The interaction between random and systematic visual stimulation and
infraslow quasi-periodic spatiotemporal patterns of whole-brain activity
Nan Xua, Derek M. Smithb,c, George Jenod, Dolly T. Seeburgerb, Eric H. Schumacherb, Shella D. Keilholza
aWallace H. Coulter Department of Biomedical Engineering, Georgia Institute of Technology and Emory University, Atlanta, GA, United States
bSchool of Psychology, Georgia Institute of Technology, Atlanta, GA, United States
cDepartment of Neurology, Division of Cognitive Neurology/Neuropsychology, The Johns Hopkins University School of Medicine, Baltimore, MD, United States
dSchool of Computer Science, Georgia Institute of Technology, Atlanta, GA, United States
Corresponding Author: Nan Xu (im.nan.xu@gmail.com)
ABSTRACT
One prominent feature of the infraslow blood oxygen level-dependent (BOLD) signal during rest or task is quasi-
periodic spatiotemporal pattern (QPP) of signal changes that involves an alternation of activity in key functional net-
works and propagation of activity across brain areas, and that is known to tie to the infraslow neural activity involved
in attention and arousal fluctuations. This ongoing whole-brain pattern of activity might potentially modify the response
to incoming stimuli or be modified itself by the induced neural activity. To investigate this, we presented checkerboard
sequences flashing at 6 Hz to subjects. This is a salient visual stimulus that is known to produce a strong response in
visual processing regions. Two different visual stimulation sequences were employed, a systematic stimulation
sequence in which the visual stimulus appeared every 20.3 s and a random stimulation sequence in which the visual
stimulus occurred randomly every 14~62.3 s. Three central observations emerged. First, the two different stimulation
conditions affect the QPP waveform in different aspects; i.e., systematic stimulation has greater effects on its phase
and random stimulation has greater effects on its magnitude. Second, the QPP was more frequent in the systematic
condition with significantly shorter intervals between consecutive QPPs compared to the random condition. Third, the
BOLD signal response to the visual stimulus across both conditions was swamped by the QPP at the stimulus onset.
These results provide novel insights into the relationship between intrinsic patterns and stimulated brain activity.
Keywords: Functional connectivity, BOLD dynamics in visual perception, Quasi-periodic patterns, Intrinsic spatiotem-
poral dynamics, Infraslow human brain activity
1.
INTRODUCTION
in blood oxygen
Spontaneous fluctuations
level-
dependent (BOLD) signals, recorded by functional mag-
netic resonance imaging (fMRI), capture the hemody-
namic response to neural activity. The infraslow (less than
0.1 Hz) BOLD fluctuations are suggested to have unique
functional and neurophysiological principles that are dis-
tinct from higher frequencies (Chen et al., 2020; Grooms
et al., 2017; Majeed et al., 2009, 2011; Mitra et al., 2018;
Pan et al., 2013; Thompson et al., 2014). The spatiotem-
poral structure of the infraslow BOLD fluctuations has
Received: 14 May 2023 Accepted: 14 May 2023 Available Online: 19 July 2023
provided novel insights into the large-scale functional
architecture of the brain, as well as its changes during
task engagement, development, and disease (Fox &
Raichle, 2007).
One type of spatiotemporal structure consists of a
reproducible pattern of spatial changes that repeat over
time, exhibiting an alternation of high and low activity in
particular areas and propagation of activity along the cor-
tex. These phase-locked quasi-periodic patterns (QPPs)
are found to characterize the intrinsic dynamics of infraslow
BOLD fluctuations in human brains (Bolt et al., 2022;
Imaging Neuroscience, Volume 1, 2023
https://doi.org/10.1162/imag_a_00002
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
© 2023 Massachusetts Institute of Technology. Published under a Creative Commons Attribution 4.0 International (CC BY 4.0) license. Research ArticleN. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
Yousefi & Keilholz, 2021). The primary (or the strongest)
QPP, in particular, displays prominent anticorrelation
between the default mode network (DMN) and task-posi-
tive network (TPN) across rodents and humans (Belloy,
Naeyaert, et al., 2018; Majeed et al., 2011; Raut et al.,
2021; Yousefi et al., 2018; Yousefi & Keilholz, 2021). It has
been shown to correlate with the infraslow neural activity
(Grooms et al., 2017; Thompson et al., 2014), which is
known to be involved in attention (Helps et al., 2010;
Monto et al., 2008) and arousal (Raut et al., 2021; Sihn &
Kim, 2022). The primary QPP can be affected by sustained
attention and other attention control/working memory
tasks (Abbas, Bassil, & Keilholz, 2019; Abbas, Belloy, et al.,
2019), as well as arousal fluctuations (Raut et al., 2021).
The interactions between the infraslow activity and
task- or stimulation-evoked brain responses have been
the focus of much research over the last decade. Several
studies (Chen et al., 2020; Fox et al., 2005; He, 2013;
Huang et al., 2017) reveal that stimulation-evoked BOLD
responses are affected by the magnitude of the sponta-
neous BOLD fluctuations at stimulus onset, namely the
prestimulus baseline, which causes the widely observed
intra-subject trial-to-trial variability in BOLD responses.
However, revealed by Chen and his colleagues, the power
of the evoked infraslow hemodynamics appeared to occur
before the power of neural dynamics (Chen et al., 2020,
Fig. 3e, f), which implies that a significant portion of the
hemodynamics may not directly arise from the neural level.
Because the primary QPP captures the major dynamics of
the intrinsic infraslow brain activity, by investigating stim-
ulus-evoked QPPs, one may probe the interaction between
the stimulation-evoked BOLD response and the sponta-
neous infraslow neural activity. A recent investigation in
stimulation-evoked QPPs was demonstrated in anesthe-
tized mice (Belloy et al., 2021), which suggests that visual
stimulation can trigger the onset of primary QPPs and that
primary QPPs with different phases prior to the visual
stimulus affect the magnitude of the subsequent visual
response. Despite this progress, more remains to be
investigated. Specifically, it is still unclear how environ-
mental perturbations affect the ongoing QPPs and how
ongoing QPPs modulate the BOLD responses to these
environmental perturbations in humans.
Expanding upon previous findings, here we describe a
comprehensive investigation into the relationship between
the primary QPPs and visual stimulation in humans. Given
that primary QPPs associated with both attention (Abbas,
Bassil, & Keilholz, 2019; Abbas, Belloy, et al., 2019) and
arousal fluctuations (Raut et al., 2021), we aimed to
explore how QPPs interact with visual stimuli under con-
ditions that held arousal fluctuations constant. We
employed two conditions of different sequences of visual
stimulation induced by flickering checkerboard flashing at
6 Hz (which is unlikely to affect arousal levels), one involv-
ing a systematic stimulation sequence where the visual
stimulus appeared every 20 s, and the other involving a
random stimulation sequence where the visual stimulus
(average
randomly between 14~62.3 s
occurred
19.95 s ± 6.37 s). Notably, a systematic sequence has
been routinely employed for evoking the infraslow sponta-
neous BOLD signals in previous studies (Belloy et al.,
2021; Duann et al., 2002). These sequences have been
shown to entrain the intrinsic rhythms of low-frequency
brain oscillation to the structure of the attended stimulus
stream (Ding et al., 2006; Lakatos et al., 2008) and
enhance attention (Ding et al., 2006; Jones et al., 2002;
Lakatos et al., 2008; Qiao et al., 2022). While our experi-
ment only involved a basic sensory stimulation paradigm
without requiring any responses or measuring attention
behaviors, our findings may provide novel insights into
how attentional processes are affected by sensory stimu-
lation for future studies.
Given that QPPs are closely associated with infraslow
neural activity (Grooms et al., 2017; Thompson et al.,
2014), the intervals between predetermined stimuli were
specially designed to ensure the frequencies of the
presentation of these noninvasive stimuli within the
infraslow range, i.e., 0.049 Hz for the systematic and
0.016 Hz~0.07 Hz for random stimulation, that was sug-
gested to modulate the infraslow neural fluctuations (Qiao
et al., 2022). Additionally, an equal number of stimuli were
presented across both systematic and random sequences.
Hence, we can investigate the interaction between QPP
and the stimuli and compare the results across two visual
stimulation conditions. Using the resting-state results as
the control, we specifically attempted to answer the fol-
lowing three questions: 1) How do QPP waveforms differ
between systematic and random visual stimulation condi-
tions? 2) How do the different visual stimulation sequences
impact the frequency and/or intervals of consecutive
QPPs? 3) How do the different QPP phases prior to the
stimulus modulate the subsequent visually evoked BOLD
responses in different visual stimulation conditions?
2. METHODS
2.1. Data acquisition and preprocessing
Functional MRI brain images of fourteen young adults (8
women, 6 men) in the Atlanta area participated in this
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
2
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
Institutional Review Board
experiment (mean age = 19.8 ± 1.7 yro; range [18–24 yro]).
The fMRI scanning was performed at the Center for
Advanced Brain Imaging (CABI) in Atlanta on a 3T Sie-
mens Trio scanner with a 12-channel radio frequency coil.
This study received research ethics approval from the
Georgia Tech
(protocol
H17227). All subjects gave informed consent. For each
subject, 7 complete gradient-echo echoplanar imaging
(EPI) scans were acquired followed by an anatomical T1
image (MPRAGE; TE = 3.98 ms, flip angle = 9°, matrix
256 × 256 (RO × PE), slice thickness = 1.0 mm, 176 slices,
and voxel size 1 × 1 × 1 mm3). Each EPI scan has 870
timepoints, i.e., Nx = 870 with the sampling rate, TR = 0.7s
for a duration of 10 min and 9 s. Other acquisition param-
eters of EPI scans include TE = 30 ms, flip angle = 90°,
matrix 64 × 64 (RO × PE), slice thickness = 3.0 mm with 22
slices, voxel size 3.4375 × 3.4375 × 3 mm3, multiband fac-
tor = 2, echo spacing = 0.51 ms, and BW = 30.637 Hz/Px.
Due to the short TR (TR = 0.7 s) of the single-shot gra-
dient echo-planar imaging (EPI), it is not possible to scan
the entire brain. Therefore, certain brain regions are
excluded from the EPI scan. As the effects of flickering
checkerboard on the visual cortex and visual processing
have been extensively studied in the past (Dale & Buckner,
1997; Engel et al., 1997; Schwartz et al., 2005; Tootell
et al., 1998), the current study focused on how this visual
stimulation, known to activate or deactivate various
regions across the whole brain (Jorge et al., 2018), affected
the remaining brain regions, such as the default and
task-positive networks. Hence, the orbital frontal cortex,
temporal pole, dorsal motor areas, and occipital lobe were
excluded from each EPI scan, and the corresponding
regions were identified using the Schaefer-Yeo Atlas
(Schaefer et al., 2018) in the final preprocessing step.
Each of the seven EPI scans for each subject fell into
one of three distinct experimental conditions. For all sub-
jects, a resting-state scan lasting 10.15 min (870TRs) was
the first functional scan collected. During the resting scan,
subjects were told to stay awake and remain still while
staring at a fixation cross. After the completion of the rest-
ing-state scan, six visual stimulation EPI scans were col-
lected using two visual stimulation conditions. During
both conditions subjects were told to focus on a red fixa-
tion cross at the center of the projection screen and that
on occasion a flashing checkerboard would appear in the
background. For both conditions, the flashing periods
were comprised of a black and white checkerboard pat-
tern that inverted every 5 refresh frames (60 Hz) for a
period of 2.1 s (3TRs). The red fixation cross remained at
the center of the screen during the checkerboard periods.
Half of the scans used a systematic stimulation sequence.
That is, a flashing checkerboard stimulus appeared for
2.1 s (3TRs) every 20.3 s (29TRs). The other three scans
used a random stimulation sequence. That is, the flashing
checkerboard stimulus appeared randomly at every
13.3~61.6 s (19~88TRs, average arrival time 19.25 s ±
6.34 s). For the systematic condition, the stimulus interval
of 20.3 s ensures at most one QPP occurring during or
after each stimulus onset. For the random condition, mul-
tiple QPPs could occur between some of the long stimu-
lus intervals. For both visual stimulations, the range of
stimulus intervals was selected to warrant a total of 30
stimulation onsets during each EPI scan, which allows us
to compare the interaction between each stimulation and
ongoing QPP between the two visual conditions. The two
types of stimulation sequences alternated in an ABABAB
order, with the order counterbalanced between subjects.
