The Effects of Frequency, Variability, and Co-occurrence
on Category Formation in Neural Systems
Daniel J. Plebanek and Karin H. James
Abstract
■ Objects are grouped into categories through a complex
combination of statistical and structural regularities. We sought
to better understand the neural responses to the structural
features of object categories that result from implicit learning.
Adult participants were exposed to 32 object categories that
contained three structural properties: frequency, variability,
and co-occurrences, during an implicit learning task. After this
exposure, participants completed a recognition task and were
then presented with blocks of learned object categories during
fMRI sessions. Analyses were performed by extracting data from
ROIs placed throughout the fusiform gyri and lateral occipital
cortex and comparing the effects of the different structural prop-
erties throughout the ROIs. Behaviorally, we found that symbol
category recognition was supported by frequency, but not vari-
ability. Neurally, we found that sensitivity to object categories
was greater in the right hemisphere and increased as ROIs were
moved posteriorly. Frequency and variability altered the brain
activation while processing object categories, although the pres-
ence of learned co-occurrences did not. Moreover, variability
and co-occurrence interacted as a function of ROI, with the
posterior fusiform gyrus being most sensitive to this relation-
ship. This result suggests that variability may guide the learner
to relevant co-occurrences and this is supported by the posterior
ventral temporal cortex. Broadly, our results suggest that the
internal features of the categories themselves are key factors
in the category learning system. ■
INTRODUCTION
As we encounter objects in our environment, we implic-
itly group them into categories. The ability to form cate-
gories of objects that are similar in a given dimension or
dimensions organizes and simplifies our knowledge.
Categorization also allows us to understand new objects
by associating them with known objects. However, un-
derstanding how we initially form categories of objects,
how category boundaries are defined, and the dynamic
nature of categories remains elusive. Understanding cat-
egory formation is further complicated by varying theo-
ries regarding the subcomponents that underlie the
structure of the categories themselves.
Perhaps the simplest way to consider category learning
is the situation where we learn the name of a new object
based on its visual appearance. In doing so, we extract
information from new events that has commonalities
and differences with previous events. For example, visual
statistical learning allows for the linking of co-occurrences
such as an object and its name. Research has demon-
strated that both infants and adults have powerful statis-
tical tracking mechanisms that allow them to overcome
ambiguity in an environment and link word–object pairs
based on these probabilistic regularities (Smith & Yu,
2008; Yu & Smith, 2007).
Indiana University, Bloomington
© 2021 Massachusetts Institute of Technology
Much of our focus in studying category learning is on
the capabilities of the learner. For example, the learner
acquires some categories by producing them by hand
( Vinci-Booher & James, 2020; Vinci-Booher, Cheng, &
James, 2019; James, 2017; James & Engelhardt, 2012)
or by physically exploring the category (Slone, Smith, &
Yu, 2019; James, Jones, Swain, Pereira, & Smith, 2014;
James & Swain, 2011; James, 2010). Nonetheless, the
learner is only one piece of the puzzle in this system.
Just as the learner has limitations and competencies that
interact with the environment to support learning, object
categories themselves also have properties that are
worthwhile to study. There is some evidence that the sta-
tistical, internal properties of categories themselves influ-
ence learning. For example, category structures such a
density and sparsity are known to affect the ease of acqui-
sition of categories, with dense categories (i.e., those
with many predictive features such as cats and dogs)
being developmentally easier to acquire than sparse cat-
egories (i.e., those with more deterministic boundaries
that have specific and necessary prerequisites like the
concept “electron”; Sloutsky, 2010; Kloos & Sloutsky,
2008). If category structure has an effect on learning mea-
sured with overt behavioral responses, then neural sys-
tems must also show a sensitivity to category structure.
A significant amount of neuroimaging research has
been devoted to understanding how categories of objects
are processed in the brain (for a review, see Grill-Spector
Journal of Cognitive Neuroscience 33:8, pp. 1397–1412
https://doi.org/10.1162/jocn_a_01738
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
& Weiner, 2014). This body of research has focused on
visual processing in the ventral temporal cortex ( VTC), a
broad neural system that has been shown to process
object properties in a nested, hierarchical manner (Grill-
Spector & Weiner, 2014). This research, however, has
focused predominately in the object properties them-
selves (shape, color, size; e.g., Vinberg & Grill-Spector,
2008), as well as the “level” of categorization that is
required of a given task (individual level, subordinate or
superordinate; e.g., Grill-Spector, Knouf, & Kanwisher,
2004). In contrast, how the structure of the category itself
affects processing in the VTC is still an open question. That
is, the exemplars in a given category have a relationship to
one another in the context of the category. When a cate-
gory is learned, there may be exemplars that occur more
frequently during learning events, but it is not known
whether the VTC is sensitive to how frequently an exem-
plar occurs within a category. Furthermore, a category may
be composed of exemplars that vary in terms of their
similarity to one another: Some categories contain highly
variable exemplars, whereas other categories may contain
exemplars that are visually very similar, even within the
same level of categorization (as defined by Rosch, Mervis,
Gray, Johnson, & Boyes-Braem, 1976). In addition, objects
rarely occur in isolation (except in experimental setups), and
the visual system is sensitive to co-occurrences of objects
both spatially and temporally (e.g., Turk-Browne, Scholl,
Chun, & Johnson, 2009). Thus, structural properties of
categories may affect overt learning and recruit different
levels of processing in the visual system. Below, we review
these three structural features of categories—frequency,
variability, and co-occurrence—that past research has
suggested play roles in learning. We then discuss an
experiment that investigates the neural basis for these
structural features after learning novel object categories.
Object Frequency
One potentially important structural feature of object cat-
egories is the frequency by which individuals encounter
category exemplars in their environment. Recent evidence
from the home environments of infants revealed that a
small number of objects were extremely frequent, demon-
strating that the distribution of visual objects in the real
world may be highly skewed (Clerkin, Hart, Rehg, Yu, &
Smith, 2017). Interestingly, the highly frequent objects
were the normatively acquired, first learned words
(Smith, Jayaraman, Clerkin, & Yu, 2018; Clerkin et al.,
2017). In behavioral categorization tasks, classification
accuracy has been found to be higher for high-frequency
exemplars (Nosofsky, 1988). Furthermore, past research
has speculated that frequency is largely related to object
typicality (Rosch & Mervis, 1975).
Recent evidence examining typicality effects in the
brain have revealed a role of object-selective brain regions
in processing typical and atypical exemplars. Specifically,
Iordan, Greene, Beck, and Fei-Fei (2016) used brain
imaging techniques to better understand neural represen-
tation of natural categories such “fish” and “dogs” that
contained exemplars that varied across typicality judg-
ments. Although Iordan et al. (2016) did not explicitly
test exemplar frequency, typicality may be a proxy for fre-
quency as the two constructs have a mutual relationship
where they alter our perception of category exemplars
(Nosofsky, 1988). Through multivoxel pattern analysis,
Iordan et al. found that atypical exemplars (e.g., a puffer-
fish) produced activation patterns that were different from
the central tendencies of other, more common category
members (e.g., a clownfish) through the lateral occipital
complex (LOC). Furthermore, this pattern was not present
in early visual areas suggesting that this effect was not
driven by lower-level, perceptual features. Thus, the brain
may process statistical information such as exemplar
frequency (although frequency has not been directly
tested) and typicality and this may be driven by the LOC.
Object Variability
The variability or distribution of exemplars within an object
category has also been noted as having impacts on learning.
For example, exposure to variable symbol forms such as
handwritten symbols results in greater categorization ability
compared to exposure to highly similar visual outputs such
as typed or traced symbols (Li & James, 2016). Similarly,
adults demonstrate faster object recognition if they were
exposed to an object from multiple, randomly sampled
viewpoints (high visual variability) as opposed to objects
that were observed from spatially continuous viewpoints
(low visual variability; Harman & Humphrey, 1999). The
underlying hypothesis is that variability may allow individ-
uals to extrapolate the central features that allow them to
form generalizable biases to support future learning
(Perry, Samuelson, Malloy, & Schiffer, 2010).
Recent evidence examining the neural mechanisms of
both letter perception and category learning have also
identified brain regions associated with variability.
Specifically, Vinci-Booher and James (2020) found that
young children who are still learning letters demonstrated
greater activation in the left middle fusiform gyrus
(FFG) for handwritten forms compared to typed letters.
However, older children and adults who were literate
did not show this effect. Vinci-Booher and James (2020)
interpreted this finding as variability contributing to initial
category formation. Thus, older children and adults did
not show this effect because they already had expert
knowledge of letter categories. Recent evidence examin-
ing novel category learning in older children and adults
have identified similar findings. Specifically, Plebanek
and James (2021) found that 8-year-olds and adults
demonstrated greater activity associated with the right
posterior fusiform gyrus when learning variable com-
pared to tight categories. Furthermore, these researchers
also found that variability led to activity associated with
the fusiform gyri driven by an invariant feature that
1398
Journal of Cognitive Neuroscience
Volume 33, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
defined the category, whereas tightly organized categories
led to responses based on overall similarity (but without a
consistent feature). Thus, the brain appears to respond to
variability among category members and the fusiform
gyrus, in particular, is highly involved in this process.
It is worth noting, however, that variability may not be
entirely beneficial for learning. Work examining chicks’
development of object invariance have suggested that
small, tightly confined viewpoint changes in an object
may be sufficient to support invariance development
( Wood, 2016). Similarly, infants’ object recognition
appears to be improved when object movement is restricted
so that they are observed from a fewer number of view-
points (Kraebel & Gerhardstein, 2006). Thus, although there
are both behavioral and brain-based effects from varying
object category exemplars, whether or not variability is
beneficial to learning remains controversial.
Objects and Co-occurrences
At the broadest level, categories, objects, and other infor-
mation present in the world is full of probabilistic regular-
ities that the learner can use to predict future events. The
brain is capable of extracting these regularities from a
young age (Saffran, Aslin, & Newport, 1996). In fact, the
brain is so attuned to these regularities that there is activa-
tion throughout category-relevant visual areas in the ven-
tral temporal cortex even though individuals do not
explicitly recall the regularities (Turk-Browne et al.,
2009). These processes may be important in formulating
links among features, dimensions spanning values of
features, or objects that co-occur across space and time.
These co-occurrences may then be the building blocks
of representations as measured by neural instantiations
of object knowledge (see Sherman, Graves, & Turk-
Browne, 2020, for a review). The neural representations
of objects appear to be driven, in part, by incidental
co-occurrences across time and space. For example,
Schapiro, Kustner, and Turk-Browne (2012) discovered
the patterns of neural representations throughout the me-
dial temporal lobe for novel objects were more similar
when the objects occurred together in time. Similar mech-
anisms may underlie object recognition more broadly. For
example, objects are organized by temporal structure that
links multiple features of objects across different views to
create composite object representations (see Wallis &
Bülthoff, 1999, for a review). This co-occurrence structure
is commonly associated with the VTC and may explain how
features play a role in object recognition ( Wallis &
Bülthoff, 1999). More specific mapping of spatial
co-occurrence sensitivity in the VTC has shown that the
anterior fusiform gyrus responds to co-occurrence more
than posterior VTC structures (Stansbury, Naselaris, &
Gallant, 2013). Therefore, recent neuroimaging work has
pointed to the VTC and specifically the anterior fusiform
gyrus for possible neural mechanisms that are sensitive
and/or support co-occurrences within a category.
