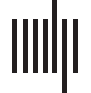REVIEW ARTICLE
The Musical Abilities, Pleiotropy, Language,
and Environment (MAPLE) Framework for
Understanding Musicality-Language
Links Across the Lifespan
Srishti Nayak1,2,4,12
Rachana Nitin1,3
, Peyton L. Coleman1
, Daniel E. Gustavson4,5,13
, Enikő Ladányi1,11
, Simon E. Fisher6,7
,
,
Cyrille L. Magne2,8
, and Reyna L. Gordon1,3,4,9,10,12
1Department of Otolaryngology – Head & Neck Surgery, Vanderbilt University Medical Center, Nashville, TN, USA
2Department of Psychology, Middle Tennessee State University, Murfreesboro, TN, USA
3Vanderbilt Brain Institute, Vanderbilt University, Nashville, TN, USA
4Vanderbilt Genetics Institute, Vanderbilt University Medical Center, Nashville, TN, USA
5Department of Medicine, Vanderbilt University Medical Center, Nashville, TN, USA
6Language and Genetics Department, Max Planck Institute for Psycholinguistics, Nijmegen, The Netherlands
7Donders Institute for Brain, Cognition and Behaviour, Radboud University, Nijmegen, The Netherlands
8PhD Program in Literacy Studies, Middle Tennessee State University, Murfreesboro, TN, USA
9Curb Center for Art, Enterprise, and Public Policy, Vanderbilt University, Nashville, TN, USA
10Vanderbilt Kennedy Center, Vanderbilt University Medical Center, TN, USA
11Department of Linguistics, Potsdam University, Potsdam, Germany
12Vanderbilt University School of Medicine, Vanderbilt University, TN, USA
13Institute for Behavioral Genetics, University of Colorado Boulder, Boulder, CO, USA
Keywords: complex trait genetics, musicality, speech and language development, pleiotropy,
individual differences, neural endophenotypes
ABSTRACT
Using individual differences approaches, a growing body of literature finds positive
associations between musicality and language-related abilities, complementing prior findings
of links between musical training and language skills. Despite these associations, musicality
has been often overlooked in mainstream models of individual differences in language
acquisition and development. To better understand the biological basis of these individual
differences, we propose the Musical Abilities, Pleiotropy, Language, and Environment (MAPLE)
framework. This novel integrative framework posits that musical and language-related abilities
likely share some common genetic architecture (i.e., genetic pleiotropy) in addition to
some degree of overlapping neural endophenotypes, and genetic influences on musically
and linguistically enriched environments. Drawing upon recent advances in genomic
methodologies for unraveling pleiotropy, we outline testable predictions for future research on
language development and how its underlying neurobiological substrates may be supported
by genetic pleiotropy with musicality. In support of the MAPLE framework, we review and
discuss findings from over seventy behavioral and neural studies, highlighting that musicality
is robustly associated with individual differences in a range of speech-language skills required
for communication and development. These include speech perception-in-noise, prosodic
perception, morphosyntactic skills, phonological skills, reading skills, and aspects of
second/foreign language learning. Overall, the current work provides a clear agenda and
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
a n o p e n a c c e s s
j o u r n a l
Citation: Nayak, S., Coleman, P. L.,
Ladányi, E., Nitin, R., Gustavson, D. E.,
Fisher, S. E., Magne, C. L., & Gordon,
R. L. (2022). The musical abilities,
pleiotropy, language, and environment
(MAPLE) framework for understanding
musicality-language links across the
lifespan. Neurobiology of Language,
3(4), 615–664. https://doi.org/10.1162
/nol_a_00079
DOI:
https://doi.org/10.1162/nol_a_00079
Supporting Information:
https://doi.org/10.1162/nol_a_00079
Received: 29 October 2021
Accepted: 8 August 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Srishti Nayak
srishti.nayak@vumc.org
Reyna L. Gordon
reyna.gordon@alumni.usc.edu
Handling Editor:
Robert Zatorre
Copyright: © 2022
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
Understanding musicality-language links
Musicality:
The myriad ways in which humans
interact with music, including
aptitude, skills, engagement,
listening, and enjoyment (Honing,
2018).
framework for studying musicality-language links using individual differences approaches,
with an emphasis on leveraging advances in the genomics of complex musicality and
language traits.
INTRODUCTION
Language and music are ubiquitous forms of communication across the world (Ujfalussy,
1993), and both make use of certain essential acoustic and perceptual parameters that facilitate
effective expression (Jackendoff, 2009; Molino, 2000; Patel, 2003). For example, the ability to
perceive rhythm (key terms are defined in the Glossary within the Supporting Information at
https://doi.org/10.1162/nol_a_00079) is important in both musical and language contexts
(Besson & Schön, 2012; Slevc, 2012). Research has also shown an overlap in the cognitive
and neural processes that are recruited during musical and language-related tasks (Fedorenko
et al., 2009; Kotz et al., 2018; Kunert et al., 2015; Merchant et al., 2015). Based on the overlaps
and dissociations in musical and language processing, many researchers have highlighted the
functional relationships between musicality and language within cognitive and neural frame-
works (Gordon & Magne, 2017; Jantzen et al., 2016; Patel, 2008).
The majority of studies have used one of the four following approaches to map associations,
overlaps, or similarities between musicality and language: (a) musical training or intervention
studies (for reviews, see Tierney & Kraus, 2013; White et al., 2013); (b) comparisons between
musicians and non-musicians on language-related tasks (for a review, see Coffey et al., 2017);
(c) comparisons of neurocognitive processing of linguistic and musical information (Sammler
& Elmer, 2020); and (d) work showing the effects of musical priming in improving language
task performance (Schön & Tillmann, 2015). Much of our current understanding of musicality-
language links comes from these informative study designs; however, these studies largely
obscure preexisting individual differences in musical and language traits. Studying individual
differences allows us to capture the range of human experiences stemming from musical and
language abilities, including social engagement, communication, academic and professional
outcomes, and overall quality of life.
While prevailing explanations for stronger musicality-language links in musicians compared
to non-musicians are often framed in terms of neuroplasticity or cognitive transfer (e.g.,
Bidelman & Alain, 2015; Patel, 2014), many authors have pointed out that individuals may
self-select into higher levels of musical experience and engagement (i.e., musical training,
instrument playing, music listening habits) based on higher levels of musical aptitude (e.g.,
Schellenberg, 2015). This pattern can be seen from very early in development (e.g., Brandt
et al., 2012). For example, both musical aptitude (e.g., music perception skills) and musical
engagement (e.g., musical listening, practice, or training) are traits that show high interindividual
variation within populations. Further, twin studies have shown that these musicality traits are all
moderately heritable—i.e., that genetic variation partly accounts for the observed individual dif-
ferences (Mosing et al., 2014; Seesjärvi et al., 2016; Ullén et al., 2014). This framing presents a
challenge to prior explanations in terms of transfer effects of music on language (Chan et al.,
1998; Kraus & Chandrasekaran, 2010; Moreno, 2009; Slater et al., 2015; Tierney & Kraus,
2013). For example, differential language abilities in groups with different levels of musical
engagement could be driven by who ends up pursuing musical interests, training, or practice.
When considering the human capacity for music and language, it is important to consider
variability in musical skills beyond variability in musical training or experience. For example,
Neurobiology of Language
616
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
studies of individual differences demonstrate positive associations between musical rhythm
abilities and grammatical skills (Gordon, Shivers, et al., 2015; Lee et al., 2020), reading-related
skills (Ozernov-Palchik et al., 2018; Woodruff Carr et al., 2014), prosodic perception (Hausen
et al., 2013; Morrill et al., 2015), and speech discrimination (Swaminathan & Schellenberg,
2020). On the other end of the spectrum, impaired musical rhythm abilities are frequently
comorbid with language-related disorders (Ladányi et al., 2020). Moreover, some frameworks
linking music and language have emphasized the need to understand genetic and develop-
mental factors (Schellenberg, 2020; Zuk & Gaab, 2018), and to account for the potential con-
founding role of genetics when studying environmental factors shaping development (Hart
et al., 2021), such as musical experiences. Given that we only have a nascent understanding
of individual differences in these traits, and of how their relationship evolves over the lifespan,
it is important to consider other developmental and biological explanations for associations
between musical and language traits.
Many traits that show high degrees of interindividual variability, and that fall along a con-
tinuous spectrum, are referred to as complex traits in the genetic literature. Complex trait
phenotypes are polygenic, meaning they are influenced by multiple genes, with effects at each
genetic locus contributing a small amount of variance (Crouch & Bodmer, 2020; Watanabe
et al., 2019). There is emerging evidence that musicality and language-related traits exhibit
polygenicity when investigated in well-powered genomic studies (Doust et al., 2022; Eising
et al., 2022; Niarchou et al., 2022). Further, as we will review in more detail below, traits related
to musicality and language are often phenotypically associated with each other, i.e., interindi-
vidual differences studies reveal significant correlations between tasks performed across the
different domains (e.g., Gordon, Shivers, et al., 2015; Morrill et al., 2015; Yu et al., 2017). These
correlations allow us to leverage principles from the field of genetics to guide discovery of
theorized biological pathways that may be shared between musicality and language traits. In
particular, Cheverud’s conjecture posits that distinct traits showing high phenotypic correla-
tions are likely inherited together and influenced by alleles at a common set of genetic loci
(Cheverud, 1988). Such biological patterns of shared genetic architecture, termed pleiotropy,
have been demonstrated in many complex traits (Sodini et al., 2018). Similarly, based on known
cross-trait correlations, musicality and language-related traits are likely influenced (at least in
part) by genetic variations at the same genes (Bulik-Sullivan et al., 2015; Wesseldijk et al.,
2021). Although, note that given their polygenic nature, this does not discount the existence
of genetic effects that contribute uniquely to musicality and language traits.
The overarching objective of this work is to present a framework for investigating biological
relationships between musical and language traits, in the context of existing behavioral, neu-
ral, and genetic evidence for associations between these domains. In this article, we introduce
a framework, as well as a set of testable hypotheses, for understanding the biological and
environmental mechanisms by which polygenic pleiotropy between musicality and speech-
language traits can result in observed correlations between musical abilities and language-
related abilities. While the environmental effects of music on cognition have traditionally been
explained via models of neuroplasticity (e.g., Bidelman & Alain, 2015; Patel, 2014), behav-
ioral genetics findings allow us to reimagine what musical environments can tell us. For exam-
ple, alongside their influence on musical and language abilities, genes also exert influence on
music-related environments (e.g., music practice: Mosing et al., 2014; music instrument
engagement: Gustavson et al., 2021).
Further, potential neural endophenotypes highlighted from current evidence can help clar-
ify cascading biological mechanisms of musicality-language links across lifespan develop-
ment. Endophenotypes can be thought of as intermediate biological phenotypes that are
617
Phenotype:
A trait of interest that can be
measured or observed (e.g., having
blue eyes, or a given score on a
quantitative measure of rhythm
abilities).
Polygenic pleiotropy:
When the same sets of genetic
variants make contributions to two or
more distinct complex traits, pointing
to shared genetic architecture.
Endophenotype:
A specific biomarker associated with
an illness/trait in the population that
is heritable and exists whether or not
the illness/trait is active (Gottesman
& Shields, 1972).
Neurobiology of Language
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Gene expression:
The process by which DNA
information is used to assemble RNA
molecules and proteins through
transcription and translation.
Heritability:
An estimate of the proportion of trait
variance accounted for by variation
at the genetic level in a particular
population, under specific
environmental circumstances.
functionally involved in the relationship between a genotype and a phenotype of interest
(Gottesman & Gould, 2003). For example, measures of basal ganglia neuroanatomy or of pre-
frontal cortex function could be explored as neural endophenotypes mediating the effects of
genetic variants on musicality and language traits. Discovery of gene expression patterns in
key brain regions and developmental processes of interest (e.g., neocortical development: Miller
et al., 2014) could help directly map associations among genetic variants, neural endopheno-
types (including gene expression and regulation patterns in the brain), and correlated musicality
and language traits (see Kong et al., 2020, for an example of identifying convergence between
functional magnetic resonance imaging (fMRI) based language circuitry and gene expression
patterns). Like musicality and language traits themselves, neural endophenotypes (including
gene expression patterns) are shaped by interacting genetic and environmental influences.
We therefore argue that future research on musicality-language associations should aim to
incorporate genetic studies of both musicality and language phenotypes as well as associated
neural endophenotypes. We suggest ways in which this can be achieved by leveraging existing
data and collaborative efforts. Specifically, genetic/genomic data can be used to (a) understand
the shared genetic architecture of musicality and language; (b) disentangle the interplay
between genes and environment; and (c) inform predictive models and intervention efforts
in the context of language-related disorders, using data available on music-related traits.
Structure and Scope of the Current Work
The structure of the current work is as follows: First, we provide an overview of heritability
estimates for musicality and speech-language phenotypes reported by twin and family-based
studies, and discuss how these estimates can inform our understanding of correlated individual
differences in musical and language abilities. Second, we propose a novel framework for
understanding links between musical and language abilities, with a focus on shared genetic
architecture and mediating neural endophenotypes. Third, we provide a specific overview of
genetics approaches that can be integrated into current investigations of musicality-language
links. Fourth, we outline specific genetic predictions that will help to systematically test our
framework, and set an agenda for how the field can pursue these directions. Last, we provide
supporting evidence for our framework, with bibliometric analyses, and a synthesis of behav-
ioral studies examining associations between musicality-language traits. We also discuss
adjacent evidence from the neural correlates of individual differences in musical or language
abilities, where these abilities are associated. The literature synthesis is organized by domain
of language ability: We discuss research on musicality and speech skills, reading skills, and
grammar skills respectively.
We constrain the scope of the current work in a few important ways. One, our discussion
and framework focus on individual differences in stable traits. Therefore, studies that investi-
gate priming effects of music on language; language in musically trained and untrained
individuals; similarities in neural processing of musical and linguistic stimuli; and effects of
musical interventions for language, are considered outside the scope of the current work, while
certainly remaining relevant for understanding relationships between musicality and language.
Similarly, we do not discuss lines of research focused on musical state-driven modulation of
neurobiology (e.g., changes in gene expression after listening to music: Nair et al., 2021),
while these studies are informative for different purposes. Two, our framework focuses mainly
on the potentially supportive role of musical abilities for language across the lifespan. We also
acknowledge that the reverse remains possible, i.e., that language abilities support musical
skills (e.g., Roncaglia-Denissen et al.’s, 2016, finding that L2 learners show enhanced musical
Neurobiology of Language
618
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
rhythm). Understanding biological relationships between musicality and language traits (e.g.,
shared genetic architecture of these skills) will be useful for understanding both musical and
language-related outcomes. Three, while we review evidence of the neural correlates of indi-
vidual differences in musicality-language skills, we generally do not focus on the many impor-
tant studies that experimentally manipulate processing of musical and linguistic stimuli and
report primarily group means in one condition versus another; such designs are not necessarily
informative about individual differences in stable musicality and language traits (see Hedge
et al., 2018, for extended discussion about trade-offs between stable individual metrics and
reliable experimental effects).
HERITABILITY OF SPEECH-LANGUAGE AND MUSICALITY PHENOTYPES
Thus far, the genetic architecture of musicality and language has been primarily studied sep-
arately, through twin and family-based methods that inform us about the heritability of specific
traits. For example, twin studies show that many speech-language traits are moderately heri-
table, as summarized in Table 1. Similarly, twin and family-based studies show that musical
abilities (e.g., pitch and rhythm sensitivity) have a significant genetic component (Drayna
et al., 2001; Seesjärvi et al., 2016; Ullén et al., 2014), as does musical engagement, including
accomplishment and training (Hambrick & Tucker-Drob, 2014), as summarized in Table 2.
Emerging evidence also finds that both musical aptitude and engagement are genetically asso-
ciated with language-related traits such as verbal ability (Gustavson et al., 2021; Wesseldijk
et al., 2021).
It is important to distinguish this twin and family-based evidence from other types of genetic
investigations that focus on altered music and/or language skills in the context of genomic
abnormalities affecting a single gene or chromosomal location, such as in individuals with
Williams, Prader-Willi, or Angelman syndromes, or in families with FOXP2 disruptions. While
ongoing work on these disorders (e.g., Williams syndrome: Kasdan, Gordon, et al., 2022;
Thakur et al., 2018; Prader-Willi and Angelman syndrome: Kotler et al., 2019; Mehr et al.,
2017; FOXP2 disruptions: Alcock et al., 2000; Fisher & Scharff, 2009) can help to reveal
causal pathways linked to rare genetic variation at particular loci, our goal here is to outline
steps for understanding the potentially overlapping genetic architecture between musicality
and language traits as it relates to common genetic variation in the broader population, which
will likely require a genome-wide perspective.
