Seeing an Auditory Object: Pupillary Light Response
Reflects Covert Attention to Auditory Space and Object
Hsin-I Liao1
, Haruna Fujihira1,2, Shimpei Yamagishi1,
Yung-Hao Yang1, and Shigeto Furukawa1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Abstract
■ Attention to the relevant object and space is the brain’s strat-
egy to effectively process the information of interest in complex
environments with limited neural resources. Numerous studies
have documented how attention is allocated in the visual
domain, whereas the nature of attention in the auditory domain
has been much less explored. Here, we show that the pupillary
light response can serve as a physiological index of auditory
attentional shift and can be used to probe the relationship
between space-based and object-based attention as well. Exper-
iments demonstrated that the pupillary response corresponds
to the luminance condition where the attended auditory object
(e.g., spoken sentence) was located, regardless of whether
attention was directed by a spatial (left or right) or nonspatial
(e.g., the gender of the talker) cue and regardless of whether
the sound was presented via headphones or loudspeakers.
These effects on the pupillary light response could not be
accounted for as a consequence of small (although observable)
biases in gaze position drifting. The overall results imply a uni-
fied audiovisual representation of spatial attention. Auditory
object-based attention contains the space representation of
the attended auditory object, even when the object is oriented
without explicit spatial guidance. ■
INTRODUCTION
The auditory world is rarely silent. It is usually full of var-
ious sounds, including background noises, and some-
times with multiple people talking at the same time.
Attention to the relevant object and space is the brain’s
strategy to effectively process the information of interest.
A massive number of studies based on behavioral mea-
surements have described how attention is distributed
in visual space, for example, as the spotlighting metaphor
(Posner, Snyder, & Davidson, 1980), the zoom-lens meta-
phor (Cave & Bichot, 1999; Eriksen & St. James, 1986), or
the gradient model (Downing, 1988). However, the
evidence for the nature of “auditory” spatial attention is
less robust and controversial (Best, Shinn-Cunningham,
Ozmeral, & Kopco, 2010; Best, Ozmeral, Kopčo, &
Shinn-Cunningham, 2008; Spence & Driver, 1994, 1996;
Mondor & Zatorre, 1995; Quinlan & Bailey, 1995; Rhodes,
1987). This makes it difficult to infer how spatial attention
functions in auditory space and for an auditory object.
Neuroimaging studies indicate that auditory spatial atten-
tion and visual spatial attention share the same neural cir-
cuit underlying the dorsal frontoparietal cortical networks
(Braga, Fu, Seemungal, Wise, & Leech, 2016; Smith et al.,
2010; Corbetta, 1998). It suggests that auditory spatial
attention operates in a way similar to visual spatial
1NTT Communication Science Laboratories, Japan, 2Japan Soci-
ety for the Promotion of Science
attention. In the current study, we aimed to demonstrate
that the internal processes related to auditory attention
can be “read out” via the pupillary light response (PLR)
as they have been shown to be in visual attention (Mathôt,
van der Linden, Grainger, & Vitu, 2013). Moreover, PLR
tracks auditory spatial attention across time, presumably
reflecting the internal processing of how attention selects
an auditory object.
Recent evidence indicates that the PLR is modulated by
neural activities from the FEF (Ebitz & Moore, 2017), and
this neural basis may explain why it reflects not only
changes in physical luminance but also top–down modu-
lated perceptual brightness (also see Strauch, Wang,
Einhäuser, Van der Stigchel, & Naber, 2022; Binda &
Gamlin, 2017). Pioneering studies have demonstrated that
pupils respond to perceived brightness even when the
physical luminance input remains the same in the case
of visual awareness to luminance in binocular rivalry
(Naber, Frässle, & Einhäuser, 2011), the bright illusion
(Suzuki, Minami, Laeng, & Nakauchi, 2019; Laeng &
Endestad, 2012), visual scene interpretation (Binda,
Pereverzeva, & Murray, 2013; Naber & Nakayama, 2013),
and mental imagery (Laeng & Sulutvedt, 2014). Remark-
ably, pupils also reflect the luminance condition of the
location to which visual attention is directed while the eyes
fixate steadily (Strauch, Romein, Naber, Van der Stigchel,
& Ten Brink, 2022; Binda, Pereverzeva, & Murray, 2014;
Mathôt, Dalmaijer, Grainger, & Van der Stigchel, 2014;
Binda et al., 2013; Mathôt et al., 2013; Haab, 1886). For
© 2022 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Journal of Cognitive Neuroscience 35:2, pp. 276–290
https://doi.org/10.1162/jocn_a_01935
instance, Mathôt et al. (2013) demonstrated that when
participants view a visual display containing luminance dis-
parity between the left and right visual hemifields (e.g.,
darkness on the left and brightness on the right, or vice
versa), pupil size is larger when they covertly attend to,
but not overtly shift the eyes to, the dark rather than bright
visual hemifield. A recent study further showed that this
attentional modulation of the PLR operates on abstract
mental content of a left-to-right spatial–numerical associa-
tion (Salvaggio, Andres, Zénon, & Masson, 2022). This
implies that the attentional modulation of the PLR does
not only serve for the anticipation of the coming percep-
tion in the visual system (Mathôt, 2018; Mathôt & Van der
Stigchel, 2015) but automatically interacts with a more
general cognitive attentional function as well. This view
also suggests the possibility of attentional modulation of
PLR in other sensory domains.
One apparent difference between visual attention and
auditory attention is that visual attention is better under-
stood in its spatial deployment, whereas auditory attention
enables us to understand more about its temporal aspect,
assuming that both operate through the same underlying
neural network (Noyce, Kwasa, & Shinn-Cunningham,
2022). Auditory objects or streams consist of acoustic fea-
tures extended through time, which can be presented in
separate or overlapped space (Shinn-Cunningham, 2008;
Fritz, Elhilali, David, & Shamma, 2007). Space is not con-
sidered an essential feature to define an auditory object.
By contrast, in natural scenes, visual objects are generally
associated with particular locations in space. Theoretically,
the location serves as the master map in visual attention
(Treisman & Gelade, 1980). Assuming the same spatial
attention mechanism operates in both the visual and
auditory domains, it would be expected that the same
attentional modulation of the PLR operates in auditory
space, particularly when the attended auditory object is
referenced by a spatial cue. However, it is unclear whether
the location information is automatically and compulsively
represented in the auditory object when it is attended and
defined via nonspatial acoustic features. Neuroimaging
studies have shown controversial evidence: Some indicate
that nonspatial auditory attention engages networks such
as the inferior frontal gyrus (Larson & Lee, 2014; Hill &
Miller, 2009); others indicate an overlapping neural circuit
between space-based and object-based auditory attention
(Bushara et al., 1999; Zatorre, Mondor, & Evans, 1999). If
nonspatial object-based auditory attention operates inde-
pendently of space-based auditory attention, it is plausible
that the auditory object can be attended and processed
without spatial attention allocated to its location. Alterna-
tively, if object-based and space-based auditory attention
share a common mechanism, spatial attention is expected
to shift to the object’s location even when the auditory
object is selected via nonspatial features. In addition, the
timing of spatial attention shifts may vary if the involve-
ment of spatial information requires further processing
depending on the circumstances.
We addressed the above issues by investigating how
PLR reflects spatial attention to auditory objects. In four
experiments, human participants listened to two concur-
rent environmental sounds or speech sentences pre-
sented dichotically to the two ears through headphones
(Experiments 1–3) or through two loudspeakers located
left and right in space (Experiment 4). They were
instructed to attend the one defined by a spatial cue (left
or right) or by a nonspatial cue, for example, the gender
of the talker (male or female). Sitting in front of a visual
display containing luminance disparity between the left
and right visual hemifields, they fixated the center of
the display while listening to the auditory stimuli. An
infrared video-based eye tracker recorded their pupillary
responses and gaze positions throughout the experi-
ments. The results consistently demonstrated that their
pupils dilated more strongly when the attended auditory
object was located on the dark side of the visual hemi-
field than when it was on the bright side. The finding
was replicated regardless of whether attention was
directed by a spatial or nonspatial cue and whether the
sound was presented via headphones or loudspeakers.
The timing of the PLR divergence (i.e., the difference
between attend-to-dark and bright conditions) occurred
earlier for the spatial cue than nonspatial cue. Local lumi-
nance differences due to gaze position drift could not
explain most of the effects. The overall results imply that
auditory attention to an object in space and visual spatial
attention recruit a common underlying mechanism.
When the auditory object is directed by the nonspatial
cue, extra time is needed to identify the auditory object’s
location before shifting spatial attention accordingly. The
finding provides profound insights into not only the neu-
ral mechanism of spatial attention across modalities but
also into brain–computer interface to predict human
auditory spatial attention by eyes.
METHODS
Participants
Seventy-four adults (54 women, age range 20–50 years,
median age 39 years) who had normal or corrected-to-
normal vision and normal hearing acuity participated in
the current study (15 in Experiment 1, 15 in Experiment 2,
27 in Experiment 3, and 17 in Experiment 4). Sample
sizes were chosen based on our previous studies with
comparable pupillometry measurements and trial numbers
per participant (Liao, Kashino, & Shimojo, 2021; Liao,
Kidani, Yoneya, Kashino, & Furukawa, 2016). In Experi-
ment 3, the number of trials in the critical dichotic condi-
tion for each participant was half of that in Experiment 2,
because of adding a diotic sound presentation condition
(see Design and Procedure for details). To have a similar
number of critical trials, more participants were recruited.
One participant in Experiment 4 was excluded because
of the < 50% accuracy in task performance (46.9%). All
were naive about the purpose of the study. The current
Liao et al.
277
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
study was approved by the NTT Communication Science
Laboratories ethics committee. All participants gave
written informed consent before the experiment and
received payment for their participation.
Overview of Experiments
Experiment 1 was modified from Mathôt et al. (2013) by
changing the main task to an auditory task to discrimi-
nate two environmental sounds. Experiments 2, 3, and
4 used spoken sentences. Participants dichotically lis-
tened to two sentences spoken by different talkers, one
male and one female, and attended the one according to
the instruction. In Experiment 2, the target sentence was
defined by space and the gender of the talker. In Exper-
iments 3 and 4, the target sentence was defined by gen-
der. In Experiment 3, half of the target sentences were
presented dichotically, and the other half was presented
diotically. In Experiment 4, the auditory stimuli were pre-
sented via two loudspeakers instead of the headphones
as in Experiments 1–3.
All the experiments had a general common structure as
follows. A trial started with presentation of a visual display
with its left and right hemifields dark and bright, respec-
tively, or vice versa. After 3 sec as an adaptation period, a
“voice cue” was presented to indicate the target dimension
(i.e., space or gender) in Japanese, followed by a “silent
period” (Experiment 1) or a “listening period” (Experi-
ments 2–4). This was followed by a “response period”
wherein participants were asked to discriminate the envi-
ronmental sound as soon and accurately as possible
(Experiment 1) or to recall the content of the target sen-
tence with no time pressure (the others). Participants
were given a written and oral explanation about the nature
of the auditory task and performed several practice trials to
familiarize themselves with the task.
