Spatial and Feature-selective Attention Have Distinct,
Interacting Effects on Population-level Tuning
Erin Goddard1,2
, Thomas A. Carlson2,3, and Alexandra Woolgar2,4
Abstract
■ Attention can be deployed in different ways: When searching
for a taxi in New York City, we can decide where to attend (e.g., to
the street) and what to attend to (e.g., yellow cars). Although we
use the same word to describe both processes, nonhuman pri-
mate data suggest that these produce distinct effects on neural
tuning. This has been challenging to assess in humans, but here
we used an opportunity afforded by multivariate decoding of
MEG data. We found that attending to an object at a particular
location and attending to a particular object feature produced
effects that interacted multiplicatively. The two types of attention
induced distinct patterns of enhancement in occipital cortex,
with feature-selective attention producing relatively more
enhancement of small feature differences and spatial attention
producing relatively larger effects for larger feature differences.
An information flow analysis further showed that stimulus repre-
sentations in occipital cortex were Granger-caused by coding in
frontal cortices earlier in time and that the timing of this feed-
back matched the onset of attention effects. The data suggest
that spatial and feature-selective attention rely on distinct neural
mechanisms that arise from frontal-occipital information
exchange, interacting multiplicatively to selectively enhance
task-relevant information. ■
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
At any moment, there is far more information available
from our senses than we can process at once. Accordingly,
only a subset of the available information is processed to a
high level, making it crucial that the brain dynamically
redistributes processing resources—selectively attends—
to the most relevant information. We can deploy our atten-
tional resources in different ways. For example, we can
decide where to attend (e.g., to the item on the left, rather
than the one on the right), and we can control what feature
of that item we attend to (e.g., its color, rather than its
shape). This allows us to dramatically reduce the compu-
tational requirements of our neural system and focus on
the information that is most pertinent to our current task.
Each of these types of attention can change behavior,
improving performance related to the attended location
or stimulus feature (Carrasco, 2011; Pestilli & Carrasco,
2005; Saenz, Buracas, & Boynton, 2003; Rossi & Paradiso,
1995). Shifts in attention also induce neural changes
consistent with greater neural resources being directed
to representing attended over unattended information.
These changes include shifts in the responses of individual
neurons (Maunsell, 2015; Sprague, Saproo, & Serences,
2015; Reynolds & Heeger, 2009), changes in the overall
responsiveness of brain regions (Gouws et al., 2014;
Lennert, Cipriani, Jolicoeur, Cheyne, & Martinez-Trujillo,
2011; Serences & Boynton, 2007; Saenz et al., 2003; Saenz,
1University of New South Wales, 2Macquarie University, Sydney,
New South Wales, Australia, 3University of Sydney, 4University
of Cambridge
Buracas, & Boynton, 2002; Chawla, Rees, & Friston, 1999;
Corbetta, Miezin, Dobmeyer, Shulman, & Petersen, 1990),
and changes in the information carried by a population
response ( Vaziri-Pashkam & Xu, 2017; Guggenmos et al.,
2015; Woolgar, Williams, & Rich, 2015).
However, neuroimaging and electrophysiological stud-
ies have often focused on attention directed in space, or to
a particular feature, in separate studies, making it difficult
to predict how the different types of attention interact.
Characterizing how different attentional mechanisms
interact is essential, however, because they rarely operate
in isolation. So far, results are mixed. Attending to a partic-
ular feature (e.g., red) changes baseline activity in the
absence of visual stimuli (Chawla et al., 1999; Corbetta
et al., 1990) and changes single-unit (McAdams &
Maunsell, 2000) and neuronal population (Bartsch,
Donohue, Strumpf, Schoenfeld, & Hopf, 2018; Cohen &
Maunsell, 2011; Jehee, Brady, & Tong, 2011) responses
across the visual field. The spatially diffuse effects of
feature-based attention are also supported by results from
visual search, where feature-based attention facilitates the
parallel identification of items matching a target feature
value (e.g., Peelen & Kastner, 2014; Bichot, Rossi, &
Desimone, 2005). Conversely, attending to an object at a
particular location can boost processing of all its features
(Haazebroek, Raffone, & Hommel, 2017; O’Craven,
Downing, & Kanwisher, 1999; Duncan, Humphreys, &
Ward, 1997; Duncan, 1984). This is consistent with results
showing these two subtypes of attention have largely
independent, additive effects (Hayden & Gallant, 2009;
Patzwahl & Treue, 2009; Treue & Martinez-Trujillo,
© 2021 the Massachusetts Institute of Technology. Published under
a Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Journal of Cognitive Neuroscience 34:2, pp. 290–312
https://doi.org/10.1162/jocn_a_01796
1999). Alternatively, others have reported more complex
interactions, where the attended feature of an attended
object was selectively enhanced whereas other informa-
tion at that location was not (e.g., Jehee et al., 2011, and
small multiplicative interaction in Hayden & Gallant,
2009).
The lack of human neuroimaging studies manipulating
both types of attention also limits our ability to directly
compare the effects of each. In nonhuman primates, sub-
stantial work suggests distinct effects of different atten-
tional strategies on the tuning of individual neurons
(e.g., reviewed by Reynolds & Heeger, 2009). However,
stimulus information carried by large-scale neuronal pop-
ulations cannot be trivially predicted from tuning curve
properties alone, because factors such as response vari-
ance and correlation can significantly impact population
information coding (Sprague et al., 2015; Moreno-Bote
et al., 2014). Suggestive data from recent MEG/EEG stud-
ies indicate that these effects on tuning curves can be used
to account for population-level responses in humans for
feature-based (Bartsch et al., 2017; Störmer & Alvarez,
2014) and spatial (Foster, Thyer, Wennberg, & Awh,
2021) attention, but a direct contrast is needed to compare
the population-level changes induced by each type of
attention.
A final question concerns the top–down mechanisms
supporting selection of information. The pFC (among
other regions) is strongly implicated as a source of atten-
tional control (Duncan, 2013; Miller & Cohen, 2001;
Dehaene, Kerszberg, & Changeux, 1998), but the content
of top–down signals is unclear. They may contain task-
level information, including maps of attentional priority
(Moore, Armstrong, & Fallah, 2003; Wolfe, 1994; Koch &
Ullman, 1985), and the participant’s criterion (Luo &
Maunsell, 2018). Alternatively, because frontoparietal
control regions prioritize coding of task-relevant stimulus
information ( Jackson & Woolgar, 2018; Jackson, Rich,
Williams, & Woolgar, 2017; Freedman & Assad, 2016;
Woolgar et al., 2015; Freedman, Riesenhuber, Poggio, &
Miller, 2001), they may bias processing elsewhere
(Duncan, 2006; Desimone & Duncan, 1995) by transfer-
ring a representation of the attended information itself
(see also Bichot, Heard, DeGennaro, & Desimone, 2015;
Liu, Hospadaruk, Zhu, & Gardner, 2011).
Here, we used multivariate decoding of human MEG
data, together with information flow analysis, to address
these questions. We found strong multiplicative effects
of where participants attended (left or right) and what
they attended to (shape or color) on stimulus-related
information coding in both frontal and occipital cortices.
We further showed that the two attentional subtypes
induced different effects on the population response, with
distinct patterns of enhancement across feature differ-
ences of increasing magnitude. Finally, our information
flow analysis tracked the exchange of information between
frontal and occipital cortices over time. We found that the
dominant direction of information flow was initially
feedforward (from visual to frontal cortex) but later
reversed and that the onset of this reversal corresponded
to the strongest attentional modulation in the occipital
lobe. We conclude that these two subtypes of attention
have demonstrably different effects on population-level
tuning in the human brain and interact multiplicatively
to bring about selective focus on task-relevant informa-
tion. The effects appear to be mediated by frontal-to-
occipital exchange of item-specific, task-relevant stimulus
information.
METHODS
Participants
Participants’ (n = 20; 14 women, 6 men) ages ranged from
18 to 32 years (mean = 22.4 years), and they were each
paid $50 as compensation for their time. All were right-
handed, had normal or corrected-to-normal vision, had
no history of neurological or psychiatric disorder, and
were naïve to the purposes of the study. Participant
recruitment and the experiment were conducted with
the approval of the Macquarie University Human Research
Ethics Committee.
Visual Stimuli
Visual stimuli were generated and presented using
MATLAB ( Version R2014b) and routines from Psychtool-
box (Kleiner, Brainard, & Pelli, 2007; Brainard, 1997; Pelli,
1997). We created novel object stimuli that varied in color
and in their shape statistics (see Figure 1B) using custom
code. The shapes were variants of “spikie” stimuli used in
previous work ( Jackson et al., 2017; Woolgar et al., 2015;
Op de Beeck, Baker, DiCarlo, & Kanwisher, 2006), pre-
sented on a black background. We varied the spike orien-
tation statistics to create four classes of “spikie” objects:
strongly or weakly “X-shaped,” and strongly or weakly
“non-X-shaped” (Figure 1B). In the shape-based task,
participants categorized the target as “X-shaped” or
“non-X-shaped.” We created 100 unique versions of each
shape class by adding random variation in the spike loca-
tions, lengths, and orientations to ensure that no single
feature was diagnostic of category and to encourage
attention to the object’s overall shape.
In color, there were also four classes (strongly or weakly
red, and strongly or weakly green; Figure 1B), which par-
ticipants categorized as either “reddish” or “greenish” in
the color task. Each object had a maximum luminance of
0 and xy chromaticity coordi-
0
108.1 cd/m2 and constant u
v
nates ( Wyszecki & Stiles, 1982), which were as follows:
0:
0
strongly red u
v
0: 0.23, 0.55
0.27, 0.54 (xy: 0.50, 0.44); weakly green u
0: 0.16, 0.56 (xy:
(xy: 0.45, 0.48); and strongly green u
0.36, 0.57).
0: 0.35, 0.53 (xy: 0.56, 0.38); weakly red u
v
v
v
0
0
0
During MEG sessions, stimuli were projected through a
customized window by an InFocus IN5108 LCD back-
Goddard, Carlson, and Woolgar
291
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
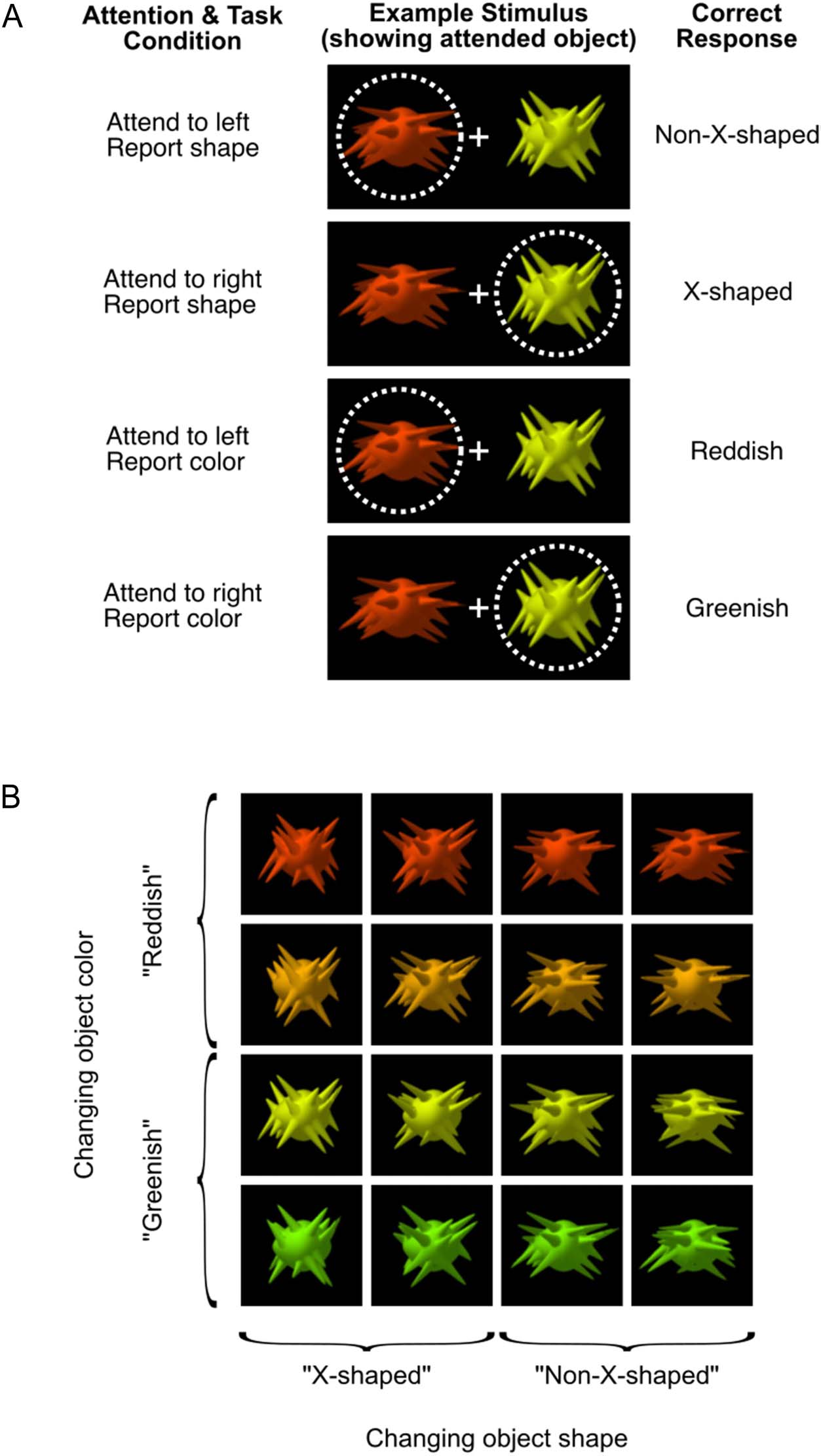
Figure 1. Visual stimuli
showing task conditions (A) and
stimulus dimensions (B).
(A) Task conditions. At the
start of each block of trials,
participants were told the
location to which they should
direct their attention (left or
right of fixation) and whether
they should report the target
object’s shape (“X-shaped” or
“non-X-shaped”) or color
(reddish or greenish). Two
objects appeared on each trial,
and participants covertly
attended to one while we
used eye tracking to monitor
their fixation. The example
illustrates how the same
stimulus configuration was
used in each of the four task
conditions. The dotted circle
indicates the location of spatial
attention and was not visible
during the experiment.
(B) Stimulus dimensions. Each
object varies systematically
along two dimensions, color
and shape. Participants
categorized the attended object
as either “greenish” or “reddish”
(when reporting color) or as
“X-shaped” or “non-X-shaped”
(when reporting shape). On
each trial, the objects were
randomly selected from 100
exemplars with the same shape
statistics but random variation
in the location, length, and
orientation of the spikes. This
variability is illustrated in the
shape variation between objects
in the same column.
projection system located outside the Faraday shield, onto
a screen located above the participant. Participants, lying
supine, viewed the screen from 113 cm. Individual “spikie”
objects each had a central body of 195 pixels (5.8 degrees
visual angle [dva]) wide × 175 pixels (5.2 dva) high. Their
total size varied with their spikes, but the spikes never
reached the border of the object image (403 × 403
pixels). Each trial’s stimulus included two “spikie” object
images side by side (total size 24 × 12 dva), with a central
white fixation cross (1 × 1 dva; Figure 1A). The display
system was characterized in situ using a Konica Minolta
CS-100A spectrophotometer and calibrated as described
previously (Goddard, Mannion, McDonald, Solomon, &
Clifford, 2010).