Genome-wide investigations can help clarify mechanisms by which shared genetic archi-
tecture between musicality and language-related traits may result in associations observed at
the phenotypic level. Further, increased access to high-throughput genetic data collected in
large cohorts, coupled with meta-analytical efforts, has greatly improved the potential for
understanding the overlapping biology of distinct complex traits related to musicality and
speech-language abilities. For example, current statistical genetics methods make it possible
to test for genetic correlations between traits even when genetic samples and phenotypic mea-
sures have been collected in separate samples (Bulik-Sullivan et al., 2015). Genetic correla-
tions inform us about the degree of phenotypic covariance in two traits explained by shared
genetic variation (Watanabe et al., 2019; Weissbrod et al., 2018).
THE MAPLE FRAMEWORK FOR CLARIFYING BIOLOGICAL RELATIONSHIPS BETWEEN
MUSICALITY AND LANGUAGE ACROSS THE LIFESPAN
The earliest stages of spoken language development (e.g., in infancy) involve learning to seg-
ment speech, recognizing vowel and consonant contrasts, and parsing rhythmic patterns in
Neurobiology of Language
619
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
N
e
u
r
o
b
o
o
g
y
i
l
o
f
L
a
n
g
u
a
g
e
6
2
0
Table 1. Heritability estimates of speech-language and reading-related phenotypes.
Language domain
Speech/Language
Construct definition
Measure description
Spoken Language /
Includes measures of word object
Oral Skills
mapping, semantics (e.g., finding
relationships between words), and
morphosyntactic skills.
Relational vocabulary (from Test of
Language Development, primary,
3rd ed.: TOLD-P:3)
Heritability
estimates
Citations
0.40–0.70
Rice et al., 2018
Verbal memory: the ability to recall
what has been heard or read.
Story memory; sentence memory;
0.48–0.87
Samuelsson et al., 2007
nonword repetition.
(3 samples)
Semantics: includes measures of
word object mapping, and
understanding conceptual
relationships between words.
Grammatic understanding task
(from TOLD-P:3).
0.10–0.70
Rice et al., 2018
Oral skills composite
Composite performance on tasks
0.34
Andreola et al., 2021
assessing expressive and receptive
vocabulary, naming abilities, and
oral language skills.
(meta-analysis of 10 studies)
Vocabulary size: receptive and
Receptive vocabulary skills; picture
0.18–0.67
expressive vocabulary size, tested
directly or via parent-report
checklist (in young children).
vocabulary comprehension;
parent-reported vocabulary
production.
Babajani-Feremi, 2017;
Dale et al., 2018;
Rice et al., 2018;
Samuelsson et al., 2007
Impaired language achievement
(in absence of other developmental
or sensory impairments).
Language impairments with speech-
language pathologist (SLP) referral.
Verbal ability
Verbal fluency: includes general
fluency and semantic-specific
fluency components.
Low performance on receptive
0.45
Tomblin & Buckwalter, 1998
language (e.g., vocabulary and
grammar), expressive language
(e.g., vocabulary, morphosyntax
and grammar).
Low performance (relative to sample
mean) on a language battery, in
addition to having received speech
language therapy or other speech
pathologist services by age 7 yrs.
Phonemic fluency (how many words
can you write beginning with one
letter and ending with another in
the time limit); semantic fluency
(list as many names of things in a
category).
0.96–0.97
Bishop & Hayiou-Thomas, 2008
0.65–0.80
Gustavson et al., 2019
U
n
d
e
r
s
t
a
n
d
i
n
g
m
u
s
i
c
a
l
i
t
y
–
l
a
n
g
u
a
g
e
l
i
n
k
s
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Verbal ability: tests of overall verbal
knowledge, comprehension, and
fluency, tested through conceptual
tests.
Language comprehension
Odd one out tests; synonym tests;
0.60
vocabulary tests; or verbal
fluency tests.
Gustavson et al., 2021;
Wesseldijk et al., 2021
Story comprehension (the ability to
listen to a story or narrative and
accurately answer questions about
its content, i.e., comprehend it).
0.32
Babajani-Feremi, 2017
Speech production
Speech articulation: ability to
Goldman-Fristoe Test of Articulation.
articulate real consonant sounds in
single words and conversational
speech, both spontaneously and
through imitation.
Speech abilities
Low performance (relative to sample
mean) on a speech composite
consisting of a speech articulation
task and a nonword repetition task
(also taps phonological working
memory).
0.25–0.60
Rice et al., 2018;
Stein et al., 2011
0.56
Bishop & Hayiou-Thomas,
2008; Hayiou-Thomas, 2008
Reading
Accuracy and speed
Reading composite
of reading
Letter-word knowledge:
the recognition and identification
of how letters form words (called
“general reading construct”
in paper).
Phonological decoding: the ability to
break written words into sounds or
syllables based on the phonemic
representations of your language to
support word recognition.
Reading comprehension: the ability
to understand and process written
text.
Combination of letter-word
knowledge, phonological
decoding, and reading
comprehension phenotypes.
Oral reading recognition;
letter/word identification.
0.66
Andreola et al., 2021
(meta-analysis of 48 studies)
0.56–0.62
Andreola et al., 2021
(meta-analysis of 32 studies);
Babajani-Feremi, 2017
Irregular word decoding;
phoneme decoding.
0.68
Andreola et al., 2021
(meta-analysis of 13 studies)
Text/story comprehension;
reading achievement.
0.68
Andreola et al., 2021
(meta-analysis of 32 studies)
Print knowledge
Knowledge about the rules of print.
Left-to-right reading; letter
0.26
Samuelsson et al., 2007
recognition, environmental print
exposure; concepts about print.
N
e
u
r
o
b
o
o
g
y
i
l
o
f
L
a
n
g
u
a
g
e
6
2
1
U
n
d
e
r
s
t
a
n
d
i
n
g
m
u
s
i
c
a
l
i
t
y
–
l
a
n
g
u
a
g
e
l
i
n
k
s
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
N
e
u
r
o
b
o
o
g
y
l
i
o
f
L
a
n
g
u
a
g
e
6
2
2
Language domain
Phonological awareness
Construct definition
Processing speech and
phonological/lexical retrieval.
Table 1.
(continued )
Measure description
Comprehensive Test of Phonological
Processing (CTOPP); syllable and
phoneme blending; word elision;
syllable and phoneme elision;
rhyme recognition; phoneme
isolation; phonemic deletion;
rhyme recognition; phonemic
segmentation.
Heritability
estimates
0.46–0.64
Citations
Andreola et al., 2021
(meta-analysis of 13 studies);
Samuelsson et al., 2007
(3 samples)
Literacy
Composite performance on
Reading fluency (reading simple
0.68–0.77
Kovas et al., 2013
standardized reading, writing,
speaking, and listening tasks,
assessed through teacher
evaluations, and testing via
telephone and internet.
sentences); literal comprehension
of sentences; analysis of written
text.
Spelling: the ability to form words
with the correct order of letters.
Orthographic decoding,
regular/irregular spelling.
Conversational language
productivity
Length and complexity of children’s
utterances in conversation, usually
computed as the mean number of
morphemes in utterances sampled.
0.80–0.91
Andreola et al., 2021
(meta-analysis of 15 studies);
Lewis et al., 2018
0.17–0.61
Dale et al., 2018;
DeThorne et al., 2012
Grammar
Morphosyntactic
Expressive and receptive skills
abilities
relating to how words are formed
and how the order of words results
in understandable phrases and
clauses. May include tasks where
whole sentences need to be
accurately recalled, tapping
morphosyntax skills.
Low performance on
grammatical/morphosyntactic
tests, or indicated by tests designed
to screen or measure grammatical
impairment.
Sentence complexity; grammatical
property of finiteness marking;
grammatical knowledge;
productive morphology.
0.36–0.92
Bishop et al., 2006;
Dale et al., 2000;
Rice et al., 2018;
Samuelsson et al., 2007
Grammatical property of finiteness
marking; third-person singular;
past tense obligatory use; receptive
grammar.
0.31–1.0
Bishop et al., 1995;
Dale et al., 2018;
Rice et al., 2018
Note. Table shows moderate heritability of speech/language and reading phenotypes. Each heritability estimate provided indicates the percentage of overall variance in the trait that is
genetically influenced, as computed by twin and family-based studies cited here. Broad definitions for constructs are provided, and corresponding phenotypic measures corresponding to
each speech-language and reading construct of interest are outlined. TOLD-P:3: Test of Language Development, Primary, 3rd ed. (Newcomer & Hammill, 1997). Goldman-Fristoe Test of
Articulation, 2nd ed. (Goldman & Fristoe, 2000). CTOPP: Comprehensive Test of Phonological Processing (Wagner et al., 1999).
U
n
d
e
r
s
t
a
n
d
i
n
g
m
u
s
i
c
a
l
i
t
y
–
l
a
n
g
u
a
g
e
l
i
n
k
s
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Table 2. Heritability estimates of musicality phenotypes.
Construct
Rhythm perception
Melody perception
Pitch perception
Measure description
Discriminating between rhythms; determining
whether a rhythm is the same as or different
than a reference rhythm.
Discriminating between melodies; identifying
which note in a given melody differs from a
reference melody.
Discriminating between pitches; identifying
whether a given pitch is higher or lower
than a reference pitch.
Rhythm production
Isochronous motor timing (self-paced).
Music practice
Duration and frequency of practicing a musical
instrument or singing.
Heritability
estimates
0.5
0.58–0.59
0.4–0.8
0.34
0.41–0.69
Citations
Mosing et al., 2016;
Ullén et al., 2014
Seesjärvi et al., 2016;
Ullén et al., 2014
Drayna et al., 2001;
Ullén et al., 2014
Mosing et al., 2016
Butkovic et al., 2015;
Mosing et al., 2014
Music flow
Degree of proneness to experiencing psychological
0.4
Butkovic et al., 2015
Musical talent
Musical aptitude
flow while engaging in musical activities.
Self-reported exceptional musical talent (singing
on playing instruments) as measured by a
self-reported talent inventory.
Self-reports of average or above average musical
aptitude (singing or playing instruments) on a
self-reported talent inventory.
For heritability of specific objectively-measured
music aptitude traits (rhythm perception, melody
perception, pitch perception), see estimates from
Ullén et al. (2014), reported above.
0.26–0.92
Hambrick &
Tucker-Drob, 2014;
Vinkhuyzen et al., 2009
0.30–0.66
Vinkhuyzen et al., 2009
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
Musical instrument
Self-reported interest, instruction, and talent with
engagement
musical instruments.
Singing
Self-reported interest, instruction, and talent with
vocal music (singing).
0.78
0.43
Gustavson et al., 2021
Gustavson et al., 2021
Note. Table shows moderate heritability of musicality phenotypes. Each heritability estimate provided indicates the percentage of overall variance in the trait
that is genetically influenced, as computed by twin and family-based studies cited here. Broad definitions for constructs are provided, and corresponding
phenotypic measures corresponding to each musicality construct of interest are outlined.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
one’s native language(s), which all highly overlap with musical listening skills (Brandt et al.,
2012; c.f. signed language development). As Brandt et al. (2012) argue, the listening skills of
very young developing humans (fetuses, newborns, and infants included) are attuned to fea-
tures of spoken language that are also crucial features of musical listening (e.g., pitch, timbre,
rhythm, dynamic stress), which form the basis of effective speech processing and lay the foun-
dation for language learning over the course of development (Sanchez-Alonso & Aslin, 2022).
The important role that processing these cues plays in language development is further
evidenced by much greater than chance prevalence of co-occurring cases of musical and
language impairments in children (Ladányi et al., 2020; Peretz & Vuvan, 2017). In particular,
children with disorders of language (e.g., developmental language disorder: DLD) and reading
(e.g., dyslexia) have higher prevalence of rhythm and melody impairments than age-matched
Neurobiology of Language
623
Understanding musicality-language links
typically developing peers, providing evidence of biological relationships between these def-
icits (Ladányi et al., 2020). As reviewed in the current article, bountiful supporting evidence
shows correlations between musicality and language skills in typically developing children
and adults, including associations for multiple levels and domains of language processing
(speech, reading, vocabulary, and grammar).
These avenues of research have clinical significance in addition to basic science value. Just
as musicality might be leveraged for early identification of risk for language-related disorders,
musical abilities can help account for resilience to speech-language problems in a variety of
common conditions of older adulthood (e.g., dementia, Alzheimer’s disease, age-related hear-
ing loss). For example, shared musicality and language-related genetic architectures could also
be implicated in inner ear function, neuroplasticity, and other adaptive mechanisms in the
context of brain damage, or natural brain aging, such that those with genetic predispositions
for enhanced musical abilities might also have genetic resilience to the magnitude or rate of
neurodegeneration associated with language-related problems. Figure 1 illustrates the relation-
ship between individual differences in musicality and language abilities, as we broadly envi-
sion it playing out across the lifespan.
Based on the hypothesized relationship between musicality and language abilities across
the lifespan (Figure 1), we propose the Musical Abilities, Pleiotropy, Language, and Environ-
ment (MAPLE) framework for clarifying biological mechanisms of this lifespan view of
musicality-language relationships (Figure 2). Briefly, the MAPLE framework proposes that
observed phenotypic associations between musicality and language-related traits are partly
driven by shared genetic architecture, and that these musicality-language links are further rein-
forced by gene-environment interactions and neuroplasticity.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Schematic illustration of the proposed role of musicality in language across the lifespan,
showing the curvilinear relationship of language abilities across ages, moderated by relatively
higher and lower levels of overall musicality (e.g., musical abilities, engagement, and environ-
ments). We propose that individuals with relatively higher musicality on a spectrum of typical
individual differences (solid curved line) will have enhanced language abilities and/or steeper
developmental trajectories in early stages of life compared to those with relatively lower musicality,
or impairments in musical abilities (dashed curved line). Similarly, we propose that in adulthood
and in the context of aging, those with relatively higher musicality will experience extended main-
tenance of peak performance, and slower decline in speech-language function later in life (e.g.,
efficiency of their speech perception in noise).
Neurobiology of Language
624
Understanding musicality-language links
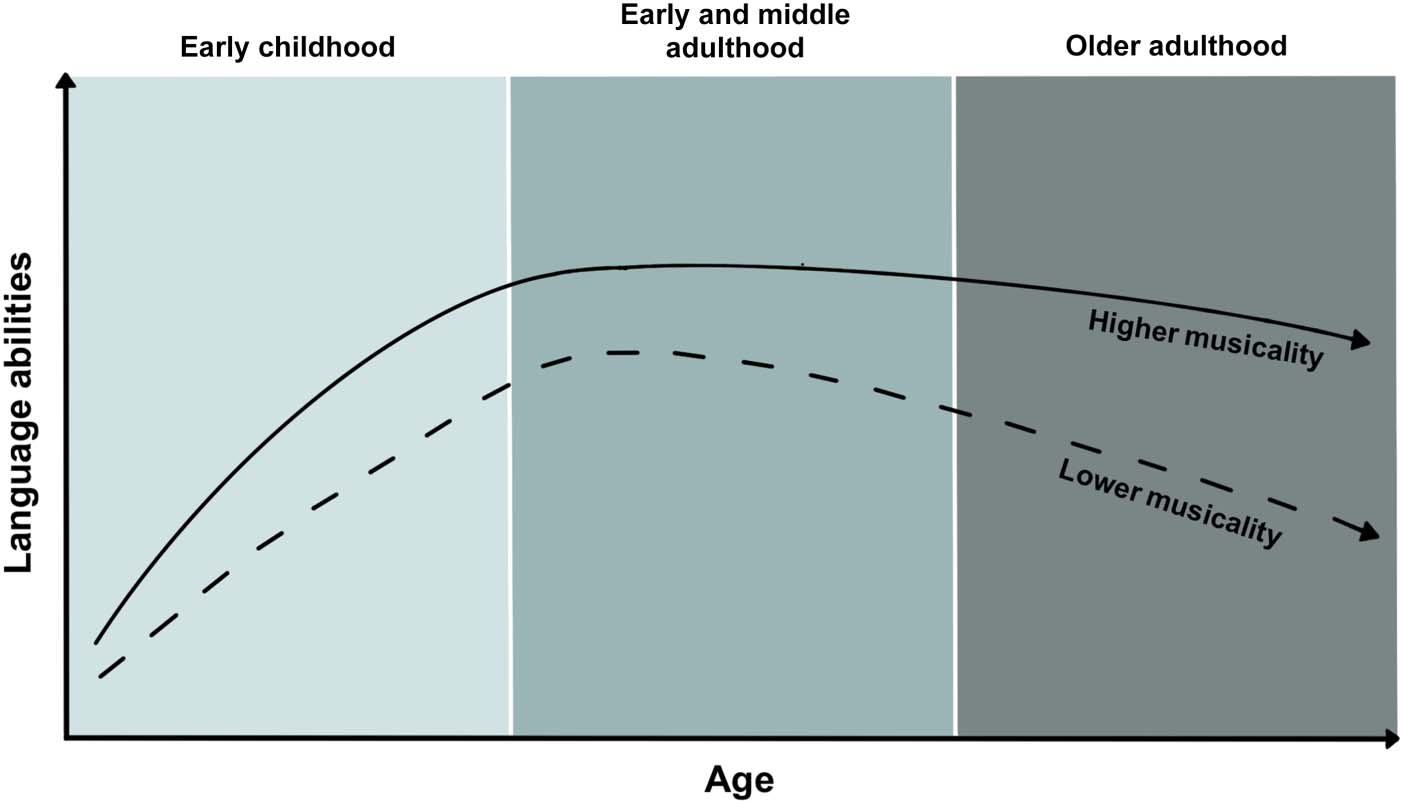
Figure 2. The MAPLE framework. Genetic pleiotropy is illustrated by overlapping maple leaves (left) signifying shared polygenic architecture
influencing musicality and language traits. Shared genes influencing musicality and language are thought to exert influence on cascading
biological processing including development, structure, and functioning of the brain, and nervous system functions relevant for musical
and language traits such as auditory processing, and sensorimotor coordination (center top). Alongside genetic influences on these neural
endophenotypes, polygenic influences on musicality and language are also thought to influence key environmental factors such as social
relationships, and home musical and language environments (center bottom). Thus, genetic pleiotropy (left) is thought to be a root biological
mechanism underlying the observable phenotypic associations between musical and language traits widespread in the literature (right). The
MAPLE framework’s predictions can be tested by examining (a) polygenic architecture underlying musicality and language traits (left), (b)
polygenic architecture and heritability of relevant neural endophenotypes, and of home and school musical and linguistic environments (cen-
ter top and bottom, respectively), and (c) phenotypic variation in musical and language traits in broad populations, and cross-trait associations
(right). Arrows illustrate the directions of influence between genes, neural endophenotypes, environment, and behavior.
