The Impact of Early Amygdala Damage on Juvenile
Rhesus Macaque Social Behavior
Eliza Bliss-Moreau, Gilda Moadab, Melissa D. Bauman,
and David G. Amaral
Abstract
■ The present experiments continue a longitudinal study of
rhesus macaque social behavior following bilateral neonatal
ibotenic acid lesions of the amygdala or hippocampus, or
sham operations. Juvenile animals (approximately 1.5–2.5 years)
were tested in four different social contexts—alone, while inter-
acting with one familiar peer, while interacting with one unfamiliar
peer, and in their permanent social groups. During infancy, the
amygdala-lesioned animals displayed more interest in conspecifics
(indexed by increased affiliative signaling) and paradoxically de-
monstrated more submission or fear (Bauman, Lavenex, Mason,
Capitanio, & Amaral, 2004a, this journal). When these animals were
assessed as juveniles, differences were less striking. Amygdala-
lesioned animals generated fewer aggressive and affiliative signals
(e.g., vocalizations, facial displays) and spent less time in social
interactions with familiar peers. When animals were observed
alone or with an unfamiliar peer, amygdala-lesioned animals,
compared with other subjects, spent more time being inactive
and physically explored the environment less. Despite the subtle,
lesion-based differences in the frequency and duration of spe-
cific social behaviors, there were lesion-based differences in the
organization of behavior such that lesion groups could be iden-
tified based on the patterning of social behaviors in a discriminant
function analysis. The findings indicate that, although overall
frequencies of many of the observed behaviors do not differ
between groups, the general patterning of social behavior may
distinguish the amygdala-lesioned animals. ■
INTRODUCTION
Damage to the adult primate amygdala disrupts affective
processing (e.g., Antoniadis, Winslow, Davis, & Amaral,
2007, 2009; Chudasama, Izquierdo, & Murray, 2009;
Machado, Kazama, & Bachevalier, 2009; Mason, Capitanio,
Machado, Mendoza, & Amaral, 2006; Izquierdo, Suda, &
Murray, 2005; Stefanacci, Clark, & Zola, 2003; Meunier,
Bachevalier, Murray, Málková, & Mishkin, 1999; Zola-
Morgan, Squire, Alverez-Royo, & Clower, 1991; Aggleton &
Passingham, 1981), which has consequences for social
behavior when interacting with conspecifics (e.g., Machado,
Emery, et al., 2008; Machado & Bachevalier, 2006; Emery
et al., 2001; Kling, 1974; Mirsky, 1960; Rosvold, Mirsky, &
Pribram, 1954). Changes in social behavior observed in
animals with adult amygdala damage have been hypothe-
sized to result from disruption of danger detection functions
of the amygdala (Amaral, 2006) rather than to an alteration
to social behavior per se. In this view, adult amygdala-
lesioned animals are hypersocial (Machado, Emery, et al.,
2008; Emery et al., 2001) because they fail to process
the potential threat of novel conspecifics. In other words,
they do not demonstrate the species-typical reluctance to
engage a novel conspecific in social interaction before clear
dominance relationships are established. The extent to
University of California, Davis
© 2013 Massachusetts Institute of Technology
which early damage to the amygdala results in alterations
in affective and social processing is less clear. The goal of
the present paper is to evaluate variation in social process-
ing in juvenile animals that received damage to the amyg-
dala as neonates as part of our ongoing study of variation
in affect (Bliss-Moreau, Bauman, & Amaral, 2011; Bliss-
Moreau, Toscano, Bauman, Mason, & Amaral, 2010, 2011)
and social behavior (Bauman, Lavenex, Mason, Capitanio,
& Amaral, 2004a, 2004b) following early amygdala damage.
Previous research from our laboratory (Bauman et al.,
2004a; Prather et al., 2001) and others (Bachevalier, 1994;
Thompson & Towfighi, 1976; Thompson, Schwartzbaum,
& Harlow, 1969; Kling & Green, 1967) demonstrated that
macaques with early damage to the amygdala are able to
generate species-typical social behaviors. In a previous
report in this journal (Bauman et al., 2004a), we reported
variation in social behaviors generated by maternally
reared, group-socialized macaques that received bilateral
neurotoxic lesions (which spare fibers of passage) of the
amygdala or hippocampus, or sham operations at ap-
proximately 2 weeks of age. During these animalsʼ first
year of life (at approximately 6, 9, and 12 months of
age) social behavior was observed in a number of social
conditions, when the subjects were alone, in dyadic inter-
actions (“dyads”) with familiar partners, in dyads with un-
familiar partners, or in groups of animals with which they
were familiar. Although there were no overall differences
Journal of Cognitive Neuroscience 25:12, pp. 2124–2140
doi:10.1162/jocn_a_00483
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
in the amount of time that amygdala-lesioned monkeys
spent interacting with their peers during the first year
of life, there were lesion-based differences in the fre-
quency of their behaviors. In two contexts, while inter-
acting one-on-one with familiar and novel partners,
amygdala-lesioned animals generated more communica-
tive signals related to affiliation (e.g., cooing, grunting,
etc.). The variety of affiliative signaling was particularly
evident when interacting with novel partners; in that con-
text, amygdala-lesioned animals also presented their bodies
to be groomed and mounted more often than controls.
One difference between animals with neonatal as com-
pared with adult amygdala damage was the expression of
apparent fear or submission behavior. During interactions
both with familiar and novel animals, in dyads and social
groups, amygdala-lesioned animals expressed more signs
of fear or submission (e.g., grimacing, screaming, etc.). This
latter observation stands in contrast to the typical pattern
of social behaviors generated by animals that receive amyg-
dala damage as adults (i.e., less signaling of fear; Machado,
Emery, et al., 2008; Emery et al., 2001). The goal of this
study was to continue the evaluation of juvenile macaque
social behavior following neonatal damage to the amygdala
or hippocampus, at a later developmental point during
which the animals were living in a more socially enriched
environment. In the time between the social behavior
experiment in the previous report (Bauman et al., 2004a)
and the experiments presented in this article, the subjects
were housed 24 hr per day in social groups with the animals
with which they had previously been socialized. Subjects
lived in these permanent social groups for approximately
3 months before the start of the present experiments.
METHODS
All experimental procedures were developed in consulta-
tion with the veterinary staff at the California National
Primate Research Center. All protocols were approved
by the University of California-Davis Institutional Animal
Care and Use Committee.
Animals and Living Conditions
Subject selection and rearing history has been fully de-
scribed in other publications (Bliss-Moreau, Toscano,
et al., 2011; Bliss-Moreau et al., 2010; Bauman et al.,
2004a, 2004b). Briefly, 24 juvenile rhesus macaque mon-
keys received bilateral ibotenic acid lesions of either the
amygdala (five females, three males) or hippocampus (five
females, three males), or sham control operations (four
females, four males) at 12–16 days of age. After surgery,
animals were returned to their mothers and housed
alone with their mothers in standard primate caging
(61 cm W × 66 cm D × 81 cm H). Following recovery,
subjects and their mothers were socialized with other
subjects and other mothers in large chain-link indoor
enclosures (2.13 m W × 3.35 m D × 2.44 m H) for 3 hr,
5 days per week. Each social group included six subjects
(two from each experimental condition) and an adult
male. Subjects were weaned and separated from their
mothers at 6 months of age, singly housed, but were so-
cialized in their groups without their mothers for 3 hr
each day. At this time, a novel adult female was added
to each social group. Subjects were permanently housed
(24 hr per day) with their social groups (i.e., peers and
the adult male and female) in the large enclosures begin-
ning at 1 year of age. The experiments described in this
article occurred while subjects were living in their perma-
nent social groups.
Indoor housing rooms were maintained on a 12-hr
light/dark cycle (lights on at 6 a.m.). Animals were fed
monkey chow (Lab Diet #5047, PMI Nutrition Interna-
tional, Inc., Brentwood, MO) twice daily, provided with
fresh fruit and vegetables twice per week, and had access
to water ad libitum.
One of the original amygdala-lesioned males died of
causes unrelated to his lesion status at approximately
1 year of age (Bauman et al., 2004a). He was replaced by
another male that underwent amygdala lesion surgery at
the same time as the present cohort. That subject was
reared by his mother for the first year of life and pair-
housed with an age-matched female after being weaned
at 1 year. He was introduced to his social group at 1 year
and 3 months of age.
Surgical Procedures
The surgical procedures have been described in detail in
previous publications (Bauman et al., 2004a, 2004b) and
are briefly summarized here. Each subjectʼs brain was
imaged on the morning before surgery to determine the
stereotaxic coordinates of the amygdala or hippocampus
for subsequent ibotenic acid injections. Subjects were
anesthetized with ketamine hydrochloride (15 mg/kg im)
and medatomidine (30 μg/kg) before being placed in an
MRI-compatible stereotaxic apparatus (Crist Instruments
Co., Inc., Damascus, MD). Brain imagining occurred on a
General Electric 1.5 T Gyroscan magnet with the following
parameters: slice thickness = 1.0 mm, T1-weighted Inver-
sion Recovery Pulse sequence, repetition time = 21, echo
time = 7.9, NEX 3, field of view = 8 cm, matrix 256 × 256.
Following the MRI, subjects were intubated so that
they could be ventilated during surgery. Subjects were
anesthetized with a combination of isoflurane (1.0%, var-
ied as needed to maintain an adequate level of anesthesia)
and intravenous infusion of fentanyl (7–10 μg/kg/hr).
Each operated subject received two craniotomies over
the left and right amygdala or hippocampus. Ibotenic
acid (IBO, Biosearch Technologies, Inc., 10 mg/ml in
0.1 M phosphate-buffered saline) was injected simulta-
neously bilaterally into the amygdala or hippocampus
using 10-μl Hamilton syringes (26-gauge beveled needles)
at a rate of 0.2 μl/min. Sham-operated controls underwent
the same presurgical preparations, received a midline
Bliss-Moreau et al.