This Study
Taken together, these studies shed light on the way the
brain processes regularities as we learn objects. Within a
single object category, these regularities take many forms.
First, at the level of the category, how frequent or typical
an exemplar is in the overall scheme of the category may
influence how it is processed. Second, the variability and
diversity of category members and features can also influ-
ence how a category is learned and generalized. Third,
features may co-occur and predict other features and
category membership. All of these regularities matter
and may guide the learner to a specific representation or
category judgment. Also of note is the regularity by which
we see subregions of the ventral temporal cortex respond
to these three (frequency, variability, and co-occurrences)
structural elements separately, but how the structural ele-
ments interact is unknown (Plebanek & James, 2021;
Iordan et al., 2016; Stansbury et al., 2013; Turk-Browne
et al., 2009).
Taken together, research supports the idea that different
structures within the VTC support different aspects of cate-
gory structure: frequency by the LOC (Iordan et al., 2016),
variability by the middle fusiform gyrus ( Vinci-Booher &
James, 2020), and co-occurrence by the anterior fusiform
gyrus (Stansbury et al., 2013). None of the past work,
however, compared these properties directly within each
of these ROIs. The present work sought to address this
gap in the literature.
Therefore, we were interested in two main questions:
(1) How does the brain process three different structural
properties that are relevant for learning new object cate-
gories: frequency, variability, and co-occurrences among
features? (2) Are these structural properties differentiated
from one another as reflected by differences in neural
responses in specified ROIs?
To answer these questions, we created a metrically
organized set of novel categories that allowed us control
these three structural elements. Participants were exposed
to the object categories over two days and then underwent
two MRI sessions that measured the brain responses to
the object categories. Given that frequency, variability,
and co-occurrence all affect object learning and have
been shown individually to recruit different regions
within the VTC, we expected to see preference in certain
ROIs for the different types of structure, but not exclu-
sivity in relative responses.
METHODS
Participants
Seventeen literate English-speaking adults (M = 23.9 years,
range = 3.3 years, 7 men) completed this study. Participants
were graduate and undergraduate students from a small,
Midwestern town and were recruited through word of
mouth. All participants were right-handed and were
screened for neurological trauma, developmental
Plebanek and James
1399
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
disorders, and magnetic resonance (MR) contraindica-
tions. Three additional participants were excluded for
the following reasons: One did not complete the study,
and two were excluded for excessive motion. All partici-
pants provided informed consent in accordance with the
Indiana University institutional review board. Participants
received $10 for each behavioral session and $25 for each
MRI session. For completing all sessions, they received a
$20 bonus, resulting in a total of $90.
Materials
A set of 90 novel object categories defined by shape were
created for this study (see Figure 1 for examples). The
objects were multistroke two-dimensional letter-like
symbols that were similar to sets previously used in
novel-object learning experiments (e.g., James & Atwood,
2009). This set was used for ease of manipulation of cate-
gory structure while still maintaining the complexity of
naturally occurring categories such as symbols and letters
(see Figure 1). They were constructed with a computer
drawing program and were composed of strokes that
occur in written letters. Thirty-two of these object catego-
ries were present during training. The remaining 58 object
categories were reserved for new categories during the
MRI sessions or the recognition test. Object categories
were composed of symbols that varied in size and color
(see Figure 1), which are labeled here as object features.
Both size and color varied metrically across 12 steps
(Figure 2). The smallest size value was 50 × 50 pixels,
and each step increased size by 25 pixels, with the largest
value ( Value 12) being approximately 325 × 325. The first
color value in red, green, blue coordinates was [255 122
122] and was a pink color. The R value incrementally
changed by −22 in each metric step until the red, green,
blue values were [13 122 122] and was a teal color.
Figure 1. Object category shapes grouped by condition. Groups of
shapes (rows) were randomly assigned to a condition defined by
frequency and variability. Symbol assignments to conditions were
counterbalanced across participants.
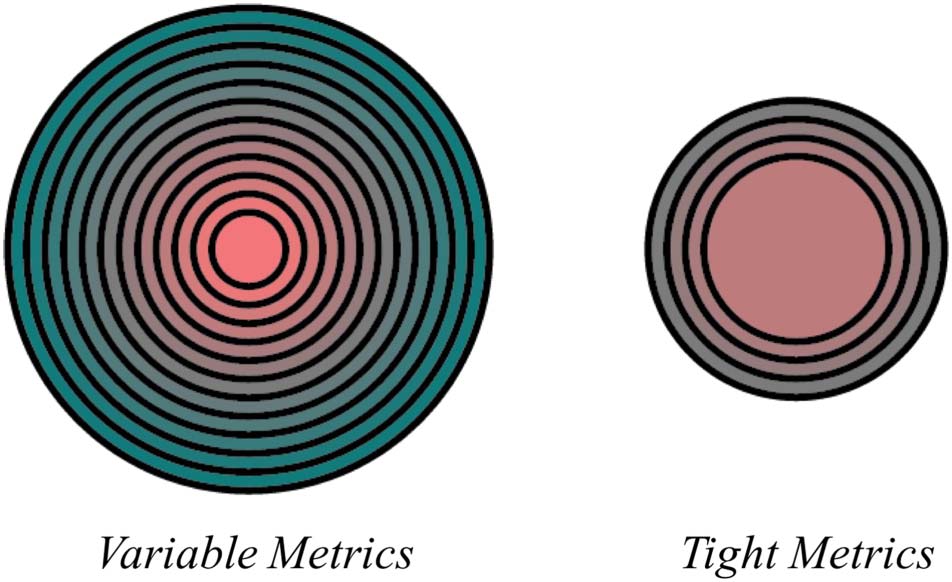
Figure 2. Size and color metric dimensions for variable (left) and tight
(right) variability structures. The center of the variable dimension
graphic is size and color value one. The furthest circle is size and color
Value 12. The center of the tight dimension graphic is size and color
value four. The furthest circle is size and color seven.
Furthermore, each object category was organized
according to three structural properties: frequency of
identical exemplars, variability among members, and
co-occurrence between features. Frequency was defined
as either high or low frequency depending on how many
times a particular exemplar from the object category was
presented during training. For high-frequency categories,
identical exemplars were presented a total of 140 times
across all training blocks. For low-frequency categories,
identical exemplars were presented a total of 40 times
across all training blocks. Variability was defined as the
distribution of features individuals saw during training
and the fMRI sessions. For training, tight categories’ fea-
ture values were only four through seven for the color
and size. For variable categories, feature values were a
broader distribution [1, 2, 3, 5, 7, 10, 11, 12] of the object
features. During the MR session, tight and variable catego-
ries were presented with the same distribution as training.
Finally, co-occurrence reflected the pairing of the values
of the features during training. During training, each
feature value was linked so that a person saw the same
numerical value for both size and color (i.e., if they saw a
value four for size, it was also a value four for color). During
the MR session, some of the blocks were unlinked—the
features of color and size were randomly paired.
Design
Object categories were then randomly assigned to a con-
dition based on these structural properties so that each
condition contained eight different object categories.
During implicit learning, participants saw four conditions:
[high, variable, linked], [low, variable, linked], [high, tight,
linked], [low, tight, linked]. During MRI, participants saw
these four conditions as well as their unlinked counter-
parts: [high, variable, unlinked], [low, variable, unlinked],
[high, tight, unlinked], [low, tight, unlinked]. Participants
1400
Journal of Cognitive Neuroscience
Volume 33, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
also saw a ninth condition consisting of new items that
were both variable and linked [new, variable, linked].
Finally, during the recognition task presented after
training, participants saw black symbols for each category
that were from Size Value 6. This was required so that
participants would not continue to associate the feature
co-occurrences after training. This resulted in a design
with the following factors and levels: frequency (levels:
high and low), variability (levels: variable and tight),
co-occurrence (levels: linked and unlinked). These factors
were combined to create eight conditions in a 2 (frequency:
high vs. low) × 2 (variability: tight vs. variable) × 2 (co-
occurrence: linked vs. unlinked) repeated-measures
design. In addition, the [new variable linked] condition
was used to examine learning and novelty effects.
Procedure
The study was completed over 4 days. During the first
day, participants completed three blocks of an implicit
training task, reflecting our interest in statistical learning.
During the second day, participants completed two
blocks of implicit training and a recognition task. The
third and fourth days consisted of MRI sessions. All ses-
sions are explained in detail below.
Implicit Training Sessions
After providing informed consent, participants were taken
to a quiet room. They were told that they were going to be
seeing some novel symbols, two-at-a-time on a computer
screen. If the participant thought the symbols were the
same, they were told to press the number “1” on the
keyboard. If they were different, they were told to press
“0.” Participants were explicitly told that there was no
correct answer to this task and to simply use their best
judgment. Therefore, any learning that occurred would
be a result of this implicit task. Although there is contro-
versy as to whether category learning should be studied
through explicit or implicit tasks (see Ashby & Valentin,
2017), we chose to use an implicit task because of our
interest in statistical learning and from demonstrations
that category learning often proceeds in this manner
(e.g., Sherman et al., 2020).
Symbols were presented so that they were vertically cen-
tered with one symbol on the left side of the screen and one
symbol on the right side of the screen (see Figure 3). Once
the symbols appeared, participants were required to wait to
make their judgment for 750 msec until the computer
prompted them for their answer. The symbols and the
prompt remained on the screen until participants re-
sponded. Combinations of symbols were organized so that
pairs matched on object category (shape) on only 28 trials
per block (9.72% of trials). Similarly, features (color and
size) matched across both objects on 28 trials per block.
Therefore, the majority of trials presented unassociated
exemplars. Co-occurrences of size and color were always
linked in this task. During Day 1, participants completed
three blocks. During Day 2, participants completed two
blocks.
Recognition Test
After completing the learning session on Day 2, participants
immediately began the recognition test. Participants were
told that they would see a briefly presented symbol in the
center of the screen followed by a static Gaussian noise
mask. They were required to press the numeral “1” button
on a keyboard if they had seen the symbol during the
training sessions, and press “0” if they had not. Each symbol
was presented for 150 msec; the mask was presented for
100 msec, followed by a response prompt. There was no
time limit to respond. Response time and sensitivity (hits −
false alarms) were measured.
MRI Sessions
Participants completed two consecutive days of imaging
sessions each lasting 45–60 min. The structure of the two
days was the same with the exception that, on their first
day, a high-resolution anatomical scan was completed prior
to the functional runs. On each day, participants completed
eight functional runs (16 total). The order of these func-
tional runs was randomized across the 2 days.
Each functional run consisted of nine blocks each lasting
20 sec. Blocks contained 20 exemplars of a symbol category,
with each symbol appearing on screen for 800 msec
followed by a 200-msec fixation cross. Thus, each block
consisted of only one object category (see Figure 4).
There was a 10-sec interblock interval that was not
analyzed. There was also a 10-sec rest period at the begin-
ning and end of each run. Thus, runs lasted approximately
4 min 40 sec. The order of blocks within each run was
randomized.