Apparatus and Stimuli
Auditory and visual stimuli were generated with MATLAB
(The MathWorks, Inc.) and PsychToolbox (Kleiner,
Brainard, & Pelli, 2007; Brainard, 1997; Pelli, 1997), con-
trolled by a personal computer (Dell OptiPlex 755). Visual
stimuli were presented on an 18.1-in. monitor (Eizo
FlexScan L685Ex) with a 60-Hz frame rate and a 1280 ×
1024 resolution in Experiments 1, 2, and 3 and a 1024 ×
768 resolution in Experiment 4. There were two types of
visual displays with luminance disparity between the left
and right visual fields: black (0.35 cd/m2) on the left
and white (94.04 cd/m2) on the right, and vice versa.
In Experiment 1, a gray fixation cross (0.5° × 0.5°,
21.04 cd/m2) was presented at the center of the visual
display against the background with luminance disparity.
In Experiments 2, 3, and 4, a gray vertical band (2.3° in
width, 21.04 cd/m2) acted as a “buffer zone” that was
superimposed at the center against the background with
luminance disparity to lessen the sharp luminance
variation in fixation. A Gabor patch (0.5° × 0.5°), which
was generated by superimposing a Gaussian and a sine-
wave function with a vertical orientation (10 cycles per
degree) and presented at the center, served as the fixation
point.
Auditory stimuli were presented through headphones
(Sennheiser HD 595) in Experiments 1, 2, and 3 and
through two custom-built loudspeakers in Experiment 4.
The loudspeakers were positioned so that their centers
were located 70 cm left and right of the fixation point on
the visual display, and the surfaces of the display and the
loudspeakers were aligned. The voice cues were recorded
by a female native-Japanese speaker in our laboratory.
They had a duration of 400–570 msec and were presented
diotically (identical signal to both ears). The target sounds
used in Experiment 1, a dog barking and a phone ringing,
were sampled from a database on a compact disc (Audio
Pro Sound Effects by Yannick Chevalier) and edited to be
500 msec in length. In Experiments 2, 3, and 4, the target
sentences were sampled from the coordinate response
measure (CRM) corpus recorded in our laboratory,
Japanese version with a slight variation in the keywords
of Bolia, Nelson, Ericson, and Simpson (2000). Each
CRM sentence consisted of three Japanese keywords,
namely, an animal name (KUMA [bear], SHIKA [deer],
TORA [tiger], INU [dog], NEKO [cat], TORI [bird], SARU
[monkey], or BUTA [pig]), a color (AKA [red], AO [blue],
SHIRO [white], KURO [black], MOMO [pink], or NIJI
[rainbow]), and a number (ZERO [zero], ICHI [one],
SAN [three], YON [four], ROKU [six], NANA [seven],
HACHI [eight], KYUU [nine], or JUU [ten]). The sentences
were spoken by 10 different native-Japanese talkers (five
of each gender) in a 2.5- to 3-sec length. The sounds were
presented at a comfortable listening level, self-adjusted
by each participant.
During the response period, participants viewed the
fixation display with luminance disparity in Experiment 1.
In Experiments 2, 3, and 4, there were three types of
response displays, corresponding to the three keywords,
namely, an animal name, color, and number. The animal-
response display consisted of eight patches (2.5° × 1.3°),
each labeled with animal names. The color-response dis-
play consisted of six patches, each labeled with color
names. The number-response display consisted of nine
patches, each labeled with a number. All the labels were
written in Japanese Katakana.
Eye movements including pupillary responses were
recorded binocularly with an infrared eye-tracker camera
(Eyelink 1000 Plus Desktop Mount, SR Research Ltd.). The
camera was positioned below the monitor. The sampling
rate of the recording was 1000 Hz. Participants sat in front
of the monitor at an 80-cm distance with their head sup-
ported on a chin rest. Before each formal experimental
session, they went through the 5-point Eyelink calibration
program to calibrate and validate their eye data. After the
calibration, they were instructed to fixate the central fixa-
tion point while performing the auditory task.
278
Journal of Cognitive Neuroscience
Volume 35, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Design and Procedure
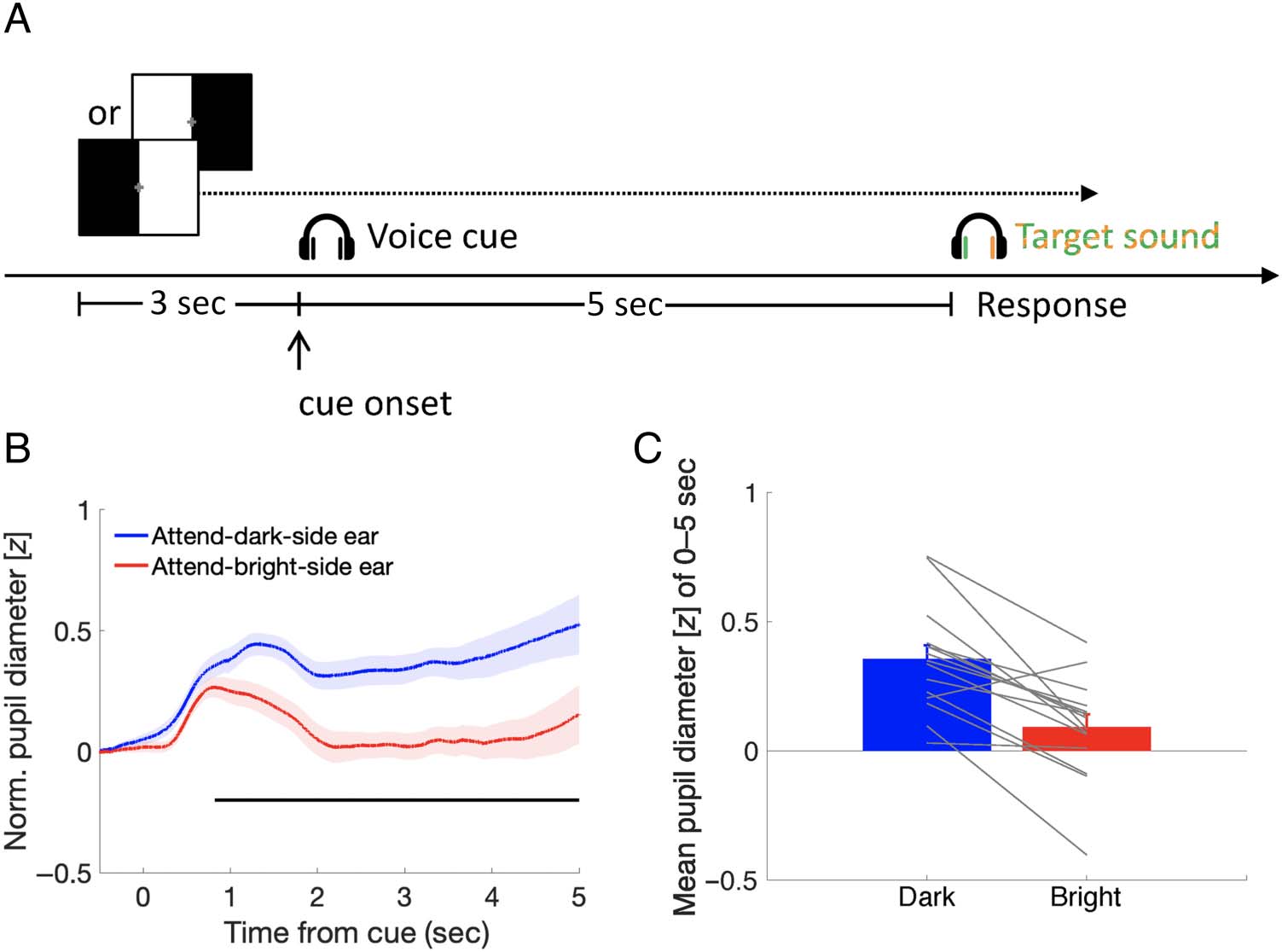
In Experiment 1 (Figure 1A), the voice cue indicating
“left” or “right” was presented, followed by a 5-sec silent
period. In the response period, the target sounds (i.e.,
dog barking and phone ringing) were presented dichoti-
cally (one to each ear). Participants were asked to
shift their attention to the left or right ear, according to
the voice cue’s instruction, and discriminate whether
the sound presented in the cued direction was “dog
barking” or “phone ringing” by pressing a designated
key on the keyboard as soon and accurately as possible.
The correspondence of the response key and target
sound was counterbalanced across the participants: Half
of the participants pressed the “M” key for “dog barking”
and the “N” key for “phone ringing,” and the other half
did the opposite. Once they pressed the response key,
the next trial started. The visual display disparity (black
on left or right), the voice cue (left or right), and the tar-
get sound (dog barking sound to the left or right ear)
were counterbalanced across trials. There were 48 trials
in each block. Each block took about 15 min, and the
participants performed two blocks with a 15-min break
in between.
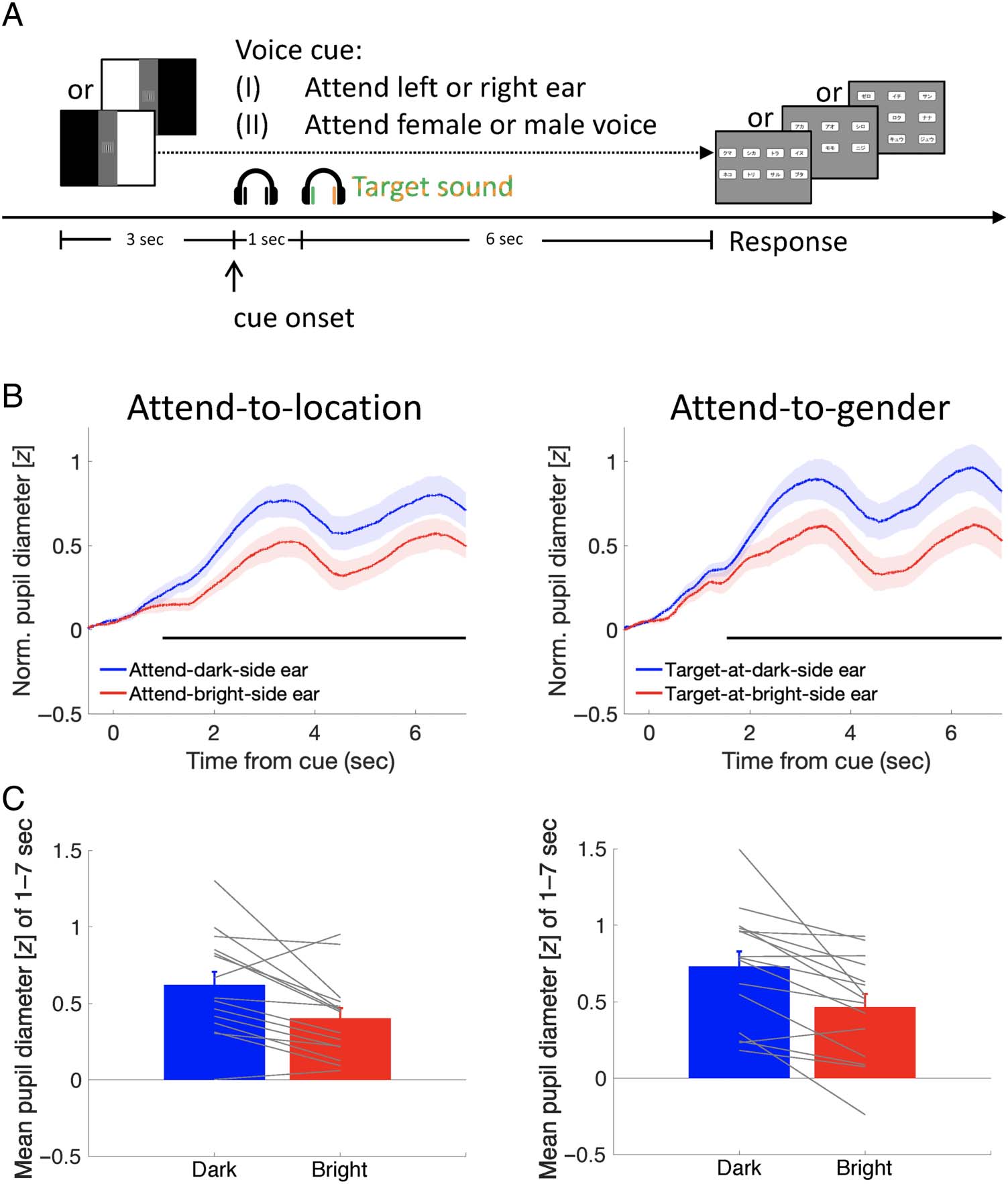
In Experiments 2, 3, and 4, 1 sec after the voice cue pre-
sentation, the listening period started, in which the two
target sentences were presented dichotically (Figure 2A).