2125
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
incision to expose the skull, and were maintained under
anesthesia for the average duration of the lesion sur-
geries. Following the surgical procedure, all infants were
monitored by a veterinarian and returned to their mothers
once they were fully alert.
frequency and duration of species typical behaviors (see
Table 1). There were three observers who were blind to
lesion conditions and had an interobserver reliability of
greater than 90%.
Lesion Analysis
This study is longitudinal, and therefore, the subjects
for this experiment continue to be tested and have
not been euthanized to complete histological analysis
of their lesions. Lesion placement was confirmed via
(1) T2-weighted MR images acquired 10 days after surgery,
(2) T1-weighted images acquired when the animals were
approximately 4 years (Machado, Snyder, Cherry, Lavenex,
& Amaral, 2008), and (3) histological analysis of the one
amygdala-lesioned subject who died during his first year
of life. First, edema associated with the brain lesions was
measured using T2-weighted MR images collected 10 days
postsurgery using a General Electric 1.5 T Gyroscan
magnet (slice thickness = 1.5 mm, T2 weighted Inversion
Recovery Pulse sequence, repetition time = 4000, echo
time = 102, NEX 3, field of view = 8 cm, matrix 256 ×
256). The hyperintense T2-weighted signal for each of
the 16 lesion animals (eight amygdala lesion, eight hip-
pocampus lesion) was evaluated to confirm the general
target and extent of the lesions (i.e., amygdala lesion
sparing the hippocampus or hippocampus lesion sparing
the amygdala). T2-weighted images of coronal sections
through the middle portion of the amygdala are illustrated
in previous publications (Bliss-Moreau, Bauman, et al.,
2011; Bauman et al., 2004a, 2004b), indicating that the
ibotenic acid was injected into and caused damage to
the amygdala or hippocampus. Second, lesion extent was
further characterized in T1-weight MRI images when ani-
mals were 4 years (Machado, Snyder, et al., 2008). Finally,
the extent of the targeted lesion was confirmed using his-
tological evaluation in the one amygdala-lesioned animal
that died because of an unrelated illness.
Experimental Design and Procedures
Behavioral Sampling Procedure
Experimental Test Cage
All observations occurred in one of the four large group
test cages in which the animals were permanently housed
(2.13 m W × 3.35 m D × 2.44 m H). The test cages were
constructed of chain link on three sides (2.13 m wide front
and back, as well as the top) and aluminum panels on the
left and right sides (3.35 m deep). Animals entered and
exited the test cages via an entry tunnel at the back of
the cage made of 1 in. metal mesh. Solo observations,
familiar dyads, and social group observations occurred
in the test cage in which the animals lived permanently.
Animals not being observed were relocated to temporary
caging in the same housing room. Novel dyad observa-
tions occurred in one of the four test cages with which
the animals were not familiar (i.e., a cage in which they
did not live).
Behavioral Observation Timing
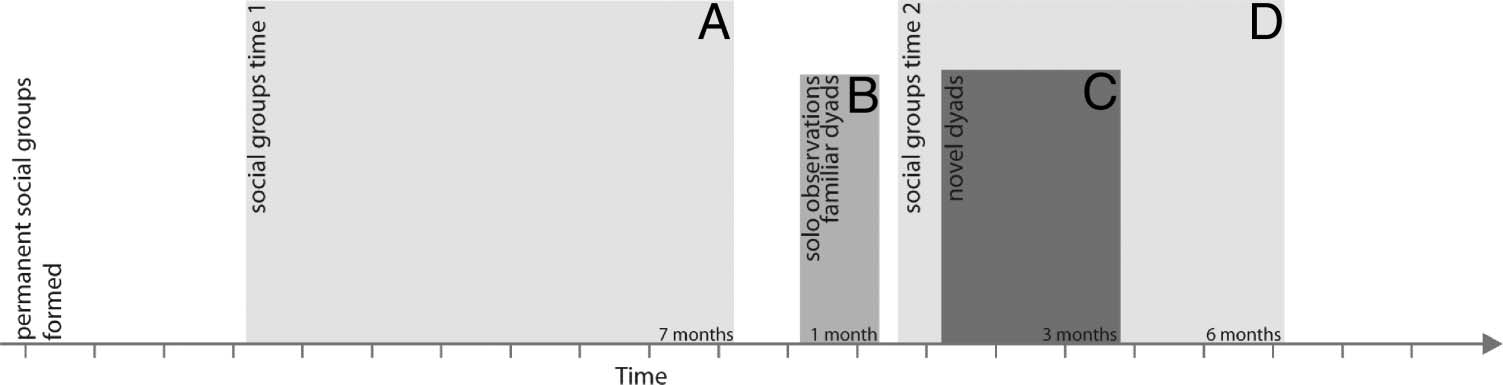
Figure 1 depicts the timing of the four behavioral experi-
ments. See the figure capture for the average ages of the
animals when each experiment was completed.
Solo observations.
Solo observations occurred on 5 con-
secutive days between 8 and 11 a.m. and 1 and 4 p.m. Each
subject was observed for two consecutive 5-min samples
during each morning and each afternoon session, yielding
a total of 20 observations per animal (note that data from
one 5-min sample was not available for one subject). Solo
observations occurred immediately before familiar dyad
observations. Because solo observations occurred without
a social interaction partner present with the focal animal,
the only state behaviors that were scored correspond to a
subset of those listed in the “nonsocial state” section of the
behavioral ethogram (Table 1). Specifically, only instances
of “nonsocial activity,” “nonsocial inactivity,” and “sleep”
were recorded for solo observations.
Social and affective behaviors generated by our experi-
mental animals were recorded in four different contexts
in a large test cage (as detailed below): (1) while each
animal was alone (“solo observations”), (2) while each
animal interacted with a series of animals from his or
her social rearing group (“familiar dyads”), (3) while each
animal interacted with a series of novel animals from a
different social rearing group (“novel dyads”), (4) while
each animal was in his or her social group (“social group
observations”). The same behavioral sampling technique
was used in all four settings. Behavioral data were col-
lected using The Observer 5.0 (Noldus, 1991) using the
focal sampling technique (Altmann, 1974) to record the
Familiar dyads.
Immediately following each solo ob-
servation, each subject was observed with a member of
his or her social rearing group for a 20-min dyadic inter-
action. During the 20-min dyadic interaction, each animal
was the focal animal (i.e., the focus of the observation)
for 10 of the 20 min. During each dyad, the focal animal
switched every 5 min, yielding a total of 20 observations
per animal. Data from one 5-min sample was not available
for one subject. Each focal animal met each partner animal
at two time points, resulting in 10 dyadic interactions per
animal. Animal testing order was counterbalanced for
testing order (test day), test session time (morning or
afternoon), interaction partner, and observer
2126
Journal of Cognitive Neuroscience
Volume 25, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 1. Behavioral Ethogram
Behavior a
States
Social states
Other contact
Proximity
Social activity
Description
Any physical contact between focal animal and other animal.
Animal is within armʼs reach of another animal.
Animal is not in proximity or other contact but is actively moving among
the group within armʼs reach or contact of the other animal(s).
Ventral–ventral contact
Ventral surface of the focal animal contacts ventral surface of another animal.
Groom
Extended play
Extended mount
Examination, picking, or licking of another animalʼs fur or body.
Rough and tumble play or chase play.
Any instance of mounting.
Extended negative
Any instance of aggression or chase.
Nonsocial states
Nonsocial activity
Animal remains out of all social states with head up, actively engaged in the environment.
Nonsocial inactivity
Animal remains out of all social states with head down, not engaged in environment.
Nonsocial avoidant
Nonsocial vigilanceb
Animal remains out of all social states and is actively avoiding other animals.
Animal remains out of all social states and is actively tracking the other animalʼs
Sleep
Events
Total communication
Barkc
Affiliative
position and placement.
Animal is asleep.
Low pitched, sharp, guttural sound.
Anogenital exploration
Oral, olfactory, or manual exploration of another animalʼs anogenital area.
Approach
Coo
Follow
Grunt
Lipsmack
Intentional movement within armʼs reach of another animal.
Clear, soft sounds, moderate in pitch and intensity; usually sounds like “whoooooo.”
Intentional follow of another animal.
Deep, muffled, low-intensity vocalization.
Rapid lip movements with pursed or puckered lips, usually accompanied by smacking sounds.
Incomplete mount
Mount that includes one or two, but not all three, of the necessary components of a “Mount.”
Inappropriate mount
An attempt to mount an inappropriate part of the body—head, side, or shoulder
instead of perineum.
Mount
Mount that includes all of the following components: appropriate positioning of partner,
hands on back, double foot clasp.
Huddle
Play threat
Present groom
Present mount
Physical contact that involves one animal ventrally touching another animal.
Open mouth threats, ear flaps, lunges, or head bobs that are more relaxed than typical threats.
Often occurs in context of Rough and Tumble Play.
Intentional presentation of neck, belly, or other part of body to another animal.
Rigid posture with rump and tail elevated and oriented toward another individual.
Rough and tumble play
Contact play consisting of mounting, tumbling, and wrestling.
Bliss-Moreau et al.
2127
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 1. (continued )
Behavior a
Agonistic/“aggression”
Aggression
Chase
Displacement
Full threat
Description
Grabbing, slapping, and biting of another animal.
Rapid pursuit of another animal lasting more than 3 sec.
Physical movement in which an animal “takes the place” of another animal.
Contains at least two or more of the following components: open mouth stare,
head bobbing, ear flaps, bark vocalizations, or lunges.
Mild threat
Contains only one of the following components: open mouth threats, ear flaps, lunges,
Toy-steal
Submission/“fear”
Crooktail
Grimace
Flee
Freeze
Scream
Exploration
Manual
Oral
Toy-play
Stress
Scratch
Self-groom
Tooth grind
Yawn
Other events
Cage shake
Crouch
or head bobs.
Deliberate and intentional taking of toy from another animal.
