Set-relevance Determines the Impact of Distractors on
Episodic Memory Retrieval
Sze Chai Kwok1, Tim Shallice2,3, and Emiliano Macaluso1
Abstract
■ We investigated the interplay between stimulus-driven atten-
tion and memory retrieval with a novel interference paradigm
that engaged both systems concurrently on each trial. Partici-
pants encoded a 45-min movie on Day 1 and, on Day 2, per-
formed a temporal order judgment task during fMRI. Each
retrieval trial comprised three images presented sequen-
tially, and the task required participants to judge the temporal
order of the first and the last images (“memory probes”) while
ignoring the second image, which was task irrelevant (“attention
distractor”). We manipulated the content relatedness and the
temporal proximity between the distractor and the memory
probes, as well as the temporal distance between two probes.
Behaviorally, short temporal distances between the probes led
to reduced retrieval performance. Distractors that at encoding
were temporally close to the first probe image reduced these
costs, specifically when the distractor was content unrelated to
the memory probes. The imaging results associated the distrac-
tor probe temporal proximity with activation of the right ventral
attention network. By contrast, the precuneus was activated for
high-content relatedness between distractors and probes and in
trials including a short distance between the two memory
probes. The engagement of the right ventral attention network
by specific types of distractors suggests a link between stimulus-
driven attention control and episodic memory retrieval, whereas
the activation pattern of the precuneus implicates this region in
memory search within knowledge/content-based hierarchies. ■
INTRODUCTION
Attention and memory are traditionally studied as sepa-
rate topics, and the interaction between the two has been
relatively neglected. Nonetheless, in most situations, the
interplay between these two systems is intimate and two
ways (Hutchinson & Turk-Browne, 2012). Memory can
serve to modulate the allocation of attention, facilitating
perception of current stimuli (Stokes, Atherton, Patai, &
Nobre, 2012; Summerfield, Rao, Garside, & Nobre, 2011;
Chun & Jiang, 2003), whereas the deployment of atten-
tion is necessary for effective memory encoding (Turk-
Browne, Golomb, & Chun, 2013; Baddeley, Lewis, Eldridge,
& Thomson, 1984), working memory maintenance (Zanto
& Gazzaley, 2009), and retrieval (Guerin, Robbins, Gilmore, &
Schacter, 2012; Craik, Govoni, Naveh-Benjamin, & Anderson,
1996). Here, we introduce a novel paradigm with the
aim of engaging both functions concurrently within the
same trial. We presented irrelevant distractors, while par-
ticipants were asked to retrieve temporal information
from episodic memory. By manipulating the temporal
and content relationships between the irrelevant sensory
distractors and the task-relevant memory probes, we
investigated the interplay between the selection/filtering
of information in the external world (attention) and
the selection of information stored internally (memory
retrieval).
1Santa Lucia Foundation, Rome, Italy, 2SISSA, Trieste, Italy,
3University College London
Selective Processing: A Common Constraint for
Attention and Memory Retrieval
Several previous neuroimaging studies highlighted that
both attention control and episodic retrieval involve
areas in the frontal and parietal cortex (see Naghavi &
Nyberg, 2005, for a review). In the memory domain,
the parietal cortex has been implicated in a wide range
of functions. Among them are its role(s) in successful
recollection of source details (Hayama, Vilberg, & Rugg,
2012), as an online “episodic buffer” of recollected infor-
mation (Vilberg & Rugg, 2008, 2012), and in the accumu-
lation of evidence leading to an eventual mnemonic
decision (Wheeler & Buckner, 2003). With respect of
the possible relationship between memory and attention,
it has been put forward that the parietal cortex may
support the directing of attention toward internal mne-
monic representations (e.g., Wagner, Shannon, Kahn,
& Buckner, 2005). Specifying this relationship further,
Ciaramelli, Grady, Levine, Ween, and Moscovitch (2010)
demonstrated a dissociation between the dorsal and
ventral parietal cortices in episodic memory retrieval,
which parallels the classical distinction between endog-
enous attention control in the intraparietal sulcus and
FEFs versus exogenous control in right TPJ (rTPJ) and
the right inferior frontal gyrus (rIFG; see Corbetta, Patel,
& Shulman, 2008; Corbetta & Shulman, 2002). The find-
ing of such correspondences has led researchers to pro-
pose that the orienting to external signals (attention) and
to internal/stored information (memory) may involve
© 2014 Massachusetts Institute of Technology Published under a
Creative Commons Attribution 3.0 Unported (CC BY 3.0) license
Journal of Cognitive Neuroscience 26:9, pp. 2070–2086
doi:10.1162/jocn_a_00601
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
some common processes (Chun & Johnson, 2011; Cabeza,
Ciaramelli, Olson, & Moscovitch, 2008; Ciaramelli, Grady, &
Moscovitch, 2008; but see Sestieri, Shulman, & Corbetta,
2010; Hutchinson, Uncapher, & Wagner, 2009).
Together with the parietal cortex, frontal regions have
also been implicated both in attention and episodic
memory retrieval. The left ventrolateral pFC is activated
during retrieval of semantic knowledge (Prince, Tsukiura,
& Cabeza, 2007). These effects are thought to reflect the
involvement of interference resolution processes (Milham
et al., 2001) and/or postretrieval selection to discriminate
among competing alternatives. In particular, the IFG
appears to support a generalized control process that
selects relevant information from among competitors
(Badre, Poldrack, Paré-Blagoev, Insler, & Wagner, 2005;
Thompson-Schill, DʼEsposito, Aguirre, & Farah, 1997).
The need to select between alternative possibilities in
the memory domain bears some similarity with attention
control mechanisms that, together with orienting/shifting
operations, are needed to process task-relevant targets in
the presence of irrelevant distractors. Examples include
visual search paradigms when participants have to identify
a target among distractors and selective attention tasks
when participants have to maintain attention to a specific
location (or feature) while ignoring distractors presented
at other locations. These tasks entail endogenous, top–
down control processes and activate intraparietal sulcus
and FEF bilaterally (Fairhall, Indovina, Driver, & Macaluso,
2009; Hopfinger, Buonocore, & Mangun, 2000; Kastner &
Ungerleider, 2000). By contrast, rTPJ and rIFG are acti-
vated when a single target is presented at an unattended
location (Corbetta & Shulman, 2002) but are deactivated
in selective attention tasks that include competing dis-
tractors (Shulman et al., 2003; see also DiQuattro & Geng,
2011). Thus, in attention/perception, the “filtering” opera-
tion that is required when participants have to maintain a
task goal in the face of distracting signals has been asso-
ciated primarily with deactivation of the rTPJ–IFG network
(Corbetta et al., 2008; see also Sestieri, Corbetta, Romani,
& Shulman, 2011; Sestieri et al., 2010; Todd, Fougnie, &
Marois, 2005).
Set Relevance: The Role of Information Shared by
Task-relevant and Task-irrelevant Stimuli
These attention-related findings appear to be discrepant
with the memory results that show increased activation
of IFG in the presence of distracting competitors (e.g.,
Badre et al., 2005). However, recent data may provide
us with a framework to reconcile these seemingly contra-
dictory results. Attention tasks that showed deactivation
of the ventral frontoparietal network made use of irrele-
vant stimuli, which do not have any relationship with
the task-relevant targets (Shulman et al., 2003; see also
Indovina & Macaluso, 2007; Kincade, Abrams, Astafiev,
Shulman, & Corbetta, 2005). In fact, studies that specifi-
cally manipulated the relationship between the target
and the distractors found the target–distractors related-
ness to be a key determinant of the response pattern
in the ventral attention system. Collectively referred as
studies of “contingent attention capture” (Folk, Leber,
& Egeth, 2002; Folk, Remington, & Johnston, 1992), sev-
eral imaging experiments found that distractors sharing
some goal-defining feature with the target activate, rather
than deactivate, the rTPJ (Natale, Marzi, & Macaluso,
2010; Serences et al., 2005). For example, in a sustained
visual task requiring participants to name red letters at
one location, the occurrence of a red nonletter distractor
at a different location will trigger contingent capture of
attention: that is, attention is pulled toward the distractor
location, because the attribute “redness” is part of the
current task set (Serences et al., 2005; see also Geng &
Mangun, 2011). Thus, in attention/perception, the ventral
attention system responds to task-irrelevant stimuli only
when sharing some aspect with the task-relevant targets
(i.e., set-relevant distractors). These paradigms high-
light the distinction between “task relevance” and “set
relevance.” Task relevance refers to some characteristics
of the experimental setting that requires a judgment/
response (e.g., a feature of the target) or that provides
some information that is potentially useful to perform
the task (e.g., a spatial cue). By contrast, “set relevance”
refers to irrelevant aspects that do not provide any infor-
mation to execute the task but are still related to the cur-
rent task set and, because of this, can influence performance
and brain activity.
In standard memory tasks, foils/competitors typically
share some feature with the memory probe: for example,
belonging to the same study list, the same semantic cate-
gory or same paired-associate (Guerin et al., 2012; Fletcher,
Shallice, Frith, Frackowiak, & Dolan, 1998). This suggests
an analogy between selection mechanisms in memory
tasks and in attention capture paradigms. Indeed, memory
studies of “proactive interference”—which formally ma-
nipulate task set during memory retrieval—showed greater
activation of ventrolateral frontal areas when participants
had to judge nontarget stimuli that were previously/
recently used as target (i.e., high set relevance) versus non-
recent trials (Badre & Wagner, 2005; Jonides, Smith,
Marshuetz, Koeppe, & Reuter-Lorenz, 1998). Within this
framework, the attention–memory correspondence ap-
pears to hold, with the ventral frontoparietal system being
engaged primarily when the task requires selection be-
tween external (attention) or internal (memory) items that
compete because they share some aspect that is relevant
for the current task set.
Task-irrelevant Distractors during
Episodic Memory Retrieval
Nonetheless, it should be noted that in proactive inter-
ference paradigms the nontarget probes are task rele-
vant, that is, participants are required to perform some
memory judgment about these stimuli (Badre & Wagner,
Kwok, Shallice, and Macaluso
2071
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
2005; Badre et al., 2005; Henson, Shallice, Josephs, &
Dolan, 2002; see also Guerin et al., 2012, for an example
concerning “shared” semantic category between target
and distractors during recognition). This type of para-
digm differs substantially from contingent capture para-
digms, where participants are told in advance what
stimuli to judge (targets) and what stimuli to ignore (exog-
enous cues, e.g., Folk et al., 1992; or stimuli at an irrelevant
location, Serences et al., 2005); and hence, the capture pro-
cess relates to fully task-irrelevant, albeit set-relevant, dis-
tractors (see also above).
A closer parallel to the perceptual situation comes
from memory studies that used fully irrelevant distractors
on memory retrieval. In a series of experiments, Wais and
colleagues reported that task-irrelevant visual distractors
reduce retrieval accuracy (Wais, Martin, & Gazzaley, 2012;
Wais, Kim, & Gazzaley, 2011; Wais, Rubens, Boccanfuso,
& Gazzaley, 2010). In these studies, participants were
instructed to direct their full attention to recall the num-
ber of items in previously studied images. The retrieval
conditions manipulated whether participantsʼ eyes were
closed, open and looking at a gray screen, or open and
looking at irrelevant visual stimuli. Behavioral data
showed diminished recollection performance in condi-
tions when the eyes were open and irrelevant visual stim-
uli were presented, compared with both gray screen and
eyes shut conditions. The imaging data associated the
diminished recollection with the disruption of functional
connectivity in a network involving the IFG and the infe-
rior parietal lobule (including TPJ). The causal role of the
IFG was subsequently confirmed in an rTMS study (Wais
et al., 2011). These results demonstrate that irrelevant
visual input can interact with memory retrieval in a
stimulus-driven manner. However, these paradigms did
not tackle the issue of whether/how shared features be-
tween memory probes and distractors affect such inter-
actions (cf. set relevance, above).
Probing the Interplay between Attention and
Memory Using Set Relevance
Here we investigated the interplay between stimulus-
driven attention and episodic retrieval by presenting irrel-
evant distractors during retrieval of temporal information
from memory and, specifically, by manipulating the rela-
tionship between the distractors and the memory probes.
On the basis of a previous paradigm (Kwok, Shallice, &
Macaluso, 2012), we used naturalistic stimuli to identify
behavioral and fMRI indices associated with the retrieval
of temporal information for episodic memory. At encod-
ing, participants were asked to watch a 45-min movie
involving complex storylines and dynamic real life-like
events, which lasted roughly the length of time of the
film. At retrieval, 24 hr later, participants were presented
with two images extracted from the film and were asked
to report which had occurred earlier at presentation. We
previously found that the temporal distance between the
probes at encoding affected retrieval performance, with
short distances yielding slower retrieval than long dis-
tances (Kwok et al., 2012; see also St. Jacques, Rubin,
LaBar, & Cabeza, 2008). We attributed the costs of retriev-
ing short instead of long temporal distances to recon-
structive search processes (cf. Friedman, 1993) and
associated these with activation of the precuneus.
