Specialized Face Perception Mechanisms Extract
Both Part and Spacing Information:
Evidence from Developmental Prosopagnosia
Galit Yovel1 and Brad Duchaine2
Abstract
& It is well established that faces are processed by mecha-
nisms that are not used with other objects. Two prominent
hypotheses have been proposed to characterize how informa-
tion is represented by these special mechanisms. The spacing
hypothesis suggests that face-specific mechanisms primarily ex-
tract information about spacing among parts rather than in-
formation about the shape of the parts. In contrast, the holistic
hypothesis suggests that faces are processed as nondecompos-
able wholes and, therefore, claims that both parts and spacing
among them are integral aspects of face representation. Here
we examined these hypotheses by testing a group of de-
velopmental prosopagnosics (DPs) who suffer from deficits
in face recognition. Subjects performed a face discrimination
task with faces that differed either in the spacing of the parts
but not the parts (spacing task), or in the parts but not the
spacing of the parts (part task). Consistent with the holistic
hypothesis, DPs showed lower performance than controls on
both the spacing and the part tasks, as long as salient contrast
differences between the parts were minimized. Furthermore,
by presenting similar spacing and part tasks with houses, we
tested whether face-processing mechanisms are specific to
faces, or whether they are used to process spacing information
from any stimulus. DPs’ normal performance on the tasks of
two houses indicates that their deficit does not result from
impairment
In
summary, our data clearly support
face-specific holistic
hypothesis by showing that
face perception mechanisms
extract both part and spacing information. &
in a general-purpose spacing mechanism.
INTRODUCTION
Extensive evidence suggests that upright faces are pro-
cessed by specialized mechanisms that are not used to
process many other object classes. Studies using func-
tional magnetic resonance imaging (fMRI) (Kanwisher,
McDermott, & Chun, 1997; McCarthy, Puce, Gore, &
Allison, 1997), event-related potentials (Eimer, 2000;
McCarthy, Puce, Belger, & Allison, 1999; Puce, Allison, &
McCarthy, 1999; Bentin, Allison, Puce, Perez, & McCarthy,
1996), and single-cell recordings (Kreiman, Koch, &
Fried, 2001; Gross, 1992; Perrett, Rolls, & Caan, 1982)
indicate neural substrates that show face-selective re-
sponses. Neuropsychological studies have shown that
face and object recognition can each be selectively im-
paired (Duchaine, Yovel, Butterworth, & Nakayama, in
press; Duchaine & Nakayama, 2005; Moscovitch &
Moscovitch, 2000; Moscovitch, Winocur, & Behrmann,
1997; Farah, 1996; Sergent & Signoret, 1992). Finally,
behavioral studies have demonstrated that faces are
processed in a more configural or in a more holistic
manner than objects (Farah, Wilson, Drain, & Tanaka,
1998; Young, Hellawell, & Hay, 1987; Tanaka & Farah,
1993, 2003; Yin, 1969). Two hypotheses have been pro-
1Tel-Aviv University, 2University College, London
posed to characterize the nature of specialized face
representation: the face-specific spacing hypothesis and
the face-specific holistic hypothesis.
Face-specific Spacing Hypothesis
Studies have shown that we are highly sensitive to subtle
displacements of face parts (e.g., eyes, nose, and mouth)
in upright faces (Haig, 1984) but not in faces that are
inverted or in faces with negative contrast (Kemp,
McManus, & Pigott, 1990). Such findings led several
researchers to suggest that face perception mechanisms
are special in that they extract a precise distance among
face parts (i.e., spacing), whereas information about the
characteristics of parts is represented by general object
recognition mechanisms (for a review, see Maurer,
Grand, & Mondloch, 2002).
This face-specific spacing hypothesis initially drew
support from studies examining face inversion effect.
The face inversion effect refers to a much larger effect
that orientation inversion has on face recognition com-
pared to object recognition. Studies that have employed
the face inversion effect to test the spacing hypothesis
have used two sets of face stimuli (Mondloch, Le Grand,
& Maurer, 2002; Le Grand, Mondloch, Maurer, & Brent,
2001; Freire, Lee, & Symons, 2000). For the spacing
D 2006 Massachusetts Institute of Technology
Journal of Cognitive Neuroscience 18:4, pp. 580–593
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
f
.
.
t
.
o
n
1
8
M
a
y
2
0
2
1
sets, the face parts (e.g., eyes and mouth) were identical
in all faces, but the spacing of the parts varied. Con-
versely, in the part sets, the face parts varied, but the
spacing of those parts remained constant. These faces
were presented in a same–different matching task in an
upright and in an inverted orientation. If face percep-
tion mechanisms are specialized for the processing of
spacing but not for the processing of part-based infor-
mation,
inversion should result in a larger cost for
matching spacing information rather than for matching
part-based information.
Several studies found support for this prediction. In
the first published study, Freire et al. (2000) found a very
large inversion effect for discrimination of spacing in-
formation (26%) but found no inversion effect for discrim-
ination of part-based information. Other studies have
also found greater inversion effects for spacing informa-
tion than for part information (Mondloch et al., 2002;
Le Grand et al., 2001). These findings suggest that dis-
tinct mechanisms mediate the processing of information
about spacing among parts and the identity of parts. Fur-
thermore, because faces are represented in a qualitatively
different manner than are objects, it has been assumed
that the mechanisms extracting spacing information are
specialized for faces, whereas those used for part repre-
sentation are used with a wide range of objects.
This account predicts that individuals with impaired
face recognition may show deficits in processing spacing
but not deficits in part-based information from faces.
Support for this prediction was found in a group of
subjects who suffered from infantile cataracts that were
removed in their first year of life. Although low-level
visual abilities in these subjects were intact, they showed
face recognition difficulties in adulthood. When tested
with a face discrimination task involving part and spac-
ing information, these individuals performed part dis-
criminations normally, but were impaired with spacing
discriminations (Le Grand, Mondloch, Maurer, & Brent,
2003; Le Grand et al., 2001). Similar results were recently
found in adults with Williams syndrome (Karmiloff-Smith
et al., 2004). The dissociability of these processes gained
more support when it was shown that children develop
sensitivity to spacing changes more slowly than they
develop sensitivity to part changes (Mondloch et al.,
2002). Note that these forgoing studies used the same
face exemplar known as the Jane face (Le Grand et al.,
2001). However, despite apparently clear results sup-
porting the face-specific spacing hypothesis, some re-
cent studies have challenged it.
Face-specific Holistic Hypothesis
The face-specific holistic hypothesis suggests that faces
are processed as nondecomposed wholes and that face
parts are processed interactively rather than indepen-
dently. Thus, in contrast to the spacing hypothesis, the
holistic hypothesis predicts that information about face
parts and spacing among parts is processed by a common
mechanism. Consistent with the holistic hypothesis,
three studies with normal subjects have found that
inversion has comparable effects on spacing and part
discrimination in faces (Malcolm, Leung, & Barton,
2005; Riesenhuber, Jarudi, Gilad, & Sinha, 2004; Yovel
& Kanwisher, 2004a). These findings are consistent with
earlier studies that examined the recognition of face parts
(i.e., eyes, nose, and mouth) in the context of a face or in
isolation (whole–part effect) (Tanaka & Sengco, 1997;
Tanaka & Farah, 1993). These studies found that when
subjects discriminated face parts in the context of a whole
face, performance was lower when faces were presented
in an inverted orientation than when they were pre-
sented in an upright orientation. Thus, in contrast to
the spacing hypothesis, these findings suggest that parts
in upright faces are processed by a mechanism different
from that used for parts in inverted faces.