Tail held in stiff “?” shape.
Exaggerated movement of lips such that lips are pulled back with teeth showing.
Rapid, intentional movement away from another animal.
Stiff body posture without any movement for more than 3 sec.
High-pitched vocalization, with extreme high intensity; sounds like “eeeeeeeeee.”
Exploration of the cage or environment with the hands.
Exploration of the cage or environment with the mouth.
Exploration of toy.
Scratches own body.
Examining, picking, or licking oneʼs own fur or skin.
Repetitive, audible rubbing of upper and lower teeth.
Yawn.
Vigorous shaking of cage bars or body slams against the cage.
Animal is quadrupedal and bending down low but not exploring, eating, or drinking.
Mount refusal
Animal who is being mounted moves away or physically pushes partner away.
Self-sex
Withdraw
Tantrum
Anogenital exploration of self.
Animal moves out of armʼs reach of another animal after being in proximity or contact.
Violent shaking or spasms of the body often accompanied by a high-pitched sticatto vocalization.
Stereotypies were also scored, but those data have been reported elsewhere (Bauman, Toscano, Babineau, Mason, & Amaral, 2008) and so are not
reported here. To be scored in a “state” behavior must occur for 3 sec.
aBehavior was not scored for any monkey during the entire study.
bNonsocial vigilance was only scored in novel dyads at a low frequency.
cIn addition to the subordinate categories, Bark was included in the Total communication category.
Novel dyads. Each subject was observed with the six
experimental animals from one other social rearing
group during novel dyads (i.e., two unlesioned control
animals, two hippocampal-lesioned animals, and two
amygdala-lesioned animals). At the start of novel dyads,
interaction partners had never had any contact and were
unfamiliar to each other (i.e., they were not interaction
partners in the novel dyads conducted in Bauman et al.,
2004a, 2004b). Each focal animal met each partner animal
six times. Six dyadic interactions occurred in the morning
2128
Journal of Cognitive Neuroscience
Volume 25, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 1. Experimental
timeline. Note: Hashmarks on
time access indicate months.
(A) Mean age at the start of
observations was 1.44 years
(SD = 0.09). Mean age at the end
of observations was 2.09 years
(SD = 0.08). (B) Mean age at
the start of observations was
2.17 (SD = 0.08). Observations
were completed 4 weeks later.
(C) Mean age at the start of
observations was 2.34 years (SD = 0.10). Mean age at the end of observations was 2.46 years (SD = 0.10). (D) Mean age at the start of observations was
2.22 years (SD = 0.09). Mean age at the end of observations was 2.69 years (SD = 0.09).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
(8–11 a.m.) and six occurred in the afternoon (1–4 p.m.) of
each test day. Animal testing order was counterbalanced
for weekly testing order (test day), test session time
(morning or afternoon), interaction partner, and observer.
As in focal dyads, observations totaled 20 min for each
dyad, alternating the focal animal every 5 min, for a total
of 72 observations per focal animal. Two animals were
each missing data from one 5-min observation.
Social group observations. There were two sets of social
group observations. Observation periods were separated
by 2.5 months (see Figure 1). Each animal was observed
for a 5-min sample, one or two times per week, for a total
of 31–34 observations per focal animal per observation
period. Social behaviors initiated by the focal animal were
qualified in terms of whether they were directed at a peer
(i.e., an amygdala-lesioned, hippocampus-lesioned, or
sham-operated control animal), an adult (i.e., the adult
male or adult female living with the social group), or the
group (i.e., behavior with nonspecific or no social target).
Observation order was pseudorandomized.
Data Analysis Strategy
Behaviors that were initiated by focal animals were
grouped into broad behavioral categories as indicated
in Table 1. Frequencies and durations were summed
across each category for each type of interaction partner
(as specified above) and then averaged across the number
of observations to create a mean per observation. ANOVA
was performed on each broad behavioral category with
focal animal lesion group as the between-subject factor.
Significant subject effects were further evaluated with post
hoc least significant difference tests and within-subject
effects were evaluated with paired t tests. Data were
log10(x + 1) transformed in cases where they were not
normally distributed. For the purposes of interpretation,
raw data (means and variance indices) are presented; log-
transformed data are available upon request. Mauchlyʼs test
of sphericity was used to assess whether the data violated
the assumption of sphericity. Degrees of freedom were
Greenhouse–Geisser corrected when necessary. Cases
that required correction are noted in the tables; the cor-
rected degrees of freedom are available upon request.
We conducted a series of ANOVA analyses to evaluate
lesion-based differences in each individual behavior at
this time point to make the results more easily compar-
able to results found at 6 and 9 months of age (Bauman
et al., 2004a) in the same animals. For the sake of brevity,
only significant results and those about which there were
a priori hypotheses (based on the findings of Bauman
et al., 2004a) are presented here. Other analyses are
available upon request.
Finally, we assessed the extent to which the organiza-
tion of behaviors of the lesioned animals in the presence
of the intact control animals varied by lesion condition
and could be used to identify lesion groups. We also
completed a series of MANOVA analyses followed by dis-
criminant function analyses on the social behaviors gen-
erated while interacting with control animals during the
dyad experiments. Beyond the lesion-based differences
in the frequency and duration of behaviors during social
interactions, we were interested in whether the organi-
zation of classes of behaviors (e.g., correlations across
multiple behaviors) might predict lesion group member-
ship. To assess whether lesion condition might influence
the relationship between behaviors, we conducted an
additional set of analyses using the data collected when
focal animals interacted with control partners. Specifi-
cally, we ran MANOVAs on the dependent variables that
constituted the behavioral categories above with lesion
condition as a between-subjects factor and then fol-
lowed those MANOVAs with discriminant function analy-
sis to examine how the relationship between dependent
variables discriminated the lesion groups. Only signifi-
cant MANOVAs are discussed. We were primarily inter-
ested in whether early damage to the amygdala might
alter patterns of close social interactions—those in which
animals actively engage each other. As such, we conducted
MANOVAs and discriminant function analyses on the
social state data (frequency and duration). Given that dif-
ferences in exploratory behaviors were found at earlier
time points, we also conducted similar analyses on the
exploration data. Those analyses are available from either
Bliss-Moreau et al.
2129
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the first author (EBM: eblissmoreau@ucdavis.edu) or
senior author (DGA: dgamaral@ucdavis.edu).
RESULTS
Solo Observations
There were lesion-based differences in the frequency of
state changes during solo observations, F(2, 21) = 5.31,
p = .014, ηp
2 = .336; A > C, H, p = .01 (log-transformed
analyses, raw means presented below). State changes
occurred more frequently for amygdala-lesioned animals
(M = 1.86, SE = 0.39) who changed their state more
frequently than did control and hippocampus-lesioned
animals (for both groups M = 1.00, SE = 0.00). Whereas
hippocampus-lesioned and control animals spent all of
each 5-min sampling period moving around the cage
(being “active”), amygdala-lesioned subjects also spent
time in states of “inactivity” (M = 0.46, SE = 0.21) and
sleep (M = 0.05, SE = 0.03). Table of all means is available
by request.
State differences were reflected in the duration data as
well. Amygdala-lesioned animals spent less time in the
active state compared with control and hippocampus-
lesioned animals, F(2, 21) = 4.04, p = .033, ηp
2 = .278
(log-transformed analyses; raw means: Amygdala-lesioned
animals: M = 284.07, SE = 7.83; control and hippocampus-
lesioned animals: M = 300.00, SE = 0.00). Five of the
eight amygdala-lesioned animals spent time in the inactive
state and/or asleep.
No significant lesion-based differences in exploration
of any kind were observed during this experiment.
Familiar Dyads
See Table 2 for frequency data and Table 3 for duration
data.
Total Number of State Changes
Replicating the finding from solo observations, lesion
conditions differed in the total number of state changes
during familiar dyads. Amygdala-lesioned animals initiated
the most state changes, and hippocampus-lesioned ani-
mals initiated the least state changes.
Social states. Amygdala-lesioned and control animals in-
itiated social states most frequently, whereas hippocampus-
lesioned animals initiated social states least frequently.
Social states were initiated most frequently with control
partner animals and least frequently with hippocampus-
lesioned animals. Control animals initiated the longest
duration of social states, although the only significant
between-group difference was between control and
hippocampus-lesioned animals. Social state durations were
also significantly longer with control animals as compared
with both amygdala-lesioned and hippocampus-lesioned
animals.
Amygdala-lesioned animals initiated the highest fre-
quency of nonsocial states, and hippocampus-lesioned
animals initiated the lowest frequency, although, once
again, only amygdala- and hippocampus-lesioned animals
differed significantly. Control animals spent the least
amount of time in nonsocial states—significantly less than
both amygdala- and hippocampus-lesioned animals.
Amygdala- and hippocampus-lesioned animals did not dif-
fer significantly in the duration of time spent in nonsocial
states. This pattern of effects was seen in the partner lesion
data as well. Nonsocial state durations were significantly
shorter when focal animals interacted with control animals,
as compared with both amygdala- and hippocampus-
lesioned animals.
Total Communication
Amygdala-lesioned animals produced the most communi-
cative signals. A significant partner lesion effect revealed
that focal animals generated fewer communicative signals
with hippocampus-lesioned animals than with animals of
the other two groups. Amygdala-lesioned animals gener-
ated fewest communicative signals with control animals,
whereas both control and hippocampus-lesioned animals gen-
erated fewest communicative signals with hippocampus-
lesioned subjects as indicated by a significant focal lesion ×
partner lesion effect.
Affiliative signals. The effect of lesion condition on
communicative signals was primarily driven by amygdala-
lesioned animalsʼ increased affiliative signaling. When the
total frequency of affiliative signals was considered alone,
amygdala-lesioned animals produced significantly more
affiliative signals than did control or hippocampus-lesioned
animals. Overall, affiliative signals were generated equally
frequently with amygdala-lesioned and control partners
and least frequently with hippocampus-lesioned partners.
