Spontaneous Preference for Slowly Moving
Objects in Visually Naïve Animals
Justin N. Wood
1
1
Department of Psychology, University of Southern California
Keywords: visual preference, newborn, object recognition, controlled rearing, chick
ABSTRACT
To perceive the world successfully, newborns need certain types of visual experiences.
The development of object recognition, for example, requires visual experience with slowly
moving objects. To date, however, it is unknown whether newborns actively seek out the
best visual experiences for developing object recognition. To address this question, I used an
automated controlled-rearing method to examine whether visually naïve animals (newborn
chicks) seek out slowly moving objects. Despite receiving equal exposure to slowly and to
quickly rotating objects, the majority of the chicks developed a preference for slowly rotating
objects. This preference was robust, producing large effect sizes across objects, experiments,
and successive test days. These results indicate that newborn brains rapidly develop
mechanisms for orienting young animals toward optimal visual experiences, thus facilitating
the development of object recognition. This study also demonstrates that automation can
be a valuable tool for studying the origins and development of visual preferences.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
An important goal in the cognitive sciences is to understand how visual preferences shape
the development of the mind. Since visual preferences influence where a newborn will look
(and thus what information will be extracted from the world), a complete understanding of
perceptual and cognitive development requires understanding the nature of early emerging
visual preferences. Due to methodological barriers, however, assessments of early visual pref-
erences have so far been based on methods that sample newborns’ behavior for short periods
of time (e.g., 5–10 min) within a single testing session. As a result, it has not been possible
to obtain precise measurements of visual preferences from newborn subjects or examine how
visual preferences change over time. To overcome these barriers, I developed an automated
controlled-rearing method that can measure newborn subjects’ visual preferences continu-
ously (24/7) from the onset of vision. This method makes it possible to measure how visual
preferences emerge and change across development, with an unprecedented degree of preci-
sion. In the present study, I tested whether newborn animals have an early emerging preference
for visual input that is more conducive for building accurate object representations.
Specifically, a growing body of research indicates that visual experience with slowly
changing features is essential for the development of vision (e.g., DiCarlo, Zoccolan, & Rust,
2012; Földiák, 1991; Rolls, 2012; Stone, 1996; Wiskott & Sejnowski, 2002). For instance,
computational models that extract slowly changing features from the visual environment can
account for the self-organization of complex-cell receptive fields (Berkes & Wiskott, 2005), the
recognition of whole objects invariant to spatial transformations (Franzius, Wilbert, & Wiskott,
2011), and the self-organization of place cells, grid cells, and head-direction cells (Franzius,
Sprekeler, & Wiskott, 2007; Wyss, König, & Verschure, 2006). Moreover, controlled-rearing
a n o p e n a c c e s s
j o u r n a l
Citation: Wood, J. N. (2017).
Spontaneous Preference for Slowly
Moving Objects in Visually Naïve
Animals. Open Mind: Discoveries in
Cognitive Science, 1(2), 111–122.
https://doi.org/10.1162/opmi_a_00012
DOI:
https://doi.org/10.1162/opmi_a_00012
Supplemental Materials:
https://doi.org/10.1162/opmi_a_00012
Received: 19 January 2017
Accepted: 08 July 2017
Competing Interests: The author
declares no conflict of interest.
Corresponding Author:
Justin N. Wood
justin.wood@usc.edu
Copyright: © 2017
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
Spontaneous Preference for Slow Objects Wood
experiments provide direct causal evidence that the development of object recognition re-
quires visual experience with slowly moving objects (Wood & Wood, 2016a). When newborn
chicks are raised with slowly moving objects, the chicks create invariant object representations
that generalize across novel viewpoints and rotation speeds. Conversely, when newborn chicks
are raised with quickly moving objects, the chicks create inaccurate object representations that
fail to generalize across novel viewing situations. Thus, visual experience with slowly moving
objects is critical for the development of object recognition.
If newborns build more accurate object representations when exposed to slowly mov-
ing objects, then natural selection might have designed newborn brains to seek out slowly
moving objects. This “selective sampling” mechanism would be useful, allowing newborns to
avoid wasting precious computational resources on visual material that cannot be transformed
into accurate object representations. To test for the existence of such a mechanism, I exam-
ined whether newborn chicks, lacking any previous visual experience, show a spontaneous
preference to approach slowly moving objects over quickly moving objects.
Newborn chicks are an ideal model system for studying the origins of visual preferences,
for several reasons. First, chicks can be raised in strictly controlled environments (e.g., en-
vironments containing no real-world objects) from the onset of vision. Thus, with chicks it is
possible to study how visual preferences emerge within strictly controlled visual environments.
Second, there is evidence that newborn chicks develop visual preferences rapidly (reviewed
by Versace & Vallortigara, 2015), and some of these preferences have been linked to specific
neural structures (e.g., Di Giorgio et al., 2016; Mayer, Rosa-Salva, Lorenzi, & Vallortigara,
2016). Third, newborn chicks develop high-level object recognition abilities. For instance,
newborn chicks can build a view-invariant representation of the first object they see in their
life (Wood, 2013, 2015; Wood & Wood, 2015), and can recognize objects rapidly, within a
fraction of a second (Wood & Wood, 2016b). Fourth, birds and mammals use homologous
neural circuits—with common structural and functional characteristics—to process sensory
input (reviewed by Jarvis et al., 2005; Karten, 2013). Accordingly, studies of chicks may
reveal general insights into the nature of vertebrate visual development.
EXPERIMENT 1
Methods
Twelve Rhode Island Red chicks of unknown sex were tested. The sample size was
Subjects
determined before the experiment was conducted, based on previous automated controlled-
rearing experiments with newborn chicks (Wood, 2013, 2014). No subjects were excluded
from the analyses. The eggs were obtained from a local distributor and incubated in darkness in
an OVA-Easy incubator (Brinsea Products Inc., Titusville, FL). The incubation room was kept in
complete darkness to avoid exposing the chicks to light through their shells. After hatching, the
chicks were moved from the incubation room to the controlled-rearing chambers in darkness
with the aid of night vision goggles. Each chick was raised singly within its own chamber. This
research was approved by the University of Southern California Institutional Animal Care and
Use Committee.
