Rhythm Complexity Modulates Behavioral and Neural
Dynamics During Auditory–Motor Synchronization
Brian Mathias1,2*, Anna Zamm1,3*, Pierre G. Gianferrara1,4
Bernhard Ross5, and Caroline Palmer1
Abstract
■ We addressed how rhythm complexity influences auditory–
motor synchronization in musically trained individuals who per-
ceived and produced complex rhythms while EEG was recorded.
Participants first listened to two-part auditory sequences (Listen
condition). Each part featured a single pitch presented at a fixed
rate; the integer ratio formed between the two rates varied in rhyth-
mic complexity from low (1:1) to moderate (1:2) to high (3:2). One
of the two parts occurred at a constant rate across conditions.
Then, participants heard the same rhythms as they synchronized
their tapping at a fixed rate (Synchronize condition). Finally, they
tapped at the same fixed rate (Motor condition). Auditory feedback
from their taps was present in all conditions. Behavioral effects of
rhythmic complexity were evidenced in all tasks; detection of
missing beats (Listen) worsened in the most complex (3:2) rhythm
condition, and tap durations (Synchronize) were most variable
and least synchronous with stimulus onsets in the 3:2 condition.
EEG power spectral density was lowest at the fixed rate during
the 3:2 rhythm and greatest during the 1:1 rhythm (Listen and
Synchronize). ERP amplitudes corresponding to an N1 time win-
dow were smallest for the 3:2 rhythm and greatest for the 1:1
rhythm (Listen). Finally, synchronization accuracy (Synchronize)
decreased as amplitudes in the N1 time window became more pos-
itive during the high rhythmic complexity condition (3:2). Thus,
measures of neural entrainment corresponded to synchronization
accuracy, and rhythmic complexity modulated the behavioral and
neural measures similarly. ■
INTRODUCTION
Many behaviors require the temporal coordination of one’s
actions with perceived auditory information, from dance
(Brown & Parsons, 2008), to athletics (Bood, Nijssen, Van
Der Kamp, & Roerdink, 2013), to music-making ( Wing,
Endo, Bradbury, & Vorberg, 2014). When dancing to music,
for example, one must coordinate the timing of body
movements with a perceived auditory rhythm, a temporally
regular acoustic pattern (Miura, Kudo, & Nakazawa, 2013;
Thaut, 2013). The temporal alignment of one’s movement
with the frequency and phase of auditory rhythms can be
described as “auditory–motor synchronization.” Musical
auditory–motor synchronization requires the simultaneous
perception of auditory rhythms and efficient coordination
of movement with those rhythms (for a review, see Repp
& Su, 2013). A major question in cognitive neuroscience
is how interactions between auditory and motor processes
give rise to accurate auditory–motor synchronization.
Emerging evidence suggests that perception of auditory
rhythms is accompanied by neural forms of entrainment,
defined here as the process by which oscillations couple
with, or alter their period in response to, another (intrinsic)
1McGill University, 2Max Planck Institute for Human Cognitive
and Brain Science, 3Central European University, Budapest,
Hungary, 4Carnegie Mellon University, 5Rotman Research
Institute, Toronto, ON, Canada
*Joint first authors.
© 2020 Massachusetts Institute of Technology
oscillation or a stimulus rhythm (Haken, Kelso, & Bunz,
1985). Neural entrainment can arise when rhythmic fluctu-
ations of electrical brain activity or neural oscillations arise
from synchronous excitability in networks of functionally
connected neurons (Buzsáki & Draguhn, 2004), which
become coupled with (adapt their period in response to)
acoustic signals. Neural entrainment with frequency com-
ponents of auditory rhythms is evidenced in EEG measures
of neural responses that occur at positions of occasional
omitted tones in rhythmic auditory sequences (Snyder &
Large, 2005) and enhanced amplitudes of neural oscilla-
tions at the periodicity of a perceived beat in auditory tone
sequences that did not contain energy at that periodicity
(Fujioka, Ross, & Trainor, 2015; Nozaradan, Peretz, &
Mouraux, 2012; Nozaradan, Peretz, Missal, & Mouraux, 2011).
The coupling of neural oscillations with acoustic rhythms
can enhance perceptual processing of stimulus events
(Large & Palmer, 2002; Engel, Fries, & Singer, 2001; Large
& Jones, 1999). EEG studies have shown that mid-latency
ERPs are enhanced in response to acoustic events that
receive enhanced perceptual processing, such as those to
which individuals voluntarily attend (Sowman, Kuusik, &
Johnson, 2012; Hillyard, Hink, Schwent, & Picton, 1973).
These effects have been observed in the N1 component, a
negative-going ERP component that peaks around 100 msec
after tone onsets (Nobre & van Ede, 2018; Lange, Rösler,
& Röder, 2003; Näätänen & Winkler, 1999). N1 amplitudes
are typically measured as the mean amplitude across a
Journal of Cognitive Neuroscience 32:10, pp. 1864–1880
https://doi.org/10.1162/jocn_a_01601
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
–
a
p
r
d
t
i
3
c
2
l
1
e
0
–
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
/
t
o
n
0
5
M
a
y
2
0
2
1
post-stimulus time window or as the negative peak within a
post-stimulus time window. Some alternatives to these
window-based measures involve defining N1 amplitude as
the voltage difference between the most prominent peak
in the N1 time window and the peak of a neighboring ERP
such as the P1 (i.e., peak-to-peak measurement). Although
these alternative peak-to-peak metrics may in some cases
be able to disambiguate multiple overlapping ERPs, the
window-based approach is less susceptible to artifactual
peaks in the data—which can arise from filtering artifacts
and other noise sources (Woodman, 2010)—and is more
widely used in the literature on auditory rhythm perception.
Moreover, P1, N1, and P2 components are often analyzed sep-
arately because they are thought to reflect different functions.
More negative amplitudes within post-stimulus time
windows associated with the N1 have been observed in
response to tones aligned with metrically strong beats
compared to metrically weak beats during the perception
of short melodies (Fitzroy & Sanders, 2015) and to sounds
that are accented by increased intensity in short rhythmic
sequences (Schaefer, Vlek, & Desain, 2011). If individuals
allocate attentional resources to tones that occur at rhyth-
mically salient frequencies in auditory sequences, then
amplitudes within an N1-related time window elicited by
these attended tones should be larger than for tones
which occur at less-attended frequencies. A final effect
on amplitudes within the timeframe of the N1 component
is repetition: Repeated sounds tend to elicit a smaller N1
response than novel, non-repeated sounds. This phenom-
enon may arise from refraction in auditory circuits (Budd,
Barry, Gordon, Rennie, & Michie, 1998), which is the tem-
poral interval across which a given neural system returns
to baseline excitability, or from sensory memory updating
(for a review, see Näätänen & Picton, 1987).
N1 amplitudes are reduced (more positive) in response
to self-generated relative to externally generated sounds,
possibly because of motor-induced suppression of audi-
tory processing (Horváth, Maess, Baess, & Tóth, 2012).
Cortical motor regions may generate templates of sounds
that we intend to produce (Bays & Wolpert, 2007); these
templates are accessible to sensory memory and are
subtracted from actual sensory input during production, re-
flected in suppressed amplitudes within a time window sur-
rounding the N1 (SanMiguel, Widmann, Bendixen, Trujillo-
Barreto, & Schröger, 2013). Therefore, N1 responses may
distinguish between sounds that one produces and sounds
that one perceives during auditory–motor synchronization.
We test here whether larger amplitudes within the typical
timeframe of the N1 are elicited in response to the frequen-
cies that participants synchronize with (hear) but do not
produce, than to the frequencies that participants produce.
Recent research suggests that production modulates
not only auditory–motor ERPs but also oscillatory brain
responses. Larger neural oscillatory responses have been
observed when individuals tap along with a rhythm compared
to only listening to the rhythm (Nozaradan, Schönwiesner,
Caron-Desrochers, & Lehmann, 2016). Moving one’s body
at a specific frequency to a rhythmic auditory sequence
can enhance amplitudes of neural oscillations at that fre-
quency while subsequently listening to the same auditory
sequence (Chemin, Mouraux, & Nozaradan, 2014). Neural
oscillations during rhythm perception may also predict syn-
chronization accuracy during rhythmic production: Stronger
oscillations at the frequency of a perceived rhythm are asso-
ciated with greater temporal prediction during sensorimotor
synchronization (Nozaradan, Peretz, & Keller, 2016).
An open question is how neural oscillations moderate
perceptual processing of complex ecological auditory
rhythms such as those occurring in multipart music. One
approach comes from the dynamical systems framework
of rhythmic entrainment (Large, Herrera, & Velasco, 2015;
Strogatz, 2001), which describes mathematically how cou-
pling arises between oscillators with different frequencies.
The stability of two rhythms (or periodic oscillations) is a
function of the ratio between their frequencies; rhythms
that form a simple integer ratio relationship (such as 1:1),
referred to here as “simple rhythms,” achieve more stability
than rhythms that form a complex integer ratio (such as
3:2), referred to here as “complex rhythms” (Glass &
Mackey, 1988). The relationships between the rhythm ra-
tios can be described by a Farey tree, which defines the re-
gions of stability for two rhythms based on their frequency
ratio (Kelso, 1991; Schroeder, 1991). Dynamical models
offer predictions for how neural oscillations respond to
simple versus complex auditory rhythms, namely, the sta-
bility of neural oscillations responding to auditory rhythms
based on their rhythmic ratios. In turn, more stable neural
oscillations may enhance perceptual processing of acoustic
rhythms. Thus, we predict that motor synchronization with
auditory rhythms containing simple or complex frequency
ratios should yield greater power of neural oscillations and
more negative amplitudes within a typical N1 time window
at the frequencies with a simple ratio than those with the
complex ratio.
We investigated the neural correlates of auditory–motor
rhythm processing during perception, production, and syn-
chronization of auditory rhythms containing simple and
complex frequency ratios. Two primary questions were
addressed: The first question was whether neural entrain-
ment with a simple-ratio auditory rhythm is enhanced com-
pared to complex-ratio rhythms, in perception, production,
and synchronization tasks. Neural entrainment was mea-
sured by assessing neural oscillations in the frequency
domain, and ERPs time-locked to auditory stimulus onsets
and to motor responses in the time domain. The second
question was whether behavioral synchronization is en-
hanced in response to simple rhythms compared to complex
rhythms and whether behavioral synchronization is asso-
ciated with neural entrainment measures. We examined
accuracy and stability of synchronization in a coordina-
tion task in which individuals tapped along with auditory
rhythms that formed simple and complex ratios.
The current study tested perception and production of
auditory rhythms by skilled musicians, who are experienced
Mathias et al.
1865
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
–
a
p
r
d
t
i
3
c
2
l
1
e
0
–
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
t
/
f
o
n
0
5
M
a
y
2
0
2
1
formal instruction on a musical instrument (range: 6–17 years;
M = 9.2 years, SD = 2.9 years) participated. Participants
currently practiced their instrument an average of 5.8 hr
a week (SD = 8.6) and averaged 12.2 years of experience
playing their instrument (SD = 4.4). All participants were
right-handed and did not possess any neurological disor-
ders. Participants passed an audiometric screening test, in
which they demonstrated hearing thresholds ≤ 20 dB for
1000-, 750-, 500-, and 250-Hz tones (representing the
range of pitches used in the experiment). Three additional
participants were recruited. Two of these participants’
data were excluded because of poor EEG signal quality,
and a third participant’s data were excluded because of
low task performance (hit rates that exceeded 3 SDs from
the group mean in the Listen task). The study was re-
viewed by the McGill University research ethics board.
Participants provided a written consent to participate after
they were fully informed about the study.
Stimuli
The auditory stimuli were sequences of repeating rhythmic
patterns (see Figure 1). The sequences consisted of low-
pitched (392-Hz) sine tones and high-pitched (660-Hz)
woodblock sounds, which were perceptually distinct in
pitch, timbre, and presentation rate. Three different Rhythm
complexity conditions were created from the high- and low-
pitched tone sequences: a 1:1 ratio, a 1:2 ratio, and a 3:2 ratio
(the first number indicates the rate of the high pitch, and
the second number indicates the rate of the low pitch).
