Seeing Sounds and Hearing Colors: An Event-related
Potential Study of Auditory–Visual Synesthesia
Aviva I. Goller1, Leun J. Otten2, and Jamie Ward1
Abstract
& In auditory–visual synesthesia, sounds automatically elicit
conscious and reliable visual experiences. It is presently un-
known whether this reflects early or late processes in the
brain. It is also unknown whether adult audiovisual synesthe-
sia resembles auditory-induced visual illusions that can some-
times occur in the general population or whether it resembles
the electrophysiological deflection over occipital sites that has
been noted in infancy and has been likened to synesthesia.
Electrical brain activity was recorded from adult synesthetes
and control participants who were played brief tones and re-
quired to monitor for an infrequent auditory target. The syn-
esthetes were instructed to attend either to the auditory or to
the visual (i.e., synesthetic) dimension of the tone, whereas
the controls attended to the auditory dimension alone. There
were clear differences between synesthetes and controls that
emerged early (100 msec after tone onset). These differences
tended to lie in deflections of the auditory-evoked potential
(e.g., the auditory N1, P2, and N2) rather than the presence
of an additional posterior deflection. The differences occurred
irrespective of what the synesthetes attended to (although
attention had a late effect). The results suggest that differences
between synesthetes and others occur early in time, and that
synesthesia is qualitatively different from similar effects found
in infants and certain auditory-induced visual illusions in adults.
In addition, we report two novel cases of synesthesia in which
colors elicit sounds, and vice versa. &
INTRODUCTION
In auditory–visual synesthesia, sounds automatically
elicit conscious visual percepts in addition to an audi-
tory percept. For example, a cello may sound like a
‘‘dark velvet or reddish-brown tree trunk-like texture’’
and a flute may be ‘‘dry and transparent with pastel
colors’’ (Mills, Boteler, & Larcombe, 2003). For some
individuals, the synesthesia is triggered solely by speech
(e.g., Nunn et al., 2002; Paulesu et al., 1995; Baron-
Cohen, Harrison, Goldstein, & Wyke, 1993), but for
others the synesthesia is triggered by all known auditory
stimuli (e.g., Thornley Head, 2006; Ward, Huckstep, &
Tsakanikos, 2006). This may reflect a qualitative dif-
ference between whether synesthesia is linked to linguis-
tic representations (e.g., graphemes) versus perceptual
properties of the stimulus such as its pitch (e.g., Simner,
Glover, & Mowat, 2006; Frith & Paulesu, 1997). Our study
considers the latter, using nonspeech tones. This variety
of synesthesia is of particular theoretical interest because
of the large literature on audiovisual interactions in the
nonsynesthetic brain (e.g., Calvert, Hansen, Iversen, &
Brammer, 2001). It raises the possibility that this type of
synesthesia reflects an adaptation of normal multisensory
processes (Ward et al., 2006). Previous research has dem-
onstrated the authenticity of this type of synesthesia by
1University of Sussex, 2University College London
showing that the auditory-color associations of these
synesthetes are more consistent than controls and by
showing that the synesthetic color of a task-irrelevant
tone interferes with color naming in a Stroop task (Ward
et al., 2006). However, in other respects there are com-
monalities between the nature of synesthetic experiences
and those reported by nonsynesthetes in imagery, match-
ing tasks, or cross-modal interference paradigms (Ward
et al., 2006; Marks, 2004). In particular, high-pitched
sounds tend to be visually lighter, higher, and smaller
than low-pitch sounds in both synesthetic experiences
and response biases of nonsynesthetes (Marks, 2004).
This suggests common processes between synesthetic
perception and audiovisual processing of nonsynesthetes.
The present study will use ERPs (small changes in the
brain’s electrical activity time locked to an event) to ad-
judicate between two different theories.
One suggestion is that the newborn infants’ experi-
ences of the world resemble a form of synesthesia
(Maurer & Mondloch, 2006; Maurer & Maurer, 1988),
in which the senses are yet to be differentiated and
in which one sense (e.g., audition) can trigger another
infants show cross-modal
(e.g., vision). For example,
habituation depending on the intensity of light and
sounds (Lewkowicz & Turkewitz, 1980). A further claim
is that, in some individuals, these early multisensory path-
ways are retained into adulthood giving rise to devel-
opmental synesthesia whereas in most other individuals
D 2008 Massachusetts Institute of Technology
Journal of Cognitive Neuroscience 21:10, pp. 1869–1881
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
e
1
0
–
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
/
t
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
they are greatly diminished (Maurer, 1997; Baron-Cohen,
1996). Electrophysiological correlates of infantile auditory–
visual ‘‘synesthesia’’ have been reported. These consist
of a large negative deflection between 100 and 500 msec
over occipital sites (absent by 30 months of age), con-
trasting with a developmentally more stable potential
over temporal sites (Neville, 1995). Although a direct
comparison between adult audiovisual synesthetes and
normal infants would be impossible to interpret (e.g.,
due to developmental changes in conductance), one can
nevertheless determine whether a qualitatively similar
electrophysiological signature is found in adult synes-
thetes to that previously documented in infants. Accord-
ing to this account, the electrophysiological responses
to auditory stimuli should include an early deflection
maximal over posterior sites.
Recent studies have shown that there are direct projec-
tions from primary auditory cortex (A1) to primary visual
cortex (V1) in the mature primate brain although they
are primarily found in regions representing peripheral
vision (Rockland & Ojima, 2003; Falchier, Clavagnier,
Barone, & Kennedy, 2002). Even in nonsynesthetes, di-
rect auditory–visual projections may play a functional
role in multisensory processing. It may even give rise to
a synesthesia-like illusion in the normal population.
Shams, Kamitani, Thompson, and Shimojo (2001) and
Shams, Kamitani, and Shimojo (2000) report that if two
beeps are played in quick succession and are accompa-
nied by a single flash, then participants often perceive
two distinct flashes instead of one: the double-flash illu-
sion. The illusion occurs predominantly in peripheral vi-
sion, consistent with the known anatomy of the direct
projections. They report that the illusory flash is accom-
panied by electrical activity over occipital sites (Oz, O1,
and O2) less than 110 msec after the onset of the second
beep (Shams et al., 2001), and a recent fMRI study shows
differences in V1 activity but not in other brain regions
(Watkins, Shams, Tanaka, Haynes, & Rees, 2006).
Anatomical studies that have attempted to look for
the reverse pathways, from V1 to A1, have not found
them (Innocenti, Berbel, & Clarke, 1988). The apparent
rarity with which vision evokes sound in synesthesia
relative to vice versa (e.g., Simner, Mulvenna, et al.,
2006; Day, 2005) could therefore relate to the relative
availability of these multisensory pathways. However,
visual–auditory synesthesia has been anecdotally noted
before. Baron-Cohen, Burt, Smith-Laittan, Harrison, and
Bolton (1996) briefly note the case of JR who sees colors
when she hears sounds and also hears sounds when
she sees colors (a situation that does cause perceptual
interference in her day-to-day life). They note that she
shows high consistency over time when assessed in
both directions although the associations are not nec-
essarily the same as each other in either direction. For
instance, a red traffic light may trigger a particular syn-
esthetic sound, but that sound may itself trigger a
synesthetic color of, say, blue (rather than red). In this
study, we retest JR using visual- and auditory-evoked
ERPs together with one other synesthete, SL, who re-
ports a similar pattern to JR.
There is, however, an alternative to the hypothesis
of direct auditory–visual connections. This has been
termed the ‘‘cross-modal transfer hypothesis’’ ( Ward
et al., 2006; Baron-Cohen, 1996). This hypothesis as-
sumes that connections between auditory and visual
regions are indirect and are mediated by multisensory
audiovisual brain regions. Activation in multisensory
neurons may feedback and influence activity in regions
traditionally considered to be unisensory (e.g., Driver
& Spence, 2000). Although multisensory processes may
normally be activated when two senses are stimulated,
it is also conceivable that they can sometimes be ac-
tivated by a unimodal stimulus in some situations
(e.g., Giraud, Price, Graham, Truy, & Frackowiak, 2001;
Calvert et al., 1997). Synesthesia may be one such
example. According to this account, an additional ERP
deflection (due to multisensory binding) would follow
the normal early auditory deflections, but would pre-
cede in time any auditory-evoked visual potential.