Procedure Newborn chicks were raised for five days within specially designed controlled-
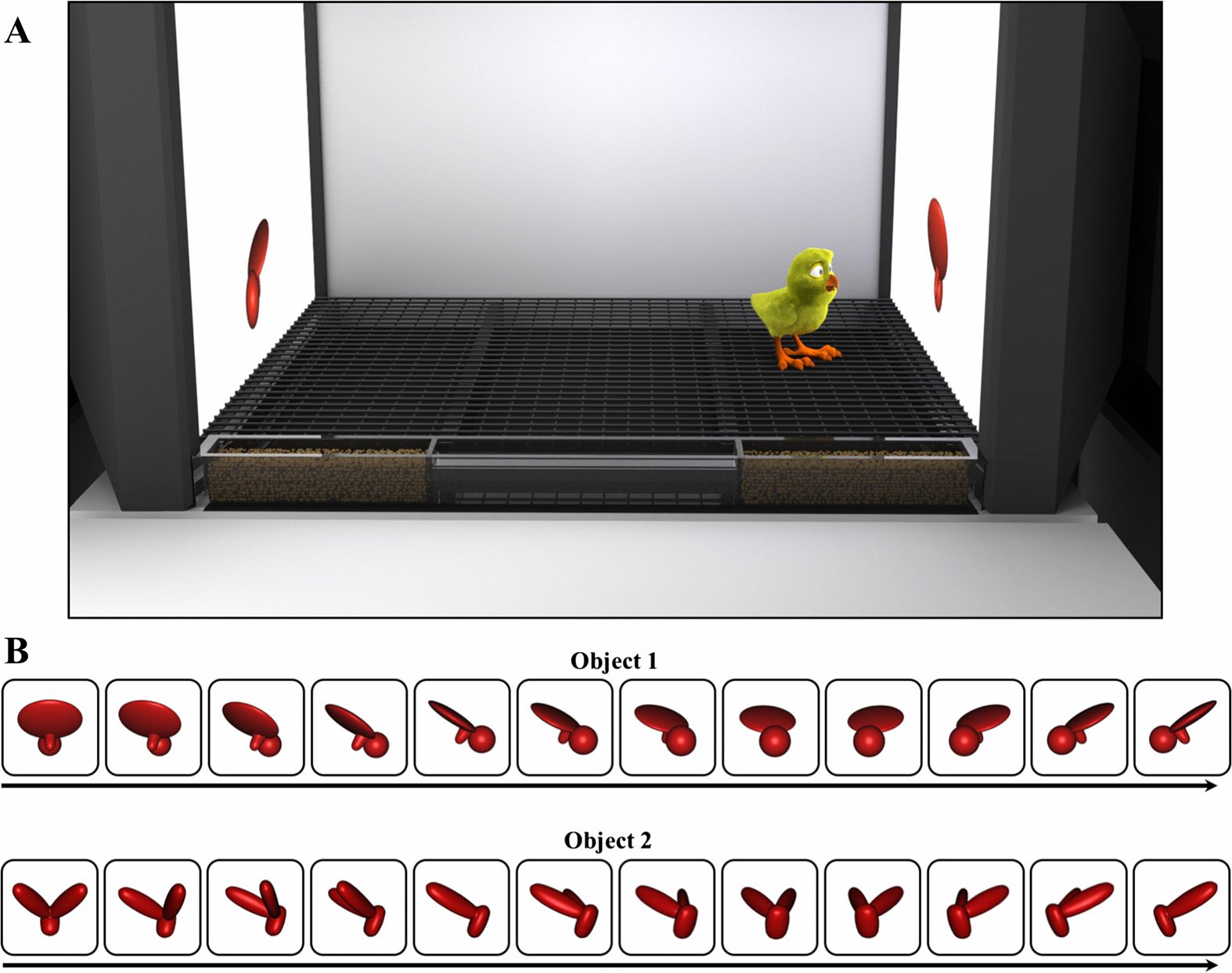
rearing chambers (66 cm length × 42 cm width × 69 cm height; Figure 1A). To present object
stimuli to the chicks, virtual objects were projected on two display walls (19” LCD monitors
with 1,440 × 900 pixel resolution) situated on opposite sides of the chamber. Food (grain) and
water were provided within transparent troughs in the ground (66 cm length × 2.5 cm width
OPEN MIND: Discoveries in Cognitive Science
112
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Spontaneous Preference for Slow Objects Wood
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. The experimental procedure.
(A) Newborn chicks were raised in controlled-rearing
chambers that contained a virtual object on each side of the chamber. The objects were identical, but
rotated at different speeds. One object rotated once every 15 s (slowly moving object), whereas the
other object rotated once per second (quickly moving object). If newborn chicks have a preference
for slowly moving objects, then the chicks should spend more time in proximity to the slowly moving
object than the quickly moving object. (B) The virtual objects. Half of the chicks were raised with
Object 1 and half of the chicks were raised with Object 2.
× 2.7 cm height). The floor of the chamber consisted of black wire mesh supported over a
black surface. All animal husbandry was performed in darkness with night vision goggles to
avoid exposing the chicks to extraneous visual input.
During each preference trial, the same object was projected on both display walls simul-
taneously (Figure 1A). On one display wall, the object moved slowly (one rotation every 15 s),
whereas on the other display wall, the object moved quickly (one rotation per second). These
two rotation speeds were used because newborn chicks build accurate object representations
when reared with objects that rotate slowly (one rotation every 15 s) and inaccurate object
representations when reared with objects that rotate quickly (one rotation per second) (Wood
& Wood, 2016a). By using the same objects and rotation speeds in the present study, it was
possible to examine directly whether newborn chicks have a preference for visual stimuli that
produces more accurate object representations.
I used rotary motion rather than lateral (picture plane) motion for two additional reasons.
First, with rotary motion, it is possible to manipulate an object’s motion speed without changing
its position in the environment. Second, it is more difficult to make compensatory head and
eye movements to adjust for rotary motion compared to lateral motion, allowing for a more
direct test of whether newborn chicks have a visual preference for slowly moving objects.
OPEN MIND: Discoveries in Cognitive Science
113
Spontaneous Preference for Slow Objects Wood
During the experiment, the chicks received equal exposure to the slowly and the quickly
moving objects. Thus, the objects were equally familiar to the chicks. If newborn chicks have
a spontaneous preference for slowly moving objects, then the chicks should spend more time
in proximity to the slowly moving object than the quickly moving object.