Several other studies provide support for the holistic
hypothesis and suggest that face mechanisms extract
both spacing-based and part-based information. In an
fMRI study, Yovel and Kanwisher (2004a) investigated
whether face-selective brain areas respond differently to
part and spacing information in faces; they found no dif-
ference in responses to spacing and parts. Additionally,
similar to the behavioral findings, the fMRI inversion effect
(i.e., lower response to inverted than upright faces) was
similar for the spacing and part task. Furthermore, Yovel
and Kanwisher (2004b) examined individual differences
in performance for part and spacing discrimination and
found that normal subjects showed a large positive cor-
relation between performance on spacing and part dis-
crimination tasks for upright faces but not for inverted
faces or houses. These findings suggest that a common
mechanism is used to process information about spacing
and parts for faces, whereas distinct mechanisms are
used for the discrimination of spacing and parts for
nonfaces. All of these findings suggest that special mech-
anisms used with faces represent information about
both parts and spacing among parts and are, therefore,
inconsistent with the face-specific spacing hypothesis.
Domain-general Spacing Hypothesis
The two hypotheses discussed above both suggest that
face perception, not object perception, is carried out by
different mechanisms. Such face-specific hypotheses are
motivated by demonstrations that faces and other types
of objects are dissociable (Duchaine et al.,
in press;
Moscovitch et al., 1997). However, in any experiment
showing a dissociation between faces and objects, recog-
nition of faces can only be compared to a limited number
of object classes. It may be that the object classes used
in such experiments did not demand spacing processing
like faces did. As a result, this leaves open the possibility
that the mechanisms that process faces are not face-
specific but are domain-general mechanisms that operate
Yovel and Duchaine
581
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
t
f
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
on any class with diagnostic spacing information. Accord-
ing to this general spacing hypothesis, impairments with
face spacing discriminations will be accompanied by non-
face spacing discriminations (Behrmann, Avidan, Marotta,
& Kimchi, 2005; Levine & Calvanio, 1989).
One direct way to examine the domain-general spac-
ing hypothesis is to compare the magnitude of the inver-
sion effect for spacing and part discriminations with faces
to other object classes. Such a comparison is essential
because an inversion effect by itself is not informative
with respect to the nature of specialized face-processing
mechanisms. Only an inversion effect that is larger for
faces than for nonfaces can provide information about
the possibility of specialized face-processing mechanisms.
Yovel and Kanwisher (2004a) manipulated spacing-based
and part-based information in house stimuli in the same
way they manipulated faces. They found large inversion
effects for face parts and face spacing, but they found
no inversion effects for house part or house spacing
discriminations. Furthermore, the fMRI response of fusi-
form face area (FFA) showed a similar response for part
and spacing discrimination in houses, and the response
was much lower than the response to faces. These find-
ings suggest that the spacing processing applied to faces
is not used with other classes of objects.
Developmental Prosopagnosia as a Test Case
for the Three Hypotheses
The conflicting results discussed above make it difficult
to determine which hypothesis best characterizes the
mechanisms performing face recognition. In this study,
our goal was to directly address these three hypotheses
by testing individuals who show severe face perception
deficits. These individuals are expected to show difficul-
ties with all tasks that are normally mediated by intact
face-processing mechanisms. Thus, to test the predic-
tions of each hypothesis, we tested developmental
prosopagnosics (DPs) on perceptual matching tasks
with faces and houses that differed in part or in spacing
information.
DPs are individuals who failed to develop normal face
recognition abilities (Behrmann & Avidan, 2005). There
are a variety of causes of this condition, including genet-
ic deficits (Behrmann et al., 2005; Duchaine & Nakayama,
2005; Duchaine, 2000; Bentin, Deouell, & Soroker, 1999;
de Haan, 1999), early brain damage (Barton, Cherkasova,
Press, Intriligator, & O’Connor, 2003), and early visual
problems (Le Grand, Mondloch, Maurer, & Brent, 2004;
Le Grand et al., 2001, 2003). Difficulties with other visual
abilities, such as recognition of facial expressions and
objects, sometimes accompany the face recognition
impairments of DPs, but some cases appear to be
in press; Duchaine
quite selective (Duchaine et al.,
& Nakayama, 2005; Duchaine, Dingle, Butterworth, &
Nakayama, 2004; Nunn, Postma, & Pearson, 2001; Bentin
et al., 1999).
To resolve the conf licting results from face dis-
crimination tasks, we used two sets of face stimuli (see
Figure 1). One set, referred to here as Alfred, yielded
similar inversion effects in controls for the spacing and
part tasks (Yovel & Kanwisher, 2004a); these results
Figure 1. The face and house
stimuli were varied in two
ways. For the part set, the eyes
and mouth in the face and the
windows and door in the
house were replaced by similar
parts from different exemplars.
The spacing of the parts was
not changed. Conversely, to
create the spacing set, the
position of the eyes and mouth
or the position of the door
and windows was varied. These
manipulations were determined
in a pilot study based on
changes that generated average
performance level in the
dynamic range (about 75%)
across the spacing and part
conditions of faces and houses.
582
Journal of Cognitive Neuroscience
Volume 18, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
t
f
.
o
n
1
8
M
a
y
2
0
2
1
support the face-specific holistic hypothesis. The other
set, known as the Jane face, was used in experiments
that revealed a larger inversion effect for the spacing
task than the part task in normal controls (Mondloch
et al., 2002; Le Grand et al., 2001). The Jane faces have
been also used in studies with patients with infantile
cataract (Le Grand et al., 2001, 2003, 2004), in a study
of adults with Williams syndrome (Karmiloff-Smith
et al., 2004), and in a developmental study that found
dissociation between spacing and parts (Mondloch
et al., 2002). In addition, we tested the prosopagnosics
with a house discrimination task with identical task
demands to address the domain of spacing processes
used with faces.
The predictions of the different hypotheses are pre-
sented in Figure 2. In (A), the face-specific spacing
hypothesis predicts that prosopagnosics will be im-
paired with face spacing items, but they will be normal
with other discriminations. In (B), the face-specific
holistic hypothesis predicts that prosopagnosics will be
Figure 2. Three hypotheses were tested. (A) The face-specific
spacing hypothesis predicts that DPs will show a deficit with spacing
information only for faces. (B) The face-specific holistic hypothesis
predicts that DPs will show a deficit on both the part and spacing tasks
for faces, but intact performance for houses. (C) The domain-general
spacing hypothesis predicts that DPs will show deficits in processing
spacing information from both faces and houses.
impaired with both face part and face spacing, but they
will perform normally with houses. In (C), the domain-
general spacing hypothesis predicts that prosopagnosics
will be impaired with spacing discriminations with both
faces and houses, but they will perform normally for
both types of part discriminations.
RESULTS
Experiment 1
Eight subjects with developmental prosopagnosia and
34 controls participated in Experiment 1. Table 1 displays
their standardized scores on three face recognition
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
t
.
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Table 1. Standardized Scores of Each of the DPs on the
Three Face Recognition Tasks
DPs
Sex
Age
(years)
Famous
Faces 1
Face
Old–
New A0
Face
Old–
New RT
CFMT
Experiment 1: Alfred–Jane experiments
A.C.
D.D.
M
F
Edward M
J.H.
K.L.
L.A.
M.L.
R.S.
M
F
F
M
F
20
32
53
23
46
40
24
27
(cid:2)4.8
(cid:2)4.7
(cid:2)10.1
(cid:2)12.5
(cid:2)6.5
(cid:2)4.9
(cid:2)15.3
(cid:2)4
(cid:2)10.6
(cid:2)4.3
(cid:2)6.4
(cid:2)7.2
(cid:2)4.1
(cid:2)3
(cid:2)6.4
(cid:2)5.6
(cid:2)1.7
(cid:2)8.4
(cid:2)4.2
(cid:2)5.1
(cid:2)5
(cid:2)6.2
(cid:2)4
0
(cid:2)2.5
(cid:2)2.3
(cid:2)2.4
(cid:2)2.6
(cid:2)2.9
(cid:2)2
(cid:2)2.4
(cid:2)2.6
Control mean
23.6/22.6
0.96 971 msec 80.4%
Control SD
1.4/1.9
0.03 205
11
Experiment 2: Ann experiments
A.M.
K.M.
D.D.
N.Z.
J.W.
B.K.