Central to the MAPLE framework is the idea that genetic architectures underlying musicality
and language traits partly overlap through the mechanism of genetic pleiotropy. Since both
musical and language traits are complex and polygenic (i.e., affected by multiple genes in
concert, rather than individual genes), we henceforth refer to this overlap in polygenic archi-
tectures as polygenic pleiotropy. An emphasis on shared genetic contributions allows us to
leverage the known variability and heritability of musical skills (Niarchou et al., 2022; Ullén
et al., 2014) and speech-language skills (Deriziotis & Fisher, 2017; Evans et al., 2015; Graham
& Fisher, 2013). Further, since there is evidence of neural overlap in music and language pro-
cessing (Atherton et al., 2018; Patel, 2012; Perani, 2012; Peretz et al., 2015), genetic variants
that influence neurobiological structure, function, and development are also expected to be
statistically overrepresented (relative to genes influencing other non-nervous system function)
among loci common to musical and language traits (see Fisher & Vernes, 2015, for a detailed
discussion of how genes affect neuronal circuits relevant for language). The MAPLE framework
therefore emphasizes the importance of neural endophenotypes that mediate the relationship
between genetic architecture and music/language phenotypes.
Alongside their influence on musical and language abilities, genes also exert influence on
music and language-related environments—including environments shaped by oneself
through increased or decreased engagement. For example, home environments during early
development partly reflect the genetic predispositions of parents, through niche-picking, as in
when musically talented parents choose to pursue musical training for their children. Since
children inherit their parents’ genetic makeup, they too may niche-pick certain experiences
Neurobiology of Language
625
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Affordances:
Properties of the environment or
neurocognitive makeup (e.g., music
perception skills) that allow
individuals to carry out actions
(e.g., frequent music listening).
compatible with their inherited aptitude and preferences (Hart et al., 2021). The effects of
parents’ genes, and the environments that are created as a result, are amplified when
children inherit stronger genetic predispositions for music and/or language traits through
the process of assortative mating (Burley, 1983), as in when individuals seek out mates with
similar levels of musicality. In addition to the genetic influences on the environment,
biological/neurocognitive affordances can also directly affect the experiences sought and
enjoyed by individuals.
The MAPLE framework is consistent with existing evidence of musical experience-
dependent neuroplasticity across the lifespan (Merrett et al., 2013; Münte et al., 2002; Schlaug
et al., 2009). Of interest here, musical training is thought to be strongly associated with higher
performance on measures of auditory skills (Kraus & Chandrasekaran, 2010), language skills
(Gordon, Fehd, & McCandliss, 2015; Tierney & Kraus, 2013; White et al., 2013), and execu-
tive functioning (George & Coch, 2011; Moreno et al., 2011). Moreover, the framework allows
us to account for genetic influences on the brain and nervous system functions, to explain how
biological mechanisms (e.g., gene regulation; neuroplasticity) interact to give rise to covari-
ance between musical and language abilities across the lifespan. The MAPLE framework com-
plements and extends existing frameworks linking musical and speech-language abilities, such
as the atypical rhythm risk hypothesis (Ladányi et al., 2020). While the atypical rhythm risk
hypothesis is primarily concerned with genetic and neural links between atypical rhythm
and risk for speech-language disorders, the MAPLE framework considers individual differences
in typical speech-language outcomes at the population-level, highlighting the role of musical
abilities and related biology. The hypotheses and predictions of the MAPLE framework are
compatible with those laid out by the atypical rhythm risk hypothesis.
The approach described here is akin to current frameworks in the field of psychiatric genet-
ics. Researchers have highlighted shared genetic influences that underlie comorbidities among
disorders both within disorders of a similar type, such as genetic overlap among mood and
anxiety disorders; and across broad categories of psychopathology, such as genetic overlap
across mood/anxiety disorders and thought disorders (Kotov et al., 2017; Selzam et al.,
2018). While these conditions often present as clinically distinct, by interrogating common
genetic influences that confer risk for psychopathology, researchers have been able to discover
shared characteristics of traditionally separate conditions. For example, overlapping genetic
architecture between psychiatric disorders can exert influences on underlying neurodevelop-
mental mechanisms that transcend diagnostic categories (Lee et al., 2019). Further, unraveling
genetic pleiotropy between psychiatric disorders can improve nosology through characteriz-
ing relevant domains (e.g., sensorimotor), constructs (e.g., action planning), and units of anal-
ysis (e.g., genes and neural circuitry) underlying different disorders, as outlined by the research
domains criteria (RDoC) framework (Cuthbert, 2014). Neurogenetic discoveries such as these
can lead to identifying novel neurobiological risk or resilience factors for various psychiatric
conditions. Similarly, this approach can be applied to musicality and language traits to help us
better characterize the neurobiological factors that give rise to individual differences in both
sets of skills and abilities, and covariation between them.
INTEGRATING GENETICS APPROACHES INTO MUSICALITY-LANGUAGE RESEARCH
Genetic designs can inform the associations between musicality and language traits in several
ways. First, the classic twin design can decompose phenotypic correlations into genetic,
shared environment, and nonshared environmental correlations. The presence of a strong
genetic correlation would indicate that a common set of genetic influences gives rise to
Neurobiology of Language
626
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
individual differences in both sets of traits. In contrast, the presence of shared or nonshared
environmental correlations could indicate that associations are driven by environmental expo-
sures, potentially including causal relationships, though there are other ways of testing for
causal associations in the context of a twin model (Heath et al., 1993). Twin and family studies
can also test for the presence of gene-environment correlations and gene-by-environment
interactions, which appear highly relevant for musical traits (Hambrick & Tucker-Drob,
2014; Wesseldijk et al., 2019). These studies have demonstrated that individual differences
in music achievement are more pronounced in those who engage in practice or had musically
enriched childhood environments. Similar work could examine whether individual differences
in language abilities are more pronounced for individuals with more musical experiences or
stronger musical abilities (for guidelines on statistical models that can be tested, see Purcell,
2002; van der Sluis et al., 2012).
One example of the utilization of twin and family studies to examine the associations
between musicality and language comes from Gustavson et al. (2021), who find that self-
reported musical instrument engagement at age 12 predicts verbal ability at age 16, controlling
for IQ, and that this relationship is likely explained by shared genetic influences. Further,
Wesseldijk et al. (2021) report recent findings in twins that demonstrate that phenotypic asso-
ciations between musical aptitude and vocabulary skills are partially attributable to shared
family influences. These methods allow for an estimate of the influences of genetics on musi-
cal and language skills in the populations being studied. Further, while the effects of music
environments have traditionally been explained via models of neuroplasticity, behavioral
genetics findings allow us to reimagine what musical environments can tell us.
Beyond the classic twin design, large-scale genomic data can also be useful to uncover
musicality-language links and inform us about the nature of their shared biology. Large-scale
genome-wide association studies (GWAS) for example, involve scanning markers across the
complete genome of many people to find common genetic variations associated with a par-
ticular trait. Consistent with the polygenic nature of complex human traits, each of these
common variations is likely to have only a small effect size, necessitating large sample sizes
(thousands of participants) to reliably detect associations. By identifying these genetic asso-
ciations with the trait of interest, we can better understand the genetic architecture of the
trait. Further, polygenic scores (PGSs; Krapohl et al., 2018) applied to individuals’ genomic
data, with weights at specific alleles derived from the results of large-scale GWAS, can be
used to develop better strategies for detecting which individuals might have genetic predis-
positions that explain variance in another phenotype (e.g., risk for dyslexia or DLD based on
relatively lower polygenic scores for a language-related trait).
While GWAS efforts for cognitive traits have largely focused on general cognitive ability
(Davies et al., 2018; Savage et al., 2018), such studies have more recently begun to shed light
on the molecular bases of speech, language, and reading (Eising et al., 2022; Graham & Fisher,
2013; see Deriziotis & Fisher, 2017, for extended discussion of language GWASs), including in
the context of developmental dyslexia (Doust et al., 2022; Gialluisi et al., 2021). The first large
GWAS of a musical trait, beat synchronization (Niarchou et al., 2022), has demonstrated highly
polygenic architecture of this rhythm-related trait. As GWAS of musical and language-related
traits become more widely available, genetic correlations (Bulik-Sullivan et al., 2015) between
these traits can be estimated even when the samples used for GWAS do not overlap.
Relatedly, we can utilize the result of extensive mapping of regions of the human genome to
biological functions, included those involved in neural development and other processes. For
example, Niarchou et al. (2022) found that the genetic architecture of beat synchronization is
Genome-wide association study
(GWAS):
A systematic, comprehensive screen
of genetic variants at many different
sites across all chromosomes of a
genome, testing each variant for
association with a trait of interest.
Polygenic scores (or Polygenic Risk
Scores):
A number reflect a person’s
estimated genetic predisposition for a
trait, by combining the estimated
effects of genetic variants at different
loci across the genome.
Neurobiology of Language
627
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Enriched:
Overrepresentation of a biological
function in the genetic architecture of
a trait, compared to chance.
enriched for brain specific regulatory regions of the genome, in both adult and fetal brain tis-
sues. In this way, statistically integrating neurogenomic data will allow us to map the genetic
architecture of musicality and language traits to neurodevelopmental and neurofunctional
endophenotypes. Application of such methods and the subsequent findings would represent
a major advance towards the goal of testing the MAPLE framework’s predictions about shared
genetic architecture between musicality and language (see the section Predictions Derived
from the Maple Framework).
Moreover, researchers can draw on the potential of genomic data by using available
polygenic scores for phenotypes of interest to examine associations with traits in target sam-
ples for whom genotyping is also available (provided the samples are of adequate size for cap-
turing polygenic signal; usually comprising tens of thousands to millions of individuals for
computing polygenic scores from discovery GWASs, and hundreds to thousands of individuals
for applying polygenic scores to target samples). For example, given cohorts with language
measures, we can examine to what extent individual differences in genetic predispositions
for musical rhythm ability (e.g., using polygenic scores computed from a large discovery
GWAS of musical rhythm) are predictive of performance on reading-related tasks (in a more
modest target sample). This approach can be incorporated into any of the behavioral/neural
designs commonly utilized in the literature on musicality-language links. Polygenic scores can
also be examined as moderators, for example to test whether individuals with high genetic risk
for language disorders show stronger language outcomes when exposed to musical experi-
ences at an early age. Given the broad and intertwined (pleiotropic) influence of genes on
(neuro)biology, genomic approaches can reveal shared biological mechanisms of musicality
and language traits that converge across various constructs, tasks, and stimuli common in the
literature (detailed in the Literature Review section).
Computational genomic approaches can also be used to investigate the evolutionary basis of
complex traits along lineages that led to present-day humans as well as comparatively across
species. For example, Gordon et al. (2021) examined overlapping genetic architecture between
beat synchronization traits in humans, and complex vocal learning traits in songbirds. That
study provided genetic evidence for Patel’s (2021) hypothesis of convergent evolution between
songbird vocal learning and human beat perception and synchronization, which posits that
musical rhythm processing piggybacks on neural circuitry evolved for complex vocal learning.
Similarly, computational approaches can be used to shed light on shared or separate evolution-
ary histories of traits of interest (Colbran et al., 2019; Tilot et al., 2021). As additional sets of
GWAS data become publicly available, the information can be used to validate and extend
results from phenotypic factor analyses, through genomic structural equation modelling
techniques that make it possible to analyze the joint genetic architecture of complex traits
(e.g., Gustavson et al., 2020) as well as the genetic loci implicated in divergence between traits
(Grotzinger et al., 2019). These advances can more directly inform our understanding of poly-
genic pleiotropy between musical and language traits proposed by the MAPLE framework.
Future GWAS efforts should target some of the behavioral and neural endophenotypes
that have been highlighted by prior studies of musicality-language links, as reviewed later in
this article. Given efforts currently underway to map the genetic architecture of neural traits
(e.g., work by the ENIGMA consortium: Smit et al., 2021) there is a timely opportunity to lever-
age existing resources to further our understanding of musicality-language links. Large-scale
data sets can now be creatively leveraged to map genetic variants to neural correlates of musi-
cality and language traits (e.g., Mekki et al., 2022, recently mapped the genetic architecture of
resting-state functional connectivity in brain regions classically associated with language
function). To identify neural endophenotypes as proposed by the MAPLE framework, future
Neurobiology of Language
628
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
research can systematically draw on existing neural data that is linked with genetic and
behavioral data, for example in biobank initiatives such as the UK Biobank (Sudlow et al.,
2015). While ideally, musical and language abilities will eventually be deeply phenotyped
in study participants in large-scale population health studies (e.g., All of Us: Kaiser, 2016), in
the meantime we can also leverage neural maps based on meta-analyses of music and language
processing or abilities (e.g., Kasdan, Burgess, et al., 2022), to guide neurogenetic investigations.
Recent advances in data-driven approaches such as data mining algorithms and machine learn-
ing models have the potential to further accelerate progress in this research area, by automating the
identification of cases with particular symptomatology in electronic heath records (e.g., automated
phenotyping tool for DLD cases, APT-DLD: Walters et al., 2020; phenome risk classifier for stut-
tering: Pruett et al., 2021), the extraction of neural features from magnetic resonance imaging
(MRI) data (e.g., toolbox for the automatic segmentation of Heschl’s gyrus, TASH toolbox: Dalboni
da Rocha et al., 2020), the extraction of features in the genetic architecture of a trait (e.g., GWAS
loci prioritization: Nicholls et al., 2020), and the integration of neuroimaging and genomic
data to predict phenotypic outcomes (e.g., Shen & Thompson, 2020).
Last, longitudinal studies investigating how early musical skills may predict aspects of lan-
guage development can additionally incorporate genetic data (i.e., via polygenic models of
musical abilities and engagement, and of brain function) to begin to chart gene-environment
interactions during development, accounting for predisposition and neural plasticity (Zuk
& Gaab, 2018).
PREDICTIONS DERIVED FROM THE MAPLE FRAMEWORK
The MAPLE framework can be used to guide many kinds of research questions, which can be
answered by utilizing genetic and genomic approaches (Box 1, numbered). For each research
question, specific predictions can be made about anticipated outcomes (Box 1, bulleted).
Testing the MAPLE Framework: An Agenda for Musicality-Language Research
Strategic directions for testing the MAPLE framework could be supported by the following
approaches:
1)
Integrating genetic approaches into music and language science:
(cid:129) Add musicality variables to genetically informative samples across a range of cogni-
tive, neurobiological, and speech-language phenotypes.
(cid:129) Investigate pleiotropy with cross-trait methods such as genetic correlations and poly-
genic score analysis. Focus on areas where there are strong phenotypic links, such as
those found in the literature synthesis here, to guide investigations of polygenic
pleiotropy.