In this study, we modified the retrieval task by present-
ing the task-irrelevant distractors interleaved between the
two probe images (Figure 1A). The distractor pictures
were also extracted from the same movie and had speci-
fic temporal and semantic relationships with the memory
probes (Figure 1B). From a theoretical perspective, exist-
ing accounts of temporal order retrieval postulate either
a “chronological organization” of the memory traces that
emphasizes temporal aspects (Friedman, 1996, 2001) or a
reconstructive “knowledge-based” process that instead
stresses the role of content (“reconstructive theories”;
Friedman, 1993, 2007). Here, we operationalized these
two viewpoints through the experimental factors of “D-I
content” (content relationship: distractor and probes
from same vs. different storylines of the movie) and “D-I
proximity” (temporal relationship: distractor extracted
from a movie time point close to the first vs. the second
memory probe), together with the original factor of tem-
poral distance (“Im1-Im2 distance”: short vs. long dis-
tance between the two memory probes).
Depending on whether the distractor taps into the
temporal or content dimension that is relevant for the
temporal retrieval task (cf. set relevance of the distrac-
tors), we would infer which type of processes (i.e., chron-
ological organization vs. reconstructive-based) better
characterize the temporal order retrieval. In line with
the reconstructive-based account (Kwok et al., 2012),
we predicted that the content relationship between the
distractors and the memory probes would affect activity
in the precuneus more than the temporal relationship (i.e.,
an effect of “content,” rather than “proximity”).
Furthermore, because of the hypothesized link be-
tween attention and memory (cf. above), we anticipated
that the type of distractor would affect activity in the ven-
tral attention network (e.g., Ciaramelli et al., 2010; Wais
et al., 2010). Specifically, if the factor “D-I content” gave
rise to a stronger modulation in the attention network
during retrieval, it would suggest that set relevance is con-
tent related; whereas if the factor “D-I proximity” resulted
in stronger changes, it would suggest that set relevance is
temporal related. These distractor factors may further
interact with the factor of distance, if the distractor-
probes relationship is important only when reconstructive
search processes are engaged (i.e., on short-distance
trials). The results would extend previous proposals about
set-related effects in attention/perception to the domain
of retrieval from episodic memory. Moreover, localization
of brain activity within attention-related areas during
memory retrieval would support recent accounts sug-
gesting a neural overlap between attention selection
2072
Journal of Cognitive Neuroscience
Volume 26, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
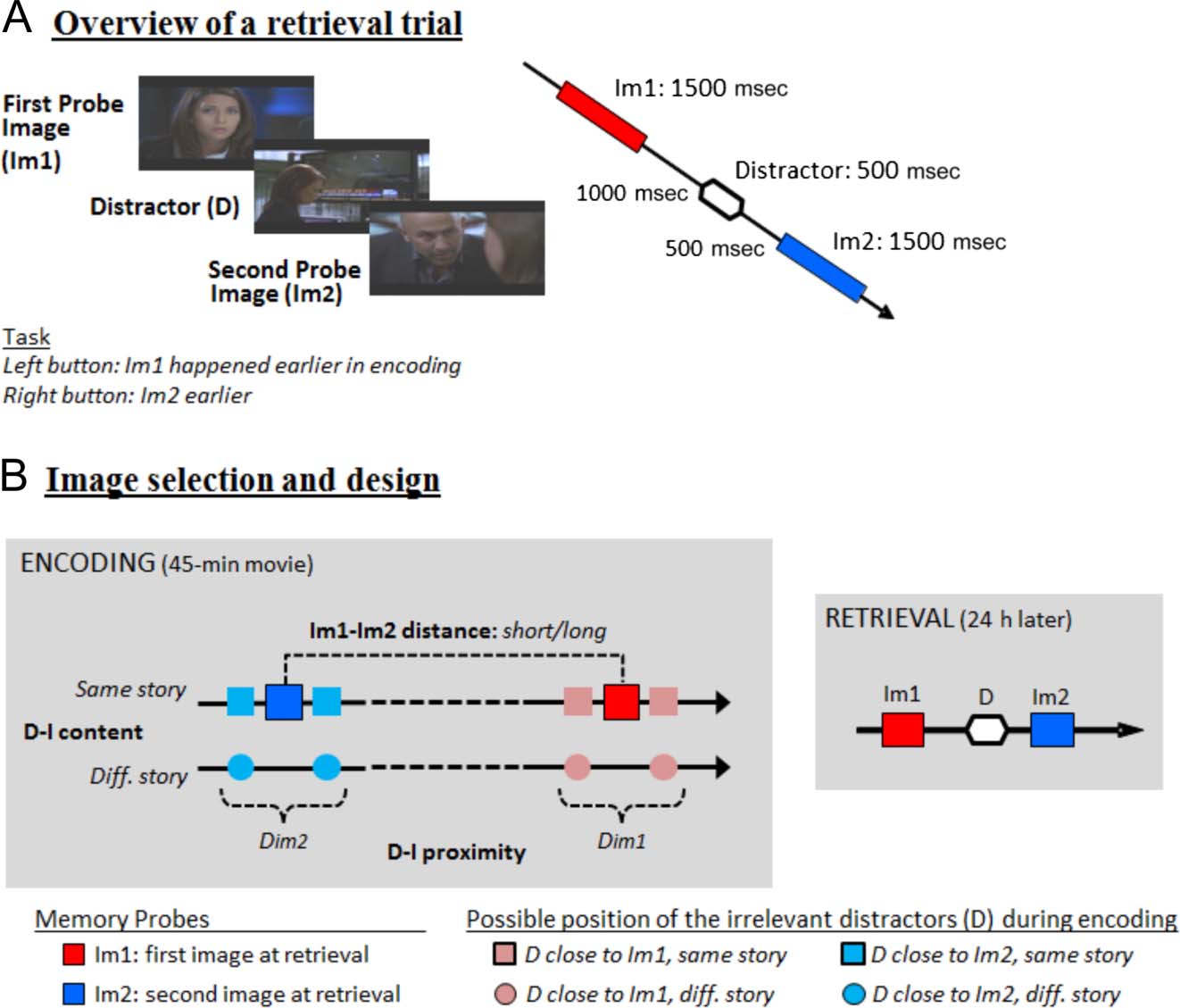
Figure 1. Retrieval task, test
images selection, and
experimental manipulation.
(A) Each retrieval trial consisted
of a triplet of images: First
Probe Image (“Im1”), Distractor
Image (“D”), and Second Probe
Image (“Im2”). The three
images were presented
sequentially. The task required
the participant to judge the
temporal order of the two
probe images and to ignore
the interleaved task-irrelevant
distractor image. (B) The main
2 × 2 × 2 factorial design
manipulated: the temporal
distance between the two probe
images during encoding (Im1-
Im2 distance: short/long), the
content relationship between
distractors and memory probes
during encoding (D-I content:
same/different storyline), and
the temporal relationship
between distractors and probes
(D-I proximity: Dim1/Dim2, with
the distractor extracted from a
time point closer to either the
probe image Im1 or Im2 at
encoding). The memory probes are labeled with respect to the order of presentation during retrieval (i.e., Im1-D-Im2, cf. A), but note that on
half of the trials Im2 occurred before Im1 during encoding (as depicted here, i.e., the dark blue square), and thus, it was the response target for
the order judgment. The control condition using scrambled-images distractors is not depicted here.
in perception and in memory (Cabeza, Ciaramelli, &
Moscovitch, 2012b; see also Nelson, McDermott, &
Petersen, 2012).
see Figure 1B). In addition, the design included a control
condition using scrambled images distractors (Cnt trials),
which did not have any relationship with the memory
probes.
METHODS
Overview
The experimental protocol consisted of two phases, en-
coding and testing, organized across two consecutive
days. On Day 1, participants were asked to watch one
single 45-min episode of a TV series. On Day 2, during
fMRI scanning, participants were tested on a temporal
order retrieval task coupled with different manipulations
of distraction (i.e., temporal/content relationship between
the probes and distracter image). Each retrieval trial
included the presentation of a triplet of static images
extracted from the film: First Probe Image (Im1), Distractor
(D), and Second Probe Image (Im2; see Figure 1A). The
participantʼs task was to choose the probe image that
had happened earlier in the film at encoding, ignoring
the interleaved distractor image. The experimental design
considered the factors of “Im1-Im2 distance” (long/short:
long or short temporal distance between the two task-
relevant probe images), “D-I content” (same/different:
distractor from the same or different storyline as the
probe images) and “D-I proximity” (Dim1/Dim2: distractors
extracted from a time point closer to either Im1 or Im2;
Participants
Fifteen subjects participated in the fMRI experiment
(mean age = 22.6 years, 20–30 years; 10 women). None
of them had seen the TV episode in question. All had
normal or corrected-to-normal visual acuity, and none
of them reported neurological impairments. The study
was approved by the Fondazione Santa Lucia (Scientific
Institute for Research Hospitalization and Health Care)
Independent Ethnics Committee, in accordance with
the Declaration of Helsinki. A different group of 12 sub-
jects (mean age = 25.4 years, 19–31 years; 10 women)
participated in the preliminary behavioral study (see also
Figure 2).
Stimuli and Task
The encoding material was one episode of the American
TV series “24” (Season 6, Disc 2, 11:00–12:00), which con-
tained five concurrent storylines portraying different char-
acters at various locations (Plot A: depiction of the president
and his team in the White House; Plot B: interactions of
Kwok, Shallice, and Macaluso
2073
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
inmates in a detention center; Plot C: happenings in the
office of the Counter Terrorism Unit; Plot D: depiction of
Agent Jack on the move; Plot E: a middleman working for
the terrorists and his girlfriend). The 45-min episode rep-
resents 1 hr of seamlessly timed events; from a temporal
perspective, watching it resonates with seeing “real-world”
events unfolding over time.
Before encoding (Day 1), participants were instructed
to concentrate on the film and memorize as much of it
as possible. They were made aware that their memories
of the film would be tested on the ensuing day, but no
information was provided regarding how and what spe-
cific memories would be targeted. Before retrieval (Day 2),
participants received detailed task instructions with a
screen display example and familiarized themselves with
using a keyboard for making responses. Each retrieval
trial included the presentation of a triplet of static images
extracted from the film (see Figure 1A and section below).