The low-pitched tones were presented at a constant rate
with an interonset interval (IOI) of 528 msec, whereas
the IOI for the high-pitched tones differed across the rate
ratios with 528 msec for the 1:1, 1056 msec for the 1:2,
and 352 msec for the 3:2 condition. Thus, the stimulus
(“high-pitched part”) frequency was defined as 1.89 Hz in
the 1:1 condition, 0.94 Hz in the 1:2 condition, and 2.84 Hz
in the 3:2 condition. The prescribed tap (“low-pitched
part”) frequency was 1.89 Hz across all Rhythm conditions
and tasks. The shared frequency (which corresponded to
with the perception and production of both simple- and
complex-ratio rhythms and therefore offer an ideal popula-
tion for studying rhythmic behavior (Collier & Wright,
1995). Participants first listened to auditory sequences com-
prising two-part rhythms (Listen task), each part presented
with a different constant pitch. One part occurred at a con-
stant (fixed) frequency across Rhythm conditions, whereas
the other part varied in frequency across conditions relative
to the fixed frequency, to form a 1:1 integer ratio (1:1 con-
dition), a 1:2 integer ratio (1:2 condition), or a 3:2 integer
ratio (3:2 condition). Rhythmic complexity therefore ranged
from low (1:1) to moderate (1:2) to high (3:2), consistent
with predictions of Farey tree hierarchies (Glass & Mackey,
1988). Participants then performed a Synchronize task in
which they tapped at the fixed frequency while aiming to syn-
chronize their movements with the other part. Stimulus-to-
tap ratios varied across three Rhythm conditions with ratios
of 1:1, 1:2, and 3:2, consistent with the rhythmic ratios in the
Listen task. To achieve a baseline measure of cortical re-
sponses during rhythmic movement in the absence of syn-
chronization, participants completed a control Motor task,
in which they tapped at the same fixed frequency as during
the Synchronization task. Neural activity was recorded at the
scalp using EEG during the Listen, Synchronize, and Motor
tasks, and sound corresponding to the stimulus and/or taps
was present in all conditions.
Following predictions from nonlinear dynamical systems,
neural oscillations should exhibit most power in the simple-
ratio (1:1) Rhythm condition and least power in the most
complex (3:2) Rhythm condition for both Listen and
Synchronize conditions. Furthermore, the amplitude of neu-
ral oscillations at the constant frequency, as well as the am-
plitude of ERP waveforms within an N1-related time window,
should increase during the Synchronization task compared
to the Listen task. Participants in the Synchronize task
should exhibit greatest synchrony of tapping with the audi-
tory stimulus in the simple-ratio (1:1) Rhythm condition and
least synchrony in the complex-ratio (3:2) Rhythm condi-
tion. Behavioral synchrony measures and the amplitude of
ERP waveforms within the N1 time window are expected to
decrease together as rhythmic complexity increases (from
1:1 to 1:2 to 3:2). Comparison of N1 amplitudes across tasks
(Listen/Synchronize/Motor) and Rhythm conditions (1:1,
1:2, 3:2) should reveal how interactions between auditory
perceptual and motor processes give rise to accurate
auditory–motor synchronization. We examine these interac-
tions under naturalistic stimulus conditions such as those
that occur during music and speech production, in which
auditory feedback from both stimuli and responses are
present.
METHODS
Participants
Twenty-nine adults (21 women, eight men; aged 18–30 years;
M = 22.6 years, SD = 3.1 years) with at least 6 years of
Figure 1. Schematic showing the alignment of participant tap onsets
(indicated by non-bold Xs) and stimulus onsets (indicated by bold Xs)
across a cycle of tap onsets for each of three Rhythm conditions. Circles
indicate the shared stimulus frequency across Rhythm conditions.
1866
Journal of Cognitive Neuroscience
Volume 32, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
–
a
p
r
d
t
i
3
c
2
l
1
e
0
–
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
f
.
t
o
n
0
5
M
a
y
2
0
2
1
points of simultaneity between the two frequencies across
all Rhythm conditions, shown by circles in Figure 1) was de-
fined as 0.94 Hz. The timbres were generated on a sound
module SD-50 SoundCanvas (Roland Inc.) using “Pre_333
Simple sine” (Synth Lead category) for the low-pitched tone
and “Rhy 001” (Drums category) for the high-pitched tone.
The attack time of the sine tone and woodblock sounds was
5 msec, and the decay time for both sounds was 36 msec.
The same percussion sound was used for a metronome cue
that signaled the tapping tempo at the start of each trial. The
sound pressure level of high- and low-pitched tones and
metronome tones was set to 75 dB SPL, confirmed with a
sound level meter.
Each trial contained four metronome clicks with IOIs of
528 msec, followed for 30 sec by one of the three Rhythms
(1:1, 1:2, and 3:2). Twelve trials were created for each of the
three Rhythm conditions in the Listen task. Two of the 12
trials contained one missing beat in either the low-pitched
part or the high-pitched part. Twelve trials were created for
each of the three Rhythm conditions in the Synchronize task;
each trial contained the four-beat metronome cue followed
by the high-pitched part (without missing beats), during
which participants heard the low-pitched part when they
tapped. Twelve trials were included in the Motor task; each
trial presented the four-beat metronome cue, after which
participants heard the low-pitched part when they tapped.
Equipment
The experiment took place in a sound-attenuated and elec-
trically shielded testing room. The audiometric screening
was administered with a diagnostic audiometer using over-
ear headphones provided by Maico (MA-40, Maico GmbH).
Auditory stimuli were presented over EEG-compatible insert
earphones (ER1-14B, Etymotic Research). Participants
tapped to the low-pitched sequence by pressing a key (note
name C3) on an electronic keyboard (PSR-500M, Yamaha
Inc.) that transmitted timing information with 1-msec reso-
lution via a musical instrument digital interface (Yamaha
Inc.). Information about the tap timing was recorded using
FTAP software (Finney, 2001) modified to transmit event
triggers (Mathias, Gehring, & Palmer, 2017).
Participants wore an EEG cap with 64 Ag/AgCl electrodes
configured according to an extension of the International
10–20 system. EEG signals were recorded by a BioSemi
ActiveTwo system at a resolution of 24 bits and a sampling
rate of 1024 Hz (BioSemi, Inc.). The EEG was grounded
using BioSemi’s combination of common mode sense
and drive right leg electrodes. Electrodes below and above
the right eye monitored vertical eye movements, and two
electrodes placed adjacent to the outer canthi of the eyes
monitored horizontal eye movements.
Design
The within-participant 2 × 3 repeated-measures design
included two factors: Task (Listen, Synchronize) and
Rhythm condition (1:1, 1:2, 3:2). The Motor condition
served as a separate control to allow identification of a
motor ROI for analysis of power spectral density (PSD).
Each participant completed the tasks in this order—
Listen, Synchronize, and Motor condition—to ensure
that tapping rates did not influence the perceptual neural
responses in the Listen task. The order of tasks was fixed
to ensure that Listen blocks were not influenced by prior
experience with producing the stimulus rhythms (via
auditory or motor imagery; cf. Brown & Palmer, 2013).
Within Listen and Synchronize tasks, the blocks of
Rhythm condition trials were presented in a fixed order
of 1:1, 1:2, and 3:2, ranging from the easiest to the most
difficult. As practice effects should favor the final (most
complex rhythm) condition within each block (Tajima
& Chosi, 2000), the order of Rhythm conditions was fixed
to bias away from the hypothesis that synchronization
performance should be best for simple relative to com-
plex rhythmic ratios.
There were two practice trials and 12 experimental tri-
als for each Rhythm condition (1:1, 1:2, 3:2) within the
Listen and Synchronize tasks and two practice trials and
12 experimental trials for the Motor task, yielding 84 ex-
perimental trials (12 × 3 × 2 + 12 = 84).
Procedure
Participants first provided written consent, completed a
questionnaire about their musical training background,
and completed an audiometry screening. Participants
who were not able to detect any tones presented at or
below 20 dB were excluded from the experiment.
Participants were then outfitted with an EEG cap and
electrodes and completed the experimental tasks.
Listen Task
Participants first listened separately to the sine tone and the
woodblock sequences to become familiarized with the audi-
tory stimuli. The woodblock sequence was referred to by the
experimenter as the “high-pitched part,” and the sine tone
sequence was referred to as the “low-pitched part.” The par-
ticipants then heard a sample of a Listen trial containing both
parts for each Rhythm condition (1:1, 1:2, and 3:2 ratios).
Participants were instructed to listen and report, at the
end of the trial, any missing sounds in either part (following
Nozaradan, Zerouali, Peretz, & Mouraux, 2015). Each
Rhythm condition began with two 38.54-sec practice trials,
one that contained an “omitted” beat and one with a “no-
omitted” beat; participants were offered more practice trials
if they desired. The participants completed 12 test trials for
each Rhythm condition, of which two contained missing
beats, as well as the two practice trials. Test trials were
38.5 sec each; therefore, each block (practice and test trials)
comprised approximately 9 min (38.5 sec × 2 practice trials +
38.5 sec × 12 test trials) of testing plus short breaks between
successive trials.
Mathias et al.
1867
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
–
a
p
r
d
t
i
3
c
2
l
1
e
0
–
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
t
f
o
n
0
5
M
a
y
2
0
2
1
Synchronization Task
Participants were first familiarized with the two rhythmic
parts in each sequence by listening to them separately
and then together for each rhythm condition. They then
completed two 38.5-sec practice trials, followed by the
12 test trials for each rhythm condition. Participants
were instructed to listen to four metronome clicks and
then, after the metronome stopped, to start tapping with
their right (dominant) hand at the rate of the metronome
while synchronizing with the presented tone sequence
(synchronization–continuation). They were told that their
goal was to synchronize their taps with the high-pitched
part such that their taps formed the specified rhythmic
ratio (1:1, 1:2, or 3:2 depending on the condition) with
the high-pitched part. Participants completed two practice
trials and 12 experimental trials for each of the 1:1, 1:2,
and 3:2 conditions.
Motor Task
Participants were presented with the four-beat metro-
nome cue at the same tapping rate (528-msec IOI) at
which they tapped across all conditions. They were asked
to tap at the rate presented by the metronome cue during
the 38.5-sec trial until their taps no longer produced
sound, which signaled the end of the trial. Participants
completed two practice trials and 12 experimental trials.
After this condition, participants removed the EEG cap
and received a small compensation.
The entire experiment lasted about 2 hr. During the ex-
periment, participants were monitored by an experimenter
who invited participants to take breaks between trials and
blocks and who offered water between blocks.
Behavioral Data Analysis
Hit rates (percentage of trials with correct detection of
missing tones) and false alarm rates (percentage of trials
with incorrect detection of missing tones) were computed
for each Rhythm condition in the Listen task. Intertap
intervals (ITIs) between consecutive taps were computed
for the Synchronize and Motor tasks as the difference be-
tween each pair of adjacent tap onsets, and mean ITIs
were computed by averaging ITIs within an analysis win-
dow that excluded the first and last four beats of each trial,
leaving 60 taps per trial. Coefficients of variation (CVs) of
ITIs were computed for each trial as the standard devia-
tion divided by the mean ITI. Absolute asynchrony of par-
ticipant taps with the auditory stimulus was computed as
the absolute difference between each tap onset and the
most temporally proximal stimulus onset (|tap onset −
stimulus onset|). Signed asynchrony was computed as
the tap onset minus the most temporally proximal stimu-
lus onset; thus, a negative value indicates anticipatory be-
havior. No participants were classified as outliers (3 SDs or
more from the group mean) in terms of their mean ITIs
or mean absolute asynchronies.
EEG Data Analysis
The EEG data were preprocessed in the EEGLAB software
package (Delorme & Makeig, 2004). Raw continuous EEG
data were first down-sampled to 512 Hz (pop_resample.m)
and referenced to the common average across electrode
sites (pop_reref.m). Independent component analysis
(ICA) was subsequently used to identify and remove stereo-
typical eye blink and lateral eye movement artifacts from
the data analysis (Jung et al., 2000; Bell & Sejnowski, 1995).
ICA was computed on a version of the original data
that was preprocessed using procedures previously shown
to optimize ICA component identification (Debener,
Thorne, Schneider, & Viola, 2010; code for those proce-
dures has been published in Stropahl, Bauer, Debener,
& Bleichner, 2018); the version of the data submitted to
ICA is hereafter referred to as the ICA set. First, the ICA
set was filtered with application of low-pass (40 Hz,
Order 100) and high-pass (1 Hz, Order 500) Hanning win-
dowed sinc finite impulse response filters (pop_firws.m).