A significant focal lesion × partner lesion effect revealed
that amygdala-lesioned animalsʼ generation of affiliative
signals was consistent across interaction partners whereas
both control and hippocampus-lesioned animals pro-
duced fewer affiliative signals with hippocampus-lesioned
animals.
Submission/“fear”-related signals. There were no lesion
group or partner lesion effects on submission or “fear”-
related behaviors.
Agonistic/“aggression”-related signals. Amygdala-
lesioned animals were significantly less agonistic than both
hippocampus-lesioned and control animals.
Exploratory Behaviors
Amygdala-lesioned animals explored the least, whereas
hippocampus-lesioned animals explored the most. A part-
ner lesion effect revealed that exploratory behavior was
2130
Journal of Cognitive Neuroscience
Volume 25, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 2. Mean Frequency per Familiar Dyad Observation
Focal Lesion Condition
Focal Lesion Effect
Partner Lesion Condition
Partner Lesion Effect
Partner Lesion
Pairwise
Comparison
Focal Lesion ×
Partner Lesion
Effect
Behavior
Total states
Sociala
AMY
CON
HIP
F(2, 21)
p
ηp
2
Post hoc
AMY
CON
HIP
F(2, 42)
p
2
ηp
t(23)
p
F(4, 42)
7.95 (0.48)
7.12 (0.32)
5.90 (0.53)
5.16
.015
3.39 (0.28)
3.02 (0.34)
2.26 (0.28)
4.62
.022
.330 A > H p = .004
C > H p = .072
.306 A > H p = .007
C > H p = .054
7.35 (0.46)
7.62 (0.46)
5.99 (0.50)
3.76
.031 .152
2.26
A > H p = .033
2.38 C > H p = .026
2.91 (0.28)
3.41 (0.30)
2.34 (0.26)
3.47
.040 .142
2.70 C > H p = .013
Nonsocial
4.56 (0.25)
4.10 (0.11)
3.64 (0.28)
4.20
.029
.286 A > H p = .009
4.44 (0.23)
4.22 (0.26)
3.65 (0.26)
2.84
.070 .119
–
–
2
ηp
–
–
p
–
–
–
–
–
–
Total communication
18.47 (1.56) 12.62 (1.41) 12.22 (1.72)
4.98
.017
eventsb
Affiliative
17.22 (1.51)
10.10 (.84) 10.18 (1.49)
9.63
.001
.322 A > C p = .015
A > H p = .010
.478 A > C p = .001
A > H p = .001
15.77 (1.17) 14.81 (1.38) 12.74 (1.31)
3.28
.048 .135
2.95
A > H p = .007
2.81
.037 .211
14.04 (1.19) 12.55 (1.13) 10.92 (1.34)
4.08
.024 .163
2.86
A > H p = .009
3.23
.021 .235
Submission/“fear”a
0.90 (0.22)
1.04 (0.59)
0.60 (0.16)
0.26
.776
.024
–
.58 (.18)
1.36 (.34)
.59 (.19)
Agonistic/“aggression”a
0.32 (0.08)
1.28 (0.19)
1.43 (0.19)
16.50
.00005 .611 C > A p = .0002
H > A p = .00002
1.05 (0.19)
0.77 (0.16)
1.22 (0.27)
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
Total exploration events
3.74 (0.78)
4.92 (0.53)
6.30 (0.62)
3.85
.038
.268 H > A p = .011
4.99 (0.55)
3.88 (0.45)
6.10 (0.63)
7.91
.001 .274
3.64 H > C p = .001
A > C p = .095
1.74
1.87 H > A p = .074
3.45
.016 .247
aStatistical analyses were performed on log-transformed data, but raw means are presented for ease of interpretation.
bIn addition to the subordinate categories, Total communication also includes Bark.
B
l
i
s
s
–
M
o
r
e
a
u
e
t
a
l
.
2
1
3
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
8
n
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
2
1
3
2
J
o
u
r
n
a
l
o
f
C
o
g
n
i
t
i
v
e
N
e
u
r
o
s
c
i
e
n
c
e
V
o
l
u
m
e
2
5
,
N
u
m
b
e
r
1
2
Table 3. Mean Duration per Familiar Dyad Observation
Focal Lesion Condition
Focal Lesion Effect
Partner Lesion Condition
Partner Lesion Effect
Partner Lesion Pairwise
Comparison
Focal Lesion ×
Partner Lesion
Effect
Behavior
AMY
CON
HIP
F(2, 21)
p
ηp
2
Post hoc
AMY
CON
HIP
F(2, 42)
p
ηp
2
t(23)
p
F(4, 42)
Total social statesa
37.60 (3.99)
59.14 (11.61)
32.39 (4.89)
4.53
.023
.302
C > H p = .007
38.56 (5.79)
59.89 (8.41)
30.69 (5.08)
6.37
.004
.233
2.40
3.98
C > A p = .025
C > H p = .001
Other contacta
Proximitya
3.48 (1.27)
3.89 (1.16)
2.90 (1.60)
24.41 (4.45)
19.53 (1.95)
15.88 (2.54)
–
–
–
–
–
–
–
–
2.69 (.58)
4.08 (1.82)
3.49 (1.55)
–
–
–
–
–
16.16 (2.36)
26.84 (3.99)
16.82 (1.87)
4.50
.017
.177
.32 (.21)
28.29 (10.42)
8.30 (3.42)
6.20
.008
.371
C > A p = .002
C > H p = .042
10.50 (4.29)
19.88 (6.88)
6.54 (4.31)
2.51
.093
.107
8.12 (1.99)
5.81 (1.77)
3.88 (1.61)
1.27 (0.87)
1.18 (0.45)
1.36 (0.57)
–
–
–
–
–
–
–
–
–
–
–
–
7.83 (1.59)
7.36 (2.30)
2.62 (.96)
3.92
.028
.157
3.35
A > H p = .003
1.04 (0.37)
1.67 (0.53)
1.11 (0.62)
0.34 (0.29)
0.06 (0.06)
0.10 (0.09)
–
–
–
–
–
–
–
–
–
–
Extended negativea
0.00 (0.00)
0.45 (0.29)
0.06 (0.06)
Total nonsocial statesb
238.24 (7.54)
194.77 (7.63)
240.38 (8.34)
10.76
.001
.506
237.63 (7.65)
193.80 (7.38)
240.11 (8.26)
11.23
.0005
.517
A > C p = .001
H > C p = .0005
A > C p = .001
H > C p = 0004
238.33 (7.04)
193.97 (10.37)
241.09 (7.67)
14.70
.00009
.412
238.28 (7.04)
192.31 (10.48)
240.97 (7.69)
15.46
.0007
.424
3.87
A > C p = .001
4.12 H > C p = .0004
3.94
A > C p = .001
4.21 H > C p = .0003
0.31 (0.17)
0.06 (0.06)
0.00 (0.00)
0.30 (0.27)
0.91 (0.62)
0.27 (0.27)
–
–
–
–
–
–
–
–
0.06 (0.06)
0.27 (0.17)
0.04 (0.04)
0.89
.387
0.00 (0.00)
1.39 (0.72)
0.08 (0.06)
3.37
.077
.041
.138
–
–
–
–
Grooma
Playa
Mounta
Activityb
Inactivitya,b
Sleepa,b
2.72
2.54
–
C > A p = .012
C > H p = .019
–
p
–
–
–
–
–
ηp
2
–
–
–
–
–
–
–
–
–
–
4.37
.005
.294
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
aStatistical analyses were performed on log-transformed data, but raw means are presented for ease of interpretation.
bDistribution violates assumptions of sphericity. Degrees of freedom were Greenhouse–Geisser corrected.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
8
n
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
greatest during interactions with hippocampus-lesioned
subjects and least during interactions with control animals.
Amygdala-lesioned animals exploratory behavior was con-
sistently low across all interaction partners, whereas con-
trol and hippocampus-lesioned animals had increased
exploratory behavior with hippocampus-lesioned interac-
tion partners.
Lesion Group Classification Based on Patterns of
Behaviors with Familiar Control Animals
Social State Frequency
There was a significant effect of lesion condition on social
state frequency, V = 1.153, F(12, 34) = 3.853, p = .00094,
indicating that the organization of social states differed
by lesion group. Separate univariate ANOVAs, however,
on the social state variables revealed a significant effect of
lesion condition only on grooming behavior, F(2, 21) =
5.690, p = .011, ηp
2 = .351 (C > A, p = .00597; C > H,
p = .0116), and nonsignificant lesion group effects on
mounting, extended negative, play, contact, and proximity.