(cid:129) Explore neural endophenotypes in relation to genes and behavior in large samples
and across traits measured in separate samples, using state-of-the-art methods and by
leveraging big data.
2)
Integrating music science with behavioral and neural approaches to language:
(cid:129) Include musical abilities in models examining individual differences in language
skills, to account for covariance.
(cid:129) Capture rich variation in language experiences, backgrounds, and engagement
alongside variation in musical traits, going beyond language outcomes and skills.
Neurobiology of Language
629
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Box 1. Research questions and predictions about musicality-language links.
1) Do musicality and language share genetic architecture? If so, which musical traits are genetically associated with which
language traits?
(cid:129) Analyses performed on the results of well-powered music and language GWAS will show significant genetic correlations
between musical and language traits that are known to be phenotypically correlated (e.g., using linkage disequilibrium
score (LDSC) regression methods: Bulik-Sullivan et al., 2015) across various constructs and stimulus types.
2) What can be predicted about phenotypic variability in language traits based on genetic variation associated with musicality,
and vice versa? How are these relationships mediated by genetic predictor models of brain activity and structure?
(cid:129) Polygenic scores (PGSs) for musical traits derived from one sample will predict behavioral speech/language/reading phe-
notypes in an independent sample, and vice versa.
(cid:129) These PGS associations will be mediated by PGSs for neural endophenotypes already shown to underlie the relation
between specific musicality and language traits.
3) Which neurobiological functions are the genes associated with musicality and language traits enriched for? Do they impli-
cate systems already known to be involved in musical and language abilities?
(cid:129) Enrichment analyses (e.g., Subramanian et al., 2005) performed on the results of well-powered GWAS of musicality
phenotypes will show that the genetic architecture of musicality is enriched for genes involved in neural endophenotypes
of language/communication. These neural endophenotypes may, for example, include the following:
(1) Hierarchical processing, music and language processing, or auditory processing: superior temporal gyrus, Heschl’s
gyrus volume; inferior frontal gyrus (Broca’s area).
(2) Temporal brain dynamics: motor system functionality (e.g., for circuits involving cerebellum, and/or basal ganglia);
resting state neural oscillations; oscillatory mechanisms of music or language perception.
(3) General cognitive abilities across the lifespan: white matter connectivity; surface area and thickness of cortical
regions; subcortical volumes; cerebral volume (see Bethlehem et al., 2022).
(cid:129) Similarly, enrichment analyses performed on the results of well-powered language-related GWAS will show that the
genetic architecture of language-related traits is enriched for genes associated with neural endophenotypes involved
in music perception and cognition.
(cid:129) Functional genomics techniques (e.g., imputed gene expression analysis; Gamazon et al., 2015, 2019) will show that
genes associated with musicality traits in well-powered GWAS are expressed in neurobiological and neurodevelopmen-
tal processes known to underly language traits.
4) How do neural endophenotypes support co-varying musical and language traits?
(cid:129) PGSs for musical traits derived from well-powered samples will mediate relationships between individual variation in
neuroimaging correlates (e.g., EEG; MEG; DTI) and language phenotypes in an independent sample, and vice versa.
(cid:129) Multivariate analyses (e.g., Grotzinger et al., 2019) will uncover the shared genetic architecture between music, lan-
guage, and relevant neural endophenotypes.
Neurobiology of Language
630
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
5) When in development are genetic predispositions most sensitive to environmental factors in influencing outcomes for
musical and language abilities? When are genes associated with musicality-language links expressed in (neuro)biological
systems of interest?
(cid:129) Longitudinal or cross-sectional developmental studies will show enrichment for genes expressed or regulated during
specific stages of neurobiological development (e.g., in fetal, neonatal, and infancy) in genes common to musical
and language traits.
(cid:129) Longitudinal or cross-sectional developmental and lifespan studies will show that genetic risk/resilience for musical traits
will differentially predict speech-language trajectories across early development (e.g., increase in performance across
childhood and adolescence) and aging (e.g., maintenance and decline across middle age and old age).
6) Who is most at-risk for speech and language difficulties? How can we better identify these individuals?
(cid:129) Individuals with higher polygenic risk scores for music-related impairments will be more likely to have clinical speech-
language difficulties.
3) Considering genetic explanations alongside transfer effects:
(cid:129) Consider hypotheses that compete with, or are complementary to, neuroplasticity-
based frameworks, when explaining behavioral outcomes studied using correlational
designs. For example, test genetic and gene-environment interaction-based hypoth-
eses, alongside neuroplasticity-based frameworks, when two behavioral traits such as
music engagement and language abilities are related.
How Can the MAPLE Framework Advance the Field?
1) Genetic architecture: The MAPLE framework can guide our understanding of the genetic
basis of the human capacity of music and language, their genetic associations, their genetic
influences on mediating biological and neural endophenotypes, and their shared evolution-
ary histories (see Box 1 and Box 2 for further discussion).
2) Individual differences: The MAPLE framework can help clarify covarying individual differ-
ences in musicality and language, including interactions between genetic, neurobiological,
cognitive, and environmental factors influencing these links. The framework can also help
better characterize known dissociations between music and language processing (Chen
et al., 2021; Clos et al., 2013). These dissociations can occur in the presence of shared
genetic architecture but non-shared neural architecture. Further, the framework allows for
temporary state-level dissociations between musicality and language, while accounting for
stable trait-level associations (see Box 2 for further discussion).
3) Clinical efforts for speech-language disorders: Given emerging evidence that individuals with
musical impairments are often at higher risk for language-related disorders (Ladányi et al.,
2020), the testable genetic predictions of the MAPLE framework can help us better understand
biological mechanisms of risk profiles for speech-language disorders, and possibly identify
biomarkers to aid early identification and timely intervention. For example, accounting for
polygenic risk scores for musical impairments (e.g., rhythm perception) could potentially
serve as a powerful transdiagnostic approach cutting across clinically distinct speech-
language disorders (Lense et al., 2021; Sauer-Zavala et al., 2017). Further, testing the MAPLE
framework will help validate neurobiological and behavioral dimensions that can bridge
Neurobiology of Language
631
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Box 2. Disentangling genetic and neural architecture of musical and language functions in the brain using the MAPLE framework.
While the MAPLE framework is primarily focused on associations between musicality and language traits, there is some evidence
from functional neuroimaging of dissociations between brain networks recruited for language and musical functions (Chen et al.,
2021; Norman-Haignere et al., 2022; Peretz et al., 2015). We argue that the MAPLE framework cannot only co-exist with findings of
neural dissociations between musical and language processing, but can also help further characterize them in the following ways.
1) The MAPLE framework is consistent with other frameworks linking music and language, which predict the dual phenomena
of neural sharing and neural specialization underlying musical and linguistic function (e.g., Patel, 2021).
There are many possible ways in which the genetic architecture of complex traits could manifest in cross-trait phenotypic asso-
ciations, but the central tenet of the MAPLE framework is that there is shared biology between musicality and language traits.
Nevertheless, we do also expect some unique/non-shared variance in individual differences in these traits, as well as in their
underlying neural mechanisms. That is, phenotypic correlations between musicality and language could arise from shared
genetic alleles at specific locations on the genome (i.e., loci) that influence separate components of neural circuitry, alongside
shared components.
Consider the following example regarding musical rhythm and reading traits:
As reviewed in the Musicality, Reading Readiness, and Literacy section, there are robust and well-replicated associations
between rhythm and reading skills, despite the signal characteristics of musical rhythms and printed word being quite distinct.
Moreover, current knowledge about the networks of brain areas activated during reading (Martin et al., 2015) and musical rhythm
tasks (Kasdan, Burgess, et al., 2022) suggests that they are minimally overlapping (although more research is needed to directly
disentangle potential neural overlap).
Yet, it is well-established that reading skills are supported by phonological learning (Snowling, 2001), which in turn has many par-
allels to complex vocal learning, the subject of Patel’s (2021) revised vocal learning hypothesis. Recent genomic results demonstrate
that songbird vocal learning exhibits shared genetic architecture with human beat synchronization (Gordon et al., 2021), thus
suggesting convergent evolution evidence in support of the revised vocal learning hypothesis. Patel describes possible delineation
within language circuitry that may have co-evolved with rhythm, specifically predicting the dorsal stream of the language network
(more so than the ventral stream) to be tuned to complex vocal learning and phonological processing, and to share circuitry with
beat perception and synchronization. The dorsal stream may be a particularly interesting candidate area to explore the relationship
between shared neural and genetic architecture of rhythm and phonological processing (Patel, personal communication).
Phenotypic correlations between musical rhythm and reading could be thus explained by the MAPLE framework in multiple ways:
(cid:129) Shared genetic architecture (i.e., the same sets of alleles at hypothesized common genetic loci between rhythm and reading
traits) acting upon shared neural circuitry (e.g., dorsal stream areas).
(cid:129) Shared genetic architecture acting on separate components of the neural circuitries of rhythm and phonological processing
(e.g., neuroimaging studies show separate neural activation patterns when processing rhythmic vs. phonological stimuli,
while genetic correlations are still found between rhythm and phonology/reading-related traits due to pleiotropy).
(cid:129) Separate genetic architecture acting on common components of these neural circuitries (e.g., genes at different loci that
influence brain structure and function in some overlapping areas/processes between rhythm and reading).
2) The MAPLE framework can account for the biological basis of associations between stable individual differences in
musicality and language traits, even in the presence of biological dissociations between musicality and linguistic states.
Here, it is important to clarify conceptual distinctions between variation in traits and variation in states. Trait-dependent variance
captures variation across individuals relative to group means, whereas state-dependent variance captures variation over repeated
Neurobiology of Language
632
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
observations of the same individual over time, allowing for changes in environmental, or developmental factors (Sanchez-Alonso &
Aslin, 2020). Genomic variation between people is known to contribute to constitutive individual differences in musicality and
language traits (e.g., rhythm discrimination abilities or morphosyntactic skills). On the other hand, experimental tasks that are
designed to observe group-level differences in neural processing, across repeated observations (e.g., syntactic congruity vs. syntac-
tic violation conditions), instead target the state-level, and are often not well suited for capturing stable individual differences in traits
(see Hedge et al., 2018, for an extended discussion about trade-offs between stable individual differences metrics and reliable
experimental effects). Thus, individual or group differences found in brain responses to musical or linguistic states do not necessarily
need to converge with the central focus on the current work: stable individual differences in musicality and language traits. Pre-
dictions of the MAPLE framework about phenotypic and genetic overlap between individual differences in musical and language
abilities are therefore consistent with evidence for neural dissociations between musical and language states, such as:
(cid:129) Chen et al.’s (2021) finding that language-specific brain regions appear to have no sensitivity to structural musical violations,
and that individuals with aphasia who showed grammatical processing impairments had intact tonal music perception.
(cid:129) Peretz et al.’s (2015) finding that there appears to be a great deal of neural specialization and neural sharing between
musical and speech processing, and their crucial reminder that neural overlap (as defined by neuroimaging studies) does
not in itself mean neural resource sharing. They argue that it is possible that neural circuits appear overlapping because
they are intermingled or adjacent.
(cid:129) Norman-Haignere et al.’s (2022) finding of a neural population that responds specifically to music with sung melodies, but
that does not respond to other musical stimuli or speech stimuli.
Future Directions: As neurogenetic advances are made to map genome-wide influences on more nuanced functional networks,
neural populations, and microstructures, the MAPLE framework will allow us to characterize the genetic influences on neural
specialization (i.e., non-shared elements), as well as neural sharing, between musicality and language traits.
current developmental disorder categories, as called for by the research domains and con-
structs (RDoC) framework (Cuthbert, 2014). This in turn may offer more nuanced or alternative
ways of diagnosing and classifying disorders (e.g., the DSM and ICD manuals), and improved
precision treatment approaches based on specific (neuro)genomic risk profiles.
4) Cognitive science debates: The grounding of the MAPLE framework in polygenic pleiotropy pro-
vides new directions for intervening productively in long-standing cognitive science debates
concerning the domain-specificity or domain-generality of aspects of music and language
(Besson & Schön, 2012; Brandt et al., 2012; Lagrois et al., 2019; Morrill et al., 2015;
Swaminathan et al., 2018), and about cognitive and neural resource sharing between music
and language processing (e.g., Abrams et al., 2011; Patel, 2012, 2021; Peretz et al., 2015). For
example, musical rhythm perception and reading-related processing may recruit partially dis-
tinct neural networks and brain regions, but these traits may still be influenced by shared genetic
architecture, which can be clarified using genomic approaches (see Box 2 for further discussion).
5) Evolutionary theories: By focusing on shared genetic architecture, the MAPLE framework is
consistent with theories that posit common evolutionary and biological roots for the musical
and language abilities of our species (Brown, 2017; Mehr et al., 2021; Morley, 2013;
Mithen, 2005; Patel, 2012, 2021). See Box 2 for further discussion.
SUPPORTING EVIDENCE FOR THE MAPLE FRAMEWORK
The remaining sections of this article provide an overview of the broader field of research exam-
ining links between musicality and language, and a detailed (non-exhaustive) thematic review of
patterns found specifically within the individual differences literature. Studies discussed below
provide supporting evidence for the MAPLE framework, and point to relevant phenotypes
Neurobiology of Language
633
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
(behavioral evidence of correlations), and potential neural endophenotypes (neural correlates of
musicality-language associations), that can become a focus of future genetics investigations.
State of the Musicality-Language Field
We conducted bibliometric mapping based on connected terms in the literature to understand
where individual differences approaches (the focus of the present work) are situated in the larger
research area of music and language, and relative to musical training approaches (see the
Supporting Information for further details on bibliometric mapping). Briefly, representative
terms related to music and language were searched in PubMed and visualized using
VOSviewer (van Eck & Waltman, 2010). The analysis showed four major emerging clusters in
the literature, based on connections between terms: (1) Music and speech perception in the
context of hearing, hearing loss, and cochlear implant users (Figure 3, yellow); (2) Music
and language processing in the context of tonal language backgrounds and second language
learning, with implications for reading-related disorders such as dyslexia (Figure 3, blue); (3)
Neuroimaging of music and language, in healthy brains as well as contexts of brain injury or
lesions (Figure 3, red); (4) Music in the context of clinical interventions and related intervention
outcomes, e.g., for treatment of speech-language pathology (Figure 3, green).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Bibliometric mapping based on co-occurring music and language terms. Figure shows the major clusters of research areas emerging
when 60% of the most relevant terms shared between published articles are shown. Search terms included common terms associated with
music and language science.
Neurobiology of Language
634
Understanding musicality-language links
We queried the term “individual differences” and found it had 84 occurrences, which indi-
cates that this area is attracting attention. In contrast, the terms “formal musical training,”
“musical training,” and “music training” together showed 360 hits, accounting for a much
larger portion of the academic interest in the links between music and language. Further,
“musical training” was linked to a wider variety of terms within each of the four clusters rel-
ative to “individual differences” (Figure 4; see Figure S1 for additional details). Taken together,
these analyses provide a bibliometric overview of musicality-language studies and highlight
“individual differences” as an emerging area.
Literature Review: Associations Between Individual Differences in Musical and Language Abilities
Individual differences literature in a nutshell
To examine phenotypic associations between musicality and language-related traits, we syn-
thesized studies reporting correlations between musical and language abilities. Specifically, we
focused on associations found in the form of significant positive correlations between at least
one musicality-related and one language-related phenotype. Studies can be broadly classified
as examining associations between musical rhythm or tonal-melodic skills, and speech, gram-
mar, or reading-related traits. (See Supporting Information for details on the literature search
terms that were used to inform the framework. Note that this literature synthesis is theoretically
driven and not meant to be exhaustive or to take the place of a systematic review or meta-
analysis.) The literature reviewed here represents a variety of samples including adults, chil-
dren, and infants; individuals with or at-risk for language-related disorders; and second
language (L2) learners, foreign language (FL) learners, or individuals with tonal language
backgrounds (e.g., Cantonese). Figure 5 provides an overview of studies that inform our frame-
work, demonstrating accumulating and converging evidence that musicality covaries with
multiple language-related traits across a broad population. The literature provides extensive
evidence for positive correlations between individual differences in musical and language
abilities, spanning both rhythm and tonal-melodic measures, and speech-language and reading
measures. Figure 5 also provides a snapshot of mixed results and areas of relative sparsity to
guide future research efforts.
Musicality and speech perception
In the context of
Behavioral evidence: Musical abilities are correlated with speech task performance.
spoken languages, successful speech perception is fundamental to language acquisition and
communication. Speech perception involves many complex processes including segmenta-
tion of auditory speech streams; parsing the embedded prosodic and rhythmic patterns of
speech to perceive tone, context, emotion, and pragmatic information during discourse; and
being able to hear speech in non-ideal or noisy talker conditions. Beyond typical develop-
ment, speech perception deficits are associated with speech-language and reading disorders
such as dyslexia (Goswami, 2011). Many studies have previously found that musicians show
greater perceptual sensitivity to speech (Bidelman & Alain, 2015; Magne et al., 2006; Marie
et al., 2011), compared to non-musicians. Here, we synthesize findings from the studies shown
in Table 3, showing associations between individual differences in musicality and speech per-
ception phenotypes.