MEG Acquisition and Eye Tracking
MEG data were collected with a whole-head MEG system
(Model PQ1160R-N2, KIT) consisting of 160 coaxial first-
order gradiometers with a 50-mm baseline (Uehara
292
Journal of Cognitive Neuroscience
Volume 34, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
et al., 2003; Kado et al., 1999). Before MEG measurements,
five marker coils were placed on the participant’s head.
Marker positions, nasion, left and right pre-auricular
points, and the participant’s head shape were recorded
with a pen digitizer (Polhemus Fastrack), using a mini-
mum of 2000 points. Each participant’s MEG data were
collected in a single session of approximately 90 min, at
a sampling frequency of 1000 Hz. On each trial, partici-
pants responded using a fiber optic response pad (fORP,
Current Designs). We tracked participant’s eye move-
ments using an EyeLink 1000 MEG-compatible remote
eye-tracking system (SR Research, 500-Hz monocular
sampling rate). Before scanning, we tested participants
for their dominant eye (usually right) and focused the
eye tracker on this eye.
Experimental Protocol
Participants were instructed to covertly attend to the
stimulus on either the left or right of fixation (“spatial
attention” manipulation), and they were required to make
a judgment based on the target object’s color or shape
(“feature-selective attention” manipulation; Chen,
Hoffmann, Albright, & Thiele, 2012). Each participant’s
MEG session was divided into eight blocks, where the
attended location and the attended feature were constant
within each block. The combination of two attended loca-
tions with two attended features yielded four different
tasks (Figure 1A). Before the experiment, each participant
was familiarized with the object shape categories
(“X-shaped” and “non-X-shaped”) and color categories
(“reddish” and “greenish”) and completed a training ses-
sion on a laptop outside the MEG scanner where they
practiced each task.
Each trial’s stimulus included two objects, one each on
the left and right of fixation, presented simultaneously.
Both spatial attention (Sundberg, Mitchell, & Reynolds,
2009; Reynolds, Chelazzi, & Desimone, 1999) and
feature-selective attention (Saenz et al., 2003) effects are
stronger when attended and unattended stimuli simulta-
neously compete for access to perceptual processing.
Within each block, every pairing of the 16 objects in
Figure 1B was included once, giving 256 (16 × 16) trials.
These 256 trials were presented in a counterbalanced
order within each block, so that objects of each shape
and color were equally likely to precede objects of all
shapes and colors. A different counterbalanced order
was used for each block, and to this sequence of 256 trials,
the last trial was added to the beginning, and the first trial
was added to the end, giving a total of 258 trials in each
block. Data from these first and last trials were discarded.
The reported feature alternated between shape and
color on every block, and the attended location alternated
after the second, fourth, and sixth blocks. Starting location
and feature were counterbalanced across participants.
Within each pair of blocks where the attention condition
was the same (e.g., Blocks 1 and 5), the buttons
corresponding to the two response options were
switched, so that response mappings were counter-
balanced across blocks. Every block commenced with an
instruction including where to attend, what feature to
report, and the response mapping for that block. Before
the first trial, participants were required to identify the
response buttons correctly with a key press and to
repeat the eye tracker’s 5-point calibration.
Every trial began with the eye tracker’s verification of fix-
ation. Participants had to fixate within 1 dva of the fixation
marker for ≥300 msec to trigger stimulus onset. During
the stimulus (maximum = 150 msec) a 50 × 50 pixel white
square was displayed in the bottom right (outside the
stimulus region), aligned with a photodetector, attached
to the mirror, whose signal was recorded with that of
the gradiometers, enabling accurate alignment MEG
recordings with stimulus timing. When eye-tracking regis-
tered participants were no longer fixating during the
150-msec stimulus presentation, the stimulus terminated
early. Eye tracker variability (e.g., eye tracker missing
frames) resulted in an unexpectedly high number of
shorter trials: The median stimulus duration was 92 msec,
and the first and third quartiles were 64 and 126 msec,
respectively. Because this affected a majority of trials, we
included all trials in our analysis, but ran an extra analysis to
check that variability in stimulus duration did not account
for our results (see below). After stimulus offset, the fixa-
tion marker remained white until participants responded
with a button press. After the response, but no sooner
than 1000 msec from the stimulus onset, the fixation
marker changed for 200 msec to provide feedback: dim-
ming to gray for “correct” or turning blue for “incorrect.”
After feedback, there was a variable intertrial interval (300–
800 msec), which comprised the fixation check for the
subsequent trial. We used a variable intertrial interval to
avoid expectancy effects.
MEG Data Analysis: Source Reconstruction
Forward modeling and source reconstruction were per-
formed using Brainstorm (Tadel, Baillet, Mosher, Pantazis,
& Leahy, 2011; neuroimage.usc.edu/brainstorm). We cre-
ated a model of each participant’s brain by manually align-
ing the ICBM152 template adult brain (Fonov et al., 2011)
to their head shape using nasion, pre-auricular points, and
head shape data. Once aligned, we applied nonlinear
warping to deform the template brain to the participant’s
head shape, which provides a superior model to an
unwarped canonical template (Henson, Mattout, Phillips,
& Friston, 2009). We generated a forward model for each
model by applying a multiple spheres model (Huang,
Mosher, & Leahy, 1999) to the individually warped tem-
plate brain and their measured head location.
Functional data were preprocessed in Brainstorm with
notch filtering (50, 100, and 150 Hz) to remove the influ-
ence of 50 Hz line noise and its harmonics, followed by
bandpass filtering (0.2–200 Hz). Cardiac and eye blink
Goddard, Carlson, and Woolgar
293
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
artifacts were removed using signal space projection:
Cardiac and eye blinks events were identified using default
filters in Brainstorm, manually verified, and then used to
estimate a small number of basis functions corresponding
to these noise components, which were removed from
the recordings (Uusitalo & Ilmoniemi, 1997). Across par-
ticipants, less than 1% (0.73%) of trials contained any eye
blink during the stimulus presentation, as detected from
the MEG signals, and there was no evidence that they were
unevenly distributed across trial types. We retained these
trials to maintain the counterbalanced design for our clas-
sification analyses. From these functional data, we
extracted two epochs for each trial: a measure of baseline
activity (−100 to −1 msec relative to stimulus onset) and
the evoked response (0–2000 msec). We estimated the
noise covariance for each run from the baseline measures,
regularized using the median eigenvalue, and then applied
a minimum norm source reconstruction to the evoked
data. For each source reconstruction, we used a 15,000
vertex cortical surface (standard for the ICBM152
template, with atlas information), dipole orientations
constrained to be orthogonal to the cortical surface. We
visually inspected the quality of the source reconstruction:
The average trial data included an initial ERP at the occip-
ital pole and subsequent ERPs at sources within the occip-
ital cortex but lateral and anterior to the occipital pole,
consistent with extrastriate areas along the ventral visual
pathway.
MEG Data Analysis: Preprocessing and Data
Set Definitions
For classification analyses, we generated three data sets:
The first included preprocessed data from all sensors,
without source reconstruction; the second included
sources in occipital, occipito-temporal, and inferior-
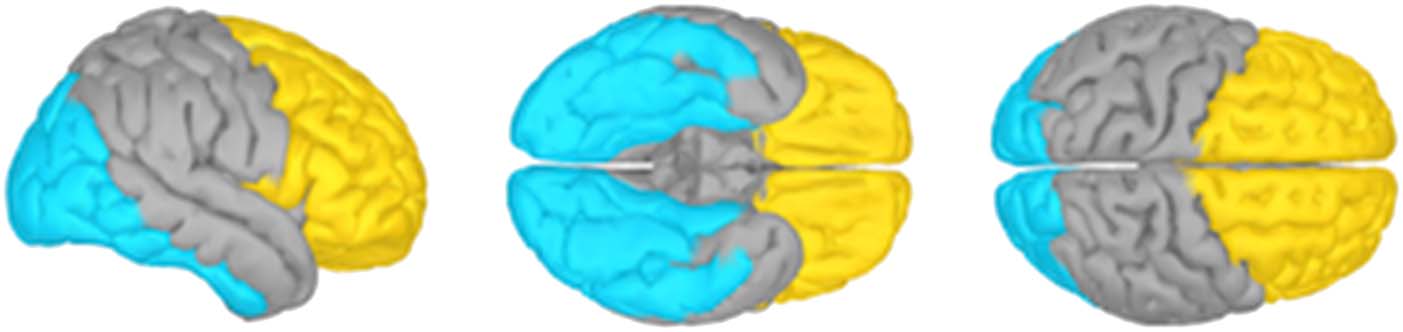
temporal cortices (“occipital” ROI, 3302 vertices) in the
atlas for the ICBM152 template; and the third included
frontal and prefrontal cortices (“frontal” ROI, 3733 verti-
ces), as shown in Figure 2A.
For each data set, we extracted data from −100 to
2000 msec relative to the stimulus onset of each trial.
We reduced each data set, comprising 2100 msec of data
for each of 2048 trials and up to 160 sensors or up to 3733
sources using PCA. We retained data from the first n com-
ponents, which accounted for 99.99% of variance (mean =
85.3, SD = 6.9 for frontal ROI; mean = 76.6, SD = 5.8 for
occipital ROI; and mean = 157.2, SD = 1.1 for whole brain
Figure 2. ROIs. The “occipital”
(cyan) and “frontal” (yellow)
ROIs shown on the partially
inflated cortical surface of the
ICBM152 template brain.
sensor data) and down-sampled to 200 Hz using the
MATLAB decimate function.
MEG Data Analysis: Classifier Analyses
We used classification analyses to measure the extent to
which brain activity could predict task condition and the
color and shape of the stimuli on each trial. For every clas-
sification, we repeated the analysis for each time sample
to capture how the information carried by the neural
response changed over time: We trained classifiers to dis-
criminate between two categories of trial and tested on
held-out data. We report results obtained with a linear sup-
port vector machine classifier, using the MATLAB function
fitcsvm with KernelFunction set to linear. We also
repeated our analyses with a linear discriminant analysis
using the MATLAB function classify with type of diagLinear
and obtained very similar results (not shown).
For each classification, we created “pseudotrials” by
averaging across trials with the same value on the dimen-
sion of interest, but with differing values along other
dimensions. We used pseudotrials to increase signal-to-
noise ratio along the dimension of interest (e.g., see
Guggenmos, Sterzer, & Cichy, 2018; Grootswagers,
Wardle, & Carlson, 2017). When training classifiers to dis-
criminate object color and shape, we trained and tested
within a single-task condition (e.g., attend left, report
color), comprising two blocks (512 trials). We trained clas-
sifiers separately on each pair of the four levels along each
feature dimension, at each object location, using pseudo-
trials to balance across irrelevant dimensions. For exam-
ple, when classifying “strongly green” versus “weakly
green” objects on the left of fixation, there were 128
“strongly green” and 128 “weakly green” trials. For classi-
fying left object color, we defined pseudotrials that were
balanced across left object shape and right object color
and shape (four levels each). Because balancing across
all three of these irrelevant dimensions would require
4 × 4 × 4 = 64 trials per pseudotrial, yielding only two
pseudotrials per category, we instead balanced across
two of three irrelevant dimensions, using 4 × 4 = 16 trials
per pseudotrial, and randomized across the third (allow-
ing eight pseudotrials per category). For each pair of irrel-
evant feature dimensions, we generated 100 sets of the
pseudotrials, each with a different randomization. Repeat-
ing this process 3 times, balancing across different pairs of
irrelevant features, gave us 300 sets of pseudotrials in
total. For each of set of pseudotrials, we trained a classifier
294
Journal of Cognitive Neuroscience
Volume 34, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
using seven of the eight pseudotrials in each condition
and tested using the remaining pair of trials, repeating
8 times, averaging classifier performance across these.
For each feature dimension (color and shape), the four
feature values gave six pairwise classifications, which we
grouped according to the feature difference between the
pair. When considering the effects of spatial and feature-
selective attention across feature difference, we grouped
classification pairs according to whether they were one
(three pairs), two (two pairs), or three (one pair) steps
apart along their feature dimension and averaged across
classifications within each group.
To summarize the effects of spatial attention (SpatAtt)
and feature-selective attention (FeatAtt), we used the fol-
0) in the
lowing metrics, based on classifier performance (d
attended location, attended feature (aLaF) condition; the
attended location, unattended feature (aLuF ) condition;
the unattended location, attended feature (uLaF ) condi-
tion; and the unattended location, unattended feature
(uLuF ) condition.
SpatAtt ¼ aLaF þ aLuF − uLaF − uLuF
FeatAtt ¼ aLaF þ uLaF − aLuF − uLuF
(1)
(2)
Statistical Testing
To generate a null distribution of chance classifier perfor-
mance for statistical testing, we repeated analyses within
each participant’s data using randomly permuted trial
labels (10 permutations of the data from every fourth time
sample, a total of 1060 samples of permuted data per
participant). For each of the 1060 permuted data sets,
we then averaged classification accuracy across partici-
pants to generate a group-level null distribution of 1060
values. We visualized the null distribution over time and
found no indication that permuted data from different
time samples varied in chance performance, so we col-
lapsed these across time in statistical testing, rather than
repeating for each time sample, to reduce the time taken
for these intensive computations. Across classifications,
0 = 0.000 to a
average chance performance varied from d
0 = 0.015. We used this group-level null
maximum of d
distribution of 1060 values to calculate one-sided nonpara-
metric p values for the probability that observed group
mean classifier performance occurred by chance and to
generate a null distribution of 1060 F statistics against
which to compare the observed F statistics from
repeated-measures ANOVAs: the main effects of Attended
Location and Attended Feature on classifier performance,
and the interaction between these main effects: F(1, 19) in
each case. Similarly, we also calculated SpatAtt and FeatAtt
using the classifier performance for data with permuted
trial labels and used these to generate null distributions
of F statistics for the interaction between feature
difference and attention type. When using repeated-
measures ANOVAs to test for main and interaction effects
(Figures 4 and 5), we report nonparametric p values based
on the proportion of F statistics in the null distribution
that exceeded the observed value. In each case, we cor-
rected these p values for multiple comparisons across
time samples using a false discovery rate (FDR) correction
(Benjamini & Hochberg, 1995).
Control Analysis: Effect of Variable Stimulus Durations
Stimuli were presented for variable durations across trials,
because trials were terminated when the participant broke
fixation or the eye tracker dropped a frame. To check that
the extent to which variability could potentially drive the
classification results reported, we repeated each classifica-
tion analysis above using the stimulus state (on or off ) for
each time sample of each trial, instead of the neural data.
For each participant, we constructed an alternate data set,
where each trial’s data were a single, binary dimension (0
and 1 according to whether the stimulus was on or off at
each time sample). Using this alternate data, we ran all
classifications following identical procedures, including
pseudotrials, following a “same analysis approach” (Görgen,
Hebart, Allefeld, & Haynes, 2018).