F
F
F
F
M
M
64
21
32
21
25
56
Control mean
Control SD
(cid:2)0.2
(cid:2)2.5
(cid:2)5.1
(cid:2)5.7
(cid:2)7.5
(cid:2)3.6
52.5
6.6
(cid:2)2.5
(cid:2)2.0
(cid:2)4.3
(cid:2)3.5
(cid:2)9.1
(cid:2)4.7
(cid:2)9.0
(cid:2)2.7
(cid:2)8.4
(cid:2)4.2
(cid:2)6.2
(cid:2)7.5
(cid:2)1.9
(cid:2)1.9
(cid:2)2.3
(cid:2)3.8
(cid:2)3.7
(cid:2)1.9
0.96 971 msec 80.4%
0.03 205
11
Z scores were computed by subtracting the control average from each
DP’s percentage of correct responses and then dividing by the control
standard deviation [(DP value (cid:2) control average)/control SD]. Values
worse than the control mean are negative. For Famous Face 1, two
averages and two standard deviations are presented. The first is for
college-age controls, whereas the second is for adults 45–55 years of
age. A.C. was not tested with the famous face test used, but scored 4.8
standard deviations below the mean on Famous Faces 2.
RT = reaction time; CFMT = Cambridge Memory Face Test.
Yovel and Duchaine
583
tasks. The tests and the control subjects are described in
Methods. The DPs had accuracy scores that were two
standard deviations or more below the control mean on
all three tests (see Table 1).
Accuracy
We first describe results with Alfred faces, which yield
similar inversion effects for the spacing and part tasks in
controls (Yovel & Kanwisher, 2004a). This face was
constructed such that performance levels are away from
ceiling or floor and are similar for upright spacing and
part discriminations (see Methods for further informa-
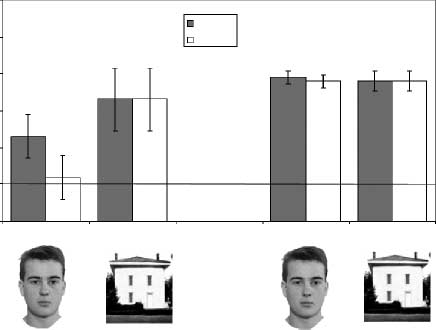
tion). Figure 3 shows proportions that are correct for
part and spacing discriminations with Alfred faces and
house stimuli. If we first consider the control results
(presented on the right side of the figure), we see that
performance was matched across all conditions so that
the difficulty of each discrimination was comparable. In
addition, because the mean proportion of correct re-
sponses ranged from 78% (SD = 10.0%) to 79% (SD =
9.6%), ceiling or floor effects had little or no influence
on control scores.
The results for the DPs show a pattern very different
from that of the controls, and this pattern is consistent
with the predictions of the face-specific holistic hypoth-
esis (see Figure 2B). The DPs’ average performance was
similar to that of the controls on the house task, but it
was much worse than that of the controls on the face
task: Group (cid:3) Stimulus interaction, F(1,39) = 20.37, p <
.0001. Whereas controls averaged 78.5% on the two face
discriminations, DPs averaged 57.4%. In contrast, the
DPs showed good performance with the houses (75%)
DPs
Spacing
Part
Controls
t
c
e
r
r
o
c
n
o
i
t
r
o
p
o
r
p
Face
House
Face
House
Figure 3. Proportion of correct responses for the DPs and the
controls on the face and house tasks. For the controls, the proportion
of correct responses was similar for all conditions. Consistent with
the face-specific holistic hypothesis, DPs show deficits on both part
and spacing discriminations of faces, but normal performance on
the house tasks. Error bars represent the standard error of the
difference between the part and spacing conditions for faces and
houses. The solid line indicates chance performance level.
relative to the controls (78%). Notably, the DPs,
like
the controls, showed no differences in performance on
the spacing and part tasks for either faces or houses: the
three-way Group (cid:3) Stimulus (cid:3) Task interaction was
not significant, F(1,40) = 1.82, p = .19. Thus, their per-
formance with spacing and part discriminations is in-
consistent with predictions of the face-specific spacing
hypothesis and the domain-general spacing hypothesis.
Because there is often substantial variability among
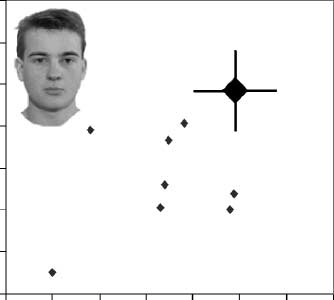
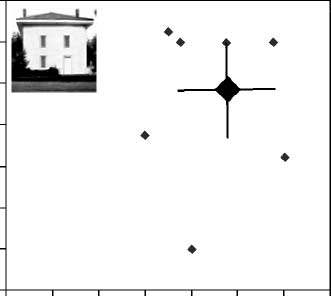
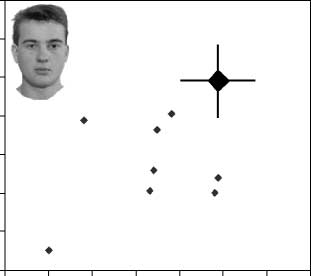
DPs, we created Figure 4 to display individual DP results.
for each DP and the
It displays performance level
average performance of the control group (larger dia-
mond) for the part and spacing tasks for faces (A) and
for houses (B). The individual results with faces make it
clear that individual DPs tended to show a similar
pattern, as the average results in that performance with
both spacing and part discriminations were usually poor.
In contrast, nearly all of the DPs were within the normal
range with both types of discriminations with houses.
These findings suggest that impairment in developmen-
tal prosopagnosia is neither a general deficit in process-
ing spacing information in faces or nonfaces (Figure 2C),
nor is it a face-specific deficit in processing spacing
information (Figure 2A) but is a specific face-processing
impairment affecting both spacing-based and part-based
information (Figure 2B).
Reaction Times
Although the primary dependent measure in our study
was accuracy, we also reported reaction time (RT) data.
Analysis of RT for correct responses (trimmed 200 msec <
RT < 1500 msec) showed slower RTs to the faces for
DPs (846 msec) than for controls (754 msec), but the
difference did not reach significance, F(1,40) = 3.22, p =
.08. RTs were slower for the face part (801 msec) than
the face spacing (741 msec) tasks for both controls and
DPs: main effect of Task, F(1,40) = 18.84, p < .0001. More
important, Group (cid:3) Task interaction was not significant,
F(1,40) < 1, which suggests that DPs showed a pattern
of response to the part and spacing face tasks similar to
that of controls.
There was no difference between DPs and controls in
RTs on the house tasks. RTs were faster on the house
part tasks (765 msec) than on the house spacing tasks
(729 msec); main effect of Task, F(1,39) = 5.01, p < .05,
for both DPs and controls. The Group (cid:3) Task interac-
tion was not significant, F(1,39) < 1, which suggests that
DPs and controls showed a pattern of response to the
part and spacing house tasks similar to that of controls.
Resolving Discrepancies with Previous Studies:
Comparing Alfred and Jane
Our data show comparable impairments for the DPs
with part and spacing processing with the Alfred faces.
584
Journal of Cognitive Neuroscience
Volume 18, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
4
-
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
t
.
f
o
n
1
8
M
a
y
2
0
2
1
sented in a mixed design, whereas the part and spacing
tasks with Jane faces were blocked. Other task parame-
ters, such as exposure duration, may also underlie these
differences.
Thus, to assess whether differences between groups
or task parameters are responsible for the discrepancy,
we compared how the DPs in our study performed with
Alfred and Jane discriminations in a mixed design. The
DPs should perform similarly with Alfred and Jane if the
discrepancy is caused by one of the above-mentioned
possibilities. However, if our DPs show better perform-
ance on the part task than on the spacing task with the
Jane faces, it will indicate that the discrepancy is due to
specific characteristics of the face stimuli that have been
used in the conflicting experiments.