To date, there have been very few ERP studies of
synesthesia. Two studies have considered grapheme-
color synesthesia using visually presented graphemes
(Schiltz et al., 1999) or spoken letter names and words
(Beeli, Esslen, & Jancke, 2008). In addition, there are
two single case studies that have specifically investigated
electrophysiological correlates of auditory–visual synes-
thesia using nonlinguistic stimuli (Rao, Nobre, Alexander,
& Cowey, 2007; Rizzo & Eslinger, 1989).
Schiltz et al. (1999) tested 17 grapheme-color synes-
thetes who were presented with runs of visual letters
and who were required to detect certain target letters
(e.g., vowels). They reported an increased positivity at
frontal and central scalp sites emerging around 150 msec
and maintained until 600 msec, relative to a nonsynes-
thetic control group. More recently, an ERP study was
reported of grapheme-color synesthesia in which spo-
ken letters and words elicit experiences of color (Beeli
et al., 2008). As noted above,
in this type of ‘‘color
hearing,’’ it has been suggested that speech activates
graphemic representation of words, which is then linked
to experiences of color (e.g., Simner, Glover, et al., 2006;
Frith & Paulesu, 1997). The fact that visual graphemes
also elicit color for these synesthetes is consistent with
this (Beeli et al., 2008). Beeli et al. (2008) found reduced
amplitudes and/or increased latencies of the auditory
N1, P2, and N2 deflections in synesthetes in response
to spoken letters and words. Although few electrode
sites were available, source localization implicated ad-
ditional activity in the synesthetes in OFC and inferior
temporal lobe.
Of the two single case studies of auditory–visual syn-
esthesia that have measured ERPs to nonlinguistic sounds,
one was an acquired case of synesthesia following blind-
ness, and the other was a case of developmental synes-
1870
Journal of Cognitive Neuroscience
Volume 21, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
e
1
0
–
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
t
f
/
.
.
o
n
1
8
M
a
y
2
0
2
1
thesia more closely related to that investigated here.
The case of Rao et al. (2007) suffered destruction of the
optic nerves following a car accident and began to re-
port phosphenes from auditory stimuli a year or so later.
A comparison of evoked potentials to sounds that did
elicit a visual experience versus those that did not re-
vealed modulation of an auditory-evoked N1 deflection
(around 100 msec) including over occipital sites. The
authors suggest that this reflects auditory activation of
visual cortex rather than volume conduction from a
distant site because sighted controls showed no com-
parable activity over the same electrode sites. Rizzo and
Eslinger (1989) conducted the only electrophysiologi-
cal study of developmental auditory–visual synesthesia
to date. Auditory clicks were not associated with abnor-
mal potentials at O1, O2, or Oz (other electrodes were
not reported), and they conclude that the synesthesia
may depend on ‘‘more anterior structures with poly-
modal connections.’’ The results of this case study were
very different to that obtained by Rao et al. (2007), sug-
gesting a possible involvement of different neural path-
ways in acquired and developmental cases of auditory–
visual synesthesia. This will be returned to again in the
discussion.
The present study is the first to consider the time
course, using ERPs, of synesthesia in a group of people
who experience colors in response to nonlinguistic
sounds. This type of synesthesia is likely to be quali-
tatively different to that based on speech sounds and
more closely resembles (in terms of phenomenology
and candidate mechanisms) synesthesia-like illusions in
the normal population (Shams et al., 2001), acquired
audiovisual synesthesia (Rao et al., 2007), and possible
infantile synesthesia (Neville, 1995). A series of pure
tones were played to a group of synesthetes (n = 10)
and controls (n = 10) who were required to detect an
infrequent target tone. In different blocks, the synes-
thetes were additionally instructed to attend either to
the pitch of the tone or to the synesthetic color of
the tone. Attention modulates synesthesia as measured
on tasks such as synesthetic Stroop interference (e.g.,
Mattingley, Payne, & Rich, 2006). However, inattention
does not necessarily eliminate synesthesia (Sagiv, Heer,
& Robertson, 2006), and it is unclear whether atten-
tion exerts its effects early or late in the induction of
synesthesia. Synesthetes and controls were additionally
shown unimodal color patches to record visual-evoked
potentials. The color patches were similar in hue to
some of their auditory-induced visual experiences al-
though only 2 of 10 of the synesthetes reported con-
scious auditory experiences from seeing color. As such,
we were able to explore differences between synes-
thetes who do and do not experience sound from color
(comparing the 2 synesthetes to the remaining 8) and
also to assess whether synesthetes have normal electro-
physiological responses to visual stimuli (by comparing
the 8 synesthetes to the 10 nonsynesthetes). It has been
suggested that colors may implicitly trigger representa-
tions of number in synesthetes (Cohen Kadosh, Cohen
Kadosh, & Henik, 2007; Cohen Kadosh & Henik, 2006;
Cohen Kadosh et al., 2005), and it is conceivable that
colors may elicit sounds.
METHODS
Participants
Ten auditory–visual synesthetes (9 women) and 10 healthy
controls (6 women) gave written informed consent to take
part in this experiment. The procedure was approved
by the University College London ethics committee.
The mean age of the synesthetes was 39.7 years with a
range of 21–68 years, and the mean age of controls was
39.9 years with a range of 20–67 years. All synesthetes
reported being right-handed; two controls reported be-
ing left-handed. Handedness was considered unlikely to
affect performance in auditory detection. Eight of the
synesthetes report their sound–vision synesthesia to be
unidirectional (i.e., sounds evoke vision but vision does
not evoke sound), whereas two claim it to be bidirec-
tional. For example, JR (also studied by Baron-Cohen
et al., 1996) reports the following sounds as she moves
her gaze around a Kandinsky painting (Composition VIII,
1923): ‘‘There is a huge splurge of sound left-hand top—
booming but also a bit vulgar! Below it a rather mousy
little meee sound which then translates into ohs and ahs
and pops at the various circles. The lines are sharp and
moving to the right with the sound of steel (like blades
scraping against one another). The triangle and boo-
merang shapes are surprised and pop up laughing with
a whooo.’’
All participants were free of known neurological illness
and reported normal hearing and normal or corrected-to-
normal vision. Color vision was assessed with the Ishihara
plates and was found to be normal in all participants.
Participants were paid at a rate of £7.50/hr and had their
travel expenses reimbursed.
The synesthetes were informally asked about the spa-
tial location of their photisms by altering the location
of a sound (e.g., a knock) and by altering their posture.
If the locations of the photisms are gaze centered, then
it would suggest dependence on retinotopically orga-
nized regions (e.g., V1), but if the locations depend on
posture, it would be more consistent with an involve-
ment of higher visual processes (e.g., Colby & Goldberg,
1999). None reported gaze-centered photisms. For
two synesthetes, the photisms appeared to be located
in front of them relative to the body trunk. Thus, the
photisms would remain ‘‘in front’’ when the eyes and/
or head were turned to the side and irrespective of
where the sound comes from. For three synesthetes,
the photism would be located in line with the loca-
tion of the sound, irrespective of their own position.
For three synesthetes, the photism was reported to be
Goller, Otten, and Ward
1871
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
–
p
r
d
t
i
2
c
1
l
e
1
0
–
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
.
f
.
/
t
o
n
1
8
M
a
y
2
0
2
1
internal, in their ‘‘mind’s eye,’’ and was unaffected by
either their own position or the location of the sound.
The remaining two synesthetes reported a combination
of the above, namely, that the photism would initially
appear to come from the location of the sound but
could be shifted in to a different spatial reference frame
(relative to their body) when attended.
A modified version of the ‘‘test of genuineness’’ used
by Ward et al. (2006) was used on a set of 40 differ-
ent tones of varying pitch and timbre (10 pure tones,
10 strings, 10 piano, and 10 notes of other timbre). Par-
ticipants were required to choose the ‘‘best’’ color on
two occasions using a standard Windows color palette
(controls were encouraged to guess). Synesthetes were
significantly more consistent (i.e., smaller differences) in
their RGB selections, t(18) = 2.55, p < .05, relative to
the controls reported by Ward et al. (2006) for the same
stimuli.
Experimental Design
The experiment was based on an oddball paradigm in
which auditory or visual stimuli were presented and
participants responded to the presence of an infrequent
target that was defined in terms of either pitch or color.