To test whether the chicks’ visual preferences would generalize across objects, the chicks
were raised with one of two objects differing in shape. Half of the chicks were raised with the
object shown in the top panel of Figure 1B (see Video S1 in the Supplemental Materials [Wood,
2017]) and half were raised with the object shown in the bottom panel of Figure 1B (see
Video S2 in the Supplemental Materials [Wood, 2017]). On average, the objects measured
8 cm (width) × 7 cm (height) and were suspended 3 cm off the ground. The object rotated in the
depth plane around a frontoparallel vertical axis (24 frames/s) and was presented at the middle
of the display wall on a uniform white background. The two objects were modeled after those
used in previous studies that tested for invariant object recognition in adult rats (Zoccolan,
Oertelt, DiCarlo, & Cox, 2009) and newborn chicks (Wood, 2013). Videos S1 and S2 in the
Supplemental Materials (Wood, 2017) show the animations used in Experiment 1.
Each chick received 120 preference trials during the experiment (one trial per hour
over 5 days). Each trial lasted 59 min, followed by a 1-min period of darkness. The slowly
and the quickly moving objects switched display walls after each preference trial. The chicks’
behavior was tracked continuously using automated image-based tracking software (EthoVi-
sion XT, Noldus Information Technology, Leesburg, VA) that calculated the amount of time
the chicks spent within zones (22 cm × 42 cm) next to each object. As a result, I was able to
examine (a) whether newborn chicks prefer slowly moving objects, (b) whether this preference
emerges at different times in different subjects, and (c) whether this preference changes over
time within subjects. In total, 1,440 hr of video footage (5 days × 24 hr/day × 12 subjects)
were collected for Experiment 1 (120 hr of preference data were collected from each chick).
By collecting large amounts of data from each chick, it was possible to achieve low measure-
ment error. For instance, the test-retest reliability of the preference scores across the five days
of the experiment was high (Cronbach’s alpha = .89).
Results and Discussion
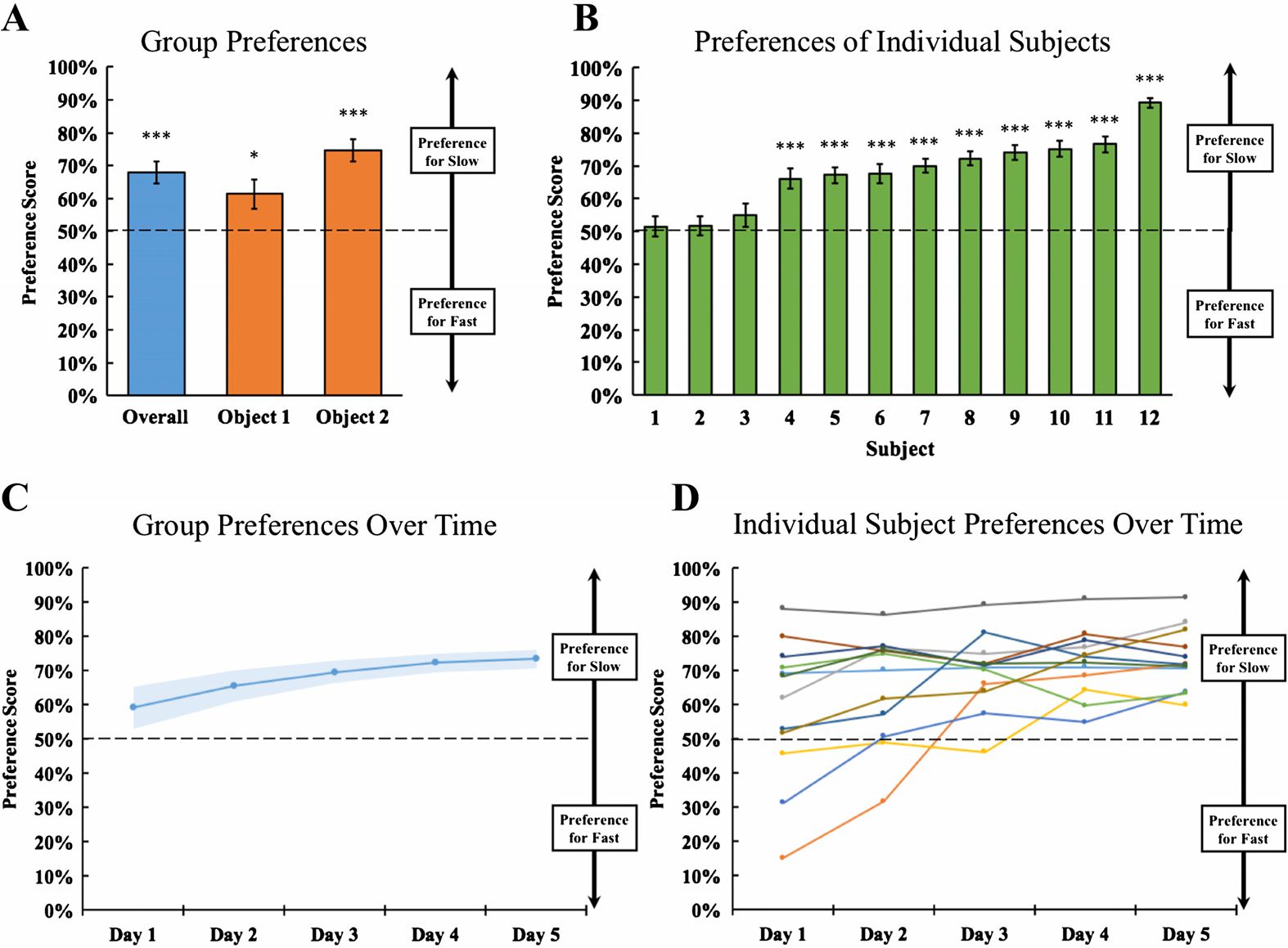
The results are shown in Figure 2. For each trial, I computed the preference score for each
chick by dividing the time spent with the slowly moving object by the time spent with both
objects. Preference scores above 50% indicate a preference for the slowly moving object and
preference scores below 50% indicate a preference for the quickly moving object. The results
were analyzed with one-sample t tests compared against chance level (50%). Analyses are
included for both the overall group and each individual subject.