Modeling the Effects of Spatial and Feature-
selective Attention on Population Representations
of Shape and Color
We examined whether a normalization model of the
effects of attention at the cellular level could capture the
differences we observed across subtypes in how attention
affected stimulus-related information in the population
response. In nonhuman primates, spatial attention’s effect
on the tuning of individual neurons has been character-
ized as multiplicative response gain (Lee & Maunsell,
2010; McAdams & Maunsell, 1999; Treue & Martinez-
Trujillo, 1999), contrast gain (Martinez-Trujillo & Treue,
2002; Reynolds, Pasternak, & Desimone, 2000), or both
( Williford & Maunsell, 2006). The effects of spatial atten-
tion on contrast response functions measured with fMRI
are also mixed (Li, Lu, Tjan, Dosher, & Chu, 2008; Buracas
& Boynton, 2007). A recent EEG study reported evidence
that covert spatial attention induces spatially selective
response gain in the population response (Foster et al.,
2021). In contrast, feature-based attention produces
single-unit effects, which should produce a “sharpening”
of the population response around the attended feature
value (Martinez-Trujillo & Treue, 2004), as was recently
reported with MEG (Bartsch et al., 2017). Intuitively, we
expected that these effects might be consistent with the
different patterns of enhancement we observed in the
present data. To formalize this intuition and to test
whether these single-unit effects could manifest in the
patterns of difference we observed, we implemented the
Reynolds and Heeger (2009) normalization model of
attention to generate predictions for our design, as illus-
trated in Figure 3.
Goddard, Carlson, and Woolgar
295
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
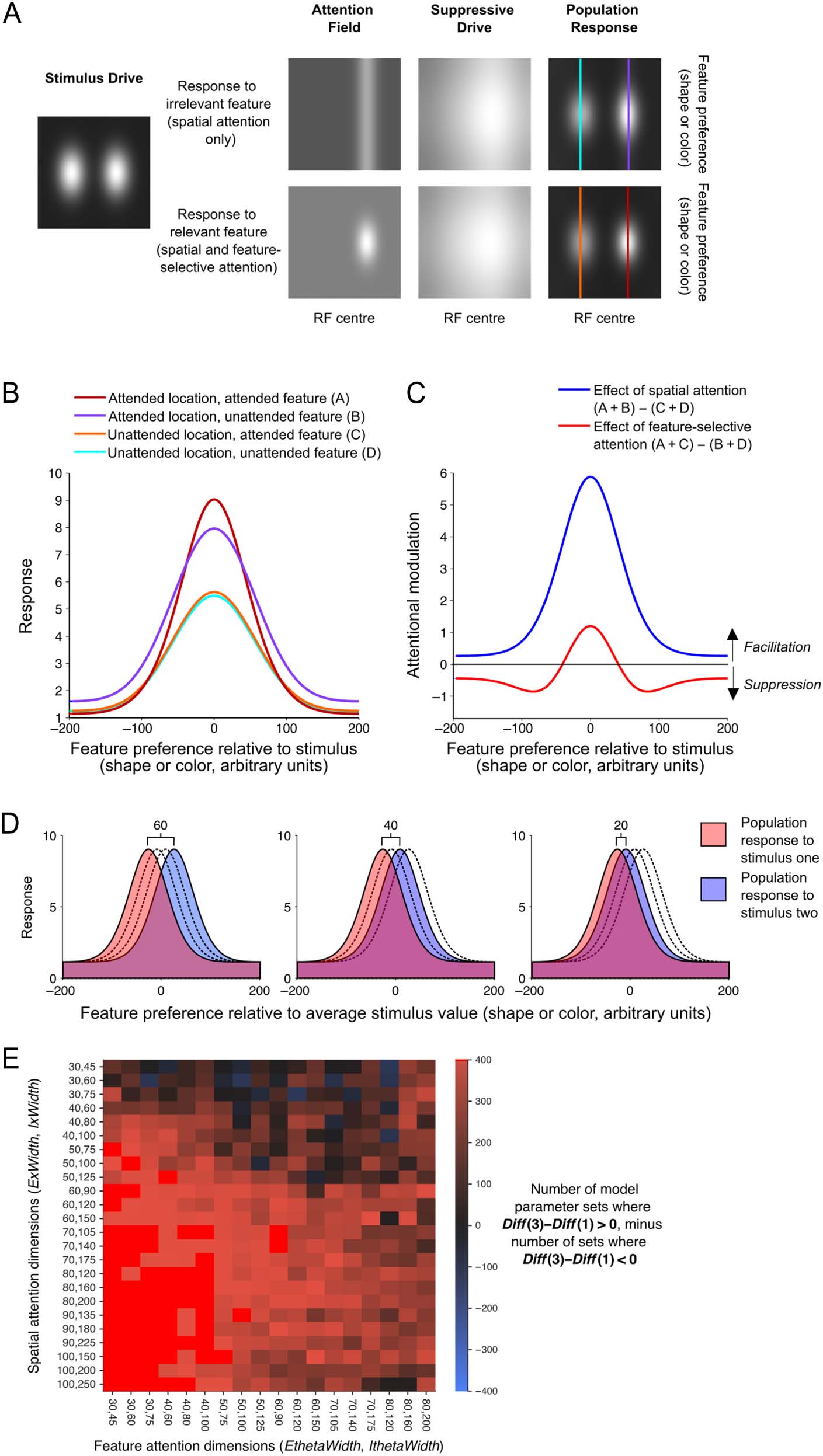
Figure 3. Normalization model
of attention. (A) Illustration of
each of the model elements
from Reynolds and Heeger
(2009, Figure 1), for a set of
example model parameters,
where each grayscale image
depicts a matrix of values
varying along a spatial
dimension (horizontally) and a
feature dimension (vertically).
For each set of model
parameters, we generated a
single “stimulus drive” and two
versions of the “attention field,”
which lead to subtly different
“suppressive drives” and
“population responses.” From
these two population
responses, we derived curves
predicting the population
response as a function of each
neuron’s preferred feature
value for each of the four
attention conditions (the
columns of the matrix indicated
with different colored vertical
lines in A). These population
responses are replotted as line
plots in B. In (C), the predicted
effects of spatial and feature-
based attention on the
population response are
summarized as the difference
between relevant population
curves from B. (D) We
predicted classifier performance
in each attention condition by
centering the population
response from B on four
different stimulus feature
values and predicting
classifier performance when
discriminating between
population responses to stimuli
of that were 60, 40, or 20
(arbitrary) units apart along the
feature dimension to simulate
the population response to
stimuli that were three, two,
or one step apart in either
color or shape. We predicted
classifier performance (d
the separation of the two
population responses, in a
manner analogous to that
used in signal detection theory.
(E) The model predictions
across four model parameters:
the excitation and inhibition
width of the spatial and feature-
based attention fields (ExWidth,
IxWidth, EthetaWidth, and IthetaWidth in Table 1). In each cell, there were 400 sets of model parameters (where other model parameters were
varied). For each set of model parameters, we calculated the difference between attention effects (Diff = SpatAtt − FeatAtt) across feature difference
(as in Figure 4). Here, we show a number of model parameter sets for which the pattern of results was qualitatively similar to the average model
prediction (Figure 4B) and to the data (e.g., Figure 4E). That is, model sets where Diff at three steps (Diff(3)) minus Diff at one step difference
(Diff(1)) was positive (red cells, 95% of cases). There were also some combinations of excitation and inhibition widths for which all 400 cases
followed this pattern (bright red cells, 16% of cases).
0) using
296
Journal of Cognitive Neuroscience
Volume 34, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
We started with the MATLAB routines from Reynolds
and Heeger (2009) available from www.cns.nyu.edu
/heegerlab/. Because we did not have strong a priori pre-
dictions for many of the model parameters, we tested a
broad range of plausible model parameters (see Table 1).
For each set of model parameters (172,800 sets in total),
we used the model to predict the response of the neural
population as a function of stimulus feature preference
(along the shape or color dimension), for each of four
cases, illustrated by lines of different colors in Figure 3A,
B. In every case, the stimulus was a single feature value (a
specific color or shape) at two fixed locations (left and
right of fixation). In two cases, we simulated attention to
one location in the absence of any feature-based attention
(simulating attention to the orthogonal feature dimen-
sion). In the other two cases, we simulated attention to
one location and attention to the feature value of the stim-
uli. From these, we predicted the population response at
attended and unattended locations, in the presence and
absence of feature-based attention. As illustrated in
Figure 3C, according to the model, spatial attention tends
to boost the population response as a multiplicative scal-
ing of the original response, whereas feature-based atten-
tion produces both facilitation and suppression of the
response, which leads to sharpening of the population
response around the attended value. Note that in the
model the spatial and feature dimensions are affected
by attention in equivalent ways, with within-dimension
attention leading to a sharpening of the population
response along the attended dimension. In this way, spa-
tial attention can also lead to a sharpening of the popula-
tion response along the spatial dimension, but only
feature-based attention leads to sharpening along the fea-
ture dimension.
One difference between the model (Reynolds &
Heeger, 2009) and our experiment is that the model is
designed to capture feature-based attention (attending
to a specific feature value, e.g., red), whereas we manipu-
lated feature-selective attention (attending to a feature
dimension, e.g., color). Although feature-based attention
has received greater attention in the electrophysiology lit-
erature, feature-selective attention has been demon-
strated to have similar effects at the level of single neurons
(Cohen & Maunsell, 2011) and to produce changes in
human EEG ( Verghese, Kim, & Wade, 2012) and fMRI
(Scolari, Byers, & Serences, 2012) responses that are sim-
ilar to the effects of feature-based attention. Furthermore,
although in human studies feature-based attention usually
refers to attention to a particular feature value that is
known before stimulus onset (e.g., visual search for red
objects), in the electrophysiological literature, there are
seminal works (e.g., Martinez-Trujillo & Treue, 2004) that
explored the effects of feature-based attention by manip-
ulating the feature value of an attended stimulus and
testing the effects on responses to unattended stimuli of
the same or different feature value. In our results (below),
the effects of feature-selective attention emerged after the
initial stimulus-induced response, making it more likely
that the observed effects were occurring after participants
had engaged their attention with the specific feature value
of the stimulus. In these ways, we felt it appropriate to see
whether the effects of feature-selective attention we
observed could be captured by modeling the effects of
attending to the feature value of the stimulus. We there-
fore implemented the feature-selective attention manipu-
lation in the model by generating population responses to
two stimuli of the same feature value and modeling the
presence of feature-selective attention as feature-based
attention to that feature value.
For every predicted population response, we predicted
classifier performance when discriminating responses to
stimuli of different feature values. To do this, we com-
pared two population responses that were identical,
except that they were centered on different feature values,
as shown in Figure 3D. To simulate the three steps of stim-
ulus difference, we considered cases where the centers of
the population responses were separated by 20, 40, or 60
in the arbitrary units of the feature dimension. In the case
of stimuli varying in color, the chromaticity coordinates of
0: 0.35, 0.53, to
the stimuli varied from strongly red u
0: 0.16, 0.56, which means that, for the
strongly green u
model, we were treating a difference of 60 arbitrary units
0 chromaticity
0
as a distance of approximately 0.19 in the u
plane. For shape, the feature dimension is defined by the
transition from “X-shaped” to “non-X-shaped.” We are not
asserting that there exist neurons tuned to this novel com-
plex shape dimension in the same way as there are neu-
rons tuned to color, but for the purposes of the model,
we treated these dimensions as equivalent. Because par-
ticipant performance was similar for the color and shape
task, we used the same distances (20, 40, and 60 in the
arbitrary units) to avoid adding another parameter to the
modeling results.
v
v
v
0
0
Using the pairs of population responses (such as those
0)
in Figure 3D), we predicted classifier performance (d
using the separation of the two population responses, in
a manner analogous to that used in signal detection
0 for these population responses,
theory. To determine d
we calculated a “hit rate” for an optimal observer detecting
a signal (stimulus two) among noise (stimulus one), where
their criterion (c) is at the midpoint between the peaks
of the two curves. We defined the “hit rate” (hits) as the
area under the blue curve to the right of c and the “false
alarm rate” (FA) as the area under the red curve to the right
of c. Then the predicted classifier performance d
=
norminv(hits) − norminv(FA). In this way, for each set
of model parameters, we predicted classifier performance
in each attention condition, for each of the three step sizes
in feature difference.
0
From the predicted classification performance, we sum-
marized the predicted effects of spatial attention and
feature-selective attention using the SpatAtt and FeatAtt
values from Equations 1 and 2. Across these different
parameter sets, there was variation in the predicted
Goddard, Carlson, and Woolgar
297
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
magnitude of the effects of spatial attention and feature-
selective attention, and there was also variation in which
stimulus pair feature distances (step sizes) showed the
greatest enhancement. However, when compared with
spatial attention, feature-selective attention tended to pro-
duce relatively more enhancement of small stimulus fea-
ture differences than larger ones, as seen in the average
difference across all model parameter sets (Figure 5E).
As seen in Figure 3E, a majority of model parameter sets
(83%) showed this qualitative pattern of relative enhance-
ment across attention subtypes.
MEG Data Analysis: Granger Analysis of
Feedforward and Feedback Information Flows
We tested for temporal dependence between the patterns
of classifier performance in occipital and frontal data sets,
seeking evidence of information flows from occipital to
frontal cortices (feedforward) and from frontal to occipital
cortices (feedback), following the rationale developed in
earlier work (Karimi-Rouzbahani, 2018; Goddard, Carlson,
Dermody, & Woolgar, 2016). Specifically, we tested for
Granger causal relationships between the patterns of clas-
sifier performance based on the occipital and frontal data
sets. We summarized the color and shape information for
each region (occipital and frontal), for each time sample,
as a 6 × 4 dissimilarity matrix (DSM) of classifier perfor-
mances. For both color and shape, the 6 × 4 DSM was
defined as each pairwise comparison (six classifications
across the four levels of the feature) by four attention con-
ditions (aLaF, aLuF, uLaF, uLuF ).
The logic of Granger causality is that time series X
“Granger causes” time series Y if X contains information
that helps predict the future of Y better than information
in the past of Y alone (for a recent review of its application
in neuroscience, see Friston, Moran, & Seth, 2013). We
performed a sliding window analysis of a simplified
(special case) of Granger causality using the partial corre-
lations in Equations 3 and 4 to define feedforward (FF)
and feedback; (FB) information flows for each time
sample (t).
FF t;d;w
ð
Þ ¼ ρ DSM frontal; t
ð
Þ DSM occipital; t;d;w
ð
ð
Þ DSM frontal; t;d;w
(3)
Þ
FB t;d;w
ð
Þ ¼ ρ DSM occipital; t
ð
Þ DSM frontal; t;d;w
ð
ð
Þ DSM occipital; t;d;w
(4)
Þ
where DSM(loc, t) is the DSM based on the sources at
location loc at time t msec post stimulus onset, and
DSM(loc, t, d, w) is the DSM based on the sensors at location
loc, averaged across all time samples from t msec to t −
(d + w) msec post stimulus onset. We calculated FF and
FB for 30 overlapping windows: for five window widths
(w = 10, 20, 30, 40, or 50 msec) for each of six delays
(d = 50, 60, 70, 80, 90, or 100). We tried a range of values
for w and d to capture interactions between occipital and
frontal cortices that may occur at different timescales. Be-
cause the results were broadly similar across values of w
and d, we report FF and FB values averaged across all
values of w and d.
We report the results of this analysis in terms of the dif-
ference between the feedforward and feedback informa-
tion flows (FF − FB). To assess whether this difference
was significantly above or below chance, we generated a
null distribution of this difference for every time sample
by performing the same analysis on 1000 bootstraps of
data from each participant where the exemplar labels
were randomly permuted for each of the DSMs used in
Equations 3 and 4.
RESULTS
We acquired MEG recordings while participants catego-
rized the color (reddish or greenish) or shape (X-shaped
or non-X-shaped) of a series of stimuli that were either
closer or farther from these decision boundaries.