This ensured that participants attended to the stimuli,
although only the frequent stimuli were analyzed. The
control participants were requested to respond to the
pitch of the auditory stimuli and the color of the visual
stimuli (i.e., two conditions). The auditory-to-visual syn-
esthetes were asked to respond to either the pitch or
the synesthetic color of the auditory stimulus or to the
color of the visual stimulus (i.e., three conditions). The
two synesthetes with bidirectional auditory–visual syn-
esthesia were given a fourth condition, namely, to
attend to the synesthetic sound of the colored stimulus.
The order of presentation of the conditions was ran-
domized. As such, our main analyses consisted of a
between-subject manipulation (presence or absence of
synesthesia) and a within-subject manipulation within
the group of synesthetes (attend auditory vs. attend
visual). This enabled us to determine when differences
between synesthetes and controls first emerge (and over
which sites) and to what extent these differences reflect
whether the synesthete was instructed to attend to the
sound or to the synesthetic vision.
Stimulus Materials
In the auditory stimulus conditions, the frequent audi-
tory stimulus consisted of five tones within a limited
pitch range. Slightly different tones were used to avoid
habituation or adaptation. The pitches of the tones were
262, 277, 294, 311, and 330 Hz (each corresponds to a
semitone difference on the Western musical scale). The
infrequent target stimulus was much higher in pitch
(1000 Hz) and was distorted with an auditory filter to
make it more distinctive. The stimuli were presented for
200 msec.
To select the colors for the visual stimulus conditions,
the sounds were played to the synesthetes in advance of
the EEG session. They used a standard Windows color
picker to choose the color that best represented their
experience to the tone. The sounds were not labeled
and were each presented twice to ensure reliability.
The order was randomized. For each sound and for
each synesthete, the two RGB values were averaged,
and this was used as the representative color for that
sound for that person. The colors were chosen in this
way to match them, approximately, to the visual reports
of their synesthesia and also to test the hypothesis that
synesthesia may be bidirectional (i.e., vision to auditory)
even in synesthetes who do not report auditory phe-
nomenology with vision. The color for the infrequent
(higher pitch) sound differed from synesthete to synes-
thete but tended to be lighter in color and was always
easily discriminable from the others. Participants were
shown the infrequent and the frequent colors before
the visual trials and informed of which one was the tar-
get. In the conditions in which visual stimuli were pre-
sented, each synesthete was presented with their own
synesthetic colors, and an age-matched control was shown
the same colors.
Procedure
In a dark cubicle, a fixation cross appeared on a black
screen for the duration of the experiment to minimize
eye movements. The screen was at a distance of 90 cm
from the participant.
In the auditory conditions, participants were pre-
sented with the tones binaurally through headphones
at intervals between 1.75 and 2.25 sec. The interval was
varied to reduce ERP components associated with prep-
aration and expectation. Participants were instructed
to press a button in response to the infrequent target.
The infrequent target was defined in terms of pitch for
controls and in terms of either pitch or color for the
synesthetes. Speed and accuracy of their responses was
recorded. The participants were first presented with
a practice block containing 10 trials, followed by two
3-min-long blocks each consisting of 80 trials, 10 of
which were infrequent targets.
The visual stimulus conditions followed the same pro-
cedure as the auditory ones, except that colored squares
were presented in place of tones and the instruction was
to respond to the infrequent target color. The colors
appeared as 2 (cid:1) 2-cm squares in the center of an other-
wise black screen (a visual angle of 1.3 degrees). We did
not predict a difference between synesthetes and con-
trols in these conditions (except in the two synesthetes
who report visual-to-auditory synesthesia) given that for
most synesthetes and all controls, colors do not elicit
sounds. However, it is also conceivable that colors could
1872
Journal of Cognitive Neuroscience
Volume 21, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
0
-
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
t
.
.
f
.
/
o
n
1
8
M
a
y
2
0
2
1
implicitly activate auditory processes in synesthetes as
similar effects have been noted in grapheme-color syn-
esthesia (Cohen Kadosh et al., 2007). Finally, the two
synesthetes who reported experiencing sounds from
color were presented with colors and were asked to at-
tend to (and respond to) the sound associated with the
infrequent color.
EEG Acquisition and ERP Analysis
The EEG was continuously recorded from 31 silver/silver
chloride electrodes. Twenty-nine of these were mounted
in an elastic cap placed according to an equidistant
montage (montage 10; www.easycap.de), and the re-
maining electrodes were situated on the right and left
mastoid processes. The recordings were made with ref-
erence to the midfrontal electrode, Fz, and were subse-
quently rereferenced off-line to the algebraic average
of the right and left mastoids. Blinks and other vertical
eye movements were monitored by recording bipolar
EOGs from a pair of electrodes placed above and below
the right eye. Horizontal eye movements were measured
by recording EOGs from electrodes on the outer right
and the left canthi. Impedances for all electrodes were
always kept below 10 k(cid:1). The EEG and the EOG sig-
nals were amplified with a bandwidth of 0.3–35 Hz (3-dB
roll-off ) and sampled at a rate of 250 Hz. The EOG ac-
tivity was visually monitored during data collection to
ensure that the participants’ eyes were kept open and
that they did not blink at regular intervals.
ERPs were averaged off-line according to condition
and electrode site for 1024 msec epochs, including a
100-msec prestimulus baseline. All ERP waveforms were
based on a minimum of 105 artifact-free trials (75% of
trials). Artifacts were both automatically detected and
monitored based on visual inspection at an individual
level without knowledge of condition. Specifically, trials
were removed if they were contaminated at any elec-
trode site by artifacts such as horizontal or nonblink
vertical eye movements, A/D saturation, or EEG drifts
larger than 50 AV. Blink artifacts were minimized by
estimating and correcting their contribution to the ERP
waveform using a standard linear regression technique
(Rugg, Mark, Gilchrist, & Roberts, 1997). Incorrect trials
were excluded from both the ERP and the behavioral
analyses.
All analyses focused on the ERPs elicited by the fre-
quent stimuli. The analyses comprised two primary
comparisons. First, ERPs elicited by tones when atten-
tion had to be directed to auditory information were
compared across synesthetes and controls. Second, in
synesthetes only, ERPs for tones were contrasted de-
pending on whether attention was paid to the auditory
stimuli or the visual experience they evoked. Additional
analyses were directed at the two bidirectional synes-
thetes. The ERPs elicited by visual stimuli in these indi-
viduals were compared with those elicited in the eight
unidirectional synesthetes and 10 controls. Auditory
ERPs were also compared across unidirectional and bi-
directional synesthetes.
RESULTS
Response Times
Participants correctly identified the infrequent stimulus
on 99.8% of trials. RTs for individual trials were removed
as outliers if they were more than three standard de-
viations above the mean RT for each subject in each
condition. The synesthetes and controls did not differ
significantly from each other for the response time
for detecting the auditory targets (synesthetes: mean =
442 msec, SD = 79; controls: mean = 419 msec, SD =
71), t(18) = .52, ns, or the visual targets (synesthetes:
mean = 497 msec, SD = 89; controls: mean = 460 msec,
SD = 74), t(18) = .78, ns. The synesthetes responded
just as fast when asked to attend to the synesthetic color
of a tone (mean = 467 msec, SD = 98) as when asked
to attend to the pitch of the tones (mean = 442 msec,
SD = 79), t(9) = 1.41, ns.
ERP Results
For the analyses of auditory stimuli, the 10 synesthetes
were treated as a single group as all reported auditory-
to-visual synesthesia. It will be shown later that the two
synesthetes who additionally report visual-to-auditory
synesthesia do not differ in their response to auditory
stimuli relative to the other eight synesthetes. For the
analyses of visual stimuli, the synesthetes are divided ac-
cording to their reported experiences.