For overall group performance, the average preference score was 67.9%
Group Performance
(SEM = 3%). The chicks spent significantly more time by the slowly moving object than by
the quickly moving object (two-tailed one-sample t test: t(11) = 5.43, p < .001, Cohen’s d =
1.57). The preference for slowly moving objects occurred both when the chicks were reared
with Object 1 [t(5) = 2.58, p = .05, Cohen’s d = 1.05] and Object 2 [t(5) = 7.30, p <
.001, Cohen’s d = 2.98]. As shown in Figure 2C, the chicks spent significantly more time by
the slowly moving object during Days 2–5 of the experiment [Day 1: t(11) = 1.50, p = .16,
Cohen’s d = .43; Day 2: t(11) = 3.42, p = .006, Cohen’s d = .99; Day 3: t(11) = 6.23,
p < .0001, Cohen’s d = 1.80; Day 4: t(11) = 7.94, p < .00001, Cohen’s d = 2.29; Day 5:
OPEN MIND: Discoveries in Cognitive Science
114
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Spontaneous Preference for Slow Objects Wood
Figure 2. Experiment 1 results. (A) Group performance. The blue bar shows the preference score
for all of the chicks and the orange bars show the preference scores for the chicks reared with each
of the two objects. The chicks had a strong preference for the slowly moving object. (B) Individual
subject performance. The bars show the preference scores for each chick (ordered by the strength
of the preference). The majority of the chicks had a statistically significant preference for the slowly
moving object.
(C) Change in preference scores across the experiment. Despite receiving equal
exposure to the slowly and quickly moving objects, the chicks developed a preference for the slowly
moving object. (D) Change in preference scores for each subject. Each line shows the preference
scores for one chick across the five days of the experiment. By the end of the experiment, all of
the chicks had developed a preference for the slowly moving object. Asterisks denote statistical
significance: ∗p < .05; ∗∗p < .01; ∗∗∗p < .001 (two-tailed one-sample t tests). Error bars indicate
standard error.
t(11) = 8.85, p < .00001, Cohen’s d = 2.55].1 Notably, these effect sizes are very large (for
reference, Cohen considered d ≥ .8 to be a large effect size; Cohen, 1988).
I also estimated the posterior probability favoring the null hypothesis using the Jeffreys–
Zellner–Siow (JZS) Bayes factor (BF10). The Bayes factor is the odds ratio comparing the
likelihood of the data fitting under the alternative hypothesis to the likelihood of the data
fitting under the null hypothesis. The BF10 for the overall preference score was 153.8 (i.e., the
alternative hypothesis was 153.8 times more likely than the null hypothesis). Thus, these data
provide strong evidence that newborn chicks prefer slowly moving objects.
1 While the group tended to prefer the slowly moving object across the first day (see Figure 3A), the group
analysis was not statistically significant for Day 1. The group analysis for Day 1 was also not statistically signif-
icant after removing the trials where the chicks were nonresponsive due to rest/sleep [t(11) = 1.14, p = .28,
Cohen’s d = .33]. Specifically, I removed trials from the analysis whenever two or more successive trials yielded
0%–100% or 100%–0% alternations in preference scores, a pattern that indicates inactivity (see Figure 3B for
details).
OPEN MIND: Discoveries in Cognitive Science
115
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Spontaneous Preference for Slow Objects Wood
Since 120 preference trials were collected from each chick, it
Individual Subject Performance
was possible to measure each chick’s visual preferences with high precision. The preference
scores for each chick are shown in Figure 2B. The majority of the chicks (9 out of 12) had a
statistically significant preference for slowly moving objects across the experiment (two-tailed
t tests: all p < .001).
Why did three of the chicks fail to show this preference across the experiment? One
possibility is that all chicks eventually develop a preference for slowly moving objects, but
that this preference emerges at different times in different subjects. To test this possibility, I
charted each chick’s preference scores across the five days of the experiment. As illustrated
in Figure 2D, there were large differences in the preference scores across chicks. Most of the
chicks (8 of the 12) preferred the slowly moving object on Day 1 (two-tailed t tests: 2 subjects,
p < .05, 2 subjects, p < .01, 4 subjects, p < .001), but a few of the chicks developed
this preference 2–3 days after hatching. Interestingly, two of the chicks started with a strong
preference for the quickly moving object, but then lost this preference and developed a new
preference for the slowly moving object.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
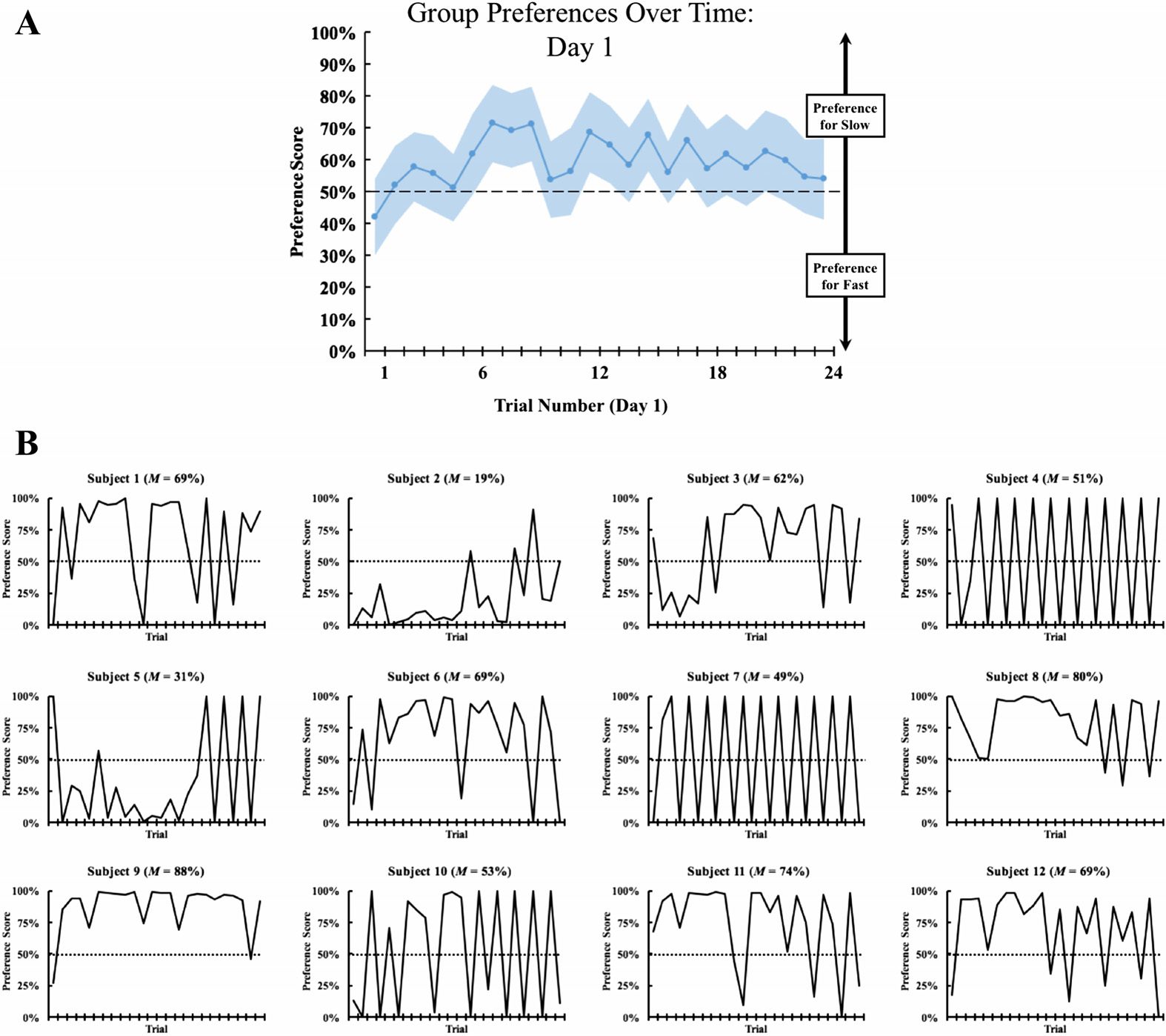
Figure 3.