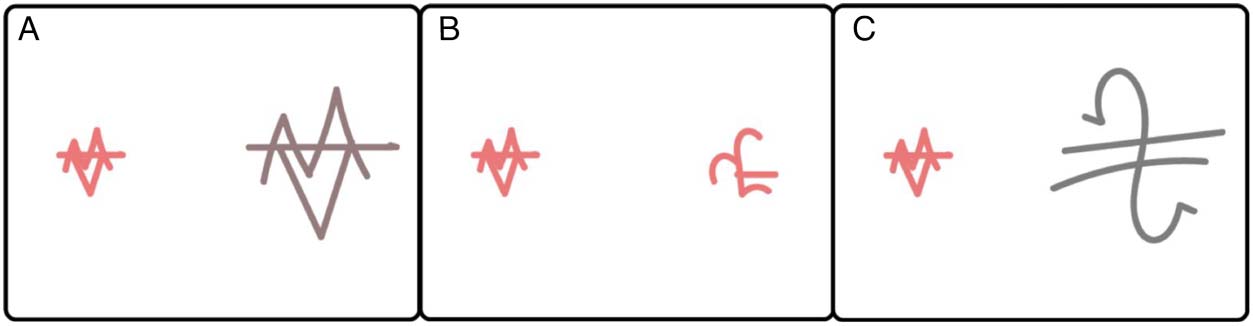
Figure 3. Examples of the trial
types present in the learning
task. (A) represents an object
category (shape) match. (B)
represents a feature match
(color and size). (C) represents
no match.
Plebanek and James
1401
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
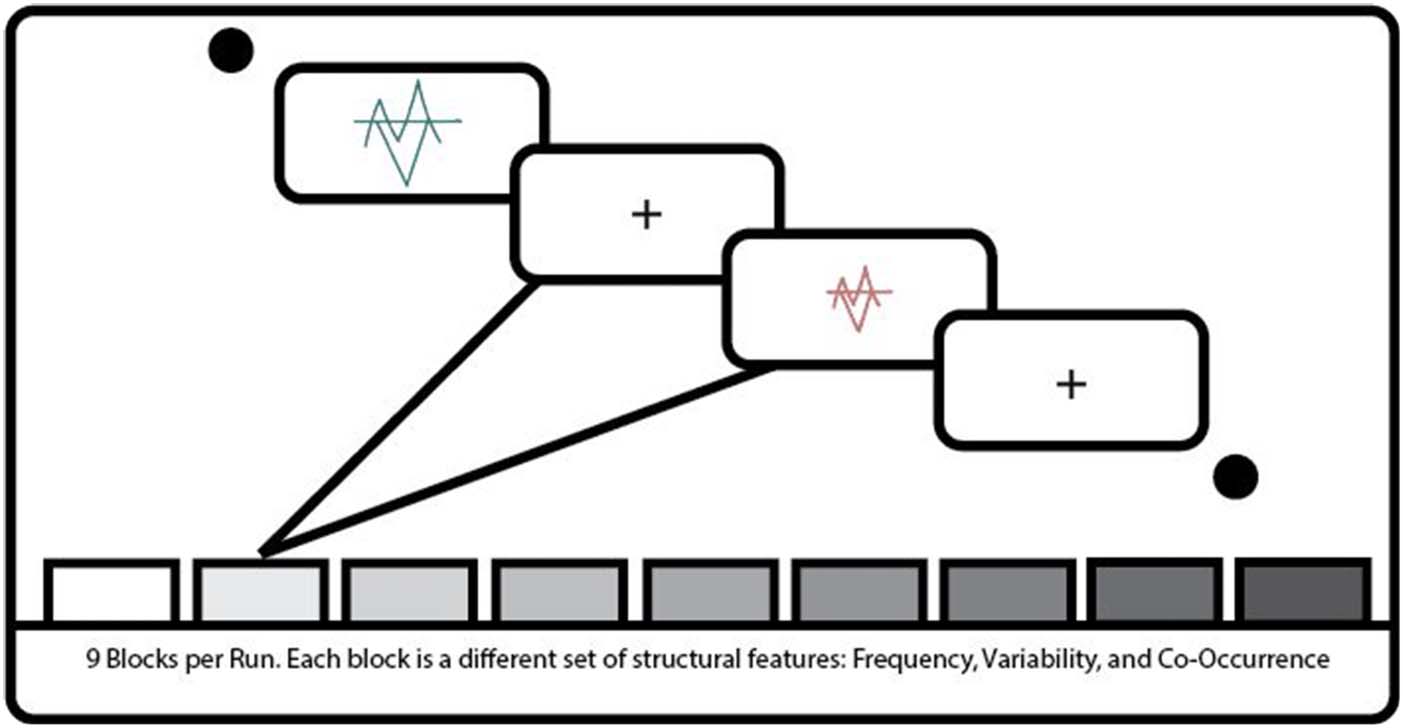
Figure 4. Schematic of the
fMRI paradigm. Participants saw
a single object category per
block. Each block consisted
of 20 presentations of object
category exemplars for
800 msec. Each exemplar
was separated by a 200-msec
fixation cross. Each block in
a run consisted of different
combinations of structural
features.
Learned categories were repeated across the experi-
ment in four blocks. Two blocks contained linked features
whereas two blocks presented unlinked features. New
object categories appeared in a total of two blocks only
and were always linked. Within each individual run, each
training condition appeared once. Each condition was a
separate block with a different object category in each
block.
Scanning Parameters
Neuroimaging was conducted using a Siemens Magnetom
Tim Trio 3-T whole-body MRI system located in the Indiana
University Imaging Research Facility at the Department of
Psychological and Brain Sciences. The high-resolution
T1-weighted anatomical scans were conducted using a
magnetization prepared rapid gradient echo sequence:
inversion time = 900 msec, echo time = 2.98 msec, rep-
etition time = 2300 msec, flip angle = 9°, with 176 sagittal
slices of 1.0-mm thickness, a field of view 256 × 248 mm,
and an isometric voxel of 1.0 mm3. For functional images,
the field of view was 220 × 220 mm, with an in-plane res-
olution of 110 × 110 pixels and 72 axial slices of 2.0-mm
thickness per volume with 0% slice gap, producing an
isometric voxel size of 2.0 mm3. Functional images were
acquired using a gradient echo EPI sequence with inter-
leaved slice order: echo time = 30 msec, repetition time =
2000 msec, flip angle = 52° for BOLD imaging.
Analyses
The main analyses consisted of standard preprocessing
pipeline for fMRI data. Analyses and preprocessing were
conducted using BrainVoyager v20.6 (Brain Innovation).
Preprocessing and Motion Correction
Each individual’s anatomical volumes were standardized to
Talairach space (Talairach & Tournoux, 1988). Preprocessing
of function volumes included slice-time correction, 3-D
motion correction using trilinear, sinc-interpolation, and
3-D Gaussian spatial smoothing at an FWHM of 6 mm.
Temporal high-pass filtering was also used with a voxel-
wise general linear model (GLM) that included a Fourier
basis with a cutoff of two sine/cosine pairs and a linear
trend predictor. A rigid body transformation was used to
coregister anatomical and functional volumes. To account
for head motion, rigid body transformation parameters
were added to the study design matrix as predictors of
no interest (Bullmore et al., 1999). As previously men-
tioned, two participants were excluded because of their
motion: one for having multiple runs with motion spikes
greater than 2 mm and one for drifting more than 3 mm
for multiple runs.
Data Analyses
Participants completed 16 functional runs. Fourteen of
these runs were randomly selected for ROIs analyses.
The remaining two runs were selected for a whole-brain
contrast that served to localize the ROIs (thus avoiding
“double-dipping” from the data). Thus, the data were
analyzed using a random-effects GLM using BrainVoyager’s
multisubject GLM module. This whole-brain analysis
served to demarcate broad regions that responded more
to objects compared with fixation (see Figure 5, Table 1).
The resultant regions were then subdivided anatomically
into ROIs for further analyses (Figure 6).
Individual brains were first normalized to the stereotaxic
space of Talairach and Tournoux (1988). After the whole-
brain contrast was performed, we divided the resultant
regions anatomically for subsequent ROI analyses. Three
of these regions corresponded to subdivisions of the fusi-
form gyrus, one corresponded to the LOC, and one served
as a control region in primary visual cortex. To subdivide the
fusiform gyri, we used similar procedures to James and
Engelhardt (2012). On the x dimension, 10 mm was used
because this is the average distance from the lateral occip-
ital sulcus and the collateral sulcus. The fusiform gyrus is
respectively bounded laterally and medially by these
1402
Journal of Cognitive Neuroscience
Volume 33, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
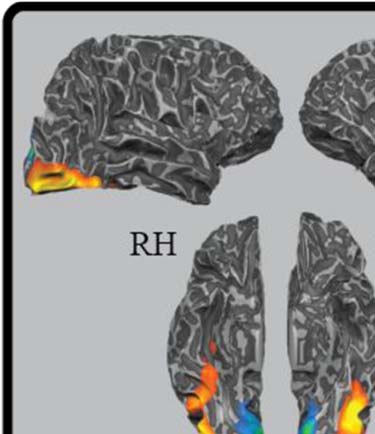
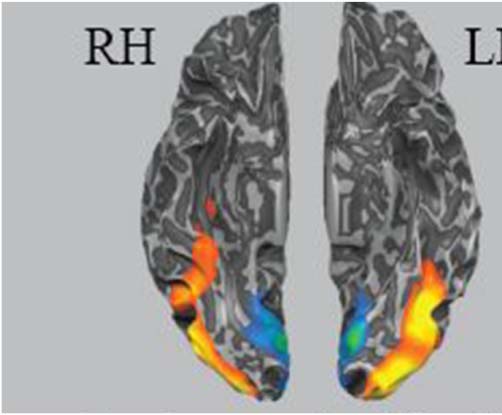
Figure 5. Results from a whole-brain contrast comparing learned object
categories and interblock fixation, p < .001, cluster corrected for six
contiguous voxels.
structures. Within the z dimension, we placed ROIs on the
ventral temporal surface that extended 10 mm dorsally. On
the y dimension, we followed the collateral sulcus
posteriorly, splitting the region in three equal portions.
The resulting ROIs were 10 × 10 × 10 mm3. For the lateral
occipital region, dimensions were kept as 10 × 10 ×
10 mm3 to maintain consistency across ROIs. On the z
dimension, the ROI was placed on the ventral occipital sur-
face and extended 10 mm dorsally. On the y dimension, the
ROI place to was posterior to the previous ROIs. This ROI
was bounded to the lateral occipital sulcus. As with the fusi-
form ROIs, this area largely corresponded to the most poste-
rior region of the brain that responded more to learned
symbols than fixation. The primary visual cortex was localized
in each individual by first locating the broad region that
responded more to fixation than learned symbols. Then,
we anatomically localized the calcarine sulcus, with the
anterior boundary of the ROI specified by the cuneal
point, and then the 10-mm3 voxel was placed posterior to
this within the calcarine folds (Hinds et al., 2008). Given
the large variability in functional localization of area V1, we
assumed that this large anatomically placed ROI would cap-
ture most of primary visual cortex and potentially
visual association areas that surround it. Because of its
role as a control area, individual retinotopic mapping was
not performed. This procedure was carried out for each
individual. Details regarding each individual’s ROIs are
present in Table 2.