Discriminant function analysis revealed that the relation-
ship between social state variables was captured by two
functions, the first which explained 84.6% of the variance
(canonical R2 = .77) and the second which explained
15.4% of the variance (canonical R2 = .38). A combination
of these discriminant functions differentiated the lesion
conditions, Λ =0 .141, χ2(12) = 36.183, p = .0003. The
correlations between the social states and discriminant
functions indicated that grooming (r = .401) and extended
negative (r = .167) loaded more highly onto the first factor,
whereas proximity (r = .588), contact (r = .435), play (r =
.323), and mounting (r = −.320) loaded most highly onto
the second factor. Given that there was no extended nega-
tive initiated by the amygdala- and hippocampus-lesioned
animals and that their rates of grooming compared with
controls were low, Function 1 likely captured the social be-
havior patterns unique to control animals. Taken together,
the two functions were able to correctly classify 83.3% of
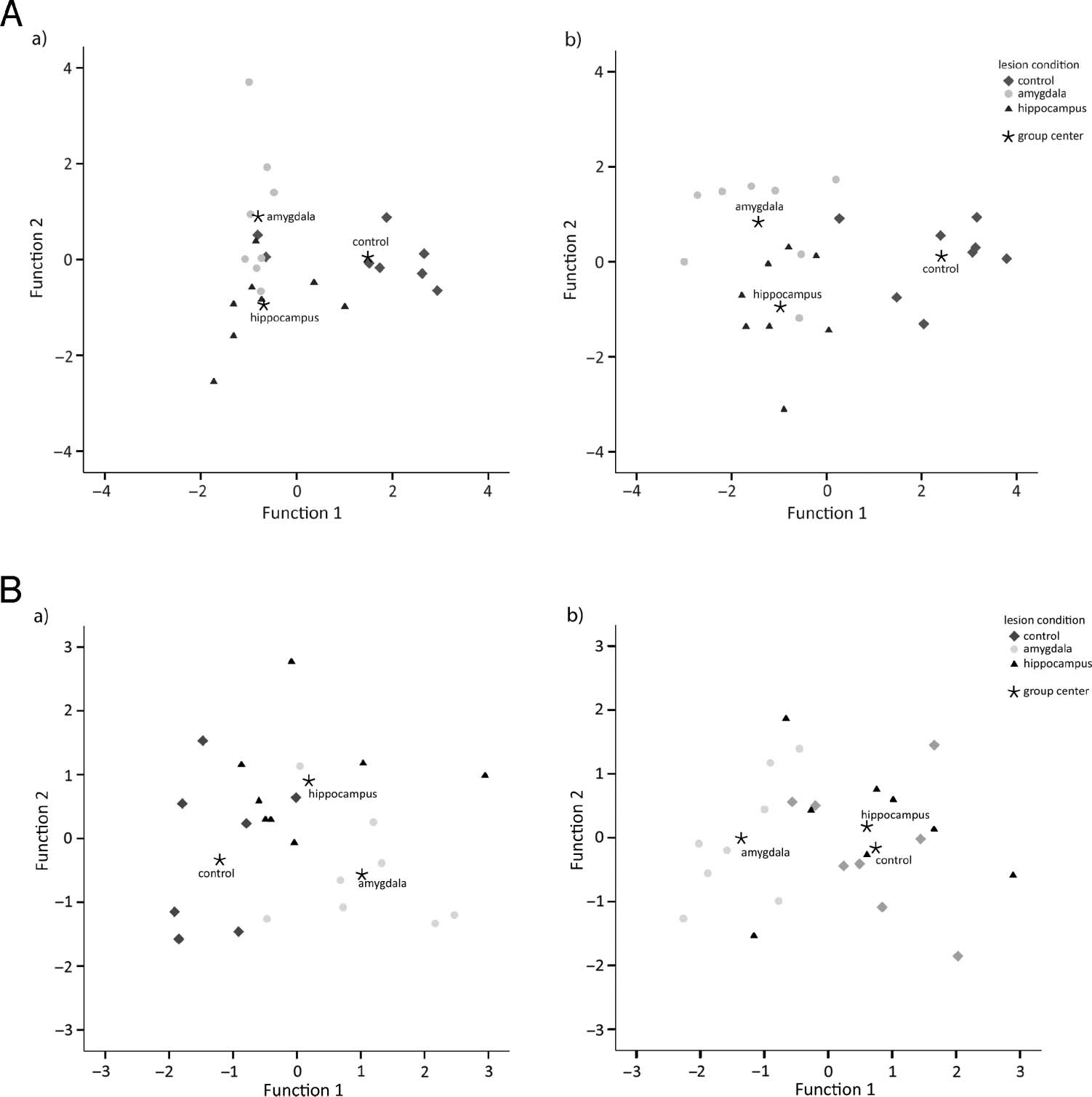
the animals into their correct lesion groups (7/8 controls,
7/8 amygdala-lesioned, 6/8 hippocampus-lesioned), Pressʼs
Q Statistic = 27, p < .001. One amygdala-lesioned subject
was misclassified as a hippocampus-lesioned subject, and
two hippocampus-lesioned subjects were misclassified as
amygdala-lesioned subjects; one control animal was mis-
classified as an amygdala-lesioned subject. See Figure 2A
for a visual depiction of the group classification.
Figure 2. Classification of
lesion groups based on (A) the
duration of time spent in social
states and (B) the frequency of
social states during familiar
dyads. Each individual data
point represents a single animal.
In both cases, Function 1
maximally separated control
from lesion groups.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Bliss-Moreau et al.
2133
Social State Duration
The analysis of social state duration paralleled the anal-
ysis of the frequency data. There was a significant ef-
fect of lesion condition on social state durations, V =
0.952, F(12, 34) = 2.572, p = .015, indicating that the
organization of social state durations differed by lesion
group. However, as in the analysis of social state fre-
quency behaviors, separate univariate ANOVAs on the
social state variables revealed a significant effect of le-
sion condition only on grooming behavior, F(2, 21) =
6.982, p = .005, ηp
2 = .399 (C > A, p = .002; C > H, p =
.009), and nonsignificant lesion group effects on mount-
ing, extended negative, play, contact, and proximity. Dis-
criminant function analysis revealed that the relationship
between social state duration variables was captured by
two functions, the first of which explained 66.2% of the
variance (canonical R2 = .56) and the second of which
explained 33.8% of the variance (canonical R2 = .39).
A combination of these discriminant functions differen-
tiated the lesion conditions, Λ = 0.268, χ2(12) = 24.369, p =
.018. The correlations between the social states and dis-
criminant functions revealed that grooming (r = .719)
and extended negative (r = .274) loaded more highly
onto the first factor whereas proximity (r = .580), contact
(r = −.173), play (r = .296), and mounting (r = −.296)
loaded most highly onto the second factor. As with the
duration data, Function 1 therefore likely captured the
social behavior patterns unique to control animals because
there were no extended negative social interactions
initiated by the amygdala and hippocampus-lesioned ani-
mals and their rates of grooming were low (compared
with controls). Taken together, the two functions were
able to correctly classify 75.0% of the animals into their
correct lesion groups (6/8 controls, 6/8 amygdala-lesioned,
and 6/8 hippocampus-lesioned), Pressʼs Q Statistic = 18.75,
p < .001. Two controls were misclassified as amygdala-
lesioned animals, two amygdala-lesioned animals were
misclassified as hippocampus-lesioned animals, and one
hippocampus-lesioned animal was misclassified as an
amygdala-lesioned animal. See Figure 2B for a visual
depiction of the group classification.
Novel Dyads
Statistics for significant analyses are presented below. All
additional means and statistics are available upon request.
Total Number of State Changes
In contrast to the findings in familiar dyads, there was no
effect of lesion condition on the total number of state
changes or the number or duration of social or nonsocial
states during novel dyads. Amygdala-lesioned animals did,
however, spend more time in the inactive state (M = 1.60,
SE = 0.56) than both control (M = 0.20, SE = 0.20)
and hippocampus-lesioned (M = 0.00, SE = 0.00) animals,
F(2, 21) = 7.20, p = .044, ηp
2 = .407; A > C, p = .007;
A > H, p = .002 (log-transformed analyses, raw means
presented).
Total Communication
In contrast to the findings in familiar dyads, there were
no lesion-based differences in the total number of com-
municative signals during novel dyads.
Affiliative signals.
In contrast to the findings in familiar
dyads, there were no lesion-based differences observed in
the total number of affiliative signals during novel dyads.
Submission/“fear”-related signals. Although there was
not a significant effect of focal animal lesion on submission-
related behaviors, there was a significant effect of partner
lesion condition, F(2, 42) = 4.19, p = .022, ηp
2 = .166;
C > A, p = .015; H > A, p = .085 (log-transformed
analyses, raw means presented below). All animals were
least submissive when interacting with amygdala-lesioned
animals (M = 1.65, SE = 0.32) as compared with when they
interacted with control animals (M = 2.56, SE = 0.36) or
hippocampus-lesioned animals (M = 2.12, SE = 0.37).
Agonistic/“aggression”-related signals. Across all three
partner lesion conditions, amygdala-lesioned animals ini-
tiated fewer agonistic behaviors (M = 0.36, SE = 0.10) than
either control (M = 1.20, SE = 0.33) or hippocampus-
lesioned animals (M = 0.96, SE = 0.17); F(2, 21) = 4.43,
p = .025, ηp
2 = .297; C > A, p = .011; H > A, p = .030
(log-transformed analyses, raw means presented).
Exploratory Behaviors
There was a main effect of lesion condition on explora-
tory behavior, F(2, 21) = 12.08, p = .0003, ηp
2 = .535
(log-transformed analyses, raw means presented below).
Rates of exploration were significantly lower for amygdala-
lesioned animals (M = 2.38, SE = 0.40) than for control
animals (M = 5.60, SE = 0.45) or hippocampus-lesioned
animals (M = 5.36, SE = 0.66).
Lesion Group Classification Based on Patterns of
Behaviors with Novel Control Animals
As in the familiar dyads, we used behaviors generated
with control animals to attempt to predict lesion group
membership. The MANOVAs on social state data were
not significant. Only the MANOVA on the exploratory be-
havior yielded significant results. It is available by request.