The target sentences were selected so that the contents of
the three keywords would not overlap. Each sentence was
repeated twice, resulting in a total duration of 6 sec. Partic-
ipants were asked to pay attention to the target sentence,
memorize its content, and recall it during the response
period. With no time pressure, they gave the answer by
using the mouse to click the patch in the response display,
which was randomly selected from the three types of
response displays for each trial. Participants did not know
in advance the response set (animal, color, or number) for
each trial and thus had to memorize the entire content to
perform the task.
Experiment 2 consisted of two conditions, namely, the
attend-to-location and attend-to-gender conditions. In the
attend-to-location condition, the voice cue was on the spa-
tial dimension (“left” or “right”). Participants paid atten-
tion to the sentence presented at the cued direction, as
in Experiment 1. In the attend-to-gender condition, the
voice cue was on the dimension of the talker’s gender
(“male” or “female”). Participants paid attention to the
sentence spoken by the male talker if the voice cue said
“male” beforehand or to the one spoken by the female
talker if the voice cue said “female” beforehand. The visual
display disparity, the voice cue (left or right in the attend-
to-location condition; male or female in the attend-to-
gender condition), and the target sound (the male’s
spoken sentence to the left or right ear) were counterba-
lanced across trials. There were 64 trials in each block.
Participants performed the two types of attention
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Experimental
procedure and pupillary
response results in Experiment 1.
(A) Schematic procedure.
The visual display with two
different luminance disparities
was randomly presented for
each trial. Participants paid
attention to their left or right
ear, depending on the voice
cue, and responded by
indicating whether the cued
target sound was a “dog
barking” or a “phone ringing”
as soon and accurately as
possible while looking at the
central fixation cross for
pupillary response recording.
(B) Pupillary response as a
function of time from the
voice cue onset, parameterized
with the cue–luminance
associations (attend dark or
bright side). The shaded
area represents standard
errors across participants.
The horizontal black line
indicates a significant difference
between the attend-dark
and attend-bright conditions (nonparametric cluster-based permutation tests, p < .05) during the period of 821–5000 msec. (C) Mean pupil diameter
as a function of the cue–luminance associations. Error bars represent standard errors among participants. Gray lines represent data of individual
participants. Norm. = Normalized.
Liao et al.
279
Figure 2. Experimental
procedure and pupillary
response results in Experiment 2.
(A) Schematic of procedure.
The visual display with two
different luminance disparities
was randomly presented for
each trial. The center vertical
gray band was presented as a
“buffer” zone. Participants
paid attention to the target
sentence, memorized its
content, and recalled it after
the sound presentation.
The target sentence was
defined by a spatial cue (I)
or a nonspatial cue (II).
(B) Pupillary response as a
function of time from the
voice cue onset in the spatial
(i.e., attend-to-location) and
nonspatial (i.e., attend-to-
gender) cue conditions,
parameterized with the cue–
luminance associations (attend
dark or bright side; left) or the
target–luminance associations
(target located on the dark or
bright side; right), respectively.
The shaded area represents
standard errors across
participants. The horizontal
black lines indicate a significant
difference between the
attend-dark and attend-bright
conditions (nonparametric
cluster-based permutation tests,
p < .05) during the period
of 978–7000 msec in the left
panel and the difference
between the target-at-dark and
target-at-bright conditions
during that of 1542–7000 msec
in the right panel. (C) Mean
pupil diameter as a function
of the cue–luminance associations (left) or target–luminance associations (right). Error bars represent standard errors among participants.
Gray lines represent data of individual participants. Norm. = Normalized.
conditions in separate sessions, with the order counter-
balanced across participants. Each attention condition
consisted of two blocks with a short break in between.
In Experiment 3, the design and procedure were the
same as in Experiment 2 except for the following. First,
only the gender voice cue (male or female) was used.
Second, the target sentences were presented dichotically
as well as diotically, where the two target sentences were
mixed into one signal and presented to the two ears. The
two types of target sound presentation (dichotic, diotic)
were presented in randomized order within a block.
There were 32 trials for each target sound presentation
type, resulting in 64 trials in each block. Participants per-
formed two blocks with a break in between. Experiment 4
followed the same design and procedure of the attend-to-
gender condition in Experiment 2, except that all the
auditory stimuli were presented via speakers instead of
headphones. Participants only performed one block.
Behavioral Data Analyses
In all experiments, trials with incorrect answers were
excluded from further analyses (1.7%, 3.4%, 4.4%, and
1.6% of trials excluded in Experiments 1, 2, 3, and 4,
respectively). Further exclusion criterion was set for
Experiment 1: Trials with an RT exceeding two times the
standard deviation from the mean within each session
were excluded (4.3% of trials). No RT criterion was set
for Experiments 2–4 because participants performed the
task with no time pressure.
280
Journal of Cognitive Neuroscience
Volume 35, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Eye Metrics Data Analyses
Because eye movements and pupillary responses are con-
sensual, only the data from the left eye were used. During
the silent or listening period, blinks accounted for 12.3%
of data points in Experiment 1, 12.2% in Experiment 2,
12.8% in Experiment 3, and 12.3% in Experiment 4. The
missing pupil-diameter data during blinks were interpo-
lated by using shape-preserving piecewise cubic interpola-
tion. To compare the pupillary response results across
participants and conditions, pupil diameter data were
normalized by z-transform using all the data recorded in
each block and then baseline-corrected trial-by-trial by
subtracting the mean of the data during the 1-sec period
before the cue onset.
Statistical Analysis
Frequentist and Bayesian Statistics
Both frequentist and Bayesian statistics were performed
on average data, including RTs (Experiment 1), accuracies,
and mean pupil diameters during the silent period (Exper-
iment 1) or the listening period (Experiments 2–4). The
Bayesian statistical analyses were computed by using the
open-source program JASP ( Jeffreys’s Amazing Statistic
Program; jasp-stats.org), referenced by Keysers, Gazzola,
and Wagenmakers (2020). Detailed parameters and tests
for each experiment are described below.
Behavioral Performance
In Experiment 1, mean RTs and accuracies were subjected
to repeated-measure ANOVAs with Display Disparity
(black on left or right), Cue Direction (left, right), and
Target Type (dog barking or phone ringing) as within-
subject factors. In Experiments 2 and 3, mean accuracies
were subjected to paired two-sample t tests on the com-
parison between attend-to-location and attend-to-gender
conditions (Experiment 2) and on the comparison
between dichotic and diotic conditions (Experiment 3).
For between-experiment comparison, mean accuracies
in the attend-to-gender condition in Experiment 2 and
mean accuracies of the dichotic trials in Experiment 3 were
subjected to independent samples t test.
repeated-measure ANOVA with the Target–Luminance
Associations (dichotic trials with the target located on
the dark or bright side, or diotic trials with the target inter-
laced at the center) as a within-subject factor. Between-
experiment comparison was conducted as follows: Mean
pupil diameters in Experiment 2 (only in the attend-to-
gender condition) and Experiment 3 (only the dichotic
presentation trials) were subjected to a mixed-design
ANOVA with the Target–Luminance Associations (target
located at the dark or bright side) as a within-subject factor
and Experiment as a between-subject factor.
Eye Metrics Data across Time
Pupil diameter change and luminance of the gazed posi-
tion were subjected to nonparametric cluster-based per-
mutation tests (Maris & Oostenveld, 2007) to examine
the difference between the conditions across time. Lumi-
nance contrast of the gazed position was smoothed by
moving averaging with a 100-msec time window for each
participant. This was done to reduce the noise across time.
The cluster-based analyses were computed by using the
Fieldtrip MATLAB toolbox (Oostenveld, Fries, Maris, &
Schoffelen, 2011) with 5000 iterations and both the
cluster-defining height threshold and FWE-corrected clus-
ter size threshold below an α level of .05.
Linear Mixed-Effects Models
Linear mixed-effects (LME) analyses were performed
at each 1-msec sampling point with luminance of the gazed
position (dark, bright in Experiment 1; dark, bright, gray in
Experiments 2–4) and attended luminance (cue–luminance
or target–luminance association as dark, bright) as fixed
effects, participant as the random effect, and pupil diameter
as the dependent variable. The LME analyses were com-
puted by using the lem4 package in R (Bates, Mächler,
Bolker, & Walker, 2015). Following a criterion similar to
Mathôt et al. (2014), the significant clusters were defined
as a t value > 2 with at least 500 consecutive samples.
Unlike Mathôt et al. (2014), who chose 200 consecutive
samples, we set the criterion stricter with larger consecu-
tive samples because the gazed luminance was noisy
across time.
Attention-Based PLR Bias
Mean pupil diameters during the silent (i.e., 0–5 sec time-
locked to the cue onset in Experiment 1) or listening (i.e.,
1–7 sec time-locked to the cue onset in Experiments 2–4)
period were subjected to paired two-sample t tests with
the cue–luminance associations (attend the dark or bright
side) for the space cue (Experiments 1 and 2) and with
the target–luminance associations (target located at the
dark or bright side) for the gender cue (Experiments 2–
4) as the independent variables. In Experiment 3, mean
pupil diameter data were further subjected to a
PLR Divergence Latency
The latency was defined as the earliest end of the period
with the cluster-based significant difference between the
attend-to- (or target-at-) dark side and attend-to- (or
target-at-) bright side conditions. Following the jackknife
resampling technique, the latency of the divergence differ-
ence and its variance were estimated. Pupil size difference
between the two attended luminance conditions was sub-
jected to nonparametric cluster-based permutation tests
by omitting the data from one participant. To guarantee
that the effective clusters could be identified for all
Liao et al.