Bad channels were then visually identified and removed to
reduce noise contributions to ICA decomposition. Filtered
data were then parsed into short (1-sec) epochs to identify
transient nonstereotypical artifacts (such as sudden body
movements), which are typically short-lasting and nonper-
iodic. Any 1-sec epochs greater than 2 SDs from the mean
activity across segments and channels were identified
(pop_jointprob.m) and removed from the data. Cleaned
data epochs were then submitted to infomax ICA (pop_
runica.m), which reconstructs continuous time series
from epoched data. To account for possible loss of rank
from common average referencing, the option “PCA”
was used in the ICA algorithm to set the number of decom-
posed components to equal one less than the number of
channels. Stereotypical eye artifacts (eye blinks and lateral
eye movements) for each participant were visually identi-
fied from their respective ICA set. ICA weights for each
participant were subsequently applied to their original
continuous data sets to remove identified eye artifacts
(pop_subcomp.m); the same bad channels that were re-
moved from ICA sets were removed from original sets be-
fore application of ICA weights to ensure consistency of
dimensions.
The application of ICA weights to original data sets re-
sulted in artifact-attenuated sets that were subsequently
preprocessed using a different set of procedures tailored
for planned time- and frequency-domain analyses. First,
data in bad channels were spherically interpolated from
neighboring channels (pop_interp.m). Second, additional
noise was removed through the application of low-pass
(20 Hz, Order 1000) and high-pass (0.1 Hz, Order 1000)
Hanning windowed sinc finite impulse response filters
tailored for the planned time- and frequency-domain
analyses (Zamm et al., 2017).
1868
Journal of Cognitive Neuroscience
Volume 32, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
–
a
p
r
d
t
i
3
c
2
l
1
e
0
–
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
/
t
o
n
0
5
M
a
y
2
0
2
1
Trials in the Listen condition with incorrect responses
regarding tone omissions were identified and excluded
from all planned EEG analyses.
PSD
Artifact-corrected EEG data were assessed for spectral
content using a procedure adapted from Zamm et al.
(2017). Continuous data in each trial were segmented into
10.56-sec consecutive (nonoverlapping) epochs, corre-
sponding to 20 taps per epoch at 528 msec per tap. Epoch
edges were multiplied with a 5407-sample (10.56-sec)
Hanning window, corresponding to the duration of each
epoch. Data segmentation allowed for the exclusion of
epochs in trials in which participants responded incorrectly
regarding tone omissions in the Listen task.
Each epoch was subsequently submitted to PSD estima-
tion (pwelch.m in MATLAB), using a window length equiv-
alent to the epoch duration (5407 samples) and no overlap
specified (overlap = []); this implementation uses no sub-
window and is therefore equivalent to Bartlett’s method
(Bartlett, 1950). The PSD was estimated at the stimulus
frequency (1.89, 0.94, and 2.84 Hz in 1:1, 1:2, and
3:2 conditions, respectively), the tap frequency (1.89 Hz
across all conditions), and the shared frequency (0.94 Hz
across all conditions), as described earlier. The resulting
power spectra were log-transformed (10*log10, dB conver-
sion) and subsequently averaged across trials for each
channel and every participant. Similar to other EEG studies
(Zamm et al., 2017; Tierney & Kraus, 2014; Nozaradan
et al., 2011, 2012), a noise reduction procedure was then
applied to ensure reduced influence of residual spectral
noise on each channel, by subtracting from each frequency
the mean power at ±3 neighboring frequency bins, corre-
sponding to ±0.1875 Hz. The noise reduction outcomes
could yield a flat spectrum centered around 0 if the signal
contained only noise or a peak resulting from the noise-
subtracted spectrum if the signal contained nonnoise com-
ponents. Immediately adjacent frequencies were included
in the noise estimates to capture a point-by-point estimate
of spectral change (Zamm et al., 2017).
Noise-subtracted spectra were averaged across all
channels for each participant (Tierney & Kraus, 2014;
Nozaradan et al., 2011, 2012), and PSDs at the target
frequencies of 1.89, 0.94, and 2.84 Hz were extracted
from each participant’s power spectrum and exported
for subsequent analyses. Auditory and motor ROIs were
defined by the electrodes that displayed maximal PSD
across participants in grand-averaged topographies for
Listen (averaged across the three Rhythm conditions)
and Motor tasks respectively, following Nozaradan et al.
(2012); these ROIs were electrode FCz in the Listen task
and electrode C3 in the Motor task. Both electrodes (FCz
and C3) were evaluated as ROI in the Synchronize task.
None of the sensors within these ROIs included interpo-
lated channels.
ERPs
Both tap- and stimulus-locked ERPs were computed. EEG
data were segmented into 600-msec epochs time-locked
to the participants’ taps and to the auditory stimuli with
a 100-msec baseline period. Epochs time-locked to miss-
ing tones in the Listen condition were excluded from the
analyses. Average ERP waveforms were computed for each
participant for the Listen and Synchronize conditions.
Amplitudes within a stereotypical N1 time window were
statistically evaluated at electrodes Fz and FCz at a latency
of 80–120 msec after previous research on auditory–motor
tasks (Mathias, Gehring, & Palmer, 2019; Mathias et al., 2017;
Horváth & Burgyán, 2013; Barry, 2009; Katahira, Abla,
Masuda, & Okanoya, 2008). The Fz and FCz sensors did
not correspond to interpolated channels for any participant.
RESULTS
Behavioral Results
Listen Task
Successful detection of missing sounds in the Listen trials
was measured by the hit rate (percentage of trials with cor-
rect detection). Responses indicating a missing sound when
one did not occur were measured by the false alarm rate
(percentage of trials with incorrect detection). The mean
false alarm rate was 1.4% in the 1:1 Rhythm condition,
6.2% in the 1:2 condition, and 4.1% in the 3:2 condition.
Because false alarm rates were very low in all conditions
(less than one trial per condition or 10%), the analysis of
Rhythm condition effects focused on hit rates. A one-way
repeated-measures ANOVA on mean hit rates by Rhythm
condition yielded a significant main effect, F(1, 28) =
1128.89, p < .001. The mean hit rate was 96.6% in the 1:1
condition (SE = 2.4%), 91.4% in the 1:2 condition (SE =
3.6%), and 86.2% in the 3:2 condition (SE = 4.2%). Tukey
post hoc tests revealed that the mean hit rate was larger in
the 1:1 condition than in the 3:2 condition (Tukey HSD =
9.46, α = .05). Thus, participants were least accurate at
detecting the missing sound onsets in the complex 3:2
Rhythm condition.
Synchronize Task
Mean ITIs were 528 msec (SE = 0.4 msec) in the 1:1 con-
dition, 529 msec (SE = 0.4 msec) in the 1:2 condition, and
530 msec (SE = 2.7 msec) in the 3:2 condition. The ITIs did
not significantly differ between conditions ( p > .05). There
was a significant effect of Rhythm condition on the mean
CV, F(2, 56) = 8.18, p < .001. The mean CV was larger in
the 3:2 Rhythm condition (mean CV = 0.075) than in the
1:1 condition (mean CV = 0.05) and the 1:2 condition
(mean CV = 0.055; Tukey HSD = 0.0195, α = .01).
Absolute asynchronies between tap and stimulus onsets
in the Synchronize trials were computed for each Rhythm
condition. There was a significant effect of Rhythm
Mathias et al.
1869
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
0
-
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
.
t
o
n
0
5
M
a
y
2
0
2
1
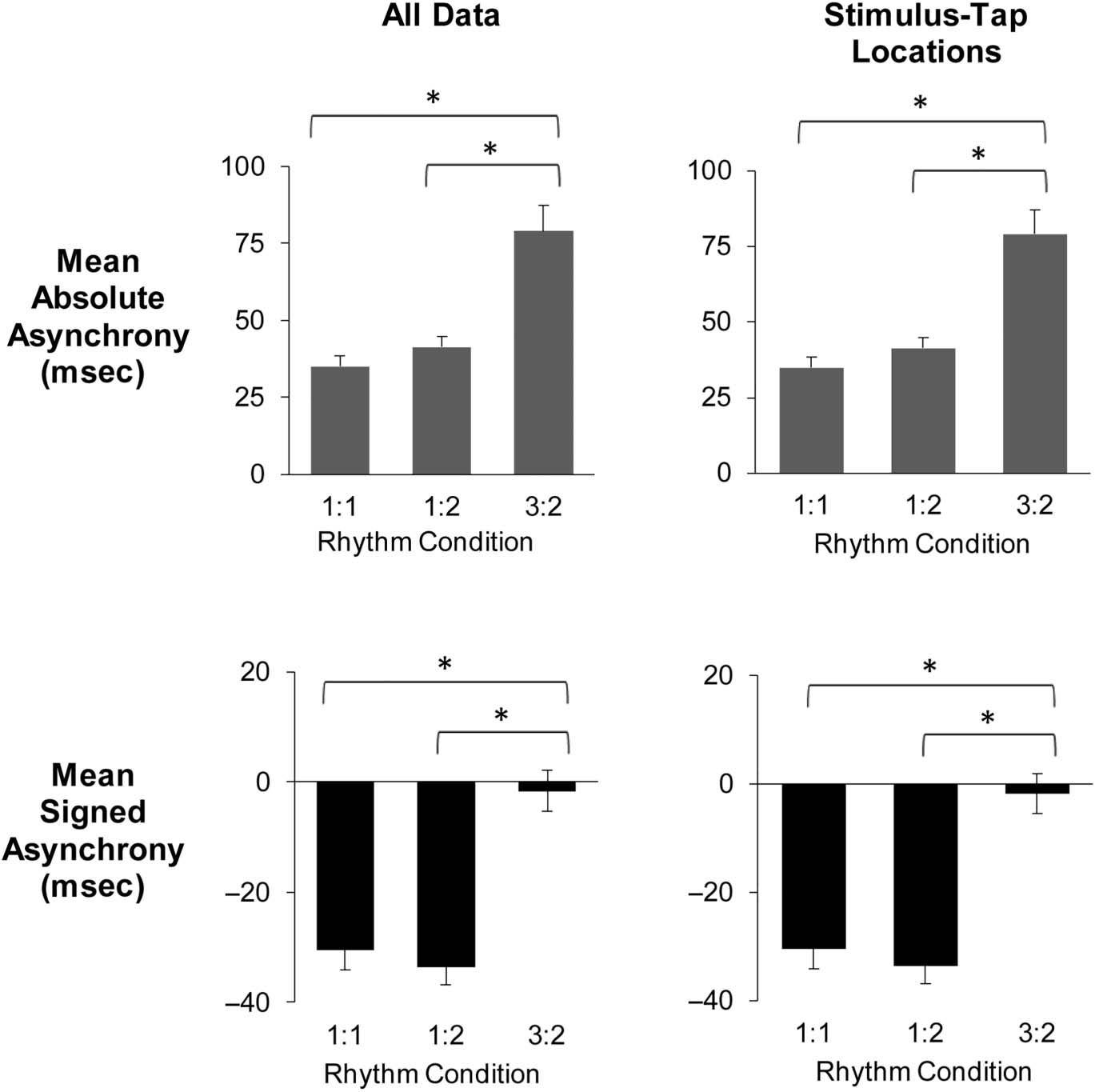
Figure 2. Mean absolute
asynchrony of participants’
taps (top) and mean signed
asynchronies (tap onset minus
stimulus onset; bottom) in the
Synchronization task by Rhythm
condition. Analyses based on all
tap events are shown in the left
column; analyses of the subset
of taps at the shared frequency
(taps that aligned with stimulus
onsets across Rhythm conditions,
circled in Figure 1) are shown in
the right column. *p < .01. Error
bars represent 1 SE.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
0
-
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
f
.
o
n
0
5
M
a
y
2
0
2
1
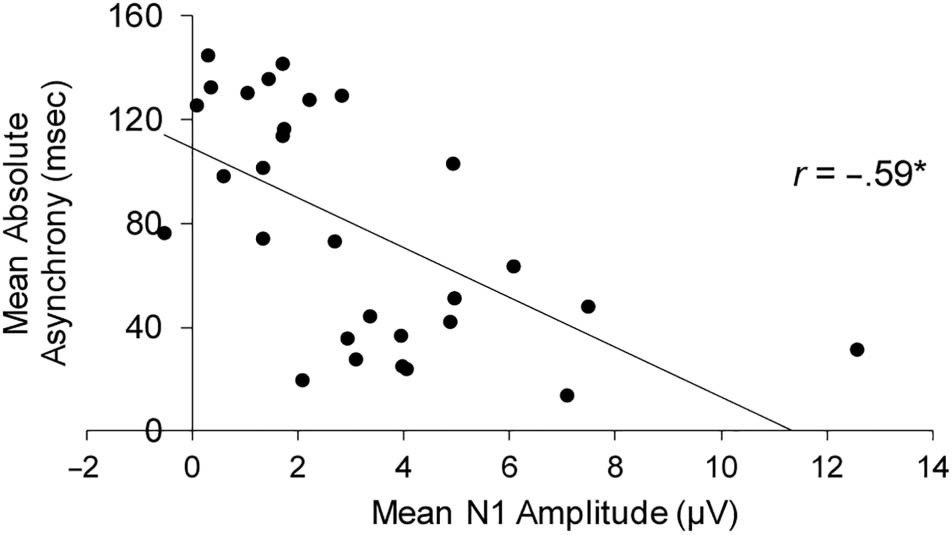
condition on absolute asynchrony values, F(2, 56) = 30.08,
p < .001. As indicated in Figure 2, participants were more
asynchronous in the 3:2 condition than in the 1:1 and 1:2
conditions (Tukey HSD = 18.59, α = .01). Thus,
participants showed greater variability as well as reduced
synchronization accuracy in the 3:2 Rhythm condition.