An illustrative example of these two different visual stimu-
lation sequences is provided in Figure S1. Between each
EPI scan, there was a roughly 30-s time gap, during which
we told the subjects to rest their eyes and remain still,
inquired on their wakefulness during the preceding scan,
and informed them of the time remaining until the end of
the scanning session.
The acquired fMRI data were preprocessed by an
automated pipeline based around SPM12 (https://www
. fil . ion . ucl . ac . uk / spm / software / spm12/), FSL (Jenkinson
et al., 2012), and AFNI (Cox, 1996; Cox & Hyde, 1997).
First, the anatomical T1 image was spatially normalized
to the 2 mm Montreal Neurological Institute (MNI) atlas.
This step includes an image reorientation to the MNI
space using FSL, a bias-field correction using FEAT
(Y. Zhang et al., 2001), and the SPM segmentation model,
which performed the tissue segmentation of gray matter,
white matter, cerebrospinal fluid (CSF), bone, soft tissue
and air/background of gray matter, and the spatial nor-
malization of these segmented tissues. The binary mask
of the white matter, CSF, and the whole brain (gray mat-
ter, white matter, and CSF) was obtained by thresholding
at the top 70% of these normalized tissues.
Next, the functional EPI timeseries were preprocessed
following procedures as described in Abbas, Belloy, et al.
(2019). Specifically, the following six steps were per-
formed. First, in order to normalize all scans of each sub-
ject to the same template, all seven EPI scans of each
subject were concatenated. Second, the concatenated
EPI data were reoriented (FSL), realigned (SPM12), and
normalized to the MNI atlas based on the estimates of the
SPM segmentation model from the anatomical data
preprocessing. In parallel, the motion parameters for the
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
3
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
concatenated EPI data, including the framewise displace-
ment (FD), were also estimated using MCFLIRT (FSL).
Here, the FD is estimated by the relative root-mean-square
movement (Jenkinson et al., 2002) in order to examine the
in-scan head motions in step six. Third, the normalized
EPI data were spatially smoothed with a Gaussian kernel
of 4 mm (SPM smooth). Fourth, the concatenated EPI
data were split back to each scan. Fifth, the EPI data of
each scan were temporally filtered at a bandwidth of 0.01
Hz~0.1 Hz (AFNI 3dBandpass), and further regressed by
the mean signals extracted from the white matter and CSF
masks. Sixth, for each scan, the head motions were exam-
ined by the FD following the criteria described in Yousefi
et al. (2018). Specifically, scans with low to moderate lev-
els of motion (i.e., mean FD < 0.2 mm and with a temporal
ratio of FD > 0.2 mm smaller than 40%) were included in
our analysis (see Fig. S2 for more details), because the low
to moderate levels of motion were found to have minimal
impact on the QPP being detected (Yousefi et al., 2018).
Note
the preprocessing procedures
described above have demonstrated success in detecting
QPPs from resting as well as task-evoked human brains
(Abbas, Belloy, et al., 2019), additional preprocessing pro-
cedures such as motion parameter regressions and vol-
ume scrubbing were not performed.
that because
Finally, the preprocessed EPI timeseries were extracted
from the brain parcels provided by the Schaefer-Yeo Atlas
(Schaefer et al., 2018) (github) and then z-scored. Due to
the incomplete brain coverage of the EPI scans (as
described in the 2nd paragraph of this section), only par-
cels with over 85% coverage across all subjects were
selected from the Schaefer-Yeo Atlas. The percentage of
EPI scanning coverage of each Schaefer-Yeo parcel of
each subject is reported in Figure S3. Notably, because
the primary and the majority of the secondary visual cortex
weren’t covered for all EPI scans, the visual network was
excluded from the analysis. In addition, because the tem-
poral lobe was not covered and most of the remaining par-
cels in the limbic system have less than 85% coverage,
the limbic network was also excluded from the analysis. As
a result, EPI timeseries from a total of 193 parcels were
extracted, which covered the 5 functional brain networks,
as described in Thomas Yeo et al. (2011), including the
somatomotor (SM), dorsal attention (DA), ventral attention
(VA), frontoparietal (FP), and default (D) networks.
2.2. Quasi-periodic pattern detection and examination
The primary QPP is a phase-locked spatiotemporal pat-
tern detected from the BOLD fluctuations, which repeats
over time. This intrinsic dynamic pattern has been found
to tie to the infraslow electrical activity (<0.1 Hz) (Grooms
et al., 2017; Raut et al., 2021; Thompson et al., 2014). In
this section, we first describe 1) the detection of the pri-
mary QPP and the corresponding QPP with a reverse
phase, and then 2) the parameters used for detecting
QPPs. Moreover, we describe 3) the rationale and
approach of detecting QPPs at a group level, and 4) the
procedures for comparing different QPPs in the final
paragraph.
2.2.1. Primary QPP, reverse-phase QPP, and their occurrences
Primary QPP and its occurrence across the entire
timeseries for each experimental condition were detected
on the EPI data using the robust QPP detection algorithm
described in Yousefi and Keilholz (2021). This is a correla-
tion-based and iterative finding algorithm, which identi-
fies similar segments of a functional timecourse and
averages them for a representative spatiotemporal tem-
plate. The algorithm detection process can be summa-
rized by the following six steps. First, an initial segment
with a preset window length (WL) was selected at the ith
timepoint (i = 1,…,Nx − WL) of the EPI timeseries of all
ROIs. This initial segment has a spatial dimension and a
temporal dimension (ROIs × WL) and was used as the ref-
erence QPP template for later steps. Second, the refer-
ence template was correlated with a segment with the
same window length across all ROIs, which was sliding
from the 1st to the Nx − WL timepoints of the timeseries at
a step of 1 timepoint, which resulted in a timecourse of
sliding correlations. Third, local maxima of this correla-
tion timecourse, which are above a preset positive
threshold and also have a minimum distance of WL, were
selected as the occurring time of the reference template,
and segments with starting points at these local maxima
were averaged to obtain an updated template. Fourth,
steps 2 and 3 were iterated until the averaged template
and the reference converge. Fifth, steps 1–5 were
repeated for all is (i = 1,…,Nx − WL), which resulted in
totally Nx − WL sets of results. Notably, the detection pro-
cess omits the final WL timepoints to avoid QPP finding
at the time boundary of different scans. Sixth, the Nx − WL
sets of results were ranked based on the summation of
local maxima of the correlation timecourse, and the set of
results with the greatest summation was selected as the
final solution. This entire process, also summarized in the
flowchart (Yousefi & Keilholz, 2021, Figs. S2–S3), pro-
duced two major outputs, one is the averaged 2D tem-
plate of timeseries, which is the primary QPP; the other is
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
4
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
the timecourse of sliding correlation, of which selected
local maxima are considered as of the occurrence of this
QPP. It is worth mentioning that the primary QPP in
resting humans displays a sinusoidal waveform (Abbas,
Belloy, et al., 2019; Belloy, Shah, et al., 2018; Yousefi
et al., 2018; Yousefi & Keilholz, 2021). Specifically, the
primary QPP in resting human brains is half-wave sym-
metric, comprising nearly identical half-cycles with oppo-
site polarities (Yousefi & Keilholz, 2021, Fig. S8b).
Each primary QPP is paired with a corresponding
QPP in the reverse phase, known as the reverse-phase
QPP.1 While the primary QPP is obtained by averaging
the segments that start from the selected local maxima
of the correlation timecourse, the reverse-phase QPP
can be obtained by averaging the segments starting
from the selected local minima of the same correlation
timecourse. These selected local minima were required
to be separated by at least WL and have a negative
magnitude below a predetermined threshold. In the rest-
ing dataset of the Human Connectome Project (HCP),
the primary QPP detected in concatenated scans of a
subject may begin from a positive magnitude (like the
sine wave) or a negative magnitude (like the −sine wave).
Remarkably, the primary QPP with a “−sine” waveform is
highly similar (Pearson correlation r > 0.88, p-value < 0.01)
to the reverse-phase QPP associated with a primary
QPP with a “sine” waveform (Yousefi & Keilholz, 2021,
Figs. S25–S26, Video 3). Hence, both the primary QPP
and its reverse-phase QPP have been utilized in the pri-
mary QPP analysis for studying resting-state popula-
tions. However, when analyzing the occurrences of
QPPs for the resting populations (see Section 2.3), we
only considered the occurrences of the primary QPPs.
This is because the QPP correlation timecourse was
selected based on the summation of the local maxima,
which were limited to the primary QPP only, and not on
its reverse-phase counterpart or the local minima.
2.2.2. Parameters for QPP detection
The QPP window length was selected based on the
duration of QPP templates observed in previous studies
(Majeed et al., 2011; Thompson et al., 2014; Yousefi
et al., 2018), and a common window length was chosen
for easy comparison across different experimental con-
ditions (e.g., for the correlation calculation described in
the final paragraph). Because the duration of QPP lasts
1 If the detected primary QPP starts from positive QPP values which is
followed by negative QPP values, then the reverse phase QPP starts from
negative QPP values and is followed by positive QPP values, and vice versa.
for approximately 20 s in both resting (Majeed et al.,
2011; Thompson et al., 2014; Yousefi et al., 2018) and
evoked human brains (Abbas, Belloy, et al., 2019), QPP
window lengths (WLs) ranging from 17.5 to 24.5 s
(WL = 25TRs~35TRs) were explored. The final window
length was determined by identifying the point at which
increasing the window length would no longer change
the appearance of the QPP across all experimental con-
ditions. This ensured that the selected window length
was appropriate for detecting the primary QPP in all
experimental conditions. In addition, the positive correla-
tion thresholds and maximum iterations were selected
based on previous studies (Abbas, Bassil, & Keilholz,
2019; Abbas, Belloy, et al., 2019; Belloy, Naeyaert, et al.,
2018; Belloy, Shah, et al., 2018; Majeed et al., 2011;
Yousefi et al., 2018). In particular, a positive QPP correla-
tion threshold of 0.1 for the first three iterations and 0.2
for subsequent iterations was selected with a maximum
of 20 iterations. A negative correlation threshold of −0.2
was set for detecting the reverse-phase QPP. The above
parameters have been shown to be effective in detecting
primary QPPs across various experimental conditions in
humans (e.g., TR = 0.3~2 s, resting state, N-back task,
and disease models) (Abbas, Bassil, & Keilholz, 2019;
Abbas, Belloy, et al., 2019; Yousefi et al., 2018).
2.2.3. Group QPP analysis for each experimental condition
Group QPP analysis is a common approach for compar-
ing primary QPPs across different experimental condi-
tions or populations (Abbas, Bassil, & Keilholz, 2019;
Abbas, Belloy, et al., 2019; Majeed et al., 2011). In this
approach, all EPI scans from each experimental condition
or population are concatenated into a single timeseries,
which is then subjected to detection of the primary QPP.
By comparing the primary QPPs across different groups,
researchers can gain insights into the differences or simi-
larities in brain dynamics between different conditions
(Abbas, Bassil, & Keilholz, 2019; Abbas, Belloy, et al.,
2019; Majeed et al., 2011). Recently, a subject-level QPP
analysis was employed on the high-quality resting-state
HCP dataset, in which 4 EPI scans of each subject were
concatenated (resulting in a 57.6-min concatenated EPI
data) for the QPP detection (Yousefi & Keilholz, 2021).
This approach could reveal inter-subject variabilities
within one experimental condition.
In our study, because we aim to investigate how differ-
ent sequences of visual stimulation (systematic or random)
influence spatiotemporal brain dynamics, and because
we have far less EPI data for each experimental condition
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
5
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
for a subject-level analysis (i.e., a total of 10.15-min EPI
recording for resting and 30.45-min EPI recording for the
systematic and for the random conditions), we have
designed a group QPP analysis with repeating groups to
assess the variabilities within each visual condition. Specif-
ically, the extracted EPI timeseries from each scan were
concatenated across all 14 subjects, which results in 1
resting group, 3 repeating groups with systematic stimuli
and 3 repeating groups with random stimuli. Each group
includes a 142.1-min concatenated EPI data, which
according to our previous study (Yousefi & Keilholz, 2021)
is sufficient for providing reproducible results under one
experimental condition. The three repeating groups with
visual stimulations will be used to test the variabilities and
reproducibility within each visual condition. In the remain-
der of the paper, we refer to this group EPI timeseries as the
resting, the systematic 1, 2, or 3, and the random 1, 2, or 3.