Given that speech prosody partially captures the rhythms embedded in speech, it is perhaps
unsurprising that musical rhythm sensitivity is emerging as a predictor of speech rhythm per-
ception (Hausen et al., 2013; Morrill et al., 2015). Beyond rhythm, individual differences in
musical pitch contour discrimination, and parsing pitch contours in speech, are also
Neurobiology of Language
635
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure mapping “individual difference” and “musical training” within bibliographic clusters. When the term “individual difference”
Figure 4.
occurred in publications, it was most commonly accompanied by the terms “speech perception,” “test,” “child,” “skill,” “year,” “processing,”
while “musical training” was linked to a much wider array of terms representing its more widespread use in the music and language science
literature. Plurals in the figure are encompassed by singular forms (e.g., “individual difference” also comprises “individual differences”).
Neurobiology of Language
636
Understanding musicality-language links
Figure 5. Studies examining within-sample associations between musicality and language traits. Each bubble corresponds to a single study,
and studies are grouped by which broad language-related construct(s) they examined (e.g., speech perception, grammatical skills, reading-
related abilities, or multiple) on the horizontal axis, and by which musicality construct(s) they examined (e.g., tonal-melodic or rhythm skills)
on the vertical axis. Studies spanning multiple musicality or language traits are represented by multiple bubbles as relevant.
associated (Cohrdes et al., 2016; Perrachione et al., 2013). This suggests that underlying neu-
ral and genetic architecture may be shared between musical and speech processing. Impor-
tantly, these associations reinforce the value of studying tonal-melodic perceptual abilities in
the context of non-tonal languages, alongside the more common study of lexical tone (illus-
trated in Figure 3), discussed further in the Musicality, Second Language Learning, and Tonal
Language Development section.
Musical rhythm skills, specifically, appear to be associated with speech discrimination
(Swaminathan & Schellenberg, 2020) and speech perception in noise (SPIN: Slater et al.,
2018; Slater & Kraus, 2016; Yates et al., 2019), in both children and adults. Yates et al.
(2019) also report positive correlations between musical experiences and training (both formal
and informal) and SPIN. These results leave open the possibility that SPIN abilities may be
shaped by musical experience, while interacting with (potentially genetically driven) indi-
vidual differences in musical and speech listening skills (e.g., Bidelman & Alain, 2015). For
example, individuals with higher musical aptitude may gravitate towards music training (i.e.,
gene-environment correlation). The heritability of speech perception task performance is
for genetic efforts
largely unknown, and information about
towards understanding music-speech links. At present, there is evidence that both speech
articulation (Table 1) and non-speech auditory perception in noise (Brewer et al., 2016) are
moderately heritable.
this would be beneficial
Alongside perceptual abilities, rhythm production and imitation are also associated with
speech encoding and SPIN (Slater et al., 2018; Woodruff Carr et al., 2014). For example, the
ability to accurately reproduce drumming sequences, drum in time to the beat of a song, and
to a metronome, were all correlated with SPIN in adults (Slater et al., 2018), pointing to the
Neurobiology of Language
637
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 3.
Studies reporting associations between musicality and speech perception.
Study
Boll-Avetisyan et al.
(2020)
Borrie et al. (2017)
Borrie et al. (2018)
Christiner and Reiterer
(2018)
Cohrdes et al. (2016)
Hausen et al. (2013)
N
46 (23 dyslexic)
50
98
35
44
64
Age
17–35
18–29
22–62
5–6
5–7
19–60
Tonal-melodic construct
Rhythm construct
Rhythm discrimination
Rhythm discrimination
Speech perception
construct
Speech rhythm sensitivity
Speech rhythm sensitivity
Speech learning
Rhythm discrimination
Melody discrimination
Speech rhythm sensitivity
Rhythm discrimination
Melody discrimination
Speech imitation
Rhythm reproduction
Melody reproduction
Speech pitch perception
Beat synchronization
Harmonic discrimination
Beat perception
Melody discrimination
Speech prosody perception
Holliman et al. (2010)
102
5–7
Rhythm reproduction
Speech rhythm sensitivity
Harmonic perception
Pitch perception
U
n
d
e
r
s
t
a
n
d
i
n
g
m
u
s
i
c
a
l
i
t
y
–
l
a
n
g
u
a
g
e
l
i
n
k
s
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
Hutchins (2018)
Mankel, Barber, and
Bidelman (2020)
Mankel and Bidelman
(2018)
90
14
28
Morrill et al. (2015)
179
Rhythm discrimination
3–6
23–27
19–25
18–49,
Beat synchronization
Singing error
Speech-in-noise perception
Beat discrimination
Melody discrimination
Speech processing
Tempo discrimination
Harmonic discrimination
Beat discrimination
Melody discrimination
Speech-in-noise perception
Tempo discrimination
Harmonic discrimination
Rhythm discrimination
Melody discrimination
Speech prosody perception
M = 19.9
Metrical perception
Musical memory
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Perrachione et al. (2013)
18
Slater and Kraus (2016)
54 males
18–30
18–35
Melody discrimination
Speech pitch perception
Rhythm discrimination
Melody discrimination
Speech-in-noise perception
N
e
u
r
o
b
o
o
g
y
i
l
o
f
L
a
n
g
u
a
g
e
6
3
8
Slater et al. (2018)
31 (17 drummers)
M = 24.7
Beat synchronization
Pitch discrimination
Speech-in-noise perception
Swaminathan and
Schellenberg (2020)
Woodruff Carr et al.
(2014)
Yates et al. (2019)
91
35
24
6–9
3–4
Rhythm discrimination
Melody discrimination
Speech perception
Musical memory (melodic)
Beat synchronization
Melody discrimination
Speech encoding
Rhythm discrimination
19–40
Beat synchronization
Melody discrimination
Speech-in-noise perception
Rhythm discrimination
Note. Rows correspond to individual studies, and columns correspond to study Ns, age ranges in years, specifics of musical rhythm and tonal-melodic constructs measured, and speech
constructs measured. When additional information is necessary about the N, specific descriptions are given in parentheses. When age range is not available, or not representative of the
sample mean, the mean is provided in addition to the age range. Only musical constructs that were reported in relation to speech perception constructs are noted.
U
n
d
e
r
s
t
a
n
d
i
n
g
m
u
s
i
c
a
l
i
t
y
–
l
a
n
g
u
a
g
e
l
i
n
k
s
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
N
e
u
r
o
b
o
o
g
y
l
i
o
f
L
a
n
g
u
a
g
e
6
3
9
Understanding musicality-language links
involvement of sensorimotor mechanisms. In the Slater et al. (2018) study, while beat synchro-
nization was associated with SPIN in both non-musicians and percussionists, reproducing or
imitating rhythmic patterns was only associated with SPIN in non-musicians, raising questions
about interactions between genetic predispositions and experience-dependent neuroplasticity.
Neural correlates of associations between musicality and speech perception. The behavioral asso-
ciations discussed above are supported by neural studies of speech encoding in subcortical
and cortical areas. For example, individuals with enhanced musical perception skills in gen-
eral (Mankel & Bidelman, 2018), and rhythm skills more specifically (Tierney et al., 2017),
show more efficient subcortical and cortical speech encoding mechanisms. In these studies,
higher music aptitude was associated with enhanced neural responses involved in both fast
(high-frequency) and slow (low-frequency) integration of auditory information over time.
Emerging neural evidence of music-speech links reflects the complex interplay between
perceptual, cognitive, and motor traits involved in rhythmic abilities (see Cannon & Patel,
2021; Kotz et al., 2018). Tierney et al. (2017) showed that rhythmic abilities can be decom-
posed into at least two constructs—sequencing and synchronization—and that these factors
are supported by differential brain networks involved in integrating auditory information. For
example, cortical responses tracked sequencing (which correlated with language traits such as
verbal memory and reading), whereas subcortical frequency-following responses tracked syn-
chronization (which correlated with non-linguistic auditory task performance).
Other studies suggest that subcortical speech encoding and reading processing are inter-
twined, and are associated with musical rhythm (Strait et al., 2011) and categorization of speech
sounds (Mankel et al., 2020). Based on evidence from preschoolers’ pre-reading skills, Woodruff
Carr et al. (2014), and others have argued for an integrative biological framework in which speech
encoding, “beat-keeping”/rhythm skills, and reading development are fundamentally linked.
Event-related potential (ERP) studies suggest that neural sensitivity to speech prosody is
associated with musical rhythm abilities. For example, Magne et al. (2016) measured partic-
ipants’ speech rhythm sensitivity using bi-syllabic words with the final word either matching or
mismatching the stress pattern of the preceding words. Participants showed an increased
fronto-central negative shift in ERPs in the stress mismatch versus match conditions, and the
magnitude of this neural response predicted speech rhythm sensitivity. Another study (Fotidzis
et al., 2018) found a similar negative shift when participants were visually presented with bi-
syllabic words with a trochaic (strong-weak) stress pattern, preceded by weak-strong tones
(i.e., mismatched stress patterns). The magnitude of the negative shift in ERPs was correlated
with both rhythm aptitude and reading comprehension skills.
The above studies suggest some degree of shared neurobiology underlies musical and speech
rhythm processing. Based on frameworks presented for the neural processing of beat and timing
in auditory and speech stimuli (Kotz et al., 2018; Kotz & Schwartze, 2010), mechanisms
underlying these correlations may involve previously identified subcortico-cortical networks
involved in processing rhythmic aspects of music and speech (see Fiveash et al., 2021).
Musicality, reading readiness, and literacy
Behavioral evidence: Musical abilities are correlated with literacy-related skills. By some accounts,
almost two-thirds of twelfth graders in the United States do not possess fundamental skills
required to master college-level reading material, and over 17% of adults have low English
literacy (National Center for Education Statistics, 2019). Literacy is a crucial driver of socio-
economic (SES) outcomes: Individuals with low literacy skills are more likely to have fewer
Neurobiology of Language
640
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
employment opportunities and earn lower wages (Decker & Decker, 1997), in addition to
increased health risks (Baker et al., 1997). A host of reading-related skills are important build-
ing blocks for reading comprehension, including decoding, fluency, and phonological
awareness skills. Table 4 provides further detail for studies examining associations between
musicality and several reading-related phenotypes.
Converging evidence shows that a variety of rhythm-related skills (e.g., rhythm production,
beat synchronization, memory for rhythm, timing/metrical perception) are robustly associated
with building blocks of reading development. Musical rhythm perception and production are
associated with phonemic skills such as word recognition and spelling (Douglas & Willatts,
1994), and letter-sound knowledge (Ozernov-Palchik et al., 2018; Ríos-López et al., 2019). Beat
synchronization and rhythm discrimination are also associated with phonological awareness,
rapid naming, and auditory working memory (Anvari et al., 2002; Moritz et al., 2013; Woodruff
Carr et al., 2014). Musicality as a whole seems to be related to children’s early literacy skills at
the start of formal education. Scores on a comprehensive music aptitude test (consisting of pitch,
rhythm, and meter discrimination and production tasks) were associated with phonological
awareness, nonword repetition, and rapid naming fluency in school-aged children even after
controlling for key demographic and nonverbal cognitive variables (Degé et al., 2015).
Longitudinal investigations show that rhythm predicts later reading readiness, further suggesting
a bootstrapping role of early musical ability for building blocks of reading development. Rhythm
imitation (tapping a rhythmic sequence from memory) at entry to the first grade was predictive of
children’s literacy skills as tested at the end of the same school year (Lundetræ & Thomson, 2018),
and rhythm at kindergarten, predicted phonological awareness and word reading in the second
grade (Moritz et al., 2013). David et al. (2007) found that rhythm skills in grade 1 predicted pho-
nological awareness in grades 2 to 5. In a large sample (N = 600), rhythm reproduction in
kindergarteners uniquely explained 14% of the variance in reading at grade 2, controlling for
demographic factors, phonological abilities, visuo-spatial attention, and processing speed
(Dellatolas et al., 2009), with children with lower SES backgrounds showing stronger rhythm-
reading links. Taken together, these findings make a compelling case for exploring musical
rhythm as a tool for reading interventions (Flaugnacco et al., 2014).
Evidence on rhythm-reading links also robustly converges at older stages of development.
For example, González-Trujillo et al. (2014) found that in school-aged readers of Spanish,
rhythm production/imitation (tapping from memory) was associated with reading fluency,
and reading with accurate lexical stress patterns. Note that lexical stress is explicitly marked
in Spanish words that diverge from canonical lexical stress patterns. Lê et al. (2020) similarly
showed a link between rhythm skills and reading and spelling skills in third graders, after
ruling out mediating contributions from both motor skills and phonological awareness.
The ability to analyze sound sequences seems specifically relevant for language/literacy.
Grube et al. (2012) found associations between auditory perception of time and phonological
skills, and between temporal sensitivity in musical sequences and phonological skills, across
several standardized language and reading assessments. Further, a small study in adults (n =
26) representing a host of different native languages found that the ability to detect a regular
rhythm within a highly irregular sequence (measured by a regularity detection threshold) predicts
reading scores (here, rapid automatized naming) in their native language (Bekius et al., 2016).
In children, studies also show associations between tonal-melodic skills such as pitch
processing, and phonemic awareness (Loui et al., 2011), nonword repetition (Flagge et al.,
2016) and phonological awareness (Steinbrink et al., 2019). Pitch production skills are also
correlated with phonemic awareness, controlling for IQ (Loui et al., 2011). Echoing their
Neurobiology of Language
641
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
N
e
u
r
o
b
o
o
g
y
i
l
o
f
L
a
n
g
u
a
g
e
6
4
2
Table 4.
Studies reporting associations between musicality and reading-related skills.
Study
Anvari et al. (2002)
N
100
Age
4–5
Rhythm construct
Rhythm discrimination
Tonal-melodic construct
Melody discrimination
Reading-related construct
Phonological awareness
Rhythm reproduction
Harmony discrimination
Reading ability
Harmony analysis
Bekius et al. (2016)
Cohrdes et al. (2016)
26
44
20–40
5–7
Beat perception
Reading fluency
Rhythm reproduction
Melody reproduction
Phonological awareness
David et al. (2007)
53
M = 6.34
Beat synchronization
Beat synchronization
Harmonic discrimination
Phonological awareness
Reading fluency
Degé et al. (2015)
55
M = 6.25
Rhythm discrimination
Melody discrimination
Phonological awareness
Beat discrimination
Pitch discrimination
Nonword repetition
Dellatolas et al. (2009)
625
5–6
Beat synchronization
Singing ability
Rhythm reproduction
Rhythm reproduction
Phonological awareness
Reading comprehension
Douglas and Willatts (1994)
Flagge et al. (2016)
González-Trujillo et al. (2014)
78
36
67
7–9
7–8
8–9
Rhythm discrimination
Pitch discrimination
Word recognition/spelling
Pitch discrimination
Phonological awareness
Rhythm reproduction
Reading fluency
Grube et al. (2012)
238
11
Rhythm discrimination
Pitch discrimination
Phonological awareness
Holliman et al. (2010)
102
5–7
Timbre discrimination
Rhythm reproduction
Rhythm discrimination
Reading ability
Phonological awareness
Hutchins (2018)
90
3–6
Beat synchronization
Singing error
Phonological awareness
Lê et al. (2020)
278
M = 8.4
Beat synchronization
Reading fluency
Phonological awareness
Reading ability
U
n
d
e
r
s
t
a
n
d
i
n
g
m
u
s
i
c
a
l
i
t
y
–
l
a
n
g
u
a
g
e
l
i
n
k
s
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
N
e
u
r
o
b
o
o
g
y
l
i
o
f
L
a
n
g
u
a
g
e
6
4
3
Loui et al. (2011)
32
M = 7.6
Pitch discrimination
Phonological awareness
Pitch production
Lundetræ and Thomson (2018)
479
6
Beat synchronization
Phonological awareness
Moritz et al. (2013)
30
Starting at age 5,
ending at age 7
(longitudinal)
Beat synchronization
Rhythm reproduction
Rhythm discrimination
Ozernov-Palchik et al. (2018)
74
M = 5.8
Rhythm discrimination
Reading fluency
Spelling
Phonological awareness
Reading ability
Phonological awareness
Reading fluency
Peynircioğlu et al. (2002) (1)
Peynircioğlu et al. (2002) (2)
Politimou et al. (2019)
Ríos-López et al. (2019)
Steinbrink et al. (2019) (1)
32
40
46
43
54
4–6
3–7
3–5
4–5
5–7
Rhythm reproduction
Melody production
Phonological awareness
Rhythm reproduction
Melody production
Phonological awareness
Rhythm discrimination
Pitch discrimination
Phonological awareness
Tempo discrimination
Melody discrimination
Beat synchronization
Melody production
Beat synchronization
Phonological awareness
Tempo discrimination
Pitch discrimination
Phonological awareness
Rhythm discrimination
Melody discrimination
Rhythm reproduction
Steinbrink et al. (2019) (2)
96
7–11
Tempo discrimination
Pitch discrimination
Reading comprehension
Strait et al. (2011)
Tierney and Kraus (2013)
Tierney et al. (2017)
Woodruff Carr et al. (2014)
42
58
64
35
8–13
14–17
Beat synchronization
M = 18.0
Beat synchronization
Rhythm reproduction
Rhythm discrimination
Melody discrimination
Spelling
Rhythm reproduction
Phonological awareness
Rhythm discrimination
Melody discrimination
Reading ability
Reading ability
Phonological awareness
Reading ability
3–4
Beat synchronization
Melody discrimination
Phonological awareness
Rhythm discrimination
Reading fluency
U
n
d
e
r
s
t
a
n
d
i
n
g
m
u
s
i
c
a
l
i
t
y
–
l
a
n
g
u
a
g
e
l
i
n
k
s
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Study
Yu et al. (2017)
Zhang et al. (2017)
N
511
54
Age
M = 20.6
8–10
Rhythm construct
Rhythm discrimination
Tonal-melodic construct
Pitch discrimination
Reading-related construct
Phonological awareness
Tempo discrimination
Pitch discrimination
Reading fluency
Table 4.