As shown in Figure 1, the subset of participants’ taps that
aligned temporally with stimulus onsets varied across the
1:1, 1:2, and 3:2 Rhythm conditions. To control for differ-
ences in number of asynchrony values among rhythm con-
ditions, we re-analyzed the subset of asynchronies for taps
that aligned with stimulus onsets. The same analysis re-
peated on these absolute asynchronies confirmed the main
effect of Rhythm condition, F(2, 56) = 30.16, p < .001.
Participants were more asynchronous in the 3:2 Rhythm
condition than in the 1:1 and 1:2 conditions (Tukey HSD =
18.58, α = .01). The standard deviations of absolute asyn-
chronies for the same subset of taps were also re-analyzed;
the ANOVA yielded the same main effect of rhythm con-
dition, F(2, 56) = 52.11, p < .001. Participants’ asyn-
chronies were more variable in the 3:2 Rhythm condition
than in the 1:1 and 1:2 conditions (Tukey HSD = 9.96,
α = .01). Thus, analysis of asynchronies that controlled
for the number of stimulus–tap events yielded the same
results as the analysis of all participant taps.
Signed asynchronies (tap onset − stimulus onset) in the
Synchronize trials were computed for each Rhythm
condition. There was a significant effect of Rhythm condi-
tion on signed asynchrony values, F(2, 56) = 32.07, p <
.001. As indicated in Figure 2, participants’ signed asyn-
chronies were significantly more anticipatory in the 1:1
and 1:2 conditions than in the 3:2 condition (Tukey HSD =
12.32, α = .01). A reanalysis of the subset of asynchronies
for the taps that aligned with stimulus onsets that are
shared across the Rhythm conditions (circled in Figure 1)
confirmed the main effect of Rhythm condition, F(2, 56) =
37.03, p < .001. Participants’ signed asynchronies were
significantly more negative in the 1:1 and 1:2 conditions
than in the 3:2 condition (Tukey HSD = 12.34, α = .01).
Thus, participants’ taps showed greater anticipation of
the auditory stimulus in the 1:1 and 1:2 Rhythm conditions
than in the 3:2 Rhythm condition.
Motor Task
Participants’ mean ITI in the Motor task was 514 msec
(SE = 3 msec), slightly shorter than the prescribed inter-
val of 528 msec. A regression analysis predicting the
mean ITI by the serial position within each trial (n =
62 ITIs) revealed that the participants sped up and short-
ened the tapping interval by 0.12 msec per ITI (r = −.76,
p < .01). The mean CV of ITIs was 0.062 (SE = 0.0067),
similar to the mean CV in Synchronize trials (M = 0.060).
These results served as a control for the accuracy and
1870
Journal of Cognitive Neuroscience
Volume 32, Number 10
precision of participants’ tapping when an external audi-
tory stimulus is absent.
In summary, both perceptual detection of missing
stimulus onsets and synchronization were enhanced in
response to simple multivoiced rhythms compared to
complex rhythms.
EEG Results
PSD
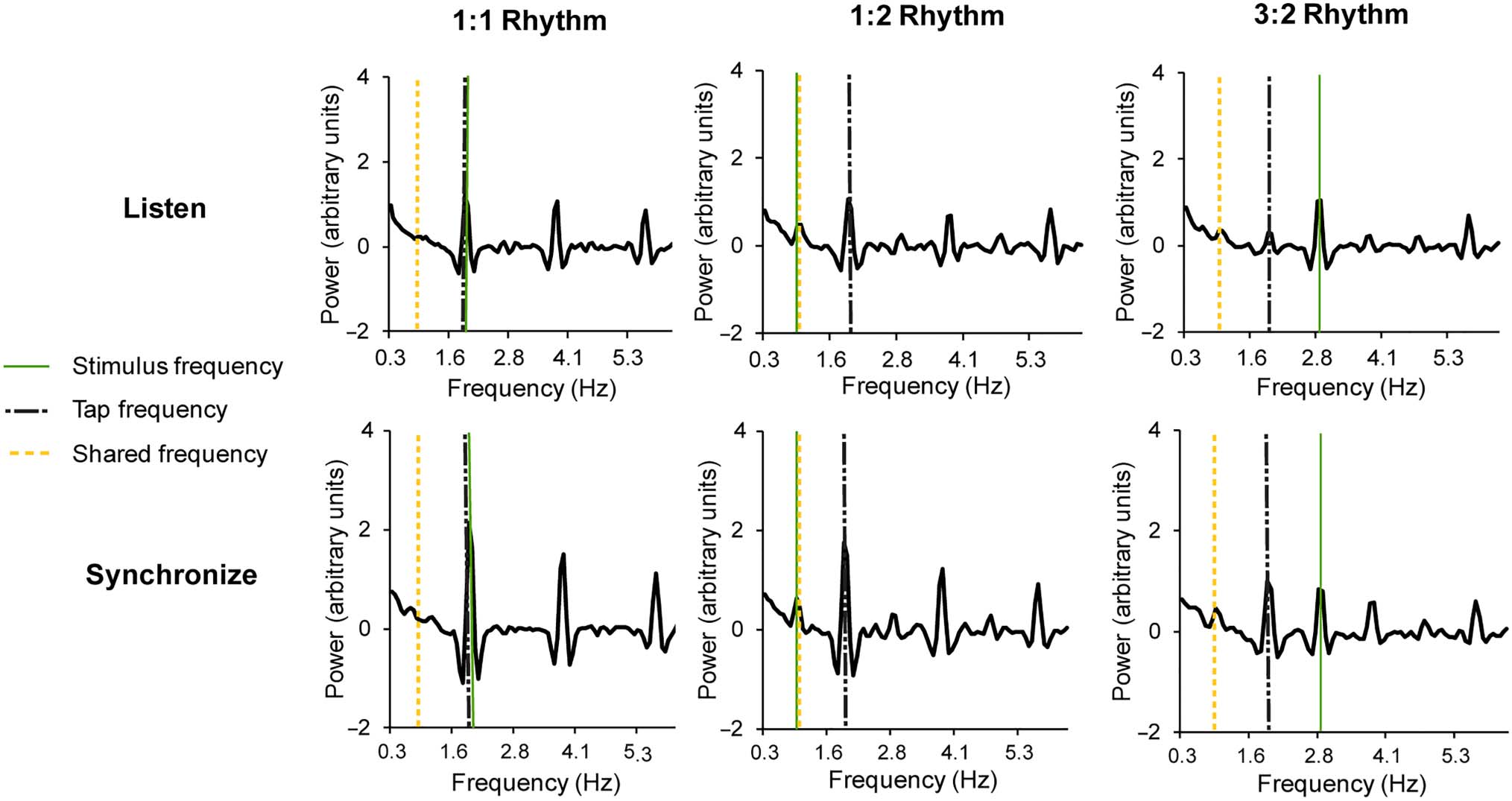
Figure 3 shows the mean power spectra for each task and
Rhythm condition averaged across electrodes. Prominent
peaks occurred at or near frequencies corresponding to
the stimulus rates (0.94, 1.89, and 2.84 Hz) and multiples
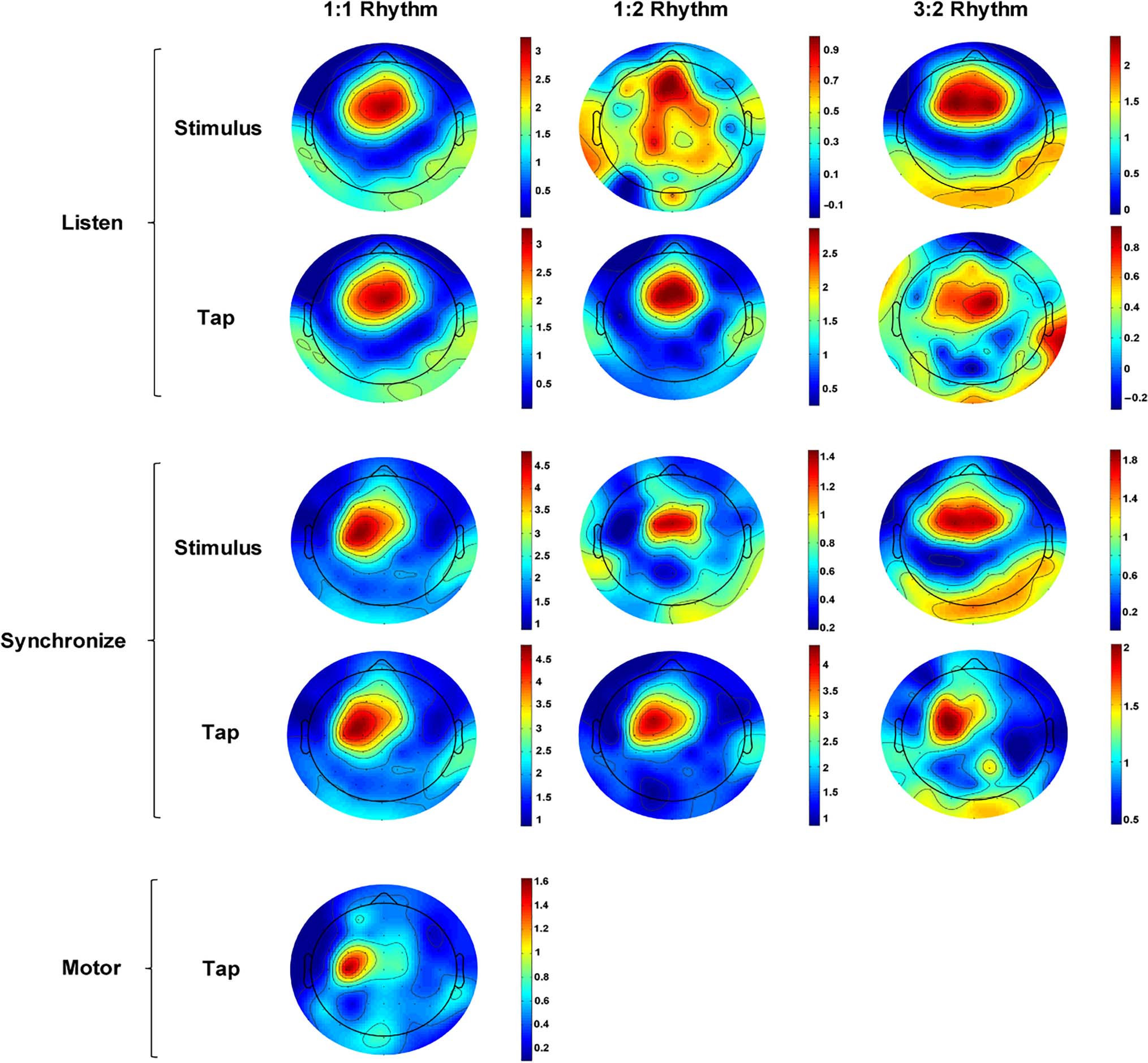
of the stimulus rate. Topographic maps of the peak PSD
for each task, Rhythm condition, and frequency of inter-
est are shown in Figure 4, which indicate characteristic
patterns of auditory cortex activity with maximum at fron-
tal midline electrodes (FCz) in the Listen task, sensorimo-
tor activity at left central electrodes (C3) in the Motor
task, and both activities in the Synchronize task.