Behavioral Accuracy and RT
Participants were faster and more accurate at identifying
color and shape for objects that were far from the decision
boundary relative to those that were near the decision
boundary. For the color task, the average accuracy was
95.6% (SD = 3.6%) on the easy trials and 85.2% (SD =
7.3%) on the hard trials. Similarly, for the shape task, the
average accuracy was 94.1% (SD = 3.5%) on the easy trials
and 74.1% (SD = 4.7%) on the hard trials. A three-way
repeated-measures ANOVA of accuracy across Task (color
or shape), Difficulty (easy or hard), and Attended Object
Location (left or right) showed significant main effects of
Task, F(1, 19) = 41.1, p < .001, and Difficulty, F(1, 19) =
328.4, p < .001, but not Location, F(1, 19) = 1.1, p = .32,
and a significant interaction between Task and Difficulty,
F(1, 19) = 47.4, p < .001. Follow-up simple main effects
showed there was a significant effect of Difficulty on accu-
racy for both the color task, F(1, 19) = 67.4, p < .001, and
the shape task, F(1, 19) = 525.1, p < .001.
RTs were also modulated by task difficulty. For the color
task, median RT was 0.69 sec on the easy trials and 0.81 sec
on the hard trials, and for the shape task, the median RT
was 0.74 sec and 0.82 sec on the easy and hard trials,
respectively. We performed a three-way repeated-
measures ANOVA of the effects of Task, Difficulty and
Attended Location on log RT. Again, there were significant
main effects of Task, F(1, 19) = 7.0, p = .016, and Diffi-
culty, F(1, 19) = 171.7, p < .001, but not Location, F(1,
19) = 1.2, p = .28, and a significant interaction between
Task and Difficulty, F(1, 19) = 16.0, p < .001. Follow-up
simple main effects showed there was a significant effect
of Difficulty on RT for both the color task, F(1, 19) = 184.5,
p < .001, and the shape task, F(1, 19) = 56.9, p < .001. On
77% of trials, the RT was shorter than 1 sec, and the feed-
back onset was 1 sec.
298
Journal of Cognitive Neuroscience
Volume 34, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Classification Analyses of MEG Data
We trained classifiers to make a series of orthogonal dis-
criminations to quantify neural information about the par-
ticipant’s task and the stimulus, within the two ROIs. We
could robustly decode the participant’s task from both
occipital and frontal sources, indicating that neural
responses differed according to the attentional set of the
participant. Decoding of attended location (left vs. right)
peaked at 270 and 390 msec after stimulus onset (occipital
and frontal ROIs, respectively) and a decoding of attended
feature (color vs. shape) peaked at 455 msec after stimulus
onset in both ROIs. Below, we present the effects of the
attentional manipulations on the representation of object
color and shape.
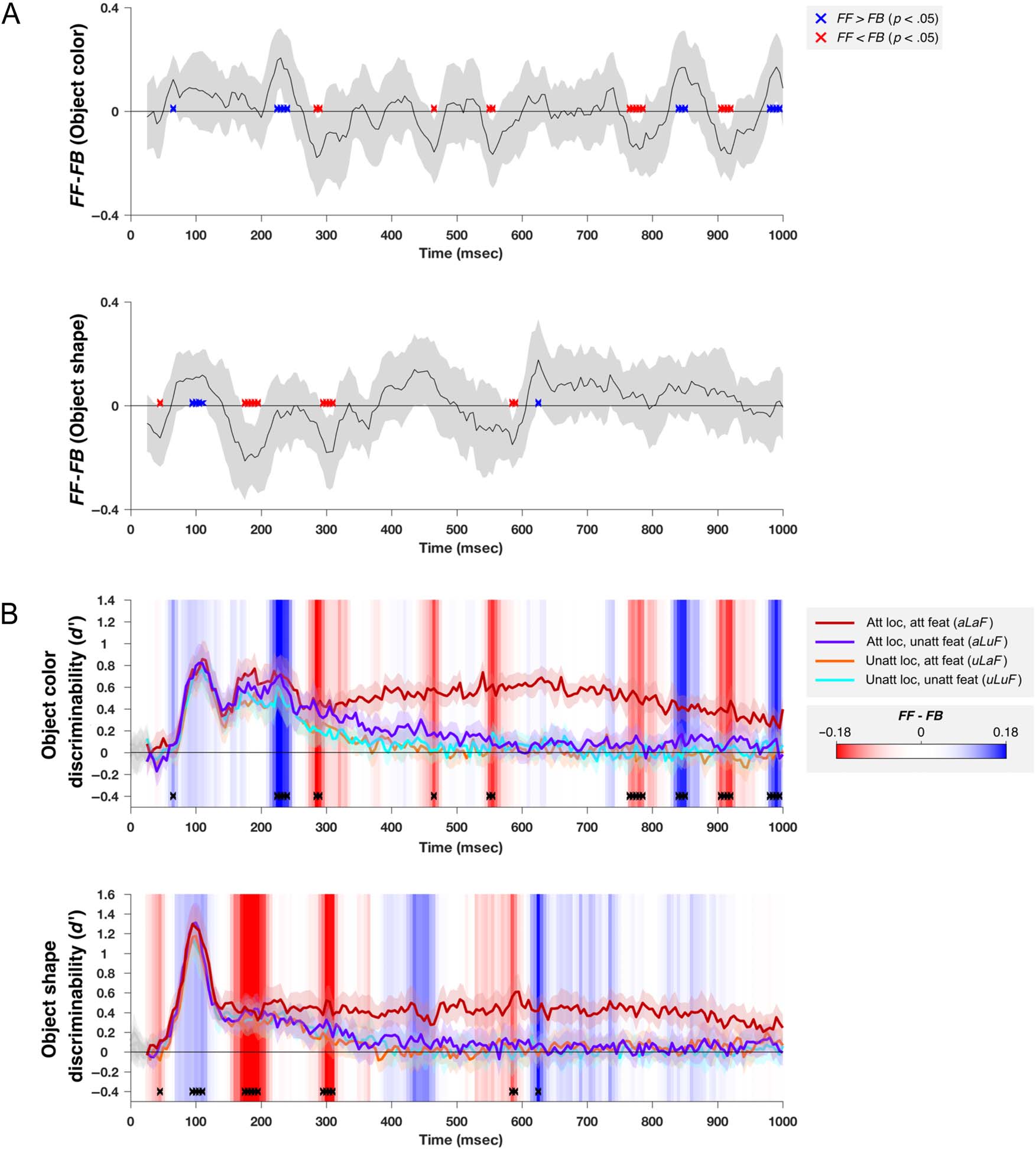
Spatial and Feature-selective Attention Interact
Multiplicatively to Boost Information Processing
First, we examined the dynamics with which spatial and
feature-selective attention affected object information
processing and how the two subtypes of attention inter-
acted in affecting this neural signal. To do so, we trained
classifiers to discriminate the color and shape of the
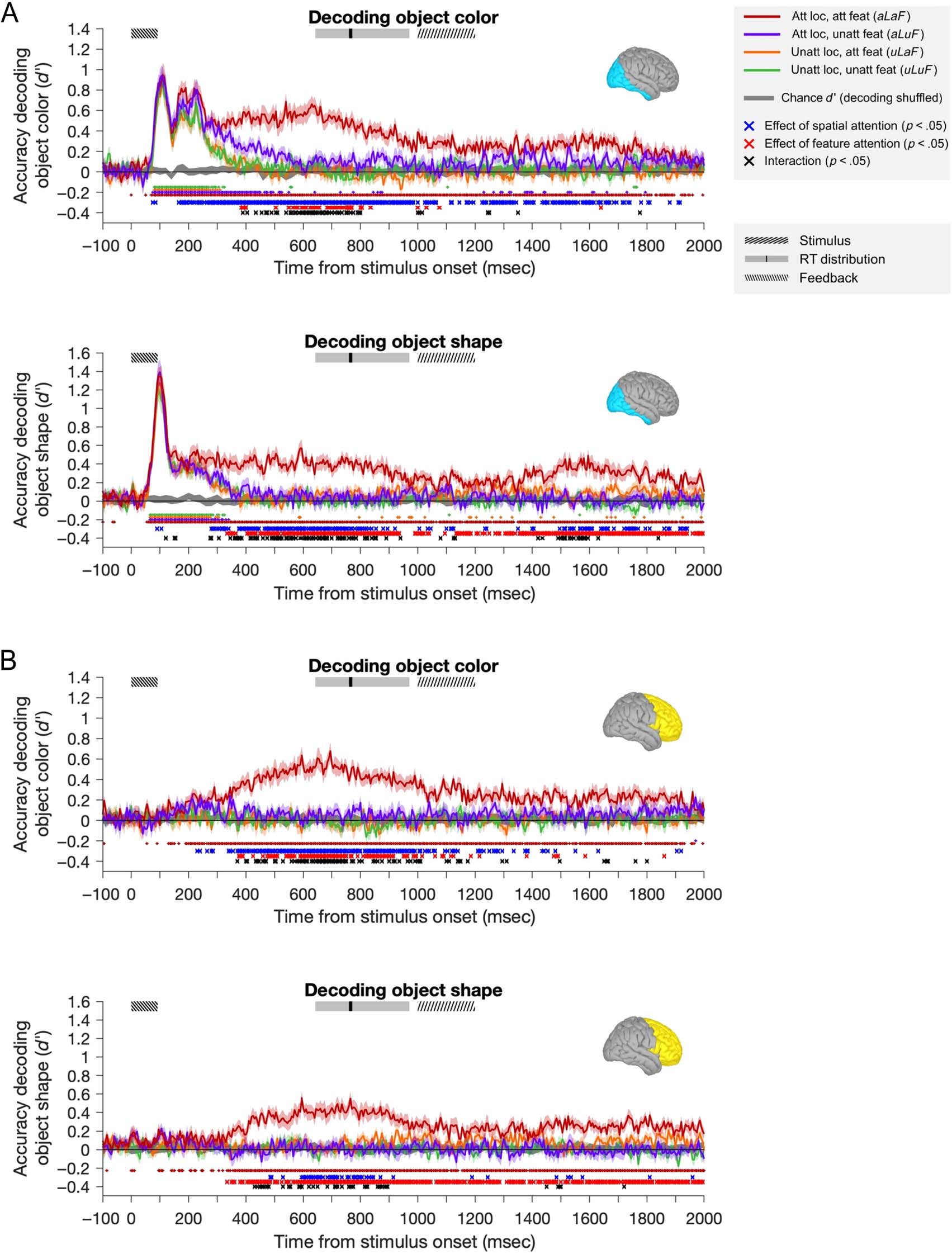
attended and nonattended objects. Figure 4 shows the
representational dynamics of object color and shape infor-
mation, for each of the four attentional conditions (2 spa-
tial locations × 2 tasks) in each ROI.
For both object color and object shape, we found signif-
icant main effects of Spatial Attention and Feature Atten-
tion and significant interactions between these effects
(at times shown in Figure 4: blue, red, and black crosses,
respectively, based on repeated-measures ANOVAs com-
pared with a permutation-based null distribution, see
Methods for details). In the occipital ROI, spatial attention
produced a small but significant increase early in the
decoding of both color and shape (blue crosses <100 msec
in Figure 4A, at 75 and 85 msec for decoding color, and
90 105 shape) or just before
the earliest peak in information processing (which was
at 105–110 color 95–100 shape).
There was no corresponding increase attributable to
feature-selective attention. For coding, there was
also a secondary early coding (∼165–240 msec),
at which time were again significant effects of Spa-
tial, but not Feature, Attention. Coding the frontal lobe
was above chance (and modulated by attention)
at these points.
For both stimulus features ROIs, attention
effects greatest magnitude emerged later, from
∼300 after onset. In occipital lobe, from
this point on, representation task-relevant
stimulus-related (Figure red traces) was
sustained, whereas equivalent all other
attentional conditions attenuated. ROI,
at this time, selective of
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
>1 sec (23%
of trials), the 200-msec feedback started at the time of response. The shaded gray region around the x-axis indicates the 95% confidence intervals of
the four classifications when performed on randomly permuted data (the empirical null distribution). Small dots below each plot indicate time
samples for which the classification of matching color was above chance level (FDR corrected, q < .05). Below these, crosses indicate time samples
for which there was a significant effect (FDR corrected, q < .05) of spatial attention (blue asterisks), feature attention (red asterisks), or an interaction
of the two (black asterisks).
300
Journal of Cognitive Neuroscience
Volume 34, Number 2
information about the attended feature at the attended
location (Figure 4B, red traces). In the occipital ROI, the
sustained effects of spatial attention preceded those of
feature-selective attention for both color (spatial from
165 msec, feature from 385 msec) and shape (spatial from
280 msec, feature from 335 msec).
From around 400 msec, for both occipital and frontal
regions (slightly earlier for shape in the occipital ROI),
there was a significant interaction between the effects of
spatial and feature-selective attention, which indicated
that the two effects combined in a multiplicative rather
than an additive manner (black crosses). In general, when-
ever both spatial and feature-selective attention had signif-
icant effects, there was also an interaction. In both cases
(color and shape), the sustained effects of spatial and
feature-selective attention interacted multiplicatively to
selectively boost in the decoding of the attended feature
at the attended location, with little enhancement in classi-
fier performance when only feature or location was
selected. That is, when location but not feature was
attended (purple lines) or when feature but not location
was attended (orange lines), decoding was closer to the
completely unattended condition (green lines) than to
the fully attended condition (dark red lines).
Information about each attended feature at the
attended location (dark red lines in Figure 4) also had later
peaks in both the occipital ROI (540–630 msec) and the
frontal ROI (595–695 msec). These peaks are well after
the offset of the stimulus (92 msec) and just before the
median RT (770 msec), suggesting they may be associated
with the participant’s decision and/or the remembered
feature value. We balanced the response mapping (by
switching the keys associated with each response pair on
half the runs and creating pseudotrials, which averaged
across equal numbers of trials from each response map-
ping), meaning that the motor preparation associated
with the participants’ response cannot have contributed
to this effect.
In summary, at early time points, all visual information
(shape and color of both objects) was represented in the
MEG trace, with some evidence for a weak modulation of
this information by spatial attention alone. At later times,
both spatial and feature-selective attention had robust
effects on coding of both shape and color in both ROIs.
These effects were multiplicative rather than additive,
leading to a selective representation of the attended fea-
ture of the attended object, which was sustained for much
of the epoch.
Control Analysis: Variable Stimulus Durations
Cannot Explain Observed Classifier Performance
Because trials were terminated when the participant broke
fixation or the eye tracker dropped a frame, stimuli were
presented for variable durations across trials. To check
that this variability could not drive the classification results
reported above, we first plotted the average duration for
each stimulus and condition and checked that there were
no identifiable differences between conditions (not
shown). Then, as a stronger test, we repeated each classi-
fication analysis above using the stimulus state (on or off )
for each time sample of each trial, instead of the neural
data (see Methods). Across time samples and classifica-
tions, the maximum group-average classifier sensitivity
0 = 0.4, indicating that variability in stimulus duration
was d
could have made a small contribution to overall classifier
performance. However, there was very little difference
between classifier accuracy for different attention condi-
tions or across step sizes. When we performed the statis-
tical tests reported in Figure 4 on the stimulus duration
data, the only significant result (effect of attended location
for decoding stimulus color) was in the opposite direction
(decoding was higher for unattended than attended
locations).
Spatial and Feature-selective Attention Have
Distinct Effects on Population Tuning Profiles
Next, we considered whether spatial and feature-selective
attention differ in the way they shape how stimulus infor-
mation in represented in population codes in human
cortex. To explore this, we considered how classifier per-
formance varied with the physical difference in the stimuli
being discriminated. Because our stimuli varied in four
“steps” along both color and shape dimensions, the pairs
of object stimuli that classifiers were trained to discrimi-
nate could be one, two, or three steps apart along either
dimension. Classifier performance generally increased
with physical difference (data not shown). Additionally,
we found that the effects of spatial and feature-selective
attention varied according to the physical discriminability
of the stimuli (Figure 5).