Social Groups
See Table 4 for the duration data. The only significant
effects in the frequency data were relative to time
2134
Journal of Cognitive Neuroscience
Volume 25, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 4. Mean Duration per Social Group Observation (Main Effects)
Focal Lesion Condition
Focal Lesion Effect
Partner Lesion Effect
Partner
Lesion
Pairwise
Comparison
Behavior
AMY
CON
HIP
F(2, 21)
p
η p
2
Post hoc
F(2, 42)
η p
2
t(23)
Total social states
67.37 (4.51)
106.92 (10.28)
92.74 (8.78)
7.51
.003
.417
C > A p = .001
H > A p = .016
(with all partners)a,b
Total social states
(with peers)a
Other contacta
Proximitya
Grooma
Playa
Mounta
27.33 (2.06)
46.61 (6.07)
33.64 (2.77)
5.78 (1.02)
11.59 (2.00)
7.27 (0.72)
12.70 (1.04)
21.65 (4.13)
14.74 (1.43)
3.70
3.25
–
.042
.261
C > A p = .014
.059
.236
–
–
–
–
1.19 (.87)
7.31 (2.12)
5.92 (2.02)
4.37
.026
.294
C > A p = .011
H > A p = .038
7.47 (1.70)
5.04 (0.85)
5.57 (0.51)
0.15 (0.08)
0.36 (0.17)
0.13 (0.05)
–
–
–
–
–
–
Extended negativea
0.01 (0.01)
0.05 (0.02)
0.01 (0.01)
2.68
.092
.203
Ventrala
0.02 (0.01)
0.61 (0.50)
0.00 (0.00)
–
–
–
Total nonsocial statesc
185.44 (5.20)
138.16 (10.34)
154.01 (7.75)
8.96
.002
.460
Activity
181.26 (5.01)
132.48 (10.53)
149.59 (8.21)
9.03
.001
.462
–
–
–
–
A > C p = .0004
A > H p = .012
A > C p = .0004
A > H p = .013
Inactivitya
Sleepa
0.51 (0.35)
0.81 (0.65)
0.79 (0.41)
3.68 (1.62)
4.87 (1.45)
3.62 (1.09)
–
–
–
–
–
–
–
–
p
–
–
–
–
–
–
–
–
–
3.02
.060
.126
–
–
–
2.65
.083
.112
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
–
p
–
–
–
–
–
–
–
–
–
–
–
–
–
Data presented are behaviors that occurred in the presence of peers, except total social (with all partners). Tables representing behaviors occurring in the presence of all animals in social groups are available upon request.
aStatistical analyses were performed on log-transformed data, but raw means are presented for ease of interpretation.
bDegrees of freedom for focal lesion effect and test time effect are 1, 21.
B
l
i
s
s
–
M
o
r
e
a
u
e
t
a
l
.
2
1
3
5
Time Condition
Test Time Effect
Time 1
Time 2
F(4, 42)
p
80.52 (5.63)
97.50 (6.59)
12.78
.002
η p
2
.378
Direction
of Effect
2 > 1
32.85 (2.89)
38.87 (3.08)
13.19
.002
.386
2 > 1
8.64 (1.30)
7.79 (1.16)
–
–
14.04 (1.48)
18.69 (2.07)
22.51
.0001
3.33 (0.93)
6.28 (1.48)
9.81
.005
–
.517
.318
–
2 > 1
2 > 1
6.29 (0.92)
5.76 (0.59)
–
–
–
–
0.10 (0.05)
0.33 (0.11)
5.16
.034
.197
2 > 1
0.03 (0.01)
0.02 (0.01)
0.42 (0.34)
0.00 (0.00)
–
–
–
–
–
–
–
–
175.25 (6.74)
143.16 (6.79)
25.97
.00005
.553
1 > 2
168.97 (7.02)
139.92 (6.83)
20.99
.0002
.500
1 > 2
0.24 (0.14)
1.17 (0.49)
6.04 (1.33)
2.08 (0.47)
4.95
7.01
.037
.015
.191
.250
2 > 1
1 > 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
8
n
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
effects (Time 1 vs. Time 2); those data are available upon
request.
Social Behaviors
Lesion-based differences in the duration of time spent
in social states was first assessed across all possible in-
teraction partners (including behaviors scored in the
presence of individual peers, adults, and the entire
group). There was a significant effect of time such that
all animals spent more time in social states during the
second as compared with the first observation period.
Amygdala-lesioned animals spent less time in social
states than control and hippocampus-lesioned animals.
There was no time × lesion effect indicating that time
effect did not vary by lesion condition nor did the lesion
effect vary by time. Animals initiated more social states
with all possible interaction partners at Time 2 as com-
pared with Time 1. There were no focal lesion condition
differences in the frequency of social states.
Lesion-based differences in social states were driven by
interactions with peers rather than with the adults, and
so only those effects are discussed further (all other ana-
lyses are available upon request). There were no signifi-
cant effects in an analysis of the social state data with
adults only.
All animals spent more time interacting with their age-
matched peers at Time 2 as compared with Time 1. This
was true for the total time spent in social states, and the
effect was driven by time spent with peers in proximity,
grooming, and mounting. The effect of time on proximity
was qualified by a complex significant focal lesion × part-
ner lesion × time effect, F(3.128, 32.841) = 3.903, p =
.016, ηp
2 = .271 (log-transformed analyses). Control ani-
mals spent the same amount of time in proximity with
amygdala-lesioned animals at both time points, yet greater
lengths of time with members of the other groups at
Time 2 as compared with Time 1. In contrast, amygdala-
and hippocampus-lesioned animals spent more time with
amygdala-lesioned animals at Time 2 than Time 1, but
the same length of time with both control animals and
hippocampus-lesioned animals.
Amygdala-lesioned animals spent less time grooming
their peers than both controls and hippocampus-lesioned
animals. Amygdala-lesioned animals also groomed less fre-
quently than control and hippocampus-lesioned animals
across both meetings, F(1, 21) = 5.035, p = .016, ηp
2 =
.324 (log-transformed analyses; raw means; Amygdala-
lesioned: M = 0.057, SE = 0.036; control animals: M =
0.171, SE = 0.039; hippocampus-lesioned: M = 0.144,
SE = 0.033), although grooming frequency increased over-
all between observation period Time 1 (M = 0.095, SE =
0.021) and Time 2 (M = 0.153, SE = 031), F(1, 21) = 6.857,
p = .016, η p
2 = .246.
Although there was not a significant effect of lesion on
extended negative behavior overall and the frequencies
of agonistic behaviors were extremely low, the general
pattern of lesion-based differences in the duration of
agnostic behavior paralleled previous findings (Bauman
et al., 2004a). The effect of focal lesion on the duration of all
initiated extended negative was not statistically significant.
However, there was a trend for control animals to have
longer durations of extended negative behavior than
amygdala- and hippocampus-lesioned animals. This was
qualified by a significant focal lesion × partner lesion
interaction, F(4, 42) = 3.18, p = .023, ηp
2 = .232
(log-transformed), in which control animals engaged
in the longest duration of extended negative towards
hippocampus-lesioned animals.
Significant lesioned-based differences in the frequency
of both displacement and aggressive grabbing paralleled
the observations with these animals earlier in their devel-
opment (Bauman et al., 2004a). Specifically, amygdala-
lesioned animals displaced their peers less frequently than
control and hippocampus-lesioned animals, F(2, 21) =
5.612, p = .011, ηp
2 = .348 (log-transformed analyses;
raw means; Amygdala-lesioned: M = 0.034, SE = 0.008;
hippocampus-lesioned: M = 0.038, SE = 0.011; control:
M = 0.038, SE = 0.011). Amygdala-lesioned animals also
initiated fewer instances of aggression than did control
and hippocampus-lesioned animals, F(2, 21) = 4.863, p =
.018, ηp
2 = .317 (log-transformed analyses; raw means;
Amygdala-lesioned: M = 0.034, SE = 0.014; hippocampus-
lesioned: M = 0.133, SE = 0.030; control: M = 0.129, SE =
0.030). The frequency of aggression decreased from
Time 1 to Time 2 for control and hippocampus-lesioned
animals, but not for amygdala-lesioned animals whose
frequency remained low over time, F(2, 21) = 4.258,
p = .028, ηp
2 = .289.
There were a number of time-related effects that speak
to the development of social behavior. Animals spent
more time in social states during the second set of ob-
servations as compared with the first set of observations
(including the total duration of social states, total proximity,
grooming, and mounting), but the frequency of commu-
nicative behaviors was lower in the second as compared
with the first set of observations. A focal lesion by time
effect that did not reach conventional levels of significance,
F(2, 21) = 2.90, p = .077, η p
2 = .216 (log-transformed),
indicated that whereas controls did not change in their
communicative signaling from Time 1 to Time 2, both
amygdala-lesioned and hippocampus-lesioned animals
tended to become less communicative.
Nonsocial Behaviors
Overall, all monkeys spent more time in nonsocial states
during the first set of observations as compared with the
second set of observations. Amygdala-lesioned animals
spent more time in nonsocial states as compared with
control and hippocampus-lesioned animals. This effect
was driven by time and lesion-based differences in the
active state. All animals spent more time being inactive
during the second as compared with the first set of
2136
Journal of Cognitive Neuroscience
Volume 25, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
observations and more time asleep during the first as
compared with second set of observations.
Table 5. Comparison of Amygdala-lesioned and Control
Animals over Time
DISCUSSION
These experiments demonstrate that early amygdala
damage results in subtle differences in juvenile behavior
across a variety of social contexts. Amygdala-lesioned ani-
mals were the least aggressive animals in the present experi-
ments. They did not show heightened submission/“fear”
signaling as seen earlier in development (Bauman et al.,
2004a). Social partners were least submissive when inter-
acting with amygdala-lesioned animals. When interacting
with familiar partners, amygdala-lesioned animals spent
less time than the other animals in social states, particu-
larly highly interactive social states like grooming. This
difference became more pronounced over the course
of the experiments. This pattern of effects is remarkable
because, despite spending less time socially interacting
and being least aggressive, amygdala-lesioned animals
had heightened communication, were the most affiliative
group, and were more likely to reciprocate play behav-
iors. In other words, the members of their established
groups should have found the amygdala-lesioned animals
to be nonthreatening and engaging social partners lead-
ing to longer social interactions, but that was not the
case. As in previous reports (Bauman, Toscano, Mason,
Lavenex, & Amaral, 2006; Bauman et al., 2004a, 2004b),
the hippocampus-lesioned subjects behaved essentially
like control animals when assessing the frequency and
duration of specific behaviors.
General developmental patterns in the maturation of
social behavior were observed for all animals over time.