At retrieval, the three images were presented in a
sequential order: First Probe Image (Im1, for 1500 msec),
Distractor (D, for 500 msec), and Second Probe Image
(Im2, for 1500 msec). There was a 1000-msec blank
screen between Im1 and D and a 500-msec blank screen
between D and Im2 (Figure 1A). The task was to choose
the target image that had happened earlier in the film at
encoding, ignoring the interleaved Distractor image. On
half of the trials, Im1 was the response target; in the other
half of the trials, Im2 was the target. Participants were
instructed to respond with a keypress as soon as they
could make a decision after the second probe image
appeared on the screen, but no more than 2.5 sec after
it disappeared. Participants indicated the target stimulus
(either Im1 or Im2) by pressing one of the two keys with
their dominant hand. The stimuli were presented using
Cogent Toolbox (www.vislab.ucl.ac.uk/cogent.php) running
under Matlab 7.4 (The MathWorks, Natick, MA). A total of
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
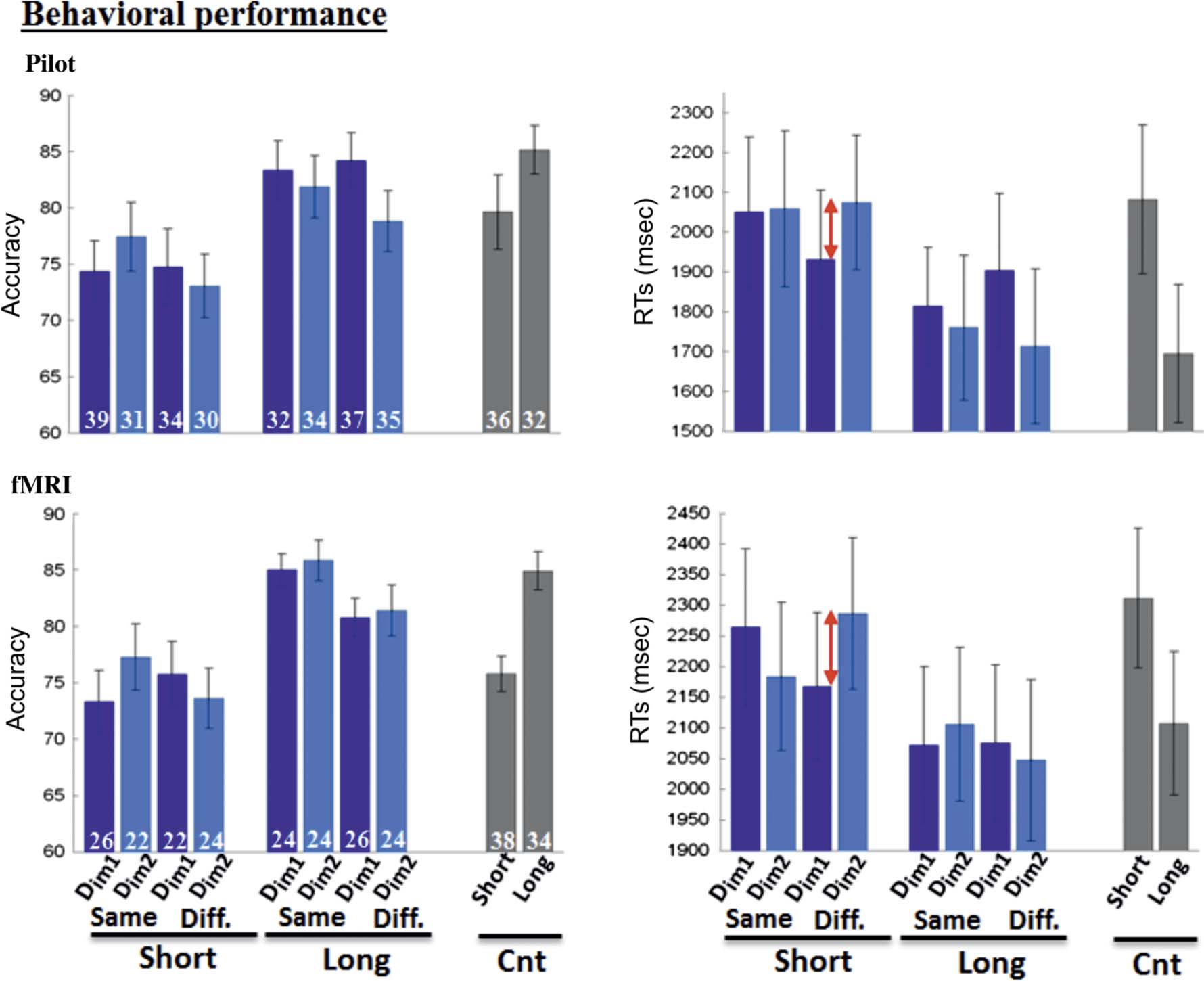
Figure 2. Behavioral performance. Accuracy (% correct) and mean RTs (msec) are plotted according to experimental conditions for the pilot
(top) and fMRI experiments (bottom). Consistently across two experiments, there was a cost of retrieving the temporal order of the two memory
probes on short- compared with long-distance trials (main effect of distance). Both the temporal and content relationships between the
distractor and the probes modulated the RT costs (Distance × Proximity × Content interaction). Specifically, distractors that at encoding were
temporally close to the first probe image (i.e., Dim1 distractors) reduced the RT costs of order judgment on short-distance trials and did so only when
they were not content related with the memory probes (indicated by red arrows contrasting “[Dim1 vs. Dim2] short/diff.” conditions on right panels).
The values in white on the left panels represent the number of trials for each of the conditions (see Methods section, for the procedure used
to assign trials to each condition). Diff., different storyline between the distractor and probe images; Cnt, control condition. Error bars: SEM.
2074
Journal of Cognitive Neuroscience
Volume 26, Number 9
264 trials were presented in four separate fMRI runs. The
ITIs were jittered and in the range of 3–5 sec, uniformly
distributed.
Extraction of the Images
Memory probes and distractor images were selected
based on a film content analysis. The film was segmented
into 103 epochs, such that each epoch contained only
one scenario/setting. Twenty-one epochs were discarded
because they contained unnaturalistic images (e.g., multi-
ple windows/scenes on a single frame), leaving 82 epochs
for the image extraction. Each epoch was assigned to one
of the five storylines. This identified 26 “clusters of
epochs,” with each cluster including a number of epochs
of the same storyline (range = 1–11) and with the end of
each cluster representing the transition from one story-
line to another. For each “epoch” (“E,” from which we
would extract a memory probe image), we created one
list including all possible epochs from where to extract
the second memory probe (Im_list) and four lists to
extract the distractor images (SamePre, SamePost, DiffPre,
DiffPost). The Im_list included all epochs of the same
storyline as epoch “E,” but belonging to a different cluster
with at least one in-between cluster of the same storyline:
for example, if the first probe belonged to an epoch E of
the first cluster of storyline A, the Im_list included all
epochs of storyline A with cluster n ≥ 3. This ensured
that the two probe images (Im1 and Im2) always belonged
to the same plot/storyline, yet there was at least one “in-
between” cluster. The Im_list included 488 possible
couplings. For each epoch E, the two same-storyline dis-
tractor lists (SamePre, SamePost) included all the epochs
of the previous or next same-storyline cluster, whereas
the different-storyline distractor lists (DiffPre, DiffPost)
included epochs belonging to a previous/following cluster
of a different storyline. This generated the following num-
bers of probe/distractor epoch couplings: SamePre = 179,
SamePost = 132, DiffPre = 306, DiffPost = 307.
For each of the 488 couplings between the two probe
images (Im_list), we selected a distractor epoch from one
of the four distractor lists or assigned the pair of probes
to the control condition. With this, we obtained a set of
triplets including two epochs for the probes and one epoch
for distractor, plus a set of probesʼ pairs for the control
condition. Then we sampled a subset of 192 triplets ensur-
ing that this final list included 96 triplets with probes and
distractor from the same storyline (48 triplets with distrac-
tor from SamePre list, 48 with distractor from SamePost list)
and 96 triplets with different storyline distractors (48 with
DiffPre and 48 with DiffPost distractors). For the control
condition, we retained 72 probesʼ pairs. This constituted
the final list of epoch associations that was employed to
extract the single frames to be used in the 264 trials.
The single frames (probes and distractors) were
extracted randomly from the selected epochs and visually
inspected to avoid any unsuitable image (e.g., a frame
including a close-up view of a wall). The distractors for
the control condition were chosen from a different epi-
sode of the same movie series. Each frame was divided in
1170 squares, which were randomly rearranged to gen-
erate the scrambled-distractor images. At this stage, we
randomly decided which of the two memory probes
would be presented first during retrieval (i.e., the Im1).
For half of the trials, Im1 was the probe image that
occurred earlier in the movie, whereas in the other half
the probe that occurred later was used as Im1. The assign-
ment of Im1/Im2 was balanced for same/different story-
line, but not for Pre/Post distractorsʼ lists.
Assignment of the Images to the
Retrieval Conditions
We assigned the 264 triplets (cf. above) to the eight con-
ditions given by the crossing of the factors of distance,
storyline, and proximity (192 trials), plus the two control
conditions (72 trials). Trials were assigned to the “short
distance” conditions when the temporal distance be-
tween the two probes was shorter than the median dis-
tance of all trials (21.2 min); otherwise, the trial was
assigned to a “long distance” condition. For the 192 trials
belonging to the main 2 × 2 × 2 design, the storyline
condition depended on whether the distractor was se-
lected from the “same” or “different” storyline as the
two memory probes (cf. Same vs. Diff distractor lists).
For the proximity factor, we considered the distance be-
tween the distractor and the two memory probes (Im1
and Im2). When the distractor was temporally closer to
Im1 than Im2, the trial was assigned to a Dim1 condition,
otherwise to a Dim2 condition. The final number of trials
for each of the conditions is reported in Figure 2 (range
for the eight main conditions = 22–26 trials).
With these procedures, we minimized idiosyncrasy in
the stimuli and in the trials assignment to the different
conditions. However, we acknowledge that with such
complex and naturalistic material we cannot exclude
the possibility that conditions differed also with respect
of some other unexplored factor.
fMRI Data Acquisition and Preprocessing
All images were acquired with a Siemens Allegra (Siemens
Medical Systems, Erlangen, Germany) 3T scanner
equipped for EPI. A quadrature volume head coil was used
for radio-frequency transmission and reception. Head
movement was minimized by mild restraint and cushion-
ing. Thirty-two slices of fMRI images were acquired using
BOLD imaging (3 × 3 mm in-plane, 2.5 mm thick, 50%
distance factor, repetition time = 2.08 sec, echo time =
30 msec, flip angle = 70°, field of view = 192 mm, acquisi-
tion order = continuous, ascending), covering the entirety
of the cortex.
Data preprocessing was performed with SPM8 ( Well-
come Department of Cognitive Neurology, London, United
Kwok, Shallice, and Macaluso
2075
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Kingdom) as implemented on MATLAB 7.4. A total of
1548 fMRI volumes for each participant were acquired
in four separate fMRI runs. After having discarded the
first four volumes in each run, images were realigned
to correct for head movements. Slice acquisition delays
were corrected using the middle slice as a reference.
Images were then normalized to the MNI EPI template,
resampled to 3 mm isotropic voxel-size, and spatially
smoothed using an isotropic Gaussian kernel of 8 mm
FWHM. Time series at each voxel were high-pass filtered
at 128 sec and prewhitened by means of autoregressive
model AR(1).
nonindependent error terms for the repeated measures
(Friston et al., 2002).
We tested the overall effect of “Im1-Im2 distance”
comparing “short vs. long” trials irrespective of distractor
condition (cf. St. Jacques et al., 2008), the main effects of
distractor “D-I proximity” and “D-I content,” and the inter-
actions between the three factors. With this model, we
also compared the eight main conditions versus the low-
level baselines (i.e., trials with scrambled distractors).
For all comparisons, the statistical threshold was set
to p-FWE < .05, whole brain-corrected at cluster level
(cluster size estimated at p-unc. = .005).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
fMRI Data Analysis
The aim of the functional imaging analyses was to identify
brain areas showing differential responses as a function
of the temporal/content relationship between the dis-
tractors and the memory probes. Accordingly, the fMRI
analyses considered eight types of trials given by the
crossing of the factors of “Im1-Im2 distance” (short/long,
temporal distance between the probes), “D-I proximity”
(Dim1/Dim2, distractor probe temporal relationship), and
“D-I content” (same/different, distractor probe content
relationship; see also Figure 1B).
The first-level multiple regression models (i.e., single-
subject analyses) included eight main conditions of interest
(“Im1-Im2 distance” × “D-I proximity” × “D-I content”),
plus two control conditions (scrambled-distractor, with
short or long temporal distance between the two probe
images). In addition, errors and movement parameters
were included as effect of no interest. Each trial was mod-
eled as a short epoch time-locked to the onset of the
first probe image (duration = 5 sec, i.e., till the offset of
the second probe image), convolved with the canonical
hemodynamic response function in SPM8.
The parameter estimates of the conditions of interest
were then assessed at the second-level for random effect
statistical inference (Penny & Holmes, 2004). The analysis
consisted of a within-subject ANOVA considering the
10 conditions described above. To account for any
potential difference in task difficulty, condition-specific
RTs for each participant were entered as a covariate.
Accordingly, here any significant difference between
conditions cannot be explained by RT differences (e.g.,
short temporal distance trials taking longer than long-
distance trials; see also Kwok et al., 2012). Moreover, we
also estimated a model that did not include the RTs as a
covariate and replicated all the main findings (i.e., main
effect of “distance” and the “Distance × Proximity” inter-
actions significant after correction for multiple com-
parisons and “Distance × Content × Proximity” at
uncorrected level), again indicating that RT differences
between conditions did not influence our results. Correc-
tion for nonsphericity was used to account for possible
differences in error variance across conditions and any
RESULTS
Behavioral Results
In a preliminary behavioral study, we found that present-
ing task-irrelevant distractors during retrieval affected the
participantsʼ performance on our temporal order judg-
ment task. ANOVAs with the factors of “Im1-Im2 dis-
tance,” “D-I content,” and “D-I proximity” revealed the
expected main effect of temporal distance between the
two memory probes: participants were faster, F(1, 11) =
45.8, p < .001, and more accurate, F(1, 11) = 27.1, p <
.001, on “long” compared with “short” trials (see Kwok
et al., 2012; St. Jacques et al., 2008). Both the accuracy
data, F(1, 11) = 5.1, p < .046, and RTs, F(1, 11) = 6.9,
p < .024, showed an interaction between proximity and
distance. The RT data showed a significant three-way
interaction between distance, content, and proximity,
highlighting the joint influence of both temporal and
content relationships between distractors and memory
probes on the temporal order judgment, F(1, 11) =
6.4, p < .029. Figure 2 shows that this complex behav-
ioral pattern arose primarily because Dim1 distractors (i.e.,
those temporally related to the first probe image) speeded
up retrieval times compared with Dim2 distractors, speci-
fically when the Im1-Im2 distance was short and the dis-
tractor was extracted from a different storyline than the
probe images (see red arrows in the panels on the right).