281
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
participants, the threshold was set to be the smallest α
level or the second smallest α level if the difference
between them was smaller than .05. The procedure was
repeated until all participants were omitted once. The esti-
mated latencies were subjected to both frequentist and
Bayesian statistics. Within-experiment comparison was
conducted in Experiment 2, in which the estimated
latencies were subjected to a paired two-sample t test with
the attention condition (attend-to-location, attend-to-
gender) as the independent variable. Between-experiment
comparisons were conducted for the attend-to-location
and attend-to-gender conditions separately. For the
attend-to-location condition, the estimated latencies in
Experiment 1 and those of the attend-to-location condi-
tion in Experiment 2 were subjected to an independent
sample t test with the experiment as the grouping
variable. For the attend-to-gender condition, the esti-
mated latencies in Experiments 3 and 4 and those of
the attend-to-gender condition in Experiment 2 were
subjected to ANOVAs with the Experiment as the fixed
factor.
RESULTS
Experiment 1: PLR Reflects the Direction of
Endogenous Auditory Spatial Attention
We started by replicating the paradigm of Mathôt et al.
(2013), but with a modification of the main task to dis-
criminate auditory objects (Experiment 1). Participants
listened to an auditory voice cue saying “left” or “right”
to shift their attention to the left or right ear, respectively.
After 5 sec, two environmental sounds, a dog barking
and a phone ringing, were presented for 500 msec
dichotically through headphones. They responded by
pressing a designated key to report whether the sound
presented in the cued ear was “dog barking” or “phone
ringing” as soon and accurately as possible (see Figure 1A
for procedure).
Behavioral results showed that RTs and accuracies were
equally fast and accurate among all the conditions (see
Table 1). For RTs, the three-way interaction of Visual Display
disparity (darkness at the left or right visual hemifield), Cue
Direction (left or right), and Target Sound (dog barking or
phone ringing) showed significance, F(1, 14) = 4.87, p =
.045, whereas the Bayes factor (BF) with the model
including only the three-way interaction was very small
(BFincl = 0.70). None of the other effects or interactions
were significant (Fs < 3.6, ps > .08, 0.1 < BFsincl < 0.6).
For accuracies, none of any main effects or interactions
were significant (Fs < 1.8, ps > .2, 0.2 < BFsincl < 0.7).
Most importantly, mean pupil size was larger when
the target sound was presented in the cued direction
where the visual hemifield was dark than when it was
bright, t(14) = 5.32, p < .001, BF+0 = 530.19, median
δ = 1.24, 95% CI [0.538, 1.976] (Figure 1C). This differ-
ence was significant 821 msec after the voice cue onset
( p < .05, nonparametric cluster-based permutation test;
see Figure 1B).
Experiment 2: PLR Reflects Continuous Covert
Attention to Auditory Object in Space
In Experiment 1, the spatial attention effect reflected in
the PLR was observed during the period in which no
auditory stimulus was presented. It could be argued that,
during this period, the participant’s spatial attention was
not directed to an auditory object but to the visual dis-
play, and thus the present result is a mere replication
of the experiment on visual attention (Mathôt et al.,
2013). We examined this possibility in Experiment 2, in
which the target sound was changed to a continuous
speech sentence presented for 6 sec, and we investigated
whether and how the PLR effect was observed during the
target sound presentation period. The stimuli for each
trial were two sentences randomly sampled from a
Japanese version of the CRM corpus (Bolia et al., 2000).
Versions of the CRM corpus have been widely used to
study speech intelligibility in competing speech or noise,
and each sentence in the corpus consists of three key-
words, namely, an animal name, color, and number in
the Japanese version. In our procedure (see Figure 2A),
two sentences by two talkers, one male and one female,
were presented dichotically through headphones. In the
attend-to-location condition, the voice cue saying “left” or
“right” (as in Experiment 1) was presented 1 sec before
the target sentence. Participants paid attention to the
cued direction (or ear), memorized the keywords in the
target sentence presented at the cued direction, and later
recalled them. In the attend-to-gender condition, the
voice cue said “male” or “female,” and participants
recalled the content of the sentence expressed by the
Table 1. Mean RTs (msec) and Accuracies (in Parentheses) Under Each Condition in Experiment 1
Cue-Left
Cue-Right
Black-left
Black-right
Phone
1128 (99%)
1151 (98%)
Dog
1149 (97%)
1188 (98%)
Phone
1213 (98%)
1144 (99%)
Dog
1154 (99%)
1178 (99%)
282
Journal of Cognitive Neuroscience
Volume 35, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
cued-gender talker. Participants performed these two
conditions in separate blocks, in a counterbalanced order
across participants.
Behavioral performance was equally good in these two
conditions (mean accuracy = 96.1% and 97.1% for the
attend-to-location and attend-to-gender conditions,
respectively, t(14) = 1.81, p = .092, BF10 = 0.97, median
δ = −0.396, 95% CI [−0.916, 0.092]. Critically, in both
conditions, mean pupil size was larger when the target
sentence appeared in the dark visual hemifield than
when it appeared at the bright one: t(14) = 3.43, p =
.004, BF+0 = 23.28, median δ = 0.775, 95% CI [0.217,
1.385], and t(14) = 3.97, p = .001, BF+0 = 58.34, median
δ = 0.907, 95% CI [0.306, 1.553], in the attend-to-location
and attend-to-gender conditions, respectively (Figure 2C).
The difference reached significance 978 msec after the
spatial cue (left or right) and 1542 msec after the nonspa-
tial cue (male or female; Figure 2B).
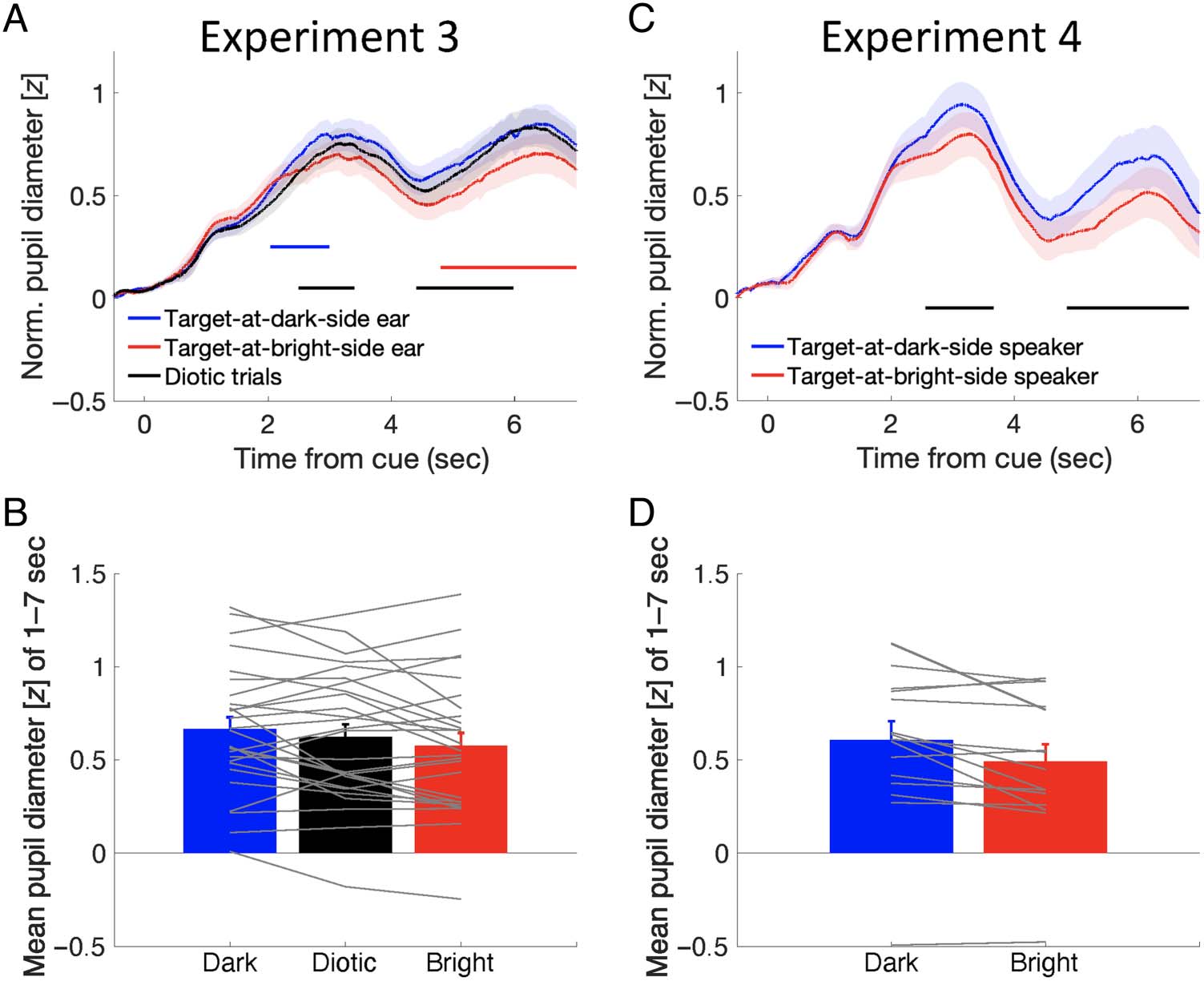
Experiment 3: Does PLR-Reflecting Object-Based
Auditory Attention Require Consistent Association
with Target Space?
An interesting finding of the above experiment (Experi-
ment 2) was that PLR reflected attention even to a nonspa-
tially guided auditory target. This suggests compulsory
involvement of space information in object-based auditory
attention. It should be noted, however, that the target
sounds were always presented in separate locations (or
ears) in that experiment. Thus, it is arguable that the
participants might have strategically used the spatial rep-
resentation of the target sound predominantly even when
the target sound was defined by a nonspatial cue and that
the observed PLR effectively reflected an intention of
attention to space. Experiment 3 was designed to discour-
age the participant from taking this strategy. We mixed trials
with dichotic stimulus presentation (as in Experiment 2)
and those with diotic presentation in a random order
within a block. In the diotic presentation trial, the two
talkers’ sentences were mixed into one signal, which was
presented to the two ears. In this case, the two voices are
not perceptually lateralized, so shifting spatial attention
toward the left or right direction would not help in the
task. We expected that the modified procedure would
encourage the participants to consistently pay more atten-
tion to the nonspatial acoustic characteristics to perform
the task.
Behavioral results showed that the accuracies of dich-
otic trials in Experiment 3 were as good as in the attend-
to-gender condition in Experiment 2 (96.6% vs. 97.1%,
t(40) = 0.28, p = .784, BF10 = 0.32, median δ = 0.067,
95% CI [−0.486, 0.633]), and they were better than
those of the diotic trials in Experiment 3 (96.6% vs.
93.5%, t(26) = 6.16, p < .001, BF10 = 11321.85, median
δ = 1.115, 95% CI [0.625, 1.621]).
Critically, mean pupil sizes were different among the
three conditions we tested (i.e., dichotic trials with target on
the dark side, dichotic trials with target on the bright side, and
diotic trials), F(1.54, 40.15)with Greenhouse–Geisser correction =
3.74, p = .043, BFM = 1.73. Post hoc multiple comparisons
indicated that mean pupil size was larger when the target
sound was located in the dark hemifield than when it was
located in the bright one ( pBonferroni = .026, BF+0 = 3.11,
median δ = 0.389, 95% CI [0.062, 0.772]), but no significant
difference was found for the other two pairs ( psBonferroni >
.4, BFs+0 < 0.94; Figure 3B). The difference among these
conditions was found during the periods of 2036–3403
and 4408–7000 msec (Figure 3A). The segment of two
significant periods corresponds to how the target sen-
tences were presented, namely, twice with a short break
in between. The result suggests that the allocation of spa-
tial attention to the auditory object matches the temporal
dynamic of the auditory object’s presentation.