For the resting-state condition, the group primary QPP
(QPPrest), and its reverse-phase QPP (QPPrest −), as
well as their occurrences were obtained from the detec-
tion algorithm. For the systematic condition, timeseries of
all 3 repeating groups were first concatenated for gener-
ating a group average QPP (QPPsys) and the correlation
timecourse across all 3 groups. Then, the QPP for each
group (systematic 1, 2, and 3) was obtained by only aver-
aging the EPI segments starting at the correlation local
maxima in each group (QPPsysi for i =1,..,3). Similarly, the
group average QPP (QPPrand) and three group QPPs
(QPPrandi for i = 1,..,3) were obtained for the random
stimulation data. The systematic and random group
relationships:
the
average QPPs have
QPPsysi
QPPsys =
.
QPPrandi
In addition, the reverse-phase QPP was also obtained for
each visual condition, denoted by QPPsys − and
QPPrand −. We refer to this set of results as group analyt-
ical results. An illustration of these results and the pro-
cess that arrived at them are shown in Figure S4. To
further test the reproducibility of the results, we also per-
formed an independent group analysis, which detected
the QPP independently for each task group, which
obtained QPP!sysi and QPP!randi for i = 1,…,3. The data
used for the group average analysis and the independent
group analysis are shown in Table S1.
following
and QPPrand =
3∑
3∑
i =1
i =1
2.2.4. Comparison between two primary QPPs
Comparing multiple primary group QPPs is a common
approach to examining spatiotemporal brain dynamics
across different experimental conditions or populations
(Abbas, Bassil, & Keilholz, 2019; Abbas, Belloy, et al.,
2019; Majeed et al., 2011). Here, we aimed to assess the
differences between the primary QPP waveforms in the
systematic and random visual conditions, however, we
found that QPPsys and QPPrand have opposite phases.
It is worth noting that the prior study (Yousefi & Keilholz,
2021) discovered that both QPPrest and QPPrest − were
suitable for primary QPP analysis in the resting state. To
facilitate comparison, we measured the differentiation
between the QPPs of each visual condition and the rest-
ing QPP with the same phase. The QPP observed during
the visual stimulation was considered the empirical result,
while the resting-state QPP used for comparison was
referred to as the null model.
More specifically, we performed the assessment in
three steps. Firstly, we calculated the correlation coeffi-
cients between the empirical (QPPsys or QPPrand) and
the null model (QPPrest or QPPrest −). Secondly, we con-
ducted a z-test to determine if the correlation of the two
visual conditions differed significantly from each other.
Specifically, the Fisher z-transformed correlation coeffi-
cient was computed from each condition and then the
z-score of their differences was computed with sample
normalization (Diedenhofen & Musch, 2015). The signifi-
cance of the difference was also tested at the average
level of the three groups. Finally, we examined the dis-
tinctions in the entrained QPP waveforms from the fol-
lowing four aspects: 1) the phase shift from the null, 2) the
amplitude changes in percentage from the null, 3) the
vertical shift in the percentage of the amplitude of the
null, and 4) the percentage changes in “peak-life” of the
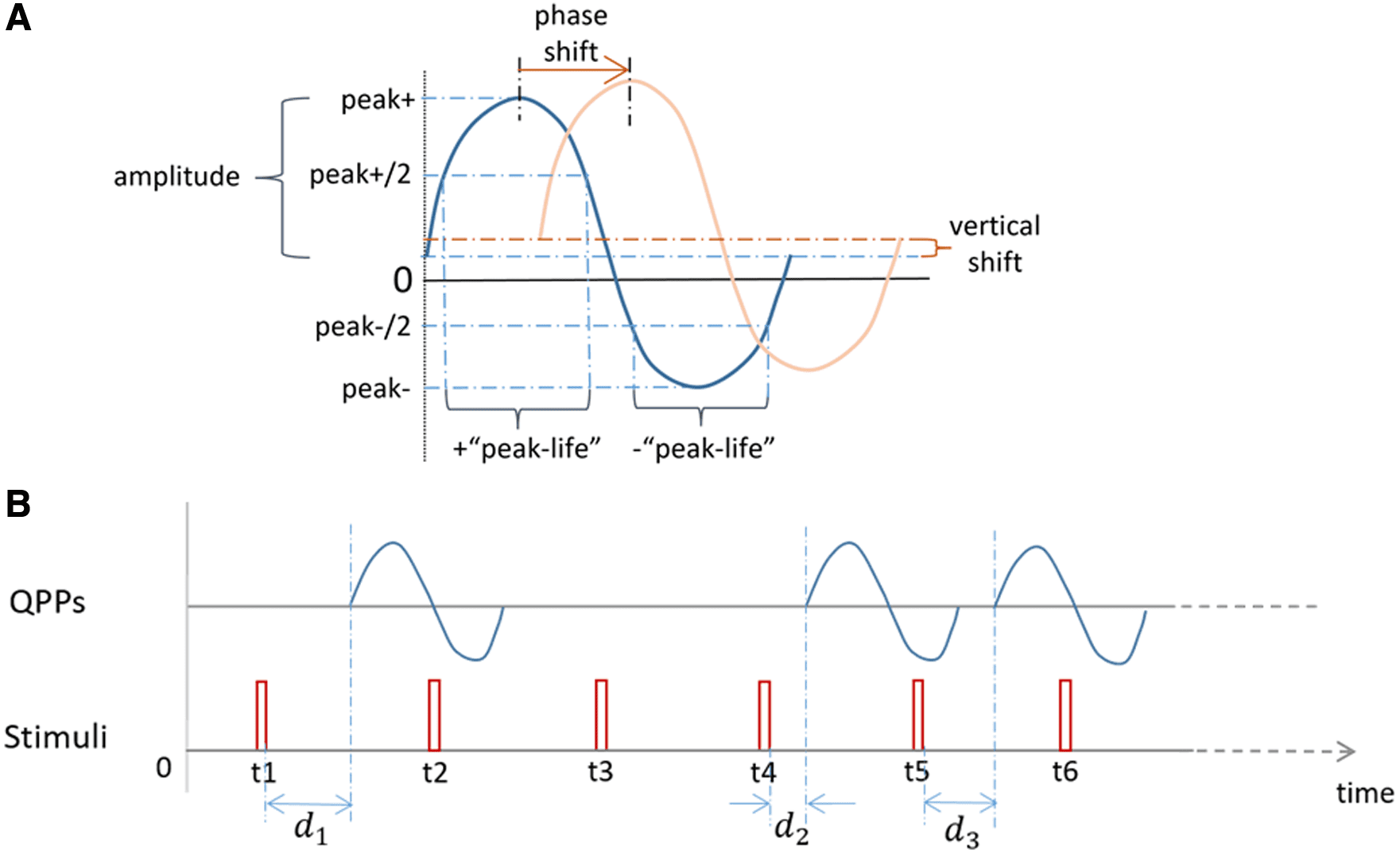
positive and negative peaks from the rest (see Fig. 1A).
Because the QPP of parcels in the same brain network
was found to share the same waveform (Yousefi et al.,
2018; Yousefi & Keilholz, 2021), the waveform examina-
tion was conducted at the network level, i.e., the network
QPP obtained by averaging the QPP of parcels in each of
the five networks described in the second to the last
paragraph of Section 2.1. More specifically, the phase
shift was estimated using Zhivomirov (2022), which
implements the algorithm in Sedlacek and Krumpholc
(2005). The “peak-life,” in particular, is defined as the
entire period that the waveform stays from its peak to half
of its peak. A 2-way ANOVA and multiple comparison
tests were used to compare the two visual conditions for
each waveform characteristic across all networks.
2.3. Entrained QPP occurrence analysis
The study also investigated the association between the
onset of visual stimuli and the occurrence of QPPs. This
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
6
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
Fig. 1. QPP waveform and time delays of QPPs occurrence followed by stimuli. (A) Characteristics that describe QPP
waveform distinctions, which include the phase and vertical shifts, the amplitude changes, and distinctions in the “peak-
life” of the positive and negative peaks. (B) Time delay of each ongoing QPP post the stimulus onset, denoted by di. Only
nonzero di’s were included in the analysis.
analysis utilized the primary QPP occurrences obtained
from both the group average and independent group
results (see Table S1). The investigation aimed to answer
two questions. The first question examined whether the
type of visual stimulation sequence, i.e., systematic or
random influences the frequency and intervals of con-
secutive QPPs. The second question aimed to investi-
gate whether the onset of stimulation in either visual
condition affects the timing of ongoing QPPs (i.e.,
advance or delay the onset of successive QPPs).
To address the first question, we calculated the fre-
quency of QPP occurrence over time for each group. This
was done by dividing the total number of QPP occurrences
by the number of timepoints in the 14 concatenated scans.
The resulting frequency was then averaged across the
repeating groups and compared between the three experi-
mental conditions. Additionally, the intervals of consecutive
QPPs were also computed, and the mean of the intervals
was compared between the systematic and random visual
conditions. A t-test was then performed to determine if the
measured mean difference between the two conditions
was significantly greater or smaller than zero.
To investigate the second question, we calculated the
time delay of QPP occurrence after each stimulus
(Fig. 1B) and formulated two hypothesis tests, one for
each visual condition. The null hypothesis was that the
visual stimuli did not affect (i.e., delay or advance) the
timing of ongoing QPPs, while the alternative hypothesis
was that they did. For each visual condition, the null
model assumed that the timing of QPP was the same as
that observed during the resting state. The null distribu-
tion of QPP delay was constructed by comparing the cor-
relation timecourse of resting QPP with the stimulation
sequence and calculating the time delay of QPP occur-
rence following each visual stimulus. Different null distri-
butions were generated for each visual condition as the
two types of stimulation sequences had different stimu-
lus onset times, which introduced different cutoff values
for QPP delays (i.e., <29TRs for the systematic condition
and <88TRs random condition). An example of null empirical
model QPP delay is shown in Fig-
ure S9. Finally, each hypothesis test, a two-sample t-test was used to determine
if mean result significantly differed from model.
Downloaded http:> 1.96) after z-scoring the aver-
aged contrasts among all 193 parcels.
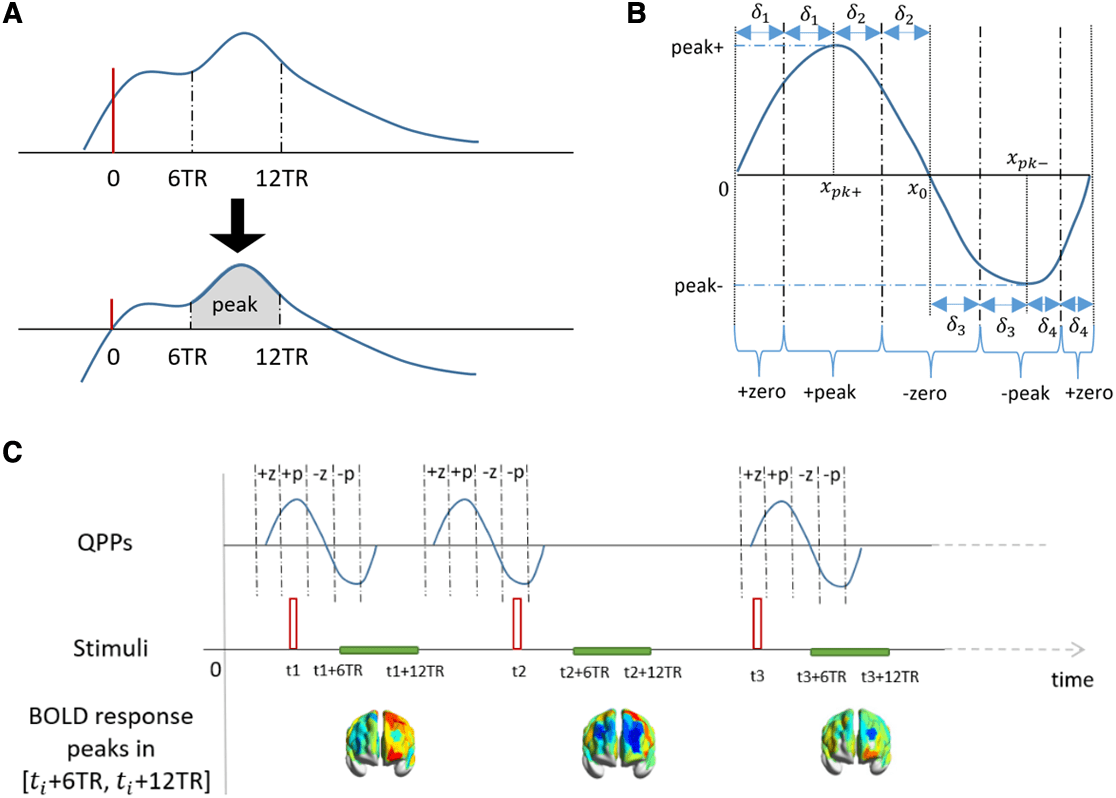
Fig. 2. Dependence of the BOLD response to visual stimulation and QPP phase. (A) Determination of peak value (shaded
area) of evoked BOLD responses. The red vertical line depicts the onset of visual stimulation. The BOLD magnitudes
in [6TR, 12TR] were subtracted by the BOLD magnitude at the stimulus onset. The gray area under the bottom curve
depicts the peak value of the stimulus response. (B) Illustration of four-phase zones of a QPP. Let xpk +, xpk −, and x0 be
the timepoint of the positive peak, negative peak, and the midpoint where the QPP wave across the zero, respectively.