(A) Change in group preference scores across Day 1 (Experiment 1). (B) Change in
preference scores across Day 1 for each subject (Experiment 1). The graphs show the preference
scores across the first 24 trials of the experiment (the chicks received one trial per hour). Large
fluctuations in performance across trials (from 0% to 100%) indicate that the chick was stationary/
sleeping (i.e., since the objects switched display walls across successive trials, stationary chicks
produced large fluctuations in preference scores across trials).
OPEN MIND: Discoveries in Cognitive Science
116
Spontaneous Preference for Slow Objects Wood
To provide a more fine-grained view of how this visual preference emerged during Day 1,
I charted the visual preference scores across the first 24 preference trials of the experiment
(Figure 3). Figure 3A shows the group preference scores, while Figure 3B shows each chick’s
preference scores. While some of the chicks developed a preference for the slowly moving
object during their first few hours of exposure to light, other chicks preferred the quickly moving
object or showed no clear preference for either object during Day 1. Together, these analyses
show that there can be substantial individual differences in the development of this visual
preference, even among subjects raised in identical visual environments. Newborn chicks can
differ both in the strength of their visual preference and in the timing of when this preference
emerges during development.
EXPERIMENT 2
The results from Experiment 1 show that newborn chicks have an early emerging preference
for slowly moving objects. However, since the chicks were presented with only two rotation
speeds, the nature of this preference remains unclear. In particular, the results are consistent
with at least three possibilities. First, newborn chicks might always prefer the slowest moving
object in their environment. Second, newborn chicks might prefer the slowest moving object
when the speeds of the objects differ by some critical ratio (e.g., when one object moves twice
as fast as the other object). Third, newborn chicks might prefer the slowest moving object
when one of the objects exceeds a particular speed threshold.
To distinguish between these possibilities, I conducted a second experiment in which the
objects rotated at five possible speeds (one rotation every 1.25 s, 2.5 s, 5 s, 10 s, or 20 s). Across
the trials, the chicks were presented with all 10 pairwise combinations of these five rotation
speeds.
If chicks always prefer the slowest moving object in their environment, then they
should have a preference for the slowly moving object across all 10 pairwise combinations.
Conversely, if chicks prefer slowly moving objects only when the object speeds differ by some
critical ratio, then the chicks’ preference for the slowly moving object should depend on the
ratio between the rotation speeds. Finally, if chicks prefer the slowly moving object only when
the quickly moving object exceeds a particular speed threshold, then the chicks should prefer
the slowly moving object only when the quickly moving object exceeds a particular rotation
speed.
Methods
The methods were identical to those used in Experiment 1, except in the following ways.
First, 11 new subjects were tested. One subject was removed from the analyses because
the camera malfunctioned during the experiment. Second, the experiment lasted for 7 days
rather than 5 days. This allowed us to collect a larger amount of data from each chick. Third,
on each preference trial, the objects could rotate at five possible speeds (one rotation every
1.25 s, 2.5 s, 5 s, 10 s, or 20 s). These successive values differ by a factor of two; thus, with
each successive increase in speed, the object rotated twice as fast. The chicks were presented
with all 10 pairwise combinations of the five rotation speeds: 1.25 s vs. 2.5 s, 1.25 s vs. 5 s,
1.25 s vs. 10 s, 1.25 s vs. 20 s, 2.5 s vs. 5 s, 2.5 s vs. 10 s, 2.5 s vs. 20 s, 5 s vs. 10 s, 5 s vs.
20 s, and 10 s vs. 20 s.
Each chick received 140 preference trials during the experiment (14 trials for each pair-
wise combination). Each preference trial lasted 68 min, followed by a 1-min period of
OPEN MIND: Discoveries in Cognitive Science
117
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Spontaneous Preference for Slow Objects Wood
In total, ~1,848 hr of video footage (7 days × 24 hr/day × 11 subjects) were
darkness.
collected for Experiment 2 (i.e., 168 hr of preference data were collected from each chick).