We compared the PLRs in comparable conditions
between Experiments 2 and 3, in which the gender-cue-
guided target and the two sentences were presented
dichotically. The purpose of this comparison was to
examine whether the PLR-reflecting object-based atten-
tion was modulated by the consistency of the availability
of the sound space information. Mean pupil sizes in the
dichotic trials in Experiment 3 and those in the attend-
to-gender condition in Experiment 2 were subjected to a
mixed-design ANOVA with the Target Sound Location
(located in the dark or bright hemifield) as a within-subject
factor and Experiment as a between-subject factor. Consis-
tent with the previous analyses conducted for individual
experiments, mean pupil size was overall larger when
the target sound was located in the dark than bright
hemifield, F(1, 40) = 23.22, p < .001, BFincl = 112.77.
Mean pupil size did not differ between the two experi-
ments, F(1, 40) = 0.04, p = .849, BFincl = 0.48. Impor-
tantly, the PLR effect was stronger in Experiment 2 than
Experiment 3, indexed by the interaction between the
target sound location and experiment, F(1, 40) = 5.75,
p = .021, BFincl = 2.27.
The overall results indicate that PLRs reflected spatial
auditory attention even when a nonspatial cue guided
attention. However, the PLR bias became weaker when
the spatial representation of the sound source did not
always help for the task at hand (as in Experiment 3).
Experiment 4: Attention-Related PLRs to Sounds
from Loudspeakers in Space
It can be argued that the attention-related PLRs reflected
the attention directed to the ear, not the space, per se, in
the previous three experiments, in which all the sounds
were presented via headphones. To test for this possibil-
ity, in Experiment 4, we replicated the attend-to-gender
condition in Experiment 2 but presented stimuli via loud-
speakers, which were located on either side of the visual
display. Participants performed the task with the same
accuracy as in Experiment 2 (98.4% vs. 97.1%, t(29) =
Liao et al.
283
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Pupillary response
results in Experiments 3 and 4.
(A) Pupillary response as a
function of time from the voice
cue onset, parameterized
with the target–luminance
associations (dichotic trials with
the target located on the dark
or bright side, or diotic trials).
The shaded area represents
standard errors across the
participants. The horizontal
black line indicates a significant
difference between the target-
at-dark and target-at-bright
conditions during periods of
2493–3403 and 4408–5986 msec.
The horizontal blue line
indicates a significant difference
between the target-at-dark and
diotic conditions during the
period of 2036–2997 msec. The
horizontal red line indicates a
significant difference between
the target-at-bright and diotic
conditions during that of 4803–
7000 msec. (B) Mean pupil
diameter as a function of the
target–luminance associations.
Error bars represent standard
errors among participants.
Gray lines represent data of
individual participants. (C) Pupillary response as a function of time from the voice cue onset, parameterized with the target–luminance associations
(target located at the dark or bright side). The shaded area represents standard errors across participants. The horizontal black lines indicate a
significant difference between the target-at-dark and target-at-bright conditions during the periods of 2559–3669 and 4856–6831 msec. (D) Mean
pupil diameter as a function of the target–luminance associations. Error bars represent standard errors among participants. Gray lines represent data
of individual participants. Norm. = Normalized.
1.27, p = .216, BF10 = 0.62, median δ = −0.340, 95% CI
[−1.016, 0.270]). Critically, the pupillary response results
showed that mean pupil size was larger when the target
sound came from the loudspeaker located next to the
dark visual hemifield than from the one next to the bright
one, t(15) = 3.04, p = .008, BF+0 = 12.62, median δ =
0.666, 95% CI [0.161, 1.227] (Figure 3D). The difference
reached significance 2559–3669 and 4856–6831 msec
after the cue presentation (Figure 3C). The segment of
the two periods showed a pattern similar to the result in
Experiment 3.
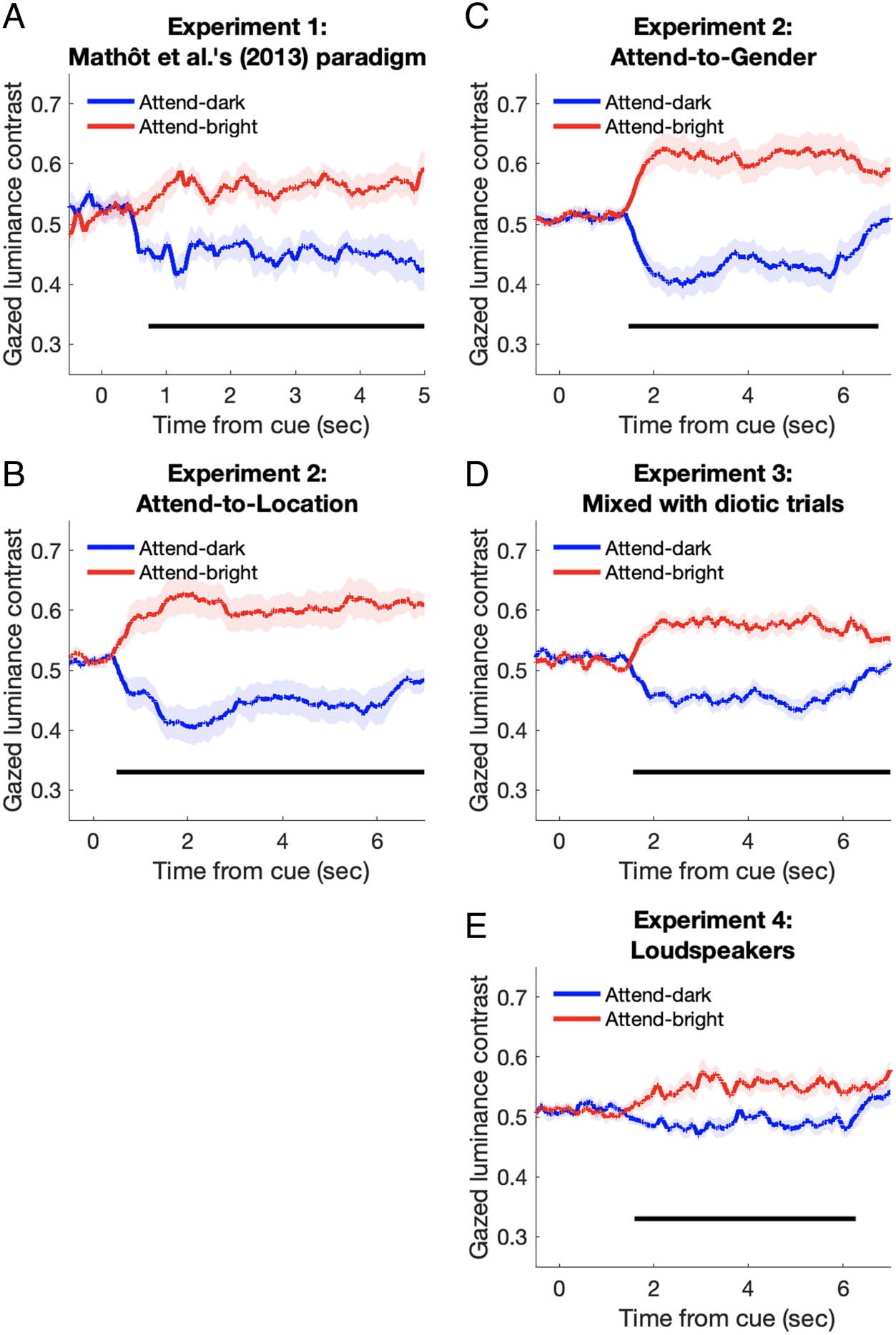
Gaze Bias and PLR
It can be argued that the attention-related PLR is because
of the change in light inputs after uncontrolled eye move-
ments such as ocular drifts. To address this issue, we first
identified the luminance of the local gazed position by
matching the gaze position at each 1-msec sampling
point and the luminance of the viewed display for each
trial. We found that although the global luminance
was controlled to be constant, there were significant
differences in the local luminance of the gazed position
between attending-dark and attending-bright conditions
in all experiments. The difference reached significance
at 726 msec after the cue presentation in Experiment 1;
494 and 1470 msec in the attend-to-location and attend-
to-gender conditions, respectively, in Experiment 2;
1564 msec in Experiment 3; and 1595 msec in Experiment
4 (see Figure 4). One unexpected finding is that the gaze
was focused more when a luminance boundary was
presented near the fixation as in Experiment 1 (93% of
the data points within 2°) than it was in Experiments 2–4
(percentage of data points within the vertical gray area:
67% for the attend-to-location condition and 70% for the
attend-to-gender condition in Experiment 2, 66% in
Experiment 3, and 74% in Experiment 4). The difference
could be also because of the nature of the task. In
any case, although the initial motivation for inserting
the gray vertical area (2.3 visual degrees in width) in
Experiments 2–4 was to make a “buffer zone” between
the hemifields to reduce drastic luminance variations in
foveal inputs accompanied by small gaze drifts, the gaze
analysis suggests that gaze drift was beyond the gray
buffer zone and thus still accompanied local luminance
variation.
284
Journal of Cognitive Neuroscience
Volume 35, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Gazed luminance
contrast as a function of time
from the voice cue onset,
parameterized with the
attended luminance condition
(dark or bright) in all experiments.