Based on these timepoints, four pairs of intervals can be determined as δ1 =
PL − xpk −
2
, where WL is the QPP window length. Then, the four phases +zero, +peak, −zero, −peak, have the
and δ4 =
following zones: [0, δ1] ∪ [PL − δ4, PL], [ xpk+ − δ1, xpk + + δ2 ], [ x0 − δ2, x0 + δ3 ], [ xpk − − δ3, xpk − + δ4 ]. (C) BOLD stimulus with
stimulus onset overlaps ongoing QPPs at different phases.
xpk +
2
, δ2 =
x0 − xpk +
2
, δ3 =
xpk − − x0
2
,
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
8
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
To investigate the effect of visually evoked BOLD
responses on overlapping QPP at different phases, we
divided the QPP of each parcel into four distinct phase
zones, referred to as “+zero,” “+peak,” “−zero,” and
“−peak.” This division was achieved through a 4-step
process. Firstly, the timepoints of the positive and nega-
tive peaks of the QPP wave, as well as the midpoints
where the wave crossed zero, were determined for each
parcel. Secondly, we identified four pairs of non-overlap-
ping time intervals (δ i for i = 1,2,3,4) covering the entire
window length [0, PL]. Thirdly, we defined the “+peak”
(“−peak”) phase zone as the interval(s) stepping away
from the positive (negative) peak within the correspond-
ing δ i. Finally, the “+zero” (“−zero”) zone was identified as
the non-overlapping interval(s) containing the QPP across
zero with an uprising (a down-falling) trend. The four-
phase zones for an exemplary QPP waveform are illus-
trated in Figure 2B and the BOLD stimulus responses
overlap different phases of the ongoing QPP are illus-
trated in Figure 2C.
The BOLD responses to the stimuli that coincide with
each phase of QPP (QPPsys, QPPrand) and of its
reverse-phase counterpart (QPPsys −, QPPrand −) were
averaged across the stimuli. Additionally, a control
group was included by averaging the BOLD responses
to stimuli that did not overlap an ongoing QPP. The
resulting averaged BOLD responses for each phase
were then compared across brain regions (parcels) and
between the two types of stimulation sequences (sys-
tematic and random), as well as between the empirical
and null results.
3. RESULTS
3.1. Differences in group-level QPPs across systemic and random
stimulations
First of all, the group average QPP affected by the sys-
tematic stimulations (in comparison to the random stim-
ulations) appeared to be more distinct from the resting
state. Differences in group average QPPs for each visual
condition in comparison to the resting QPP were shown
both among all parcels (Fig. 3A) and for each network
(Fig. 3B). Numerically, the group average resting QPP
has a significantly lower correlation to QPPsys than to
QPPrand (i.e., z-score = −36.76, p-value < 0.01), as the
Fisher z-transformed correlation between QPPrest and
QPPsys is 1.003 (Pearson correlation r = 0.763, p-value
< 0.01), and it is 1.673 (Pearson correlation r = 0.932,
p-value < 0.01) between QPPrest − and QPPrand. This
significantly lower correlation value also appears in
the between-group calculations (Table S2, z-score =
−32.402, p-value < 0.05), as well as in the average of
independent group analysis (Fig. S7, z-score = −12.396,
p-value < 0.05). The group average QPPsys − and
QPPrand − are shown in Figure S5. The resting QPPs
demonstrate (anti-)correlations between different net-
works that are similar to the previous findings on the
HCP dataset (see Fig. S6). The positive and negative
phases of both QPPrest and QPPrest − appeared to
have same duration, which is also consistent with the
previous findings (Yousefi & Keilholz, 2021). QPPs
detected for each independent group are shown in
Figure S7. For each experimental condition, the QPP
window length of 21.7 s (WL = 31TRs) was selected fol-
lowing the procedure as described in Section 2.2.
Secondly, systematic stimulations versus random
stimulations appear to affect the network QPP wave-
forms in different ways. Compared to the random stimuli,
the systematic stimuli have a more significant effect
(Bonferroni corrected p-value < 1e-4) on the phase of
QPPs. On the other hand, the random stimuli have a
more significant effect (Bonferroni corrected p-value
< 1e-4) on the magnitude of QPPs (Fig. 3C and Table S3).
Specifically, in the random condition, the magnitude of
QPP shifts more positively in the somatomotor and the
three task-positive networks, but shifts more negatively
in the default network compared to the systematic condi-
tion. This QPP vertical shift is also combined with changes
in amplitude. For example, QPP amplitudes increase
more in dorsal and ventral attention and decrease more
in frontoparietal and default for the random than the sys-
tematic condition. In addition, the phase shift affected by
the systematic stimuli (an average of 29.18 degrees) is
greater than the one affected by the random stimuli (an
average of |3.65| degrees). This greater phase shift by
systematic stimulations is also reflected as a squeezed
+peak in the task-positive networks for both stimulation
sequences. For example, the QPP wave’s +“peak-life” in
dorsal attention, ventral attention, and frontoparietal net-
works is shortened by 35.41% on average by the sys-
tematic stimuli but only shortened by 18.48% on average
by the random stimuli.
3.2. Effect of visual stimulation on the incidence of QPPs
On average, the systematic visual condition had a higher
frequency of QPP occurrences compared to the random
condition, while the resting state was in between (see
Table 1 for the group average frequency). This finding is
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
9
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
Fig. 3. Group average QPP for resting, systematic, and random visual stimuli. (A) The global spatiotemporal QPP across
different experimental conditions. For each visual condition, both empirical and null QPPs are shown. These QPPs were
simultaneously detected from all 193 parcels, covering five networks, somatomotor (SM), dorsal attention (DA), ventral
attention (VA), frontoparietal (FP), and default (D). The y-axis of each pattern corresponds to the spatial dimension, while
the x-axis corresponds to the temporal dimension. (B) Network QPP averaged among parcels in each of the five networks.
Both the null and empirical QPP curves are displayed in each plot, with alignment at the first timepoint. To emphasize the
changes in waveform of the empirical QPP for each network, the timepoint where the magnitude of the null QPP curve
crosses zero was selected as the reference point (0 s). (C) Bar plots of the average changes in three characteristics of the
QPP waveform, including the amplitude changes (%), vertical shift (%), and phase shift (deg), as perturbed by the visual
stimulation (empirical) when compared to the resting QPP (null). In the case of the frontoparietal network, because the
amplitude of QPPrand is attenuated by over 50% and is almost close to zeros, the phase shift cannot be appropriately
determined, and hence a “NA” is reported. The significance level of the multiple comparison test between the two visual
conditions is denoted above each characteristic, with “ns” and “****” representing Bonferroni corrected p-values greater
than 0.05 and less than 1e-4, respectively. Bar plots of the average changes in all five waveform characteristics, including
the + and −“peak-life” changes (%) and the above three characteristics are shown together with the numerical values for
these changes and the multiple comparison test results in Table S3.
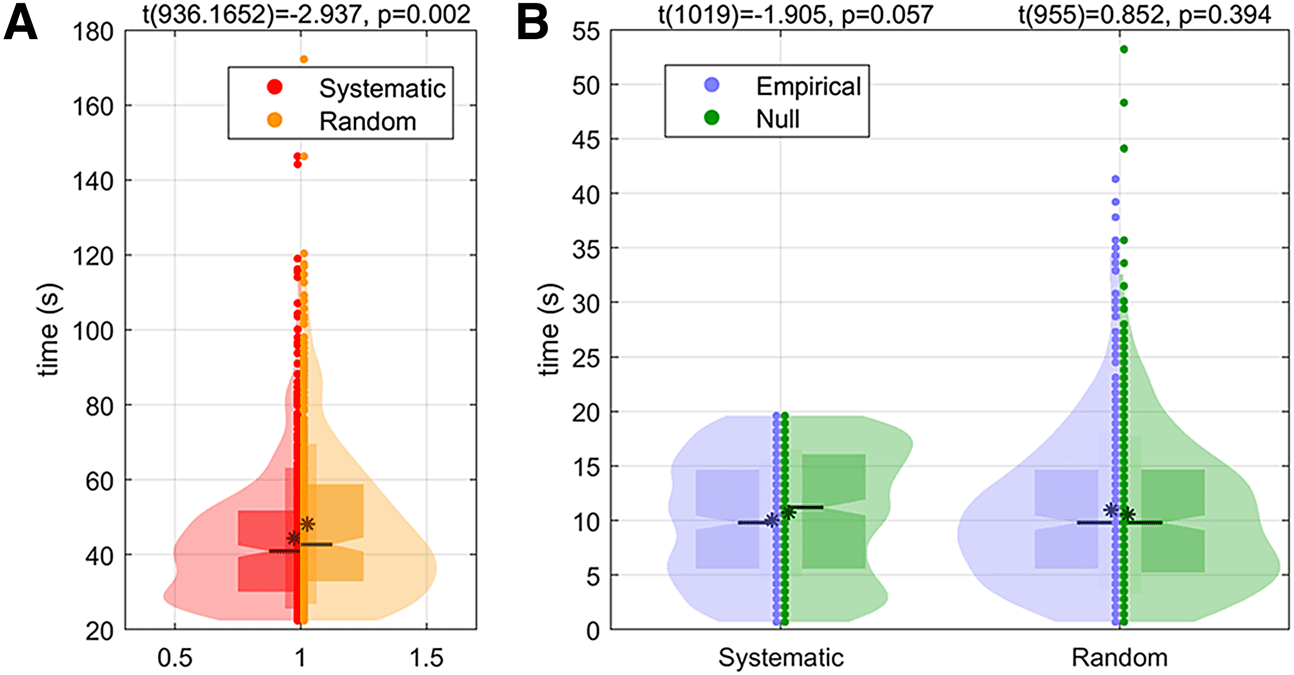
also supported by the shorter QPP intervals observed in
the systematic condition compared to the random condi-
tion, as illustrated in Figure 4A. In particular, as shown in
Figure 4A, the mean of the random condition QPP inter-
vals is significantly greater than the systematic condition
(i.e., p-value = 0.002 < 1%). In the independent group
analysis, the mean of QPP intervals for the three random
groups is also greater than for the three systematic
groups (see Fig. S8, i.e., p-value = 0.007 < 5%).
On the other hand, the QPP time delay in each visual
condition exhibits no significant difference from its null
model (see Fig. 4B). In Figure 4B, the probability density
function of the time interval between the stimulus and the
onset of a subsequent QPP is shown. These QPP delays
were compared to the null model (computed from the
resting data as described in Section 2.3) for each visual
stimulation condition. Numerically, despite the slight dif-
ference in the mean of QPP delays for each condition,
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
10
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
Table 1. Frequency of QPP occurrences for each group
Group average
results
Independent group
results
Resting
Systematic
Random
1.46%
1.47% ± 0.03%
1.41% ± 0.07%
1.46%
1.55% ± 0.03%
1.45% ± 0.05%
For each experimental condition, the group average result is
averaged by the frequency of the group average QPP that
occurred in each group, whereas the independent group result is
averaged by the frequency of QPP that is independently detected
in each group.
neither difference is significant (i.e., the systematic condi-
tion has p-value = 0.057 > 5%, and the random condition
has p-value = 0.394 > 5%, and also see Fig. S10 for the
independent group results).