In other words, the Dim1 distractors reduced the costs of
performing the order judgment on short-distance trials,
but did so only when they were not semantically/content
related with the memory probes.
The behavioral data collected during fMRI confirmed
the main findings of the preliminary pilot study. The
main effect of distance was highly significant both in
terms of accuracy, F(1, 14) = 17.6, p < .002, and RTs,
F(1, 14) = 23.2, p < .001, and the RT data replicated
the three-way interaction between distance, content,
and proximity, F(1, 14) = 8.9, p < .011. The RT data also
showed a Proximity × Storyline interaction, F(1, 14) =
5.6, p < .034, that was not found in the preliminary study.
Tukeyʼs HSD post hoc tests on the RTs confirmed that
Dim1 distractors speeded up retrieval compared with
Dim2 distractors on short-distance, different-storyline
2076
Journal of Cognitive Neuroscience
Volume 26, Number 9
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
trials ( p < .017), whereas this difference was not signifi-
cant for any of the other distance/storyline combinations
(all p > .1).
In an additional set of analyses, we also verified whether
the correspondence between the order of presentation of
the images during encoding and retrieval had any effect
on memory performance. First we considered only the
task-relevant memory probes (Im1 and Im2), comparing
all trials when Im1 was presented before Im2 at encoding
(i.e., “congruent order” between encoding and retrieval)
versus Im2 before Im1 (i.e., “incongruent order”). In the
preliminary experiment, we found slower RTs when
Im1 was first (1937 vs. 1866 msec; t(11) = 2.9, p < .015),
but this was not fully significant during fMRI (2187 vs.
2138 msec; t(14) = 1.7, p < .122). Moreover, the accu-
racy data indicated some speed–accuracy trade-off, with
greater accuracy on “congruent order” trials (prelim-
inary study: 80.8 vs. 78.6%; fMRI: 80.7 vs. 78.4%), but
again the statistical evidence was inconsistent across the
two data sets (preliminary study: p < .067; fMRI: p <
.229). Next, we considered also the distractor images and
compared trials with a “fully congruent image order”
(sequence of events at encoding: Im1-D-Im2) versus all
the other “incongruent” sequences (i.e., Im2 before Im1,
D-Im1-Im2, or Im1-Im2-D). This revealed that participants
tended to be slower and more accurate on “congruent”
than “incongruent” trials, but again the statistics was not
reliable across the two data sets.
In summary, consistently in two independent data
sets, the behavioral data showed that (A) participants
were slower and less accurate to judge short- compared
with long-distance trials and (B) the costs on short-
distance trials were reduced by task-irrelevant distractors
that—at encoding—were temporally closer to the first
than the second probe image (i.e., the Dim1 distractors),
especially when the distractors belonged to a different
storyline than the two probe images, that is, three-way
interaction. These results demonstrate that, although
the distractors were fully task irrelevant, some informa-
tion about this to-be-disregarded stimulus continued to
influence participantsʼ evaluation of the Im1/Im2 order
(cf. “retrieval inhibition” effects in Bjork & Bjork, 1996).
Whether the irrelevant information could affect memory
performance depended on both the temporal relation-
ship (“D-I proximity”) and the content relationship (“D-I
content”) between the task-irrelevant distractor and the
probe images. By the use of functional imaging, we then
sought to determine whether this multifaceted behavioral
outcome maps onto the activity of one specific brain area/
network or can be separately mapped to different areas/
networks.
(Dim1/Dim2), and the interactions between these factors.
The main aim of the fMRI analyses was to map the effects
of temporal proximity and content relationship between
the perceptual distractors and memory probes. Specifi-
cally, we asked whether these would include areas where
previous studies reported an effect of temporal distance
(i.e., the precuneus; see Kwok et al., 2012; St. Jacques
et al., 2008) and/or areas previously associated with
filtering/selection in stimulus-driven attention and mem-
ory (i.e., regions in ventrolateral frontal and parietal cor-
tices; e.g., Cabeza, Ciaramelli, & Moscovitch, 2012a;
Ciaramelli et al., 2010; Badre & Wagner, 2007; Corbetta
& Shulman, 2002).
The Role of the Precuneus during
Temporal Order Retrieval
First, we sought to replicate our previous findings and
confirm greater activation of the precuneus when com-
paring trials including “short > long” temporal distance
between the two probe images (Im1 and Im2). Table 1
Table 1. Main Effects of Temporal Distance between Two
Probe Images and of Distractors Content
Cluster
Voxel
Brain Region
k
p-corr.
Z
x y z
Main Effect of Temporal Distance
Short > long
Precuneus
2457 < .001 5.24 0 −66 53 Medial occipital cortex R 4.85 6 −96 −1 Long > short
Superior temporal gyrus L
221 < .046 4.04 −51 3 −4 Main Effect of Content Same > different
Superior parietal gyrus L
479 < .001 4.92 −24 −63 53
Precuneus
4.25
6 −51 56
Medial occipital cortex R
702 < .001 4.84
27 −63 17
Medial occipital cortex L
Middle occipital gyrus R
Medial occipital cortex
Middle frontal gyrus L
Inferior frontal gyrus L
4.27 −36 −66 20
3.61
3.99
51 −75 11
12 −93 23
225
120
.001
.017
4.40 −42 24 14
4.43 −36 3 53
fMRI Results
The fMRI analyses considered the overall effect of “Im1-
Im2 distance” (short/ long), the main effects of “D-I
content” (same/different storyline) and “D-I proximity”
Statistical thresholds set to p-FWE = .05, whole brain-corrected at clus-
ter level (cluster size estimated at p-unc. = .005, but for the main effect
of “same > different storyline” for which the cluster size was estimated
at p-unc. = .001, see main text). L/R = left/right hemisphere; x y z =
coordinates in the standard MNI space of the activation peaks in the
clusters; k = number of voxels in each cluster.
Kwok, Shallice, and Macaluso
2077
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
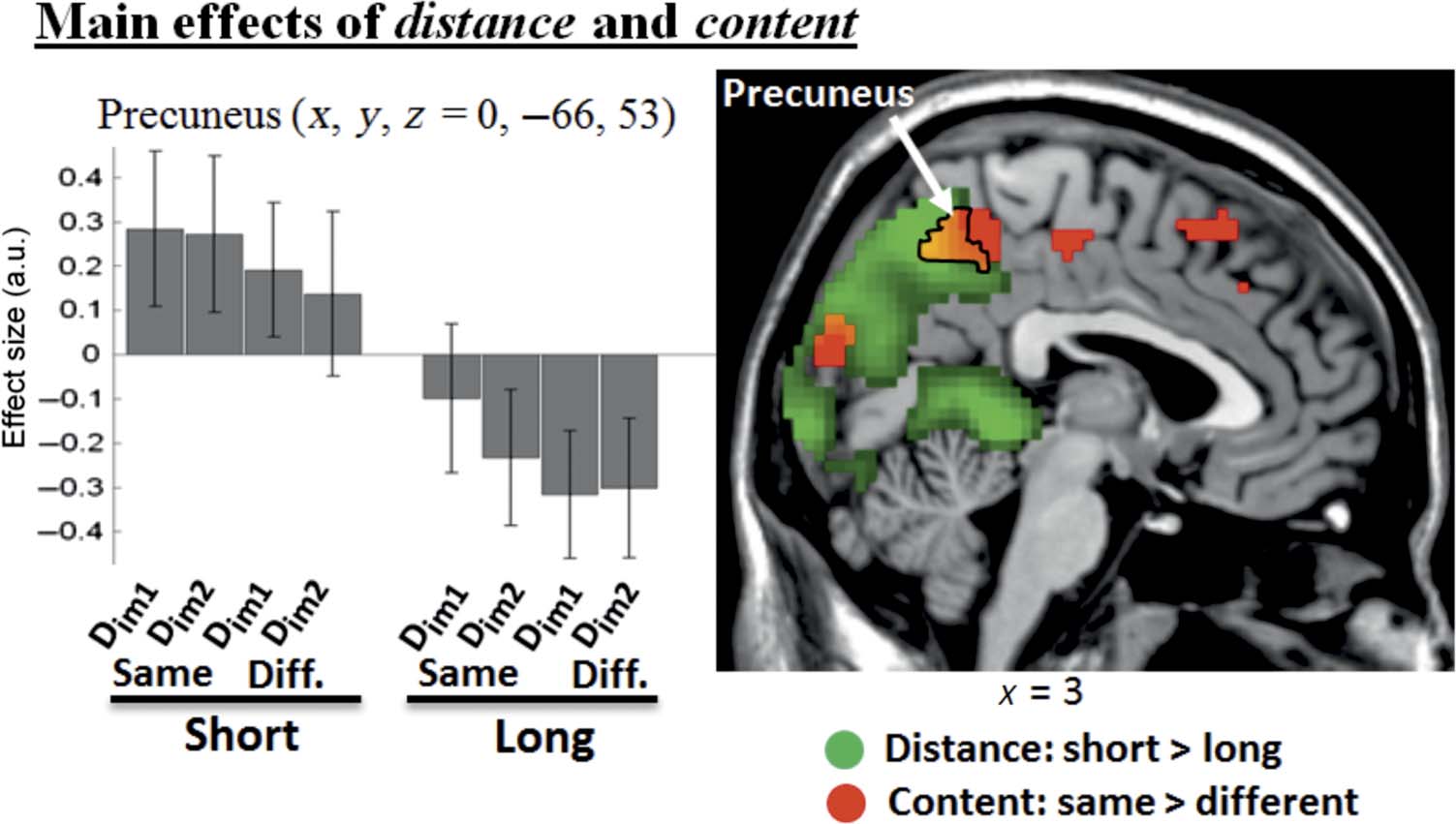
Figure 3. Main effects of
distance and content activate
common area in medial parietal
cortex. The “short > long
distance” comparison revealed
activation of the precuneus
(shown in green). The same
area in the precuneus also
showed an effect of content
(“same > different storyline,”
shown in red), with greater
activation for trials including a
task-irrelevant distractor
belonging to the same storyline
as the two memory probes
than otherwise. The signal plot
shows activity at a local maxima
of the distance effect, located
within the region also exhibiting
the main effect of content. The
activation clusters are displayed
at a threshold of p-FWE = .05,
whole brain-corrected at cluster
level (cluster size estimated at
p-unc. = .005). Effect sizes are
mean adjusted (sum to zero) and
expressed in arbitrary units
(a.u. ± 90% CI). Diff., different
storyline between a distractor
and probe images.
lists the regions activated in this contrast. As expected,
the main cluster of activation was found in the precuneus
(Figure 3, in green), with greater BOLD response for
short than long trials, irrespective of distractor conditions
(see signal plots in Figure 3). The reverse comparison
(long > short) highlighted activation in left anterior
superior temporal gyrus (Table 1).
We then turned to test for changes of brain activity as a
function of the relationship between the type of distrac-
tor and the memory probes. First, we considered the
effect of “content” (D-I content) comparing trials includ-
ing irrelevant distractors extracted from the same versus
different storylines. The contrast of “same > different”
storyline revealed activation of the precuneus (Figure 3,
in red). This was part of a widespread cluster that in-
cluded adjacent areas in the superior parietal gyrus, but
also anterior regions in the frontal cortex. Because of this,
we retested the same contrast but using a more stringent
voxel-level cluster-defining threshold ( p-unc. = .001,
rather than .005) that enabled us to better localize the
activated areas. The activation of the precuneus and the
left superior parietal cortex remained significant, and we
localized other activation clusters in the left superior and
the left inferior frontal gyri, plus the occipital cortex (see
Table 1). The reverse contrast of “different > same” did
not reveal any significant activation. Accordingly, the
precuneus showed both an effect of the temporal dis-
tance between the two task-relevant probes (“Im1-Im2
distance”: short > long) and an effect of the content
relationship between the memory probes and the task-
irrelevant distractors (“D-I content”: same > different
storyline; cf. also Figure 3). Nonetheless, no interaction
was found between these two factors. We then also
examined the effect of the temporal proximity between
the distractors and the memory probes (“D-I proximity”).
Contrasts comparing trials including a distractor extracted
from a time point close to Im1 vs. Im2 (Dim1 > Dim2) and
vice versa (Dim2 > Dim1) did not reveal any significant
activation.