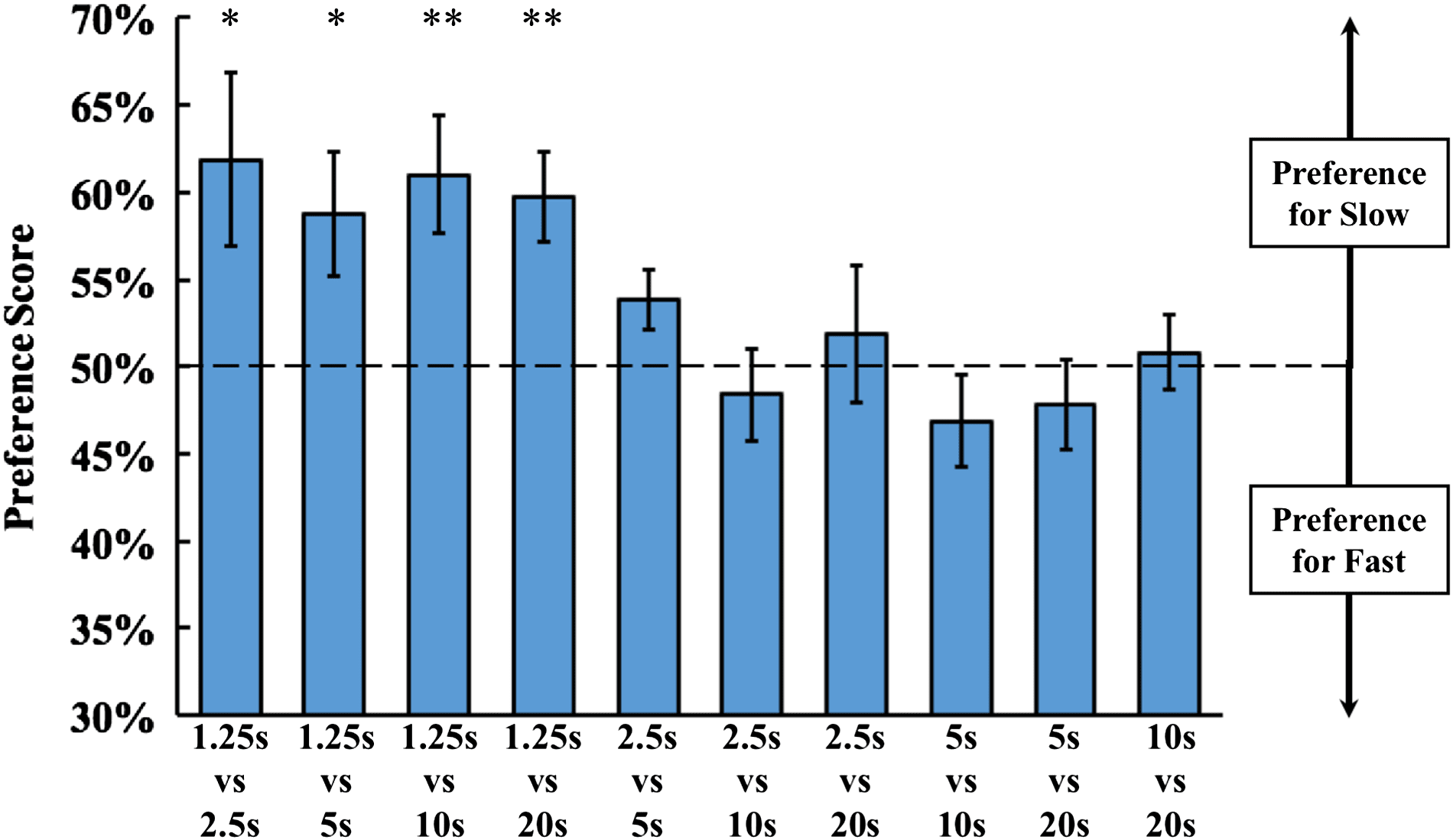
Results and Discussion
The results are shown in Figure 4. As in Experiment 1, preference scores above 50% indi-
cate a preference for the slowly moving object and preference scores below 50% indicate a
preference for the quickly moving object. Across the 10 pairwise combinations, the average
preference score was 54.1% (SEM = 1%). The chicks spent significantly more time by the
slowly moving object than the quickly moving object [two-tailed t test: t(10) = 4.87, p <
.001, Cohen’s d = 1.40]. The preference for slowly moving objects occurred both when the
chicks were reared with Object 1 [t(5) = 3.28, p = .02, Cohen’s d = 1.31] and Object 2
[t(4) = 3.45, p = .03, Cohen’s d = 1.54]. The Bayes factor (BF10) across the 10 pairwise
combinations was 58.1 (i.e., the alternative hypothesis that chicks preferred the slowly moving
object over the quickly moving object was 58.1 times more likely than the null hypothesis).
These data replicate Experiment 1 and provide additional evidence that newborn chicks have
a spontaneous preference for slowly moving objects.
To distinguish between the three possibilities described above, I computed the chicks’
preference scores for each of the 10 pairwise combinations. As shown in Figure 4, the chicks’
preference scores did not vary as a function of the ratio between the rotation speeds. For exam-
ple, on the trials where there was a 1:4 ratio between the rotation speeds, the chicks preferred
the slowly moving object on the 1.25 s vs. 5 s comparison, but not on the 5 s vs. 20 s com-
parison. The chicks preferred the slowly moving object only when the quickly moving object
rotated at the fastest (1.25 s) rotation speed. Thus, these results support the third possibility:
newborn chicks prefer slowly moving objects when other objects in the environment exceed
a particular speed threshold.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Experiment 2 results. The graph shows the average preference scores for each of
the 10 pairwise combinations of the five rotation speeds. Asterisks denote statistical significance:
∗p < .05; ∗∗p < .01; ∗∗∗p < .001 (two-tailed t tests). Error bars indicate standard error.
OPEN MIND: Discoveries in Cognitive Science
118
Spontaneous Preference for Slow Objects Wood
GENERAL DISCUSSION
Behavioral and computational research indicate that visual experience with slowly chang-
ing objects is essential for the development of object recognition (e.g., DiCarlo et al., 2012;
Földiák, 1991; Wiskott & Sejnowski, 2002). Newborn chicks, for instance, require experi-
ence with slowly moving objects to develop object recognition (Wood & Wood, 2016a). In
the present study, I examined whether newborn chicks actively seek out these optimal (slowly
moving) object features.
In Experiment 1, newborn chicks were reared with one object that rotated slowly and one
object that rotated quickly. The majority of the chicks had a strong preference for the slowly
rotating object on their first day of exposure to light. By the end of the experiment, all of the
chicks had developed a preference for the slowly rotating object. Experiment 2 replicated this
finding and showed that the preference is subject to a threshold effect. Together, these exper-
iments indicate that newborn chicks spontaneously develop a preference for slowly moving
objects. This preference was robust, yielding large effect sizes across objects, experiments,
and successive test days. Thus, this visual preference for slowly moving objects appears to be
a strong driver of behavior. The results also revealed substantial individual differences across
subjects, both in terms of the strength of the visual preference and the timing at which this
preference emerged during development. To my knowledge, this is the first study revealing
significant individual differences in the visual preferences of newborn subjects.
It is important to note that newborn chicks will readily spend time with quickly moving
objects when the environment lacks slowly moving objects (Wood & Wood, 2016a), which
rules out the possibility that newborn chicks simply avoid quickly moving objects in general.
A large literature on avian imprinting also shows that newborn chicks prefer moving objects
over stationary objects (reviewed by Horn, 2004). Thus, it is not the case that chicks simply
prefer objects that undergo the least amount of visual change. Finally, previous experiments
have shown that newborn chicks can recognize these virtual objects when the objects rotate
quickly, provided that the objects rotated slowly when being encoded into memory (Wood &
Wood, 2016a). Consequently, this preference for slowly moving objects cannot be explained
simply as a limitation in the speed of chicks’ perceptual abilities.