3.3. QPP phase dependence of BOLD response to visual stimulation
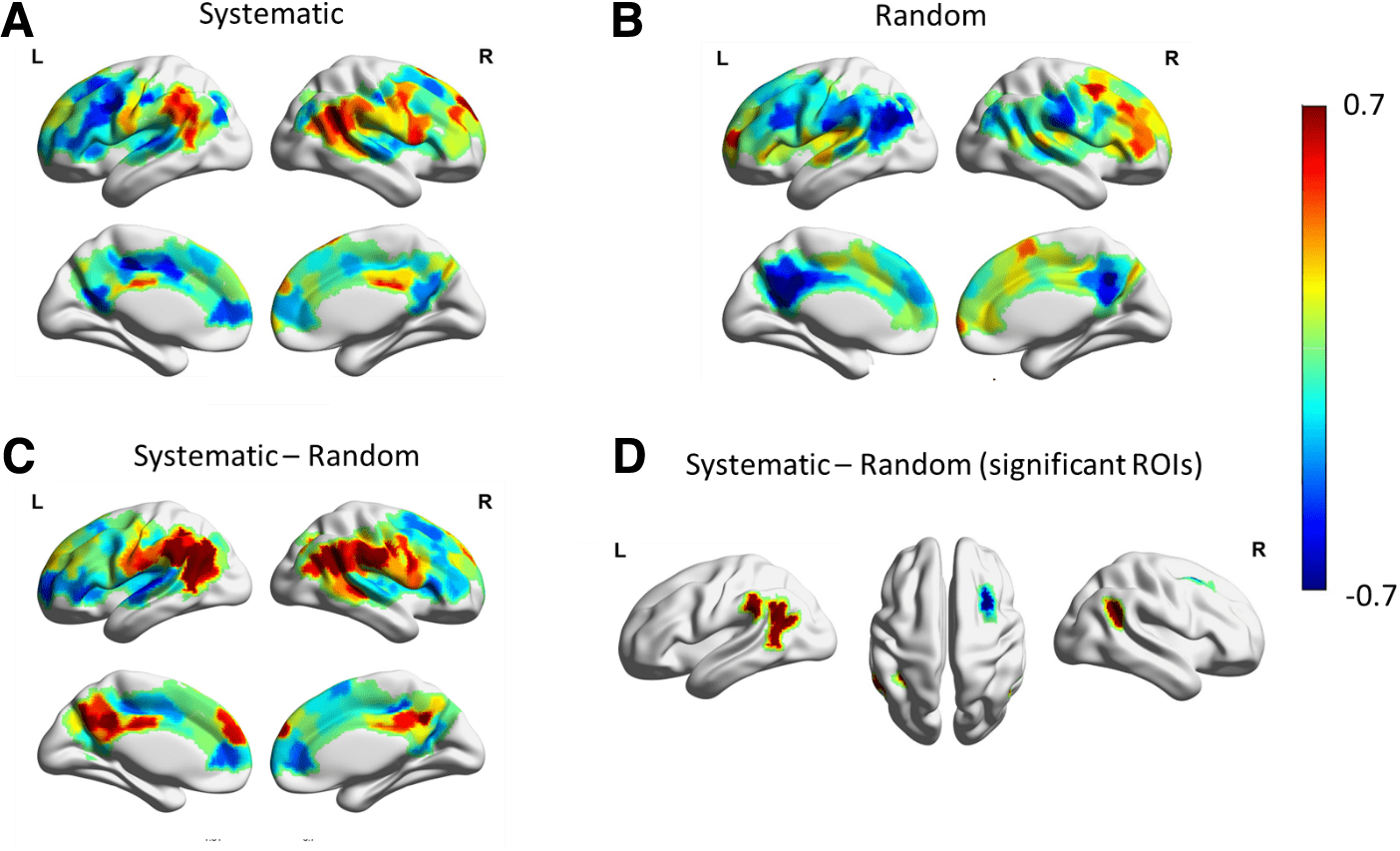
The systematic versus the random visual stimulations
appear to evoke different BOLD responses in several
brain regions. In particular, results (Fig. 5 and Fig. S11)
suggest that both types of visual stimulation sequences
activated the prefrontal lobe. However, the temporoparietal
junctions were activated by the systematic stimuli but
were deactivated by the random stimuli, whereas the
middle frontal gyrus was deactivated by systematic stim-
uli but was activated by random stimuli. Additionally, sys-
tematic stimuli significantly deactivated the ventral
regions in the posterior cingulate cortex (PCC)-precentral
gyrus of the posteromedial cortex whereas the random
stimuli significantly deactivated the default regions in
PCC-precuneus.
More specifically, six brain parcels demonstrated sig-
nificant averaged systematic-random contrast (p-value
< 0.05, as shown in Fig. 5, in which the averaged peak
value of BOLD responses for the two visual conditions,
as well as for their contrast are demonstrated). Five of
them have positive contrast values and all lie in the bilat-
eral temporoparietal junctions spanning across the ven-
tral attention and default networks. On average, these
parcels have strong positive BOLD responses to system-
atic stimuli (e.g., among the top 40% parcels with posi-
tive mean of BOLD peaks), but have strong negative
responses to random stimuli (e.g., among the bottom
30.8% parcels with negative mean of BOLD peaks). On
the other hand, one brain parcel with significant negative
systematic-random contrast (z-score < −1.96) is located
at the right dorsolateral prefrontal cortex (in the frontopa-
rietal network). On average, this parcel has strong nega-
tive BOLD responses to systematic stimuli (i.e., at the
bottom 19.44% of parcels with negative mean peak val-
ues), but has strong positive BOLD responses to random
stimuli (i.e., at the top 6.58% of all positive mean peaks).
Among these six parcels, the three task-negative parcels
(the three default network parcels) also demonstrate a
much more depressed amplitude in averaged QPPs by
random stimulations than by systematic stimulations
Fig. 4. Violin plot of QPP intervals and QPP time delays. (A) Distribution of QPP intervals for all groups for each visual
condition. Here, the QPP intervals are contrasted between the two stimulation sequences. (B) Distribution of QPP time
delay followed by visual stimuli for all groups in systematic stimulation sequences. Here, the distribution of QPP time delay
in each visual condition is contrasted to its null model (derived from the resting data as described in Section 2.3). The t-test
statistics, degree of freedom (noted as t(df)), and p-values are reported above each pair of violin distributions. Please refer
to the right panel of Figure S8 for a thorough explanation of all marks present in the violin plot.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
11
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
Fig. 5. Averaged BOLD peak values in response to systematic and random visual stimulation. See Figure 2A for the
calculation of a stimulated BOLD peak value. (A) The average of BOLD peak values in response to the systematic stimuli.
(B) The average of BOLD peaks in response to random stimuli. Parcels with significant (|z-score| > 1.96) BOLD peak
response for systematic and for random are shown in Figure S11. (C) The average of systematic-random peak contrast
for all selected ROIs. (D) The average of systematic-random peak contrast parcels with significant averaged contrast
(|z-score| > 1.96). Note that the light gray areas in the brain maps are non-covered regions (see Fig. S3 for the covered
and non-covered brain regions in the analysis).
(Fig. S12), which is consistent with the sign of systematic-
random contrast in the BOLD response peaks. However,
opposite to the systematic-random contrast in the BOLD
response peaks, the three task-positive parcels (the two
ventral attention parcels as well as the right dorsolateral
prefrontal parcel) demonstrate a much more elevated
QPP amplitude by random stimulations (Fig. S12).
In addition, the BOLD responses in task-positive and
task-negative networks (including the dorsal attention,
ventral attention, frontoparietal, and default networks)
as well as in the somatomotor network are found to be
dominated by the waveform of the overlapping ongoing
QPPs. The averaged BOLD responses to stimuli, which
were presented at different phases of QPP for each par-
cel, are organized in the five networks. There are 70.56%
of systematic stimuli and 69.84% of random stimuli
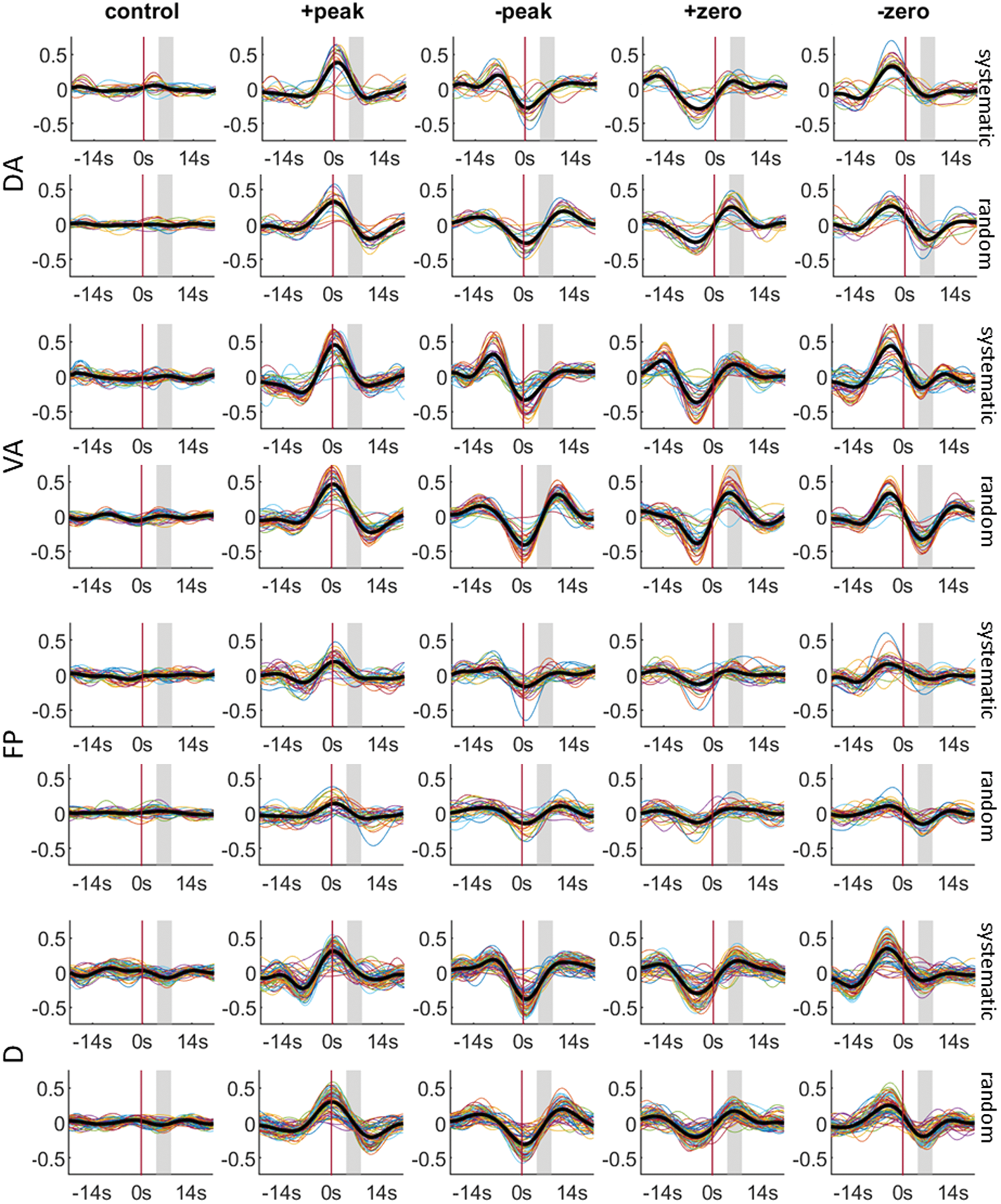
overlapping with ongoing QPPs. As shown in Figure 6
and Figure S15, for both visual stimulation conditions,
the BOLD response to the checkerboard is swamped by
the ongoing QPP signals no matter which of the four
QPP phases overlap. For example, the stimulation onset
in the “+peak” (“−peak”) range will follow by a down
tread (uprising) BOLD response. In contrast, when the
stimulation onset does not meet the ongoing QPPs, an
average with more moderate BOLD responses appeared,
which covers 29.44% or 30.16% of the entire system-
atic or random stimuli. The averaged BOLD responses
of the six brain regions with a significant contrast
between the systematic and random conditions were
also linked to the four QPP phases. As illustrated in Fig-
ure S13, both positive and negative contrast values
between systematic and random conditions were pri-
marily related to the comparison of ongoing QPPs in
each visual condition.