Attentional Capture in Ventral Parietal Cortex
We further assessed the influence of irrelevant dis-
tractors on temporal order retrieval by testing for the
“Proximity × Distance” and the “Content × Distance”
interactions. With these we aimed to identify any temporal-
or content-related effect that was specific for the retrieval
of either short- or long-distance trials, that is, the inter-
actions between distractor-related set relevance and
probe-related reconstructive processes that we associated
specifically with short-distance trials (cf. Introduction;
Kwok et al., 2012).
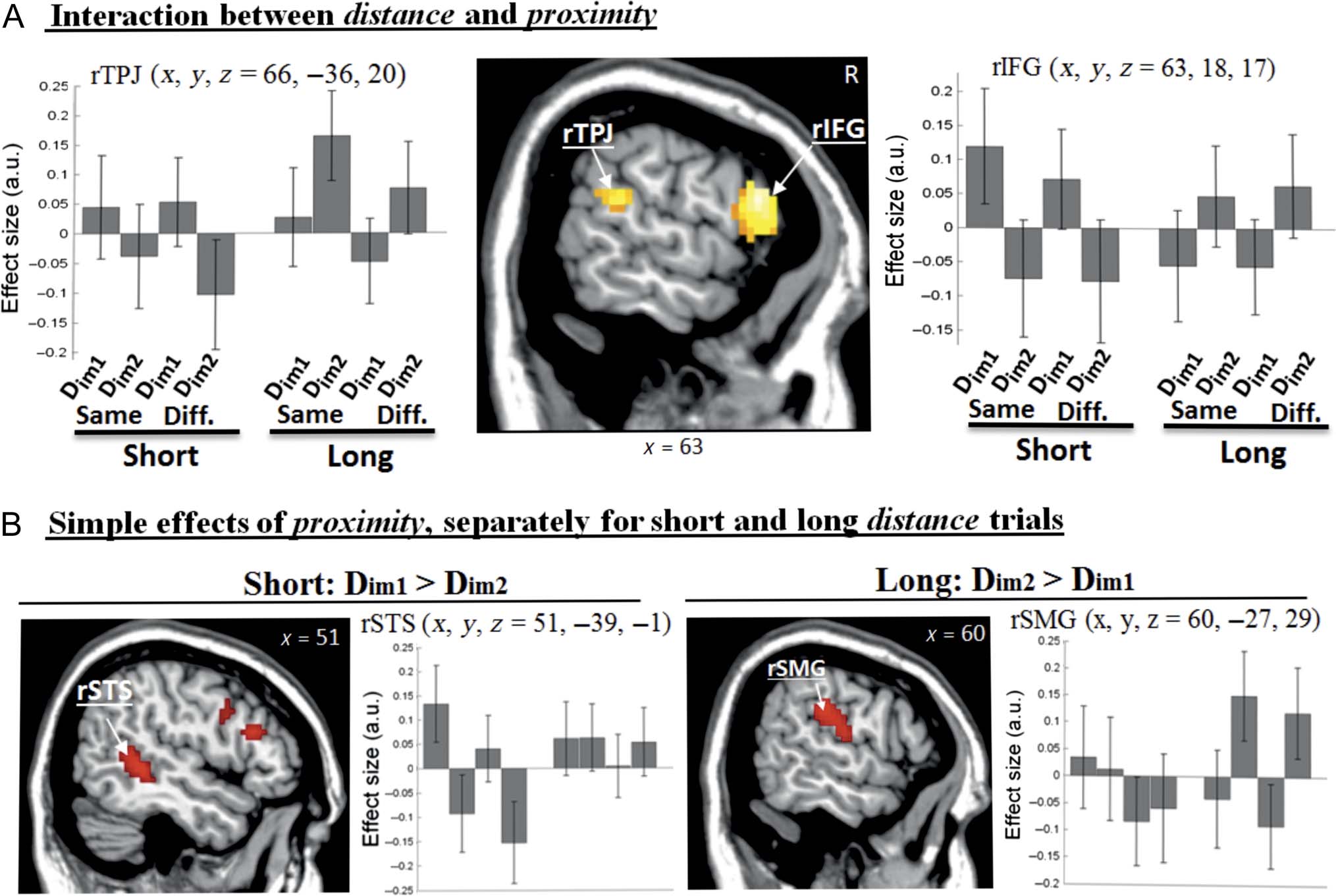
The contrast testing for the interaction between
“proximity” and “distance” (i.e., [Dim1 − Dim2] short >
[Dim1 − Dim2] long) revealed three distinct clusters of
activation: the rIFG, the rTPJ, plus the left inferior occipi-
tal gyrus (see Figure 4A and Table 2). The rTPJ cluster
extended ventrally to the STS, and the signal plots of the
different peaks within this cluster appeared to show differ-
ent patterns of activation (note that several conditions
2078
Journal of Cognitive Neuroscience
Volume 26, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
contribute to the proximity by distance interaction). We
explored this further by comparing separately the sim-
ple effects of [Dim1 − Dim2] in “short” trials and [Dim2 −
Dim1] in “long” trials. This revealed that the Dim1 distractors
in short trials activated the right STS (rSTS), whereas the
Dim2 distractors in long trials activated the right supra-
marginal gyrus (rSMG; see Figure 4B). The reverse con-
trast ([Dim1 − Dim2] long > [Dim1 − Dim2] short) and
interaction between “proximity” and “content” did not
give rise to any significant activation.
Additional Analyses Regarding Possible Correspondences
between fMRI and Behavior
First, in parallel with the three-way interaction observed
in the behavioral data, the highest-order interaction
(Distance × Content × Proximity) was associated with
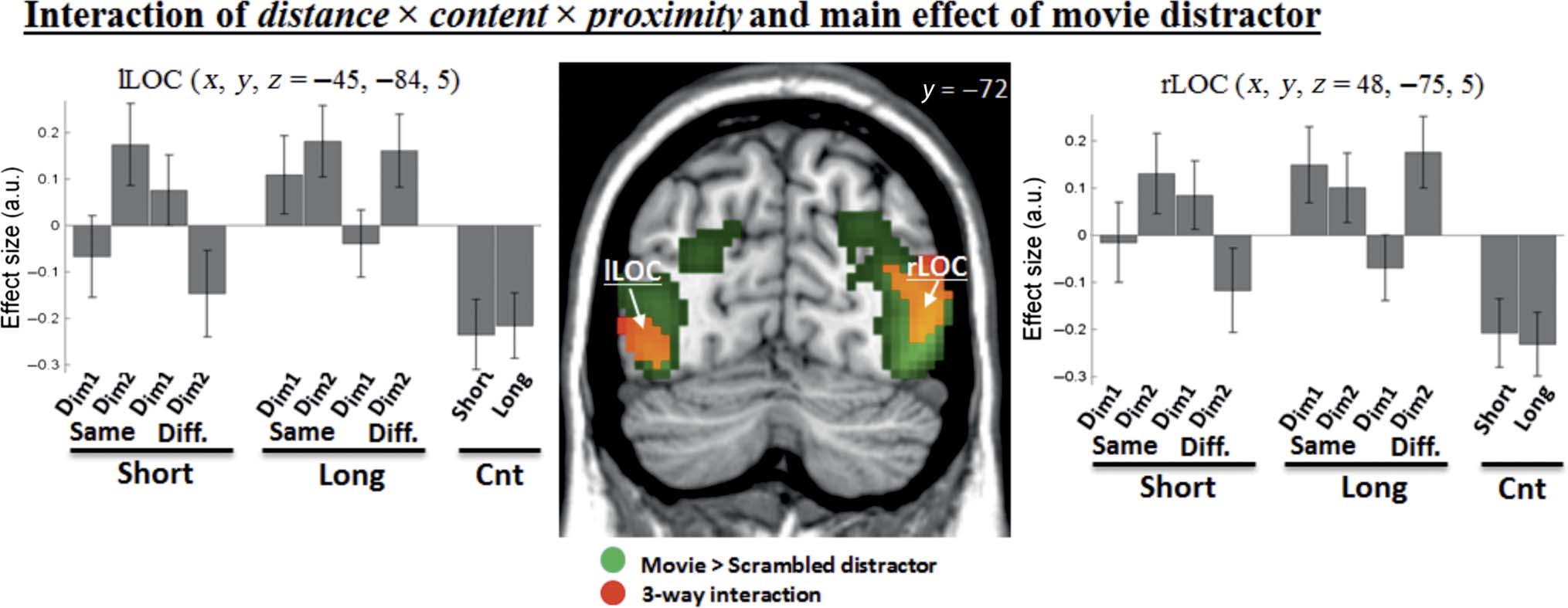
changes of activity in the posterior occipital regions. Spe-
cifically, the contrast [(Dim1 − Dim2) short > (Dim1 −
Dim2) long]DIFFERENT > [(Dim1 − Dim2) short > (Dim1 −
Dim2) long]SAME revealed activation of the middle occipi-
tal gyrus, bilaterally (see Figure 5, in red, and Table 2).
These regions showed a complex pattern of activity,
including larger activation for “Dim1 vs. Dim2 distractors”
in short-distance, different-storyline trials (cf. “bar 3 >
bar 4,” signals plots in Figure 5). The reverse contrast
[(Dim1 − Dim2) short > (Dim1 − Dim2) long]SAME >
[(Dim1 − Dim2) short > (Dim1 − Dim2) long]DIFFERENT
did not reveal any significant effect.
Second, we assessed whether performance on task
covaried with the fMRI responses. For this, we made
use of the RT covariate that was initially included in our
main statistical model to remove any potential RT con-
founds (see also Methods section). The effect of the RT
covariate revealed a positive covariation between the
BOLD signal and RTs in the ventrolateral temporal cortex,
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 4. Interaction between distance and proximity, and the simple effects of proximity separately in “short”- and “long”-distance trials.
(A) Clusters of activation and signal plots for the “Distance × Proximity” interaction. Activation clusters are displayed at a threshold p-FWE = .05,
whole brain-corrected at cluster level (cluster size estimated at p-unc. = .005). The section is taken through the main peak of the cluster in the
rIFG and the signal plot refers to the corresponding voxel. The main peak of the posterior cluster was located more medially and ventrally in the rSTS
(cf. also B) and the signal plot in this panel refers to a secondary peak (rTPJ, see also Table 2). (B) Additional tests assessing separately the
simple effects of [Dim1 − Dim2] in “short” trials and [Dim2 − Dim1] in “long” trials showed distinct effects in posterior regions in the rSTS
(x y z = 51 −39 −1, Z = 4.02, p-unc. < .001; left) and the rSMG (x y z = 60 −27 29, Z = 3.76, p-unc. < .001; right). The rSTS cluster overlapped
substantially with the main interaction cluster, whereas the overlap between the rSMG and the interaction clusters was seen only when lowering
the statistical thresholds ( p-unc. = .005). Effect sizes are mean adjusted (sum to zero) and expressed in arbitrary units (a.u. ± 90% CI). Diff.,
different storyline between a distractor and probe images.
Kwok, Shallice, and Macaluso
2079
Table 2. Interactions among Temporal Distance, Content,
and Proximity
Brain Region
k
p-corr.
Z
x y z
Cluster
Voxel
cuneus and the right ventral attention system that, we
suggest, jointly contribute to the final behavioral out-
come on task.
Distance × Proximity
DISCUSSION
Inferior frontal gyrus R
351
.006
4.18
63 18 17
Superior temporal sulcus R 385
.003
4.22
Lateral occipital cortex R
Temporoparietal junction R
3.70
3.64
45 −42 5
33 −75 17
66 −36 20
Inferior occipital gyrus L
415
.002
4.30 −30 −96 −4
Distance × Content × Proximity
Middle occipital gyrus R
Middle occipital gyrus L
275
198
.018
4.43
48 −75 5
.069
3.95 −45 −84 5
Statistical thresholds set to p-FWE = .05, whole brain-corrected at clus-
ter level, cluster size estimated at p-unc. = .005. L/R = left/right hemi-
sphere; x y z = coordinates in the standard MNI space of activation
peaks in the clusters; k = number of voxels in each cluster.
bilaterally (right hemisphere: x y z = 74 −36 −10, Z =
5.17, p-FWE-corr. < .001; left hemisphere: x y z = −57
−21 −22, Z = 4.37, p-FWE-corr. < .049). In these regions,
activity increased with increasing RTs suggesting a pos-
sible link between these areas and performance on task.
Because these areas did not show any condition-specific
effect, we will focus our discussion primarily on the pre-
We investigated the interplay between stimulus-driven
attention and memory retrieval by presenting task-irrelevant
visual distractors during retrieval of temporal information
about complex events. Behaviorally, trials where there
was only a short temporal distance between the two mem-
ory probes incurred a retrieval cost on the order judgment
task (lower accuracy and longer RTs). The RT costs were
modulated by both temporal and content relationships be-
tween the irrelevant distractors and the memory probes.