Auditory Stimuli: Differences between Synesthetes
and Controls
At debriefing, the synesthetes did report color experi-
ences to the auditory stimuli. The group average ERP
waveforms elicited by the auditory stimuli when attend-
ing to the pitch of tones in synesthetes and controls at
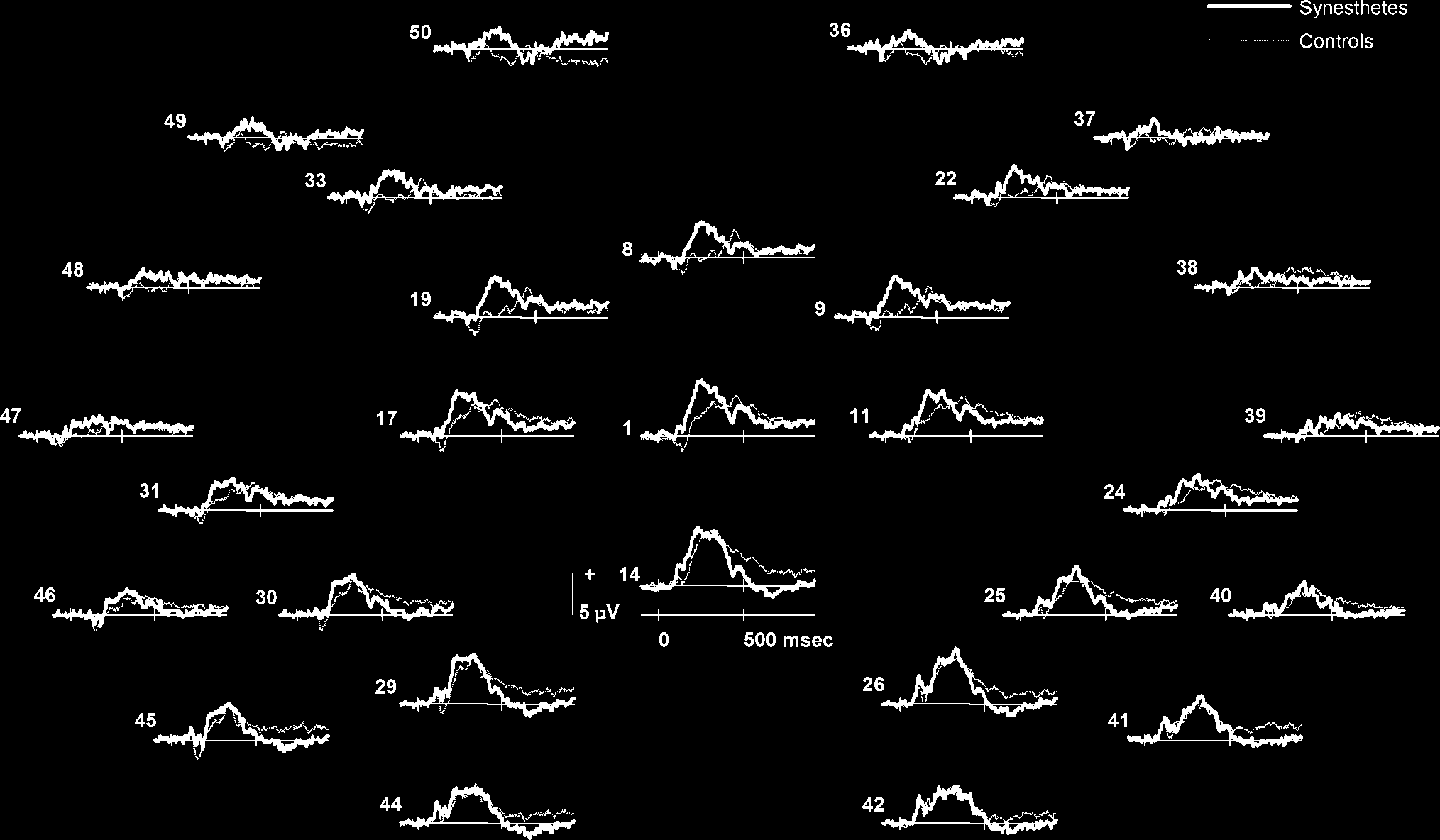
all electrode sites are presented in Figure 1. Both syn-
esthetes and controls showed identifiable N1, P2, and
N2 peaks that are characteristic of auditory-evoked po-
tentials (AEP; Picton, 1990). For both groups, the N1 was
maximal over midline frontal sites, the P2 over midline
central sites, and the N2 over frontal sites (see Figure 2).
A small positive deflection after 200 msec corresponds
to the offset of the sound. The waveforms of the con-
trols were generally more negative going than those
of synesthetes, particularly at frontocentral electrode
sites. Synesthetes elicited a smaller N1 and N2 than
controls, and a negative slow wave around 400–800 msec
apparent in the controls was virtually absent in the
synesthetes.
Goller, Otten, and Ward
1873
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
0
-
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
f
t
.
/
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 1. Group-averaged ERP waveforms at all 29 recorded electrode sites for synesthetes and controls when attending to the pitch of an
auditory stimulus (thick line = controls, thin line = synesthetes). Positive values are plotted upward.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
0
-
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
/
.
f
.
.
t
o
n
1
8
M
a
y
2
0
2
1
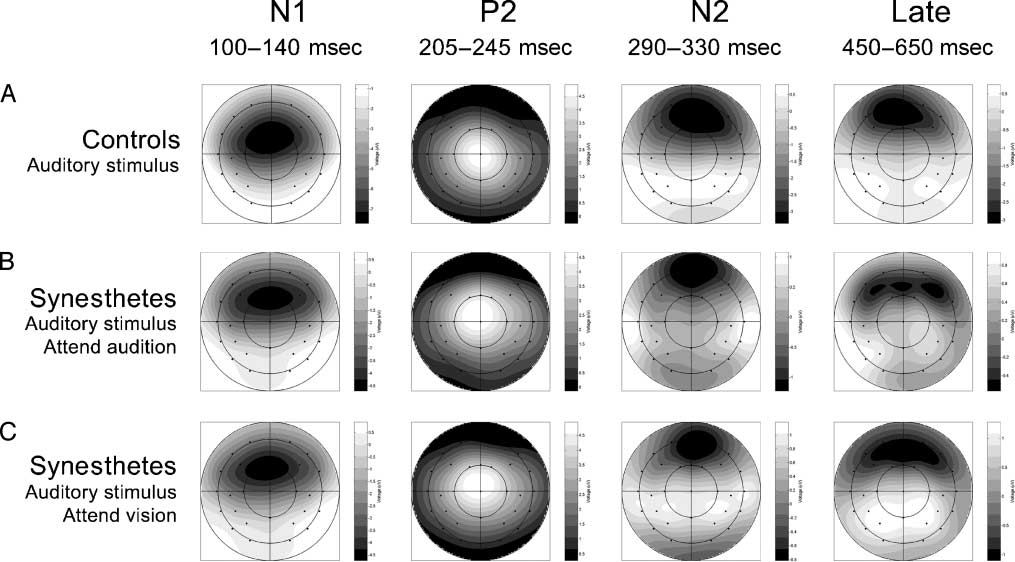
Figure 2. Voltage spline maps showing the distribution of the N1, P2, N2, and late negative deflection elicited by unimodal auditory stimuli
across the scalp. The maps are range scaled. Warm colors represent positive values; cold colors represent negative values. Maps are shown
separately for controls and synesthetes, when synesthetes attend to either the auditory stimulus or the visual illusion.
1874
Journal of Cognitive Neuroscience
Volume 21, Number 10
The analyses of the ERP data focused on the three
known AEP deflections, N1, P2, and N2. The auditory
P1 was not prominent and so was not considered. These
deflections were quantified by centering 40-msec-wide
latency regions on the deflections’ maximum peaks
based on visual inspection of the grand average (100–
140, 205–245, and 290–330 msec, respectively) and mea-
suring the mean amplitudes with relation to the mean
of the 100-msec prestimulus baseline. A late latency
region of 450–650 msec was additionally chosen for
analysis to capture the visible differences on the later
negative slow wave. The spline maps illustrating the scalp
distributions of the deflections for synesthetes and con-
trols are shown in Figure 2. The analyses were per-
formed across all 29 electrode sites to consider changes
in amplitude as well as scalp distribution.