Accuracy
In our face experiment, we included the same Jane faces
that were used in past experiments (Karmiloff-Smith
et al., 2004; Mondloch et al., 2002; Le Grand et al.,
2001, 2003). This allows us to directly compare the
Alfred faces with the Jane faces under the same exper-
imental conditions in the same sample of subjects.
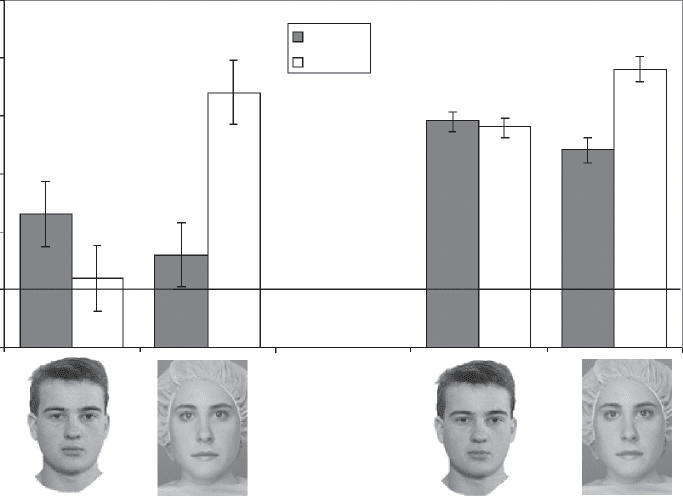
Figure 5 displays the proportion of correct responses
for the spacing and part tasks with Alfred and Jane
faces for the DPs and the controls. As is evident from
the figure, the DPs’ performance showed a different
pattern with Jane and Alfred faces. Our controls, like
those in other studies with Jane faces (Karmiloff-Smith
et al., 2004; Mondloch et al., 2002; Le Grand et al.,
2001, erratum), scored better on the part discrimina-
tions (88%) than on the spacing discriminations (74%).
Like Le Grand et al. (2001), performance for the Jane
part task did not differ between the DPs (83%) and
the controls (88%), t(40) = 2.72, p = .11. Again, as with
the patients with cataract and subjects with Williams
syndrome, the DPs performed more poorly with the
Jane spacing task (62%) than controls (74%), t(40) =
8.67, p < .006. This dissociation between discrimi-
nation of parts and spacing is inconsistent with our
findings with Alfred faces, which yielded lower per-
formance for DPs than for controls on both the part
tasks, t(40) = 30.78, p < .0001, and the spacing tasks,
t(40) = 14.42, p < .001. Accordingly, an analysis of
variance (ANOVA) on proportions of correct responses
with Group as a between-subject factor and with Task
and Face Stimuli (Alfred, Jane) as a repeated measure
factor yielded a significant three-way Group (cid:3) Face
Stimulus (cid:3) Task interaction, F(1,40) = 4.73, p < .05 (see
Figure 5).
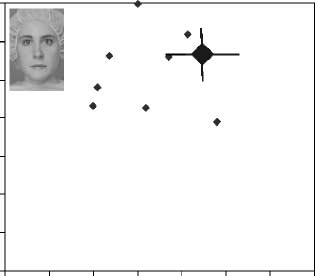
Figure 6 shows the performance of each DP and the
average performance of the control group on the spac-
ing and part tasks for the Jane and Alfred faces. Whereas
the DPs are shifted down relative to the control mean
on both the spacing axis and the part axis with Alfred
faces, we see that they are shifted down only on the
Yovel and Duchaine
585
Figure 4. The performance of each DP is presented along with
the average performance of controls (large diamond) on the (A) face
tasks and on the (B) house tasks. Error bars represent one standard
deviation above and below the mean for the spacing and part tasks
in the control group (see Appendix I).
In contrast, studies that used the Jane faces found
that individuals with face-processing deficits have im-
pairments with spacing but not with part-based face
information (Karmiloff-Smith et al., 2004; Le Grand
et al., 2001). There are a number of differences be-
tween these studies that may account for these incon-
sistent findings.
First, this discrepancy could simply be the result of
testing groups with qualitatively different face recogni-
tion impairments. For instance,
it could be that the
DPs that we have tested have impairments with both
dissociable mechanisms hypothesized to process face
spacing and face parts, whereas individuals with face
difficulties due to visual deprivation are only impaired
with the mechanism used with face spacing. Another
possible explanation for the discrepancy is that the
spacing and part discriminations with Alfred were pre-
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
4
-
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
f
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 5. The proportion of
correct responses of DPs and
controls on the discrimination
of spacing and parts in Alfred
and Jane. Consistent with
previous reports that used
the Jane faces, we find higher
performance on the part tasks
than on the spacing tasks
for both controls and DPs.
However, when performance
on the part and spacing tasks
is matched away from ceiling
effects in controls (Alfred
faces), DPs show deficits on
both face tasks. Error bars
represent the standard error
of the difference between the
part and spacing conditions
for faces and houses. The
solid line indicates chance
performance level.
t
c
e
r
r
o
c
n
o
i
t
r
o
p
o
r
p
DPs
Spacing
Parts
Controls
D
o
w
n
l
o
a
d
e
d
f
r
o
m
spacing axis with Jane faces. On the Jane task, the con-
trol group and seven of eight DPs showed a higher
percentage of correct responses on the part task than
on the spacing task. Thus, our data for the Jane faces
are consistent with previous reports that used these
faces. More important, these data show that the dis-
crepancies between our findings with Alfred faces and
prior findings with Jane faces are not due to differ-
ences between our subject groups or specific task
parameters, but lie in the nature of the face stimulus
sets that were used in the different studies. We will
consider these differences and their implications in
Experiment 2.
Reaction Times
Consistent with the accuracy results, response times
were faster for the Jane part task (731 msec) than for
the Jane spacing task (805 msec), F(1,40) = 6.43, p <
.02, for both controls and DPs. Whereas DPs were worse
than controls on the spacing tasks but not on the part
tasks for accuracy data, DPs were slower than controls
on both the part tasks (848 msec) and the spacing
tasks (898 msec): main effect of Task, F(1,40) = 10.84,
p < .005, but no Group (cid:3) Task interaction. Because
Le Grand et al. (2001) did not report the response time
data of their prosopagnosic group, we cannot determine
whether our findings with Jane are in agreement with
their observations.
In summary, our findings show that prosopagnosic
individuals showed deficits in both spacing-based and
part-based processing for Alfred faces but not for Jane
faces. In Experiment 2, we investigate the source of the
discrepancy between the two face exemplars.
Experiment 2
One important difference between the manipulation of
part information in Alfred and Jane faces was that Alfred
part changes primarily involved the shape of the face
parts. In contrast, the Jane part changes involved the
shape of the parts and salient changes in the brightness
and contrast of the different parts. The most notable
brightness/contrast difference is that two of the women
are wearing eyeliner or lipstick, whereas two are not.
Leder and Bruce (2000) have shown that discrimination
of faces that differ in color information does not pro-
duce the face inversion effect, which suggests that such
information does not involve specialized face-processing
mechanisms.
To resolve the discrepancies between the results we
obtained with the Jane face and with the Alfred face, we
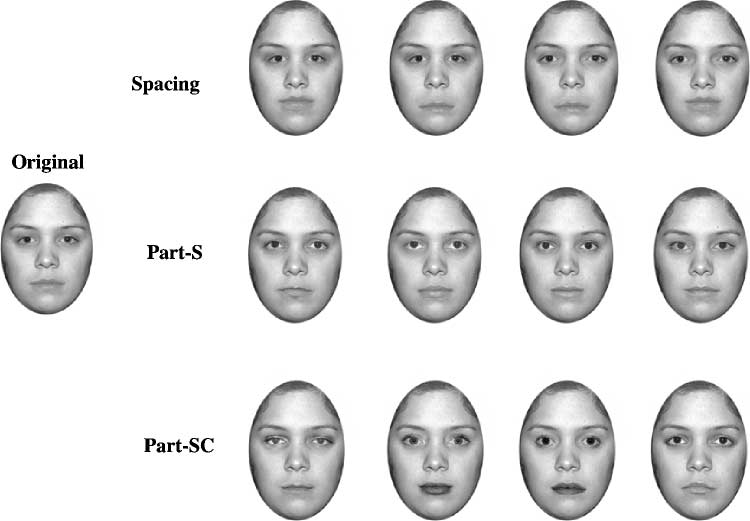
generated a new face (Ann), which, like Jane, was a female
face in which the hair was cropped. The Ann face was
manipulated in three ways (see Figure 7 and Methods):
1. Spacing: Similar to the spacing manipulation of the
Alfred and Jane faces, we generated four faces that
differed in the distance between the eyes and the
distance between the nose and the mouth.