4. DISCUSSION
The dynamics of intrinsic brain activity can be captured
by several quasi-periodic spatiotemporal patterns (QPPs)
(Bolt et al., 2022; Yousefi & Keilholz, 2021). The primary
QPP captures the major dynamics of infraslow intrinsic
neural activity (Grooms et al., 2017; Thompson et al.,
2014), which is known to be involved in attention (Helps
et al., 2010; Monto et al., 2008) and arousal (Raut et al.,
2021; Sihn & Kim, 2022). The interaction between the
ongoing primary QPPs in the brain and visual stimula-
tions was investigated in this study. More specifically, we
investigated how different sequences of visual stimuli
affect the primary QPP in awake humans, and how spon-
taneous QPP prior to each stimulus modifies the subse-
quent visually evoked BOLD response. Two different
types of stimulation sequences induced by flickering
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
12
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
Fig. 6. BOLD responses of systematic (upper) and random (bottom) stimulation patterns that are associated with four
QPP phases for each of the task-positive and task-negative networks. The control presented in the 1st column includes
BOLD responses with no intrinsic primary QPP and the reverse-phase QPP. In each plot, the colorful lines represent the
BOLD signal of each parcel within the network, while the bold black line represents the average of all the parcels within
the network. The vertical axis represents the magnitude of BOLD response whereas the horizontal axis represents the time
interval before and after the stimulus onset at 0 s—depicted by the red vertical line. The gray-shaded area in each plot
depicts the peak range [6TR, 12TR] of the hemodynamic response.
checkerboard were presented to the subjects, a system-
atic stimulation sequence in which the visual stimulus
appeared every 20.3 s and a random stimulation
sequence which has the visual stimulus occurring ran-
domly every 14~62.3 s. Finally, the results of the two
types of stimulation sequences were contrasted to the
resting-state results, which were then compared with
each other.
Due to the limited brain coverage resulting from the
use of a short TR and single-shot gradient EPI, this study
mainly examined the interaction between visual stimula-
tion and the default, task-positive, and somatomotor net-
works. This is because flickering checkerboard visual
stimuli have been shown to activate and deactivate numer-
ous regions throughout the brain, not just in the visual
system (Jorge et al., 2018). In our study, the flickering
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
13
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
checkerboard stimuli were designed to have minimal
impact on arousal fluctuations, but there may be a slight
increase in arousal due to uncertainty (Critchley et al.,
2001; Ramsøy et al., 2012; Urai et al., 2017; Zhao et al.,
2019) surrounding the random stimulus intervals. While
the sequence of visual stimulation is consistent across all
systematic scans, it differs across all random scans.
Although the designed stimulation sequences may intro-
duce increased variability in arousal levels in the random
condition, the standard deviation of QPP correlations in
both the group average analysis (see Table S2 bottom)
and the independent group analysis (noted in the caption
of Fig. S7) indicates that the variability of QPP within the
random condition is actually smaller than that within the
systematic condition. Thus, we believe that the effect of
arousal caused by the uncertain stimulus intervals in ran-
dom sequences is minimal, or not captured by the QPP.
Three central observations of this study are discussed
below.
4.1. Sequences of visual stimulation modify the group averaged
QPPs
The QPPs during the systematic visual condition are sig-
nificantly different from the ones during the random visual
condition, which is more similar to the resting QPPs.
These differences are primarily reflected by a phase mod-
ulation. This is consistent with existing literature on high-
er-frequency activity. For example, in a theta frequency
band, the phase of spontaneous oscillations was found
to be significantly modulated by only predictable (or
attended) but not unpredictable (or unattended) visual
stimuli (Busch & VanRullen, 2010). Similarly, in a fre-
quency band of ~8 Hz, the phase coherence was found
to be strengthened by a systematic visual attentional task
(Zareian et al., 2020).
For the random condition, visual stimulation was found
to affect the QPP magnitude at a network level. Specifi-
cally, the QPP magnitude was much elevated in the ven-
tral and dorsal attention networks but attenuated in the
frontoparietal network. The default network QPP was
also more attenuated by random stimuli than systematic
stimuli. Similar magnitude changes in infraslow dynamics
have also been observed in patients with ADHD (Abbas,
Bassil, & Keilholz, 2019; Helps et al., 2010), suggesting
that sustained attention was distracted by random visual
stimuli. More specifically, the systematic sequence may
entrain intrinsic neural oscillations related to generating
expectancies for future events and allocating attention,
while a random presentation of stimuli typically involves a
longer reaction time and may indicate less sustained
attention (e.g., Jones et al., 2002; Lakatos et al., 2008).
However, as the study did not include a perfor-
mance-based measure of attention, these speculations
are based on reverse inference and should only serve as
a starting point for further research.
4.2. Visual stimulation affects both the frequency of QPP
occurrence and the BOLD response to the stimulus
Even though the visual conditions have exactly the same
number of stimuli, the systematic stimulation produces
more frequent QPPs with significantly shorter consecu-
tive QPP intervals than the random stimulation (Fig. 4A).
However, neither stimulation sequence significantly per-
turbs the onset of QPPs. This seems to contradict the
previous findings in mice that the primary QPPs are more
likely to be triggered at the onset of stimulus (Belloy et al.,
2021). There are several possible reasons for this discrep-
ancy. First, a very different visual stimulation sequence
was employed in Belloy et al. (2021). Particularly, a stim-
ulation of “ON” (30 s) and “OFF” (60 s) cycle that repeats
over time includes a flickering light constantly flashing
at 4 Hz before becoming silent. Second, anesthetized,
instead of awake, mice were studied in Belloy et al. (2021),
and anesthesia is known to affect infraslow brain dynam-
ics (Pan et al., 2013). Finally, this difference may suggest
that the intrinsic QPPs in humans are more robust and
less likely to be disrupted by environmental perturbations
compared to anesthetized mice.
In addition, we also found that the two visual stimula-
tion sequences evoked distinct patterns in the BOLD
response. For example, the bilateral temporoparietal junc-
tion (spanning the ventral attention and the default net-
works) was significantly activated by the systematic
condition but not the random condition (Fig. 5D). This is
consistent with reports that this region is involved in tem-
poral order judgment (Davis et al., 2009) and lack of pre-
dictability in the random condition (Wu et al., 2015). On the
other hand, the right dorsolateral prefrontal region (around
Brodmann area 9) was strongly activated by random stim-
ulation but not systematic stimulation (Fig. 5D), which con-
trasts with the waveform distinctions of the group average
QPP in these two visual conditions. This region has been
linked to working memory, planning, and evaluating
recency, which may be more active in the random than the
systematic condition (Fincham et al., 2002; J. X. Zhang
et al., 2003; Zorrilla et al., 1996). However, one should be
cautious with these reverse inferences due to the signifi-
cant differences between the visual stimuli used in
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
14
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
our study and the tasks employed in previous research.
Relevant to this, a significant deactivation in the default
network in the PCC-precuneus was observed only in the
random condition but not in the systematic condition. This
significant visually-evoked deactivation was also observed
in a previous study using a flickering checkerboard with a
different stimulation sequence (Jorge et al., 2018). One
possible explanation for the increased (decreased) engage-
ment of the dorsolateral prefrontal region (PCC-precuneus)
in the random condition observed in our study is that sub-
jects may be less/more engaged in mind wandering during
the random/systematic condition due to the focus of antic-
ipation of the arrival of stimuli. Consequently, the default
network, known to be activated during mind wandering
(Godwin et al., 2017), becomes more suppressed, while
the dorsolateral prefrontal region becomes more engaged
during random stimulations. Among these 5 parcels, the
activation of the task-negative (default) network regions and
the deactivation of the task-positive network regions appear
to associate with a greater amplitude of group average
QPP, which remains to be investigated in the future.
4.3. The BOLD response is dominated by the QPP waveform when
visual stimulation overlaps with ongoing infraslow brain activity
Flashing checkerboards are prominent visual stimuli
known to produce extensive brain activity well beyond the
visual system (Gonzalez-Castillo et al., 2012; Jorge et al.,
2018). In our specific experiments, across both visual
conditions, nearly 70% of the BOLD stimulus responses
overlap with and are overwhelmed by the waveform of
ongoing primary QPPs. The BOLD response in this set is
significantly greater than the 30% of trials where the stim-
ulation does not overlap. This observation is distinct from
the finding of visually stimulated BOLD response in anes-
thetized mice (Belloy et al., 2021), which observed the
ongoing QPP only moderately affected the magnitude of
subsequent stimulus BOLD (Belloy et al., 2021, Fig. 2D).
This result further confirms our conjecture about the
robustness of intrinsic QPP in awake humans in compari-
son to anesthetized mice. In other words, the dynamics of
the spontaneous infraslow brain activity in the human
brain that supports attention and modulates arousal is
highly robust and less likely to be disrupted by environ-
mental perturbations, though the overall dynamic wave-
form can be perturbed by stimulations in various ways
(discussed in the 2nd paragraph in this section). More-
over, the distinct patterns in the BOLD response to the
different sequences of visual stimulations, demonstrated
by brain parcels with significant averaged systematic-
random contrast (Fig. S13), can also be captured by the
distinctions of evoked ongoing QPPs between the two
visual conditions. Our findings suggest that the intrinsic
QPPs influenced by the flickering checkerboard may also
provide a new explanation for previously reported activa-
tions and deactivations of brain regions located outside of
the visual system (Jorge et al., 2018).
The widely known trial-to-trial variability in stimulated
BOLD responses was popularly examined in a micro-
scopic view in previous studies. A detailed excitation
model is often described based on each stimulus and the
prestimulus baseline (Chen et al., 2020; Fox et al., 2005;
He, 2013; Huang et al., 2017). One influential fMRI study
suggested that the observed BOLD response is a linear
combination of the stimulated response and the prestim-
ulus baseline (Fox et al., 2005). Yet, later works (Chen
et al., 2020; He, 2013; Huang et al., 2017) suggest a non-
additive but inverse modulation between the stimulation
and the prestimulus baseline. Specifically, a higher (lower)
pre-stimulus baseline results in less (more) activation
across widespread human brain regions (Huang et al.,
2017) and rodent brains (Chen et al., 2020).
In these fMRI studies, even though the BOLD
responses in the temporally filtered infraslow frequency
range (Huang et al., 2017) or the broader low-frequency
range (Fox et al., 2005; He, 2013) were studied, their
underlying neurophysiological correlates remain to be
investigated. The pioneering study (Chen et al., 2020)
used concurrent calcium and hemodynamic imaging in
the somatosensory cortical area of anesthetized rats and
found a correlation between the evoked infraslow hemo-
dynamic response and the evoked infraslow neuronal
activity. However, Chen and colleagues also found that
the infraslow hemodynamic power occurred before the
neuronal dynamic power (Chen et al., 2020, Fig. 3e, f),
which implies that a significant portion of the hemody-
namics may not arise from the neuronal level.
Complementing these studies with detailed activation
models, our results explain this trial-to-trial variability
from a macroscopic view. In particular, the varying mag-
nitude of BOLD stimulus responses is largely controlled
by the intrinsic global fluctuations of QPP—a BOLD
dynamic pattern that was found to arise from the infraslow
neural activity. In addition, our results provide novel
insights into these non-additive activation models (Chen
et al., 2020; He, 2013; Huang et al., 2017). Specifically,
due to the sinusoidal nature of primary QPPs (Abbas,
Belloy, et al., 2019; Belloy, Shah, et al., 2018; Yousefi
et al., 2018; Yousefi & Keilholz, 2021) and its window
length of ~20 s, the hemodynamic peak range would
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
15
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
likely fall into a QPP phase right after the QPP phase at
the prestimulus baseline, resulting in an inverse modula-
tion between these two factors.