These findings complement previous studies reporting that newborn chicks have visual
preferences for faces (Rosa-Salva, Farroni, Regolin, Vallortigara, & Johnson, 2011), biologi-
cal motion (Vallortigara, Regolin, & Marconato, 2005), self-propelled motion (Mascalzoni,
Regolin, & Vallortigara, 2010), and speed changes (Rosa-Salva, Grassi, Lorenzi, Regolin, &
Vallortigara, 2016), and extend this literature by demonstrating that newborn chicks have a
visual preference for slowly moving objects. This is an important addition to the literature
because experience with slowly moving objects is required for the development of object
recognition (Wood & Wood, 2016a). This visual preference may drive the development
of object recognition by motivating newborn animals to seek out the best visual inputs for
constructing accurate mental models of objects. It would be interesting for future studies to
measure newborn chicks’ visual preferences for faces, biological motion, and self-propelled
motion with the automated controlled-rearing method described here, thereby allowing for a
direct comparison of the strength of these different early emerging visual preferences.
Additional research is needed to characterize the development of this visual preference
more precisely. For example, chicks might develop a preference for slowly moving objects from
their experiences with a slowly changing environment or from their experiences with their own
movements. Future studies are also needed to characterize the nature of this visual preference
OPEN MIND: Discoveries in Cognitive Science
119
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Spontaneous Preference for Slow Objects Wood
more formally. For instance, there is evidence that human infants prefer visual experiences
that are optimal in their complexity (e.g., Kidd, Piantadosi, & Aslin, 2012; Oudeyer & Smith,
2016). Likewise, it is possible that quickly moving objects are more complex (difficult to
process) than slowly moving objects, and that newborn chicks prefer objects that are optimal
in their complexity.2
In what ways do these results illuminate visual development in humans? On the neu-
ral level, there is growing evidence that birds and mammals use homologous neural circuits
to process sensory input (reviewed by Jarvis et al., 2005; Karten, 2013). Although these cir-
cuits are organized differently in birds and mammals (nuclear vs. layered organization, respec-
tively), the circuits share similarities in terms of cell morphology (Reiner, Yamamoto, & Karten,
2005), the connectivity pattern of the input and output neurons (Wang, Brzozowska-Prechtl,
& Karten, 2010), gene expression (Dugas-Ford, Rowell, & Ragsdale, 2012), and function
(Calabrese & Woolley, 2015). If newborn chicks and human infants use homologous neural
circuits to process sensory input—as these studies suggest—then chicks and infants might share
common visual preferences.
On the behavioral level, these results accord with a large body of work in developmental
psychology showing that human infants prefer slow speech over fast speech (e.g., Cooper &
Aslin, 1990; Fernald et al., 1989). When talking to infants, adults typically use a unique
speech register known as infant-directed speech (IDS), and a slow speaking rate is one of the
most distinctive characteristics of IDS. There is also evidence that infants learn words more
effectively when adults use slower speaking rates (Song, Demuth, & Morgan, 2010), akin
to how newborn chicks build more accurate object representations when raised with slowly
moving objects (Wood & Wood, 2016a). Thus, human infants and newborn chicks both prefer
slowly changing sensory input and show enhanced learning from slowly changing sensory
input.
Finally, an important contribution of this study is to introduce an automated controlled-
rearing method for studying visual preferences in newborn subjects. Using automation, it
is possible to sample behavior continuously (24/7) and chart how visual preferences change
across the first days of life. Automation also allows large amounts of data to be collected from
each subject, which reduces measurement error and increases the power of the design. Given
growing evidence that published findings are more likely to be false when measurement error
is high (e.g., Ioannidis, 2005; Loken & Gelman, 2017), automation may be a valuable tool
for combating the replication crisis in psychology.
In summary, the present study provides evidence for a spontaneous visual preference for
slowly moving objects in visually naïve newborn chicks. This study also introduces a pow-
erful automated controlled-rearing method for probing the origins and development of visual
preferences in a newborn model system.
2 Another possibility is that the fast rotation speed impaired the chicks’ ability to perceive continuous mo-
tion. Since both objects were presented at 24 frames per second, the quickly moving object rotated a greater
distance across successive frames than the slowly moving object. Thus, the slowly moving objects were more
temporally smooth than the quickly moving objects, a factor that influences the development of object recog-
nition (Wood, 2016). It would be interesting for future studies to systematically manipulate both the speed and
temporal smoothness of objects to examine how these factors influence newborn chicks’ early emerging visual
preferences.
OPEN MIND: Discoveries in Cognitive Science
120
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Spontaneous Preference for Slow Objects Wood
ACKNOWLEDGMENTS
Funded by NSF CAREER Grant BCS-1351892. I thank Samantha M. W. Wood and Brian Wood
for helpful comments on the manuscript.
AUTHOR CONTRIBUTIONS
JNW designed the research, performed the research, analyzed the data, and wrote the article.
REFERENCES
Berkes, P., & Wiskott, L.
rich repertoire of complex cell properties.
579–602.
(2005). Slow feature analysis yields a
Journal of Vision, 5,
Calabrese, A., & Woolley, S. M. N. (2015). Coding principles of the
canonical cortical microcircuit in the avian brain. Proceedings
of the National Academy of Sciences, 112, 3517–3522.
Cohen, J.
(1988). Statistical power analysis for the behavioral sci-
ences (2nd ed.). Hillsdale, NJ: Lawrence Erlbaum.
Cooper, R. P., & Aslin, R. N. (1990). Preference for infant-directed
speech in the first month after birth. Child Development, 61,
1584–1595.
DiCarlo, J. J., Zoccolan, D., & Rust, N. C.
(2012). How does the
brain solve visual object recognition? Neuron, 73, 415–434.
Di Giorgio, E., Loveland, J. L., Mayer, U., Rosa-Salva, O., Versace,
(2016). Filial responses as predisposed
E., & Vallortigara, G.
and learned preferences: Early attachment in chicks and babies.
Behavioural Brain Research, 325(Pt. B), 90–104. doi:10.1016/j.
bbr.2016.09.018
Dugas-Ford, J., Rowell, J. J., & Ragsdale, C. W.
(2012). Cell-type
homologies and the origins of the neocortex. Proceedings of the
National Academy of Sciences, 109, 16974–16979.
Fernald, A., Taeschner, T., Dunn, J., Papousek, M., de Boysson-
Bardies, B., & Fukui, I. (1989). A cross-language study of prosodic
modifications in mothers’ and fathers’ speech to preverbal in-
fants. Journal of Child Language, 16, 477–501.
Földiák, P.
(1991). Learning invariance from transformation se-
quences. Neural Computation, 3, 194–200.