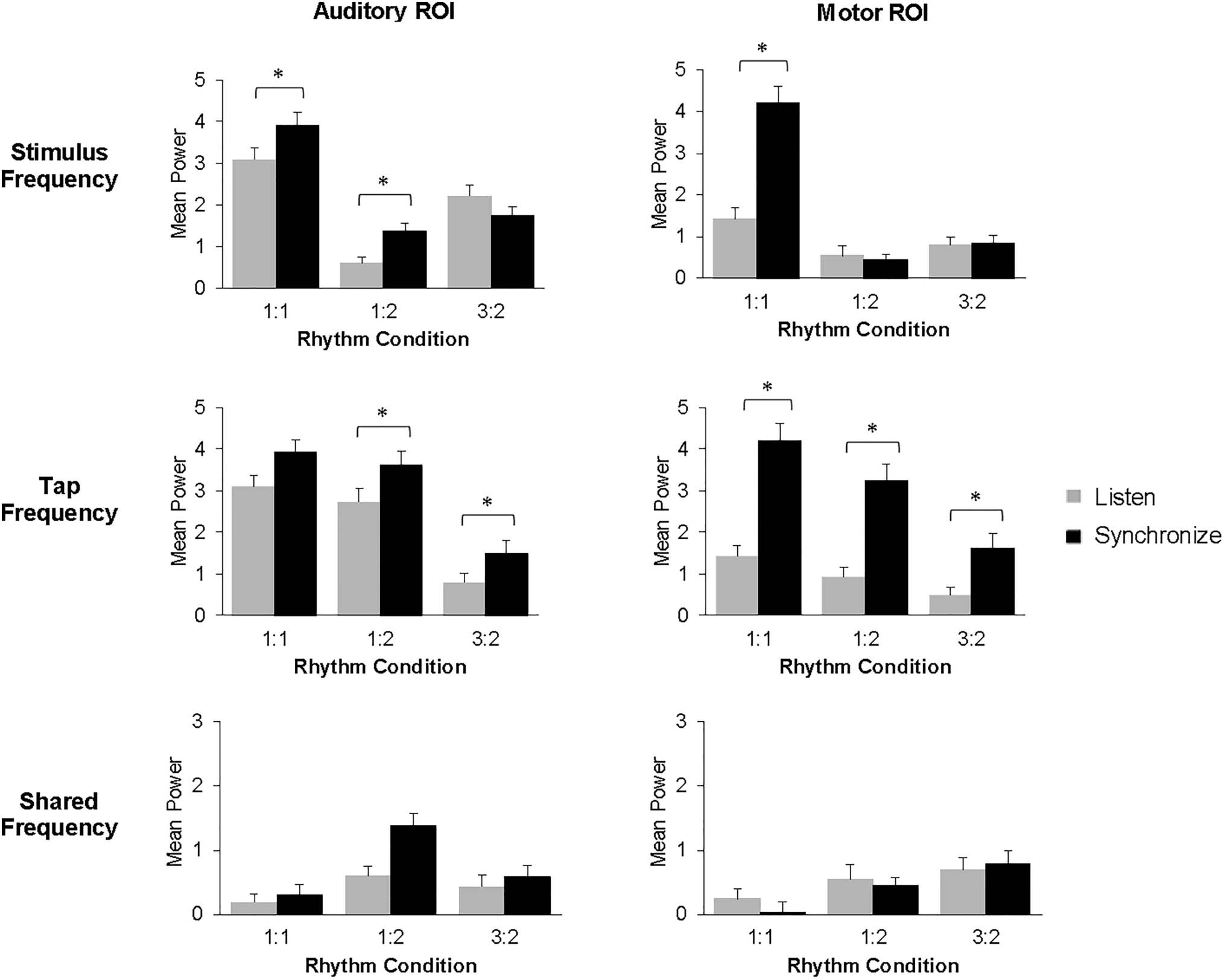
Stimulus frequency effects. A three-way ANOVA was
conducted on the mean spectral power at the stimulus fre-
quency with the factors Task (Listen/Synchronize),
Rhythm condition, and ROI (auditory/motor). We com-
pared the frequency of the stimulus voice that participants
tapped along with (did not produce) in the Synchronize
task with the corresponding stimulus frequency in the
Listen task.
The ANOVA revealed a main effect of Task, F(1, 28) =
29.35, p < .001. Spectral power at the stimulus frequency
was greater during the Synchronize than during the Listen
tasks. The ANOVA also revealed main effects of Rhythm
condition, F(2, 56) = 74.53, p < .001, and ROI, F(1, 28) =
31.54, p < .001. Mean spectral power was significantly
greater at the 1:1 Rhythm condition than at the 3:2 condi-
tion, and the 3:2 condition was significantly greater than
the 1:2 condition (Tukey HSD = 0.49, p < .05). There
was also a significant Task × Rhythm condition interac-
tion, F(2, 56) = 23.93, p < .001, and significant Task ×
Rhythm condition × ROI interaction, F(2, 56) = 13.67,
p < .001. To pursue the interactions, two-way ANOVAs
were conducted on spectral power at the stimulus fre-
quency for each ROI separately.
The mean spectral power present at the stimulus fre-
quency in the auditory ROI differed significantly by Task,
F(1, 28) = 6.38, p < .05, and by Rhythm condition, F(2,
56) = 55.92, p < .001, with greater power during
Synchronize than Listen tasks, and for the 1:1 Rhythm than
for other Rhythm conditions (Tukey HSD = 0.74, p < .01).
The interaction of Task and Rhythm condition was also sig-
nificant, F(2, 56) = 6.96, p < .01. As shown in Figure 5, the
power for the Listen task was greatest in 1:1, followed by
3:2, and least in 1:2 Rhythms; power in the Synchronize task
was greater for 1:1 than for other rhythms (Tukey HSD =
0.82, p < .05).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
0
-
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
f
t
/
o
n
0
5
M
a
y
2
0
2
1
Figure 3. Mean power spectra for the Listen task (top) and the Synchronize task (bottom) by Rhythm condition (1:1, 1:2, and 3:2 ratios). Stimulus
frequencies for each condition are shown with a continuous vertical line, the tapping frequency (1.89 Hz) is shown with a dotted vertical line, and the
shared frequency (0.94 Hz) is indicated with a dashed vertical line.
Mathias et al.
1871
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
0
-
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
.
f
t
o
n
0
5
M
a
y
2
0
2
1
Figure 4. Mean PSD scalp topographies at the stimulus frequencies (1.89 Hz in the 1:1 Rhythm condition, 0.94 Hz in the 1:2 Rhythm condition, and
2.84 Hz in the 3:2 Rhythm condition), Tap frequency (1.89 Hz) by Rhythm condition. Top: Listen task. Center: Synchronize task. Bottom: Motor task.
Scalp topographies are scaled within condition, ranging from the minimum PSD to the maximum PSD.
The same two-way ANOVA on the spectral power pres-
ent in the motor ROI also indicated main effects of Task,
F(1, 28) = 29.23, p < .001, and of Rhythm condition,
F(2, 56) = 49.01, p < .001. Mean spectral power was greater
for Synchronize than for Listen tasks and for 1:1 Rhythm
conditions than for 1:2 or 3:2 Rhythms (Tukey HSD =
0.77, p < .01). There was a significant interaction of Task
with Rhythm condition, F(2, 56) = 30.38, p < .001. As
shown in Figure 5, the 1:1 Rhythm condition in the Listen
task yielded more power than the 1:2 and 3:2 conditions
(Tukey HSD = 1.04, p < .01). The 1:1 Rhythm condition
in the Synchronize task also yielded significantly greater
power than all other rhythms in the Synchronize task and
all Rhythm conditions in the Listen task (Tukey HSD = 1.04,
p < .01).
In summary, effects of the stimulus frequency indicated
that the motor ROI exhibited greater mean power during
the Synchronize task with the 1:1 Rhythm, compared to
the other Rhythm conditions and to the Listen task. As
expected, the auditory ROI indicated greater power for
the 1:1 Rhythm compared to other rhythms in both the
Listen and Synchronize tasks.
Tap frequency effects. The same ANOVA on mean spec-
tral power at the tap frequency revealed a main effect of
Task, F(1, 28) = 44.19, p < .001. Spectral power at the tap
1872
Journal of Cognitive Neuroscience
Volume 32, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
0
-
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
f
.
o
n
0
5
M
a
y
2
0
2
1
Figure 5. Mean spectral power at the stimulus frequencies (top), tap frequency (middle), and shared frequency (bottom), by Task, ROI, and Rhythm
condition. The left column shows mean power within the auditory ROI, and the right column shows mean power within the motor ROI. Gray bars
represent the Listen task, and black bars represent the Synchronize task. Significant pairwise comparisons are shown between means that differed
only for the stimulus and tap frequencies, as those frequencies showed a significant three-way Task × Rhythm × ROI interaction. *p < .05. Error bars
represent 1 SE.
frequency was greater during the Synchronize condition
than during the Listen condition, as expected. There was
also a main effect of Rhythm condition, F(2, 56) = 65.49,
p < .001, and ROI, F(1, 28) = 8.06, p < .01. Similar to find-
ings at the stimulus frequencies, mean spectral power was
greatest at the 1:1 Rhythm condition; however, power at
the tap frequency was greater in the 3:2 condition than in
the 1:2 condition (Tukey HSD = 0.45, p < .05). There
were significant Task × Rhythm condition interaction,
F(2, 56) = 4.99, p = .01, Task × ROI interaction, F(1, 28) =
15.40, p = .001, Rhythm condition × ROI interaction,
F(2, 56) = 8.13, p = .001, and three-way Task × Rhythm
condition × ROI interaction, F(2, 56) = 3.94, p < .05.
Two-way ANOVAs were conducted at each ROI to address
the complex interactions.
The mean spectral power measured at the auditory ROI
for the tap frequency indicated significant effects of both
tasks, F(1, 28) = 14.33, p < .01, and of Rhythm condition,
F(2, 56) = 60.97, p < .001. As expected, spectral power at
the tap frequency was greater in Synchronize than in
Listen conditions; power was greater in the 1:1 and 1:2
conditions compared to the 3:2 condition (Tukey HSD =
0.70, p < .01). There was no significant interaction.
The mean spectral power measured at the motor ROI
for the tap frequency also yielded significant main effects
of Task, F(1, 28) = 43.02, p < .001), and of Rhythm condi-
tion, F(2, 56) = 32.96, p < .001. In addition, the interaction
of Task and Rhythm condition was significant, F(2, 56) =
10.40, p < .001). As shown in Figure 5, spectral power
at the motor ROI was greater for Synchronize tasks than
for Listen tasks and greater for the 1:1 Rhythm, followed
by the 1:2 Rhythm and the 3:2 Rhythm (Tukey HSD =
0.66, p < .01). In addition, spectral power was greater in
the Synchronize task for all three Rhythm conditions than
Mathias et al.
1873
in the Listen task, with the largest difference between
tasks in the 1:1 Rhythm condition (Tukey HSD = 0.94,
p < .01).
In summary, analyses of spectral power at the tap fre-
quency showed similar findings to analyses at the stimu-
lus frequency. Spectral power was greater in response to
the simple 1:1 Rhythm than the more complex rhythms.
Both auditory and motor ROIs showed greater power for
the Synchronize task compared with the Listen task.
Shared frequency results. The same three-way ANOVA
conducted on spectral power at the shared frequency
(0.94 Hz) across Rhythm conditions (circled in Figure 1)
revealed a main effect of Rhythm condition, F(2, 56) =
8.78, p < .001. Spectral power was significantly greater
for the 1:2 and 3:2 Rhythm conditions compared to the
1:1 condition (Tukey HSD = 0.42, p < .001). There were
also significant Task × ROI interaction, F(1, 28) = 1.54,
p < .05, and Rhythm condition × ROI interaction, F(2,
56) = 4.78, p < .05. Two-way ANOVAs were conducted
on each ROI to pursue the complex interactions.
The two-way ANOVA on spectral power in the auditory
ROI at the shared frequency indicated significant main effects
of Task, F(1, 28) = 4.82, p < .05, and Rhythm condition,
F(2, 56) = 9.96, p < .001; there were no significant inter-
actions. As shown in Figure 5, spectral power was greater
in Synchronize tasks than in Listen tasks. In contrast to
stimulus frequency and tap frequency findings, spectral
power at the shared frequency was greatest in the 1:2
Rhythm condition and less in the 1:1 and 3:2 conditions
(Tukey HSD = 0.38, p < .05).