At each latency region, 2 (cid:1) 29 ANOVAs were con-
ducted to assess the between-subjects effect of group
(synesthetes vs. controls), the within-subjects effect of
electrode site (29 levels), and the interaction between
them. All ANOVAs used the Greenhouse–Geisser cor-
rections for sphericity violations (Keselman & Rogan,
1980). Significant main effects of electrode site were
found at all latency regions ( p < .001). The ANOVA on
the N1 latency region found a main effect of group,
F(1,18) = 6.46, p = .02, but no interaction between
group and electrode site, F(1.97,35.42) = 1.35, ns. This
indicates that the amplitude of the N1 deflection was
significantly larger in controls than in synesthetes and
that this did not vary as a function of scalp location. The
ANOVAs on the P2 and the N2 latency regions demon-
strated no group main effects, F(1,18) = .17, ns and
F(1,18) = 2.55, ns, respectively, or Group (cid:1) Site inter-
actions, F(2.03,36.48) = .57, ns and F(1.73,31.10) = 3.21,
p = .06, respectively. The final ANOVA on the 450-
to 650-msec latency region revealed both a group main
effect, F(1,18) = 4.91, p = .04, and an interaction be-
tween group and electrode site, F(2.29,41.18) = 4.95,
p = .009. The interaction did not remain significant,
F(2.66,47.85) = .49, ns, after the data were normalized
using the max/min scaling procedure of McCarthy and
Wood (1985). This suggests that the interaction was due
to amplitude differences between synesthetes and con-
trols, with synesthetes showing a virtual absence of this
late negative deflection.
To look more closely for evidence of a visual potential
evoked by auditory information in synesthetes relative
to controls, we constructed difference waves at 50-msec
intervals over the four most posterior scalp locations
(sites 26, 29, 42, and 44 of montage M10). These liberal
analyses only revealed significant differences in two in-
tervals at one or two sites [site 29 between 250–300 and
350–400 msec, t(18) = 2.30 and 3.26, respectively, both
p < .034; site 26 between 350 and 400 msec, t(18) =
2.18, p < .05]. Importantly, in all instances, there was a
reduced amplitude in synesthetes. This is qualitatively
different to reports of auditory-evoked visual potentials
found in multisensory illusions (Shams et al., 2001),
in acquired synesthesia (Rao et al., 2007), and in early
infancy (Neville, 1995). Thus, there is no evidence in
the present data that tones evoke a visual potential in
synesthetes.
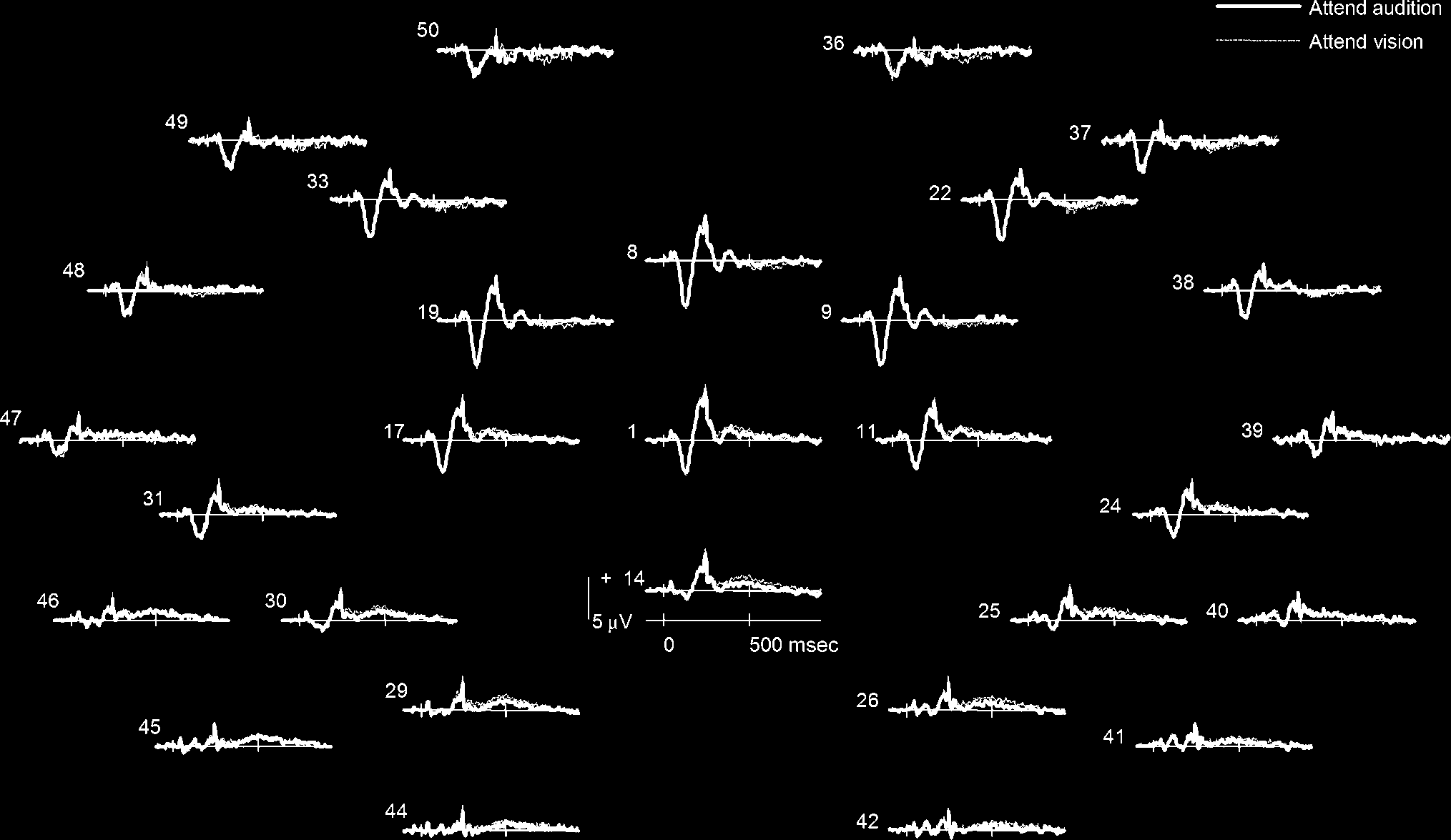
Auditory Stimuli: Effects of Attention on Synesthesia
Further analyses compared the conditions in which syn-
esthetes attended to either the auditory percept or their
visual synesthetic experience when they were presented
with a unimodal auditory stimulus. The group-averaged
ERP waveforms for these conditions are presented in
Figure 3. The first thing to note is that there are few
visible differences between these conditions. To deter-
mine any effects of attention, we carried a 2 (attend
auditory vs. visual) (cid:1) 29 (electrode sites) ANOVA out at
each latency region previously used in the analysis of
the AEP. No difference was found for attention at any
of the latency regions, and only the late latency region,
450–650 msec, demonstrated a significant interaction
between electrode site and attention, F(3.01,27.12) =
5.08, p = .006. This reflects increased positivity at more
posterior sites accompanied by increased negativity at
more anterior sites (i.e., attention increases the ampli-
tude of this late deflection). After scaling the data, this
interaction was no longer significant, F(2.32,20.88) =
1.04, ns. This suggests that there is an effect of attention,
but the effect is to modulate a deflection rather than
introduce a new deflection. All latency regions showed
an electrode site main effect ( p < .05), except for the
N2 region, F(1.96,17.65) = 2.22, ns.
These findings suggest that attention toward or
away from their synesthesia has a late modulatory in-
fluence on the ERP to auditory stimuli, but differences
between synesthetes and controls emerge far earlier (from
100 msec). These differences are thus unlikely to be
due to between-group differences in attention. The fact
that some attention-related differences were observed
implies that synesthetes were complying with instruc-
tions. A visual inspection of the data from those syn-
esthetes who reported colors in their mind’s eye (n =
3) versus elsewhere (n = 7) did not reveal extra de-
flections, although we lacked the power to assess this
statistically.
Visual Stimuli
The group-averaged waveforms for visual-evoked po-
tentials are shown in Figure 4. As with the auditory
potentials, the analysis of the visual-evoked potentials
concentrated on two known deflections, the visual P1
and the N1, together with a later positive deflection that
was particularly prominent in the synesthetes. In both
groups, the P1 was maximal at occipital sites, the N1 at
left temporal sites, and the late positive deflection at
central/parietal sites. These deflections were quantified
Goller, Otten, and Ward
1875
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
0
-
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
f
/
.
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 3. Group-averaged ERP waveforms for synesthetes when presented with an auditory stimulus and attending to either the pitch (thick line)
or the synesthetic color (thin line).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
0
-
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
/
f
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 4. Group-averaged ERP waveforms for synesthetes (n = 8) experiencing auditory–visual synesthesia (but not visual–auditory synesthesia)
and controls when attending to the color of a visual stimulus (thick line = controls, thin line = synesthetes).