Figure 6. Schematic of ROI placement (performed individually, this
depicts average placement). Blue: anterior fusiform; green: middle
fusiform; pink: posterior fusiform; gray: LOC; red: primary visual cortex.
Data were extracted from the primary visual cortex and
compared across conditions, but was not used in the
overall data analyses for the remaining ROIs given that
it was simply used as a control region to determine sen-
sitivity to category structure within a region that would
not be predicted to have such sensitivity. We then ex-
tracted each individual’s data from three ROIs within
the left and right fusiform gyri and one ROI within the
left and right LOC (eight ROIs in total). Average activation
across the time course (excluding first and last three time
points) from these data was used as the dependent measure
in a 2 (Frequency: high vs. low) × 2 (Variability: tight vs.
variable) × 2 (Co-occurrence: linked vs. unlinked) × 4
(Region: anterior FFG vs. mid FFG vs. posterior FFG vs.
LOC) × 2 (Hemisphere: left vs. right) ANOVA was per-
formed on the resultant data. Follow-up analyses on simple
effects and a priori t tests were also conducted.
RESULTS
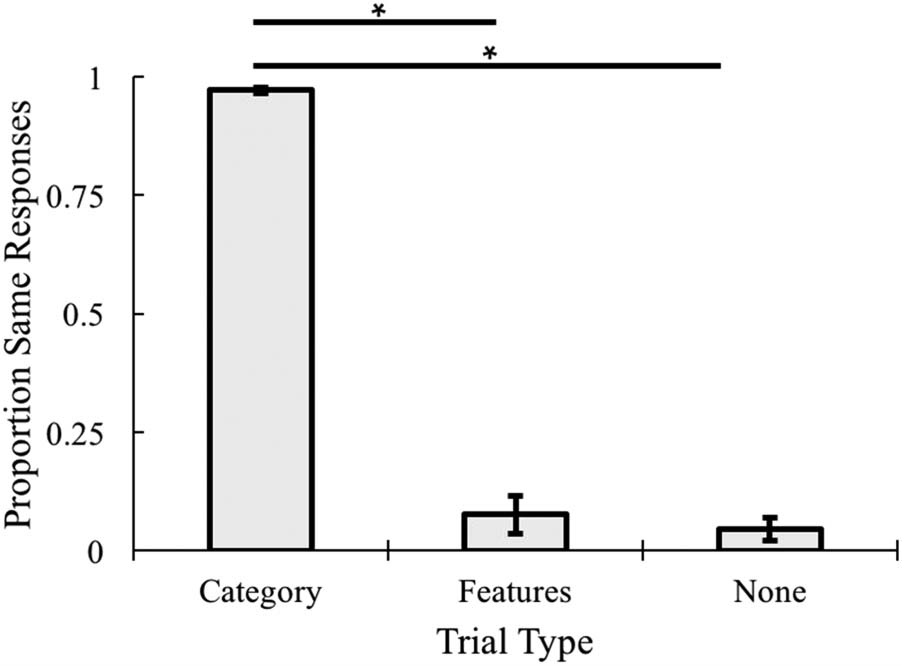
Categorization Performance
Proportions of “same” responses in the learning sessions
was calculated for the three conditions: same object shape,
same features, and no match items (Figure 7). These data
were submitted to a one-way ANOVA. There was a signifi-
cant effect of Trial type, F(2, 32) = 650.41, p < .001, η2 =
.946. Follow-up comparisons revealed that participants
were more likely to categorize objects together when the
object shape matched than when color and size matched,
t(16) = 21.84, p < .001, d = 5.40, or when there was no
Table 1. Region of Interest Localizer Analysis
Contrast
Learned > Fixation
Fixation > Learned
Cluster Size
( Voxels)
21586
21147
1477
Talairach Coordinates
Peak x
Peak y
Peak z
Peak t
Anatomical Location
−42
39
12
−64
−70
94
−20
−20
1
12.70
12.70
10.34
Left ventral temporal
Right ventral temporal cortex
Bilateral lingual gyrus
This table presents cluster sizes, peak coordinates, and peak t values for regions that were significant with our localizer contrast.
Plebanek and James
1403
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 2. Region of Interest Coordinates
Table 2. (continued )
Participant
Region
x-Range
y-Range
z-Range
Participant
Region
x-Range
y-Range
z-Range
TK
LaFFG −44…−35 −41…−32 −21…−12
RLOC
34…43
−81…−73 −15…−6
LmFFG −45…−36 −53…−44 −21…−12
DL
LaFFG −47…−38 −37…−28 −26…−17
LpFFG −42…−33 −65…−56 −21…−12
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−32….23
−92…−84 −21…−12
34…43
34…43
32…41
25…34
−45…−36 −21…−12
−58…−49 −21…−12
−69…−60 −21…−12
−88…−79 −21…−12
VR
LaFFG −35…−26 −43…−35 −22…−13
LmFFG −44…−35 −51…−42 −26…−17
LpFFG −44…−35 −51…−42 −26…−17
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−36…−26 −85…−76 −26…−17
36…45
36…45
36…45
23…30
−39…−30 −27…−18
−53…−44 −27…−18
−67…−58 −27…−18
−85…−76 −27…−17
LmFFG −36…−27 −55…−46 −22…−13
AB
LaFFG −44…−34 −38…−29 −21…−12
LpFFG −30…−21 −60…69
−22…−13
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−30…21
−79…−70 −22…−13
37…46
41…50
38…47
32…41
−36…−45 −22…−13
−55…−46 −22…−13
−66…−57 −22…−13
−86…−77 −22…13
MT
LaFFG −38…−29 −43…−34 −21…−12
LmFFG −41…−32 −51…−42 −21…−12
LpFFG −40…−31 −65…−56 −21…−12
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−34…−25 −83…−74 −23…−14
41…50
38…47
38…47
34…43
−26…−17 −23…−14
−40…−31 −23…−14
−54…−45 −23…−14
−82…−73 −23…−14
LmFFG −36…−27 −54…−46 −21…−12
EM
LaFFG −42…−33 −36…−27 −26…−17
LpFFG −34…−25 −65…−56 −21…−12
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−29…−20 −85…−77 −21…−12
32…41
32…41
28…37
19…28
−44…−35 −24…−15
−56…−47 −24…−15
−69…−60 −24…−15
−91…−82 −24…−15
EC
LaFFG −44…−35 −41…−32 −25…−16
LmFFG −45…−36 −47…−38 −26…−17
LpFFG −44…−35 −62…−54 −26…−17
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−39…−30 −87…−78 −17…−8
37…48
36…45
35…44
30…39
−32…−23 −26…−17
−49…−40 −26…−17
−63…−54 −26…−17
−87…−78 −17…−8
LmFFG −42…−33 −55…−46 −25…−16
BM
LaFFG −42…−33 −38…−29 −26…−17
LpFFG −41…−32 −65…−56 −25…−16
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−32…−23 −90…−81 −25…−16
37…47
38…47
37…46
25…34
−42…−33 −25…−16
−55…−46 −25…−16
−67…−58 −25…−16
−92…−82 −25…−16
JF
LaFFG −28…−19 −42…−33 −16…−7
LmFFG −41…−32 −52…−43 −26…−17
LpFFG −40…−31 −70…−61 −26…−17
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−31…−22 −87…−78 −26…−17
36…45
36…45
34…43
24…33
−37…−28 −26…−17
−42…−50 −26…−17
−63…−54 −26…−17
−84…−75 −22…−13
LmFFG −25…−16 −53…−45 −16…−7
PM
LaFFG −41…−32 −37…−27 −34…−25
LpFFG −24…−15 −66…57
−16…−7
LLOC
RaFFG
RmFFG
RpFFG
−24…−15 −91…−82 −13…−4
41…40
39…48
38…47
−36…−27 −15…−6
−49…40
−15…−6
−63…−54 −15…−6
LmFFG −41…−32 −50…−41 −34…−25
LpFFG −40…−31 −65…−56 −34…−25
LLOC
RaFFG
RmFFG
−34…−25 −86…−77 −34…−25
38…47
38…47
−34…−25 −29…−20
−46…−36 −29…−20
1404
Journal of Cognitive Neuroscience
Volume 33, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
LLOC
RaFFG
RmFFG
RpFFG
RLOC
LLOC
RaFFG
RmFFG
RpFFG
RLOC
Table 2. (continued )
Table 2. (continued )
Participant
Region
x-Range
y-Range
z-Range
Participant
Region
x-Range
y-Range
z-Range
RpFFG
RLOC
38…47
35…44
−57…−48 −29…−20
−80…−71 −29…−20
ML
LaFFG −37…−28 −41…−32 −26…−17
RmFFG
RpFFG
RLOC
35…44
34…43
35…43
−43…−34 −23…−14
−55…−46 −23…−14
−80…−71 −19…−10
LmFFG −36…−27 −55…−47 −26…−17
AM2
LaFFG −38…−29 −43…−34 −21…−12
LpFFG −35…−26 −68…−59 −26…−17
LmFFG −36…−27 −55…−46 −21…−12
−38…−29 −84…−76 −26…−17
LpFFG −36…−27 −66…−58 −21…−12
35…44
35…44
34…44
24…33
−30…−21 −30…−21
−44…−35 −30…−21
−60…−51 −30…−21
−87…−78 −30…−21
BC
LaFFG −42…−33 −37…−26 −20…−11
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−44…−35 −82…−73 −21…−12
32…41
33…42
31…40
28…37
−41…−32 −24…−15
−55…−46 −24…−15
−66…−57 −17…−8
−86…−77 −17…−8
LmFFG −42…−33 −50…−41 −20…−11
CC
LaFFG −35…−26 −42…−33 −22…−13
LpFFG −38…−29 −61…−52 −20…−11
LmFFG −34…−25 −55…−46 −22…−13
−29…−20 −91…−82 −16…−7
LpFFG −33…−24 −65…−56 −22…−13
38…47
37…46
37…46
33…42
−37…−28 −20…−11
−50…−41 −20…−11
−63…−55 −20…−11
−88…−79 −13…−4
AM
LaFFG −38…−29 −34…−25 −25…−16
LmFFG −40…−31 −45…−36 −25…−16
LpFFG −35…−26 −54…−46 −25…−16
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−51…−42 −73…−64 −25…−16
37…46
37…46
34…43
−33…−24 −25…−16
−44…−35 −25…−16
−54…−46 −25…−16
43…−52 −72…−64 −19…−10
IE
LaFFG −41…−32 −40…−31 −23…−14
LmFFG −38…−29 −51…−42 −23…−14
LpFFG −37…−28 −62…−53 −23…−14
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−45…−26 −77…−68 −23…−14
38…47
34…43
29…38
25…34
−38…−29 −23…−14
−49…−40 −23…−14
−64…−55 −26…−17
−89…80
−22…−13
KH
LaFFG −42…−33 −32…−25 −21…−12
LmFFG −41…−32 −45…−36 −23…−14
LpFFG −39…−30 −56…−47 −23…−14
LLOC
RaFFG
−47…−38 −73…−64 −23…14
34…43
−32…−23 −22…−13
LLOC
RaFFG
RmFFG
RpFFG
RLOC
−46…−37 −81…−72 −22…−13
34…43
32…41
27…37
36…45
−44…−35 −22…−13
−56…−47 −22…−13
−67…−58 −22…−13
−87…−78 −22…−13
This table presents individual participants’ ROI coordinates.
match across the objects, t(16) = 39.99, p < .001, d = 9.61.