For example, all animals spent more time socially inter-
acting and were less aggressive in social groups during
the second set of observations as compared with the first
set, which were separated by 2.5 months. Communicative
signaling decreased from the first to second set of ob-
servations, particularly for amygdala- and hippocampus-
lesioned subjects. Changes in communicative signaling
were also observed across experimental test phases (i.e.,
infancy to present). See Table 5 for a summary compar-
ing amygdala-lesioned and control animals as infants
(Bauman et al., 2004a) and the present test period. As in-
fants, amygdala-lesioned animals, compared with control
animals, had both heightened submissive/“fear” and affilia-
tive signaling (Bauman et al., 2004a). As juveniles, amygdala-
lesioned animals had heightened affiliative signaling during
familiar dyadic interactions but no evidence of heightened
affiliative signaling in other contexts or submissive signaling
in any context. These changes over time likely reflect gen-
eral social development, stabilization of the social dynamics
in the permanently house social groups, as well as experi-
ence-dependent brain plasticity.
The present findings also illustrate that, although early
brain damage may not cause profound variation in specific
Behavioral Categories
Solo Observations
Exploration
Activity
Familiar Dyads
Affiliation
Submission/“Fear”
Agonistic/“Aggression”
Exploration
Novel Dyads
Affiliation
Submission/“Fear”
Agonistic/“Aggression”
Exploration
Social Groups
Submission/“Fear”
Agonistic/“Aggression”c
Exploration
Social Duration
Effects during
Infancy after
Weaning a
Effects during
Juvenile Period b
C > A
–
A > C
A > C
–
C > A
A > C
A > C
C > A
C > A
A > C
C > A
C > A
–
–
C > A
A > C
–
C > A
–
–
–
C > A
C > A
–
C > A
–
C > A
For ease of interpretation, only the differences between the amygdala-
lesioned and control groups are presented here. The hippocampus-
lesioned group did not differ consistently from the control group in
either period of evaluation. Note that Bauman et al. (2004a) evaluated
each behavior in individual analyses rather than grouping behaviors
into broad categories. Lesion group variation in the broad categories of
behavior is presented here for consistency with the present report.
aPreviously reported in Bauman et al. (2004a).
bReported in this article.
cLesion group difference only in displacement behavior, rather than
broad category.
social behaviors, it does impact the patterns of expressed
behavior. Typical statistical analyses used to assess varia-
tion in individual social behaviors (as in Machado, Emery,
et al., 2008; Machado & Bachevalier, 2006; Bachevalier,
Málková, & Mishkin, 2001; Emery et al., 2001) do not ac-
count for how brain damage may impact the covariation
between social behaviors. MANOVA evaluates the covaria-
tion of behaviors across groups but is typically not suited
for the small sample sizes in most nonhuman primate stu-
dies. By subjecting small subsets of behaviors to MANOVA,
we were able to observe lesion-based differences in the
organization of social states and exploratory behavior.
Bliss-Moreau et al.
2137
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Furthermore, in cases where there were significant lesion-
based behavioral organization differences, we were able
to use those data to classify, with high fidelity, the sub-
jects into groups that reflected their experimental status.
Notably, despite the fact that hippocampus-lesioned ani-
mals appeared to behave like controls when the average fre-
quency and duration of behaviors were considered, when
patterning of behavior is considered, the hippocampus-
animals appear to be unique. The classifier analyses were
accurate at classifying all three groups. This finding sug-
gests that, whereas the overall rates of behavior may be
comparable between control and hippocampus-lesioned
animals, the organization of behavior is not. Overall, group
classification based on the social state data was more
accurate than classification based on the exploration data,
suggesting the importance of social interactions in the
daily life of the rhesus macaque.
The relatively minor impact of early amygdala damage
on the frequency and duration of specific social behaviors
is remarkable given the impact of damage to the amygdala
in adulthood and how these same animals behave in tests
of nonsocial threat responding at the same time point.
When interacting with novel objects and objects thought
to engender threat responding at 18 months of age,
amygdala-lesioned animals physically explored objects
whereas control animals did not (Bliss-Moreau et al.,
2010, Experiment 2), indicating that amygdala-lesioned
animalsʼ affective processing was perturbed in nonsocial
contexts. This supports the view that the amygdala is
not necessary for social processing per se, but rather
serves a broader function related to evaluating threat.
One notable finding in the present experiments is that
the amygdala-lesioned animals had periods of time during
which they disengaged from their environments as evi-
denced by long durations of inactivity and low frequen-
cies of environmental exploration. This disengagement
occurred while they were in contexts where social en-
gagement was nonexistent (solo observations) or low by
design (during the introduction of novel animals, where
the base rates of behaviors were extremely low for all
animals). One possible explanation for these results is that
socially engaging contexts provide amygdala-lesioned
animals with signals that they use to regulate their
own behavior. This possibility is consistent with findings
that in the presence of “mammal-like” objects (e.g., stuffed
animals with eyes and fur), neonatally amygdala -lesioned
animals were as slow as control animals to retrieve concur-
rently presented food rewards (Bliss-Moreau, Toscano,
et al., 2011). In this view, features of social stimuli (e.g.,
mammalian eyes) might provide animals with early amyg-
dala damage cues that they need to regulate their behavior
in a fashion more comparable to control animals.
The idea that social context allows amygdala-lesioned
animals to regulate their behavior is consistent with the
idea that a rich early social environment may be impor-
tant for ameliorating the impact of early damage on social
behavior. Other laboratories that have conducted similar
experiments, but have isolate-reared or peer-reared (rather
than mother-reared) their subjects, have found that early
perturbations in social behavior persist across the de-
velopmental
trajectory (e.g., Málková, Mishkin, Suomi,
& Bachevalier, 2010; Bachevalier et al., 2001; Thompson
& Towfighi, 1976; Thompson et al., 1969). Similarly, social
isolation exacerbates the impact of neonatal damage to the
rat amygdala, such that amygdala-lesioned animals that
spend time socially isolated when they were young spent
less time interacting with peers than amygdala-lesioned
animals raised socially later in life (Diergaarde, Gerrits, Stuy,
Spruijt, & van Ree, 2004). In this view, mother-rearing in
combination with early group socialization and then perma-
nent group housing may have potentiated brain plasticity
resulting in varied patterns of socioaffective behavior across
the developmental trajectory. Investigating the impact of
social environment on neural and behavioral plasticity is
an important area for further research.
The present findings, in concert with experimental evi-
dence collected in other laboratories and from the current
experimental group, illustrate that the impact of neonatal
brain damage may differ from that of brain damage during
adulthood and that such impact may be brain region spe-
cific. For example, neonatal damage to the medial-temporal
lobe (including the amygdala, entorhinal cortex, hippo-
campal formation, and parhippocampal gyrus) prevents
prefrontal down-regulation of striatal dopamine release
(Saunders, Kolachana, Bachevalier, & Weinberger, 1998)
and reduces the binding of a dopamine antagonist to
D2 receptors in the striatum (Heinz et al., 1999), although
damage to the same structures in adulthood does not
alter dopamine regulation. In some cases, putative func-
tions of a structure can be completely accommodated
for during neural development. For example, animals
that receive damage to the hippocampus as adults are
unable to use spatial relational cues to locate food rewards
(Banta Lavenex, Amaral, & Lavenex, 2006), yet subjects
that received bilateral hippocampus lesions as neonates,
like sham-operated control animals, are able to use these
cues to locate food rewards (Lavenex, Banta Lavenex, &
Amaral, 2007). Further evidence for neural reorganization
following early damage in these animals comes from a
functional neuroimagining study conducted approximately
2 years after the present experiments (Machado, Snyder,
et al., 2008). Resting state glucose metabolism was indexed
using PET imagining. Compared with control animals,
amygdala-lesioned animals had greater glucose metabo-
lism in the cerebellum, but lower glucose metabolism
in the orbital, ventromedial, and dorsolateral frontal cor-
tices and in the ACC as well as in the caudate nucleus and
hippocampus. Hippocampus-lesioned animals only dif-
fered from controls in terms of lower glucose metabolism
in the retrosplenial cortex. Taken together, these findings
suggest that the behavioral and neural consequences of
early brain damage may vary based on the area damaged.
Our findings speak to the longstanding debate about
whether phenotypic recovery is better following brain
2138
Journal of Cognitive Neuroscience
Volume 25, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
damage that occurs early as compared with late in life
(see Kolb, 2010, for a review). Although our amygdala-
lesioned subjectsʼ social behavior did differ from control
animals, their heightened affiliative signaling and mildly
reduced propensity for social interaction is not particu-
larly remarkable given the robust social behavior deficits
observed in macaques that receive amygdala lesions as
adults. Damage-related variation in social behavior appears
to have been partially ameliorated with age and potentially
as a result of housing our subjects in fulltime social groups
before the start of these experiments. The question remains
whether neurodevelopment that occurs after this time
point will continue to accommodate early damage and
therefore further ameliorate variation in social behavior
and threat responding, rendering the amygdala-lesioned
animals more “normal” (i.e., like controls) at later points
in development. Future experiments will explore whether
early damage-related variation in social behavior may
emerge at various critical points in development such as
puberty and sexual maturity.
Acknowledgments
This research was supported by funding from the National
Institutes of Mental Health (R37MH57502 to D. G. A.) and by
the base grant of the California National Primate Research
Center (RR00169). E. B. M. was supported by F32MH087067.
This work was also supported through the Early Experience and
Brain Development Network of the MacArthur Foundation. We
thank the veterinary and husbandry staff of the California Na-
tional Primate Research Center for excellent care of the animal
subjects. We thank Dr. Pierre Lavenex, Jeffrey Bennett, and
Pamela Tennant for assistance with surgical procedures and
Dr. Christopher Machado for comments on an earlier draft.
Reprint requests should be sent to Eliza Bliss-Moreau, Depart-
ment of Psychiatry and Behavioral Sciences, California National
Primate Research Center, University of California, Davis, One
Shields Ave., Davis, CA 95616, or via e-mail: eblissmoreau@
ucdavis.edu or David G. Amaral, The M.I.N.D. Institute, University
of California, Davis, 2825 50th St., Sacramento, CA 95817, or
via e-mail: dgamaral@ucdavis.edu.