The two-way ANOVA on spectral power in the motor
ROI at the shared frequency indicated a significant main
effect of Rhythm condition, F(2, 56) = 4.94, p < .05.
Spectral power was significantly greater in the 3:2 Rhythm
condition than in the 1:1 Rhythm condition (Tukey HSD =
0.59, p < .01). There were no significant main effects of
Task or interaction. Thus, in contrast to the auditory ROI
findings of greatest power at the shared frequency for the
1:2 Rhythm, the motor ROI indicated increased power for
the 3:2 (most difficult) condition.
ERPs
Effects of rhythmic complexity on the amplitude of ERP
waveforms were examined in the Listen and Synchronize
tasks. Figure 6 shows the grand-averaged ERP waveforms
time-locked to tap onsets and to stimulus onsets. To con-
trol for potential differences in the number of stimulus
tones between Rhythm conditions, we analyzed event-
related responses elicited by taps and stimuli for only the
subset of locations at which taps and stimuli aligned across
Rhythm conditions (circled in Figure 1). This ensured that
the same number of events was included across Rhythm
conditions as well as in stimulus-locked and tap-locked
analyses in the Synchronization condition. The onset times
for stimulus-locked ERPs elicited by high- and low-pitched
stimuli (the two parts of the auditory stimuli) were identi-
cal in the Listen condition; onset times for stimulus- and
tap-locked ERPs were not identical for the Synchronize
condition, because participants did not always tap synchro-
nously with the stimulus.
Listen task. We first assessed ERP amplitudes in the N1
time window across Rhythm conditions in the Listen task,
shown in Figure 7. A one-way ANOVA on stimulus-locked
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
0
-
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
t
.
f
o
n
0
5
M
a
y
2
0
2
1
Figure 6. Grand-averaged ERPs elicited during the Listen (top left), Motor (top right), and Synchronize (bottom) tasks by Rhythm condition (1:1, 1:2,
and 3:2) at electrode Fz. ERPs time-locked to the stimulus onsets are shown in the left column. ERPs time-locked to participant taps are shown in the
right column. Shaded areas indicate the N1 ERP component. Negative values are plotted upward.
1874
Journal of Cognitive Neuroscience
Volume 32, Number 10
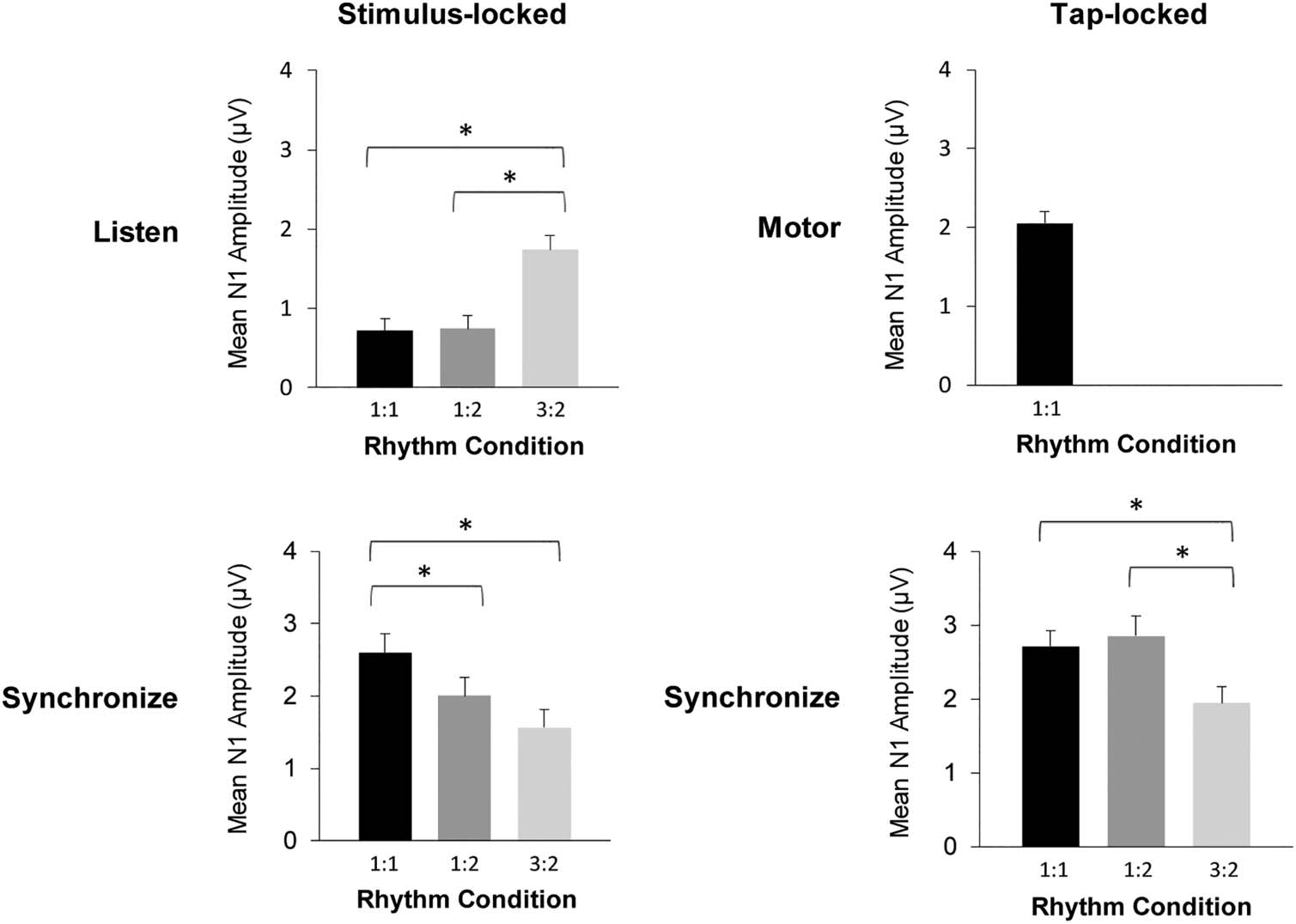
Figure 7. Mean N1 amplitude
values in the Listen (top left),
Motor (top right), and
Synchronize (bottom) tasks by
Rhythm condition (1:1, 1:2, and
3:2). Stimulus-locked N1
amplitudes are shown in the left
column, and tap-locked N1
amplitudes are shown in the
right column. Positive values are
plotted upward. *p < .05. Error
bars represent 1 SE.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
1
e
0
-
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
t
.
o
n
0
5
M
a
y
2
0
2
1
mean amplitudes in the N1 time window yielded a main
effect of Rhythm, F(2, 56) = 24.53, p < .001. The stimulus
tones in the 3:2 Rhythm condition elicited less negative
N1 amplitudes than in both the 1:1 and 1:2 Rhythm con-
ditions (Tukey HSD = 0.50, p < .01). Thus, participants
demonstrated a reduction in mean amplitudes within the
N1 time window while listening to the 3:2 Rhythm com-
pared to the 1:1 and 1:2 Rhythms.
Synchronization task. We assessed mean amplitudes in
the N1 time window across rhythms in the Synchronize
task (also shown in Figure 7). A one-way ANOVA on
stimulus-locked amplitudes yielded a main effect of
Rhythm, F(2, 56) = 13.11, p < .001. Both the 3:2 and 1:2
Rhythm conditions elicited more negative mean ampli-
tudes than the 1:1 condition (Tukey HSD = 0.49, p <
.05). The tap-locked amplitudes in the synchronization
task also yielded a main effect of Rhythm, F(2, 56) =
10.08, p < .001. The 3:2 Rhythm condition elicited more
negative amplitudes in the N1 time window than both
the 1:1 and 1:2 conditions (Tukey HSD = 0.66, p < .01).
Thus, amplitudes in the N1 time window that were time-
locked to both taps and to stimuli were more negative for
the 3:2 Rhythm condition than the 1:1 condition.
Motor task. We compared mean amplitudes in the N1
time window elicited during the Motor task with mean
amplitudes observed in the Listen and Synchronize tasks.
A one-way ANOVA conducted on mean amplitudes for the
1:1 Rhythm (tap-locked N1 amplitudes in the Motor and
Synchronize tasks and stimulus-locked amplitudes in the
Listen task) yielded a main effect of Task, F(2, 56) = 56.8,
p < .001. Amplitudes in the N1 time window were more
suppressed (more positive) in the Motor and Synchronize
conditions compared to the Listen condition and less sup-
pressed (more negative) in the Motor condition compared
to the Synchronize condition (Tukey HSD = 0.58, p < .01).
Correlations between behavioral asynchrony, N1
amplitudes, and individual differences in musical
practice. We tested the relationship between partici-
pants’ asynchronies, amplitudes in the N1 time window,
and amount of weekly musical practice (which ranged from
0 to 30 hr/week) in the most complex synchronization con-
dition, the 3:2 Rhythm condition. A multiple linear regres-
sion was conducted to predict absolute asynchronies in
the 3:2 condition from participants’ hours of weekly instru-
mental practice, stimulus- and tap-locked ERP amplitudes
within the N1 time window in the 3:2 condition, and PSD
measures at the stimulus and tap frequencies in the 3:2
condition. A significant regression was observed, R2 = .74,
F(5, 23) = 5.50, p < .005. Semi-partial correlations indicated
significant contributions to asynchrony from stimulus-
locked amplitudes, β = −.73, t(23) = 2.76, p < .05, and
from hours of weekly practice, β = −.38, t(23) = 2.39,
p < .05. As stimulus-locked amplitudes in the N1 time win-
dow became more positive, absolute asynchrony in the 3:2
Rhythm condition decreased, and as weekly practice in-
creased, absolute asynchrony in the 3:2 Rhythm condition
decreased. The two significant predictors did not correlate
with each other, r(27) = −.12, p = .53. Thus, musical prac-
tice and stimulus-locked amplitudes in the N1 time window
yielded independent contributions to absolute asynchrony
in the 3:2 Synchronize condition.
Mathias et al.
1875
accompanies movement in natural synchronization tasks
such as music performance (for a review, see Palmer,
2013), the current design provides a step forward in iden-
tifying mechanisms of auditory–motor coordination un-
der more natural feedback conditions comparable to
musicians’ performance.
The musicians performed at high levels of overall
accuracy in both perceptual and production tasks.
Nonetheless, their behavioral responses indicated effects
of rhythmic complexity in both perceptual and produc-
tion tasks. Detection rates for missing beats in the
Listen task worsened in the most complex (3:2) rhythm
condition. Participants were most variable in tapping du-
rations and least synchronous with stimulus onsets in the
3:2 rhythm condition in the Synchronize task, consistent
with previous findings that behavioral entrainment de-
creases as rhythmic complexity increases (Chapin et al.,
2010; Collier & Wright, 1995). Finally, participants’ tap-
ping accuracy remained high in the Motor task (in the
absence of other auditory stimuli). Importantly, partici-
pants’ accuracy of tapping the fixed frequency was equiv-
alently high across Synchronize and Motor tasks, allowing
comparison of neural entrainment in the presence of
equivalent behavior. Several studies document musi-
cians’ greater accuracy and precision in producing
rhythms (Summers, Rosenbaum, Burns, & Ford, 1993;
Summers & Kennedy, 1992) and perceiving rhythms
(Manning & Schutz, 2016). Thus, musicians’ rhythmic be-
havior, often near ceiling, provides a conservative test of
the hypothesis that rhythmic complexity modulates syn-
chronization performance.
Neural measures of entrainment to the stimulus and
tap frequencies also decreased as rhythmic complexity in-
creased during Listen and Synchronize tasks. Specifically,
the 1:1 rhythm condition elicited greater entrainment
(higher PSD) at stimulus and tap frequencies relative to
other rhythm conditions (1:1, 1:2), at both auditory and
motor ROIs. This finding supports dynamical systems
models of rhythmic entrainment (Large et al., 2015;
Strogatz, 2001), specifically Farey tree frameworks of
rhythmic stability, which suggest that rhythms featuring
simple integer ratios should display more stable entrain-
ment relative to rhythms featuring complex integer ratios
(Bouvet, Varlet, Dalla Bella, Keller, & Bardy, 2017; Peper,
Beek, & Van Wieringen, 1991). The observed relationship
between neural entrainment and rhythmic complexity is
consistent with our behavioral results, which also indicated
enhanced entrainment for simple relative to complex
rhythms. Future research could address how rhythmic
complexity modulates other characteristics of entrainment
than period coupling of neural oscillations with an external
stimulus. For example, entrainment is also characterized by
the dynamic alignment of oscillatory phase with a stimulus
(Bauer, Bleichner, Jaeger, Thorne, & Debener, 2018); the
dynamics of phase alignment may vary as a function of
rhythmic complexity, whereby phase alignment occurs
more rapidly with simple relative to complex rhythms.