1876
Journal of Cognitive Neuroscience
Volume 21, Number 10
by centering 40-msec-wide latency regions on the max-
imum peaks based on visual inspection of the group
averages (P1 = 80–120 msec; N1 = 120–160 msec; late
positive = 230–270 msec). These latency regions are in
line with those reported elsewhere (Luck, 2005). The
mean amplitudes were measured in relation to the
mean of the 100-msec prestimulus baseline. Given that
two of our synesthetes report conscious auditory ex-
periences from vision, these synesthetes were consid-
ered separately (see next section). Thus, the analyses
compared 10 controls and 8 synesthetes using a 2 (cid:1)
29 ANOVA contrasting group and electrode site. In
general, the differences between synesthetes and con-
trols were less pronounced than for auditory stimuli,
consistent with their phenomenological reports. None-
theless, some differences were found. The synesthetes
showed a reduced visual N1 deflection, which mani-
fested itself as a main effect of group, F(1,16) = 8.62,
p = .01, but the effect of group did not interact with
site. The late positive deflection showed a Group (cid:1)
Site interaction that was of borderline significance,
F(2.93,46.82) = 2.79, p = .052. There were no significant
group effects for the other deflections and no inter-
actions between group and site. The main effect of site
was significant for all three deflections. In general, the
results suggest that the visual perception of color in
synesthesia is atypical even if color does not evoke overt
synesthetic perceptions. This could either reflect implicit
bidirectionality between colors and sounds based on
their lifelong association or lack of inhibition between
areas (Cohen Kadosh et al., 2005) or more fundamental
differences in color processing. It is important for fu-
ture studies to contrast colors that are associated with
sounds relative to colors that are not to distinguish be-
tween these interpretations.
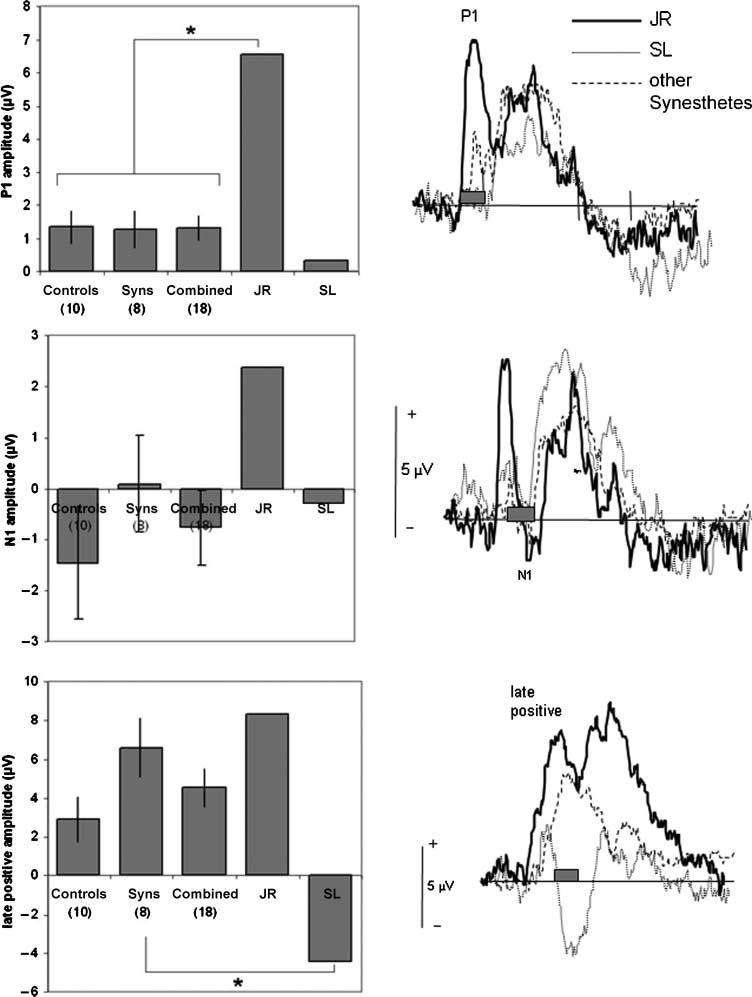
Visual–Auditory Synesthetes
For the two synesthetes who report auditory experi-
ences to color, the analyses focused on the same three
deflections as described above. However, these de-
flections were analyzed over a limited selection of elec-
trodes to minimize a type II error. The electrodes
chosen were those in which the visual-evoked potential
was noted to be maximal in the grand average of the
controls (electrodes 42, 19, and 14 for the visual P1,
N1, and late positive deflection, respectively). The mean
amplitudes for the two synesthetes were compared with
the other synesthetes (n = 8), the controls (n = 10),
and the combined group (n = 18) using the modified
t test reported by Crawford and Howell (1998). This
procedure compares a single score to a group mean,
taking into account the size of the group (the degrees
of freedom is N (cid:2) 1, where N is the size of the com-
parison group). The data are summarized in Figure 5.
One of the synesthetes, JR, had a significantly accentu-
ated P1 deflection (compared with the combined group
and synesthete group, respectively), t(17) = 3.33, p <
.001 and t(7) = 3.14, p < .05. In contrast, the other
synesthete, SL, had a significantly more negative-going
late deflection at 230–270 msec compared with the syn-
esthete group, t(7) = 2.45, p < .05. This could be due
to a longer latency of the earlier N1 deflection. The
same pattern was found when these two synesthetes
were explicitly instructed to attend to the auditory (i.e.,
synesthetic) component of the color (JR had a visual
P1 of +7.59 AV and SL had a late ‘‘positive’’ deflection
of (cid:2)2.83 AV). Thus, the basic finding is unlikely to be
due to the focus of attention per se. It is to be noted
that these two synesthetes did not stand out as anom-
alous on the auditory tasks. We reanalyzed the data
for the four auditory deflections (N1, P2, N2, and late
negative deflection) over their maximal electrode sites
comparing JR and SL to the other synesthetes for both
auditory presentation conditions (attend visual and at-
tend auditory). No significant differences were found.
As such, these two synesthetes do not appear to be
globally different. The differences that they manifest are
limited to the condition in which they report a differ-
ent perceptual experience to the other synesthetes.
This provides the first evidence for the authenticity of
this type of synesthesia. We shall consider potential rea-
sons for the discrepancy between the two vision-to-
auditory synesthetes in the Discussion section.
DISCUSSION
In summary, our results demonstrate significant electro-
physiological differences between synesthetes and con-
trols presented with unimodal auditory stimuli, in which
the auditory tone is reliably associated with a visual ex-
perience in the synesthetes but not the controls. These
differences reflect modulations of deflections of the
AEP (i.e., N1) together with a greatly attenuated late slow
negative deflection in the synesthetes. These differ-
ences are found irrespective of whether the synesthetes
were instructed to attend to the pitch of the tone or the
color of the tone (i.e., attention directed away or toward
the synesthetic experience), although there were some
late differences between these conditions. There was
no evidence of an auditory-evoked visual potential over
occipital electrode sites.
Our results differ from those showing greater electro-
physiological responses over occipital sites in response
to sounds in infants (Neville, 1995), in the ‘‘double-flash’’
illusion (Shams et al., 2001), and in a case of acquired
synesthesia following blindness (Rao et al., 2007). As
such, we suggest that a different mechanism is in opera-
tion in developmental synesthesia to that which is tapped
in these studies. That is, we suggest that developmental
synesthesia does not reflect long-range projections be-
tween early auditory and early visual areas (e.g., A1 to V1).
We also did not find evidence that auditory stimuli elic-
ited a distinct multisensory ERP deflection in synesthetes.
Goller, Otten, and Ward
1877
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
0
-
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
t
.
.
.
f
/
o
n
1
8
M
a
y
2
0
2
1
Figure 5. Group-averaged
ERP waveforms for three
electrode sites when presented
with unimodal visual stimuli
(colors). The graphs on the
left show average amplitudes
over a 40-msec time window
relative to a 100-msec
prestimulus baseline for
2 synesthetes who report
conscious experiences to tones
(SL and JR; *p < .05), for
8 synesthetes who do not
but who show the reverse
type of synesthesia (sounds
trigger colors), and for 10
control participants who
lack synesthesia. Error bars
show 1 SEM.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
0
-
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
f
.
t
.