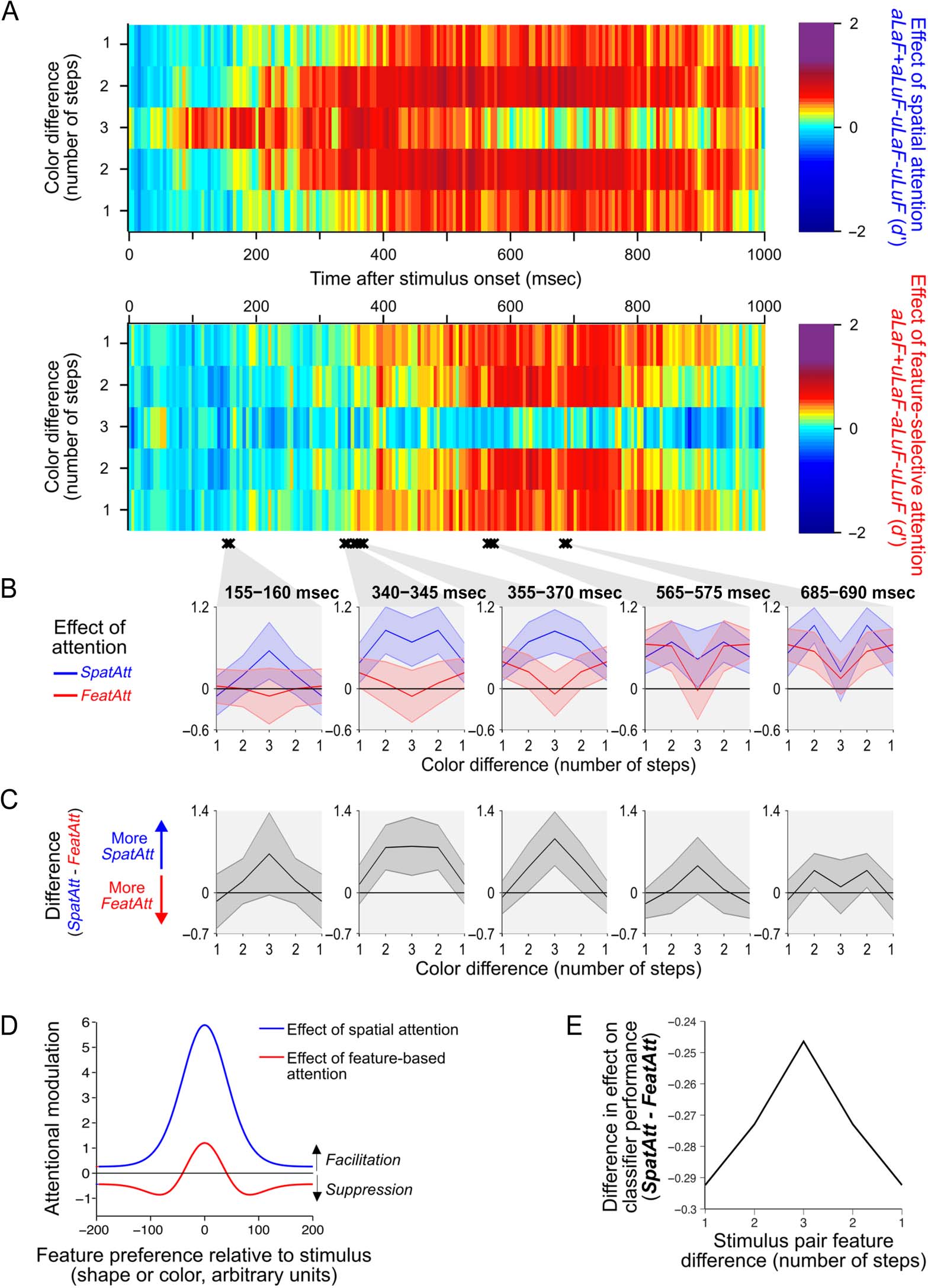
Figure 5A shows the effect of each type of attention sep-
arately, across step size and time, for coding of object color
in the occipital ROI. The effects of attention are here
expressed as the change in classifier performance between
attended and unattended conditions, so a difference of
zero (light green in Figure 5A) shows no difference
between attention conditions, rather than an overall clas-
sifier performance of zero. If spatial and feature-selective
attention produced similar effects on neural responses,
then the two plots in Figure 5A should look similar, and
the regions of yellow–red (largest improvements in
decoding with attention) should have a similar shape.
Instead, there are systematic differences between the
two in their relative effects on classifier performance
across step size. This is seen most clearly in the “convex”
versus “concave” shape of the yellow–red regions from
300 msec after stimulus onset. Although spatial attention
tended to produce greatest enhancements for stimuli sep-
arated by two steps in feature space, feature-selective
attention tended to produce greatest enhancements for
stimuli only one step apart.
Goddard, Carlson, and Woolgar
301
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Effects of spatial and feature-selective attention on the decoding of object color in the occipital ROI. (A) The effects of spatial attention (top
plot) and feature-selective attention (bottom plot) on decoding of stimulus color were calculated by taking the difference in classifier performance
(d0) between the relevant attended and unattended conditions for each step size (see Equations 1 and 2). Two-way repeated-measures ANOVAs for
each time sample revealed times where there was a significant interaction (compared with a permutation-based null distribution) between Attention
Condition and Step Size (black crosses show clusters of at least two time samples where p < .05). Data from four epochs of interest, with significant
interactions, were averaged and plotted in the insets below B. In C, the difference between the two attention effects (from the same time bins as in B)
is plotted. Data in A–C are mirror-reversed for illustration only; statistical analyses were performed on data without mirror reversals. Shaded error
bars indicate the 95% confidence interval of the between-subject mean. (D) The predicted change in simulated population response induced by
spatial and feature-based attention on a population of neuronal responses, for an example set of normalization model parameters. According to the
model, spatial attention tends to boost the response of all neurons as a multiplicative scaling of the original response, whereas feature-based
attention produces both facilitation of neurons, which prefer the attended value, and suppression of neurons preferring nearby values, which leads to
sharpening of the population response around the attended value. (E) Predicted difference between the effects of spatial (SpatAtt, Equation 1) and
feature-selective attention (FeatAtt, Equation 2) on classifier performance across pairs of stimuli with different physical differences, averaged over all
172,800 sets of model parameters we tested. The difference values plotted in C correspond to the prediction from the model in E.
302
Journal of Cognitive Neuroscience
Volume 34, Number 2
To identify times at which spatial and feature-selective
attention differed statistically in their effects across step
size, we performed a two-way repeated-measures ANOVA
compared with a permutation-based null distribution for
each time sample (see Figure 5A, black crosses, for times
of significant interaction between Attention Type and Step
Size). Then, for each cluster of time samples with signifi-
cant interactions, we plotted the average effects of spatial
and feature-selective attention (Figure 5B). We found that
the effect went in the same direction for every cluster: spa-
tial attention had a greater effect than feature-selective
attention at the largest step size (size 3), whereas
feature-selective attention had a larger effect than spatial
attention at the smallest step size (size 1). This is illustrated
most clearly in the difference plots of Figure 5C. As an
additional control, we confirmed that the same pattern
of results persisted when excluding participants with any
bias to fixate toward the attended location (data not
shown). These data suggest a robust difference between
spatial and feature-selective attention in the way they
enhance the color information in occipital areas.
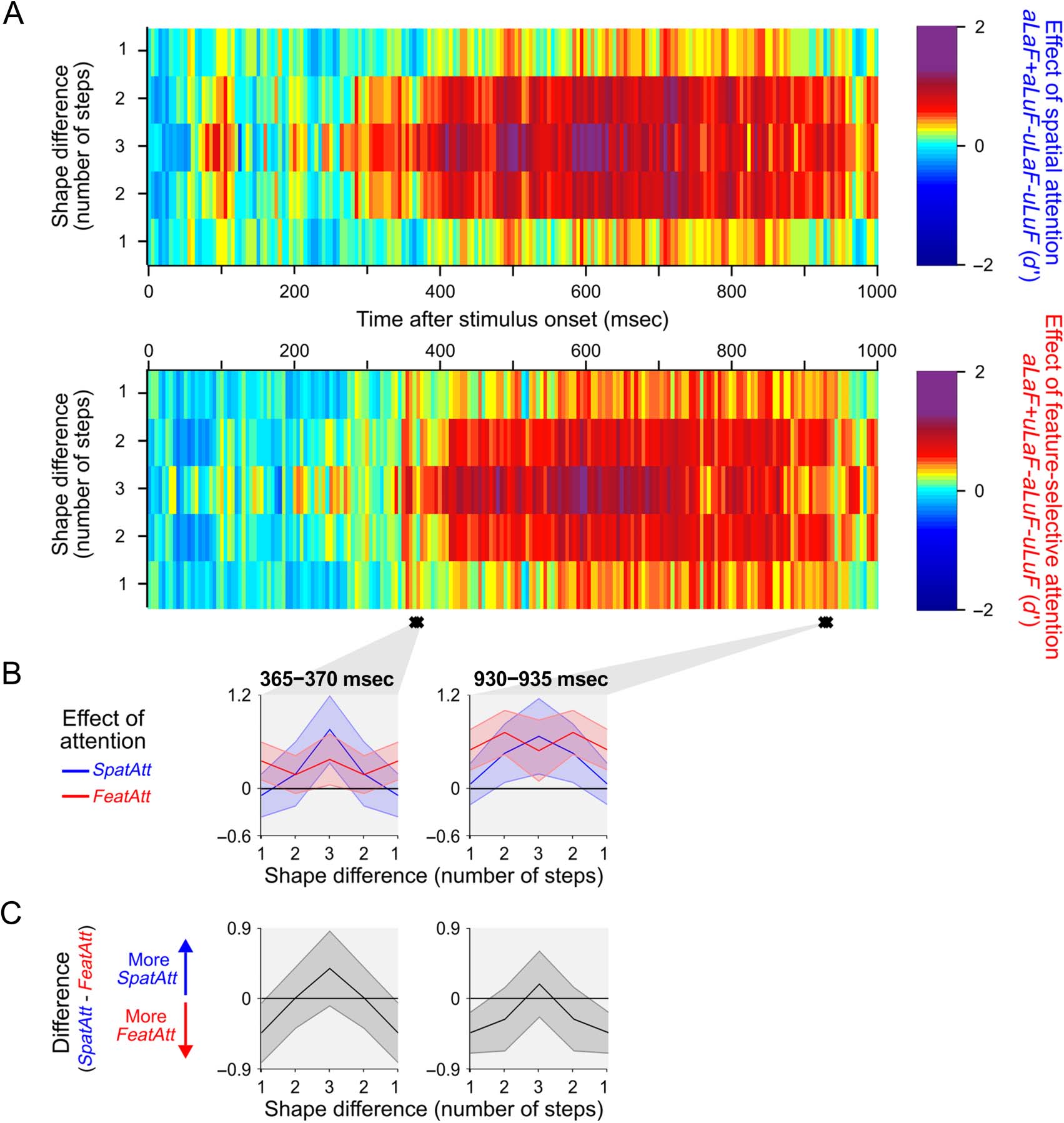
Next, we asked whether the same pattern of effects was
seen for coding of shape information. In both the occipital
and frontal ROIs, the effects of spatial and feature-selective
attention were more uniform across step sizes, and there
were no clusters of time samples with a significant interac-
tion between attention subtype and step size (data not
shown). In a more powerful analysis pooling over data
from the whole brain (sensor level; see Methods), there
were two clusters of consecutive time samples where
there was a significant interaction between attention
subtype and step size (Figure 6). Overall, any pattern of
difference was much weaker for shape than for color
(see Figure 5A vs. Figure 6A). However, where these inter-
actions occurred, the pattern of effects was in the same
direction as that shown in Figure 5C. The data, though less
definitive than for color, offer preliminary support for the
notion that there may be a general difference between
spatial and feature-selective attention in their effect on
population-level tuning.
We were interested to know whether this distinction
between the effects of spatial and feature-selective atten-
tion at the population level might reflect differences
between spatial and feature attention in their effects on
the tuning of individual neurons. To test this idea, we used
a normalization model of attention (Reynolds & Heeger,
2009; see Methods for details). A number of groups have
proposed models including normalization to describe the
effects of attention on neuronal response properties
(Boynton, 2009; Lee & Maunsell, 2009; Reynolds &
Heeger, 2009).
Model predictions for our experimental design are illus-
trated in Figure 5D–E. Details of the model predictions,
including further illustrations, are found in Figure 3.
Because the model of Reynolds and Heeger (2009) is
descriptive, with a large number of free parameters, we
systematically generated model predictions for a wide
range of model parameter sets, 172,800 in total. The large
space of model parameter sets generally converged on the
prediction that when compared with spatial attention,
feature-selective attention would produce relatively more
enhancement of small physical stimulus differences than
larger ones (Figure 5E), matching the pattern of difference
across attention types in our data.
Frontal Activity Influences the Occipital
Representation of Object Shape and Color
with a Time Course Matching the Strongest
Attentional Effects
To characterize the exchange of stimulus-related informa-
tion between the occipital and frontal ROIs, we used an
information flow analysis (Goddard et al., 2016). Because
we have fine temporal resolution measures of each pair-
wise classification, in each attention condition, we used
the pattern of classification performance across these
measures as a summary of the structure of representa-
tional space for each time sample and tested for evidence
of Granger causal interactions between the ROIs (see
Methods for details). Note that by applying this analysis
to patterns of classification performance (rather than
raw signals), we are not simply testing for evidence of
connectivity between brain regions but are specifically
testing for evidence of the exchange of stimulus-related
information between areas.
The results of this analysis are plotted in Figure 7. For
both color and shape, the earliest time samples were dom-
inated by feedforward information flow (FF > FB), consis-
tent with the early visual responses in occipital cortex
being relayed to frontal regions. These were followed by
periods of feedback information flow, starting at 285 and
185 msec for color and shape, respectively. In both cases,
information flow was biased toward the feedback direction
until ∼400 msec after stimulus onset. Interestingly, for
both color and shape, the timing of the feedback informa-
tion flows align with the onsets of the largest differences in
stimulus decoding across attention condition, despite the
later onset of these effect for color than for shape. This is
seen in Figure 7B, where the large divergence between the
dark red line (task-relevant information) and the other
conditions starts around the onset of the first red region
(FB > FF ), for both color (top panel) and shape (bottom
panel). This is compatible with the suggestion that frontal
feedback to occipital regions drives the large attentional
effects observed in occipital cortex after about 300 msec.
Moreover, it suggests that the exchange of stimulus-
related information, specifically, is important in driving
the selection of attended information in occipital cortex.
Information exchange followed different time courses
for color and shape information. For color, the early dom-
inance of feedforward information persisted for longer
(until 240 msec) than that for shape (until 115 msec). This
extra period of feedforward information flow for color
appears to correspond to the second early peak in
Goddard, Carlson, and Woolgar
303
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
decoding performance (∼165–240 msec after stimulus
onset) and could be related to higher order processing
of color information by occipital cortex at this time, such
as the ventral temporal occipital areas (Lafer-Sousa, Con-
way, & Kanwisher, 2016; Mullen, Dumoulin, McMahon, de
Zubicaray, & Hess, 2007). Conversely, because the shape
dimension we constructed for this study is highly artificial
and unlikely to correspond to a feature dimension of rele-
vance in the occipital cortex, it could be that the earlier
feedback signal in this case is related to the frontal cortex’s
involvement in storing information about the shape task
and in modifying the responses of occipital areas in such
a way that the object’s position along the shape dimension
can be read out.