2. Part-SC (shape + contrast) (‘‘Jane-like’’): The eyes
and mouth from four different faces replaced the eyes
and mouth of the original
face. Like the Jane part
manipulation, the parts differed in both shape and
contrast/brightness (e.g., lipstick).
586
Journal of Cognitive Neuroscience
Volume 18, Number 4
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
4
-
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
f
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Six subjects with developmental prosopagnosia and
15 controls participated in Experiment 2. Five of these
subjects did not participate in Experiment 1. Table 1
displays their standardized scores on three face recog-
nition tasks.
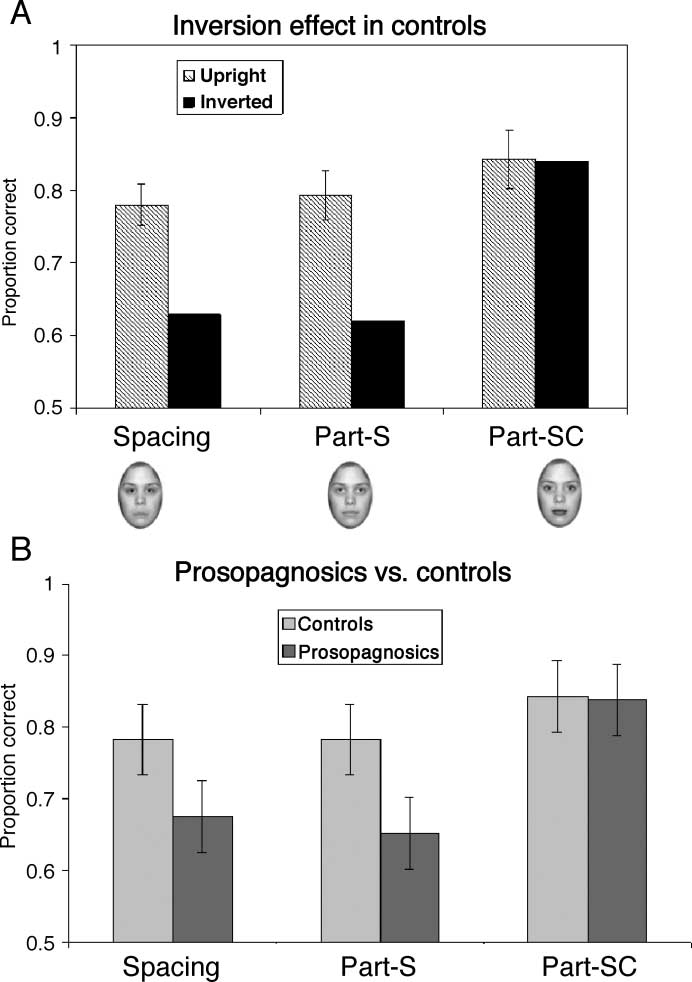
Face Inversion Effect in Controls
Subjects first performed a discrimination task with the
spacing and part-S tasks, followed by a discrimination
task that included the spacing and part-SC tasks. Control
subjects were presented with both upright and inverted
face tasks. The prosopagnosic subjects performed only
the upright face tasks.
Consistent with our predictions, we found an inver-
sion effect in the spacing and part-S (Alfred-like) tasks,
but not in the part-SC ( Jane-like) task (Figure 8A).
The Type (spacing, part-S, part-SC) by Orientation
(upright,
inverted) interaction was highly significant,
F(2,14) = 13.23, p < .0001, which reflects a signifi-
cantly larger inversion effect for part-S than for the
part-SC task, F(2,14) = 14.75, p < .001, and for spacing
than for the part-SC task, F(2,14) = 20.15, p < .001,
but no difference in the magnitude of the inversion
effect for the part-S and spacing tasks, F(2,14) < 1
(Figure 8A).
Performance of Prosopagnosic Subjects
Consistent with the inversion effect findings, prosop-
agnosic subjects performed significantly worse in the
spacing task (prosopagnosics, 64%; controls, 78%),
t(1,19) = 3.76, p < .001, and in the part-S task (prosop-
agnosics, 68%; controls, 80%), t(1,19) = 2.98, p < .01,
than the controls. However, they did not differ from
the control group in their performance for the part-SC
task (prosopagnosics, 84%; controls, 85%) ( Jane-like),
t(1,19) = 0.11 p > .90 (Figure 8B). These findings
suggest that prosopagnosics can discriminate faces that
differ in brightness/contrast as well as controls, but
not between faces that primarily differ in the shape
of the part.
DISCUSSION
It is often argued that specialized face perception mech-
anisms primarily process the spacing among face parts,
but not the parts themselves (Maurer et al., 2002; Leder
& Bruce, 2000). The data we present in this article
challenge this idea. We found that individuals who
suffer from developmental prosopagnosia show face
discrimination deficits for faces that differ in spacing
among parts and for faces that differ in parts in which
contrast/brightness differences are minimal. This in-
version effect demonstrates that the stimulus has en-
Yovel and Duchaine
587
Figure 6. Performance on the (A) Alfred task and on the (B) Jane
task for each DP is presented along with the average performance
of controls (large diamond). Error bars represent one standard
deviation above and below the mean for each task in the control
group (see Appendix I).
3. Part-S (shape) (‘‘Alfred-like’’): The eyes and
mouth from four different faces replaced the eyes and
mouth of the original face, but differences in contrast/
brightness were minimal.
We hypothesized that face mechanisms are not nec-
essary to discriminate faces that differ in salient contrast/
brightness information, which can be performed by
lower-level visual processing mechanisms. Therefore,
we expect no inversion effect in controls and no deficit
in discrimination of the part-SC Ann task in prosopag-
nosic subjects. In contrast, we hypothesized that face
mechanisms are as necessary for the discrimination of
parts that primarily differ in shape as they are for the
extraction of spacing information. Thus, we expect
normal subjects to show an inversion effect in the
part-S Ann task that is as large as in the spacing task,
and we predict that prosopagnosic subjects will have
difficulty in discriminating faces that differ in such part
information.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
t
.
.
f
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 7. The three Ann sets
used in Experiment 2. The Ann
spacing set was used in both
blocks involving Ann faces.
Eye spacing and the distance
between the nose and mouth
were manipulated. In the
Alfred-like part-S set, the eyes
and mouth were varied by
substituting eyes and mouths
that differed in shape, but
not in contrast/brightness. In
contrast, the eyes and mouths
in the Jane-like part-SC set
varied in shape and contrast/
brightness. By inverting the
figure, readers can experience
the effect of inversion on the
different sets.
gaged face-specific mechanisms. Thus, these findings
suggest that specialized face perception mechanisms
process both part and spacing information. Our find-
ings are consistent with recent reports of a similar-sized
inversion effect on part and spacing discrimination
tasks for faces (Malcolm et al., 2005; Riesenhuber
et al., 2004; Yovel & Kanwisher, 2004a) and a similar
fMRI response in face-selective brain areas (i.e., FFA)
for the spacing and part tasks (Yovel & Kanwisher,
2004a).
In addition, our results indicate that the deficit that
these DPs show on the discrimination of part and spac-
ing information is not a general deficit in the extrac-
tion of spacing information because they performed
as well as controls did in a closely matched task with
houses (Figure 3). This dissociation is consistent with
Yovel and Kanwisher (2004a), who found an inversion
effect for the spacing and part tasks for faces but not
for houses, and a similar FFA response on both tasks
for houses that was lower than that for faces. Thus,
our data are inconsistent with the general spacing
hypothesis.