The horizontal black lines
indicate a significant difference
(nonparametric cluster-based
permutation tests, p < .05)
between the two attended
luminance conditions during
the periods of 726–5000 msec
in (A), 494–7000 msec in (B),
1470–6747 msec in (C),
1564–7000 msec in (D), and
1595–6268 msec in (E).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
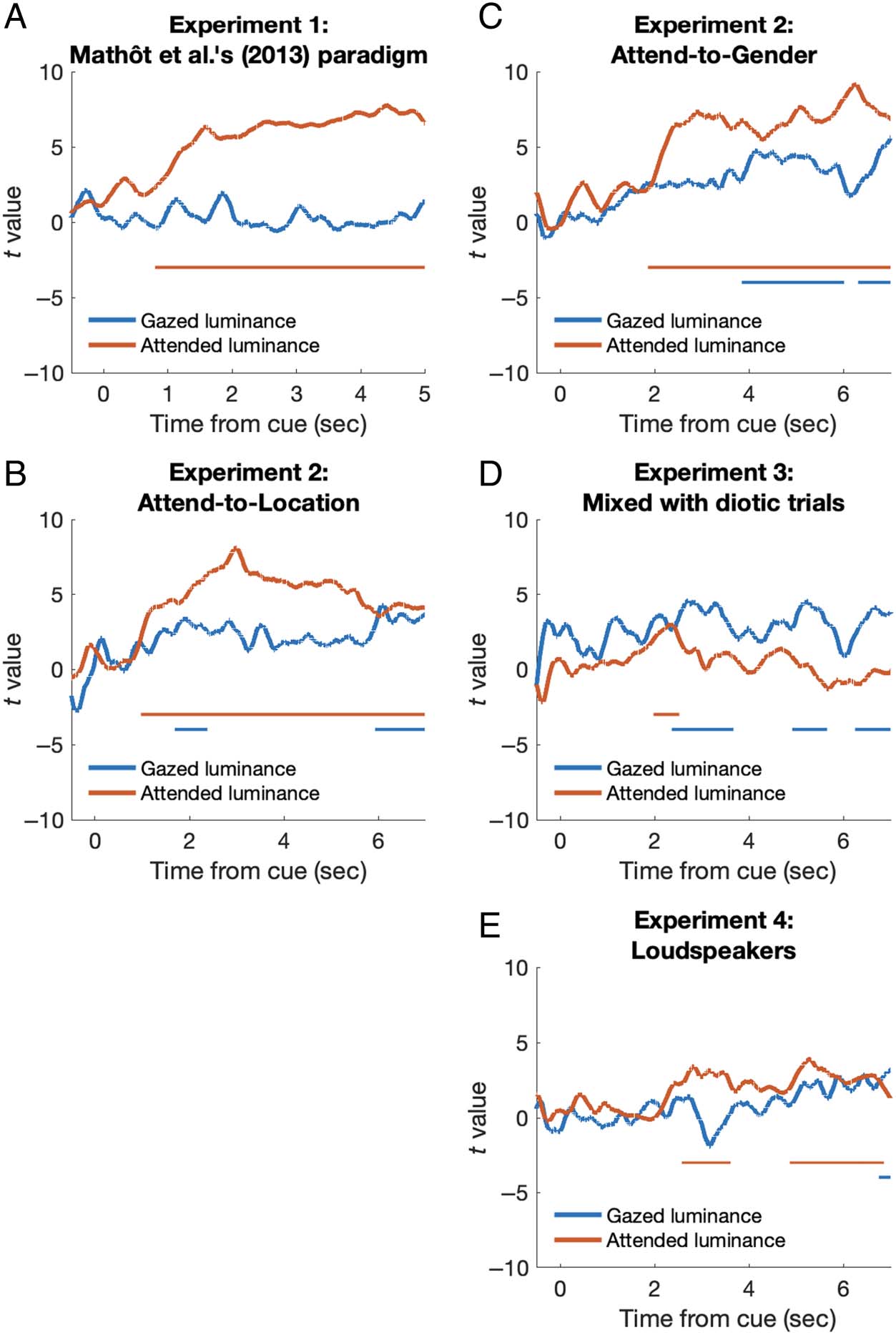
We therefore performed LME analyses to examine
whether gazed local luminance or attended luminance
could better explain the changes in pupil size. LME analy-
ses were conducted at each 1-msec sampling point with
gazed luminance and attended luminance as fixed effects,
participant as the random effect, and pupil size as the
dependent measure. Results showed that, except for
Experiment 3, pupil size was better predicted by attended
luminance than gazed luminance. In Experiment 1, only
attended luminance, not gazed luminance, significantly
predicted pupil size. The t value reached significance
800 msec after the cue onset. In Experiments 2 and 4,
attended luminance, compared with gazed luminance,
predicted pupil size with earlier timings: 971 versus
1690 msec in the attend-to-location condition, 1861 versus
3848 msec in the attend-to-gender condition in Experi-
ment 2, and 2579 versus 6755 msec in Experiment 4. The
significant periods were also longer and more stable for
attended luminance than for gazed luminance (see
Figure 5). By contrast, in Experiment 3, although attended
luminance showed earlier prediction timing than gazed
luminance (1977 vs. 2364 msec), the effect did not last long.
The gazed luminance instead showed more stable and
larger t values than attended luminance. The results suggest
that when the spatial representation of the sound source
was not always clear and lateralized, as we mixed dichotic
trials and diotic trials here, the PLR was better explained
by the gazed local luminance than the attended luminance.
The uncertainty of the spatial property may evoke unex-
pected gaze drift, obscuring the attention-evoked effect.
Liao et al.
285
Figure 5. Result of the LME
analysis in all experiments.
t Value as a function of time
from the voice cue onset,
parameterized with regression
variables of gazed luminance
(blue lines) and attended
luminance (orange lines)
predicting pupil size. The plot
was smoothed by moving
averaging with a 200-msec time
window. The horizontal color
lines indicate the significant
period (t > 2) with at least
500 consecutive samples.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
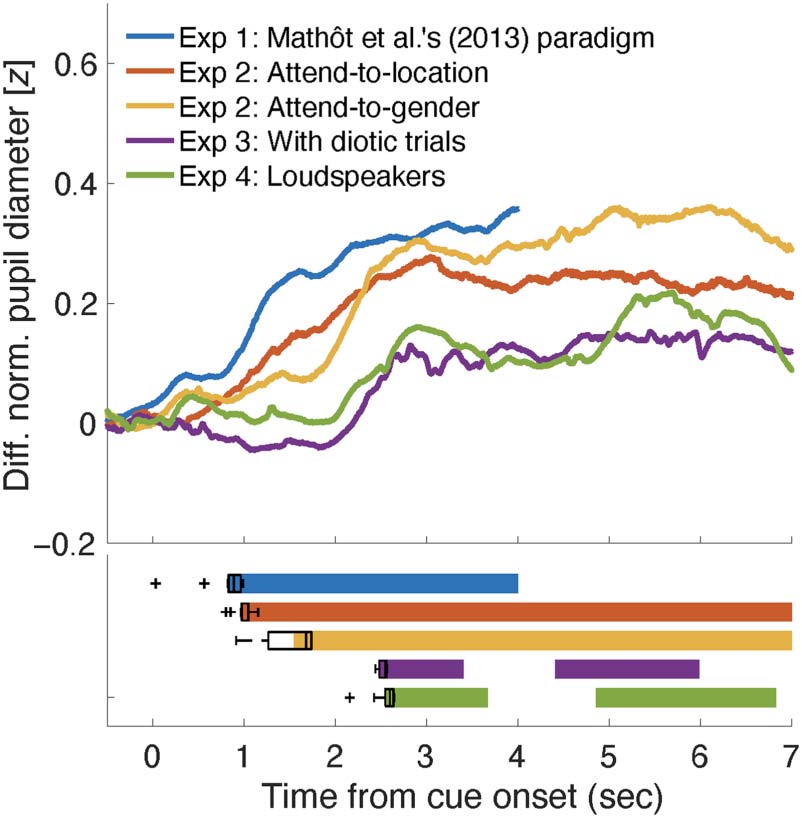
Tracking the Time Course of Attentional Shift by
PLR Divergence
On the assumption that the instantaneous change in pupil
size reflects the state of spatial attention at a time point
(with some delay), we may be able to infer the partici-
pants’ internal process for directing spatial attention. That
is, the divergence of the PLR between the attend-to-dark
and attend-to-bright conditions can be regarded as reflect-
ing the existence or degree of attention-direction bias.
Figure 6 represents the time course of the difference (in
z score) between the pupil size for the attend-to- (or
target-at-) dark and bright side conditions, summarizing
the results of Experiments 1–4. The horizontal color bars
at the bottom indicate the time ranges in which the
difference (or divergence) was significantly above zero.
We adopted the jackknife resampling technique to esti-
mate the variance of the onset latency for statistical analy-
ses (see Methods for details). Results showed that, in
Experiment 2, the onset latency was shorter in the
attend-to-location condition than in the attend-to-gender
condition, t(14) = 6.38, p < .001, BF10 = 1358.88, median
δ = 1.496, 95% CI [0.724, 2.320]. For the attend-to-
location condition, the onset latency was shorter in Exper-
iment 1 than in Experiment 2, t(28) = 2.74, p = .011,
BF10 = 4.89, median δ = 0.814, 95% CI [0.103, 1.592]. For
the attend-to-gender condition, the onset latency was differ-
ent among the three experiments (i.e., Experiments 2, 3,
and 4; F(2, 55) = 179.00, p < .001, BFM = 2.463e+21).
Post hoc analysis indicated that the latency was shorter
286
Journal of Cognitive Neuroscience
Volume 35, Number 2
Experiment 1 may reflect the complexity or difficulty of
the task, or the onset of a target sentence, which delays
attentional shift. When the cue was nonspatial (i.e., the
attend-to-gender condition in Experiments 2, 3, and 4),
participants needed extra time to consciously or uncon-
sciously identify the location of the target talker before
shifting their spatial attention to the auditory object
accordingly. Within a comparable attend-to-gender condi-
tion, the latency was longer when the sounds were pre-
sented via loudspeakers in space (Experiment 4) than
when presented directly to the ears through headphones
(Experiment 2). It should be noted that the size of the PLR
difference was also smaller in Experiment 4 ( loud-
speakers) than in Experiment 2 (headphones). The
slower and smaller divergence in Experiment 4 may be
because of a smaller difference between the two sound
sources in the internal representation. With the loud-
speaker presentation, the azimuthal angles of the sources
were smaller, and the sound from a loudspeaker reached
both ears.
The observed PLR effect in Experiment 3 (dichotic and
diotic sound presentations mixed in an experimental
block in random order) was generally weaker (compared
with Experiment 2) and better explained by the local gazed
luminance caused by unstable ocular drifting than by
attended luminance (Figure 5). This could be because of
the complexity of the stimulus presentation procedure, in
which the auditory objects were sometimes presented in
overlapping space. Participants may have, in general,
decreased their incentives for using space representation
of the auditory object. Alternatively, they may have changed
their strategy trial-by-trial, depending on the availability
of a clear space presentation of the auditory object (i.e.,
left or right vs. center/unknown). Such a trial-by-trial
switching of strategy is implied by the behavioral perfor-
mance in that the accuracy was higher when the sounds
were presented in separate spaces (i.e., dichotic trials)
than mixed into one signal (i.e., diotic trials). In any case,
the Experiment 3 results suggest that, despite the possibil-
ity of a coactivation of a spatial-attention-related mecha-
nism by object-based attentional orientation, the spatial
attention may more strongly affect fixational eye move-
ments than the PLR per se. These movements can be
regarded as ocular drifts or microtremors rather than sac-
cades, because most of the samples of the gaze position
fell within 1 visual degree around the fixation point, albeit
it is difficult to classify the detailed characteristics with the
video-based eye-tracking system used in the current study
(Ko, Snodderly, & Poletti, 2016). As a result of the fixa-
tional eye movements, together with the nature of the
visual display we used here, the local luminance is input
to the eyes differently and thus affects pupil sizes. Future
study should investigate to use of a wider gray area around
the fixation to avoid drastic luminance input differences
because of gaze position drifting or to better control the
participant’s fixation within a designed area by online
contingent-gaze position monitoring.
Liao et al.
287
Figure 6. Differences in pupillary responses between the attend-to-
(or target-at-) dark and attend-to- (or target-at-) bright conditions, as
a function of the time from the cue onset. The horizontal color bars
indicate the significance (nonparametric cluster-based permutation
tests, p < .05) as the pupillary response divergence deviated from zero.
The boxplots show the median values, the interquartile range (IQR),
IQR × 1.5, and outliers of the divergence latency estimated by the
jackknife resampling technique. Exp = Experiment.
in Experiment 2 than in Experiment 3 ( pTukey < .001,
BF10 = 3.600e+15) or in Experiment 4 ( pTukey < .001,
BF10 = 3.981e+9).