Specifically, when distractors were closer in time to the first
than the second probe image (i.e., the Dim1 distractors),
these costs were significantly reduced. This effect of “dis-
traction” on retrieval performance took place only when
the irrelevant distractors were semantically less related to
the task-relevant probes (i.e., different storyline trials).
The fMRI results revealed distinct effects of temporal
versus content relationship between the distractors and
the memory probes. The temporal relationship was
found to affect activity in the right ventral attention net-
work (rTPJ-rIFG), dependent on both how temporally
close the distractor was to the first probe image and
the temporal separation between the two probe images.
Within this system, the rSTS was found to be activated
when the distractor was close to the first probe image
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 5. Interaction of Distance × Content × Proximity and main effect of movie distractor. Clusters of activation and signal plots for the
lateral occipital cortex showing a significant “Distance × Content × Proximity” interaction (shown in red). The pattern of brain activity in
these regions was modulated in correspondence with the three-way interaction observed in the behavioral measures. The figure also shows the
results of the comparison between the “movie distractor” vs. “scrambled-images distractors” (shown in green). This comparison showed
bilateral activation of occipital visual areas, likely to reflect the overall increase of meaningful visual input (objects/people) provided by the
3 vs. 2 visual images presented in movie distractor vs. scrambled-distractor trials. All activation clusters are displayed at a threshold
p-FWE = .05, whole brain-corrected at cluster level (cluster size estimated at p-unc. = .005). Effect sizes are mean adjusted (sum to zero)
and expressed in arbitrary units (a.u. ± 90% CI). Cnt, control condition (scrambled-images distractor); l/r LOC, left/right lateral occipital cortex.
2080
Journal of Cognitive Neuroscience
Volume 26, Number 9
(i.e., Dim1 > Dim2) in short-distance trials, whereas the
rSMG showed an effect of Dim2 > Dim1 in long trials.
By contrast, the content relationship between distrac-
tors and memory probes was found to affect activity in
the precuneus. This region showed increased activation
when the trials included distractors extracted from the
same storyline as the probe images and also showed an
overall effect of temporal distance, with greater activation
for trials with short compared with long distances be-
tween the two memory probes. Finally, temporal and
content relationships jointly modulated activity in the
lateral occipital cortex, where the pattern of BOLD signal
exhibited some correspondence with the RT data.
These results demonstrate that task-irrelevant distrac-
tors can influence memory retrieval and that this crucially
depends on both the temporal and content relationships
between the distractors and the memory probes, thus
highlighting the relevance of the current task set. The
interaction between perceptual distractors and memory
probes in rTPJ and rIFG suggests a link between selec-
tion/filtering in the attention and memory domains,
whereas the response pattern of the precuneus impli-
cates this region in memory search within knowledge/
content-based hierarchies (cf. “reconstructive theories,”
Friedman, 1993, 2007). We discuss these points below.
Contingent Capture on a Memory
Representation Level
Linking our current paradigm—which crossed perceptual
(distractors) and memory (temporal retrieval) factors—to
previous attention research, we predicted that specific
probe–distractor combinations would lead to a contingent
capture of attention on a memory representation level (see
also Ciaramelli et al., 2010; Wagner et al., 2005). Our be-
havioral measures indicate that the temporal- and content-
related relationship between distractors and memory
probes affected the retrieval performance in a complex
manner (cf. a three-way interaction). Specifically, on seeing
the first probe image (Im1), we assume that participants
lodged themselves in a mode of retrieving information
from a specific time point within their memory of the
encoded stimuli (i.e., the top–down goal). In Dim1 trials,
the temporal closeness of the distractor to Im1 generated
a contingency between the current top–down goal and
the task-irrelevant distractor, that is, the time point of pre-
sentation of the distractor during encoding matching the
currently relevant Im1 time point. We suggest that this
correspondence helps to capture or anchor the partici-
pantʼs attention toward that point in time in the memory
representation (i.e., the “Im1-D” moment in the movie).
This would in turn facilitate the temporal order judgment
when the forthcoming second probe image (Im2) is near—
at presentation—this “Im1-D” anchor (i.e., the short-
distance trials). By virtue of these temporal relationships,
attention capture by the Dim1 distractors led to faster
responses in short-distance trials.
A somewhat related account of this temporal effect
concerns the classical memory finding that the recall of an
item is facilitated by the presentation of another item that
occurred close in time during encoding (temporal congru-
ity effect). Howard and Kahana (2002) proposed a temporal
context model postulating that the encoding of new items
dynamically changes the representation of the current tem-
poral context. When an item is retrieved at test, this reinstates
the temporal context associated with that item during en-
coding. As the temporal context will include several neigh-
boring items, the contextual model accounts for the temporal
contiguity effect (Sederberg, Howard, & Kahana, 2008;
Howard & Kahana, 2002). In the current paradigm, the pre-
sentation of the first probe image (Im1) would reactivate a
specific temporal context associated with the encoding
phase. In Dim1 trials, the distractor would coincide with the
reactivated context, which would further strengthen the reac-
tivation and give rise to the improved retrieval performance.
fMRI Correlates of Contingent Capture in
Episodic Memory
In accord with the attention capture interpretation, the
fMRI analyses found that the Dim1 versus Dim2 distractor
conditions modulated activity in the right ventral atten-
tion network (rTPJ-rIFG). Our main analysis showed a
Distance × Proximity interaction, which was then
further differentiated into an effect of “Dim1 > Dim2 on
short trials” in rSTS and of “Dim2 > Dim1 on long trials”
in rSMG (see Figure 3B). Our finding of an interaction
between “D-I proximity” and “Im1-Im2 distance,” rather
than just a main effect of proximity (Dim1 > Dim2 or Dim2 >
Dim1) indicates that activity in the ventral attention system
depends both on the relationship between the distractor
(D) and the memory probe (Im1), as well as the temporal
distance between the two memory probes. In visuospatial
attention research, contingent capture paradigms showed
activation of the rTPJ when a task-relevant target is pre-
sented at a different position compared with the task-
irrelevant but “set-relevant” cue (i.e., spatially invalid trials;
Natale et al., 2010). The interpretation of this is that the
set-relevant cue captures attention, but that rTPJ is acti-
vated because additional operations are required during
target processing (e.g., spatial reorienting). In our cur-
rent account (cf. above), the distractor would corre-
spond to the task-irrelevant “cue,” and on Dim1 trials,
its temporal proximity to Im1 would correspond to the
“feature” that makes that distractor set relevant. Im2
would correspond to the task-relevant target that triggers
activation of the ventral attention system, specifically when
it followed a D-Im1 pair that captured/anchored attention.
As noted above, separate tests on “short” and “long”
distance trials showed that, in opposite fashion to the
rSTS, the rSMG was activated more for Dim2 compared
with Dim1 distractors, specifically in long-distance trials.
This could be interpreted within a framework “reorient-
ing within memory representations” (Cabeza et al., 2012a)
Kwok, Shallice, and Macaluso
2081
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
with the right SMG being engaged because the temporal
association between the distractor and the second probe
(Dim2) triggered a shift of attention away (and more so
on long-distance trials) from the temporal position of
the first probe. This is different from our main account
regarding the Dim1 distractors, but in this case the effect
of proximity would also depend on both the temporal
relationship between the distractor and the probe, as
well as the temporal distance between the two probes.
With respect to this account, both the rSTS and rSMG
were found to process the task-irrelevant distractors as
a function of their temporal relationship with the probes
(i.e., according to their set relevance).
Here we wish to emphasize that the effects of proxi-
mity (main interaction term, Figure 4A, and the two sim-
ple effects on short and long trials, Figure 4B) were all
found in ventral regions of the right hemisphere. This
localization appears consistent with the bottom–up atten-
tion hypothesis (Cabeza et al., 2012a) and attention-to-
memory models (e.g., Cabeza et al., 2008). Our findings help
to extend these proposals as they suggest that the ventral
attention system can capitalize on external, task-irrelevant
signals to modulate memory temporal order retrieval. We
nonetheless acknowledge that our account of set-dependent
contingent capture in the memory domain is still some-
what speculative and that additional work, possibly includ-
ing additional within-participant attention control tasks,
could provide more specific evidence about the role of dif-
ferent subregions around the rTPJ (e.g., see Guerin et al.,
2012; Sestieri et al., 2010; Hutchinson et al., 2009).1
Contingent Capture and Retrieval Specification
Our contingent capture account for the “Dim1 short-
distance trials” also has similarities to the notion of “re-
trieval specifications,” whereby external cues (here, the
“Im1-D” anchor) can provide the participant with a mem-
ory search “description” (Fletcher et al., 1998; Burgess &
Shallice, 1996). Related to this, Fletcher, Shallice, and
Dolan (2000) proposed a mechanism of retrieval from
memory involving Frithʼs (2000) concept of the “sculpting”
of the response space, with retrieval cues modulating lower-
level processes to help to ensure that task-appropriate
responses are selected. Both accounts have been asso-
ciated with activation of the IFG. More broadly, the strong
temporal relationship between the distractor and the two
memory probes on Dim1 short-distance trials (i.e., all
three events were temporally close during encoding) may
trigger specific postretrieval selective processes, again
a function previously associated with the IFG (Badre &
Wagner, 2007; Badre et al., 2005; Thompson-Schill et al.,
2002; Fletcher et al., 2000). Nonetheless, in the current
framework, the IFG would not be merely implicated in
selecting suitable alternatives (Badre et al., 2005) or sup-
pressing unwanted contents (Levy & Anderson, 2012;
Depue, Curran, & Banich, 2007), but rather, it would make
use of potentially distracting task-irrelevant stimuli to help
access relevant information in episodic memory (i.e., the
Im1-Im2 temporal order).
In addition to these time-related effects (Distance ×
Proximity interaction), both behavioral and imaging re-
sults revealed an influence of the content relationship
between the distractors and the memory probes. Behav-
iorally, we found that the influence of Dim1 distractors on
short trials took place only when they belonged to a
different storyline compared with the memory probes
(i.e., Distance × Proximity × Content interaction). Nota-
bly, at the neurophysiological level D-I content and D-I
proximity effects were largely dissociated. Content did
not modulate activities in the right ventral attention net-
work but instead modulated responses in the medial parie-
tal cortex including the precuneus. In this area, we found an
overlap between the main effect of D-I content, with greater
activation for “same” than “different” storylines, and the
main effect of Im1-Im2 distance (“short > long”).
Event Reconstruction and the Precuneus
We predicted that the precuneus would be involved in
reconstructive-based retrieval processes. Confirming this
hypothesis and replicating previous studies that also used
temporal judgment tasks with naturalistic material (Kwok
et al., 2012; St. Jacques et al., 2008), we found modula-
tion of activity in the precuneus as a function of the tem-
poral distance between the two probe images. Activity in
the precuneus increased with decreasing temporal dis-
tance. Previous studies interpreted this effect, suggesting
that the retrieval of temporal events involves search pro-
cesses within knowledge hierarchies organized by eco-
logically meaningful grains (Kwok et al., 2012) and/or
effortful reconstruction processes (St. Jacques et al.,
2008). Here, we found that the content relationship
between memory probes and the distractors modulated
activity in the precuneus, showing that even a brief expo-
sure to semantically more related images during retrieval
affected brain activity in the medial parietal cortex. This
highlights the role of the precuneus in dealing with mean-
ingful, related events during retrieval, lending stronger
support to content-based reconstructive theories (Friedman,
1993, 2004) than chronological search processes (Friedman,
1996, 2001). Moreover, our current results regarding the
precuneus support proposals that implicate this region in
parsing streams of content-related complex events into
meaningful units during perception (Kurby & Zacks,
2008; Zacks, Tversky, & Iyer, 2001) and structuring these
into knowledge-based memory hierarchies (Zacks, Speer,
Swallow, & Maley, 2010; Speer, Zacks, & Reynolds, 2007).
Overall, these findings contribute to a greater specification
of the role of the medial parietal cortex during the retrieval
of temporal information about complex memories and
highlight the central role of content/semantics rather than
mere temporal aspects.
The association of the precuneus with effortful mem-
ory search within knowledge hierarchies also fits with
2082
Journal of Cognitive Neuroscience
Volume 26, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
current hypotheses about the role of top–down (volun-
tary) attention during memory retrieval (AtoM: attention-
to-memory; Cabeza et al., 2008; Ciaramelli et al., 2008).