(continued )
Zuk et al. (2013)
45
6–8
Rhythm reproduction
Musical memory
Reading fluency
Phonological awareness
Note. Rows correspond to individual studies, and columns correspond to study Ns, age ranges in years, specifics of musical rhythm and tonal-melodic constructs measured, and reading
constructs measured. When additional information is necessary about the N, specific descriptions are given in parentheses. When age range is not available, or not representative of the
sample mean, the mean is provided in addition to the age range. Only musical constructs that were reported in relation to reading-related constructs are noted.
Tonal sensitivity (Mandarin)
Phonological awareness
U
n
d
e
r
s
t
a
n
d
i
n
g
m
u
s
i
c
a
l
i
t
y
–
l
a
n
g
u
a
g
e
l
i
n
k
s
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
N
e
u
r
o
b
o
o
g
y
l
i
o
f
L
a
n
g
u
a
g
e
6
4
4
Understanding musicality-language links
findings in the rhythm and temporal processing domain, auditory sequencing skills related to
pitch and timbre correlated with phonological awareness abilities (Grube et al., 2012).
Musical listening and production (e.g., when playing an instrument) involves encoding/
decoding hierarchically embedded information such as notes and chords, parallel to the
encoding/decoding of phonemes, syllables, words, and sentences involved in reading. Indeed,
two cohorts of preschoolers (American and Turkish) showed correlations between perfor-
mance on a tone-deletion task with melodic excerpts and an equivalent phoneme deletion
task with words and pseudowords (Peynircioğlu et al., 2002). Further, weaker performance
at transcribing musical sequences from memory has been found to be associated with reading
more slowly and less accurately (Zuk et al., 2013). Reading-related decoding skills in turn
highly predicted performance on other linguistic tasks in a cognitive battery, consistent with
the proposed role of musicality in language development (Figure 1).
Two studies shed light on the relationship between music and reading in the context of
non-alphabetic languages (e.g., Chinese languages), where the building blocks of reading
development involve other types of decoding than is typically measured in alphabetic lan-
guages. For example, melodic skills (but not rhythmic skills) were associated with semantic
processing while reading Chinese (Yu et al., 2017). Further, English phonological awareness
correlated with musical meter and pitch discrimination in English-Chinese bilinguals, medi-
ating the relationship between musical perception and English reading. However, Chinese
reading skills were not associated with musicality (Zhang et al., 2017).
Neural correlates of associations between musicality and literacy. Both cortical and subcortical
neural endophenotypes may be relevant for understanding music-reading links. Auditory
brainstem responses (ABRs)—subcortical markers of auditory encoding—have been linked
with speech perception skills in both children and adults. School-aged children showed
enhanced ABRs to syllables in a predictable versus variable speech context, and ABR magni-
tudes correlated with both musical aptitude and word reading skills (Strait et al., 2011). These
associations may exist even prior to literacy acquisition; for example, preschoolers’ beat syn-
chronization abilities were associated with ABRs to syllables, and to behavioral preliteracy
measures (e.g., phonological awareness, auditory short-term memory, and rapid automatized
naming: Woodruff Carr et al., 2014). In adults, associations between efficient neural markers of
auditory perception (e.g., ABRs to auditory clicks used in hearing screenings) are correlated
with greater musical experience, as well as enhanced phonemic decoding (Tichko & Skoe,
2018).
As noted throughout this article, greater musical experience could itself be a function of
both environmental and genetic influences. Similarly, evoked cortical responses to speech
sounds have been associated with enhanced musical rhythm and reading skills (Tierney
et al., 2017). Structural neural endophenotypes that might be involved in links between
musicality and literacy include gray matter volume of primary auditory cortex structures.
Specifically, larger relative gray matter volumes of Heschl’s gyrus compared to planum
temporale, in both hemispheres, were correlated with reading and spelling skills in 7- to
9-year-old children (Seither-Preisler et al., 2014). Larger right Heschl’s gyrus volumes also
correlated with more efficient processing of musical instrument sounds and tones (as measured
by evoked P1 responses through magnetoencephalography, MEG), and strongly correlated
with musical aptitude scores (Seither-Preisler et al., 2014). Beyond phonemic and phonolo-
gical building blocks of reading, musical rhythm abilities are also associated with individual
differences in electrophysiological correlates of reading (Fotidzis et al., 2018; Ríos-López
et al., 2019).
Neurobiology of Language
645
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Musicality and grammatical skills
Behavioral evidence: Musical abilities are correlated with morphosyntactic skills. Morphosyntactic
skills are foundational to language development across spoken and signed languages. Mor-
phological manipulations allow children to form words from smaller units of meaning, and
syntactic skills allow them to connect elements of language together to parse and produce
sentences at multiple levels of syntactic hierarchy. These skills also make it possible to connect
ideas within and across sentences, which facilitates other aspects of communication, e.g.,
expressing and comprehending ideas in speech and written language. Successful complex
syntax acquisition enables social relationships, school-readiness, educational outcomes, and
life skills (Brimo et al., 2017; Fujiki et al., 1999), and morphosyntactic disruptions are often a
key feature of DLD. Table 5 summarizes studies examining associations between musicality
and spoken grammar-related phenotypes.
Studies focused on preschoolers, school-aged children, and adolescents, find converging
evidence for associations between musical rhythm traits and grammatical development. For
example, in a sample of primary school-aged children (N = 25), Gordon, Shivers, et al.
(2015) demonstrated that individuals who were more accurate at distinguishing between musi-
cal rhythm sequences, also performed more accurately on tasks that probed children’s ability
to use particular morphosyntactic formations. Phonological awareness did not explain the
association between musical rhythm and grammatical abilities, and in a follow-up study in
a larger sample (N = 121), neither prosody perception skills nor working memory were found
to mediate the relationship between musical rhythm and morphosyntactic skills (Nitin et al.,
in press). As seen in Table 5, there are as yet very few studies exploring the relationship
between musical rhythm sensitivity and grammatical abilities.
These findings were extended by Swaminathan and Schellenberg (2020), who showed a
strong association between musical beat perception and receptive grammar in ~100 school-
aged children performing the Test for Reception of Grammar (TROG: Bishop, 2003). The
TROG is a grammatical comprehension task commonly used to measure grammatical impair-
ments in children with DLD. This test, which asks children to point to the picture that corre-
sponds with the sentence they hear, is commonly used to measure grammatical skill. Further,
Politimou et al. (2019) showed that rhythm perception was associated with performance on a
sentence imitation task (which encompasses both the receptive and expressive nature of
grammar) in preschoolers. In keeping with these observations, preschoolers who had stronger
abilities to synchronize to an external beat, had higher scores on tests of reading and sentence
imitation (Woodruff Carr et al., 2014). Moreover, Lee et al. (2020) also found a correlation
between rhythm discrimination and receptive grammar via a language comprehension task
that required participants to identify the agent of the sentence, in a wider age range of partic-
ipants (7- to 17-year-olds), while controlling for working memory, age, and musical training,
showing that the relationship between musical rhythm sensitivity and grammar cuts across the
developmental arc.
Melodic skills are also associated with grammatical abilities, and this relationship may be
moderated by the richness of musical exposure and level of musical engagement in children’s
home environments. For example, Politimou et al. (2019) found that children’s melody skills
and degrees of music in the home were associated with grammar skills, particularly with tasks
testing word structure awareness and sentence imitation. Moreover, in their study, the best
model predicting grammar further included parental musicality measures, suggesting a poten-
tial genetic influence on music and grammar traits in children. Relatedly, memory for melodic
patterns is associated with both musical rhythm and grammar ability (Swaminathan &
Schellenberg, 2020).
Neurobiology of Language
646
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
N
e
u
r
o
b
o
o
g
y
l
i
Table 5.
Studies reporting associations between musicality and morphosyntactic skills.
Study
Cohrdes et al. (2016)
Gordon, Jacobs, et al. (2015)
Gordon, Shivers, et al. (2015)
Lee et al. (2020) Exp 1
Lee et al. (2020) Exp 2
Nitin et al. (in press)
N
44
25
25
98
96
121
Age
5–7
5–7
5–7
7–17
7–17
5–8
Rhythm construct
Rhythm reproduction
Tonal-melodic construct
Grammar-related construct
Melody reproduction
Receptive grammar
Beat synchronization
Harmonic discrimination
Rhythm discrimination
Rhythm discrimination
Rhythm discrimination
Rhythm discrimination
Rhythm discrimination
Complex syntax
Expressive grammar
Receptive grammar
Receptive grammar
Expressive grammar
Complex syntax
Politimou et al. (2019)
46
3–5
Rhythm discrimination
Pitch discrimination
Expressive/receptive grammar
Swaminathan and Schellenberg
(2020)
Woodruff Carr et al. (2014)
91
35
6–9
3–4
Tempo discrimination
Melody discrimination
Beat synchronization
Melody production
Rhythm discrimination
Melody discrimination
Receptive grammar
Beat synchronization
Rhythm discrimination
Musical memory (melodic)
Expressive/receptive grammar
Note. Rows correspond to individual studies, and columns correspond to study Ns, age ranges in years, specifics of musical rhythm and tonal-melodic constructs measured, and morpho-
syntactic constructs measured. When additional information is necessary about the N, specific descriptions are given in parentheses. When age range is not available, or not representative
of the sample mean, means M are provided in addition to the age range. Only musical constructs that were reported in relation to grammatical/mophosyntactic constructs are noted.
o
f
L
a
n
g
u
a
g
e
6
4
7
U
n
d
e
r
s
t
a
n
d
i
n
g
m
u
s
i
c
a
l
i
t
y
–
l
a
n
g
u
a
g
e
l
i
n
k
s
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Neural correlates of associations between musicality and grammatical skills. A recent electroen-
cephalography (EEG) study found that individual differences in children’s neural entrainment
to the beat of rhythmic patterns (measured with evoked time-frequency analyses in beta and
gamma bands) predicted variation in their expressive grammar abilities (Persici et al., 2022).
These results are consistent with what many have proposed—that the overlap between musi-
cal rhythm and linguistic syntax is related to the processing of hierarchically organized syntax
in language sharing common biology with the metrical aspect of rhythm processing (Asano
et al., 2021; Fitch, 2017; Heard & Lee, 2020), for example, shared cognitive control circuitry
in the frontal cortex and basal ganglia (Asano et al., 2021). However, see Fedorenko and
Varley (2016) and Chen et al. (2021) for evidence that certain musical stimuli do not mean-
ingfully activate language regions.
Musicality, second language learning, and tonal language development
Behavioral evidence: Musical abilities predict second language (L2) learning and tonal language
development. Second language (L2) and foreign language learning have long been heralded
as a window into cognition. Findings about musicality from L2 and foreign language contexts
converge with studies in native speakers. For example, musical aptitude (including specific
abilities such as singing and harmonic discrimination) is associated with more accurate pho-
nemic perception and production abilities (see Milovanov & Tervaniemi, 2011, for a review;
Christiner & Reiterer, 2018), pronunciation (Turker et al., 2017) in a foreign language, and
phonological abilities in an L2 (Slevc & Miyake, 2006). Further, musical aptitude predicts
foreign language silent reading fluency better than other reading-related skills or auditory
working memory (Foncubierta et al., 2020). Musicality was also associated with more accurate
grammatical judgments of complex syntax in an artificial grammar learning task (Brod &
Opitz, 2012). A school-based study of educational achievement found that Italian students’
grades in music classes were correlated with their grades in English and French classes at
school (Picciotti et al., 2018). Given the relatively large sample size (N = 500), the naturalistic
setting (the school system), and assessment of two foreign languages, this evidence suggests
that individual differences in musical traits explain a proportion of the variation in foreign lan-
guage learning achievement.
What about languages in which musical skills (e.g., pitch perception) are also language skills
(e.g., lexical tone perception)? In this case, pitch skills might be a better predictor of language
development than musical rhythm skills, in children acquiring tonal languages (Antoniou et al.,
2015). However, pitch perception of auditory tones embedded in non-hierarchical contexts
(e.g., non-musical/non-linguistic stimuli) did not predict Cantonese lexical tone perception
(Wong & Cheng, 2020), suggesting that it is the hierarchical structures of musical rhythm that
are specifically relevant for morphosyntactic parsing, rather than musical rhythm perception
on the whole. This converges with other evidence about dissociations in tone versus lexical
tone perception, e.g., only a subset of Mandarin Chinese speakers with congenital amusia
show lexical tone perception deficits (Huang et al., 2015; Nan et al., 2010).
Neural evidence: The role of Heschl’s gyrus in musical and foreign language aptitude. Structural brain
imaging results have shown that individual differences in the morphology of the right Heschl’s
gyrus are associated with both phonetic coding in an unknown foreign language, and with
musical aptitude (Turker et al., 2017). Turker et al. argue that since the morphology of Heschl’s
gyrus tends to be stable, musical and language abilities have relatively reduced flexibility and
are privy to genetic influences or environmental influences that shape very early development
(e.g., at the prenatal or very early postnatal stages). Thus, Heschl’s gyrus could be a key brain
Neurobiology of Language
648
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
area of interest for examining genetic influences on phenotypic links between musical and
foreign language aptitude. This result converges with other findings implicating Heschl’s gyrus
in musicality-language links (e.g., Seither-Preisler et al., 2014, discussed in the Musicality,
Reading Readiness, and Literacy section; Sutherland et al., 2012, discussed in the Building
Blocks of Musicality and Language in Infancy section.
Building blocks of musicality and language in infancy
Infant-
Behavioral evidence: Emerging musical and language abilities are correlated in infancy.
directed speech (Fernald et al., 1989; Leong et al., 2017), nursery rhymes, and infant-directed
singing (Trehub & Trainor, 1998) are prevalent cross-culturally, and musical speech may in fact
support language processing at the earliest stages of life. Accumulating evidence shows that the
earliest stages of music and language development follow parallel tracks, and support culture-
specific attunement to both music and speech (Brandt et al., 2012). Here, we synthesize infant
studies examining correlations between early precursors of musical and language traits.
Infant studies contribute to understanding variability in musical and auditory traits that
are present at the foundational stages of music and language development. For example,
infants’ sensitivity to pitch (Leppänen et al., 2010) and to amplitude rise times of pure tones
(Kalashnikova et al., 2019) predict later language-related traits such as phonological aware-
ness, reading speed, and vocabulary development. Temporal auditory processing in infancy
has also been shown to predict subsequent language production (Rocha et al., 2020), reading
fluency (van Zuijen et al., 2012), and morphosyntactic skills (Höhle et al., 2014), consistent
with previously discussed studies in children and adults. Höhle et al. (2014) found that infants’
rhythmic discrimination predicted sentence comprehension and morphological ability at
age 5. Importantly, infants with family risk of language impairment were less able to discrim-
inate rhythmic patterns, versus typically developing infants, consistent with studies in older
children at-risk for language and reading disorders (see the Musicality and Developmental
Disorders of Reading and Language section).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neural evidence: Correlates of listening to tones, music, and sung speech. A handful of neural
investigations in infants have shown that individual differences in neural markers of temporal
auditory processing predict language and reading outcomes later in development (Fava et al.,
2014; François et al., 2017; van Zuijen et al., 2012). For example, newborns’ cortical responses
to word form violations in sung speech streams predict vocabulary size at 18 months (François
et al., 2017). Further, ERP responses to standard versus deviant inter-tone intervals in infancy are
associated with reading fluency in later life (van Zuijen et al., 2012), and infants’ hemodynamic
brain responses to both music and speech can be categorized into “high responder” and “low
responder” groups based on neural engagement patterns (Fava et al., 2014).