/
.
The data thus do not strongly support the idea of cross-
model transfer in this type of synesthesia. Nonetheless,
our results are more consistent with this account. A
remarkable feature of our results is that no differences
in scalp distribution were observed for synesthetes rela-
tive to controls. If multisensory AV regions are spatially
close to those normally involved in auditory percep-
tion, this would explain the lack of a Group (cid:1) Site
interaction in addition to the significant group effects
that were observed. For instance, there could be anom-
alous cross-activation between adjacent regions of audi-
tory cortex and regions in the superior temporal guys/
sulcus that are implicated in audiovisual perception
(e.g., Calvert, 2001). It is also to be noted that tradition-
ally defined unimodal auditory areas can sometimes
respond to nonauditory events. Brosch, Selezneva, and
Scheich (2005) report that neurons in the monkey pri-
mary auditory cortex and posterior belt areas respond
to the presence of visual stimuli when the visual stimu-
lus is predictive of a subsequent auditory event (but not
in other circumstances). These neurons also respond to
unimodal auditory stimuli. That is, they have audiovi-
sual response properties rather than being strictly au-
ditory or visual. It is conceivable that neurons such as
these, which lie in or around the cortical auditory path-
ways, contribute to audiovisual synesthesia. Specifically,
synesthetes may contain far more neurons with audio-
visual response properties than unimodal auditory re-
o
n
1
8
M
a
y
2
0
2
1
1878
Journal of Cognitive Neuroscience
Volume 21, Number 10
sponses. Although direct evidence is lacking, the present
research is consistent with this view given our findings
of early modulations of the AEP. Our results are also
compatible with the idea that there is disinhibition or
unmasking of visual neurons within predominantly audi-
tory regions (Cohen Kadosh & Walsh, 2006).
Our research does not disprove the notion that in-
fants may have some form of synesthesia, but it does
raise questions about whether this type of synesthesia is
directly comparable to that found in adults even when
the synesthesia is for relatively simple pairings (pure
tones and color). Our results are entirely consistent with
a single case study of developmental synesthesia that
also recorded auditory-evoked ERPs (Rizzo & Eslinger,
1989). They are also broadly consistent with recent re-
sults reported by Beeli et al. (2008) showing reduced
amplitudes and/or longer latencies in the auditory N1
and P2 deflections for colors induced for spoken gra-
phemes and words. The question of why synesthetes
should have reduced deflections when they are report-
ing an ‘‘extra’’ experience remains to be fully explored,
but it is by no means a general feature of synesthesia
(Cohen Kadosh et al., 2007; Schiltz et al., 1999). It is
possible that the reduced negativity actually reflects an
additional presence of a positive-going deflection. It is
also possible that early auditory processing is attenu-
ated as a result of their habitual experiences of accom-
panying vision.
A novel aspect of the present study concerns the pro-
cessing of colored visual stimuli in synesthesia. For most
of the synesthetes studied here, colors do not induce
any synesthetic experiences. Nevertheless, there were
significant early (120–160 msec) differences in the visual-
evoked potential between synesthetes and controls. This
suggests that the presence of synesthesia has repercus-
sions for visual perception even when the visual stimuli
do not themselves elicit synesthesia (although there
could still be implicit bidirectionality; for example, see
Cohen Kadosh et al., 2005). This is consistent with a
recent study showing that grapheme-color synesthetes
have better perceptual discrimination of color and better
memory for colors than nonsynesthetic controls ( Yaro
& Ward, 2007), and synesthetes who experience touch
have enhanced spatial tactile discrimination even when
the task does not involve their synesthesia (Banissy &
Ward, 2008). These results suggest that there are more
fundamental differences in perceptual processing in
people with synesthesia that reveal themselves both
behaviorally and electrophysiologically rather than syn-
esthesia being solely the presence of some additional
sensory attribute.
We also report the first empirical evidence to support
the existence of vision-to-auditory synesthesia, although
this pattern has been briefly noted before for one of the
participants that we studied (Baron-Cohen et al., 1996).
The two synesthetes who report this symptom showed
abnormal visual-evoked potentials (relative to other syn-
esthetes who do not report this experience) but showed
normal AEPs relative to other synesthetes. This suggests
that they are not outliers on all measures. The two syn-
esthetes did, however, differ from each other. JR showed
a greater amplitude of visual P1 (80–120 msec) and SL
showed a negative-going later deflection (230–270 msec)
that tended to be positive in other participants. Individ-
ual differences in the timing of synesthetic experiences
have been postulated in grapheme-color synesthesia
(Ward, Salih, Li, & Sagiv, 2007; Dixon, Smilek, & Merikle,
2004) with one suggestion being that externalized color
percepts occur earlier than those reported in ‘‘the mind’s
eye.’’ This could be one explanation of the difference
that we find (JR experiences external photisms for sound-
induced vision but SL does not). For the present pur-
poses, it is sufficient to note that this type of synesthesia
can occur. It poses a potential challenge to present
theories (Hubbard & Ramachandran, 2005) that fail to
account for why it is more common in one direction
(auditory to visual) than the other (visual to auditory).
In conclusion, the present findings suggest that adult
forms of auditory–visual synesthesia with a develop-
mental origin utilizes pathways based on cross-modal
transfer in and around the normal auditory processes
rather than direct audiovisual pathways.
Acknowledgments
All research was conducted at the Institute of Cognitive Neuro-
science, University College London. Stimulus presentation was
programmed with the Cogent2000 software of the physics group
of the Wellcome Trust Centre for Neuroimaging. Both L. J. O.
and J. W. are supported by the Wellcome Trust.
Reprint requests should be sent to Jamie Ward, Department of
Psychology, University of Sussex, Falmer, Brighton, BN1 9QH,
UK, or via e-mail: jamiew@sussex.ac.uk.
REFERENCES
Banissy, M., Walsh, V., & Ward, J. (2008). ‘‘Mirror-touch’’
synaesthesia is associated with enhanced tactile
discrimination. Journal of Cognitive Neuroscience Annual
Meeting Program (Supplement). Abstract presented at 15th
annual meeting of Cognitive Neuroscience Society, p. 52.
Baron-Cohen, S. (1996). Is there a normal phase of
synaesthesia in development? Psyche, 2.
Baron-Cohen, S., Burt, L., Smith-Laittan, F., Harrison, J.,
& Bolton, P. (1996). Synaesthesia: Prevalence and
familiality. Perception, 25, 1073–1079.
Baron-Cohen, S., Harrison, J., Goldstein, L. H., & Wyke, M.
(1993). Colored speech perception: Is synaesthesia what
happens when modularity breaks down? Perception, 22,
419–426.
Beeli, G., Esslen, M., & Jancke, L. (2008). Time course of
neural activity correlated with colored-hearing synesthesia.
Cerebral Cortex, 18, 375–381.
Brosch, M., Selezneva, E., & Scheich, H. (2005). Nonauditory
events of a behavioral procedure activate auditory cortex
of highly trained monkeys. Journal of Neuroscience, 25,
6797–6806.
Goller, Otten, and Ward
1879
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
0
-
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
/
f
.
t
.
o
n
1
8
M
a
y
2
0
2
1
Calvert, G. A. (2001). Crossmodal processing in the human
brain: Insights from functional neuroimaging studies.
Cerebral Cortex, 11, 1110–1123.
Calvert, G. A., Bullmore, E. T., Brammer, M. J., Campbell, R.,
Williams, S. C. R., McGuire, P. K., et al. (1997). Activation
of auditory cortex during silent lipreading. Science, 276,
593–596.
Calvert, G. A., Hansen, P. C., Iversen, S. D., & Brammer,
M. J. (2001). Detection of audio-visual integration sites
in humans by application of electrophysiological criteria
to the BOLD effect. Neuroimage, 14, 427–438.
Cohen Kadosh, R., Cohen Kadosh, K., & Henik, A. (2007).
The neuronal correlates of bi-directional synaesthesia:
A combined ERP and fMRI study. Journal of Cognitive
Neuroscience, 19, 2050–2059.
Cohen Kadosh, R., & Henik, A. (2006). When a line is
a number: Color yields magnitude information in a
digit-color synaesthete. Neuroscience, 137, 3–5.