These theories postulate an attention-related involvement
of dorsal parietal regions—including the precuneus—
during retrieval search, monitoring, and verification
(Cabeza et al., 2008; Ciaramelli et al., 2008). Nonetheless,
to our knowledge, these previous models have not yet
put forward a detailed characterization mapping specific
retrieval operations to subregions within the dorsal parietal
cortex. It is therefore possible that different subsets of
attention processes are utilized by different top–down
memory retrieval operations/tasks. The empirical evidence
presented here suggests that the precuneus may be pref-
erentially engaged for operations acting on discrimi-
nating the content relationship between episodic events
(cf. “reconstructive theories,” Friedman, 1993, 2007).
Together with the precuneus, the content relatedness
of the distractor with respect to the memory probes
(same vs. different storyline) also showed increased acti-
vation in the left pFC. This prefrontal activation is con-
sistent with previous evidence showing that memory
for temporal order is dependent on the pFC (Konishi
et al., 2002; Suzuki et al., 2002; see also Moscovitch,
1992; Milner, Corsi, & Leonard, 1991, for related work
in patients). These frontal effects may be of super-
visory/monitoring nature acting upon some “organiza-
tional” structure (Shallice & Burgess, 1996). In the
context of the current paradigm, we suggest that the
same-storyline trials required additional supervisory/
monitoring processes to correctly sort the three images
(Im1/D/Im2) in relevant memory probes versus the to-be-
ignored distractor. This would occur because the high-level
of content association between the probes and the dis-
tractor in these trials (compared with the different-storyline
trials) generated an additional potential source of confusion
for the retrieval of the temporal order of the two probes.
Behavioral Outcome Is Multifaceted
Finally, the content relationship between the distractors
and the memory probes was found to modulate activity
in the visual cortex. In the lateral occipital cortex, we
found greater activity for Dim1 than Dim2 distractors spe-
cifically in short-distance, different-storylines trials. Thus,
in the visual cortex, the pattern of brain activity paralleled
the interaction between distance, content, and proximity
found at the behavioral level. This result was unexpected,
and we can only speculate about the possible underlying
processes. On each trial, the task required the participant
to suppress the processing of the interleaved distractor
image, focusing selectively on the first and third images
for the memory judgment. The behavioral advantage for
trials including Dim1 distractors indicates that the sup-
pressed representation of the task-irrelevant distractor
was reduced (cf. also “directed forgetting,” Bjork & Bjork,
1996). One possibility is that the modulation of activity in
visual cortex reflects some aspect of this recovery pro-
cess, the strength of which may depend both on sensory
details of the three pictures presented sequentially (Im1-
D-Im2: “repetition priming”; Buckner et al., 1998) as well
as inhibitory mechanisms related to the filtering/selection
requirements of the current memory task (“negative
priming”; see Nee & Jonides, 2008).
Despite the similarity between the pattern of activation
in occipital cortex and the behavioral data (three-way
interaction), we wish to emphasize that the behavioral
performance represents only the final output of multiple
operations/processes, likely to be carried out by different
areas of the brain. In particular, the performance for the
“Dim1/short-distance/different-storyline” trials ought to
depend on processes carried out by both the rTPJ-IFG
network (cf. our contingent capture account) and the
medial parietal cortex, which were activated when par-
ticipants have to judge on short-distance trials (see also
Kwok et al., 2012) as well as being differentially involved
when the distractor and the memory probes belonged to
the same versus different storyline.
Conclusions
We conclude that bottom–up attention and memory re-
trieval entail common selection processes and that this
involves a multifaceted mechanism. The degree to which
task-irrelevant distractor stimuli presented during re-
trieval affected memory performance, depended on the
specific relationship between the irrelevant perceptual
signals and the memory information that participants
were asked to retrieve (i.e., the task set). Both temporal-
and content-related factors were found to play a role.
Behaviorally, this led to a complex pattern of retrieval
performance, where task-irrelevant distractors that were
close in time—at presentation—to the first memory
probe reduced the RT costs of retrieving temporal infor-
mation on short-distance trials. We link this effect of tem-
poral proximity to set-related contingent capture of
attention on a memory representation level and associate
this with the activation of the rTPJ-IFG network. The con-
tent relationship between the distractor and memory
probes also influenced the retrieval performance, but in
a different way from the temporal effects at the neuro-
physiological level. The influence of the distractor probe
content relationship was found to overlap with the effect
of temporal distance between the two probes in the pre-
cuneus. This is consistent with the proposal that the in-
volvement of the medial parietal cortex during memory
retrieval reflects search processes within knowledge/
content-based memory hierarchies.
Acknowledgments
This work, conducted in The Neuroimaging Laboratory, Santa
Lucia Foundation, was supported by The Italian Ministry of
Health. This work has also received funding from the European
Kwok, Shallice, and Macaluso
2083
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Research Council under the European Unionʼs Seventh Frame-
work Programme (FP7/2007-2013)/ ERC Grant Agreement
n.242809. The authors thank Paolo Alessandrini, Steve Gazzitano,
Fabio Mollo, and Domenico Itri for their assistance.
Reprint requests should be sent to Sze Chai Kwok, Neuroimaging
Laboratory, Santa Lucia Foundation, Via Ardeatina 306, 00179
Rome, Italy, or via e-mail: sc.kwok@hsantalucia.it.
Note
1. Distance and proximity were found to interact in the right
posterior/superior temporal cortex (rSTS) and the right anterior
part of the inferior parietal cortex (rSMG). The possible dis-
tinction between anterior and posterior regions of parietal cortex
(i.e., SMG vs. AG) and the temporal cortex (STS and SMG) has
been long debated in the attention literature (e.g., Karnath, Ferber,
& Himmelbach, 2001) and, more recently, in the literature on
attention and memory (e.g., Cabeza et al., 2012a; Nelson et al.,
2012). Studies on attention reorienting have reported peaks in
SMG (Kincade et al., 2005), AG (Arrington, Carr, Mayer, & Rao,
2000), as well as temporal regions including the posterior STS
(Macaluso & Patria, 2007; Gitelman et al., 1999), and in the absence
of any specific within-experiment dissociations the term TPJ is
often employed (see Carter & Huettel, 2013; Jakobs et al., 2012,
for recent examples). Adding further complexity to this issue, the
caudal part of STS extends into the inferior parietal lobule, possibly
including cytoarchitectonic areas corresponding to AG (see Segal
& Petrides, 2012; Caspers et al., 2006). To draw any distinctions
between the functions of these regions around the TPJ would be
beyond the aim of the current study (but see Daselaar, Huijbers,
Eklund, Moscovitch, & Cabeza, 2013; Nelson, McDermott, Wig,
Schlaggar, & Petersen, 2013, for studies specifically designed to
address this issue).
REFERENCES
Arrington, C. M., Carr, T. H., Mayer, A. R., & Rao, S. M. (2000).
Neural mechanisms of visual attention: Object-based
selection of a region in space. Journal of Cognitive
Neuroscience, 12(Suppl. 2), 106–117.
Baddeley, A., Lewis, V., Eldridge, M., & Thomson, N. (1984).
Attention and retrieval from long-term memory. Journal of
Experimental Psychology: General, 113, 518.
Badre, D., Poldrack, R. A., Paré-Blagoev, E. J., Insler, R. Z., &
Wagner, A. D. (2005). Dissociable controlled retrieval and
generalized selection mechanisms in ventrolateral prefrontal
cortex. Neuron, 47, 907–918.
Badre, D., & Wagner, A. D. (2005). Frontal lobe mechanisms
that resolve proactive interference. Cerebral Cortex, 15,
2003–2012.
Badre, D., & Wagner, A. D. (2007). Left ventrolateral prefrontal
cortex and the cognitive control of memory.
Neuropsychologia, 45, 2883–2901.
Bjork, E. L., & Bjork, R. A. (1996). Continuing influences of
to-be-forgotten information. Consciousness and Cognition,
5, 176–196.
Buckner, R. L., Goodman, J., Burock, M., Rotte, M., Koutstaal,
W., Schacter, D., et al. (1998). Functional-anatomic correlates
of object priming in humans revealed by rapid presentation
event-related fMRI. Neuron, 20, 285–296.
Burgess, P. W., & Shallice, T. (1996). Confabulation and the
control of recollection. Memory, 4, 359–411.
Cabeza, R., Ciaramelli, E., & Moscovitch, M. (2012a). Cognitive
contributions of the ventral parietal cortex: An integrative
theoretical account. Trends in Cognitive Sciences, 16,
338–352.
Cabeza, R., Ciaramelli, E., & Moscovitch, M. (2012b). Response
to Nelson et al.: Ventral parietal subdivisions are not
incompatible with an overarching function. Trends in
Cognitive Sciences, 16, 400–401.
Cabeza, R., Ciaramelli, E., Olson, I. R., & Moscovitch, M. (2008).
The parietal cortex and episodic memory: An attentional
account. Nature Reviews Neuroscience, 9, 613–625.
Carter, R. M., & Huettel, S. A. (2013). A nexus model of the
temporal-parietal junction. Trends in Cognitive Sciences, 17,
328–336.
Caspers, S., Geyer, S., Schleicher, A., Mohlberg, H., Amunts,
K., & Zilles, K. (2006). The human inferior parietal cortex:
Cytoarchitectonic parcellation and interindividual variability.
Neuroimage, 33, 430–448.
Chun, M. M., & Jiang, Y. (2003). Implicit, long-term spatial
contextual memory. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 29, 224–234.
Chun, M. M., & Johnson, M. K. (2011). Memory: Enduring traces
of perceptual and reflective attention. Neuron, 72, 520–535.
Ciaramelli, E., Grady, C., Levine, B., Ween, J., & Moscovitch, M.
(2010). Top–down and bottom–up attention to memory are
dissociated in posterior parietal cortex: Neuroimaging and
neuropsychological evidence. The Journal of Neuroscience,
30, 4943–4956.
Ciaramelli, E., Grady, C. L., & Moscovitch, M. (2008). Top–down
and bottom–up attention to memory: A hypothesis (AtoM)
on the role of the posterior parietal cortex in memory
retrieval. Neuropsychologia, 46, 1828–1851.
Corbetta, M., Patel, G., & Shulman, G. L. (2008). The reorienting
system of the human brain: From environment to theory of
mind. Neuron, 58, 306–324.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews
Neuroscience, 3, 201–215.
Craik, F. I. M., Govoni, R., Naveh-Benjamin, M., & Anderson,
N. D. (1996). The effects of divided attention on encoding
and retrieval processes in human memory. Journal of
Experimental Psychology: General, 125, 159–180.
Daselaar, S. M., Huijbers, W., Eklund, K., Moscovitch, M., &
Cabeza, R. (2013). Resting-state functional connectivity of
ventral parietal regions associated with attention reorienting
and episodic recollection. Frontiers in Human Neuroscience,
7, 1–9.
Depue, B. E., Curran, T., & Banich, M. T. (2007). Prefrontal
regions orchestrate suppression of emotional memories via a
two-phase process. Science, 317, 215–219.
DiQuattro, N. E., & Geng, J. J. (2011). Contextual knowledge
configures attentional control networks. The Journal of
Neuroscience, 31, 18026–18035.
Fairhall, S. L., Indovina, I., Driver, J., & Macaluso, E. (2009). The
brain network underlying serial visual search: Comparing
overt and covert spatial orienting, for activations and for
effective connectivity. Cerebral Cortex, 19, 2946–2958.
Fletcher, P. C., Shallice, T., & Dolan, R. J. (2000). “Sculpting the
response space“—An account of left prefrontal activation at
encoding. Neuroimage, 12, 404–417.
Fletcher, P. C., Shallice, T., Frith, C. D., Frackowiak, R. S., &
Dolan, R. J. (1998). The functional roles of prefrontal cortex
in episodic memory. II. Retrieval. Brain, 121, 1249–1256.
Folk, C. L., Leber, A. B., & Egeth, H. E. (2002). Made you blink!
Contingent attentional capture produces a spatial blink.
Perception & Psychophysics, 64, 741–753.
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992).
Involuntary covert orienting is contingent on attentional
control settings. Journal of Experimental Psychology:
Human Perception and Performance, 18, 1030–1044.
Friedman, W. J. (1993). Memory for the time of past events.
Psychological Bulletin, 113, 44–66.
2084
Journal of Cognitive Neuroscience
Volume 26, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Friedman, W. J. (1996). Distance and location processes in
memory for the times of past events. In D. L. Medin (Ed.),
The psychology of learning and motivation ( Vol. 35,
pp. 1–41). San Diego, CA: Academic Press.
Friedman, W. J. (2001). Memory processes underlying humansʼ
chronological sense of the past. In C. Hoerl & T. McCormack
(Eds.), Time and memory: Issues in philosophy and
psychology (pp. 139–167). Oxford, England: Oxford
University Press.
Friedman, W. J. (2004). Time in autobiographical memory.
Social Cognition, 22, 591–605.
Friedman, W. J. (2007). The role of reminding in long-term
memory for temporal order. Memory & Cognition, 35, 66–72.