DISCUSSION
PLR Reflects Spatial Attentional Shift to
Auditory Object
In four experiments, we demonstrated that the PLR
reflects the focus of covert auditory attention not only
when attention is directed to a particular space by an
endogenous spatial cue (Experiments 1 and 2) but also
when it is allocated to a particular auditory object via non-
spatial characteristics (e.g., gender; Experiments 2, 3, and
4). The finding was replicated regardless of whether the
sounds were presented via headphones (Experiments 1,
2, and 3) or loudspeakers (Experiment 4).
The time course of the PLR divergence (i.e., the differ-
ence between attend-to-dark and attend-to-bright condi-
tions; Figure 6) can be assumed to reflect the timing
of the auditory spatial attention shift in participant’s
planning process of goal-directed action. Given this
assumption, we can explain the observed differences in
PLR-divergence latencies among the conditions by the fol-
lowing scenarios. When the spatial cue was provided
explicitly (i.e., Experiment 1 and the attend-to-location
condition in Experiment 2), participants started shifting
their attention immediately after the cue presentation.
The somewhat longer latency in Experiment 2 than in
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Possible Neural Mechanism of PLR Reflecting
Auditory Spatial Attention and Its Implications
The PLR has been considered primarily as a reflex, but it is
not entirely accounted for by physical inputs. The neural
pathway from the optic nerves to the oculomotor nerves
controls the “reflex” response. When the photosensitive
ganglion cells in the retina are activated by light, the infor-
mation is transmitted via the optic nerves to the olivary
pretectal nucleus (OPN) and projected to the Edinger–
Westphal nucleus (EWN) in the midbrain. The EWN sup-
plies preganglionic parasympathetic fibers to the eye,
which exit with the oculomotor nerves and form synapses
with neurons in the ciliary ganglion. Postsynaptic fibers of
the parasympathetic root leave the ciliary ganglion in short
ciliary nerves, which innervate the pupillary sphincter
muscle to induce pupil constriction.
In addition to this direct “reflex pathway,” several other
modulatory inputs exist. Developmentally, the optic
nerves are derived from the diencephalon, a division of
the forebrain, although they are categorized as cranial
nerves involved in sensory and somatic motor functions.
Anatomically, the OPN receives inputs from the FEF in
pFC (McDougal & Gamlin, 2015; Leichnetz, 1990; Huerta,
Krubitzer, & Jon, 1986; Künzle & Akert, 1977). Recent
evidence has further demonstrated that subthreshold
electrical microstimulation of the FEF modulates the PLR
(Ebitz & Moore, 2017). It has been proposed that the
PLR is modulated by multiple cortical and subcortical pro-
jections, including a direct projection from the FEF to
OPN, or indirect projections involving the relayed areas
such as occipital visual cortical areas or superior colliculus
(SC) to the OPN or EWN ( Joshi & Gold, 2020; Binda &
Gamlin, 2017).
Recent evidence supports the account that emphasizes
the contribution of the SC, especially in relation to orient-
ing responses (Strauch, Wang, et al., 2022; Wang & Munoz,
2018). This SC-centered circuit receives top–down modu-
lations from the FEF, ACC, and lateral intraparietal cortex
and projects neural activities to the OPN and EWN, as men-
tioned above. Microstimulation of the intermediate layers
of the SC evokes not only pupil dilation orienting
responses ( Wang, Boehnke, White, & Munoz, 2012) but
also location-specific pupil luminance modulation ( Wang
& Munoz, 2018). Particularly, Wang and Munoz (2018)
used the visual display with constant global luminance,
similar to the current study, and demonstrated that
through altering activity of the intermediate layers of the
SC via electrical stimulation (facilitation) and lidocaine
injection (inhibition), pupil size was modulated by local
luminance at the next fixated location (i.e., attended loca-
tion). This serves as the neural basis of top–down modu-
lated pupillary responses to the attended or expected
luminance condition. The SC, together with the FEF and
lateral intraparietal cortex, constitutes the neural network
that takes charge of the control of visual selective attention
and eye movements (Maunsell, 2015; Knudsen, 2011). In
addition to visual attention, the SC is known to integrate
multisensory information and have a cross-modal space
map. The inferior colliculus, a major brainstem nucleus
in the auditory pathway, transmits information that is
essential for forming the space map in the SC (Cohen &
Knudsen, 1999). The inferior colliculus also receives
descending projections from the auditory cortex and thus
could be under the influence of cognitive states, including
attention (Huffman & Henson, 1990). Our finding of the
PLR’s reflecting auditory spatial attention ties closely with
the SC-centered account. Assuming that attention-
modulated PLR serves for the preparation of the upcoming
perception, in particular in the visual domain (Mathôt,
2018; Mathôt & Van der Stigchel, 2015), our finding
implies that the spatial representation in auditory space
is merged with the pupil-related spatial map in the visual
domain.
Conclusions
The current study provides a method to infer auditory spa-
tial attention by examining the luminance condition of the
environment and pupillary response. The attention-
modulated PLR reflects the dynamics of attentional shift
to auditory objects. The overall results imply a unified
audiovisual representation of spatial attention. Auditory
object-based attention contains a space representation
of the attended auditory object, even when the object is
oriented without explicit spatial guidance.
Reprint requests should be sent to Hsin-I Liao, NTT Communi-
cation Science Laboratories, NTT Corporation, 3-1, Morinosato
Wakamiya, Atsugi, Kanagawa 243-0198, Japan, or via e-mail:
hsini.liao.pb@hco.ntt.co.jp.
Data Availability Statement
Data and analysis code are available at github.com/hsiniliao
/PLR_AuditoryObject.
Author Contributions
H.-I. L. and S. F. developed the study concept, contributed
to the study design, and wrote the manuscript. H.-I. L., H. F.,
and Y.-H. Y. conducted experiments and collected the
data. H.-I. L. performed the data analysis. H. F., S. Y. and
Y.-H. Y. provided critical comments. All authors approved
the final version of the manuscript for submission.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions
of authorship teams (categorized by estimated gender
288
Journal of Cognitive Neuroscience
Volume 35, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
identification of first author/last author) publishing in the
Journal of Cognitive Neuroscience ( JoCN ) during this
period were M(an)/M = .407, W(oman)/M = .32, M/ W =
.115, and W/ W = .159, the comparable proportions
for the articles that these authorship teams cited were
M/M = .549, W/M = .257, M/ W = .109, and W/ W = .085
(Postle and Fulvio, JoCN, 34:1, pp. 1–3). Consequently,
JoCN encourages all authors to consider gender balance
explicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender
citation balance. The authors of this article report its pro-
portions of citations by gender category to be as follows:
M/M = .707; W/M = .122; M/W = .049; W/W = .122.
REFERENCES
Bates, D., Mächler, M., Bolker, B., & Walker, S. (2015). Fitting
linear mixed-effects models using lme4. Journal of Statistical
Software, 67, 1–48. https://doi.org/10.18637/jss.v067.i01
Best, V., Ozmeral, E. J., Kopčo, N., & Shinn-Cunningham, B. G.
(2008). Object continuity enhances selective auditory
attention. Proceedings of the National Academy of Sciences,
U.S.A., 105, 13174–13178. https://doi.org/10.1073/pnas
.0803718105, PubMed: 18719099
Best, V., Shinn-Cunningham, B., Ozmeral, E., & Kopco, N. (2010).
Exploring the benefit of auditory spatial continuity. Journal
of the Acoustical Society of America, 127, EL258–EL264.
https://doi.org/10.1121/1.3431093, PubMed: 20550229
Binda, P., & Gamlin, P. D. (2017). Renewed attention on the
pupil light reflex. Trends in Neurosciences, 40, 455–457.
https://doi.org/10.1016/j.tins.2017.06.007, PubMed: 28693846
Binda, P., Pereverzeva, M., & Murray, S. O. (2013). Attention to
bright surfaces enhances the pupillary light reflex. Journal of
Neuroscience, 33, 2199–2204. https://doi.org/10.1523
/jneurosci.3440-12.2013, PubMed: 23365255
Binda, P., Pereverzeva, M., & Murray, S. (2014). Pupil size
reflects the focus of feature-based attention. Journal of
Neurophysiology, 112, 3046–3052. https://doi.org/10.1152/jn
.00502.2014, PubMed: 25231615
Bolia, R. S., Nelson, W. T., Ericson, M. A., & Simpson, B. D. (2000).
A speech corpus for multitalker communications research.
Journal of the Acoustical Society of America, 107, 1065–1066.
https://doi.org/10.1121/1.428288, PubMed: 10687719
Braga, R. M., Fu, R. Z., Seemungal, B. M., Wise, R. J. S., & Leech,
R. (2016). Eye movements during auditory attention predict
individual differences in dorsal attention network activity.
Frontiers in Human Neuroscience, 10, 164. https://doi.org
/10.3389/fnhum.2016.00164, PubMed: 27242465
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 433–436. https://doi.org/10.1163
/156856897X00357, PubMed: 9176952
Bushara, K. O., Weeks, R. A., Ishii, K., Catalan, M.-J., Tian, B.,
Rauschecker, J. P., et al. (1999). Modality-specific frontal and
parietal areas for auditory and visual spatial localization in
humans. Nature Neuroscience, 2, 759–766. https://doi.org/10
.1038/11239, PubMed: 10412067
Cave, K. R., & Bichot, N. P. (1999). Visuospatial attention: Beyond
a spotlight model. Psychonomic Bulletin & Review, 6, 204–223.
https://doi.org/10.3758/BF03212327, PubMed: 12199208
Cohen, Y. E., & Knudsen, E. I. (1999). Maps versus clusters:
Different representations of auditory space in the midbrain
and forebrain. Trends in Neurosciences, 22, 128–135. https://
doi.org/10.1016/S0166-2236(98)01295-8, PubMed: 10199638
Corbetta, M. (1998). Frontoparietal cortical networks for
directing attention and the eye to visual locations: Identical,
independent, or overlapping neural systems? Proceedings
of the National Academy of Sciences, U.S.A., 95, 831–838.
https://doi.org/10.1073/pnas.95.3.831, PubMed: 9448248
Downing, C. J. (1988). Expectancy and visual–spatial attention:
Effects on perceptual quality. Journal of Experimental
Psychology: Human Perception and Performance, 14,
188–202. https://doi.org/10.1037/0096-1523.14.2.188,
PubMed: 2967876
Ebitz, R. B., & Moore, T. (2017). Selective modulation of the
pupil light reflex by microstimulation of prefrontal cortex.
Journal of Neuroscience, 37, 5008–5018. https://doi.org/10
.1523/jneurosci.2433-16.2017, PubMed: 28432136
Eriksen, C. W., & St. James, J. D. (1986). Visual attention within
and around the field of focal attention: A zoom lens model.
Perception & Psychophysics, 40, 225–240. https://doi.org/10
.3758/BF03211502, PubMed: 3786090
Fritz, J. B., Elhilali, M., David, S. V., & Shamma, S. A. (2007).
Auditory attention—Focusing the searchlight on sound.
Current Opinion in Neurobiology, 17, 437–455. https://doi
.org/10.1016/j.conb.2007.07.011, PubMed: 17714933
Haab, O. (1886). Vortrag gesellschaft der ärzte in zürich am 21.