There were no differences between feature matches and no
matches, although there was a trend, t(16) = 1.833, p =
.086, d = 0.503, with numerically higher “same” responses
for feature match items. However, the proportions of same
matches on both feature match and no match items were
extremely low. Thus, not surprisingly, category formation
reflected a preference for shape similarity in these overt
behavioral responses.
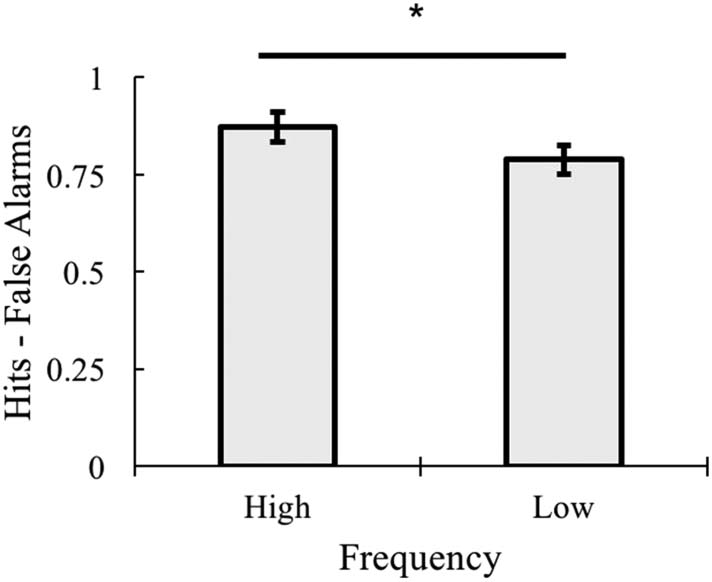
Recognition Performance
Sensitivity was calculated for each participant as hits −
false alarms separately for each condition (Figure 8).
Sensitivity was then submitted to a 2 (Frequency: high vs.
low) × 2 (Variability: variable vs. tight) repeated-measures
ANOVA. There was no main effect of Variability, F(1, 16) =
1.225, p = .285, η2 = .071. There was, however, a main
effect of Frequency, F(1, 16) = 13.88, p = .002, η2 =
.464, with high-frequency items having higher accuracy
than low-frequency items. This interaction was not signif-
icant, F(1, 16) = 0.585, p = .455, η2 = .035.
We also examined RTs on correct responses for learned
items by performing a 2 (Frequency: high vs. low) × 2
Plebanek and James
1405
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Proportion of “same” responses in the categorization task
across each trial type, *p < .001. Error bars represent ±1 standard error
of the mean.
(Variability: variable vs. tight) repeated-measures ANOVA
(Figure 9). There were no main effects ( ps > .25) or inter-
actions, F(1, 16) = 1.62, p = .221.
fMRI Data
Data Localization
The whole-brain contrast of learned object categories >
fixation revealed significant activation differences in two
large clusters spanning the left and right ventral temporal
and occipital cortices (see Figure 5 and Table 1) when
using a voxel-wise error rate p < .001. We corrected for
multiple comparisons by using the BrainVoyager cluster
threshold estimator plug in tool. We chose to use a
whole-brain false-positive discovery rate of p < .05, which
resulted in a cluster correction of six contiguous voxels,
and a voxel-wise error rate of p < .001. Thus, there
is evidence that ventral temporal regions such as the fusi-
form gyrus and the LOC are involved in learning these
Figure 9. Comparison of high, low, and new across variable and linked
items in the VTC ROIs (Figure 5, *p < .05). Error bars represent ±1
standard error of the mean.
novel symbols. This therefore served as our justification
for placing ROIs throughout these regions. See Figure 6
for a schematic of the ROI placement within these regions.
ROI Analyses
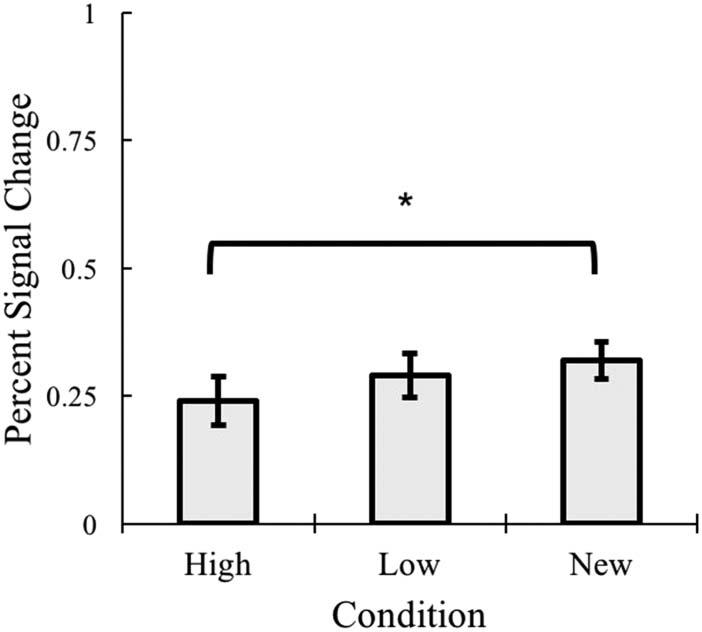
Familiar versus new. We first examined the overall
effect of familiarity to determine if the neural ROIs were
distinguishing between the implicitly learned and previ-
ously unseen objects. Data were therefore extracted from
the eight ROIs for three conditions: [high variable
linked], [low variable linked], and [new variable linked].
These conditions were selected to examine learning
differences while equating variability and co-occurrences.
Resultant data were then analyzed via a one-way ANOVA.
Planned comparisons were then performed to better un-
derstand the role of familiarity.
There was a significant effect of Condition (violated sphe-
ricity p = .022, Greenhouse Geisser: F(1.43, 22.90) = 4.47,
p = .034, η2 = .217; see Figure 10). Follow-up comparisons
revealed no difference between high- and low-frequency
items, although activity for low was numerically greater,
t(16) = 1.53 p = .145, d = 0.383. New items demonstrated
significantly greater activity than high items, t(16) = 2.33,
p = .033, d = 0.565. Similarly, new items demonstrated
greater activation than low items, but this only trended
toward significance, t(16) = 2.06, p = .056, d = 0.477.
These initial results served to suggest that the VTC was
sensitive to the difference between the implicitly learned
items and unseen items, but did not reveal differential
responding to high versus low frequency, implying that
there was no potential effects of adaptation.
Comparison of Structural Features
Figure 8. Sensitivity (hits − false alarms) by frequency collapsing
across variability in the recognition task, *p < .01. Error bars represent
±1 standard error of the mean.
Our main goal was to examine how the three structural
features that were learned during the training sessions
(frequency, variability, and co-occurrences) impacted the
1406
Journal of Cognitive Neuroscience
Volume 33, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 10. Main effect of Variability collapsed across both hemispheres,
all regions, and all other structural features, p < .01. Error bars represent
±1 standard error of the mean.
Figure 11. Main effect of Region collapsing across hemispheres and
structural features with follow-up comparisons, *p < .05. Error bars
represent ±1 standard error of the mean.
brain regions involved in processing object categories.
ROIs were the same as the previous analysis except that,
here, we also analyzed the data from primary visual cortex
separately from the overall model to test whether there
was differential responding based on category structure
in this region. In primary visual cortex, we first performed
a 2 (Frequency: high vs. low) × 2 (Variability: tight vs.
variable) × 2 (Co-occurrence: linked vs. unlinked) × 2
(Hemisphere: left vs. right) repeated-measures ANOVA.
In primary visual cortex, there was no main effects for
Frequency, F(1, 16) = 0.54, ns; Variability, F(1, 16) =
0.32, ns; Co-occurrence, F(1, 16) = 0.21, ns; or Hemisphere,
F(1, 16) = 0.43, ns; and no interaction among the vari-
ables (all Fs < 1.0). Thus, primary visual areas did not
show sensitivity to category structure in this design, but
responded with a similar amplitude to all the presented
objects.
The data extracted from the remaining ROIs were then
analyzed via a 2 (Frequency: high vs. low) × 2 (Variability:
tight vs. variable) × 2 (Co-occurrence: linked vs. un-
linked) × 4 (Region: anterior FFG vs. mid FFG vs. poste-
rior FFG vs. LOC) × 2 (Hemisphere: left vs. right)
repeated-measures ANOVA. Planned follow-up compari-
sons were performed for significant interactions and
main effects.
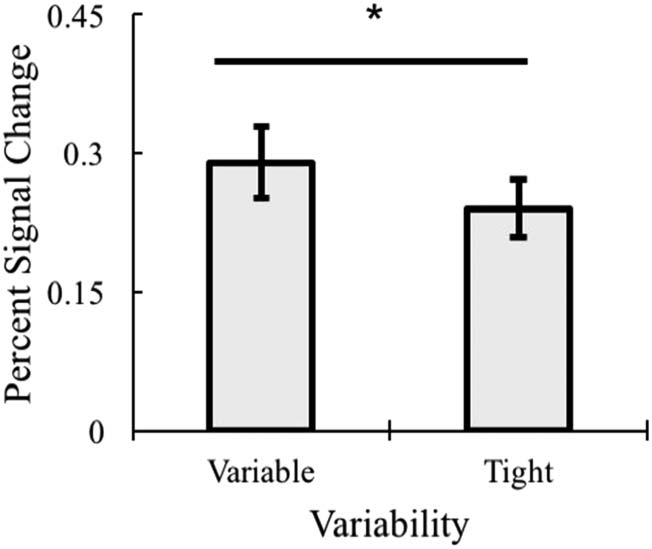
First, frequency trended toward significance, F(1, 16) =
3.61, p =.071, η2 = .184, with low-frequency object catego-
ries having numerically greater BOLD activation than high-
frequency object categories. There was a main effect of
Variability, F(1, 16) = 11.97, p = .003, η2 = .428, with greater
activation for variable object categories compared to tight
object categories (Figure 11). There was no main effect of
Co-occurrence ( p > .20).