Friston, K. J., Penny, W., Phillips, C., Kiebel, S., Hinton, G., &
Ashburner, J. (2002). Classical and Bayesian inference in
neuroimaging: Theory. Neuroimage, 16, 465–483.
Frith, C. D. (2000). The role of the dorsolateral prefrontal cortex
in the selection of action as revealed by functional imaging. In
S. Monsell & J. Driver (Eds.), Control of cognitive processes:
Attention and performance XVIII (pp. 549–565). Cambridge,
MA: MIT Press.
Kastner, S., & Ungerleider, L. G. (2000). Mechanisms of visual
attention in the human cortex. Annual Review of Neuroscience,
23, 315–341.
Kincade, J. M., Abrams, R. A., Astafiev, S. V., Shulman, G. L., &
Corbetta, M. (2005). An event-related functional magnetic
resonance imaging study of voluntary and stimulus-driven
orienting of attention. The Journal of Neuroscience, 25,
4593–4604.
Konishi, S., Uchida, I., Okuaki, T., Machida, T., Shirouzu, I., &
Miyashita, Y. (2002). Neural correlates of recency judgment.
The Journal of Neuroscience, 22, 9549–9555.
Kurby, C. A., & Zacks, J. M. (2008). Segmentation in the
perception and memory of events. Trends in Cognitive
Sciences, 12, 72–79.
Kwok, S. C., Shallice, T., & Macaluso, E. (2012). Functional
anatomy of temporal organisation and domain-specificity
of episodic memory retrieval. Neuropsychologia, 50,
2943–2955.
Levy, B. J., & Anderson, M. C. (2012). Purging of memories from
conscious awareness tracked in the human brain. The
Journal of Neuroscience, 32, 16785–16794.
Geng, J. J., & Mangun, G. R. (2011). Right temporoparietal
Macaluso, E., & Patria, F. (2007). Spatial re-orienting of visual
junction activation by a salient contextual cue facilitates target
discrimination. Neuroimage, 54, 594–601.
Gitelman, D. R., Nobre, A. C., Parrish, T. B., LaBar, K. S.,
Kim, Y.-H., Meyer, J. R., et al. (1999). A large-scale
distributed network for covert spatial attention. Brain, 122,
1093–1106.
Guerin, S. A., Robbins, C. A., Gilmore, A. W., & Schacter, D. L.
(2012). Interactions between visual attention and episodic
retrieval: Dissociable contributions of parietal regions during
gist-based false recognition. Neuron, 75, 1122–1134.
Hayama, H. R., Vilberg, K. L., & Rugg, M. D. (2012). Overlap
between the neural correlates of cued recall and source
memory: Evidence for a generic recollection network?
Journal of Cognitive Neuroscience, 24, 1127–1137.
Henson, R. N. A., Shallice, T., Josephs, O., & Dolan, R. J. (2002).
Functional magnetic resonance imaging of proactive
interference during spoken cued recall. Neuroimage, 17,
543–558.
Hopfinger, J. B., Buonocore, M. H., & Mangun, G. R. (2000).
The neural mechanisms of top–down attentional control.
Nature Neuroscience, 3, 284–291.
Howard, M. W., & Kahana, M. J. (2002). A distributed
representation of temporal context. Journal of
Mathematical Psychology, 46, 269–299.
Hutchinson, J. B., & Turk-Browne, N. B. (2012). Memory-guided
attention: Control from multiple memory systems. Trends in
Cognitive Sciences, 16, 576–579.
Hutchinson, J. B., Uncapher, M. R., & Wagner, A. D. (2009).
Posterior parietal cortex and episodic retrieval: Convergent
and divergent effects of attention and memory. Learning &
Memory, 16, 343–356.
Indovina, I., & Macaluso, E. (2007). Dissociation of stimulus
relevance and saliency factors during shifts of visuospatial
attention. Cerebral Cortex, 17, 1701–1711.
Jakobs, O., Langner, R., Caspers, S., Roski, C., Cieslik, E. C.,
Zilles, K., et al. (2012). Across-study and within-subject
functional connectivity of a right temporo-parietal junction
subregion involved in stimulus-context integration.
Neuroimage, 60, 2389–2398.
Jonides, J., Smith, E. E., Marshuetz, C., Koeppe, R. A., & Reuter-
Lorenz, P. A. (1998). Inhibition in verbal working memory
revealed by brain activation. Proceedings of the National
Academy of Sciences, 95, 8410–8413.
Karnath, H.-O., Ferber, S., & Himmelbach, M. (2001). Spatial
awareness is a function of the temporal not the posterior
parietal lobe. Nature, 411, 950–953.
attention along the horizontal or the vertical axis.
Experimental Brain Research, 180, 23–34.
Milham, M. P., Banich, M. T., Webb, A., Barad, V., Cohen, N. J.,
Wszalek, T., et al. (2001). The relative involvement of anterior
cingulate and prefrontal cortex in attentional control depends
on nature of conflict. Cognitive Brain Research, 12, 467–473.
Milner, B., Corsi, P., & Leonard, G. (1991). Frontal-lobe
contribution to recency judgements. Neuropsychologia, 29,
601–618.
Moscovitch, M. (1992). Memory and working-with-memory: A
component process model based on modules and central
systems. Journal of Cognitive Neuroscience, 4, 257–267.
Naghavi, H. R., & Nyberg, L. (2005). Common fronto-parietal
activity in attention, memory, and consciousness: Shared
demands on integration? Consciousness and Cognition, 14,
390–425.
Natale, E., Marzi, C. A., & Macaluso, E. (2010). Right temporal-
parietal junction engagement during spatial reorienting
does not depend on strategic attention control.
Neuropsychologia, 48, 1160–1164.
Nee, D. E., & Jonides, J. (2008). Dissociable interference-control
processes in perception and memory. Psychological Science,
19, 490–500.
Nelson, S. M., McDermott, K. B., & Petersen, S. E. (2012).
In favor of a “fractionation” view of ventral parietal cortex:
Comment on Cabeza et al. Trends in Cognitive Sciences, 16,
399–400.
Nelson, S. M., McDermott, K. B., Wig, G. S., Schlaggar, B. L., &
Petersen, S. E. (2013). The critical roles of localization and
physiology for understanding parietal contributions to
memory retrieval. The Neuroscientist, 19, 578–591.
Penny, W., & Holmes, A. (2004). Random effects analysis. In
R. S. Frackowiak, K. J. Friston, C. D. Frith, R. J. Dolan, C. J.
Price, S. Zeki, et al. (Eds.), Human brain function (2nd ed.,
pp. 843–851). San Diego, CA: Elsevier.
Prince, S. E., Tsukiura, T., & Cabeza, R. (2007). Distinguishing
the neural correlates of episodic memory encoding and
semantic memory retrieval. Psychological Science, 18,
144–151.
Sederberg, P. B., Howard, M. W., & Kahana, M. J. (2008). A
context-based theory of recency and contiguity in free recall.
Psychological Review, 115, 893.
Segal, E., & Petrides, M. (2012). The anterior superior parietal
lobule and its interactions with language and motor areas
during writing. European Journal of Neuroscience, 35,
309–322.
Kwok, Shallice, and Macaluso
2085
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth,
H. E., & Yantis, S. (2005). Coordination of voluntary and
stimulus-driven attentional control in human cortex.
Psychological Science, 16, 114–122.
Sestieri, C., Corbetta, M., Romani, G. L., & Shulman, G. L.
(2011). Episodic memory retrieval, parietal cortex, and the
default mode network: Functional and topographic analyses.
The Journal of Neuroscience, 31, 4407–4420.
Sestieri, C., Shulman, G. L., & Corbetta, M. (2010). Attention to
memory and the environment: Functional specialization and
dynamic competition in human posterior parietal cortex.
The Journal of Neuroscience, 30, 8445–8456.
Shallice, T., & Burgess, P. (1996). The domain of supervisory
processes and temporal organization of behaviour [and
discussion]. Philosophical Transactions of the Royal Society
of London, Series B, Biological Sciences, 351, 1405–1412.
Shulman, G. L., McAvoy, M. P., Cowan, M. C., Astafiev, S. V.,
Tansy, A. P., dʼAvossa, G., et al. (2003). Quantitative analysis
of attention and detection signals during visual search.
Journal of Neurophysiology, 90, 3384–3397.
Speer, N. K., Zacks, J. M., & Reynolds, J. R. (2007). Human brain
activity time-locked to narrative event boundaries.
Psychological Science, 18, 449–455.
St. Jacques, P., Rubin, D. C., LaBar, K. S., & Cabeza, R. (2008).
The short and long of it: Neural correlates of temporal-order
memory for autobiographical events. Journal of Cognitive
Neuroscience, 20, 1327–1341.
Stokes, M. G., Atherton, K., Patai, E. Z., & Nobre, A. C. (2012).
Long-term memory prepares neural activity for perception.
Proceedings of the National Academy of Sciences, 109,
E360–E367.
Summerfield, J. J., Rao, A., Garside, N., & Nobre, A. C. (2011).
Biasing perception by spatial long-term memory. The Journal
of Neuroscience, 31, 14952–14960.
Suzuki, M., Fujii, T., Tsukiura, T., Okuda, J., Umetsu, A., Nagasaka,
T., et al. (2002). Neural basis of temporal context memory:
A functional MRI study. Neuroimage, 17, 1790–1796.
Thompson-Schill, S. L., DʼEsposito, M., Aguirre, G. K., & Farah,
M. J. (1997). Role of left inferior prefrontal cortex in retrieval
of semantic knowledge: A reevaluation. Proceedings of the
National Academy of Sciences, 94, 14792–14797.
Thompson-Schill, S. L., Jonides, J., Marshuetz, C., Smith, E. E.,
DʼEsposito, M., Kan, I. P., et al. (2002). Effects of frontal lobe
damage on interference effects in working memory.
Cognitive, Affective, & Behavioral Neuroscience, 2, 109–120.
Todd, J. J., Fougnie, D., & Marois, R. (2005). Visual short-term
memory load suppresses temporo-parietal junction activity
and induces inattentional blindness. Psychological Science,
16, 965–972.
Turk-Browne, N. B., Golomb, J. D., & Chun, M. M. (2013).
Complementary attentional components of successful
memory encoding. Neuroimage, 66, 553–562.
Vilberg, K. L., & Rugg, M. D. (2008). Memory retrieval and the
parietal cortex: A review of evidence from a dual-process
perspective. Neuropsychologia, 46, 1787–1799.
Vilberg, K. L., & Rugg, M. D. (2012). The neural correlates
of recollection: Transient versus sustained fMRI effects.
The Journal of Neuroscience, 32, 15679–15687.
Wagner, A. R., Shannon, B., Kahn, I., & Buckner, R. (2005).
Parietal lobe contributions to episodic memory retrieval.
Trends in Cognitive Sciences, 9, 445–453.
Wais, P. E., Kim, O. Y., & Gazzaley, A. (2011). Distractibility
during episodic retrieval is exacerbated by perturbation of
left ventrolateral prefrontal cortex. Cerebral Cortex, 22,
717–724.
Wais, P. E., Martin, G. M., & Gazzaley, A. (2012). The impact of
visual distraction on episodic retrieval in older adults. Brain
Research, 1430, 78–85.
Wais, P. E., Rubens, M. T., Boccanfuso, J., & Gazzaley, A. (2010).
Neural mechanisms underlying the impact of visual
distraction on retrieval of long-term memory. The Journal of
Neuroscience, 30, 8541–8550.
Wheeler, M. E., & Buckner, R. L. (2003). Functional dissociation
among components of remembering: Control, perceived
oldness, and content. The Journal of Neuroscience, 23,
3869–3880.
Zacks, J. M., Speer, N. K., Swallow, K. M., & Maley, C. J.
(2010). The brainʼs cutting-room floor: Segmentation of
narrative cinema. Frontiers in Human Neuroscience, 4,
Article 168.
Zacks, J. M., Tversky, B., & Iyer, G. (2001). Perceiving,
remembering, and communicating structure in events.
Journal of Experimental Psychology: General, 130, 29–58.
Zanto, T. P., & Gazzaley, A. (2009). Neural suppression of
irrelevant information underlies optimal working memory
performance. The Journal of Neuroscience, 29, 3059–3066.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
–
p
–
d
p
d
2
f
6
/
9
2
6
2
/
0
9
7
/
0
2
1
0
9
7
4
0
7
/
7
1
5
7
4
8
o
1
c
7
n
0
_
1
a
/
_
j
0
o
0
c
6
n
0
1
_
a
p
_
d
0
0
b
6
y
0
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
2086
Journal of Cognitive Neuroscience
Volume 26, Number 9