REFERENCES
Aggleton, J. P., & Passingham, R. E. (1981). Syndrome produced
by lesions of the amygdala in monkeys (Macaca mulatta).
Journal of Comparative and Physiological Psychology,
95, 961–977.
Altmann, J. (1974). Observational study of behavior: Sampling
methods. Behavior, 49, 227–267.
Amaral, D. G. (2006). The amygdala, social behavior, and
danger detection. Annals of the New York Academy of
Sciences, 1000, 337–347.
Antoniadis, E. A., Winslow, J. T., Davis, M., & Amaral D. G.
(2007). Role of the primate amygdala in fear-potentiated
startle: Effects of chronic lesions in the rhesus monkey.
Journal of Neuroscience, 27, 7386–7396.
Antoniadis, E. A., Winslow, J. T., Davis, M., & Amaral D. G.
Bachevalier, J. (1994). Medial temporal lobe structures and
autism: A review of clinical and experimental findings.
Neuropsychologia, 32, 627–648.
Bachevalier, J., Málková, L., & Mishkin, M. (2001). Effects of
selective neonatal temporal lobe lesions on socioemotional
behavior in infant rhesus monkeys (Macaca mulatta).
Behavioral Neuroscience, 115, 545–559.
Banta Lavenex, P., Amaral, D. G., & Lavenex, P. (2006).
Hippocampal lesion prevents spatial relational learning in
adult macaque monkeys. Journal of Neuroscience, 26,
4546–4558.
Bauman, M. D., Lavenex, P., Mason, W. A., Capitanio, J. P.,
& Amaral, D. G. (2004a). The development of social
behavior following neonatal amygdala lesions in rhesus
monkeys. Journal of Cognitive Neuroscience, 16,
1388–1411.
Bauman, M. D., Lavenex, P., Mason, W. A., Capitanio, J. P., &
Amaral, D. G. (2004b). The development of mother-infant
interactions after neonatal amygdala lesions in rhesus
monkeys. Journal of Neuroscience, 24, 711–721.
Bauman, M. D., Toscano, J. E., Babineau, B. A., Mason, W. A.,
& Amaral, D. G. (2008). Emergency of stereotypies in
juvenile monkeys (Macaca mulatta) with neonatal
amygdala or hippocampus lesions. Behavioral
Neuroscience, 122, 1005–1015.
Bauman, M. D., Toscano, J. E., Mason, W. A., Lavenex, P., &
Amaral, D. G. (2006). The expression of social dominance
following neonatal lesions of the amygdala or hippocampus
in rhesus monkeys (Macaca mulatta). Behavioral
Neuroscience, 120, 749–760.
Bliss-Moreau, E., Bauman, M. D., & Amaral, D. G. (2011).
Neonatal amygdala lesions result in globally blunted adult
affect in adult rhesus macaques. Behavioral Neuroscience,
125, 848–858.
Bliss-Moreau, E., Toscano, J. E., Bauman, M. D., Mason, W. A., &
Amaral, D. G. (2010). Neonatal amygdala or hippocampus
lesions influence responsiveness to objects. Developmental
Psychobiology, 52, 487–503.
Bliss-Moreau, E., Toscano, J. E., Bauman, M. D., Mason, W. A., &
Amaral, D. G. (2011). Neonatal amygdala lesions alter
responsiveness to objects in juvenile rhesus macaques.
Neuroscience, 178, 123–132.
Chudasama, Y., Izquierdo, A., & Murray, E. A. (2009). Distinct
contributions of the amygdala and hippocampus to fear
expression. European Journal of Neuroscience, 30,
2327–2337.
Diergaarde, L., Gerrits, M. A. F. M., Stuy, A., Spruijt, B. M., &
van Ree, J. M. (2004). Neonatal amygdala lesions and juvenile
isolation in the rate: Differential effects on locomotor and
social behavior in later life. Behavioral Neuroscience, 118,
298–305.
Emery, N. J., Capitanio, J. P., Mason, W. A., Machado, C. J.,
Mendoza, S. P., & Amaral, D. G. (2001). The effects of
bilateral lesions of the amygdala on dyadic social interactions
in rhesus monkeys (Macaca mulatta). Behavioral
Neuroscience, 115, 515–544.
Heinz, A., Saunders, R. C., Kolachana, B. S., Jones, D. W.,
Gorey, J. G., Bachevalier, J., et al. (1999). Striatal dopamine
receptors and transporters in monkeys with neonatal
temporal limbic damage. Synapse, 32, 71–79.
Izquierdo, A., Suda, R. K., & Murray, E. A. (2005).
Comparison of the effects of bilateral orbital prefrontal
cortex lesions and amygdala lesions on emotional
responses in rhesus monkeys. The Journal of
Neuroscience, 25, 8534–8542.
(2009). The nonhuman primate amygdala is necessary for the
acquisition but not the retention of fear potentiated startle.
Biological Psychiatry, 65, 241–248.
Kling, A. (1974). Differential effects of amygdalectomy in male
and female nonhuman primates. Archives of Sexual
Behavior, 3, 1974.
Bliss-Moreau et al.
2139
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Kling, A., & Green, P. C. (1967). Effects of neonatal
amygdalectomy in the maternally reared and maternally
deprived macaque. Nature, 213, 742–743.
Kolb, B. (2010). Effects of neonatal medial versus lateral
temporal cortex injury: Theoretical comment on Málková
et al. (2010). Behavioral Neuroscience, 124, 873–876.
Lavenex, P., Banta Lavenex, P., & Amaral, D. G. (2007). Spatial
relational learning persists following neonatal lesions in
macaque monkeys. Nature Neuroscience, 10, 234–239.
Machado, C. J., & Bachevalier, J. (2006). The impact of selective
amygdala, orbital frontal cortex, or hippocampal formation
lesions on established social relationships in rhesus monkeys
(Macaca mulatta). Behavioral Neuroscience, 120, 761–786.
Machado, C. J., Emery, N. J., Capitanio, J. P., Mason, W. A.,
Mendoza, S. P., & Amaral, D. G. (2008). Bilateral neurotoxic
amygdala lesions in rhesus monkeys (Macaca mulatta):
Consistent pattern of behavior across different social
contexts. Behavioral Neuroscience, 22, 251–266.
Machado, C. J., Kazama, A. M., & Bachevalier, J. (2009). Impact
of amygdala, orbitofrontal, or hippocampal lesions on threat
avoidance and emotional reactivity in nonhuman primates.
Emotion, 9, 147–163.
Machado, C. J., Snyder, A. Z., Cherry, S. R., Lavenex, P., &
Amaral, D. G. (2008). Effects of neonatal amygdala or
hippocampus lesions on resting brain metabolism in the
macaque monkey: A microPET imagining study.
Neuroimage, 15, 832–846.
Málková, L., Mishkin, M., Suomi, S. J., & Bachevalier, J. (2010).
Long-term effects of neonatal medial temporal ablations on
socieoemotional behavior in monkeys (Macaca mulatta).
Behavioral Neuroscience, 124, 742–760.
Mason, W. A., Capitanio, J. P., Machado, C. J., Mendoza, S. P., &
Amaral, D. G. (2006). Amygdalectomy and responsiveness
to novelty in Rhesus monkeys (Macaca mulatta): Generality
and individual consistency of effects. Emotion, 6, 73–81.
Meunier, M., Bachevalier, J., Murray, E. A., Málková, L., &
Mishkin, M. (1999). Effects of aspiration versus neurotoxic
lesions of the amygdala on emotional responses in monkeys.
European Journal of Neuroscience, 11, 4403–4418.
Mirsky, A. F. (1960). Studies of the effects of brain lesions on
social behavior in Macaca mulatta: Methodological and
theoretical considerations. Annals of the New York Academy
of Sciences, 85, 785–794.
Noldus, L. P. (1991). The observer: S software system for
collection and analysis of observational data. Behavior
Research Methods, Instruments and Computers, 23,
415–429.
Prather, M. D., Lavenex, P., Mauldin-Jourdain, M. L.,
Mason, W. A., Capitanio, J. P., Mendoza, S. P., et al. (2001).
Increased social fear and decreased fear of objects in
monkeys with neonatal amygdala lesions. Neuroscience,
106, 653–658.
Rosvold, H. E., Mirsky, A. F., & Pribram, K. H. (1954). Influence
of amygdalectomy on social behavior in monkeys. Journal
of Comparative and Physiological Psychology, 47, 173–178.
Saunders, R. C., Kolachana, B. S., Bachevalier, J., & Weinberger,
D. R. (1998). Neonatal lesions of the medial temporal lobe
disrupt prefrontal cortical regulation of striatal dopamine.
Nature, 393, 169–171.
Stefanacci, L., Clark, R. E., & Zola, S. M. (2003). Selective
neurotoxic amygdala lesions in monkeys disrupt reactivity
to food and object stimuli and have limited effects on
memory. Behavioral Neuroscience, 117, 1029–1043.
Thompson, C. I., Schwartzbaum, J. S., & Harlow, H. F. (1969).
Development of social fear after amygdalectomy in infant
rhesus monkeys. Physiology & Behavior, 4, 249–254.
Thompson, C. I., & Towfighi, J. T. (1976). Social behavior of
juvenile rhesus monkeys after amygdalectomy in infancy.
Physiology & Behavior, 17, 831–836.
Zola-Morgan, S., Squire, L. R., Alverez-Royo, P., & Clower,
R. P. (1991). Independence of memory functions
and emotional behavior: Separate contributions of
the hippocampal formation and the amygdala.
Hippocampus, 1, 207–220.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
5
/
1
2
2
5
/
2
1
1
2
2
/
4
2
1
1
9
2
4
4
6
/
0
1
8
1
7
7
o
9
c
9
n
3
_
2
a
/
_
j
0
o
0
c
4
n
8
3
_
a
p
_
d
0
0
b
4
y
8
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
2140
Journal of Cognitive Neuroscience
Volume 25, Number 12