4.4. Limitation and future study
The primary constraint of our study is the incomplete
brain coverage caused by using a short TR and sin-
gle-shot gradient EPI. This limits our ability to directly
compare findings in non-visual areas to the visual sys-
tem, which is most responsive to the stimuli. Additionally,
while neural-BOLD adaptation to repeated visual stimuli
has been well-observed in the visual cortex (Grill-Spector
et al., 2006; Krekelberg et al., 2006), it is unclear how it
contributes to QPPs among all brain regions. Although
we did not investigate BOLD adaptation in the current
study, any changes in QPPs due to neural adaptation
would be reflected in the overall pattern of QPP, which is
an averaged pattern across all concatenated runs. Future
studies may use 7T multi-echo EPI to verify if all regions
significantly activated or deactivated by visual stimuli are
genuinely caused by intrinsic QPPs and if BOLD adapta-
tion is reflected in the ongoing QPPs over time.
4.5.
Implications for BOLD fMRI
Spontaneous fluctuations in BOLD signals recorded by
fMRI link to the underlying neuronal activity through com-
plex neurovascular coupling. Yet, leveraging the multi-
modal imaging of BOLD and neuronal recordings, various
studies reveal BOLD fluctuations directly reflecting the
dynamics of neural activity in various frequency bands
(Chen et al., 2020; Grooms et al., 2017; Pan et al., 2013;
Raut et al., 2021; Thompson et al., 2014, 2015; X. Zhang
et al., 2020). In particular, BOLD signals preserve rich
information in the infraslow frequency range of brain
activity. This frequency range was initially dismissed as
“noise,” artifact, or epi-phenomena in previous studies of
circuit-level neural activity (Fox & Raichle, 2007), but has
been found to have a unique neurophysiological basis
closely linked to attention (Helps et al., 2010; Monto
et al., 2008) and arousal (Raut et al., 2021; Sihn & Kim,
2022). More specifically, a quasi-period dynamic pattern
(QPP) detected from the infraslow BOLD fluctuations was
found to relate to the infraslow neuronal activity (Chen
et al., 2020; Grooms et al., 2017; Pan et al., 2013; Raut
et al., 2021; Thompson et al., 2014, 2015; X. Zhang et al.,
2020), and also can be affected by attention (Abbas, Bassil,
& Keilholz, 2019; Abbas, Belloy, et al., 2019) and arousal
fluctuations (Raut et al., 2021). Thus, investigating QPP
allows us to infer the dynamics of infraslow neural activ-
ity. This sheds light on understanding the interaction of
environmental perturbation and evoked brain response
that directly ties to the neuronal level using this noninva-
sive imaging technique. In addition, our results reveal that
the QPP waveform is not likely to be disrupted but can
still be affected by visual stimulation in various ways. This
raises the possibility of developing novel non-invasive
sensory stimulation procedures to perturb the dynamics
of infraslow brain activity to enhance attention in humans.
DATA AND CODE AVAILABILITY
The code for processing the partial brain fMRI data is
available at https://github . com / GT – EmoryMINDlab / Partial
HumanBrain _ preprocessing, and the software toolbox
for detecting and analyzing primary QPP is available at
https://github . com / GT – EmoryMINDlab / QPPLab. The fMRI
imaging data is available upon request.
AUTHOR CONTRIBUTIONS
Nan Xu: Conceptualization; data curation; data prepro-
cessing; methodology; formal analysis; writing, original
draft; and writing, review and editing. Derek M. Smith:
Data acquisition; data curation; and writing, review and
editing. George Jeno: Data preprocessing. Dolly T. See-
burger: Data curation. Eric H. Schumacher: Conceptual-
ization; data acquisition; writing, review and editing;
funding acquisition; and supervision. Shella D. Keilholz:
Conceptualization; methodology; writing, review and
editing; funding acquisition; and supervision.
DECLARATION OF COMPETING INTEREST
The authors declare that they have no situation of real,
potential, or apparent conflict of interest and that there is
no financial/personal interest or belief that could affect
their objectivity.
ACKNOWLEDGMENTS
All the authors thank the National Science Foundation
(NSF grant 1533260) for funding support. Nan Xu and
Shella D. Keilholz thank the National Institutes of Health
(NIH grant R01NS078095) for funding support. Nan Xu
also thanks the National Institutes of Health (NIH grant
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
16
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
1K99NS123113) for funding support. Derek M. Smith
would like to thank the Therapeutic Cognitive Neuroscience
Fund. The authors also thank Dr. Ying Guo for her helpful
discussions.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available with
the online version here: https://doi.org/10.1162/imag_a_
00002.
REFERENCES
Abbas, A., Bassil, Y., & Keilholz, S. (2019). Quasi-periodic
patterns of brain activity in individuals with attention-
deficit/hyperactivity disorder. Neuroimage: Clinical, 21,
101653. https://doi . org / 10 . 1016 / j . nicl . 2019 . 101653
Abbas, A., Belloy, M., Kashyap, A., Billings, J., Nezafati, M.,
Schumacher, E. H., & Keilholz, S. (2019). Quasi-periodic
patterns contribute to functional connectivity in the brain.
Neuroimage, 191, 193–204. https://doi . org / 10 . 1016 / j
. neuroimage . 2019 . 01 . 076
Belloy, M. E., Billings, J., Abbas, A., Kashyap, A., Pan,
W.-J., Hinz, R., Vanreusel, V., Van Audekerke, J., Van der
Linden, A., Keilholz, S. D., Verhoye, M., & Keliris, G. A.
(2021). Resting brain fluctuations are intrinsically coupled
to visual response dynamics. Cerebral Cortex, 31(3),
1511–1522. https://doi . org / 10 . 1093 / cercor / bhaa305
Belloy, M. E., Naeyaert, M., Abbas, A., Shah, D., Vanreusel,
V., van Audekerke, J., Keilholz, S. D., Keliris, G. A., Van
der Linden, A., & Verhoye, M. (2018). Dynamic resting
state fMRI analysis in mice reveals a set of quasi-
periodic patterns and illustrates their relationship with
the global signal. Neuroimage, 180, 463–484. https://doi
. org / 10 . 1016 / j . neuroimage . 2018 . 01 . 075
Belloy, M. E., Shah, D., Abbas, A., Kashyap, A., Roßner,
S., Van Der Linden, A., Keilholz, S. D., Keliris, G. A., &
Verhoye, M. (2018). Quasi-periodic patterns of neural
activity improve classification of Alzheimer’s disease in
mice. Scientific Reports, 8(1), 10024. https://doi . org / 10
. 1038 / s41598 – 018 – 28237 – 9
Bolt, T., Nomi, J. S., Bzdok, D., Salas, J. A., Chang, C.,
Thomas Yeo, B. T., Uddin, L. Q., & Keilholz, S. D. (2022).
A parsimonious description of global functional brain
organization in three spatiotemporal patterns. Nature
Neuroscience, 25(8), 1093–1103. https://doi . org / 10 . 1038
/ s41593 – 022 – 01118 – 1
Busch, N. A., & VanRullen, R. (2010). Spontaneous EEG
oscillations reveal periodic sampling of visual attention.
Proceedings of the National Academy of Sciences of the
United States of America, 107(37), 16048–16053. https://
doi . org / 10 . 1073 / PNAS . 1004801107 / SUPPL _ FILE / PNAS
. 201004801SI . PDF
Chen, W., Park, K., Pan, Y., Koretsky, A. P., & Du, C. (2020).
Interactions between stimuli-evoked cortical activity and
spontaneous low frequency oscillations measured with
neuronal calcium. Neuroimage, 210, 116554. https://doi
. org / 10 . 1016 / J . NEUROIMAGE . 2020 . 116554
Cox, R. W. (1996). AFNI: Software for analysis and
visualization of functional magnetic resonance
neuroimages. Computers and Biomedical Research,
29(3), 162–173. https://doi . org / 10 . 1006 / cbmr . 1996 . 0014
Cox, R. W., & Hyde, J. S. (1997). Software tools for analysis
and visualization of fMRI data. NMR in Biomedicine,
10(4–5), 171–178. https://doi . org / 10 . 1002 / (SICI)1099
– 1492(199706 / 08)10:4 / 5<171::AID - NBM453>3 . 0 . CO;2 – L
Critchley, H. D., Mathias, C. J., & Dolan, R. J. (2001). Neural
activity in the human brain relating to uncertainty and
arousal during anticipation. Neuron, 29(2), 537–545.
https://doi . org / 10 . 1016 / S0896 – 6273(01)00225 – 2
Dale, A. M., & Buckner, R. L. (1997). Selective averaging
of rapidly presented individual trials using fMRI. Human
Brain Mapping, 5, 329–340. https://onlinelibrary . wiley
. com / doi / 10 . 1002 / (SICI)1097 – 0193(1997)5:5%3C329::AID
– HBM1%3E3 . 0 . CO;2 – 5
Davis, B., Christie, J., & Rorden, C. (2009). Temporal order
judgments activate temporal parietal junction. The
Journal of Neuroscience, 29(10), 3182. https://doi . org / 10
. 1523 / JNEUROSCI . 5793 – 08 . 2009
Diedenhofen, B., & Musch, J. (2015). cocor: A
comprehensive solution for the statistical comparison of
correlations. PLoS One, 10(4), e0121945. https://doi . org
/ 10 . 1371 / JOURNAL . PONE . 0121945
Ding, J., Sperling, G., & Srinivasan, R. (2006). Attentional
modulation of SSVEP power depends on the network
tagged by the flicker frequency. Cerebral Cortex, 16(7),
1016–1029. https://doi . org / 10 . 1093 / CERCOR / BHJ044
Duann, J. R., Jung, T. P., Kuo, W. J., Yeh, T. C., Makeig,
S., Hsieh, J. C., & Sejnowski, T. J. (2002). Single-trial
variability in event-related BOLD signals. Neuroimage,
15(4), 823–835. https://doi . org / 10 . 1006 / NIMG . 2001 . 1049
Engel, S. A., Glover, G. H., & Wandell, B. A. (1997).
Retinotopic organization in human visual cortex and the
spatial precision of functional MRI. Cerebral Cortex, 7(2),
181–192. https://doi . org / 10 . 1093 / CERCOR / 7 . 2 . 181
Fincham, J. M., Carter, C. S., Van Veen, V., Stenger, V. A., &
Anderson, J. R. (2002). Neural mechanisms of planning:
A computational analysis using event-related fMRI.
Proceedings of the National Academy of Sciences of the
United States of America, 99(5), 3346–3351. https://doi
. org / 10 . 1073 / PNAS . 052703399
Fox, M. D., & Raichle, M. E. (2007). Spontaneous
fluctuations in brain activity observed with functional
magnetic resonance imaging. Nature Reviews
Neuroscience, 8(9), 700–711. https://doi . org / 10 . 1038
/ nrn2201
Fox, M. D., Snyder, A. Z., Zacks, J. M., & Raichle, M. E.
(2005). Coherent spontaneous activity accounts for
trial-to-trial variability in human evoked brain responses.
Nature Neuroscience, 9(1), 23–25. https://doi . org / 10
. 1038 / nn1616
Godwin, C. A., Hunter, M. A., Bezdek, M. A., Lieberman, G.,
Elkin-Frankston, S., Romero, V. L., Witkiewitz, K., Clark,
V. P., & Schumacher, E. H. (2017). Functional connectivity
within and between intrinsic brain networks correlates
with trait mind wandering. Neuropsychologia, 103, 140–
153. https://doi . org / 10 . 1016 / J . NEUROPSYCHOLOGIA
. 2017 . 07 . 006
Gonzalez-Castillo, J., Saad, Z. S., Handwerker, D. A., Inati,
S. J., Brenowitz, N., & Bandettini, P. A. (2012). Whole-
brain, time-locked activation with simple tasks revealed
using massive averaging and model-free analysis.
Proceedings of the National Academy of Sciences of the
United States of America, 109(14), 5487–5492. https://
www . pnas . org / action / downloadSupplement ? doi = 10
. 1073%2Fpnas . 1121049109 & file = pnas . 201121049si . pdf
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
17
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
Grill-Spector, K., Henson, R., & Martin, A. (2006). Repetition
and the brain: Neural models of stimulus-specific effects.
Trends in Cognitive Sciences, 10(1), 14–23. https://doi
. org / 10 . 1016 / J . TICS . 2005 . 11 . 006
Grooms, J. K., Thompson, G. J., Pan, W. J., Billings, J.,
Schumacher, E. H., Epstein, C. M., & Keilholz, S. D.