Franzius, M., Sprekeler, H., & Wiskott, L.
(2007). Slowness and
sparseness lead to place, head-direction, and spatial-view cells.
PLoS Computational Biology, 3(8), e166.
Franzius, M., Wilbert, N., & Wiskott, L. (2011). Invariant object
recognition and pose estimation with slow feature analysis.
Neural Computation, 23, 2289–2323.
Horn, G.
(2004). Pathways of the past: The imprint of memory.
Nature Reviews Neuroscience, 5, 108–120.
Ioannidis, J. P. A. (2005). Why most published research findings
are false. PLoS Med, 2(8), e124. doi:10.1371/journal.pmed.
0020124
Jarvis, E. D., Gunturkun, O., Bruce, L., Csillag, A., Karten, H., Kuen-
zel, W., . . . Butlet, A. B.
(2005). Avian brains and a new
understanding of vertebrate brain evolution. Nature Reviews
Neuroscience, 6, 151–159.
Karten, H. J. (2013). Neocortical evolution: Neuronal circuits arise
independently of lamination. Current Biology, 23, R12–15.
Kidd, C., Piantadosi, S. T., & Aslin, R. N.
(2012). The Goldilocks
effect: Human infants allocate attention to visual sequences
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
that are neither too simple nor too complex. PLoS ONE, 7(5),
e36399.
Loken, E., & Gelman, A. (2017). Measurement error and the repli-
cation crisis. Science, 355, 584–585.
Mascalzoni, E., Regolin, L., & Vallortigara, G. (2010). Innate sen-
sitivity for self-propelled causal agency in newly hatched chicks.
Proceedings of the National Academy of Sciences, 107, 4483–
4485.
Mayer, U., Rosa-Salva, O., Lorenzi, E., & Vallortigara, G.
(2016).
Social predisposition dependent neuronal activity in the inter-
mediate medial mesopallium of domestic chicks (Gallus gallus
domesticus). Behavioral Brain Research, 310, 93–102.
Oudeyer, P.-Y., & Smith, L. B.
(2016). How evolution may work
through curiosity-driven developmental process. Topics in Cog-
nitive Science, 8, 492–502.
Reiner, A., Yamamoto, K., & Karten, H. J. (2005). Organization and
evolution of the avian forebrain. The Anatomical Record Part
A: Discoveries in Molecular, Cellular, and Evolutionary Biology,
287A, 1080–1102.
Rolls, E. T.
(2012).
Invariant visual object and face recognition:
Neural and computational bases, and a model, VisNet. Frontiers
in Computational Neuroscience, 6, 1–70.
Rosa-Salva, O., Farroni, T., Regolin, L., Vallortigara, G., & Johnson,
M. H. (2011). The evolution of social orienting: Evidence from
chicks (Gallus gallus) and human newborns. PLoS ONE, 6(4),
e18802.
Rosa-Salva, O., Grassi, M., Lorenzi, E., Regolin, L., & Vallortigara,
G.
(2016). Spontaneous preference for visual cues of animacy
in naïve domestic chicks: The case of speed changes. Cognition,
157, 49–60.
Song, J. Y., Demuth, K., & Morgan,
Effects of the
acoustic properties of infant-directed speech on infant word
recognition. Journal of the Acoustical Society of America, 128,
389–400.
(2010).
J.
Stone, J. V. (1996). Learning perceptually salient visual parameters
using spatiotemporal smoothness constraints. Neural Computa-
tion, 8, 1463–1492.
Vallortigara, G., Regolin, L., & Marconato, F. (2005). Visually inex-
perienced chicks exhibit spontaneous preference for biological
motion patterns. PLoS Biology, 3(7), e208.
Versace, E., & Vallortigara, G.
(2015). Origins of knowledge: In-
sights from precocial species. Frontiers in Behavioral Neuro-
science, 9, 338. doi:10.3389/fnbeh.2015.00338
Wang, Y., Brzozowska-Prechtl, A., & Karten, H. J. (2010). Laminar
and columnar auditory cortex in avian brain. Proceedings of the
National Academy of Sciences, 107, 12676–12681.
OPEN MIND: Discoveries in Cognitive Science
121
Spontaneous Preference for Slow Objects Wood
Wiskott, L., & Sejnowski, T. J.
(2002). Slow feature analysis: Un-
supervised learning of invariances. Neural Computation, 14,
715–770.
Wood, J. N.
(2013). Newborn chickens generate invariant object
representations at the onset of visual object experience. Proceed-
ings of the National Academy of Sciences, 110, 14000–14005.
Wood, J. N. (2014). Newly hatched chicks solve the visual binding
Wood, J. N., & Wood, S. M. W.
(2016a). The development of
newborn object recognition in fast and slow visual worlds. Pro-
ceedings of the Royal Society B: Biological Sciences, 283(1829).
doi:10.1098/rspb.2016.0166
Wood, J. N., & Wood, S. M. W.
(2016b). Measuring the speed of
newborn object recognition in controlled visual worlds. Devel-
opmental Science, 20, e12470. doi:10.1111/desc.12470
problem. Psychological Science, 25, 1475–1481.
Wood, J. N.
(2015). Characterizing the information content of a
newly hatched chick’s first visual object representation. Devel-
opmental Science, 18, 194–205.
Wood, J. N. (2016). A smoothness constraint on the development
of object recognition. Cognition, 153, 140–145.
Wood, J. N. (2017). Supplementary videos: The controlled-rearing
chambers with Objects 1 and 2. Supplemental material
for
“Spontaneous preference for slowly moving objects in visually
naive animals.” Open Mind: Discoveries in Cognitive Science,
1(2), 111–122. doi:10.1162/opmi_a_00012
Wood, S. M. W., & Wood, J. N. (2015). A chicken model for study-
ing the emergence of invariant object recognition. Frontiers in
Neural Circuits, 9(7), 1–12. doi:10.3389/fncir.2015.00007
Wyss, R., König, P., & Verschure, P.
(2006). A model of the ven-
tral visual system based on temporal stability and local memory.
PLoS Biology, 4(5), e120.
Zoccolan, D., Oertelt, N., DiCarlo, J. J., & Cox, D. D.
(2009).
A rodent model for the study of invariant visual object recog-
nition. Proceedings of the National Academy of Sciences, 106,
8748–8753.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
e
d
u
o
p
m
i
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
2
1
1
1
1
8
6
8
2
8
9
o
p
m
_
a
_
0
0
0
1
2
p
d
.
i
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
OPEN MIND: Discoveries in Cognitive Science
122