Cohen Kadosh, R., Sagiv, N., Linden, D. E. J., Robertson,
L. C., Elinger, G., & Henik, A. (2005). Do colors influence
numerical cognition in synaesthesia? Journal of Cognitive
Neuroscience, 17, 1766–1773.
Cohen Kadosh, R., & Walsh, V. (2006). Rewired or cross-wired
brains? Current Biology, 16, R962–R963.
Colby, C. L., & Goldberg, M. E. (1999). Space and attention
in parietal cortex. Annual Review of Neuroscience, 22,
319–349.
Crawford, J. R., & Howell, D. C. (1998). Comparing an
individual’s test score against norms derived from small
samples. Clinical Neuropsychologist, 11, 482–486.
Day, S. (2005). Some demographic and socio-cultural aspects
of synesthesia. In L. C. Robertson & N. Sagiv (Eds.),
Synesthesia: Perspectives from cognitive neuroscience.
Oxford: Oxford University Press.
Dixon, M. J., Smilek, D., & Merikle, P. M. (2004). Not all
synaesthetes are created equal: Projector vs. associator
synaesthetes. Cognitive, Affective, and Behavioral
Neuroscience, 4, 335–343.
Driver, J., & Spence, C. (2000). Multisensory perception:
Beyond modularity and convergence. Current Biology,
10, R731–R735.
Falchier, A., Clavagnier, S., Barone, P., & Kennedy, H.
(2002). Anatomical evidence of multimodal integration
in primate striate cortex. Journal of Neuroscience, 22,
5749–5759.
Frith, C. D., & Paulesu, E. (1997). The physiological basis of
synaesthesia. In S. Baron-Cohen & J. E. Harrison (Eds.),
Synaesthesia: Classic and contemporary readings.
Oxford: Blackwell.
Giraud, A. L., Price, C. J., Graham, J. M., Truy, E., &
Frackowiak, R. S. J. (2001). Cross-modal plasticity underpins
language recovery after cochlear implantation. Neuron,
30, 657–663.
Hubbard, E. M., & Ramachandran, V. S. (2005). Neurocognitive
mechanisms of synesthesia. Neuron, 48, 509–520.
Innocenti, G. M., Berbel, P., & Clarke, S. (1988). Development
of projections from auditory to visual areas in the cat.
Journal of Comparative Neurology, 272, 242–259.
Keselman, H. J., & Rogan, J. C. (1980). Repeated measures
F tests and psychophysiological research: Controlling the
number of false positives. Psychophysiology, 17, 499–503.
Lewkowicz, D., & Turkewitz, G. (1980). Cross-modal
equivalence in infancy: Auditory–visual intensity matching.
Developmental Psychology, 16, 597–607.
Luck, S. J. (2005). An introduction to the event-related
potential technique. Cambridge, MA: MIT Press.
The handbook of multisensory processes. Cambridge, MA:
MIT Press.
Mattingley, J. B., Payne, J., & Rich, A. N. (2006). Attentional load
attenuates synaesthetic priming effects in grapheme-color
synaesthesia. Cortex, 42, 213–221.
Maurer, D. (1997). Neonatal synaesthesia: Implications for
the processing of speech and faces. In S. Baron-Cohen
& J. E. Harrison (Eds.), Synaesthesia: Classic and
contemporary readings. Oxford: Blackwell.
Maurer, D., & Maurer, C. (1988). The world of the newborn.
New York: Basic Books.
Maurer, D., & Mondloch, C. J. (2006). The infant as synesthete?
Attention and performance XXI (pp. 449–471).
McCarthy, G., & Wood, C. C. (1985). Scalp distributions of
event-related potentials: An ambiguity associated with
analysis of variance models. Electroencephalography
and Clinical Neurophysiology, 62, 203–208.
Mills, C. B., Boteler, E. H., & Larcombe, G. K. (2003).
‘‘Seeing things in my head’’: A synesthete’s images for
music and notes. Perception, 32, 1359–1376.
Neville, H. J. (1995). Developmental specificity in
neurocognitive development in humans. In
M. Gazzaniga (Ed.), The cognitive neurosciences.
Cambridge, MA: MIT Press.
Nunn, J. A., Gregory, L. J., Brammer, M., Williams, S. C. R.,
Parslow, D. M., Morgan, M. J., et al. (2002). Functional
magnetic resonance imaging of synesthesia: Activation
of V4/ V8 by spoken words. Nature Neuroscience, 5,
371–375.
Paulesu, E., Harrison, J., Baron-Cohen, S., Watson, J. D. G.,
Goldstein, L., Heather, J., et al. (1995). The physiology
of colored hearing: A PET activation study of color-word
synaesthesia. Brain, 118, 661–676.
Picton, T. W. (1990). Auditory evoked potentials. In
D. D. Daly & T. A. Pedley (Eds.), Current practice of
clinical electroencephalography (2nd ed.). New York:
Raven Press.
Rao, A. L., Nobre, A. C., Alexander, I., & Cowey, A. (2007).
Auditory evoked visual awareness following sudden ocular
blindness: An EEG and TMS investigation. Experimental
Brain Research, 176, 288–298.
Rizzo, M., & Eslinger, P. J. (1989). Colored hearing
synesthesia: An investigation of neural factors.
Neurology, 39, 781–784.
Rockland, K. S., & Ojima, H. (2003). Multisensory convergence
in calcarine visual areas in macaque monkey. International
Journal of Psychophysiology, 50, 19–26.
Rugg, M. D., Mark, R. E., Gilchrist, J., & Roberts, R. C.
(1997). ERP repetition effects in indirect and direct
tasks: Effects of age and interitem lag. Psychophysiology,
34, 572–586.
Sagiv, N., Heer, J., & Robertson, L. (2006). Does binding
of synesthetic color to the evoking grapheme require
attention? Cortex, 42, 232–242.
Schiltz, K., Trocha, K., Wieringa, B. M., Emrich, H. M., Johannes,
S., & Munte, T. F. (1999). Neurophysiological aspects of
synesthetic experience. Journal of Neuropsychiatry and
Clinical Neuroscience, 11, 58–65.
Shams, L., Kamitani, Y., & Shimojo, S. (2000). Illusions.
What you see is what you hear. Nature, 408, 788.
Shams, L., Kamitani, Y., Thompson, S., & Shimojo, S.
(2001). Sound alters visual evoked potentials in
humans. NeuroReport, 12, 3849–3852.
Simner, J., Glover, L., & Mowat, A. (2006). Linguistic
determinants of word colouring in grapheme-colour
synaesthesia. Cortex, 42, 281–289.
Marks, L. E. (2004). Cross-modal interactions in speeded
Simner, J., Mulvenna, C., Sagiv, N., Tsakanikos, E., Witherby,
classification. In G. Calvert, C. Spence, & B. E. Stein (Eds.),
S. A., Fraser, C., et al. (2006). Synaesthesia: The prevalence
1880
Journal of Cognitive Neuroscience
Volume 21, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
0
-
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
f
/
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
of atypical cross-modal experiences. Perception, 35,
1024–1033.
Thornley Head, P. D. (2006). Synaesthesia: Pitch-color
isomorphism in RGB-space? Cortex, 42, 164–174.
Ward, J., Huckstep, B., & Tsakanikos, E. (2006).
Sound-color synaesthesia: To what extent does it use
cross-modal mechanisms common to us all? Cortex, 42,
264–280.
Ward, J., Salih, S., Li, R., & Sagiv, N. (2007). Varieties
of grapheme-color synaesthesia: A new theory of
phenomenological and behavioural differences.
Consciousness and Cognition.
Watkins, S., Shams, L., Tanaka, S., Haynes, J. D., & Rees, G.
(2006). Sound alters activity in human V1 in association
with illusory visual perception. Neuroimage, 31,
1247–1256.
Yaro, C., & Ward, J. (2007). Searching for Shereshevskii:
What is superior about the memory of synaesthetes?
Quarterly Journal of Experimental Psychology, 60,
682–696.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
0
-
p
1
d
8
f
6
/
9
2
1
1
9
/
3
1
7
0
5
/
0
1
0
8
o
6
c
9
n
/
2
1
0
7
0
5
9
9
7
2
8
1
1
7
3
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
4
7
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
f
.
t
.
/
o
n
1
8
M
a
y
2
0
2
1
Goller, Otten, and Ward
1881