Our findings of a similar impairment for both spac-
ing and part processing in DPs are inconsistent with
reports that used the Jane faces to study part and
spacing discrimination in neuropsychological subjects
(Karmiloff-Smith et al., 2004; Le Grand et al., 2001).
These individuals showed a deficit in processing spac-
ing-based but not part-based face information. One
important difference between the manipulation of
part information in Alfred and Jane was that Alfred
part changes involved primarily the shape of the face
parts. In contrast, the Jane part changes involved sa-
lient changes in brightness/contrast. The most notable
brightness/contrast differences are that two of the wo-
men are wearing eyeliner, whereas two are not, and two
are wearing lipstick, whereas two are not. To directly
address the inconsistent findings with the two face sets,
we generated a new face stimulus that was designed
to be like the Jane-part (part-SC) set and the Alfred-part
(part-S) set (see Figure 7). In other words, in the Jane-
like manipulation, the faces differed in both shape and
contrast/brightness information, whereas in the Alfred-
like part manipulation, contrast differences between
the face parts were minimized and the parts primarily
differed by shape. Our findings clearly show that con-
trol subjects did not show inversion effect on this task.
These findings are consistent with those of Leder and
Bruce (2000), who reported no inversion effect for the
discrimination of faces that differed in color informa-
tion. Furthermore, we found that our prosopagnosic
subjects did as well as controls in discriminating these
faces. In contrast, when contrast/brightness differences
across the faces were minimized, the controls showed a
similar inversion effect on the spacing and part tasks,
and prosopagnosics were as impaired on the part task
intact face perception
as on the spacing task. Thus,
mechanisms are not necessary for discriminating faces
that differ by salient contrast information. However,
they are as necessary for the representation of shape-
based part information as they are for the represen-
tation of spacing information. This accounts for the
observation that prosopagnosics often report that they
are able to recognize people by way of distinctive facial
characteristics, such as Mikhail Gorbachev’s port-wine
stain.
588
Journal of Cognitive Neuroscience
Volume 18, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
t
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Figure 8. Results for
Experiment 2 involving the
Ann faces. (A) The inversion
effect for normal subjects in
the three conditions. Error
bars represent the standard
error of the difference between
the upright and inverted
conditions. (B) A comparison
of the performance of the
normal subjects and the
prosopagnosics for the three
conditions. Error bars
represent the standard error
of the difference between the
DPs and the control group.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
f
.
t
.
In conclusion, our results suggest that specialized face
perception mechanisms extract both part and spacing in-
formation from faces only. These findings challenge the
spacing hypothesis but support holistic theories of face
perception, which suggest that face perception mecha-
nisms are special in that they interactively process facial
information as a nondecomposable whole.
METHODS
Prosopagnosic Subjects
We tested 13 prosopagnosics individuals who contacted
the Center for Prosopagnosia Research at the Harvard
o
n
1
8
M
a
y
2
0
2
1
University Web site (www.faceblind.org) because they
reported significant problems with face recognition in
daily life. None except R.S. was aware of any events that
may have caused brain damage, and all reported lifelong
problems with face recognition. R.S. reported that she
collided with a refrigerator as a young child, and al-
though she remained conscious, her mother told her
that immediately after the collision, R.S. asked, ‘‘Why are
the trees white?’’ as she looked out the window. Four of
the DPs have been reported on in other articles, and
these articles and their designation in these articles are
listed in Appendix II. Eight prosopagnosics participated
in Experiment 1. One prosopagnosic from this group
(D.D.) and five new prosopagnosics participated in
Yovel and Duchaine
589
Experiment 2. Each subject was tested with a battery of
face-processing tasks in order to evaluate their face-
processing abilities. To determine whether they did, in
fact, suffer from face recognition impairments, we tested
them with three types of tests.
Famous Faces
Two different famous face tests were used with the two
groups. Subjects in Experiment 1 were presented with
photos of 25 celebrities drawn from entertainment and
politics (Duchaine et al.,
in press; Duchaine, 2000),
whereas those in Experiment 2 were presented with
60 faces. The images were cropped so that little hair
or clothing was visible, and each image was presented
for 10 sec. Two groups of controls were used for the
test with 25 images. College-age controls were com-
pared to DPs in their twenties, and controls between
45 and 55 years of age were compared to older DPs.
The college-age controls averaged 23.6 (SD = 1.4) on
the test, and the middle-aged adult controls averaged
22.6 (SD = 1.9). Controls for the 60-face test were be-
tween 35 and 45 years of age, and they averaged 52.5
(SD = 6.6).
Cambridge Memory Test for Faces
In the test, subjects were introduced to six target faces
with study images (Duchaine & Nakayama, 2006). Each
face is shown in three different views;
immediately
after presentation of those study images, subjects were
presented with three forced-choice items, each of
which consisted of one of the study images paired with
two other faces in the same pose. After this introduc-
tory phase, subjects were tested with 54 forced-choice
items consisting of novel views of one of the target
faces along with two nontarget faces. Fifty college-age
subjects served as our controls, and they averaged 80.4%
(SD = 11.0). We also tested nine adults with a mean age
of 47 years, and their average was slightly higher than
that of the college students; thus, to be conservative, all
DPs were compared to the college-age group.
Face Old–New Recognition Memory Tests
In each old–new recognition memory test, 40 faces were
used (Duchaine & Nakayama, 2005). Ten faces were
target faces, and they were shown twice during the
study phase of the experiment. During the test phase,
each target face was shown twice along with 30 nontar-
get faces. Two versions of the test, which used separate
sets of faces, were created. Grayscale yearbook photo-
graphs of women’s faces were cropped so that very
little or no hair was visible. Graduate students in their
twenties to mid-thirties served as controls. Although
these controls are not age-matched with Edward, K.L.,
A.M., and B.K., all scored normally on a number of
parallel object recognition tests when compared to the
same control group (for Edward’s data, see Duchaine
et al., in press; for K.L.’s data, see Duchaine & Nakayama,
2005). A0 was used as the measure of discrimination,
and the control mean was 0.96 (SD = .03). An average
A0 score and RT were computed for each subject (ex-
cept J.W.) by combining performance on the two ver-
sions of the test. J.W. was only tested with one of the
tests, but we were unable to retest him. His scores
on all face tests were so poor that it is clear that he is
prosopagnosic.
Standardized scores for each prosopagnosic are pre-
sented in Table 1. The face recognition impairments of
prosopagnosics are evident from their extremely poor
scores.
Control Subjects for Experiments 1 and 2
The control group in Experiment 1 (N = 34) that is
included in this report is a subset of the control group
that was reported in Yovel and Kanwisher (2004a).
Stimulus order in Yovel and Kanwisher was counter-
balanced, and the controls used in this article were
those who, like the DPs, performed the face task before
the house task. Fifteen age-matched control subjects
(age, 18–54 years) participated in Experiment 2.
Stimuli and Apparatus
Photoshop was used to create the spacing and part sets
for the face and house stimuli. Stimuli were presented
using Superlab 1.2. Subjects viewed the stimuli from a
distance of approximately 50 cm. The stimuli subtended
4.68 of visual angle in width and 7.48 in length.
Face Stimuli
Alfred face stimuli. Two sets of four face stimuli were
generated from a picture of a male face. For the spac-
ing set, four faces,
in which eyes were either closer
to or farther apart from each other and in which the
mouth was either closer to or farther from the nose,
were constructed. For the part set, the two eyes and
the mouth were replaced in each of the four faces
by eyes and mouths of a similar shape from differ-
ent original
face photos. By piloting the part and
spacing manipulations prior to the current experiment,
we determined manipulations that produced a similar
average performance for the part and spacing tasks
and that were in the dynamic range away from ceiling
or floor effects. Figure 1 shows a face stimulus gen-
erated by the same procedure, which yielded be-
havioral findings similar to those of the face stimulus
used in the experiment (which is not presented in
the figure because we did not obtain permission to
publish it).