Relationships between auditory processing in infancy and longitudinal language outcomes
(e.g., Rocha et al., 2020) appear to persist in school-aged children. For example, Sutherland
et al. (2012) found in a study of school-aged children, that changes in volume/structure in the
auditory cortex (specifically Heschl’s gyrus) were associated with frequency modulation detec-
tion at 2 Hz—a measure of auditory perceptual sensitivity—and with reading and spelling
abilities.
As suggested by the MAPLE framework, genetic investigations of neural endophenotypes
associated with foundational musicality and language traits can help clarify the cascading
biological and developmental effects of musicality-language links, in infancy and later in
development.
Neurobiology of Language
649
Understanding musicality-language links
Musicality and developmental disorders of reading and language
Behavioral evidence: Musical abilities are correlated with language abilities in developmental disorders
of reading and language. Developmental reading and language disorders (e.g., dyslexia; DLD)
are primarily characterized by difficulties with written and/or spoken language abilities (e.g.,
reading, phonology, morphology, syntax). A recent theoretical framework synthesizes abun-
dant evidence that individuals with such disorders also often show deficits in musical rhythm
skills (Ladányi et al., 2020), and posits that atypical rhythm is a comorbid risk factor for child-
hood developmental speech and language disorders, potentially implicating shared genetic
architecture (the atypical-rhythm risk hypothesis). Studies of musicality in language disorder
contexts often emphasize group comparisons relative to typically developing individuals.
Boorom et al. (2022) provide an extended discussion of the role of musical abilities and musi-
cal interventions in developmental language disorders. Here, we synthesize studies using indi-
vidual differences approaches, which can provide more granular views of the entire spectrum
of musical and language-related abilities represented in the population.
Studies of individual differences in cohorts of children with DLD have found associations
between musical rhythm skills, and grammar skills, expressive and receptive language skills,
and artificial grammar learning abilities. For example, an early study of children with DLD
found associations between musical rhythm sensitivity and grammar skills across multiple
standardized language tests (Weinert, 1992). This association also extended to an artificial
grammar learning task with prosodically enriched stimuli. Across the spectrum of language
ability (e.g., in children with DLD as well as in typically developing children), studies have
found associations between musical beat perception and beat synchronization (i.e., ability
to tap to a beat), and receptive and expressive language skills and reading skills (Corriveau
& Goswami, 2009; Cumming et al., 2015), controlling for age and nonverbal IQ. Reading-
related skills investigated include phonological awareness, word and nonword reading,
reading comprehension, spelling, reading fluency, and nonword repetition.
There is also some evidence suggesting that tonal-melodic skills are associated with lan-
guage abilities in preschoolers with DLD (Sallat & Jentschke, 2015). Here, morphosyntactic
skills (measured by a sentence repetition task) were also found to correlate with performance
on melody recognition, melody discrimination, and rhythmic-melodic perception tasks.
Although, results should be interpreted cautiously because overall performance on the lan-
guage task was at chance levels, and analyses did not control for demographic or cognitive
factors.
Similarly, in the context of dyslexia, musical rhythm measures such as metrical sensitivity or
rhythm reproduction are associated with reading-related skills, both in children (Flaugnacco
et al., 2014; Huss et al., 2011; Overy, 2003) and in adults (Boll-Avetisyan et al., 2020).
Forgeard et al. (2008) also reported significant correlations between melody discrimination
abilities and phonemic awareness in a small sample of school-aged children with dyslexia. In
a large-scale online cohort, time-based but not-pitch based congenital amusia was associated
with self-reported dyslexia and speech disorder (Peretz & Vuvan, 2017), further highlighting
the unique role that biological mechanisms supporting rhythm/temporal processing may play
in driving musicality-language links.
Insights from the literature
Converging evidence thus suggests that musical abilities are highly relevant for our under-
standing of individual differences across the continuum of language abilities, in both typical
development and language disorder contexts, and in both native and L2 contexts. Specifically,
Neurobiology of Language
650
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
our literature synthesis highlighted the importance of multiple dimensions of musicality,
including beat synchronization, rhythm and melody discrimination, and pitch discrimination
abilities. Language-related phenotypes for which variance is explained in part by musical
abilities include nonword repetition, complex syntax, sentence recall, speech perception-in-
noise, speech rhythm discrimination, and L2 phonemic perception and production.
Studies examining associations between musical and language abilities have often exam-
ined and/or controlled for environmental factors such as family SES and degree of formal
and/or informal musical training. However, recall from the MAPLE framework (Figure 2) that
so-called environmental factors can themselves show significant heritability. For example,
musicality can be influenced by gene-environment interactions (when genetic predispositions
moderate environmental effects on a phenotype or endophenotype), or gene-environment cor-
relations (when environments are selected or created in ways that are complementary to
genetic predispositions). Therefore, the study of environmental effects can potentially be con-
founded by genetic factors (see Hart et al., 2021, for an expanded discussion of the issue).
Future research on musicality-language associations should therefore aim to incorporate
gene-environment correlations and gene-environment interactions into models where genetic
and environmental data are available. Alternatively, where genetic information is not avail-
able, one option is to incorporate Hart et al.’s (2021) familial control method as a kind of
genetic proxy. Here, the same trait is measured in parent and child and included as a covariate
to estimate the effect of the rearing environment. Note that while this method can be included
in some developmental work going forward, it may be currently difficult to account for genetic
confounds in this way where child and adult-friendly measures of a music or language con-
struct do not yet exist.
Prior studies have commonly included working memory as a measure of executive function
(EF) skills, but without considering other aspects of EF (e.g., inhibitory control; attentional
shifting). EF skills are implicated in both language (Gooch et al., 2016; Kapa & Erikson,
2020; Woodard et al., 2016) and musicality (Clayton et al., 2016; Moreno et al., 2011), and
are some of the most heritable psychological traits (Friedman et al., 2008). In fact, cognitive
control processes may have a particularly relevant role in musicality-language links, partic-
ularly for further understanding relationships between rhythm and grammar. For example,
the coordinated hierarchical control hypothesis (Asano et al., 2021) points to coordination
among the frontal cortex areas, for example, inferior frontal gyrus (IFG), dorsolateral prefron-
tal cortex (dlPFC), pre-supplemental motor area (pre-SMA), and the basal ganglia as a shared
mechanism underlying musical rhythm and linguistic syntax processing, and specifically
argues for the central importance of control processes such as inhibition, selection, and
maintenance. While it is common to study EF in the context of both musical and language
abilities, more robust investigations of individual differences in various measures of
EF—perhaps especially inhibitory control—at behavioral, neural, and genetic levels, would
allow for better developed predictions about musicality-language links across the lifespan.
A noteworthy pattern that emerged from the literature on musical abilities and reading-
related skills was that current investigations generally focus on smaller building blocks of lan-
guage (e.g., phonemes, syllables, words), but lack naturalistic reading tasks (e.g., reading
sentences/passages; reading comprehension). Including these reading contexts in future stud-
ies of musicality-language links can shed light on the range of reading functions that may be
supported by biological mechanisms involved in music and language, and may provide a
novel focus for developing music-based reading interventions.
Further, many studies on reading highlighted that performance on nonword repetition tasks
is robustly associated with both rhythmic and tonal-melodic musical abilities. Given this
Neurobiology of Language
651
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
pattern, and studies suggesting that nonword repetition is a good proxy for assessing overall
spoken language skills (i.e., higher-order language skills in the auditory-vocal modality), we
argue that future models examining musicality-language links should consider nonword
repetition as a key language-related trait of interest, beyond viewing it as a measure of pho-
nological awareness or working memory in the manner that it has sometimes been designated
in the phenotypic literature (e.g., Degé et al., 2015; Flagge et al., 2016). This point may be
especially important for advancing the genetic study of musicality-language links within the
MAPLE framework, as nonword repetition is an existing language-related phenotype in several
large-scale genetic cohorts, and its heritability (Table 1) and genetic architecture (Eising et al.,
2022) have been examined.
Our synthesis of the literature did not find a critical mass of studies focused on older adult-
hood reporting behavioral associations between musical and language abilities (although some
studies have older adults in their broader adult samples). However, there are known associations
between musical training and protective effects against age-related declines in speech percep-
tion outcomes in older adulthood (e.g., Bidelman & Alain, 2015), and interest in music-based
interventions for healthy aging (James et al., 2020), age-related neurodegenerative condi-
tions such as dementia or Alzheimer’s disease (Leggieri et al., 2019; Moreno-Morales et al.,
2020), or stroke rehabilitation (Grau-Sánchez et al., 2020). The MAPLE framework takes a life-
span view, proposing that individuals “high” on a spectrum of musical abilities may show
greater resilience to age-related decline in normal speech-language functions, e.g., SPIN or
hearing acuity (Figure 1). Systematic efforts to understand individual differences in
musicality-language associations in older adulthood, can create a strong foundation for a per-
sonalized approach to music-based interventions in the context of neurodegenerative con-
ditions. Importantly, a lifespan approach allows us to probe genetic risk and resilience to
speech-language outcomes based on individual differences in musicality.
Last, since musicality-language links are likely shaped by the diversity of both musical and
language experiences, future research should pay special attention to capturing individuals’
language backgrounds, including years of simultaneous or sequential multilingual exposure;
native tonal language backgrounds; and extent of engagement in language learning beyond
native language(s). Capturing variability in both language and musical experiences will
certainly provide a more complete picture of the relationship between musical and language
abilities, and variation across the spectrum of these abilities. Ultimately, to map population-
level relationships between genetic and neural mechanisms, environmental factors, and
musicality and language traits, it will be imperative for research efforts to represent diverse
cultures, languages, and musical systems.
Compared to the behavioral evidence showing covarying individual differences in musical-
ity and language traits, our literature review found that studies testing for neural correlates of
these associations were relatively sparse. Since the MAPLE framework proposes that the
behavioral associations commonly found between musical and language abilities are partly
driven by shared genetic influences, it is crucial to understand the neurobiological mecha-
nisms through which these genetic influences are exerted, to meaningfully test the framework’s
hypotheses. With the goal of mapping the relationships among genetics, neurobiology, and
perception/cognition, future research should aim to integrate neuroscience and genetics
approaches to test for potential neural endophenotypes of the (shared) genetic influences on
musicality and language traits.
Potential neural endophenotypes involving brain structures are already available as part of
large-scale data sets that include both neural and genetic information, such as the UK
Neurobiology of Language
652
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Biobank (Sudlow et al., 2015) or the Cohorts for Heart and Aging Research in Genomic Epi-
demiology (CHARGE) Consortium (Psaty et al., 2009). We can also define functional neural
endophenotypes of interest based on available neuroimaging studies of musical and language
processing, extracting genomic or gene expression data associated with functional activation
data (Kong et al., 2020; Mekki et al., 2022). For example, Mekki et al. (2022) defined a lan-
guage network based on an existing meta-analysis of fMRI-based language-related function,
then used the network to explore the genetic architecture of resting state functional connec-
tivity in the inferred language network, using individual genotypes from the UK Biobank
(Sudlow et al., 2015). Therefore, even though neuroimaging during a language task was
not conducted in UK Biobank participants, this triangulation among genes, brain, and behav-
ior, makes it possible to gain insights into the genetic architecture of the relevant circuitry
(although we do not yet know to what degree genetic associations with resting-state data
reflect those that would be seen with task-based functional activation data from the same
participants). Similarly, resources that focus on developmental trajectories, such as data from
the Adolescent Brain Cognitive Development study (ABCD; Volkow et al., 2018) can be har-
nessed to understand neural mechanisms of musicality and language traits as they unfold
during development.
In tandem, primary experimental research in the lab, with a focus on neuroimaging and
electrophysiological approaches, can directly investigate structural, functional, and develop-
mental correlates of musicality-language links at the intersection of specific traits of interest. In-
lab approaches allow for “deeper” measurement or phenotyping in smaller samples, which are
important for validating and qualifying findings from larger samples using the aforementioned
triangulation methods.
CONCLUSIONS
We introduced the Musical Abilities, Pleiotropy, Language, and Environment (MAPLE) frame-
work, which integrates genetic and environmental explanations for the many behavioral
associations between musicality and language skills. The MAPLE framework provides a
bird’s eye view of individual differences that work together to explain covariation of musicality
and language traits. Individual variation in shared genetic architecture between musicality and
language may result in variation at the level of neurobiological development, structure, and
function, along with effects on other biological systems potentially relevant for music and/or
language abilities. This in turn explains covariance in cognitive and neural measures of music
and language. Additionally, neurobiology and other biological systems supporting language
traits are thought to be influenced by genetic and environmental factors that also influence
musicality. We also introduced genetic approaches that can be leveraged in several ways
to investigate associations between music and language, including to understand how musi-
cality traits may function as risk factors or promotive factors for language development and
speech processing abilities. In particular, since genetic studies in this area are still nascent,
the behavioral and neural findings of interest described here help generate predictions that
can be tested using a variety of genetics approaches, as groundwork is established in the
genetics of musicality and language. Beyond fundamental discoveries of shared genetic archi-
tecture, incorporating genetic/genomic data into music cognition and language science
approaches can also help disentangle genetic and environmental factors underlying
musicality-language associations, and improve identification and intervention efforts for
language-related disorders through a richer understanding of musicality correlates of
language-related abilities.
Neurobiology of Language
653
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
As a basis for MAPLE’s predictions about shared genetic architecture, we synthesized pat-
terns of findings in the behavioral and neural evidence linking musicality and language skills.
By synthesizing the evidence for musicality-language links across multiple domains of
language-related traits (i.e., speech perception; reading-related skills; grammatical skills), this
literature review provides a timely overview as individual differences approaches gain traction
in their ability to shed light on common biological origins of musicality and language. The
predictions derived from the MAPLE framework can be tested by integrating behavioral
and/or neural methods with genetics approaches, and highlight the incredible potential of
multidisciplinary collaboration.
ACKNOWLEDGMENTS
The authors wish to acknowledge Russ Beebe and Navya Thakkar for their assistance with
illustrations; Jonathan Z. Liu and Gabija Zilinskaite for technical assistance; Nancy J. Cox
for thoughtful discussions about the framework; and Miriam D. Lense, Anna V. Kasdan, and
two anonymous reviewers for helpful feedback on earlier versions of the manuscript.
FUNDING INFORMATION
Reyna L. Gordon, National Science Foundation (https://dx.doi.org/10.13039/100000001),
Award ID: 1926794. Cyrille L. Magne, National Science Foundation (https://dx.doi.org/10
.13039/100000001), Award ID: 1926786. Reyna L. Gordon, National Institutes of Health
(https://dx.doi.org/10.13039/100000002), Award ID: R01DC016977. Reyna L. Gordon,
National Institutes of Health (https://dx.doi.org/10.13039/100000002), Award ID: DP2
HD098859. Simon E. Fisher, Max Planck Society (no award ID). The content is solely
the responsibility of the authors and does not necessarily represent the official views of the
funders.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
AUTHOR CONTRIBUTIONS
Srishti Nayak: Conceptualization: Supporting; Data curation: Equal; Formal analysis: Lead;
Investigation: Equal; Methodology: Equal; Project administration: Equal; Visualization: Sup-
porting; Writing – original draft: Lead; Writing – review & editing: Lead. Peyton L. Coleman:
Data curation: Equal, Formal analysis: Supporting; Investigation: Equal; Project administration:
Supporting; Software: Lead; Visualization: Lead; Writing – original draft: Supporting; Writing –
review & editing: Supporting. Enikő Ladányi: Formal analysis: Supporting; Methodology:
Supporting; Writing – original draft: Supporting; Writing – review & editing: Supporting.
Rachana Nitin: Formal analysis: Supporting, Methodology: Supporting; Writing – original
draft: Supporting; Writing – review & editing: Supporting. Daniel E. Gustavson: Formal
analysis: Supporting; Investigation: Supporting; Methodology: Supporting; Writing – original
draft: Supporting; Writing – review & editing: Supporting. Simon E. Fisher: Funding acquisi-
tion: Equal; Methodology: Supporting; Project administration: Equal; Writing – original draft:
Supporting; Writing – review & editing: Equal. Cyrille L. Magne: Formal analysis: Supporting;
Funding acquisition: Equal; Investigation: Supporting; Methodology: Supporting; Writing –
original draft: Supporting. Reyna L. Gordon: Conceptualization: Lead; Formal analysis: Equal;
Funding acquisition: Lead; Methodology: Equal; Project administration: Lead; Supervision:
Lead; Visualization: Supporting; Writing – original draft: Supporting; Writing – review &
editing: Equal.