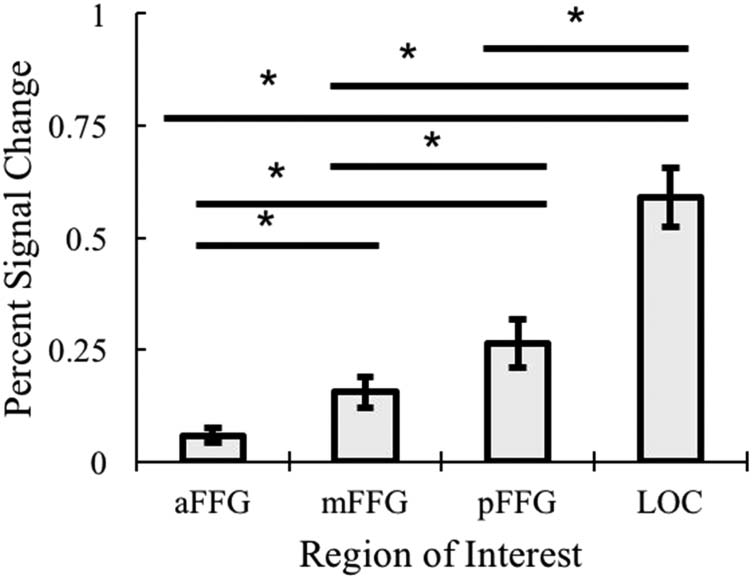
In terms of brain areas, there was a significant main effect
of Region (violated sphericity, p = .001, Greenhouse
Geisser: F(1.92, 30.71) = 40.29, p < .001, η2 = .716; see
Figure 12). We then performed planned comparisons to
better understand the effect of region in processing object
categories. In general, this main effect reflected the pattern
that activation while processing object categories increased
as the ROIs were placed more posteriorly. Specifically, the
LOC showed higher activation than all other regions,
ts(16) > 5.30, ps < .001, ds > 1.22. The posterior fusiform
gyrus was significantly greater than both the mid and
anterior fusiform gyri, ts(16) > 2.70, ps < .016, ds > 0.657.
Finally, the mid fusiform was greater than the anterior fusi-
form, t(16) = 4.66, p < .001, d = 1.18. There was also a
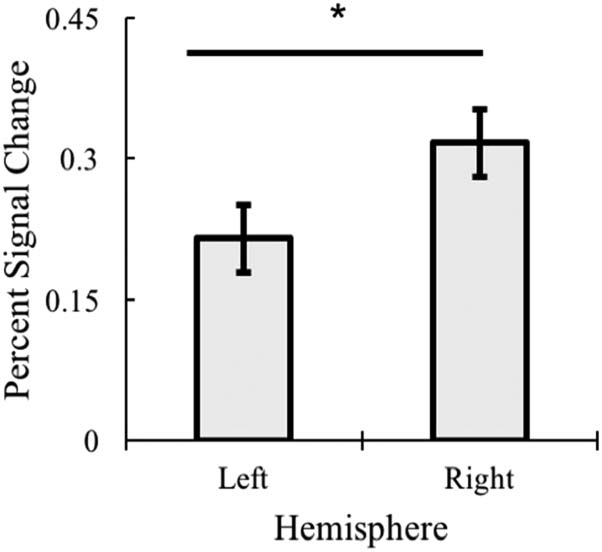
main effect of hemisphere, F(1, 16) = 11.54, p = .004, η2 =
.419, with greater activation across the right hemisphere
while processing object categories (see Figure 12).
We then examined interactions among our factors. No
five-way or four-way interactions were significant. Two in-
teractions were significant. First, there was a significant
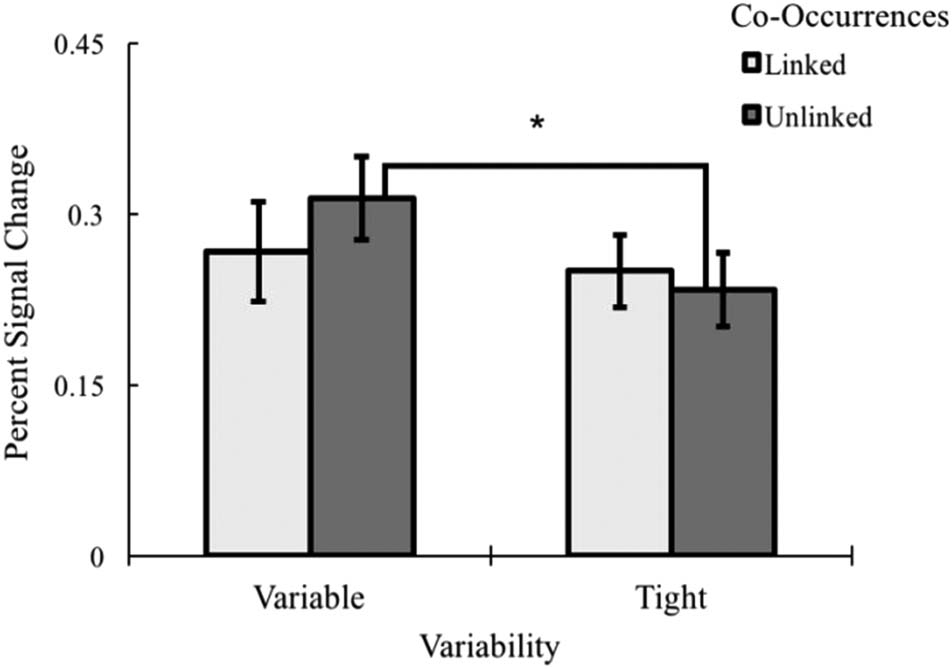
variability by co-occurrence interaction, F(1, 16) = 7.60,
p = .014, η2 = .322 (Figure 13). Specifically, there was
significantly greater activation for processing variable
compared to tight object categories when the object fea-
tures were also unlinked, t(16) = 4.23, p < .001, d =
1.10. No other differences were significant ( ps > .075).
In short, the interaction was driven by greater activity for
variability-unlinked compared to variable-linked items.
Figure 12. Main effect of Hemisphere collapsing across regions and
structural features, p < .01. Error bars represent ±1 standard error of
the mean.
Plebanek and James
1407
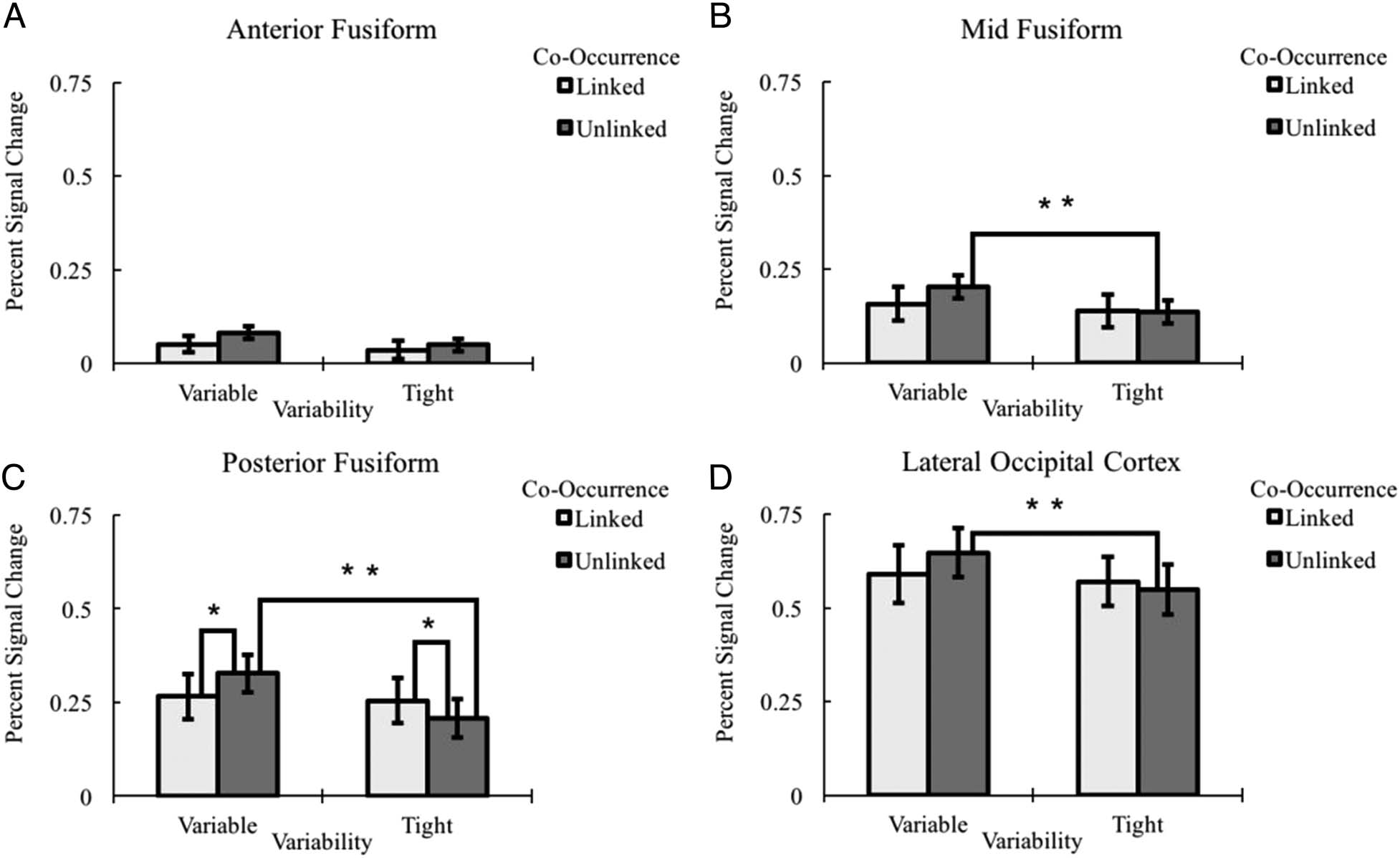
was no significant interaction in the anterior fusiform gyrus
( p = .403). However, there were significant Variability ×
Co-occurrence interactions within the middle fusiform,
F(1, 16) = 4.82, p = .043, η2 = .221; posterior fusiform,
F(1, 16) = 11.26, p = .004, η2 = .413; and the LOC, F(1,
16) = 4.98, p = .040, η2 = .237. Within the middle frontal
gyrus, the interaction was driven by greater activity for
variable-unlinked items compared to tight-unlinked items,
t(16) = 2.93, p = .010, d = 0.710). No other comparisons
were significant in this region, ( ps > .10). Within the pos-
terior fusiform gyrus, the pattern of the interaction was
more complex. First, variability resulted in greater activation
for unlinked items, t(16) = 4.46, p < .001, d = 1.09, but no
differences for linked items ( p > .150). Comparing across
co-occurrence levels revealed greater activation for un-
linked variable compared to linked variable items, t(16) =
2.37, p = .030, d = 0.466. However, tight categories showed
the reverse patterns with tight linked being greater than
tight unlinked, t(16) = 2.332, p = .033, d = 0.565. Within
the LOC, the pattern of the interaction was similar to the
middle fusiform gyrus. Specifically, the interaction was
driven by greater activity for variable-unlinked items com-
pared to tight-unlinked items, t(16) = 4.27, p = .001, d =
1.05. No other comparisons were significant in this region,
( ps > .10). In summary, variability is related to detecting
the relevant co-occurrences of object features and this
detection is primarily associated with the posterior fusi-
form gyrus.
Figure 13. A 2 (Variability: variability vs. tight) × 2 (Co-occurrence:
linked vs. unlinked) interaction collapsing across co-occurrence, region,
and hemisphere with follow-up comparisons, *p < .001. Error bars
represent ±1 standard error of the mean.
This suggests that variability may impact the detection of
relevant co-occurrences of features.