590
Journal of Cognitive Neuroscience
Volume 18, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
f
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Jane face stimuli. The Jane face stimuli were adapted
from the studies conducted by the Maurer laboratory
(Mondloch et al., 2002; Le Grand et al., 2001; Freire
et al., 2000). In contrast to the Alfred stimuli, the part
and spacing tasks with the Jane stimuli were not
matched for performance, and performance on the part
task was significantly higher than performance on the
spacing task.
Ann face stimuli. Three sets of faces (spacing, part-S,
and part-SC), which each included four different stimuli,
were generated. For the spacing set, four faces, in which
the eyes were either closer to or farther apart from
each other and in which the mouth was either closer
to or farther from the nose, were constructed. For
the part-SC set, the eyes and mouth were replaced
in each of the four faces by eyes and mouths that
differed in both shape and contrast/brightness infor-
mation (makeup). For the part-S set, the eyes and
mouth were replaced in each of four faces by eyes and
mouths that differed in shape, but contrast differences
were minimized. By piloting the part and spacing ma-
nipulations prior to the current experiment, we deter-
mined manipulations that produced a similar average
performance for the part-S task and the spacing task
that were in the dynamic range away from ceiling or
floor effects.
House stimuli. House stimuli were created using a
method similar to that used for the face stimuli. For
the spacing set, four houses were constructed in which
we manipulated the location of the windows and the
door (the two left-side windows and the right-side
window and door were closer together or farther
apart, or the two upper windows were closer to or
farther from the roof ). For the part set, the windows
and the door were replaced by windows and a door
of similar overall shape but of a different texture (see
Figure 1). Pilot studies determined manipulations that
produced similar performance levels on the part and
spacing tasks that were comparable to those of the
face task.
Procedure
Experiment 1
Subjects were presented with a sequential same–different
matching task. Each trial started with a 500-msec fixa-
tion dot at the center of the screen. A first stimulus
was presented for 250 msec followed by a 1000-msec
interstimulus interval during which time the fixation dot
was on the screen. The second face was on the screen
for 250 msec. The part and spacing trials were presented
in a randomized order within each block. The face
task preceded the house task for both the DPs and the
controls. Trials with Alfred and Jane were randomly
interleaved. Subjects were not informed that the stimuli
would differ in spacing or parts. Each task (upright face,
upright house) included a total of 80 stimuli: 20 pairs
of different stimuli and 20 pairs of the same stimuli for
the part and the spacing tasks.
Experiment 2
The procedure for the Ann task was similar to the
procedure used for the Alfred task except for the fol-
lowing differences: To obtain a performance level of
75% correct in the Ann task, the face stimuli were
presented for 500 msec. Pilot studies showed that when
the part-SC trials were mixed in the same block with
the part-S trials, normal subjects’ performance with
part-S and spacing tasks was too low to allow valid tests
of the inversion effect (floor effects). However, when
part-SC trials were presented in a separate block, the
proportion of correct responses for the part-S task
and the spacing task improved and allowed us to val-
idly examine the effect of inversion. It seems that
the salient difference between the faces in the part-
SC manipulation (see Figure 7) modified subjects’ dis-
crimination criterion, and we missed the more subtle
differences between the spacing and the part-S manip-
ulations relative to the case in which these stimuli
were presented in a separate block. Thus, we presented
the two part tasks in separate blocks. In each block,
the part trials were randomly mixed with the spacing
trials. Subjects were not informed about the nature
in which the face stimuli were manipulated. Note that
all controls and prosopagnosic subjects were tested
with the same procedure, so the blocking (or lack
thereof in Experiment 1) cannot account for the con-
ditions in which the prosopagnosics showed impaired
performance.
Data Analysis
Experiment 1
We computed the proportion of correct responses
and RT for the spacing and part tasks for faces and
houses. ANOVA was performed with Group (DP, con-
trols) as a between-subject factor and with Stimulus
(face, house) and Task (part, spacing) as within-subject
factors.
Experiment 2
We computed the proportion of correct responses for
the spacing task, the Jane-like part (part-SC), and the
Alfred-like part (part-S) tasks. A repeated measures
ANOVA with Orientation and Type (spacing, part-SC,
and part-S) as within-subject factors was performed to
compare the inversion effects for the three stimulus
manipulations in controls.
Yovel and Duchaine
591
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
f
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
APPENDIX I
REFERENCES
Performance of each of the DPs on the face and house
tasks, average performance, and standard deviation of a
control group (n = 34).
Houses
Alfred Face
Jane Face
Spacing Parts
Spacing Parts
Spacing Parts
A.C.
Edward
K.L.
L.A.
M.L.
R.S.
D.D.
J.H.
0.70
0.68
0.65
0.78
0.60
0.88
0.90
–
0.40
0.90
0.93
0.90
0.68
0.90
0.63
–
Average
0.74
0.76
Controls
Average
SD
0.78
0.12
0.78
0.11
APPENDIX II
0.40
0.48
0.68
0.79
0.63
0.64
0.78
0.65
0.63
0.79
0.10
0.35
0.69
0.71
0.54
0.51
0.56
0.50
0.67
0.55
0.78
0.10
0.60
0.51
0.54
0.71
0.50
0.78
0.67
0.62
0.62
0.74
0.10
1.00
0.78
0.86
0.92
0.73
0.69
0.86
0.73
0.83
0.88
0.08
Four of the DPs have been reported on in other articles.
Next to each subject’s designation in this article, we list
other articles that they have appeared in, along with
their designation in those articles (in parentheses).
J.H.: Duchaine & Nakayama, 2005, Journal of Cognitive
Neuroscience (M1)
Edward: Duchaine et al., in press, Cognitive Neuropsy-
chology (Edward); Duchaine et al., 2004, Neuron
(Edward)
K.L.: Duchaine & Nakayama, 2005, Journal of Cognitive
Neuroscience (F2)
M.L.: Duchaine & Nakayama, 2005, Journal of Cognitive
Neuroscience (M2)
Acknowledgments
We thank Daphne Maurer, Cathy Mondloch, and Richard
Le Grand for providing us with the Jane face stimuli. We are
thankful to Alison Harris and Javid Sadr for their suggestions.
Ken Nakayama and Nancy Kanwisher provided invaluable sup-
port for this project. We also thank Arielle Tambini, Kathleen
Cui, Nao Gamo, Gayle Speck, and Kerry Dingle for their help
with control data collection, and Stephanie Chow and Kathleen
Cui for help with stimulus generation. This work was sup-
ported by grants from the National Institutes of Health (F32
MH64246-02 and RO1 EY13602).
Reprint requests should be sent to Galit Yovel, Department of
Brain and Cognitive Sciences, Massachusetts Institute of Tech-
nology, 77 Massachusetts Avenue, NE20-443, Cambridge, MA
02139, or via e-mail: galit@mit.edu.
Barton, J. J., Cherkasova, M. V., Press, D. Z., Intriligator, J. M.,
& O’Connor, M. (2003). Developmental prosopagnosia: A
study of three patients. Brain and Cognition, 51, 12–30.
Behrmann, M., & Avidan, G. (2005). Congenital prosopagnosia:
Face-blind from birth. Trends in Cognitive Sciences, 9,
180–187.
Behrmann, M., Avidan, G., Marotta, J. J., & Kimchi, R.
(2005). Detailed exploration of face-related processing in
congenital prosopagnosia: 1. Behavioral findings. Journal
of Cognitive Neuroscience, 17, 1130–1149.
Bentin, S., Allison, T., Puce, A., Perez, E., & McCarthy, G.
(1996). Electrophysiological studies of face perception in
humans. Journal of Cognitive Neuroscience, 8, 551–565.
Bentin, S., Deouell, L. Y., & Soroker, N. (1999). Selective
visual streaming in face recognition: Evidence from
developmental prosopagnosia. NeuroReport: An
International Journal for the Rapid Communication
of Research in Neuroscience, 10, 823–827.
de Haan, E. (1999). A familial factor in the development
of face recognition deficits. Journal of Clinical and
Experimental Neuropsychology, 21, 312–315.