As with any correlation, it is possible that our partial cor-
relations reflect correlation with another (untested) area.
Therefore, although our results are consistent with a late
dominance of feedback from frontal to occipital regions, it
is possible that the feedback could originate in another
area (e.g., parietal cortex; see Lauritzen, D’Esposito,
Heeger, & Silver, 2009). It is also possible that our source
reconstruction did not accurately isolate frontal and occip-
ital regions and that either of these includes signals from
nearby regions. However, note that if, for example, any
parietal signals were present in both frontal and occcipital
ROIs, or in the unlikely event that frontal signals were
present in the occipital ROI or vice versa, this would tend
to reduce the measures of feedfoward and feedback
information flows, rather than introduce false positives,
making this a conservative analysis. Indeed, the presence
of significant feedfoward and feedback information
flows at all provides evidence that the ROIs were well
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Effects of spatial and feature-selective attention across decoding of object shape for all MEG sensors. Plotting conventions for A–C are as in
Figure 5A–C.
304
Journal of Cognitive Neuroscience
Volume 34, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Analysis of feedforward and feedback interactions between occipital and frontal cortices. (A) FF (see Equation 3) minus FB (see Equation 4)
based on classification performance on decoding stimulus color (top plot) and shape (bottom plot). Time samples at which the difference is
significantly above or below zero (FF > FB, or FF < FB) are shown in blue and red, respectively ( p values based on bootstrapped distribution, FDR
corrected to q < .05). Shaded error bars indicate the 95% confidence interval of the between-subject mean. In (B), the occipital classification
performance in each attention condition is replotted from Figure 4A. The background of the plot is colored according to the data from A, as indicated
by the color bar. Time samples where FF − FB was significantly different from zero are also replotted, here with black crosses.
segregated from one another, as does the absence of early
classification performance in the frontal ROI.
DISCUSSION
We set out to address three open questions about the
neural mechanisms supporting selective attention: the
interaction between subtypes of attention on information
coding, whether they induce similar or distinct effects on
population tuning, and the dynamics of interregional
information exchange giving rise to them. We found, first,
that both spatial and feature-selective attention robustly
boosted the stimulus-related information and that, when
the effects of both were present, they interacted multipli-
catively. Second, we found systematic differences in their
pattern of enhancement across fine and coarse feature dis-
criminations, which are consistent with differences from
single-unit work in nonhuman primates. Third, through
our information flow analysis of Granger causal relation-
ships, we found evidence for the influence of frontal codes
Goddard, Carlson, and Woolgar
305
on occipital ones, with the onset of this influence coincid-
ing with the onset of large attentional effects in occipital
regions. We consider each of these findings below.
Spatial and Feature-selective Attention Interact
Multiplicatively to Enhance Coding of Relevant
Stimulus Information
For the decoding of both color and shape, we found that
spatial and feature-selective attention interacted multiplica-
tively, rather than having additive effects, resulting in a
selective representation of task-relevant information. Addi-
tive effects are suggested by the integrated competition
hypothesis of attention (Haazebroek et al., 2017; O’Craven
et al., 1999; Duncan et al., 1997; Duncan, 1984), which pre-
dicts that both relevant and irrelevant features of an
attended object will be boosted (object-based attention).
Additive effects are also suggested by the empirical obser-
vation that feature-selective attention can sometimes mod-
ulate responses at unattended locations (e.g., Jehee et al.,
2011; McAdams & Maunsell, 2000). However, in our data,
when the effects of both types of attention were present,
there was a clear multiplicative effect, with only the
attended feature of the attended object prioritized and no
advantage for unattended features of attended objects or
attended features of unattended objects.
It has been suggested elsewhere that spatial and feature-
based attention could combine additively in cases of low
stimulus competition (when stimuli are dissimilar, e.g.,
McAdams & Maunsell, 2000), as well as in the earliest part
of the stimulus-induced response to more similar stimuli,
followed by multiplicative interactions later in the time
course when stimuli compete to control the response
(e.g., White, Rolfs, & Carrasco, 2015). This account seeks
to reconcile the electrophysiological evidence of additive
effects with psychophysical evidence of multiplicative inter-
actions (e.g., White et al., 2015; Kingstone, 1992). In line with
this possibility, we did observe an effect of spatial attention
alone at earlier time points. Particularly for color, there
appeared to be an advantage for representation of the unat-
tended feature of the attended object (relative to features of
the unattended object) in line with object-based accounts.
However, the absence of feature-selective attention effects
in this earliest stage of the response means that our data
do not provide definitive evidence in favor of early additive
effects. The effects of feature-selective attention emerged
relatively late (from around 300 msec) and interacted with
the effects of spatial attention, again consistent with a later
emergence of multiplicative interactions. This late emer-
gence of feature-selective attention effect might reasonably
reflect processes associated with the maintenance of the
relevant feature value in working memory. Previous fMRI
(Serences, Ester, Vogel, & Awh, 2009) and ERP (Woodman &
Vogel, 2008) studies demonstrate that relevant feature infor-
mation is selectively maintained during working memory.
The absence of feature-selective attention effects in the
earliest part of the response is consistent with previous
reports of feature-selective attention effects that emerge after
the initial transient response (Mirabella et al., 2007; Hillyard
& Münte, 1984) or become stronger over time (Chen et al.,
2012). There are also similar reports for feature-based atten-
tion where the earliest responses in occipital and frontal
areas show little modulation (Bartsch et al., 2017; Bichot
et al., 2015; Zhou & Desimone, 2011). However, there are
conditions under which feature-based attention has been
shown to modulate responses from the earliest onset (Zhang
& Luck, 2009), which was not the case here.
Differential Effects of Spatial and Feature-
selective Attention
We found systematic differences between spatial and
feature-selective attention in their patterns of enhance-
ment across stimulus difference. For decoding of color
(and to a lesser extent for shape), feature-selective atten-
tion produced a relatively greater enhancement of classi-
fier performance for small physical differences than for
large differences, as compared with the effects of spatial
attention. We tested whether these differences in stimulus
coding by the population might reflect differences propa-
gated from the single-cell level by modeling population-
level effects using a normalization model (Reynolds &
Heeger, 2009). Normalization models of attention can
account for a range of the effects of attention observed at
the level of a single neuron (Ni & Maunsell, 2019; Ni, Ray,
& Maunsell, 2012; Boynton, 2005, 2009; Lee & Maunsell,
2009; Reynolds & Heeger, 2009). Here, we adapted a
normalization model to see if it could also capture atten-
tion-based changes in the information carried by the popula-
tion response. Although the Reynolds and Heeger (2009)
model is designed to model the effects of feature-based
attention (attending to a feature value), feature-selective
attention (attending to a feature-dimension) has been
demonstrated to have similar effects at the level of single
neurons (Cohen & Maunsell, 2011), and here we found
that model predictions for feature-based attention cap-
tured the qualitative effects of feature-selective attention
in our data.
Normalization models are based on the average effect of
attention on the responses of single neurons, without
modeling the heterogeneity of effects across neurons. Fur-
thermore, attention can induce population-level changes
beyond those reflected in individual tuning curves. One
example is altering the correlation structure of a popula-
tion response, which can significantly affect the informa-
tion carried by the population ( Verhoef & Maunsell,
2017; Sprague et al., 2015; Moreno-Bote et al., 2014;
Cohen & Maunsell, 2009). Despite these simplifications,
we found that a normalization model (Reynolds & Heeger,
2009) captured the different patterns of attentional
enhancement in our data. Specifically, because the model
predicts that feature-selective attention “sharpens” popu-
lation tuning along the feature dimension, whereas spatial
attention does not, it captured the main difference here
306
Journal of Cognitive Neuroscience
Volume 34, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
that feature-selective attention produces a relatively
greater enhancement of classifier performance for small
physical differences than for large differences, as com-
pared with the effects of spatial attention. In this way,
our data show that these differences between spatial and
feature-selective attention, demonstrated at a single-unit
level, may also be the source of differences observed at
the level of the population response in human cortex.
What does the success of the normalization model
imply for how these features are coded by human cortex?
We found the most marked difference between attention
subtypes in the decoding of stimulus color in the occipital
ROI. Of the two feature dimensions we manipulated
(shape and color), it is more plausible for color that there
are single units with response functions that approximate
those in the normalization model. Neurons in a range of
visual cortical areas are tuned for color (e.g., Hanazawa,
Komatsu, & Murakami, 2000; Komatsu, Ideura, Kaji, &
Yamane, 1992), and attention to color is a form of
feature-based and feature-selective attention that has been
investigated in single-unit work (Chen et al., 2012;
Mirabella et al., 2007; Bichot et al., 2005; Motter, 1994).
In contrast, the shape dimension is a more artificial, com-
plex dimension, which could align with the feature selec-
tivity of neurons in an area with intermediate to high-level
shape selectivity, (e.g., V4; see Pasupathy, 2006) but is
unlikely to correspond to a population code in the same
way as for color. We found only subtle differences between
spatial and feature-selective attention for the shape dimen-
sion, and these only reached significance at a few time
points. However, where attention subtype differences
were significant for shape (in the sensor-level decoding),
the effect was in the same direction as for color, in line with
the model predictions. This preliminary evidence suggests
that a population tuning curve framework may also be
helpful for understanding the effects of attention on
arbitrary, higher level feature dimensions. It opens the
possibility of using such models more generally as an
explanatory bridge between the single-unit and popula-
tion levels of description and further characterizing the
similarities and differences between these levels of
description. Further predictions of the model could also
be tested at a population level, for instance, the prediction
that spatial attention should induce more sharpening
along the spatial dimension than feature-selective atten-
tion. When model parameters are further constrained by
data, another direction for future work is to test quantita-
tive as well as qualitative predictions of these models.
Information Flow Analysis: The Role of Frontal
Feedback in Attentional Modulation
The earliest occipital response was primarily stimulus-
driven, showing little modulation with attention, but
strong attention effects emerged after about 300 msec.
Our information flow analysis suggests a key role for infor-
mation exchange from frontal areas, with the pattern of
stimulus-related information coding in occipital cortex
Granger-caused by information coding in the frontal lobe
shortly beforehand. Moreover, the onset of feedback dom-
inating the flow of information corresponded to the time
at which the occipital lobes showed a divergence between
task-relevant and task-irrelevant information. For decod-
ing color, where there was a second early peak in classifier
performance, this period was later (285 msec) than for
decoding shape (185 msec), but in both cases, it aligned
with the time at which information processing in the
occipital lobes became dominated by the task-relevant
information.
Seminal theories of prefrontal function converge on the
proposal that pFC implements cognitive control by affect-
ing processing in more specialized cortices (Duncan,
2001; Miller & Cohen, 2001; Dehaene et al., 1998;
Desimone & Duncan, 1995). For example, one prominent
model ( biased competition model; Duncan, 2006;
Desimone & Duncan, 1995) proposes that feedback from
the pFC biases local competition in more specialized cor-
tices in favor of task-relevant information. However, these
important proposals do not specify the nature or content
of this feedback. Our data build on these proposals by sug-
gesting a particular role for exchange of stimulus-related
information. Because our information flow analysis specif-
ically tracks the representation of stimuli, rather than sim-
ple activation, we can specify that selective processing in
occipital cortex arises, at least in part, from feedback of
stimulus-related information.
Our suggestion is consistent with work demonstrating
that the responses of frontoparietal regions contain
stimulus-related information (e.g., Freedman et al., 2001)
that increases with spatial ( Woolgar et al., 2015) and
feature-selective (Jackson et al., 2017) attention. Attentional
effects on stimulus responses by rhesus monkey prefrontal
cells emerge over a timescale that is broadly consistent with
our results (Kadohisa et al., 2013), as is the observation that
attentional effects in frontal cortices can precede those in
sensory cortex (e.g., Lennert & Martinez-Trujillo, 2013). It
goes beyond these observations, however, in specifying
that—with particular time courses—frontal codes both
result from and, in turn, influence representation in occip-
ital cortex. This also goes beyond the proposal that frontal
regions induce a preparatory biasing of sensory regions
toward the attended content or a target template (see the
review by Battistoni, Stein, & Peelen, 2017), because our
results suggest that the feedback information contains stim-
ulus information, rather than a purely attentional template.
At the point that frontal codes dominantly exert (rather than
receive) influence, selective processing begins to arise.
This is consistent with an interactive system in which selec-
tive attention arises from the dynamic exchange of stim-
ulus information favoring task-relevant processing.
Future work could build on these findings in two ways.
First, because of the spatial uncertainty of MEG source
reconstruction, we chose not to resolve into more fine-
grained parcellations of the frontal lobe. However, this
Goddard, Carlson, and Woolgar
307
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
would be an interesting avenue for future work, ideally
with concurrent EEG and individual MRI scans to help con-
strain the inverse problem. Second, with better source
estimation, it would be interesting to examine the role
of other brain regions, particularly the parietal lobe (which
is known to have important roles in attention; e.g., Hebart,
Bankson, Harel, Baker, & Cichy, 2018; Jerde, Merriam,
Riggall, Hedges, & Curtis, 2012; Lennert et al., 2011;
Woolgar, Hampshire, Thompson, & Duncan, 2011; Duncan,
2010). Our information flow analysis could be extended to
a multiple regression framework (Kietzmann et al., 2019)
to allow comparison between multiple regions.
Conclusions
We found that both spatial and feature-selective attention
enhanced the representation of visual information in
human occipital and frontal cortices and that the two sub-
types of attention interacted in a multiplicative way to yield
selective processing of task-relevant stimulus aspects. We
found differences in how spatial and feature-selective atten-
tion enhanced information across feature differences,
which were consistent with modeling based on the distinct
effects of spatial and feature-selective attention at the level
of single cells. This suggests that changes in the tuning of
single units may propagate to population-level coding, even
if the latter is also affected by changes, for example, in
correlation and covariance. An information flow analysis
specified the dynamics of information exchange between
occipital and frontal lobes and suggested that the largest
attentional effects in occipital areas may be driven by feed-
back of stimulus-related information from frontal areas.
Acknowledgments
We thank Erika Contini and Elizabeth Magdas for their assis-
tance with MEG data collection.
Reprint requests should be sent to Erin Goddard, School of
Psychology, Mathews Bldg., University of New South Wales,
Kensington, Sydney, New South Wales 2052, Australia, or via
e-mail: erin.goddard@unsw.edu.au.
Author Contributions
Erin Goddard: Conceptualization; Data curation; Formal
analysis; Methodology; Writing—Original draft; Writing—
Review & editing. Thomas A. Carlson: Conceptualization;
Funding acquisition; Writing—Review & editing. Alexandra
Woolgar: Conceptualization; Funding acquisition; Supervi-
sion; Writing—Review & editing.
Funding Information
This project was funded under an Australian Research
Council (https://dx.doi.org/10.13039/501100000923) Dis-
covery Projects (DP160101300) awarded to T. C. and
(DP170101840) awarded to A. W., an ARC Future
Fellowship (FT170100105) awarded to A. W., Medical
R e s e a r c h C o u n c i l ( h t t p s : / / d x . d o i . o r g / 1 0 . 1 3 0 3 9
/501100000265; U. K.) intramural funding SUAG/052/
G101400 awarded to A. W., and an ARC DECRA Fellowship
(DE200100139) to E. G.