There was also a significant Variability × Co-Occurrence ×
Region interaction (violated sphericity, p = .021, F(2.25,
36) = 4.11, p = 021, η2 = .204). We performed further
analyses examining the Variability × Co-Occurrence inter-
action within individual regions (see Figure 14). There
Figure 14. A 2 (Variability: variable vs. tight) × 2 (Co-occurrence: linked vs. unlinked) × 4 (Region: anterior vs. middle vs. posterior vs. LOC)
interaction with follow-up comparisons, *p < .05, **p < .01. Error bars represent ±1 standard error of the mean.
1408
Journal of Cognitive Neuroscience
Volume 33, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
DISCUSSION
To better understand the role that structural aspects of
categories play in forming object categories, we examined
the responsiveness of the ventral temporal cortex to three
previously learned structural aspects: frequency of exem-
plars, variability among exemplars, and co-occurrences
among features among exemplars. To achieve this goal,
we created a set of metrically organized object categories
through which we could manipulate and quantify these
properties. Participants were then exposed to the object
categories during an implicit learning task. Following this
task, participants completed a recognition task as well as
neuroimaging sessions during which participants ob-
served learned an unlearned object categories composed
of different structures. Through this paradigm, we were
able to not only demonstrate that the fusiform gyrus and
the LOC are sensitive to variability (and to a lesser extent
frequency). We also demonstrated that some structural
elements interact to impact how the brain regions process
the categories. Our results can be summarized by three
critical contributions: (1) Variability among category mem-
bers influences the detection of co-occurrences between
object features. (2) This detection is also modulated by the
brain region, with the posterior fusiform gyrus being
especially sensitive to the variability–co-occurrence rela-
tionship. (3) Although shape frequency within a category
affects overt measures of recognition and has some effect
on BOLD signal, it does not interact in the same manner as
variability and co-occurrence.
Frequency
Frequency and object typicality have long been known
to play a role in the perception and learning of object
categories. As previously noted, frequency and typicality
have a mutual relationship and typicality judgments
increase as category members are presented at higher
frequencies (Nosofsky, 1988). Throughout development,
frequency appears to be an important factor in under-
standing the early acquisition of infant’s words to adult’s
categorization accuracy (Clerkin et al., 2017; Nosofsky,
1988). Past neuroimaging findings have implicated the
fusiform gyrus and the LOC in typicality measures of cate-
gory representation. For example, Davis and Poldrack
(2014) created a category stimulus space that allowed for
the manipulation of category exemplar features such as
typicality. Patterns of activation for typical members were
more similar to each other throughout the ventral–
temporal and occipital regions than they were to atypical
exemplars (Davis & Poldrack, 2014). In addition, Iordan
et al. (2016) found that representational similarity associ-
ated with the LOC decreased as typicality of exemplars
decreased. Our results corroborate these findings by sug-
gesting that “surprising” or atypical object categories may
activate object category learning brain systems more than
well-learned categories as evidenced by the greater
activation associated with low frequency and new catego-
ries compared to higher frequency categories. This inter-
pretation of our neural data is in line with our behavioral
data as well, where lower frequency items resulted in
poorer accuracy suggesting that they leave weaker
memory traces. Thus, the decreased responsiveness of
the high-frequency (and low frequency to a lesser extent)
categories compared to new categories may reflect more
established representations of the learned categories. In
summary, typicality and frequency may be proxies for
object familiarity and can impact the brain systems re-
sponsible for processing object categories and shape
their neural representations as well as overt recognition.
Variability and Co-occurrences
Variability has also been established as a factor in the mech-
anisms supporting category representation. Behaviorally,
variability has been known to support object recognition,
categorization, and generalization across the life span (Li
& James, 2016; Perry et al., 2010; Harman & Humphreys,
1999). Our findings demonstrate that the ventral temporal
cortex is involved in processing this variability and corrob-
orates previous findings in symbol and category formation
(Plebanek & James, 2021; Vinci-Booher et al., 2019; James,
2017; James & Engelhardt, 2012). We believe that this vari-
ability is most important in forming the initial representa-
tion of category.
However, our findings point to another role for variability:
identifying relevant co-occurrences within the category
structure. Research has already established the brain is
primed to extract structural regularities even though the
learner may not be explicitly aware of such regularities
(Turk-Browne et al., 2009). Our findings suggest that vari-
ability may make the representation of the feature co-
occurrences a stronger component of the object category
representation. Specifically, unlinked features resulted in
greater activation when the categories were also variable
whereas linked features were more equivalently processed
for variable and tight category structures.
Previous findings in category generalization have sup-
ported this role. Plebanek and James (2021) found that pro-
viding adults and 8-year-olds with variability during category
learning leads to generalization (based on increased brain
activation) via the feature that was invariant. In contrast,
highly similar exposure led to generalization based on
the overall appearance of the exemplar (Plebanek &
James, 2021). Taken together, these and our current
findings suggest that, from variability, category structure
emerges. Furthermore, this structure may be representa-
tive of co-occurrences: be they co-occurrences of time and
space, features and category membership, or features to
other features.
Categories and the Brain
The neural correlates of category learning have long been
debated. At the heart of this debate is the origin of
Plebanek and James
1409
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
category representations in the brain. Some researchers
propose functionally specialized centers such as the fusi-
form face area (Kanwisher, McDermott, & Chun, 1997)
whereas others propose process-driven expertise with
categories drive neural specialization (Gauthier,
Skudlarski, Gore, & Anderson, 2000). An additional can-
didate theory proposes that the brain represents informa-
tion about categories that may overlap throughout the
ventral temporal cortex (Haxby et al., 2001). These theo-
ries place different burdens on the role of the category.
Specialization theories suggest something inherent about
the category triggers domain-specific brain systems
(Kanwisher, 2017). Alternatively, these specialized re-
gions may encode other information regarding nonspe-
cialized categories (Haxby et al., 2001). Thus, there is a
conflict in the relationship between the brain regions
and categories in dictating category formation.
Our findings suggest a different pathway toward cate-
gory representation. The internal structure of categories
recruit different neural systems. Specifically, we identified
sensitivity to variability and feature co-occurrence that in-
creased in the posterior fusiform gyrus relative to other
regions. This finding parallels past research profiling the
fusiform gyrus’ responsiveness to letters and letter
strings. For example, James, James, Jobard, Wong, and
Gauthier (2005) found that the left anterior fusiform gy-
rus was selective for individual letters whereas the poste-
rior fusiform gyrus was selective for strings of letters.
Other works on the organization of the brain regions
involved in processing letters have supported a gradient-
style organization, although the exact distribution of sen-
sitivity is disagreed upon ( Vinckier et al., 2007). More
broadly, the occipito-temporal cortex may also show
graded sensitivity to the eccentricity of objects (Hasson,
Levy, Behrmann, Hendler, & Malach, 2002). Our results
suggest that a potential explanation for this heterogeneity
within brain regions is the subtle statistical differences,
most likely those stemming from variability, present in
object categories.
Thus, the recruitment of different regions throughout
the ventral temporal cortices may also reflect the extrac-
tion of the internal, statistical structures of categories that
guides the formation of representations. In our study,
structural elements such as variability may be closely tied
to perceptual features and thus guide the patterns of sim-
ilarity in the brain as well as the systems that process cat-
egories. Therefore, future evaluation of this theory may
be supplemented with the more distributed accounts of
category representations, which are guided, in part, by
perceptual elements that reflect the acquired stimulus
space (Kriegeskorte, Mur, & Bandettini, 2008).
Conclusions
Object categories have rich, internal structures that may
impact the brain systems recruited to learn novel catego-
ries. Here, we created metrically organized categories that
allowed us to operationalize variability, frequency, and
co-occurrence structures of novel categories. We have
demonstrated that the fusiform gyrus and the LOC are
sensitive to these structural elements. Moreover, as
regions move posteriorly through the fusiform gyrus,
sensitivity to structural properties (in particular the
variability-to-co-occurrence relationship) increases. In
short, we have demonstrated that the internal statistics
of object categories are critical in learning. Thus, future
research should not take for granted the role of structure
in exploring category learning dynamics.
Acknowledgments
This project was supported by National Institute of Health 2
T32 grant HD 007475-21 and by the Indiana University Office of
the Vice President for Research Emerging Area of Research
Initiative, Learning: Brains, Machines, and Children. We thank
Annie Abioye, Julia Lambert, and Lauren Wilkins for their assistance
on this project. No funding sources were involved in the study
design, analysis, or interpretation of the data, in the writing of
this paper, or in the decision to submit this paper for publication.
The final version of this work is published after the death of the
first author, Daniel Plebanek. Science has lost a bright young
light in his passing.
Reprint requests should be sent to Karin H. James, Department
of Psychological and Brain Sciences, Indiana University
Bloomington, 1101 East 10th St., Bloomington, IN 47405-
7000, or via e-mail: khjames@indiana.edu.
Author Contributions
Daniel J. Plebanek: Conceptualization; Data curation;
Formal analysis; Investigation; Methodology; Project ad-
ministration; Software; Validation; Visualization; Writing—
Original draft; Writing—Review & editing. Karin H. James:
Conceptualization; Formal analysis; Funding acquisition;
Methodology; Resources; Supervision; Visualization;
Writing—Review & editing.
Funding Information
Karin H. James, National Institute of Health 2 T32, grant
number: HD 007475-21. Karin H. James, Indiana
University Office of the Vice President for Research
Emerging Area of Research Initiative, grant number:
Learning: Brains, Machines, and Children.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed
a persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by esti-
mated gender identification of first author/last author) pub-
lishing in the Journal of Cognitive Neuroscience ( JoCN)
during this period were M(an)/M = .408, W(oman)/M =
.335, M/ W = .108, and W/ W = .149, the comparable pro-
portions for the articles that these authorship teams cited
1410
Journal of Cognitive Neuroscience
Volume 33, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
were M/M = .579, W/M = .243, M/ W = .102, and W/ W =
.076 (Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
REFERENCES
Ashby, F. G., & Valentin, V. V. (2017). Multiple systems of
perceptual category learning: Theory and cognitive tests. In H.
Cohen and C. Lefebvre (Eds.), Handbook of categorization in
cognitive science (2nd ed., pp. 157–188). Cambridge, MA:
Elsevier. https://doi.org/10.1016/B978-0-08-101107-2.00007-5
Bullmore, E. T., Brammer, M. J., Rabe-Hesketh, S., Curtis, V. A.,
Morris, R. G., Williams, S. C., et al. (1999). Methods for
diagnosis and treatment of stimulus-correlated motion in
generic brain activation studies using fMRI. Human Brain
Mapping, 7, 38–48. https://doi.org/10.1002/(SICI)1097-0193
(1999)7:1<38::AID-HBM4>3.0.CO;2-Q, PubMed: 9882089
Clerkin, E. M., Hart, E., Rehg, J. M., Yu, C., & Smith, L. B. (2017).
Real-world visual statistics and infants’ first-learned object
names. Philosophical Transactions of the Royal Society of
London, Series B: Biological Sciences, 372, 20160055. https://
doi.org/10.1098/rstb.2016.0055, PubMed: 27872373
Davis, T., & Poldrack, R. A. (2014). Quantifying the internal
structure of categories using a neural typicality measure.
Cerebral Cortex, 24, 1720–1737. https://doi.org/10.1093
/cercor/bht014, PubMed: 23442348
Gauthier, I., Skudlarski, P., Gore, J. C., & Anderson, A. W.