Duchaine, B., & Nakayama, K. (2005). Dissociations of face
and object recognition in developmental prosopagnosia.
Journal of Cognitive Neuroscience, 17, 249–261.
Duchaine, B., & Nakayama, K. (2006). The Cambridge Face
Memory Test: Results for neurologically intact individuals
and an investigation of its validity using inverted face
stimuli and prosopagnosic subjects. Neuropsychologia,
44, 576–585.
Duchaine, B., Yovel, G., Butterworth, E. J., & Nakayama, K.
(in press). Prosopagnosia as an impairment to face-specific
recognition mechanisms: Elimination of the testable
domain-general hypotheses. Cognitive Neuropsychology.
Duchaine, B. C. (2000). Developmental prosopagnosia
with normal configural processing. NeuroReport, 11,
79–83.
Duchaine, B. C., Dingle, K., Butterworth, E., & Nakayama,
K. (2004). Normal greeble learning in a severe case of
developmental prosopagnosia. Neuron, 43.
Eimer, M. (2000). Event-related brain potentials distinguish
processing stages involved in face perception and
recognition. Clinical Neurophysiology, 111, 694–705.
Farah, M. J. (1996). Is face recognition ‘‘special’’? Evidence
from neuropsychology. Behavioural Brain Research, 76,
181–189.
Farah, M. J., Wilson, K. D., Drain, M., & Tanaka, J. N. (1998).
What is ‘‘special’’ about face perception? Psychological
Review, 105, 482–498.
Freire, A., Lee, K., & Symons, L. A. (2000). The face-inversion
effect as a deficit in the encoding of configural information:
Direct evidence. Perception, 29, 159–170.
Gross, C. G. (1992). Representation of visual stimuli in inferior
temporal cortex. Philosophical Transactions of the Royal
Society of London, Series B, Biological Sciences, 335, 3–10.
Haig, N. D. (1984). The effect of feature displacement on
face recognition. Perception, 13, 505–512.
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). The
fusiform face area: A module in human extrastriate cortex
specialized for face perception. Journal of Neuroscience,
17, 4302–4311.
Karmiloff-Smith, A., Thomas, M., Annaz, D., Humphreys, K.,
Ewing, S., Brace, N., Duuren, M., Pike, G., Grice, S., &
Campbell, R. (2004). Exploring the Williams syndrome
face-processing debate: The importance of building
developmental trajectories. Journal of Child Psychology
and Psychiatry, 45, 1258–1274.
592
Journal of Cognitive Neuroscience
Volume 18, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
t
.
f
.
.
o
n
1
8
M
a
y
2
0
2
1
Kemp, R., McManus, C., & Pigott, T. (1990). Sensitivity to
the displacement of facial features in negative and inverted
images. Perception, 19, 531–543.
Kreiman, G., Koch, C., & Fried, I. (2001). Category-specific
visual responses of single neurons in the human medial
temporal lobe. Nature Neuroscience, 3, 946–953.
Le Grand, R., Mondloch, C. J., Maurer, D., & Brent, H. P.
(2001). Neuroperception. Early visual experience and
face processing. Nature, 410, 890.
Le Grand, R., Mondloch, C. J., Maurer, D., & Brent, H. P.
(2003). Expert face processing requires visual input to the
right hemisphere during infancy. Nature Neuroscience, 6,
1108–1112.
external features, and for fractured faces. Cognitive
Neuropsychology, 17, 201–219.
Moscovitch, M., Winocur, G., & Behrmann, M. (1997).
What is special about face recognition? Nineteen
experiments on a person with visual object agnosia
and dyslexia but normal face recognition. Journal of
Cognitive Neuroscience, 9, 555–604.
Nunn, J. A., Postma, P., & Pearson, R. (2001). Developmental
prosopagnosia: Should it be taken at face value?
Neurocase, 7, 15–27.
Perrett, D. I., Rolls, E. T., & Caan, W. (1982). Visual
neurones responsive to faces in the monkey temporal
cortex. Experimental Brain Research, 47, 329–342.
Le Grand, R., Mondloch, C. J., Maurer, D., & Brent, H. P.
Puce, A., Allison, T., & McCarthy, G. (1999).
(2004). Impairment in holistic face processing following
early visual deprivation. Psychological Science, 15,
762–768.
Leder, H., & Bruce, V. (2000). When inverted faces are
recognized: The role of configural information in
face recognition. Quarterly Journal of Experimental
Psychology, A, Human Experimental Psychology, 53,
513–536.
Levine, D. N., & Calvanio, R. (1989). Prosopagnosia: A defect
in visual configural processing. Brain and Cognition, 10,
149–170.
Malcolm, G., Leung, C., & Barton, J. J. S. (2005). Regional
variations in the inversion effect for faces: Differential
effects for feature shape, spatial relations, and external
contour. Perception, 33, 1221–1231.
Maurer, D., Grand, R. L., & Mondloch, C. J. (2002). The
many faces of configural processing. Trends in Cognitive
Sciences, 6, 255–260.
McCarthy, G., Puce, A., Belger, A., & Allison, T. (1999).
Electrophysiological studies of human face perception:
II. Response properties of face-specific potentials
generated in occipitotemporal cortex. Cerebral Cortex,
9, 431–444.
McCarthy, G., Puce, A., Gore, J. C., & Allison, T. (1997).
Face-specific processing in the human fusiform gyrus.
Journal of Cognitive Neuroscience, 9, 605–610.
Mondloch, C. J., Le Grand, R., & Maurer, D. (2002).
Configural face processing develops more slowly than
featural face processing. Perception, 31, 553–566.
Moscovitch, M., & Moscovitch, D. A. (2000). Super
face-inversion effects for isolated internal or
Electrophysiological studies of human face perception: III.
Effects of top–down processing on face-specific potentials.
Cerebral Cortex, 9, 445–458.
Riesenhuber, M., Jarudi, I., Gilad, S., & Sinha, P. (2004).
Face processing in humans is compatible with a simple
shape-based model of vision. Proceedings of the Royal
Society of London, B, Biological Sciences, 271, S448–S450.
Sergent, J., & Signoret, J.-L. (1992). Varieties of functional
deficits in prosopagnosia. Cerebral Cortex, 2, 375–388.
Tanaka, J. W., & Farah, M. J. (1993). Parts and wholes in
face recognition. Quarterly Journal of Experimental
Psychology, A, Human Experimental Psychology, 46A,
225–245.
Tanaka, J. W., & Farah, M. (2003). The holistic representation
of faces. In M. A. Peterson & G. Rhodes (Eds.), Perception
of faces, objects and scenes: Analytic and holistic processes.
New York: Oxford University Press.
Tanaka, J. W., & Sengco, J. A. (1997). Features and their
configuration in face recognition. Memory & Cognition,
25, 583–592.
Yin, R. (1969). Looking at upside down faces. Journal of
Experimental Psychology, 81, 141–145.
Young, A. W., Hellawell, D., & Hay, D. C. (1987).
Configurational information in face perception. Perception,
16, 747–759.
Yovel, G., & Kanwisher, N. (2004a). Face perception:
Domain specific, not process specific. Neuron, 44, 1–20.
Yovel, G., & Kanwisher, N. (2004b). Face perception engages
a domain-specific system for processing both configural
and part-based information about faces [Abstract]. Journal
of Vision, 4, 133a.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
–
r
p
t
d
i
c
1
l
8
e
4
–
5
p
8
d
0
f
/
1
1
9
8
3
5
/
7
4
8
/
1
5
8
o
0
c
/
n
1
2
7
0
5
0
6
6
1
1
2
8
7
/
4
j
5
o
8
c
0
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
4
o
.
n
5
0
8
8
0
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
t
.
.
f
.
.
o
n
1
8
M
a
y
2
0
2
1
Yovel and Duchaine
593