Data Availability
All the raw data and the results of our classification analy-
ses are available on an Open Science Framework project
(https://doi.org/10.17605/OSF.IO/V893T).
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed
a persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by esti-
mated gender identification of first author/last author)
publishing in the Journal of Cognitive Neuroscience ( JoCN)
during this period were M(an)/M = .408, W(oman)/M =
.335, M/ W = .108, and W/ W = .149, the comparable pro-
portions for the articles that these authorship teams cited
were M/M = .579, W/M = .243, M/ W = .102, and W/ W =
.076 (Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently,
JoCN encourages all authors to consider gender balance
explicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender cita-
tion balance.
REFERENCES
Bartsch, M. V., Donohue, S. E., Strumpf, H., Schoenfeld, M. A.,
& Hopf, J.-M. (2018). Enhanced spatial focusing increases
feature-based selection in unattended locations. Scientific
Reports, 8, 16132. https://doi.org/10.1038/s41598-018-34424-5,
PubMed: 30382137
Bartsch, M. V., Loewe, K., Merkel, C., Heinze, H.-J., Schoenfeld,
M. A., Tsotsos, J. K., et al. (2017). Attention to color sharpens
neural population tuning via feedback processing in the
human visual cortex hierarchy. Journal of Neuroscience, 37,
10346–10357. https://doi.org/10.1523/JNEUROSCI.0666-17
.2017, PubMed: 28947573
Battistoni, E., Stein, T., & Peelen, M. V. (2017). Preparatory
attention in visual cortex. Annals of the New York Academy
of Sciences, 1396, 92–107. https://doi.org/10.1111/nyas.13320,
PubMed: 28253445
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false
discovery rate: A practical and powerful approach to multiple
testing. Journal of the Royal Statistical Society, Series B:
Methodological, 57, 289–300. https://doi.org/10.1111/j.2517
-6161.1995.tb02031.x
Bichot, N. P., Heard, M. T., DeGennaro, E. M., & Desimone, R.
(2015). A source for feature-based attention in the prefrontal
cortex. Neuron, 88, 832–844. https://doi.org/10.1016/j.neuron
.2015.10.001, PubMed: 26526392
Bichot, N. P., Rossi, A. F., & Desimone, R. (2005). Parallel and
serial neural mechanisms for visual search in macaque area
V4. Science, 308, 529–534. https://doi.org/10.1126/science
.1109676, PubMed: 15845848
308
Journal of Cognitive Neuroscience
Volume 34, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Boynton, G. M. (2005). Attention and visual perception.
Current Opinion in Neurobiology, 15, 465–469. https://doi
.org/10.1016/j.conb.2005.06.009, PubMed: 16023853
Boynton, G. M. (2009). A framework for describing the effects
of attention on visual responses. Vision Research, 49,
1129–1143. https://doi.org/10.1016/j.visres.2008.11.001,
PubMed: 19038281
Brainard, D. H. (1997). The Psychophysics toolbox. Spatial
Vision, 10, 433–436. https://doi.org/10.1163/156856897X00357,
PubMed: 9176952
Buracas, G. T., & Boynton, G. M. (2007). The effect of spatial
attention on contrast response functions in human visual
cortex. Journal of Neuroscience, 27, 93–97. https://doi.org
/10.1523/JNEUROSCI.3162-06.2007, PubMed: 17202476
Carrasco, M. (2011). Visual attention: The past 25 years. Vision
Research, 51, 1484–1525. https://doi.org/10.1016/j.visres.2011
.04.012, PubMed: 21549742
Chawla, D., Rees, G., & Friston, K. J. (1999). The physiological
basis of attentional modulation in extrastriate visual areas.
Nature Neuroscience, 2, 671–676. https://doi.org/10.1038
/10230, PubMed: 10404202
Chen, X., Hoffmann, K.-P., Albright, T., & Thiele, A. (2012).
Effect of feature-selective attention on neuronal responses
in macaque area MT. Journal of Neurophysiology, 107,
1530–1543. https://doi.org/10.1152/jn.01042.2010, PubMed:
22170961
Cohen, M. R., & Maunsell, J. H. R. (2009). Attention improves
performance primarily by reducing interneuronal correlations.
Nature Neuroscience, 12, 1594–1600. https://doi.org/10.1038
/nn.2439, PubMed: 19915566
Cohen, M. R., & Maunsell, J. H. R. (2011). Using neuronal
populations to study the mechanisms underlying spatial and
feature attention. Neuron, 70, 1192–1204. https://doi.org/10
.1016/j.neuron.2011.04.029, PubMed: 21689604
Corbetta, M., Miezin, F. M., Dobmeyer, S., Shulman, G. L., &
Petersen, S. E. (1990). Attentional modulation of neural
processing of shape, color, and velocity in humans. Science,
248, 1556–1559. https://doi.org/10.1126/science.2360050,
PubMed: 2360050
Dehaene, S., Kerszberg, M., & Changeux, J. P. (1998). A
neuronal model of a global workspace in effortful cognitive
tasks. Proceedings of the National Academy of Sciences, U.S.A.,
95, 14529–14534. https://doi.org/10.1073/pnas.95.24.14529,
PubMed: 9826734
Desimone, R., & Duncan, J. (1995). Neural mechanisms of
selective visual attention. Annual Review of Neuroscience,
18, 193–222. https://doi.org/10.1146/annurev.ne.18.030195
.001205, PubMed: 7605061
Duncan, J. (1984). Selective attention and the organization of
visual information. Journal of Experimental Psychology:
General, 113, 501–517. https://doi.org/10.1037/0096-3445
.113.4.501, PubMed: 6240521
Duncan, J. (2001). An adaptive coding model of neural function
in prefrontal cortex. Nature Reviews Neuroscience, 2, 820–829.
https://doi.org/10.1038/35097575, PubMed: 11715058
Duncan, J. (2006). EPS mid-career award 2004: Brain mechanisms
of attention. Quarterly Journal of Experimental Psychology,
59, 2–27. https://doi.org/10.1080/17470210500260674,
PubMed: 16556554
Duncan, J. (2010). The multiple-demand (MD) system of the
primate brain: Mental programs for intelligent behaviour.
Trends in Cognitive Sciences, 14, 172–179. https://doi.org/10
.1016/j.tics.2010.01.004, PubMed: 20171926
Duncan, J. (2013). The structure of cognition: Attentional
episodes in mind and brain. Neuron, 80, 35–50. https://doi
.org/10.1016/j.neuron.2013.09.015, PubMed: 24094101
Duncan, J., Humphreys, G., & Ward, R. (1997). Competitive
brain activity in visual attention. Current Opinion in
Neurobiology, 7, 255–261. https://doi.org/10.1016/s0959-4388
(97)80014-1, PubMed: 9142748
Fonov, V., Evans, A. C., Botteron, K., Almli, C. R., McKinstry, R. C.,
Collins, D. L., et al. (2011). Unbiased average age-appropriate
atlases for pediatric studies. Neuroimage, 54, 313–327.
https://doi.org/10.1016/j.neuroimage.2010.07.033, PubMed:
20656036
Foster, J. J., Thyer, W., Wennberg, J. W., & Awh, E. (2021).
Covert attention increases the gain of stimulus-evoked
population codes. Journal of Neuroscience, 41, 1802–1815.
https://doi.org/10.1523/JNEUROSCI.2186-20.2020, PubMed:
33441434
Freedman, D. J., & Assad, J. A. (2016). Neuronal mechanisms of
visual categorization: An abstract view on decision making.
Annual Review of Neuroscience, 39, 129–147. https://doi.org
/10.1146/annurev-neuro-071714-033919, PubMed: 27070552
Freedman, D. J., Riesenhuber, M., Poggio, T., & Miller, E. K.
(2001). Categorical representation of visual stimuli in the
primate prefrontal cortex. Science, 291, 312–316. https://doi
.org/10.1126/science.291.5502.312, PubMed: 11209083
Friston, K., Moran, R., & Seth, A. K. (2013). Analysing
connectivity with Granger causality and dynamic causal
modelling. Current Opinion in Neurobiology, 23, 172–178.
https://doi.org/10.1016/j.conb.2012.11.010, PubMed:
23265964
Goddard, E., Carlson, T. A., Dermody, N., & Woolgar, A. (2016).
Representational dynamics of object recognition: Feedforward
and feedback information flows. Neuroimage, 128, 385–397.
https://doi.org/10.1016/j.neuroimage.2016.01.006, PubMed:
26806290
Goddard, E., Mannion, D. M., McDonald, J. S., Solomon, S. G.,
& Clifford, C. W. G. (2010). Combination of subcortical color
channels in human visual cortex. Journal of Vision, 10, 1–17.
https://doi.org/10.1167/10.5.25, PubMed: 20616126
Görgen, K., Hebart, M. N., Allefeld, C., & Haynes, J.-D. (2018).
The same analysis approach: Practical protection against the
pitfalls of novel neuroimaging analysis methods. Neuroimage,
180, 19–30. https://doi.org/10.1016/j.neuroimage.2017.12.083,
PubMed: 29288130
Gouws, A. D., Alvarez, I., Watson, D. M., Uesaki, M., Rodgers, J.,
Rogers, J., et al. (2014). On the role of suppression in spatial
attention: Evidence from negative BOLD in human
subcortical and cortical structures. Journal of Neuroscience,
34, 10347–10360. https://doi.org/10.1523/JNEUROSCI.0164
-14.2014, PubMed: 25080595
Grootswagers, T., Wardle, S. G., & Carlson, T. A. (2017). Decoding
dynamic brain patterns from evoked responses: A tutorial on
multivariate pattern analysis applied to time series neuroimaging
data. Journal of Cognitive Neuroscience, 29, 677–697.
https://doi.org/10.1162/jocn_a_01068, PubMed: 27779910
Guggenmos, M., Sterzer, P., & Cichy, R. M. (2018). Multivariate
pattern analysis for MEG: A comparison of dissimilarity
measures. Neuroimage, 173, 434–447. https://doi.org/10
.1016/j.neuroimage.2018.02.044, PubMed: 29499313
Guggenmos, M., Thoma, V., Cichy, R. M., Haynes, J.-D., Sterzer,
P., & Richardson-Klavehn, A. (2015). Non-holistic coding
of objects in lateral occipital complex with and without
attention. Neuroimage, 107, 356–363. https://doi.org/10.1016
/j.neuroimage.2014.12.013, PubMed: 25512039
Haazebroek, P., Raffone, A., & Hommel, B. (2017). HiTEC: A
connectionist model of the interaction between perception
and action planning. Psychological Research, 81, 1085–1109.
https://doi.org/10.1007/s00426-016-0803-0, PubMed: 27620189
Hanazawa, A., Komatsu, H., & Murakami, I. (2000). Neural
selectivity for hue and saturation of colour in the primary
visual cortex of the monkey. European Journal of
Neuroscience, 12, 1753–1763. https://doi.org/10.1046/j.1460
-9568.2000.00041.x, PubMed: 10792452
Goddard, Carlson, and Woolgar
309
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Hayden, B. Y., & Gallant, J. L. (2009). Combined effects of
spatial and feature-based attention on responses of V4
neurons. Vision Research, 49, 1182–1187. https://doi.org
/10.1016/j.visres.2008.06.011, PubMed: 18619996
Hebart, M. N., Bankson, B. B., Harel, A., Baker, C. I., & Cichy,
R. M. (2018). The representational dynamics of task and
object processing in humans. eLife, 7. https://doi.org/10.7554
/eLife.32816, PubMed: 29384473
Henson, R. N., Mattout, J., Phillips, C., & Friston, K. J. (2009).
Selecting forward models for MEG source-reconstruction
using model-evidence. Neuroimage, 46, 168–176. https://doi
.org/10.1016/j.neuroimage.2009.01.062, PubMed: 19457358
Hillyard, S. A., & Münte, T. F. (1984). Selective attention to
color and location: An analysis with event-related brain
potentials. Perception & Psychophysics, 36, 185–198. https://
doi.org/10.3758/bf03202679, PubMed: 6514528
Huang, M. X., Mosher, J. C., & Leahy, R. M. (1999). A
sensor-weighted overlapping-sphere head model and
exhaustive head model comparison for MEG. Physics in
Medicine and Biology, 44, 423–440. https://doi.org/10.1088
/0031-9155/44/2/010, PubMed: 10070792
Jackson, J., Rich, A. N., Williams, M. A., & Woolgar, A. (2017).
Feature-selective attention in fronto-parietal cortex:
Multivoxel codes adjust to prioritize task-relevant
information. Journal of Cognitive Neuroscience, 29, 310–321.
https://doi.org/10.1162/jocn_a_01039, PubMed: 27626230
Jackson, J. B., & Woolgar, A. (2018). Adaptive coding in the
human brain: Distinct object features are encoded by
overlapping voxels in frontoparietal cortex. Cortex, 108,
25–34. https://doi.org/10.1016/j.cortex.2018.07.006, PubMed:
30121000
Jehee, J. F. M., Brady, D. K., & Tong, F. (2011). Attention
improves encoding of task-relevant features in the human
visual cortex. Journal of Neuroscience, 31, 8210–8219.
https://doi.org/10.1523/JNEUROSCI.6153-09.2011, PubMed:
21632942
Jerde, T. A., Merriam, E. P., Riggall, A. C., Hedges, J. H., &
Curtis, C. E. (2012). Prioritized maps of space in human
frontoparietal cortex. Journal of Neuroscience, 32,
17382–17390. https://doi.org/10.1523/JNEUROSCI.3810-12
.2012, PubMed: 23197729
Kado, H., Higuchi, M., Shimogawara, M., Haruta, Y., Adachi, Y., Kawai,
J., et al. (1999). Magnetoencephalogram systems developed
at KIT. IEEE Transactions on Applied Superconductivity, 9,
4057–4062. https://doi.org/10.1109/77.783918
Kadohisa, M., Petrov, P., Stokes, M., Sigala, N., Buckley, M.,
Gaffan, D., et al. (2013). Dynamic construction of a coherent
attentional state in a prefrontal cell population. Neuron,
80, 235–246. https://doi.org/10.1016/j.neuron.2013.07.041,
PubMed: 24035763
Karimi-Rouzbahani, H. (2018). Three-stage processing of
category and variation information by entangled interactive
mechanisms of peri-occipital and peri-frontal cortices.
Scientific Reports, 8, 12213. https://doi.org/10.1038/s41598
-018-30601-8, PubMed: 30111859
Kietzmann, T. C., Spoerer, C. J., Sörensen, L. K. A., Cichy, R. M.,
Hauk, O., & Kriegeskorte, N. (2019). Recurrence is required
to capture the representational dynamics of the human visual
system. Proceedings of the National Academy of Sciences,
U.S.A., 116, 21854–21863. https://doi.org/10.1073/pnas
.1905544116, PubMed: 31591217
Kingstone, A. (1992). Combining expectancies. Quarterly
Journal of Experimental Psychology A: Human
Experimental Psychology, 44, 69–104. https://doi.org/10.1080
/14640749208401284
Kleiner, M., Brainard, D., & Pelli, D. G. (2007). What’s new
in Psychtoolbox-3? Perception, 36, 1–16, ECVP Abstract
Supplement.