Figure 8. Correlation between mean stimulus-locked N1 amplitudes
and mean absolute asynchronies in the 3:2 Rhythm condition of the
Synchronize task. *p < .05.
Figure 8 shows the simple correlation of mean absolute
asynchronies in the 3:2 Rhythm condition with mean
stimulus-locked amplitudes in the N1 time window,
r(27) = −.59, p < .05. Simple correlations of the absolute
asynchronies with mean amplitudes in the N1 time win-
dow and with hours of weekly musical practice did not
reach significance in the 1:1 (stimulus-locked: r(27) =
−.09, p > .05; musical practice: r(27) = −.33, p = .08)
or 1:2 (stimulus-locked: r(27) =.003; musical practice:
r(27) = −.28; ps > .05) Rhythm conditions. No significant
simple correlations between hours of weekly musical prac-
tice and stimulus-locked amplitudes in the N1 time win-
dow were observed. Moreover, there were no significant
correlations between amplitudes in the N1 time window
and PSD amplitudes.
In summary, multiple regression analyses revealed that
more negative amplitudes in the N1 time window and
greater amounts of weekly musical practice were inde-
pendently associated with greater synchronization accu-
racy for complex rhythms.
DISCUSSION
We examined behavioral and neural entrainment during
musicians’ naturalistic perception and production of sim-
ple and complex auditory rhythms. Participants listened
to two-part auditory sequences whose rhythms formed
integer ratios varying in complexity from low (1:1) to
moderate (1:2) and high (3:2) complexity. One of the
two parts was fixed in rate across all rhythmic complexity
conditions, allowing us to compare neural responses to
the two parts under similar conditions. Participants also
performed a synchronization task with the same rhythms
in which they tapped at the fixed rate while synchroniz-
ing with the other auditory part. Finally, participants per-
formed a motor task in which they tapped the same fixed
rate in the absence of other auditory stimuli. In contrast
to many studies of auditory–motor synchronization, par-
ticipants’ taps resulted in auditory feedback in all condi-
tions of the current study. As auditory feedback typically
1876
Journal of Cognitive Neuroscience
Volume 32, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
–
a
p
r
d
t
i
3
c
2
l
1
e
0
–
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
.
f
o
n
0
5
M
a
y
2
0
2
1
Neural entrainment was also modulated by task:
Enhanced EEG spectral density was observed at stimulus
and tap frequencies during synchronization relative to
perception (Listen task), for both auditory and motor
ROIs. This finding is consistent with previous studies
showing enhanced amplitudes of neural oscillations at
musical beat frequencies in perceived and produced tone
sequences (Fujioka et al., 2015; Nozaradan et al., 2011,
2012). Moreover, the current findings extend this work
by revealing consistent effects of synchronization on en-
trainment across simple and complex rhythms.
We also assessed neural entrainment to the frequency at
which stimulus and taps aligned (referred to as the shared
frequency; see Figure 1). This frequency corresponded to
either the stimulus or tap frequency in the 1:1 and 1:2 con-
ditions (stimulus and tap frequencies were integer multi-
ples) but did not correspond to either stimulus or tap
frequency in the 3:2 condition. Most importantly, in the
3:2 condition, the shared frequency emerged from the poly-
rhythmic relationship between stimulus and tap frequen-
cies. Enhanced auditory neural entrainment to the shared
frequency occurred in the Synchronize relative to the
Listen task, suggesting that tapping along with a stimulus
amplifies one’s perception of the frequency at which indi-
viduals align taps with a rhythmic stimulus. Importantly, this
effect cannot be explained by the increased acoustic ampli-
tude at the shared frequency arising from temporally coin-
cident stimulus and tap onsets, as this was controlled
across rhythm conditions and tasks.
We also investigated how rhythmic complexity modulated
event-related cortical responses to tone onsets, specifically
within the time window of the N1 ERP component. The
N1 component has been linked to enhanced auditory
perceptual processing (Nobre & van Ede, 2018; Lange
et al., 2003; Näätänen & Winkler, 1999), as would be
expected for attended frequencies in perceived and pro-
duced rhythms. Increases in rhythmic complexity in both
Listen and Synchronize tasks resulted in more positive
amplitudes in the N1 time window in the 3:2 rhythm con-
dition relative to the 1:1 and 1:2 conditions. Modulation of
the N1 time window by rhythmic complexity contributes
to a growing literature that specifies the mechanisms un-
derlying the occurrence and stability of spontaneous
movement synchronization to auditory rhythms (Bouvet
et al., 2019).
Changes in ERP amplitudes within the N1 time window
were also modulated by whether participants perceived
or produced rhythms. Amplitudes in the Motor and
Synchronize task for the simplest rhythm condition (1:1)
were more positive relative to the Listen task. In addition,
amplitudes in the Motor task were more negative relative to
the Synchronize task. These findings are consistent with the
view that N1 changes reflect motor-induced suppression of
auditory cortical processing (Horváth, 2015; SanMiguel,
Todd, & Schröger, 2013). They also fit with evidence that
the N1 wave of auditory evoked responses is attenuated
in response to sound that is not selectively attended to,
relative to attended sound (Snyder, Alain, & Picton, 2006;
Hillyard et al., 1973). The current findings further indicate
that task complexity modulates the N1; we propose that the
Synchronize task (which presented one auditory part to
track) required a higher level of auditory processing than
was required in the Motor task (with no parts to track)
and a lower level of auditory processing than was required
in the Listen task (with two parts to track).
The observed pattern of amplitudes within the N1 time
window may also be influenced by other mechanisms. It is
possible that overlap of ERP responses to temporally prox-
imal tone onsets may have resulted in waveform cancella-
tion in the N1 analysis time window (carryover effects). For
example, the P1 elicited by a given tap onset could have
overlapped in time with the N1 response to a preceding
stimulus tone, which may have resulted in voltage cancel-
lation and potentially reduced amplitudes within the N1
time window. Amplitudes in stereotypical N1 time win-
dows may also be influenced by refractory effects from
stimulus rates. It is well established that tones presented
in shorter intervals elicit N1 ERPs of a smaller magnitude
than tones presented at longer intervals (Budd et al.,
1998), whereas in contrast, the P1 and P2 waves are less af-
fected by increases in stimulus rate (Gutschalk, Patterson,
Uppenkamp, Scherg, & Rupp, 2004). Decreases in N1 ERP
magnitude at faster stimulus rates may arise from de-
creased excitability of neural generators at intervals smaller
than the refractory period of the underlying network
(Gutschalk et al., 2004). The observed pattern of results
cannot be fully accounted for by any one of these mecha-
nisms; future work should aim to disentangle how each
mechanism may differentially contribute to ERP responses
elicited by rhythms of varying complexity. This could po-
tentially be accomplished using alternative measures of
ERP amplitude such as peak-to-peak approaches (Snyder
et al., 2006), which may disambiguate neighboring ERPs
within an analysis time window.
Finally, musicians’ behavioral asynchrony measures in the
high-complexity (3:2) Synchronize task decreased as ampli-
tudes within the N1 time window became more positive and
the amount of musical practice increased. The more posi-
tive amplitudes were associated with smaller asynchronies
in the 3:2 rhythm condition, suggesting that motor-induced
suppression of auditory cortical processing aided synchroni-
zation; this effect was larger in participants with greater
amounts of musical training. Although amplitudes within
the N1 time window can be sensitive to learning effects,
decreasing as listeners adapt to stimulus repetition over
multiple blocks (Ross, Barat, & Fujioka, 2017), the
Synchronization condition in the current study occurred af-
ter the Listen condition, by which time participants had be-
come familiar with the auditory stimuli from all rhythmic
complexity conditions. Thus, learning-related adaptation is
less likely to account for the decreased amplitudes within
the N1 time window observed during the Synchronize con-
dition. Another experimental ordering consideration is the
role of imagery; a music production task followed by a
Mathias et al.
1877
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
–
a
p
r
d
t
i
3
c
2
l
1
e
0
–
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
t
.
o
n
0
5
M
a
y
2
0
2
1
perceptual task that relies on similar stimulus material can
lead to the use of imagery (auditory or motor) during the
later perceptual task (Mathias, Palmer, Perrin, & Tillmann,
2015; Brown & Palmer, 2013). To avoid these potential im-
agery effects, the current study ordered the Listen task be-
fore the Synchronize and Motor tasks, and participants
were not informed about the Synchronize and Motor tasks
until after the Listen task had been completed. Future
studies may manipulate the order of perception and pro-
duction tasks in the presence of auditory feedback to fur-
ther evaluate learning effects on amplitudes within the N1
time window.
In summary, the current study compared behavioral and
neural responses across perception and production tasks
and levels of rhythm complexity while controlling for the
participants’ tapping rate and while providing auditory
feedback associated with both stimuli and responses.
These controls allowed us to compare directly the neural
responses to simple and complex rhythms presented in
comparable naturalistic conditions in terms of perceptual
stimulation and response rates. Although previous studies
have attempted to control auditory feedback across percep-
tion conditions (Nozaradan et al., 2011, 2012) or response
rates across motor conditions (Mathias et al., 2017), to our
knowledge, no single study has simultaneously controlled
motor production rate and the presence of self-generated
auditory feedback. This study has demonstrated that behav-
ioral and neural entrainment underlies accurate auditory–
motor synchronization and is modulated in similar ways
by rhythmic complexity. Many real-world auditory synchro-
nization tasks—such as group music performance—contain
actions that are accompanied by auditory feedback; the
current study represents a step toward understanding more
naturalistic sensorimotor synchronization behaviors. Future
studies may address how rhythmic complexity modulates
entrainment across individuals in multiperson synchroniza-
tion tasks (such as group music-making) as well as how
nonexperts (such as musical novices) learn to entrain to
complex auditory rhythms.
Acknowledgments
This research was funded in part by an NSF Graduate Fellowship
to B. Mathias, a PBEEE Graduate award from FRQNT to A.
Zamm, an NSERC-USRA award to P. Gianferrara, and NSERC
Grant 298173 and a Canada Research Chair to C. Palmer. We
thank Shelby Trapid, James O’Callaghan, Jamie Dunkle, and
Frances Spidle for assistance.
Reprint requests should be sent to Caroline Palmer, Department
of Psychology, McGill University, Montreal, Quebec, Canada
H3A 1B1, or via e-mail: caroline.palmer@mcgill.ca.
REFERENCES
Barry, R. J. (2009). Evoked activity and EEG phase resetting in
the genesis of auditory Go/NoGo ERPs. Biological Psychology,
80, 292–299.
Bartlett, M. S. (1950). Periodogram analysis and continuous
spectra. Biometrika, 37, 1–16.
Bauer, A. R., Bleichner, M. G., Jaeger, M., Thorne, J. D., &
Debener, S. (2018). Dynamic phase alignment of ongoing
auditory cortex oscillations. Neuroimage, 167, 396–407.
Bays, P. M., & Wolpert, D. M. (2007). Computational principles
of sensorimotor control that minimize uncertainty and
variability. Journal of Physiology, 578, 387–396.
Bell, A. J., & Sejnowski, T. J. (1995). An information-maximization
approach to blind separation and blind deconvolution. Neural
Computation, 7, 1129–1159.
Bood, R. J., Nijssen, M., Van Der Kamp, J., & Roerdink, M.
(2013). The power of auditory–motor synchronization in
sports: Enhancing running performance by coupling cadence
with the right beats. PLoS One, 8, e70758.
Bouvet, C. J., Varlet, M., Dalla Bella, S., Keller, P. E., & Bardy, B. G.
(2017). Auditory–motor entrainment to complex frequency
ratios. In J. A. Weast-Knapp & G. J. Pepping (Eds.), Studies
in Perception and Action XIV: Nineteenth International
Conference on Perception and Action (pp. 45–48). New York:
Routledge.
