.7554/eLife.05025, PubMed: 25584461
Robin, J. (2018). Spatial scaffold effects in event memory and
imagination. Wiley Interdisciplinary Reviews: Cognitive Science,
9, e1462. https://doi.org/10.1002/wcs.1462, PubMed: 29485243
Robin, J., Buchsbaum, B. R., & Moscovitch, M. (2018). The
primacy of spatial context in the neural representation of
events. Journal of Neuroscience, 38, 2755–2765. https://doi
.org/10.1523/JNEUROSCI.1638-17.2018, PubMed: 29440386
Schacter, D. L., Benoit, R. G., & Szpunar, K. K. (2017). Episodic
future thinking: Mechanisms and functions. Current Opinion
in Behavioral Sciences, 17, 41–50. https://doi.org/10.1016/j
.cobeha.2017.06.002, PubMed: 29130061
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo,
X. N., Holmes, A. J., et al. (2018). Local-global parcellation of
the human cerebral cortex from intrinsic functional
connectivity MRI. Cerebral Cortex, 28, 3095–3114. https://
doi.org/10.1093/cercor/bhx179
Sherman, B. E., & Turk-Browne, N. B. (2020). Statistical
prediction of the future impairs episodic encoding of the
present. Proceedings of the National Academy of Sciences,
U.S.A., 117, 22760–22770. https://doi.org/10.1073/pnas
.2013291117, PubMed: 32859755
Sievers, C., Bird, C. M., & Renoult, L. (2019). Predicting memory
formation over multiple study episodes. Learning & Memory,
26, 465–472. https://doi.org/10.1101/lm.049791.119, PubMed:
31732707
Smith, T. A., Hasinski, A. E., & Sederberg, P. B. (2013). The
context repetition effect: Predicted events are remembered
better, even when they don’t happen. Journal of
Experimental Psychology: General, 142, 1298. https://doi.org
/10.1037/a0034067, PubMed: 23957285
Smith, S. M., Jenkinson, M., Woolrich, M. W., Beckmann, C. F.,
Behrens, T. E. J., Johansen-Berg, H., et al. (2004). Advances in
functional and structural MR image analysis and implementation
as FSL. Neuroimage, 23, S208–S219. https://doi.org/10.1016/j
.neuroimage.2004.07.051, PubMed: 15501092
Sohoglu, E., & Davis, M. H. (2020). Rapid computations of
spectrotemporal prediction error support perception of
degraded speech. eLife, 9, e58077. https://doi.org/10.7554
/eLife.58077, PubMed: 33147138
St-Laurent, M., Abdi, H., & Buchsbaum, B. R. (2015). Distributed
patterns of reactivation predict vividness of recollection.
Journal of Cognitive Neuroscience, 27, 2000–2018. https://
doi.org/10.1162/jocn_a_00839, PubMed: 26102224
Sun, W., Advani, M., Spruston, N., Saxe, A., & Fitzgerald, J. E. (2021).
Organizing memories for generalization in complementary
learning systems. BioRxiv.
Tulving, E. (1983). Elements of episodic memory. Oxford:
Oxford University Press.
Van Strien, J. W., Hagenbeek, R. E., Stam, C. J., Rombouts,
S. A. R. B., & Barkhof, F. (2005). Changes in brain electrical
activity during extended continuous word recognition.
Neuroimage, 26, 952–959. https://doi.org/10.1016/j
.neuroimage.2005.03.003, PubMed: 15955505
Wechsler, D. (1945). A standardized memory scale for clinical
use. Journal of Psychology, 19, 87–95. https://doi.org/10.1080
/00223980.1945.9917223
Wittkuhn, L., Chien, S., Hall-McMaster, S., & Schuck, N. W.
(2021). Replay in minds and machines. Neuroscience &
Biobehavioral Reviews, 129, 367–388. https://doi.org/10.1016
/j.neubiorev.2021.08.002
Yassa, M. A., & Reagh, Z. M. (2013). Competitive trace theory: A
role for the hippocampus in contextual interference during
retrieval. Frontiers in Behavioral Neuroscience, 7, 107.
https://doi.org/10.3389/fnbeh.2013.00107, PubMed: 23964216
Yeshurun, Y., Nguyen, M., & Hasson, U. (2021). The default mode
network: Where the idiosyncratic self meets the shared social
world. Nature Reviews Neuroscience, 22, 181–192. https://doi
.org/10.1038/s41583-020-00420-w, PubMed: 33483717
Zadbood, A., Chen, J., Leong, Y. C., Norman, K. A., & Hasson, U.
(2017). How we transmit memories to other brains:
Constructing shared neural representations via
communication. Cerebral Cortex, 27, 4988–5000. https://doi
.org/10.1093/cercor/bhx202, PubMed: 28922834
Bromis et al.
531
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
3
5
1
7
1
9
8
4
8
5
8
/
j
o
c
n
_
a
_
0
1
8
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3