November 1885. Korrespondenzblatt für Schweizer Ärzte,
16, 153.
Hill, K. T., & Miller, L. M. (2009). Auditory attentional control
and selection during cocktail party listening. Cerebral Cortex,
20, 583–590. https://doi.org/10.1093/cercor/bhp124, PubMed:
19574393
Huerta, M. F., Krubitzer, L. A., & Jon, H. C. K. (1986). Frontal
Intracortical eye field as defined by microstimulation in
squirrel monkeys, owl monkeys, and macaque monkeys: I.
subcortical connections. Journal of Comparative Neurology,
253, 415–439. https://doi.org/10.1002/cne.902530402,
PubMed: 3793998
Huffman, R. F., & Henson, O. W. (1990). The descending
auditory pathway and acousticomotor systems: Connections
with the inferior colliculus. Brain Research Reviews, 15,
295–323. https://doi.org/10.1016/0165-0173(90)90005-9,
PubMed: 2289088
Joshi, S., & Gold, J. I. (2020). Pupil size as a window on neural
substrates of cognition. Trends in Cognitive Sciences, 24,
466–480. https://doi.org/10.1016/j.tics.2020.03.005, PubMed:
32331857
Keysers, C., Gazzola, V., & Wagenmakers, E.-J. (2020). Using
Bayes factor hypothesis testing in neuroscience to establish
evidence of absence. Nature Neuroscience, 23, 788–799.
https://doi.org/10.1038/s41593-020-0660-4, PubMed: 32601411
Kleiner, M., Brainard, D., & Pelli, D. (2007). What’s new in
Psychtoolbox-3? Perception, 36, 1–16.
Knudsen, E. I. (2011). Control from below: The role of a
midbrain network in spatial attention. European Journal of
Neuroscience, 33, 1961–1972. https://doi.org/10.1111/j.1460
-9568.2011.07696.x, PubMed: 21645092
Ko, H.-K., Snodderly, D. M., & Poletti, M. (2016). Eye
movements between saccades: Measuring ocular drift and
tremor. Vision Research, 122, 93–104. https://doi.org/10.1016
/j.visres.2016.03.006, PubMed: 27068415
Künzle, H., & Akert, K. (1977). Efferent connections of cortical,
area 8 (frontal eye field) in Macaca fascicularis. A
reinvestigation using the autoradiographic technique.
Journal of Comparative Neurology, 173, 147–164. https://doi
.org/10.1002/cne.901730108, PubMed: 403205
Laeng, B., & Endestad, T. (2012). Bright illusions reduce the
eye’s pupil. Proceedings of the National Academy of
Sciences, U.S.A., 109, 2162–2167. https://doi.org/10.1073/pnas
.1118298109, PubMed: 22308422
Laeng, B., & Sulutvedt, U. (2014). The eye pupil adjusts to
imaginary light. Psychological Science, 25, 188–197. https://
doi.org/10.1177/0956797613503556, PubMed: 24285432
Liao et al.
289
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Larson, E., & Lee, A. K. C. (2014). Switching auditory attention
Pelli, D. G. (1997). The VideoToolbox software for visual
using spatial and non-spatial features recruits different
cortical networks. Neuroimage, 84, 681–687. https://doi.org
/10.1016/j.neuroimage.2013.09.061, PubMed: 24096028
Leichnetz, G. R. (1990). Preoccipital cortex receives a
differential input from the frontal eye field and projects to the
pretectal olivary nucleus and other visuomotor-related
structures in the rhesus monkey. Visual Neuroscience, 5,
123–133. https://doi.org/10.1017/S095252380000016X,
PubMed: 2177637
Liao, H.-I., Kashino, M., & Shimojo, S. (2021). Attractiveness in
the eyes: A possibility of positive loop between transient
pupil constriction and facial attraction. Journal of Cognitive
Neuroscience, 33, 315–340. https://doi.org/10.1162/jocn_a
_01649, PubMed: 33166194
Liao, H.-I., Kidani, S., Yoneya, M., Kashino, M., & Furukawa, S.
(2016). Correspondences among pupillary dilation response,
subjective salience of sounds, and loudness. Psychonomic
Bulletin & Review, 23, 412–425. https://doi.org/10.3758
/s13423-015-0898-0, PubMed: 26163191
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190. https://doi.org/10.1016/j.jneumeth
.2007.03.024, PubMed: 17517438
Mathôt, S. (2018). Pupillometry: Psychology, physiology, and
function. Journal of Cognition, 1, 16. https://doi.org/10.5334
/joc.18, PubMed: 31517190
Mathôt, S., Dalmaijer, E., Grainger, J., & Van der Stigchel, S.
(2014). The pupillary light response reflects exogenous
attention and inhibition of return. Journal of Vision, 14, 7.
https://doi.org/10.1167/14.14.7, PubMed: 25761284
Mathôt, S., van der Linden, L., Grainger, J., & Vitu, F. (2013).
The pupillary light response reveals the focus of covert visual
attention. PLoS One, 8, e78168. https://doi.org/10.1371
/journal.pone.0078168, PubMed: 24205144
Mathôt, S., & Van der Stigchel, S. (2015). New light on the
mind’s eye: The pupillary light response as active vision.
Current Directions in Psychological Science, 24, 374–378.
https://doi.org/10.1177/0963721415593725, PubMed: 26494950
Maunsell, J. H. R. (2015). Neuronal mechanisms of visual
attention. Annual Review of Vision Science, 1, 373–391.
https://doi.org/10.1146/annurev-vision-082114-035431,
PubMed: 28532368
McDougal, D. H., & Gamlin, P. D. (2015). Autonomic control of
the eye. Comprehensive Physiology, 5, 439–473. https://doi
.org/10.1002/cphy.c140014, PubMed: 25589275
Mondor, T. A., & Zatorre, R. J. (1995). Shifting and focusing
auditory spatial attention. Journal of Experimental Psychology:
Human Perception and Performance, 21, 387–409. https://doi
.org/10.1037//0096-1523.21.2.387, PubMed: 7714479
Naber, M., Frässle, S., & Einhäuser, W. (2011). Perceptual
rivalry: Reflexes reveal the gradual nature of visual awareness.
PLoS One, 6, e20910. https://doi.org/10.1371/journal.pone
.0020910, PubMed: 21677786
Naber, M., & Nakayama, K. (2013). Pupil responses to high-level
image content. Journal of Vision, 13, 7. https://doi.org/10
.1167/13.6.7, PubMed: 23685390
Noyce, A. L., Kwasa, J. A. C., & Shinn-Cunningham, B. G. (2022).
Defining attention from an auditory perspective. WIREs
Cognitive Science, e1610. https://doi.org/10.1002/wcs.1610,
PubMed: 35642475
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011).
FieldTrip: Open source software for advanced analysis of
MEG, EEG, and invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011,
156869. https://doi.org/10.1155/2011/156869, PubMed:
21253357
psychophysics: Transforming numbers into movies. Spatial
Vision, 10, 437–442. https://doi.org/10.1163/156856897X00366,
PubMed: 9176953
Posner, M., Snyder, C., & Davidson, B. (1980). Attention and the
detection of signals. Journal of Experimental Psychology:
General, 109, 160–174. https://doi.org/10.1037/0096-3445
.109.2.160, PubMed: 7381367
Quinlan, P. T., & Bailey, P. J. (1995). An examination of
attentional control in the auditory modality: Further evidence
for auditory orienting. Perception & Psychophysics, 57, 614–628.
https://doi.org/10.3758/BF03213267, PubMed: 7644322
Rhodes, G. (1987). Auditory attention and the representation of
spatial information. Perception & Psychophysics, 42, 1–14.
https://doi.org/10.3758/BF03211508, PubMed: 3658631
Salvaggio, S., Andres, M., Zénon, A., & Masson, N. (2022). Pupil
size variations reveal covert shifts of attention induced by
numbers. Psychonomic Bulletin & Review, 29, 1844–1853.
https://doi.org/10.3758/s13423-022-02094-0, PubMed:
35384595
Shinn-Cunningham, B. G. (2008). Object-based auditory and
visual attention. Trends in Cognitive Sciences, 12, 182–186.
https://doi.org/10.1016/j.tics.2008.02.003, PubMed: 18396091
Smith, D. V., Davis, B., Niu, K., Healy, E. W., Bonilha, L.,
Fridriksson, J., et al. (2010). Spatial attention evokes similar
activation patterns for visual and auditory stimuli. Journal of
Cognitive Neuroscience, 22, 347–361. https://doi.org/10.1162
/jocn.2009.21241, PubMed: 19400684
Spence, C. J., & Driver, J. (1994). Covert spatial orienting in
audition: Exogenous and endogenous mechanisms. Journal
of Experimental Psychology: Human Perception and
Performance, 20, 555–574. https://doi.org/10.1037/0096-1523
.20.3.555
Spence, C., & Driver, J. (1996). Audiovisual links in endogenous
covert spatial attention. Journal of Experimental Psychology:
Human Perception and Performance, 22, 1005–1030,
https://doi.org/10.1037/0096-1523.22.4.1005, PubMed: 8756965
Strauch, C., Romein, C., Naber, M., Van der Stigchel, S., & Ten
Brink, A. F. (2022). The orienting response drives
pseudoneglect—Evidence from an objective pupillometric
method. Cortex, 151, 259–271. https://doi.org/10.1016/j
.cortex.2022.03.006, PubMed: 35462203
Strauch, C., Wang, C.-A., Einhäuser, W., Van der Stigchel, S., &
Naber, M. (2022). Pupillometry as an integrated readout of
distinct attentional networks. Trends in Neurosciences, 45,
635–647. https://doi.org/10.1016/j.tins.2022.05.003, PubMed:
35662511
Suzuki, Y., Minami, T., Laeng, B., & Nakauchi, S. (2019). Colorful
glares: Effects of colors on brightness illusions measured with
pupillometry. Acta Psychologica, 198, 102882. https://doi.org
/10.1016/j.actpsy.2019.102882, PubMed: 31288107
Treisman, A. M., & Gelade, G. (1980). A feature-integration
theory of attention. Cognitive Psychology, 12, 97–136. https://
doi.org/10.1016/0010-0285(80)90005-5, PubMed: 7351125
Wang, C. A., Boehnke, S. E., White, B. J., & Munoz, D. P. (2012).
Microstimulation of the monkey superior colliculus induces
pupil dilation without evoking saccades. Journal of
Neuroscience, 32, 3629–3636. https://doi.org/10.1523
/jneurosci.5512-11.2012, PubMed: 22423086
Wang, C.-A., & Munoz, D. P. (2018). Neural basis of
location-specific pupil luminance modulation. Proceedings of
the National Academy of Sciences, U.S.A., 115, 10446–10451.
https://doi.org/10.1073/pnas.1809668115, PubMed: 30249636
Zatorre, R. J., Mondor, T. A., & Evans, A. C. (1999). Auditory
attention to space and frequency activates similar cerebral
systems. Neuroimage, 10, 544–554. https://doi.org/10.1006
/nimg.1999.0491, PubMed: 10547331
290
Journal of Cognitive Neuroscience
Volume 35, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
2
2
7
6
2
0
6
7
0
1
8
/
j
o
c
n
_
a
_
0
1
9
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3