(2017). Infraslow electroencephalographic and dynamic
resting state network activity. Brain Connectivity, 7(5),
265. https://doi . org / 10 . 1089 / BRAIN . 2017 . 0492
He, B. J. (2013). Spontaneous and task-evoked brain
activity negatively interact. Journal of Neuroscience,
33(11), 4672–4682. https://doi . org / 10 . 1523 / JNEUROSCI
. 2922 – 12 . 2013
Helps, S. K., Broyd, S. J., James, C. J., Karl, A., Chen, W.,
& Sonuga-Barke, E. J. S. (2010). Altered spontaneous
low frequency brain activity in attention deficit/
hyperactivity disorder. Brain Research, 1322, 134–143.
https://doi . org / 10 . 1016 / j . brainres . 2010 . 01 . 057
Huang, Z., Zhang, J., Longtin, A., Dumont, G., Duncan,
N. W., Pokorny, J., Qin, P., Dai, R., Ferri, F., Weng, X.,
& Northoff, G. (2017). Is there a nonadditive interaction
between spontaneous and evoked activity? Phase-
dependence and its relation to the temporal structure of
scale-free brain activity. Cerebral Cortex, 27(2), 1037–
1059. https://doi . org / 10 . 1093 / CERCOR / BHV288
Jenkinson, M., Bannister, P., Brady, M., & Smith, S. (2002).
Improved optimization for the robust and accurate linear
registration and motion correction of brain images.
Neuroimage, 17(2), 825–841. https://doi . org / 10 . 1006
/ NIMG . 2002 . 1132
Jenkinson, M., Beckmann, C. F., Behrens, T. E. J., Woolrich,
M. W., & Smith, S. M. (2012). FSL. Neuroimage, 62(2),
782–790. https://doi . org / 10 . 1016 / j . neuroimage . 2011 . 09
. 015
Jones, M. R., Moynihan, H., MacKenzie, N., & Puente, J.
(2002). Temporal aspects of stimulus-driven attending in
dynamic arrays. Psychological Science, 13(4), 313–319.
https://doi . org / 10 . 1111 / 1467 – 9280 . 00458
Jorge, J., Figueiredo, P., Gruetter, R., & van der Zwaag,
W. (2018). Mapping and characterization of positive
and negative BOLD responses to visual stimulation in
multiple brain regions at 7T. Human Brain Mapping,
39(6), 2426–2441. https://doi . org / 10 . 1002 / HBM . 24012
Krekelberg, B., Boynton, G. M., & van Wezel, R. J. A.
(2006). Adaptation: From single cells to BOLD signals.
Trends in Neurosciences, 29(5), 250–256. https://doi . org
/ 10 . 1016 / J . TINS . 2006 . 02 . 008
Lakatos, P., Karmos, G., Mehta, A. D., Ulbert, I., &
Schroeder, C. E. (2008). Entrainment of neuronal
oscillations as a mechanism of attentional selection.
Science, 320(5872), 110–113. https://doi . org / 10 . 1126
/ SCIENCE . 1154735 / SUPPL _ FILE / LAKATOS . SOM . PDF
Majeed, W., Magnuson, M., Hasenkamp, W., Schwarb,
H., Schumacher, E. H., Barsalou, L., & Keilholz, S. D.
(2011). Spatiotemporal dynamics of low frequency BOLD
fluctuations in rats and humans. Neuroimage, 54(2),
1140–1150. https://doi . org / 10 . 1016 / j . neuroimage . 2010
. 08 . 030
Majeed, W., Magnuson, M., & Keilholz, S. D. (2009).
Spatiotemporal dynamics of low frequency fluctuations
in BOLD fMRI of the rat. Journal of Magnetic Resonance
Imaging, 30(2), 384–393. https://doi . org / 10 . 1002 / jmri
. 21848
Mitra, A., Kraft, A., Wright, P., Acland, B., Snyder, A.
Z., Rosenthal, Z., Czerniewski, L., Bauer, A., Snyder,
L., Culver, J., Lee, J. M., & Raichle, M. E. (2018).
Spontaneous infra-slow brain activity has unique
spatiotemporal dynamics and laminar structure. Neuron,
98(2), 297.e6–305.e6. https://doi . org / 10 . 1016 / j . neuron
. 2018 . 03 . 015
Monto, S., Palva, S., Voipio, J., & Palva, J. M. (2008). Very
slow EEG fluctuations predict the dynamics of stimulus
detection and oscillation amplitudes in humans. Journal
of Neuroscience, 28(33), 8268–8272. https://doi . org / 10
. 1523 / JNEUROSCI . 1910 – 08 . 2008
Pan, W.-J., Thompson, G. J., Magnuson, M. E., Jaeger, D.,
& Keilholz, S. (2013). Infraslow LFP correlates to resting-
state fMRI BOLD signals. Neuroimage, 74(0), 288–297.
https://doi . org / 10 . 1016 / j . neuroimage . 2013 . 02 . 035
Qiao, J., Li, X., Wang, Y., Wang, Y., Li, G., Lu, P., & Wang, S.
(2022). The infraslow frequency oscillatory transcranial
direct current stimulation over the left dorsolateral
prefrontal cortex enhances sustained attention. Frontiers
in Aging Neuroscience, 14, 879006. https://doi . org / 10
. 3389 / FNAGI . 2022 . 879006
Ramsøy, T. Z., Friis-Olivarius, M., Jacobsen, C., Jensen, S.
B., & Skov, M. (2012). Effects of perceptual uncertainty on
arousal and preference across different visual domains.
Journal of Neuroscience, Psychology, and Economics,
5(4), 212–226. https://doi . org / 10 . 1037 / A0030198
Raut, R. V., Snyder, A. Z., Mitra, A., Yellin, D., Fujii, N.,
Malach, R., & Raichle, M. E. (2021). Global waves
synchronize the brain’s functional systems with
fluctuating arousal. Science Advances, 7(30), eabf2709.
https://doi . org / 10 . 1126 / SCIADV . ABF2709 / SUPPL _ FILE
/ SCIADV . ABF2709 _ SM . PDF
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O.,
Zuo, X.-N., Holmes, A. J., Eickhoff, S. B., & Yeo, B.
T. T. (2018). Local-global parcellation of the human
cerebral cortex from intrinsic functional connectivity MRI.
Cerebral Cortex, 28(9), 3095–3114. https://doi . org / 10
. 1093 / cercor / bhx179
Schwartz, S., Vuilleumier, P., Hutton, C., Maravita, A.,
Dolan, R. J., & Driver, J. (2005). Attentional load and
sensory competition in human vision: Modulation of
fMRI responses by load at fixation during task-irrelevant
stimulation in the peripheral visual field. Cerebral Cortex,
15(6), 770–786. https://doi . org / 10 . 1093 / CERCOR / BHH178
Sedlacek, M., & Krumpholc, M. (2005). Digital measurement
of phase difference—A comparative study of DSP
algorithms. Metrology and Measurement Systems, 12(4),
427–448.
Sihn, D., & Kim, S. P. (2022). Brain infraslow activity
correlates with arousal levels. Frontiers in Neuroscience,
16, 230. https://doi . org / 10 . 3389 / FNINS . 2022 . 765585
/ BIBTEX
Thomas Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu,
M. R., Lashkari, D., Hollinshead, M., Roffman, J. L.,
Smoller, J. W., Zöllei, L., Polimeni, J. R., Fisch, B., Liu,
H., Buckner, R. L., Fischl, B., Liu, H., Buckner, R. L.,
Yeo, B. T., Krienen, F. M., Sepulcre, J., … Buckner, R. L.
(2011). The organization of the human cerebral cortex
estimated by intrinsic functional connectivity. Journal of
Neurophysiology, 106(3), 1125–1165. https://doi . org / 10
. 1152 / jn . 00338 . 2011
Thompson, G. J., Pan, W. J., & Keilholz, S. D. (2015).
Different dynamic resting state fMRI patterns are linked
to different frequencies of neural activity. Journal of
Neurophysiology, 114(1), 114–124. https://doi . org / 10
. 1152 / jn . 00235 . 2015
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
18
N. Xu, D.M. Smith, G. Jeno et al.
Imaging Neuroscience, Volume 1, 2023
Thompson, G. J., Pan, W. J., Magnuson, M. E., Jaeger, D.,
& Keilholz, S. D. (2014). Quasi-periodic patterns (QPP):
Large-scale dynamics in resting state fMRI that correlate
with local infraslow electrical activity. Neuroimage, 84,
1018–1031. https://doi . org / 10 . 1016 / j . neuroimage . 2013
. 09 . 029
Zareian, B., Maboudi, K., Daliri, M. R., Abrishami
Moghaddam, H., Treue, S., & Esghaei, M. (2020).
Attention strengthens across-trial pre-stimulus phase
coherence in visual cortex, enhancing stimulus
processing. Scientific Reports, 10(1), 1–12. https://doi
. org / 10 . 1038 / s41598 – 020 – 61359 – 7
Tootell, R. B. H., Hadjikhani, N., Hall, E. K., Marrett, S.,
Vanduffel, W., Vaughan, J. T., & Dale, A. M. (1998).
The retinotopy of visual spatial attention. Neuron,
21(6), 1409–1422. https://doi . org / 10 . 1016 / S0896
– 6273(00)80659 – 5
Urai, A. E., Braun, A., & Donner, T. H. (2017). Pupil-linked
arousal is driven by decision uncertainty and alters
serial choice bias. Nature Communications, 8(1), 14637.
https://doi . org / 10 . 1038 / ncomms14637
Zhang, J. X., Leung, H. C., & Johnson, M. K. (2003). Frontal
activations associated with accessing and evaluating
information in working memory: An fMRI study.
Neuroimage, 20(3), 1531–1539. https://doi . org / 10 . 1016 / j
. neuroimage . 2003 . 07 . 016
Zhang, X., Pan, W. J., & Keilholz, S. D. (2020). The
relationship between BOLD and neural activity arises from
temporally sparse events. Neuroimage, 207, 116390.
https://doi . org / 10 . 1016 / j . neuroimage . 2019 . 116390
Wald, L. L., & Polimeni, J. R. (2015). High-speed, high-
Zhang, Y., Brady, M., & Smith, S. (2001). Segmentation of
resolution acquisitions. Brain Mapping: An Encyclopedic
Reference, 1, 103–116. https://doi . org / 10 . 1016 / B978 – 0
– 12 – 397025 – 1 . 00011 – 7
Wu, Q., Chang, C. F., Xi, S., Huang, I. W., Liu, Z., Juan, C. H.,
Wu, Y., & Fan, J. (2015). A critical role of temporoparietal
junction in the integration of top-down and bottom-up
attentional control. Human Brain Mapping, 36(11),
4317–4333. https://doi . org / 10 . 1002 / HBM . 22919
Yousefi, B., & Keilholz, S. (2021). Propagating patterns of
intrinsic activity along macroscale gradients coordinate
functional connections across the whole brain.
Neuroimage, 231, 117827. https://doi . org / 10 . 1016 / j
. neuroimage . 2021 . 117827
brain MR images through a hidden Markov random field
model and the expectation-maximization algorithm. IEEE
Transactions on Medical Imaging, 20(1), 45–57. https://
doi . org / 10 . 1109 / 42 . 906424
Zhao, S., Chait, M., Dick, F., Dayan, P., Furukawa, S., &
Liao, H. I. (2019). Pupil-linked phasic arousal evoked by
violation but not emergence of regularity within rapid
sound sequences. Nature Communications, 10(1), 1–16.
https://doi . org / 10 . 1038 / s41467 – 019 – 12048 – 1
Zhivomirov, H. (2022). Phase difference measurement with
Matlab. MATLAB Central File Exchange. https://www
. mathworks . com / matlabcentral / fileexchange / 48025
– phase – difference – measurement – with – matlab
Yousefi, B., Shin, J., Schumacher, E. H., & Keilholz, S. D.
Zorrilla, L. T., Aguirre, G. K., Zarahn, E., Cannon, T. D., &
(2018). Quasi-periodic patterns of intrinsic brain activity
in individuals and their relationship to global signal.
Neuroimage, 167, 297–308. https://doi . org / 10 . 1016 / j
. neuroimage . 2017 . 11 . 043
D’Esposito, M. (1996). Activation of the prefrontal cortex
during judgments of recency: A functional MRI study.
Neuroreport, 7(15–17), 2803–2806. https://doi . org / 10
. 1097 / 00001756 – 199611040 – 00079
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00002/2154702/imag_a_00002.pdf by guest on 09 September 2023
19