Bouvet, C. J., Varlet, M., Dalla Bella, S., Keller, P. E., Zelic, G., &
Bardy, B. G. (2019). Preferred frequency ratios for spontaneous
auditory–motor synchronization: Dynamical stability and
hysteresis. Acta Psychologica, 196, 33–41.
Brown, R. M., & Palmer, C. (2013). Auditory and motor imagery
modulate learning in music performance. Frontiers in Human
Neuroscience, 7, 320.
Brown, S., & Parsons, L. M. (2008). The neuroscience of dance.
Scientific American, 299, 78–83.
Budd, T. W., Barry, R. J., Gordon, E., Rennie, C., & Michie, P. T.
(1998). Decrement of the N1 auditory event-related potential
with stimulus repetition: Habituation vs. refractoriness.
International Journal of Psychophysiology, 31, 51–68.
Buzsáki, G., & Draguhn, A. (2004). Neuronal oscillations in
cortical networks. Science, 304, 1926–1929.
Chapin, H. L., Zanto, T., Jantzen, K. J., Kelso, S. J., Steinberg, F.,
& Large, E. W. (2010). Neural responses to complex auditory
rhythms: The role of attending. Frontiers in Psychology,
1, 224.
Chemin, B., Mouraux, A., & Nozaradan, S. (2014). Body
movement selectively shapes the neural representation of
musical rhythms. Psychological Science, 25, 2147–2159.
Collier, G. L., & Wright, C. E. (1995). Temporal rescaling of
simple and complex ratios in rhythmic tapping. Journal of
Experimental Psychology: Human Perception and
Performance, 21, 602–627.
Debener, S., Thorne, J. D., Schneider, T. R., & Viola, F. C.
(2010). Using ICA for the analysis of multi-channel EEG Data.
In M. Ullsperger & S. Debener (Eds.), Simultaneous EEG and
fMRI: Recording, analysis, and application (pp. 121–133).
New York: Oxford Scholarship Online.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
Engel, A. K., Fries, P., & Singer, W. (2001). Dynamic predictions:
Oscillations and synchrony in top–down processing. Nature
Reviews Neuroscience, 2, 704–716.
Finney, S. A. (2001). FTAP: A Linux-based program for tapping
and music experiments. Behavior Research Methods,
Instruments, & Computers, 33, 65–72.
Fitzroy, A. B., & Sanders, L. D. (2015). Musical meter modulates
the allocation of attention across time. Journal of Cognitive
Neuroscience, 27, 2339–2351.
Fujioka, T., Ross, B., & Trainor, L. J. (2015). Beta-band oscillations
represent auditory beat and its metrical hierarchy in perception
and imagery. Journal of Neuroscience, 35, 15187–15198.
Glass, L., & Mackey, M. C. (1988). From clocks to chaos: The
rhythms of life. Princeton, NJ: Princeton University Press.
1878
Journal of Cognitive Neuroscience
Volume 32, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
–
a
p
r
d
t
i
3
c
2
l
1
e
0
–
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
.
t
o
n
0
5
M
a
y
2
0
2
1
Gutschalk, A., Patterson, R. D., Uppenkamp, S., Scherg, M., &
Rupp, A. (2004). Recovery and refractoriness of auditory
evoked fields after gaps in click trains. European Journal of
Neuroscience, 20, 3141–3147.
Haken, H., Kelso, J. A. S., & Bunz, H. (1985). A theoretical
model of phase transitions in human hand movements.
Biological Cybernetics, 51, 347–356.
Hillyard, S. A., Hink, R. F., Schwent, V. L., & Picton, T. W. (1973).
Electrical signs of selective attention in the human brain.
Science, 182, 177–180.
Horváth, J. (2015). Action-related auditory ERP attenuation:
Paradigms and hypotheses. Brain Research, 1626, 54–65.
Horváth, J., & Burgyán, A. (2013). No evidence for peripheral
mechanism attenuating auditory ERPs to self-induced tones.
Psychophysiology, 50, 563–569.
Horváth, J., Maess, B., Baess, P., & Tóth, A. (2012). Action–sound
coincidences suppress evoked responses of the human auditory
cortex in EEG and MEG. Journal of Cognitive Neuroscience,
24, 1919–1931.
Jung, T.-P., Makeig, S., Humphries, C., Lee, T., McKeown, M. J.,
Iragui, I., et al. (2000). Removing electroencephalographic
artefacts by blind source separation. Psychophysiology, 37,
163–178.
Katahira, K., Abla, D., Masuda, S., & Okanoya, K. (2008). Feedback-
based error monitoring processes during musical performance:
An ERP study. Neuroscience Research, 61, 120–128.
Kelso, J. A. S. (1991). Multifrequency behavioural patterns and the
phase attractive circle map. Biological Cybernetics, 64, 485–495.
Lange, K., Rösler, F., & Röder, B. (2003). Early processing stages
are modulated when auditory stimuli are presented at an
attended moment in time: An event-related potential study.
Psychophysiology, 40, 806–817.
Large, E. W., Herrera, J. A., & Velasco, M. J. (2015). Neural
networks for beat perception in musical rhythm. Frontiers in
Systems Neuroscience, 9, 159.
Large, E. W., & Jones, M. R. (1999). The dynamics of attending:
How people track time-varying events. Psychological Review,
106, 119–159.
Large, E. W., & Palmer, C. (2002). Perceiving temporal regularity
in music. Cognitive Science, 26, 1–37.
Manning, F. C., & Schutz, M. (2016). Trained to keep a beat:
Movement-related enhancements to timing perception in
percussionists and non-percussionists. Psychological Research,
80, 532–542.
Mathias, B., Gehring, W. J., & Palmer, C. (2017). Auditory N1
reveals planning and monitoring processes during music
performance. Psychophysiology, 54, 235–247.
Mathias, B., Gehring, W. J., & Palmer, C. (2019). Electrical brain
responses reveal sequential constraints on planning during
music performance. Brain Sciences, 9, 25.
Mathias, B., Palmer, C., Perrin, F., & Tillmann, B. (2015).
Sensorimotor learning enhances expectations during
auditory perception. Cerebral Cortex, 15, 2238–2254.
Miura, A., Kudo, K., & Nakazawa, K. (2013). Action–perception
coordination dynamics of whole-body rhythmic movement in
stance: A comparison study of street dancers and non-dancers.
Neuroscience Letters, 544, 157–162.
Näätänen, R., & Picton, T. (1987). The N1 wave of the human electric
and magnetic response to sound: A review and an analysis of the
component structure. Psychophysiology, 24, 375–425.
Näätänen, R., & Winkler, I. (1999). The concept of auditory
stimulus representation in cognitive neuroscience. Psychological
Bulletin, 125, 826–859.
Nobre, A. C., & van Ede, F. (2018). Anticipated moments:
Temporal structure in attention. Nature Reviews Neuroscience,
19, 34–48.
Nozaradan, S., Peretz, I., & Keller, P. E. (2016). Individual
differences in rhythmic cortical entrainment correlate with
predictive behaviour in sensorimotor synchronization.
Scientific Reports, 6, 20612.
Nozaradan, S., Peretz, I., Missal, M., & Mouraux, A. (2011).
Tagging the neuronal entrainment to beat and meter.
Journal of Neuroscience, 31, 10234–10240.
Nozaradan, S., Peretz, I., & Mouraux, A. (2012). Selective
neuronal entrainment to the beat and meter embedded in a
musical rhythm. Journal of Neuroscience, 32, 17572–17581.
Nozaradan, S., Schönwiesner, M., Caron-Desrochers, L., &
Lehmann, A. (2016). Enhanced brainstem and cortical encoding
of sound during synchronized movement. Neuroimage, 142,
231–240.
Nozaradan, S., Zerouali, Y., Peretz, I., & Mouraux, A. (2015).
Capturing with EEG the neural entrainment and coupling
underlying sensorimotor synchronization to the beat.
Cerebral Cortex, 25, 736–747.
Palmer, C. (2013). Music performance: Movement and
coordination. In D. Deutsch (Ed.), The psychology of music
(3rd ed., pp. 405–422). Amsterdam: Elsevier Press.
Peper, C. E., Beek, P. J., & Van Wieringen, P. C. W. (1991).
Bifurcations in polyrhythmic tapping: In search of Farey
principles. In J. Requin & G. Stelmach (Eds.), Tutorials in
motor neuroscience (pp. 413–431). Dordrecht, The
Netherlands: Springer.
Repp, B. H., & Su, Y. H. (2013). Sensorimotor synchronization:
A review of recent research (2006–2012). Psychonomic
Bulletin & Review, 20, 403–452.
Ross, B., Barat, M., & Fujioka, T. (2017). Sound-making actions
lead to immediate plastic changes of neuromagnetic evoked
responses and induced β-band oscillations during perception.
Journal of Neuroscience, 37, 5948–5959.
SanMiguel, I., Todd, J., & Schröger, E. (2013). Sensory suppression
effects to self-initiated sounds reflect the attenuation of the
unspecific N1 component of the auditory ERP. Psychophysiology,
50, 334–343.
SanMiguel, I., Widmann, A., Bendixen, A., Trujillo-Barreto, N., &
Schröger, E. (2013). Hearing silences: Human auditory
processing relies on preactivation of sound-specific brain
activity patterns. Journal of Neuroscience, 33, 8633–8639.
Schaefer, R. S., Vlek, R. J., & Desain, P. (2011). Decomposing
rhythm processing: Electroencephalography of perceived
and self-imposed rhythmic patterns. Psychological Research,
75, 95–106.
Schroeder, M. (1991). Fractals, chaos, power laws. New York:
Freeman.
Snyder, J. S., Alain, C., & Picton, T. W. (2006). Effects of
attention on neuroelectric correlates of auditory stream
segregation. Journal of Cognitive Neuroscience, 18, 1–13.
Snyder, J. S., & Large, E. W. (2005). Gamma-band activity
reflects the metric structure of rhythmic tone sequences.
Cognitive Brain Research, 24, 117–126.
Sowman, P. F., Kuusik, A., & Johnson, B. W. (2012). Self-initiation
and temporal cueing of monaural tones reduce the auditory N1
and P2. Experimental Brain Research, 222, 149–157.
Strogatz, S. H. (2001). Exploring complex networks. Nature,
410, 268–276.
Stropahl, M., Bauer, A.-K., Debener, S., & Bleichner, M. G. (2018).
Source-modeling auditory processes of EEG data using EEGLAB
and Brainstorm. Frontiers in Neuroscience, 12, 309.
Summers, J. J., & Kennedy, T. M. (1992). Strategies in the
production of a 5:3 polyrhythm. Human Movement Science,
11, 101–112.
Summers, J. J., Rosenbaum, D., Burns, B., & Ford, S. (1993).
Production of polyrhythms. Journal of Experimental Psychology:
Human Perception and Performance, 19, 416–428.
Tajima, M., & Choshi, K. (2000). Effects of learning and
movement frequency on polyrhythmic tapping performance.
Perceptual and Motor Skills, 90, 675–690.
Mathias et al.
1879
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
–
a
p
r
d
t
i
3
c
2
l
1
e
0
–
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
t
f
.
o
n
0
5
M
a
y
2
0
2
1
Thaut, M. (2013). Rhythm, music, and the brain: Scientific
Woodman, G. F. (2010). A brief introduction to the use
foundations and clinical applications. New York: Routledge.
Tierney, A., & Kraus, N. (2014). Neural entrainment to the
rhythmic structure of music. Journal of Cognitive Neuroscience,
27, 400–408.
Wing, A. M., Endo, S., Bradbury, A., & Vorberg, D. (2014).
Optimal feedback correction in string quartet synchronization.
Journal of the Royal Society Interface, 11, 20131125.
of event-related potentials in studies of perception and
attention. Attention, Perception, & Psychophysics, 72,
2031–2046.
Zamm, A., Palmer, C., Bauer, A.-K. R., Bleichner, M. G., Demos,
A. P., & Debener, S. (2017). Synchronizing MIDI and wireless
EEG measurements during natural piano performance. Brain
Research, 1716, 27–38.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
–
a
p
r
d
t
i
3
c
2
l
1
e
0
–
1
p
8
d
6
f
4
/
2
3
0
2
1
/
3
1
9
0
6
/
6
1
8
o
6
c
4
n
_
/
a
1
_
8
0
6
1
2
6
0
0
5
1
3
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
6
n
0
0
1
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
/
t
o
n
0
5
M
a
y
2
0
2
1
1880
Journal of Cognitive Neuroscience
Volume 32, Number 10