Koch, C., & Ullman, S. (1985). Shifts in selective visual
attention: Towards the underlying neural circuitry. Human
Neurobiology, 4, 219–227. PubMed: 3836989
Komatsu, H., Ideura, Y., Kaji, S., & Yamane, S. (1992). Color
selectivity of neurons in the inferior temporal cortex of the
awake macaque monkey. Journal of Neuroscience, 12,
408–424. https://doi.org/10.1523/JNEUROSCI.12-02-00408
.1992, PubMed: 1740688
Lafer-Sousa, R., Conway, B. R., & Kanwisher, N. G. (2016).
Color-biased regions of the ventral visual pathway lie
between face- and place-selective regions in humans, as in
macaques. Journal of Neuroscience, 36, 1682–1697. https://
doi.org/10.1523/JNEUROSCI.3164-15.2016, PubMed:
26843649
Lauritzen, T. Z., D’Esposito, M., Heeger, D. J., & Silver, M. A.
(2009). Top–down flow of visual spatial attention signals from
parietal to occipital cortex. Journal of Vision, 9, 18.1–18.14.
https://doi.org/10.1167/9.13.18, PubMed: 20055551
Lee, J., & Maunsell, J. H. R. (2009). A normalization model of
attentional modulation of single unit responses. PLoS One, 4,
e4651. https://doi.org/10.1371/journal.pone.0004651,
PubMed: 19247494
Lee, J., & Maunsell, J. H. R. (2010). The effect of attention on
neuronal responses to high and low contrast stimuli. Journal
of Neurophysiology, 104, 960–971. https://doi.org/10.1152/jn
.01019.2009, PubMed: 20538780
Lennert, T., Cipriani, R., Jolicoeur, P., Cheyne, D., & Martinez-
Trujillo, J. C. (2011). Attentional modulation of
neuromagnetic evoked responses in early human visual
cortex and parietal lobe following a rank-order rule. Journal
of Neuroscience, 31, 17622–17636. https://doi.org/10.1523
/JNEUROSCI.4781-11.2011, PubMed: 22131423
Lennert, T., & Martinez-Trujillo, J. C. (2013). Prefrontal neurons
of opposite spatial preference display distinct target selection
dynamics. Journal of Neuroscience, 33, 9520–9529. https://
doi.org/10.1523/JNEUROSCI.5156-12.2013, PubMed:
23719818
Li, X., Lu, Z.-L., Tjan, B. S., Dosher, B. A., & Chu, W. (2008).
Blood oxygenation level-dependent contrast response
functions identify mechanisms of covert attention in early
visual areas. Proceedings of the National Academy of
Sciences, U.S.A., 105, 6202–6207. https://doi.org/10.1073/pnas
.0801390105, PubMed: 18413602
Liu, T., Hospadaruk, L., Zhu, D. C., & Gardner, J. L. (2011).
Feature-specific attentional priority signals in human cortex.
Journal of Neuroscience, 31, 4484–4495. https://doi.org/10
.1523/JNEUROSCI.5745-10.2011, PubMed: 21430149
Luo, T. Z., & Maunsell, J. H. R. (2018). Attentional changes in
either criterion or sensitivity are associated with robust
modulations in lateral prefrontal cortex. Neuron, 97,
1382–1393. https://doi.org/10.1016/j.neuron.2018.02.007,
PubMed: 29503191
Martinez-Trujillo, J., & Treue, S. (2002). Attentional modulation
strength in cortical area MT depends on stimulus contrast.
Neuron, 35, 365–370. https://doi.org/10.1016/s0896-6273(02)
00778-x
Martinez-Trujillo, J. C., & Treue, S. (2004). Feature-based
attention increases the selectivity of population responses in
primate visual cortex. Current Biology, 14, 744–751. https://
doi.org/10.1016/j.cub.2004.04.028, PubMed: 15120065
Maunsell, J. (2015). Neuronal mechanisms of visual attention.
Annual Review of Vision Science, 1, 373–391. https://doi.org
/10.1146/annurev-vision-082114-035431, PubMed: 28532368
McAdams, C. J., & Maunsell, J. H. (1999). Effects of attention on
orientation-tuning functions of single neurons in macaque
cortical area V4. Journal of Neuroscience, 19, 431–441.
https://doi.org/10.1523/JNEUROSCI.19-01-00431.1999,
PubMed: 9870971
310
Journal of Cognitive Neuroscience
Volume 34, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
McAdams, C. J., & Maunsell, J. H. (2000). Attention to both
space and feature modulates neuronal responses in macaque
area V4. Journal of Neurophysiology, 83, 1751–1755. https://
doi.org/10.1152/jn.2000.83.3.1751, PubMed: 10712494
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of
prefrontal cortex function. Annual Review of Neuroscience,
24, 167–202. https://doi.org/10.1146/annurev.neuro.24.1.167,
PubMed: 11283309
Mirabella, G., Bertini, G., Samengo, I., Kilavik, B. E., Frilli, D.,
Libera, C. D., et al. (2007). Neurons in area V4 of the macaque
translate attended visual features into behaviorally relevant
categories. Neuron, 54, 303–318. https://doi.org/10.1016/j
.neuron.2007.04.007, PubMed: 17442250
Moore, T., Armstrong, K. M., & Fallah, M. (2003). Visuomotor
origins of covert spatial attention. Neuron, 40, 671–683. https://
doi.org/10.1016/s0896-6273(03)00716-5, PubMed: 14622573
Moreno-Bote, R., Beck, J., Kanitscheider, I., Pitkow, X., Latham,
P., & Pouget, A. (2014). Information-limiting correlations.
Nature Neuroscience, 17, 1410–1417. https://doi.org/10.1038
/nn.3807, PubMed: 25195105
Journal of Neuroscience, 19, 1736–1753. https://doi.org/10
.1523/JNEUROSCI.19-05-01736.1999, PubMed: 10024360
Reynolds, J. H., & Heeger, D. J. (2009). The normalization
model of attention. Neuron, 61, 168–185. https://doi.org/10
.1016/j.neuron.2009.01.002, PubMed: 19186161
Reynolds, J. H., Pasternak, T., & Desimone, R. (2000). Attention
increases sensitivity of V4 neurons. Neuron, 26, 703–714.
https://doi.org/10.1016/s0896-6273(00)81206-4, PubMed:
10896165
Rossi, A., & Paradiso, M. (1995). Feature-specific effects of
selective visual attention. Vision Research, 35, 621–634. https://
doi.org/10.1016/0042-6989(94)00156-g, PubMed: 7900301
Saenz, M., Buracas, G. T., & Boynton, G. M. (2002). Global
effects of feature-based attention in human visual cortex.
Nature Neuroscience, 5, 631–632. https://doi.org/10.1038
/nn876, PubMed: 12068304
Saenz, M., Buracas, G. T., & Boynton, G. M. (2003). Global
feature-based attention for motion and color. Vision
Research, 43, 629–637. https://doi.org/10.1016/s0042-6989
(02)00595-3, PubMed: 12604099
Motter, B. C. (1994). Neural correlates of attentive selection for
Scolari, M., Byers, A., & Serences, J. T. (2012). Optimal
color or luminance in extrastriate area V4. Journal of
Neuroscience, 14, 2178–2189. https://doi.org/10.1523
/JNEUROSCI.14-04-02178.1994, PubMed: 8158264
Mullen, K. T., Dumoulin, S. O., McMahon, K. L., de Zubicaray,
G. I., & Hess, R. F. (2007). Selectivity of human retinotopic
visual cortex to S-cone-opponent, L/M-cone-opponent and
achromatic stimulation. European Journal of Neuroscience,
25, 491–502. https://doi.org/10.1111/j.1460-9568.2007.05302
.x, PubMed: 17284191
Ni, A. M., & Maunsell, J. H. R. (2019). Neuronal effects of spatial
and feature attention differ due to normalization. Journal
of Neuroscience, 39, 5493–5505. https://doi.org/10.1523
/JNEUROSCI.2106-18.2019, PubMed: 31068439
Ni, A. M., Ray, S., & Maunsell, J. H. R. (2012). Tuned
normalization explains the size of attention modulations.
Neuron, 73, 803–813. https://doi.org/10.1016/j.neuron.2012
.01.006, PubMed: 22365552
O’Craven, K. M., Downing, P. E., & Kanwisher, N. (1999). fMRI
evidence for objects as the units of attentional selection.
Nature, 401, 584–587. https://doi.org/10.1038/44134,
PubMed: 10524624
Op de Beeck, H. P., Baker, C. I., DiCarlo, J. J., & Kanwisher, N. G.
(2006). Discrimination training alters object representations
in human extrastriate cortex. Journal of Neuroscience, 26,
13025–13036. https://doi.org/10.1523/JNEUROSCI.2481-06
.2006, PubMed: 17167092
Pasupathy, A. (2006). Neural basis of shape representation in
the primate brain. Progress in Brain Research, 154, 293–313.
https://doi.org/10.1016/S0079-6123(06)54016-6
Patzwahl, D. R., & Treue, S. (2009). Combining spatial and
feature-based attention within the receptive field of MT
neurons. Vision Research, 49, 1188–1193. https://doi.org/10
.1016/j.visres.2009.04.003, PubMed: 19362573
Peelen, M. V., & Kastner, S. (2014). Attention in the real world:
Toward understanding its neural basis. Trends in Cognitive
Sciences, 18, 242–250. https://doi.org/10.1016/j.tics.2014.02
.004, PubMed: 24630872
Pelli, D. G. (1997). The videotoolbox software for visual
psychophysics: Transforming numbers into movies. Spatial
Vision, 10, 437–442. https://doi.org/10.1163/156856897X00366,
PubMed: 9176953
Pestilli, F., & Carrasco, M. (2005). Attention enhances contrast
sensitivity at cued and impairs it at uncued locations. Vision
Research, 45, 1867–1875. https://doi.org/10.1016/j.visres.2005
.01.019, PubMed: 15797776
Reynolds, J. H., Chelazzi, L., & Desimone, R. (1999). Competitive
mechanisms subserve attention in macaque areas V2 and V4.
deployment of attentional gain during fine
discriminations. Journal of Neuroscience, 32, 7723–7733.
https://doi.org/10.1523/ JNEUROSCI.5558-11.2012,
PubMed: 22649250
Serences, J. T., & Boynton, G. M. (2007). Feature-based
attentional modulations in the absence of direct visual
stimulation. Neuron, 55, 301–312. https://doi.org/10.1016/j
.neuron.2007.06.015, PubMed: 17640530
Serences, J. T., Ester, E. F., Vogel, E. K., & Awh, E. (2009).
Stimulus-specific delay activity in human primary visual
cortex. Psychological Science, 20, 207–214. https://doi.org/10
.1111/j.1467-9280.2009.02276.x, PubMed: 19170936
Sprague, T. C., Saproo, S., & Serences, J. T. (2015). Visual
attention mitigates information loss in small- and
large-scale neural codes. Trends in Cognitive Sciences, 19,
215–226. https://doi.org/10.1016/j.tics.2015.02.005,
PubMed: 25769502
Störmer, V. S., & Alvarez, G. A. (2014). Feature-based attention
elicits surround suppression in feature space. Current
Biology, 24, 1985–1988. https://doi.org/10.1016/j.cub.2014.07
.030, PubMed: 25155510
Sundberg, K. A., Mitchell, J. F., & Reynolds, J. H. (2009). Spatial
attention modulates center-surround interactions in macaque
visual area V4. Neuron, 61, 952–963. https://doi.org/10.1016/j
.neuron.2009.02.023, PubMed: 19324003
Tadel, F., Baillet, S., Mosher, J. C., Pantazis, D., & Leahy, R. M.
(2011). Brainstorm: A user-friendly application for MEG/EEG
analysis. Computational Intelligence and Neuroscience,
2011, 879716. https://doi.org/10.1155/2011/879716, PubMed:
21584256
Treue, S., & Martinez-Trujillo, J. C. (1999). Feature-based
attention influences motion processing gain in macaque
visual cortex. Nature, 399, 575–579. https://doi.org/10.1038
/21176, PubMed: 10376597
Uehara, G., Adachi, Y., Kawai, J., Shimogawara, M., Higuchi, M.,
Haruta, Y., et al. (2003). Multi-channel SQUID systems for
biomagnetic measurement. IEICE Transactions on
Electronics, E86-C, 43–54.
Uusitalo, M. A., & Ilmoniemi, R. J. (1997). Signal-space projection
method for separating MEG or EEG into components.
Medical & Biological Engineering & Computing, 35, 135–140.
https://doi.org/10.1007/BF02534144, PubMed: 9136207
Vaziri-Pashkam, M., & Xu, Y. (2017). Goal-directed visual
processing differentially impacts human ventral and dorsal
visual representations. Journal of Neuroscience, 37,
8767–8782. https://doi.org/10.1523/JNEUROSCI.3392-16
.2017, PubMed: 28821655
Goddard, Carlson, and Woolgar
311
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Verghese, P., Kim, Y.-J., & Wade, A. R. (2012). Attention selects
informative neural populations in human V1. Journal of
Neuroscience, 32, 16379–16390. https://doi.org/10.1523
/JNEUROSCI.1174-12.2012, PubMed: 23152620
Verhoef, B.-E., & Maunsell, J. H. R. (2017). Attention-related
changes in correlated neuronal activity arise from
normalization mechanisms. Nature Neuroscience, 20,
969–977. https://doi.org/10.1038/nn.4572, PubMed: 28553943
White, A. L., Rolfs, M., & Carrasco, M. (2015). Stimulus
competition mediates the joint effects of spatial and
feature-based attention. Journal of Vision, 15, 7. https://doi
.org/10.1167/15.14.7, PubMed: 26473316
Williford, T., & Maunsell, J. H. R. (2006). Effects of spatial
attention on contrast response functions in macaque area V4.
Journal of Neurophysiology, 96, 40–54. https://doi.org/10
.1152/jn.01207.2005, PubMed: 16772516
Wolfe, J. M. (1994). Guided Search 2.0: A revised model of
visual search. Psychonomic Bulletin & Review, 1, 202–238.
https://doi.org/10.3758/BF03200774, PubMed: 24203471
Woodman, G. F., & Vogel, E. K. (2008). Selective storage and
maintenance of an object’s features in visual working
memory. Psychonomic Bulletin & Review, 15, 223–229.
https://doi.org/10.3758/pbr.15.1.223, PubMed: 18605507
Woolgar, A., Hampshire, A., Thompson, R., & Duncan, J. (2011).
Adaptive coding of task-relevant information in human
frontoparietal cortex. Journal of Neuroscience, 31,
14592–14599. https://doi.org/10.1523/JNEUROSCI.2616-11
.2011, PubMed: 21994375
Woolgar, A., Williams, M. A., & Rich, A. N. (2015). Attention
enhances multi-voxel representation of novel objects in frontal,
parietal and visual cortices. Neuroimage, 109, 429–437. https://
doi.org/10.1016/j.neuroimage.2014.12.083, PubMed: 25583612
Wyszecki, G., & Stiles, W. S. (1982). Color science: Concepts and
methods, quantitative data and formulas. New York: Wiley.
Zhang, W., & Luck, S. J. (2009). Feature-based attention
modulates feedforward visual processing. Nature
Neuroscience, 12, 24–25. https://doi.org/10.1038/nn.2223,
PubMed: 19029890
Zhou, H., & Desimone, R. (2011). Feature-based attention in
the frontal eye field and area V4 during visual search.
Neuron, 70, 1205–1217. https://doi.org/10.1016/j.neuron
.2011.04.032, PubMed: 21689605
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
2
2
9
0
1
9
8
0
9
8
9
/
j
o
c
n
_
a
_
0
1
7
9
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
312
Journal of Cognitive Neuroscience
Volume 34, Number 2