(2000). Expertise for cars and birds recruits brain areas
involved in face recognition. Nature Neuroscience, 3,
191–197. https://doi.org/10.1038/72140, PubMed: 10649576
Grill-Spector, K., Knouf, N., & Kanwisher, N. (2004). The
fusiform face area subserves face perception, not generic
within-category identification. Nature Neuroscience, 7,
555–562. https://doi.org/10.1038/nn1224, PubMed: 15077112
Grill-Spector, K., & Weiner, K. S. (2014). The functional
architecture of the ventral temporal cortex and its role in
categorization. Nature Reviews Neuroscience, 15, 536–548.
https://doi.org/10.1038/nrn3747, PubMed: 24962370
Harman, K. L., & Humphrey, G. K. (1999). Encoding ‘regular’
and ‘random’ sequences of views of novel three-dimensional
objects. Perception, 28, 601–615. https://doi.org/10.1068
/p2924, PubMed: 10664756
Hasson, U., Levy, I., Behrmann, M., Hendler, T., & Malach, R.
(2002). Eccentricity bias as an organizing principle for human
high-order object areas. Neuron, 34, 479–490. https://doi.org
/10.1016/S0896-6273(02)00662-1, PubMed: 11988177
Haxby, J. V., Gobbini, M. I., Furey, M. L., Ishai, A., Schouten, J. L., &
Pietrini, P. (2001). Distributed and overlapping representations
of faces and objects in ventral temporal cortex. Science, 293,
2425–2430. https://doi.org/10.1126/science.1063736, PubMed:
11577229
Hinds, O. P., Rajendran, N., Polimeni, J. R., Augustinack, J. C.,
Wiggins, G., Wald, L. L., et al. (2008). Accurate prediction
of V1 location from cortical folds in a surface coordinate
system. Neuroimage, 39, 1585–1599. https://doi.org/10.1016
/j.neuroimage.2007.10.033, PubMed: 18055222
Iordan, M. C., Greene, M. R., Beck, D. M., & Fei-Fei, L. (2016).
Typicality sharpens category representations in object-selective
cortex. Neuroimage, 134, 170–179. https://doi.org/10.1016/j
.neuroimage.2016.04.012, PubMed: 27079531
James, K. H. (2010). Sensori-motor experience leads to changes
in visual processing in the developing brain. Developmental
Science, 13, 279–288. https://doi.org/10.1111/j.1467-7687
.2009.00883.x, PubMed: 20136924
James, K. H. (2017). The importance of handwriting experience
on the development of the literate brain. Current Directions
in Psychological Science, 26, 502–508. https://doi.org/10
.1177/0963721417709821
James, K. H., & Atwood, T. P. (2009). The role of sensorimotor
learning in the perception of letter-like forms: Tracking
the causes of neural specialization for letters. Cognitive
Neuropsychology, 26, 91–110. https://doi.org/10.1080
/02643290802425914, PubMed: 18830859
James, K. H., & Engelhardt, L. (2012). The effects of
handwriting experience on functional brain development
in pre-literate children. Trends in Neuroscience and
Education, 1, 32–42. https://doi.org/10.1016/j.tine.2012.08
.001, PubMed: 25541600
James, K. H., James, T. W., Jobard, G., Wong, A. C. N., &
Gauthier, I. (2005). Letter processing in the visual system:
Different activation patterns for single letters and strings.
Cognitive, Affective, & Behavioral Neuroscience, 5,
452–466. https://doi.org/10.3758/CABN.5.4.452, PubMed:
16541814
James, K. H., Jones, S. S., Swain, S., Pereira, A., & Smith, L. B.
(2014). Some views are better than others: Evidence for
a visual bias in object views self-generated by toddlers.
Developmental Science, 17, 338–351. https://doi.org/10.1111
/desc.12124, PubMed: 24410976
James, K. H., & Swain, S. N. (2011). Only self-generated actions
create sensori-motor systems in the developing brain.
Developmental Science, 14, 673–678. https://doi.org/10.1111
/j.1467-7687.2010.01011.x, PubMed: 21676088
Kanwisher, N. (2017). The quest for the FFA and where it led.
Journal of Neuroscience, 37, 1056–1061. https://doi.org/10
.1523/JNEUROSCI.1706-16.2016, PubMed: 28148806
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). The
fusiform face area: A module in human extrastriate cortex
specialized for face perception. Journal of Neuroscience, 17,
4302–4311. https://doi.org/10.1523/JNEUROSCI.17-11-04302
.1997, PubMed: 9151747
Kloos, H., & Sloutsky, V. M. (2008). What’s behind different
kinds of kinds: Effects of statistical density on learning and
representation of categories. Journal of Experimental
Psychology: General, 137, 52–72. https://doi.org/10.1037
/0096-3445.137.1.52, PubMed: 18248129
Kraebel, K. S., & Gerhardstein, P. C. (2006). Three-month-old
infants’ object recognition across changes in viewpoint using an
operant learning procedure. Infant Behavior & Development,
29, 11–23. https://doi.org/10.1016/j.infbeh.2005.10.002,
PubMed: 17138257
Kriegeskorte, N., Mur, M., & Bandettini, P. (2008). Representational
similarity analysis—Connecting the branches of systems
neuroscience. Frontiers in Systems Neuroscience, 2, 4. https://
doi.org/10.3389/neuro.06.004.2008, PubMed: 19104670
Li, J. X., & James, K. H. (2016). Handwriting generates variable
visual output to facilitate symbol learning. Journal of
Experimental Psychology: General, 145, 298–313. https://doi
.org/10.1037/xge0000134, PubMed: 26726913
Nosofsky, R. M. (1988). Similarity, frequency, and category
representations. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 14, 54–65. https://doi.org
/10.1037/0278-7393.14.1.54
Perry, L. K., Samuelson, L. K., Malloy, L. M., & Schiffer, R. N.
(2010). Learn locally, think globally: Exemplar variability
supports higher-order generalization and word learning.
Psychological Science, 21, 1894–1902. https://doi.org/10.1177
/0956797610389189, PubMed: 21106892
Plebanek, D. J., & James, K. H. (2021). Category structure
guides the formation of neural representations. Experimental
Brain Research, 239, 1667–1684. https://doi.org/10.1007
/s00221-021-06088-7, PubMed: 33782786
Plebanek and James
1411
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Rosch, E., & Mervis, C. B. (1975). Family resemblances: Studies
in the internal structure of categories. Cognitive Psychology,
7, 573–605. https://doi.org/10.1016/0010-0285(75)90024-9
Rosch, E., Mervis, C. B., Gray, W. D., Johnson, D. M., &
Boyes-Braem, P. (1976). Basic objects in natural categories.
Cognitive Psychology, 8, 382–439. https://doi.org/10.1016
/0010-0285(76)90013-X
Saffran, J. R., Aslin, R. N., & Newport, E. L. (1996). Statistical
learning by 8-month-old infants. Science, 274, 1926–1928. https://
doi.org/10.1126/science.274.5294.1926, PubMed: 8943209
Schapiro, A. C., Kustner, L. V., & Turk-Browne, N. B. (2012).
Shaping of object representations in the human medial
temporal lobe based on temporal regularities. Current
Biology, 22, 1622–1627. https://doi.org/10.1016/j.cub.2012.06
.056, PubMed: 22885059
Sherman, B. E., Graves, K. N., & Turk-Browne, N. B. (2020). The
prevalence and importance of statistical learning in human
cognition and behavior. Current Opinion in Behavioral
Sciences, 32, 15–20. https://doi.org/10.1016/j.cobeha.2020.01
.015, PubMed: 32258249
Slone, L. K., Smith, L. B., & Yu, C. (2019). Self-generated
variability in object images predicts vocabulary growth.
Developmental Science, 22, e12816. https://doi.org/10.1111
/desc.12816, PubMed: 30770597
Sloutsky, V. M. (2010). From perceptual categories to concepts:
What develops? Cognitive Science, 34, 1244–1286. https://doi
.org/10.1111/j.1551-6709.2010.01129.x, PubMed: 21116483
Smith, L. B., Jayaraman, S., Clerkin, E., & Yu, C. (2018). The
developing infant creates a curriculum for statistical learning.
Trends in Cognitive Sciences, 22, 325–336. https://doi.org/10
.1016/j.tics.2018.02.004, PubMed: 29519675
Smith, L., & Yu, C. (2008). Infants rapidly learn word-referent
mappings via cross-situational statistics. Cognition, 106,
1558–1568. https://doi.org/10.1016/j.cognition.2007.06.010,
PubMed: 17692305
Stansbury, D. E., Naselaris, T., & Gallant, J. L. (2013). Natural
scene statistics account for the representation of scene
categories in human visual cortex. Neuron, 79, 1025–1034.
https://doi.org/10.1016/j.neuron.2013.06.034, PubMed:
23932491
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic
atlas of the human brain. 3-Dimensional proportional
system: An approach to cerebral imaging. Stuttgart: Thieme.
Turk-Browne, N. B., Scholl, B. J., Chun, M. M., & Johnson, M. K.
(2009). Neural evidence of statistical learning: Efficient
detection of visual regularities without awareness. Journal of
Cognitive Neuroscience, 21, 1934–1945. https://doi.org/10
.1162/jocn.2009.21131, PubMed: 18823241
Vinberg, J., & Grill-Spector, K. (2008). Representation of shapes,
edges, and surfaces across multiple cues in the human visual
cortex. Journal of Neurophysiology, 99, 1380–1393. https://
doi.org/10.1152/jn.01223.2007, PubMed: 18171705
Vinckier, F., Dehaene, S., Jobert, A., Dubus, J. P., Sigman, M., &
Cohen, L. (2007). Hierarchical coding of letter strings in the
ventral stream: Dissecting the inner organization of the visual
word-form system. Neuron, 55, 143–156. https://doi.org/10
.1016/j.neuron.2007.05.031, PubMed: 17610823
Vinci-Booher, S., Cheng, H., & James, K. H. (2019). An analysis
of the brain systems involved with producing letters by hand.
Journal of Cognitive Neuroscience, 31, 138–154. https://doi
.org/10.1162/jocn_a_01340, PubMed: 30240307
Vinci-Booher, S., & James, K. H. (2020). Visual experiences
during letter production contribute to the development of
the neural systems supporting letter perception. Developmental
Science, 23, e12965. https://doi.org/10.1111/desc.12965,
PubMed: 32176426
Wallis, G., & Bülthoff, H. (1999). Learning to recognize objects.
Trends in Cognitive Sciences, 3, 22–31. https://doi.org/10
.1016/S1364-6613(98)01261-3
Wood, J. N. (2016). A smoothness constraint on the development
of object recognition. Cognition, 153, 140–145. https://doi.org
/10.1016/j.cognition.2016.04.013, PubMed: 27208825
Yu, C., & Smith, L. B. (2007). Rapid word learning under
uncertainty via cross-situational statistics. Psychological
Science, 18, 414–420. https://doi.org/10.1111/j.1467-9280
.2007.01915.x, PubMed: 17576281
1412
Journal of Cognitive Neuroscience
Volume 33, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
8
1
3
9
7
1
9
2
7
6
6
8
/
/
j
o
c
n
_
a
_
0
1
7
3
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3