Neurobiology of Language
654
Understanding musicality-language links
REFERENCES
Abrams, D. A., Bhatara, A., Ryali, S., Balaban, E., Levitin, D. J., &
Menon, V. (2011). Decoding temporal structure in music and
speech relies on shared brain resources but elicits different
fine-scale spatial patterns. Cerebral Cortex, 21(7), 1507–1518.
https://doi.org/10.1093/cercor/bhq198, PubMed: 21071617
Alcock, K. J., Passingham, R. E., Watkins, K., & Vargha-Khadem, F.
(2000). Pitch and timing abilities in inherited speech and lan-
guage impairment. Brain and Language, 75(1), 34–46. https://
doi.org/10.1006/brln.2000.2323, PubMed: 11023637
Andreola, C., Mascheretti, S., Belotti, R., Ogliari, A., Marino, C.,
Battaglia, M., & Scaini, S. (2021). The heritability of reading
and reading-related neurocognitive components: A multi-level
meta-analysis. Neuroscience and Biobehavioral Reviews, 121,
175–200. https://doi.org/10.1016/j.neubiorev.2020.11.016,
PubMed: 33246020
Antoniou, M., To, C. K. S., & Wong, P. C. M. (2015). Auditory cues
that drive language development are language specific: Evidence
from Cantonese. Applied Psycholinguistics, 36(6), 1493–1507.
https://doi.org/10.1017/S0142716414000514
Anvari, S. H., Trainor, L. J., Woodside, J., & Levy, B. A. (2002). Rela-
tions among musical skills, phonological processing, and early
reading ability in preschool children. Journal of Experimental
Child Psychology, 83(2), 111–130. https://doi.org/10.1016
/S0022-0965(02)00124-8, PubMed: 12408958
Asano, R., Boeckx, C., & Seifert, U. (2021). Hierarchical control as
a shared neurocognitive mechanism for language and music.
Cognition, 216, Article 104847. https://doi.org/10.1016/j
.cognition.2021.104847, PubMed: 34311153
Atherton, R. P., Chrobak, Q. M., Rauscher, F. H., Karst, A. T., Hanson,
M. D., Steinert, S. W., & Bowe, K. L. (2018). Shared processing of
language and music. Experimental Psychology, 65, 40–48. https://
doi.org/10.1027/1618-3169/a000388, PubMed: 29415645
Babajani-Feremi, A. (2017). Neural mechanism underling compre-
hension of narrative speech and its heritability: Study in a large
population. Brain Topography, 30(5), 592–609. https://doi.org/10
.1007/s10548-017-0550-6, PubMed: 28214981
Baker, D. W., Parker, R. M., Williams, M. V., Clark, W. S., & Nurss,
J. (1997). The relationship of patient reading ability to
self-reported health and use of health services. American Journal
of Public Health, 87(6), 1027–1030. https://doi.org/10.2105
/AJPH.87.6.1027, PubMed: 9224190
Bekius, A., Cope, T. E., & Grube, M. (2016). The beat to read: A
cross-lingual link between rhythmic regularity perception and
reading skill. Frontiers in Human Neuroscience, 10, Article 425.
https://doi.org/10.3389/fnhum.2016.00425, PubMed: 27630551
Besson, M., & Schön, D. (2012). Comparison between language
and music. In I. Peretz & R. Zatorre (Eds.), The cognitive neuro-
science of music (pp. 232–258). Oxford Scholarship Online.
https://doi.org/10.1093/acprof:oso/9780198525202.003.0018
Bethlehem, R. A. I., Seidlitz, J., White, S. R., Vogel, J. W., Anderson,
K. M., Adamson, C., Adler, S., Alexopoulos, G. S., Anagnostou,
E., Areces-Gonzalez, A., Astle, D. E., Auyeung, B., Ayub, M.,
Bae, J., Ball, G., Baron-Cohen, S., Beare, R., Bedford, S. A.,
Benegal, V., … Alexander-Bloch, A. F. (2022). Brain charts for
the human lifespan. Nature, 604, 525–533. https://doi.org/10
.1038/s41586-022-04554-y, PubMed: 35388223
Bidelman, G. M., & Alain, C. (2015). Musical training orchestrates
coordinated neuroplasticity in auditory brainstem and cortex to
counteract age-related declines in categorical vowel perception.
Journal of Neuroscience, 35(3), 1240–1249. https://doi.org/10
.1523/JNEUROSCI.3292-14.2015, PubMed: 25609638
Bishop, D. V. M. (2003). Test for reception of grammar: TROG-2:
Manual. Pearson.
Bishop, D. V. M., Adams, C. V., & Norbury, C. F. (2006). Distinct
genetic influences on grammar and phonological short-term
memory deficits: Evidence from 6-year-old twins. Genes, Brain
and Behavior, 5(2), 158–169. https://doi.org/10.1111/j.1601
-183X.2005.00148.x, PubMed: 16507007
Bishop, D. V. M., & Hayiou-Thomas, M. E. (2008). Heritability of
specific language impairment depends on diagnostic criteria.
Genes, Brain and Behavior, 7(3), 365–372. https://doi.org/10
.1111/j.1601-183X.2007.00360.x, PubMed: 17919296
Bishop, D. V. M., North, T., & Donlan, C. (1995). Genetic basis of
specific language impairment: Evidence from a twin study.
Developmental Medicine & Child Neurology, 37(1), 56–71.
https://doi.org/10.1111/j.1469-8749.1995.tb11932.x, PubMed:
7828787
Boll-Avetisyan, N., Bhatara, A., & Höhle, B. (2020). Processing of
rhythm in speech and music in adult dyslexia. Brain Sciences,
10(5), Article 261. https://doi.org/10.3390/ brainsci10050261,
PubMed: 32365799
Boorom, O., Nayak, S., Ladányi, E., Magne, C. L., & Gordon, R. L.
(2022). Music and developmental disorders of reading and
spoken language. PsyArXiv. https://psyarxiv.com/r5t4c
Borrie, S. A., Baese-Berk, M., Van Engen, K., & Bent, T. (2017). A
relationship between processing speech in noise and dysarthric
speech. The Journal of the Acoustical Society of America, 141(6),
4660–4667. https://doi.org/10.1121/1.4986746, PubMed:
28679257
Borrie, S. A., Lansford, K. L., & Barrett, T. S. (2018). Understanding
dysrhythmic speech: When rhythm does not matter and learning
does not happen. The Journal of the Acoustical Society of Amer-
ica, 143(5), EL379–EL385. https://doi.org/10.1121/1.5037620,
PubMed: 29857710
Brandt, A., Gebrian, M., & Slevc, L. R. (2012). Music and early lan-
guage acquisition. Frontiers in Psychology, 3, Article 327. https://
doi.org/10.3389/fpsyg.2012.00327, PubMed: 22973254
Brewer, C. C., Zalewski, C. K., King, K. A., Zobay, O., Riley, A.,
Ferguson, M. A., Bird, J. E., McCabe, M. M., Hood, L. J., Drayna,
D., Griffith, A. J., Morell, R. J., Friedman, T. B., & Moore, D. R.
(2016). Heritability of non-speech auditory processing skills.
European Journal of Human Genetics, 24(8), 1137–1144.
https://doi.org/10.1038/ejhg.2015.277, PubMed: 26883091
Brimo, D., Apel, K., & Fountain, T. (2017). Examining the contribu-
tions of syntactic awareness and syntactic knowledge to reading
comprehension. Journal of Research in Reading, 40(1), 57–74.
https://doi.org/10.1111/1467-9817.12050
Brod, G., & Opitz, B. (2012). Does it really matter? Separating the
effects of musical training on syntax acquisition. Frontiers in
Psychology, 3, Article 543. https://doi.org/10.3389/fpsyg.2012
.00543, PubMed: 23248608
Brown, S. (2017). A joint prosodic origin of language and music.
Frontiers in Psychology, 8, Article 1894. https://doi.org/10.3389
/fpsyg.2017.01894, PubMed: 29163276
Bulik-Sullivan, B., Finucane, H. K., Anttila, V., Gusev, A., Day, F. R.,
Loh, P.-R., ReProGen Consortium, Psychiatric Genomics
Consortium, Genetic Consortium for Anorexia Nervosa of the
Wellcome Trust Case Control Consortium 3, Duncan, L., Perry,
J. R. B., Patterson, N., Robinson, E. B., Daly, M. J., Price, A. L., &
Neale, B. M. (2015). An atlas of genetic correlations across human
diseases and traits. Nature Genetics, 47(11), 1236–1241. https://
doi.org/10.1038/ng.3406, PubMed: 26414676
Neurobiology of Language
655
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Burley, N. (1983). The meaning of assortative mating. Ethology and
Sociobiology, 4(4), 191–203. https://doi.org/10.1016/0162-3095
(83)90009-2
Butkovic, A., Ullén, F., & Mosing, M. A. (2015). Personality related
traits as predictors of music practice: Underlying environmental
and genetic influences. Personality and Individual Differences,
74, 133–138. https://doi.org/10.1016/j.paid.2014.10.006
Cannon, J. J., & Patel, A. D. (2021). How beat perception co-opts motor
neurophysiology. Trends in Cognitive Sciences, 25(2), 137–150.
https://doi.org/10.1016/j.tics.2020.11.002, PubMed: 33353800
Chan, A. S., Ho, Y. C., & Cheung, M. C. (1998). Music training
improves verbal memory. Nature, 396(6707), 128. https://doi
.org/10.1038/24075, PubMed: 9823892
Chen, X., Affourtit, J., Ryskin, R., Regev, T. I., Norman-Haignere, S.,
Jouravlev, O., Malik-Moraleda, S., Kean, H., Varley, R., &
Fedorenko, E. (2021). The human language system does not
support music processing. bioRxiv. https://doi.org/10.1101/2021
.06.01.446439
Cheverud, J. M. (1988). A comparison of genetic and phenotypic
correlations. Evolution, 42(5), 958–968. https://doi.org/10.1111
/j.1558-5646.1988.tb02514.x, PubMed: 28581166
Christiner, M., & Reiterer, S. M. (2018). Early influence of musical
abilities and working memory on speech imitation abilities:
Study with pre-school children. Brain Sciences, 8(9), Article 169.
https://doi.org/10.3390/brainsci8090169, PubMed: 30200479
Clayton, K. K., Swaminathan, J., Yazdanbakhsh, A., Zuk, J., Patel,
A. D., & Kidd, G. (2016). Executive function, visual attention
and the cocktail party problem in musicians and non-musicians.
PLOS ONE, 11(7), Article e0157638. https://doi.org/10.1371
/journal.pone.0157638, PubMed: 27384330
Clos, M., Amunts, K., Laird, A. R., Fox, P. T., & Eickhoff, S. B.
(2013). Tackling the multifunctional nature of Broca’s region
meta-analytically: Co-activation-based parcellation of area 44.
N e u ro I m a g e , 8 3 , 1 7 4 – 1 8 8 . h t t p s : / / d o i . o rg / 1 0 . 1 0 1 6 / j
.neuroimage.2013.06.041, PubMed: 23791915
Coffey, E. B. J., Mogilever, N. B., & Zatorre, R. J. (2017). Speech-in-
noise perception in musicians: A review. Hearing Research, 352,
49–69. https://doi.org/10.1016/j.heares.2017.02.006, PubMed:
28213134
Cohrdes, C., Grolig, L., & Schroeder, S. (2016). Relating language
and music skills in young children: A first approach to systemize
and compare distinct competencies on different levels. Frontiers
in Psychology, 7, Article 1616. https://doi.org/10.3389/fpsyg
.2016.01616, PubMed: 27826266
Colbran, L. L., Gamazon, E. R., Zhou, D., Evans, P., Cox, N. J., &
Capra, J. A. (2019). Inferred divergent gene regulation in archaic
hominins reveals potential phenotypic differences. Nature
Ecology & Evolution, 3(11), 1598–1606. https://doi.org/10.1038
/s41559-019-0996-x, PubMed: 31591491.
Corriveau, K. H., & Goswami, U. (2009). Rhythmic motor entrain-
ment in children with speech and language impairments:
Tapping to the beat. Cortex, 45(1), 119–130. https://doi.org/10
.1016/j.cortex.2007.09.008, PubMed: 19046744
Crouch, D. J., & Bodmer, W. F. (2020). Polygenic inheritance,
GWAS, polygenic risk scores, and the search for functional
variants. Proceedings of the National Academy of Sciences,
11 7 ( 3 2 ) , 1 8 9 2 4 –1 8 9 3 3 . h t t ps : / / d o i . o rg / 1 0 . 1 0 7 3 / pn a s
.2005634117, PubMed: 32753378
Cumming, R., Wilson, A., Leong, V., Colling, L. J., & Goswami, U.
(2015). Awareness of rhythm patterns in speech and music in
children with specific language impairments. Frontiers in Human
Neuroscience, 9, Article 672. https://doi.org/10.3389/fnhum
.2015.00672, PubMed: 26733848
Cuthbert, B. N. (2014). The RDoC framework: Facilitating transition
from ICD/DSM to dimensional approaches that integrate neuro-
science and psychopathology. World Psychiatry, 13(1), 28–35.
https://doi.org/10.1002/wps.20087, PubMed: 24497240
Dalboni da Rocha, J. L., Schneider, P., Benner, J., Santoro, R.,
Atanasova, T., Van De Ville, D., & Golestani, N. (2020). TASH:
Toolbox for the automated segmentation of Heschl’s gyrus.
Scientific Reports, 10(1), Article 3887. https://doi.org/10.1038
/s41598-020-60609-y, PubMed: 32127593
Dale, P. S., Dionne, G., Eley, T. C., & Plomin, R. (2000). Lexical and
grammatical development: A behavioural genetic perspective.
Journal of Child Language, 27(3), 619–642. https://doi.org/10
.1017/S0305000900004281, PubMed: 11089341
Dale, P. S., Rice, M. L., Rimfeld, K., & Hayiou-Thomas, M. E.
(2018). Grammar clinical marker yields substantial heritability
for language impairments in 16-year-old twins. Journal of
Speech, Language, and Hearing Research, 61(1), 66–78. https://
doi.org/10.1044/2017_JSLHR-L-16-0364, PubMed: 29305597
David, D., Wade-Woolley, L., Kirby, J. R., & Smithrim, K. (2007).
Rhythm and reading development in school-age children: A lon-
gitudinal study. Journal of Research in Reading, 30(2), 169–183.
https://doi.org/10.1111/j.1467-9817.2006.00323.x
Davies, G., Lam, M., Harris, S. E., Trampush, J. W., Luciano, M.,
Hill, W. D., Hagenaars, S. P., Ritchie, S. J., Marioni, R. E.,
Fawns-Ritchie, C., Liewald, D. C. M., Okely, J. A., Ahola-Olli,
A. V., Barnes, C. L. K., Bertram, L., Bis, J. C., Burdick, K. E.,
Christoforou, A., DeRosse, P., … Deary, I. J. (2018). Study of
300,486 individuals identifies 148 independent genetic loci
influencing general cognitive function. Nature Communications,
9(1), Article 2098. https://doi.org/10.1038/s41467-018-04362-x,
PubMed: 29844566
Decker, C. A., & Decker, J. R. 1997. Planning and administering
early childhood programs (6th ed.). Merrill Prentice Hall.
Degé, F., Kubicek, C., & Schwarzer, G. (2015). Associations
between musical abilities and precursors of reading in preschool
aged children. Frontiers in Psychology, 6, Article 1220. https://doi
.org/10.3389/fpsyg.2015.01220, PubMed: 26347687
Dellatolas, G., Watier, L., Le Normand, M.-T., Lubart, T., &
Chevrie-Muller, C. (2009). Rhythm reproduction in kindergar-
ten, reading performance at second grade, and developmental
dyslexia theories. Archives of Clinical Neuropsychology, 24(6),
555–563. https://doi.org/10.1093/arclin/acp044, PubMed:
19628461
Deriziotis, P., & Fisher, S. E. (2017). Speech and language: Translat-
ing the genome. Trends in Genetics, 33(9), 642–656. https://doi
.org/10.1016/j.tig.2017.07.002, PubMed: 28781152
DeThorne, L. S., Harlaar, N., Petrill, S. A., & Deater-Deckard, K.
(2012). Longitudinal stability in genetic effects on children’s con-
versational language productivity. Journal of Speech, Language,
and Hearing Research, 55(3), 739–753. https://doi.org/10.1044
/1092-4388(2011/11-0014), PubMed: 22232406
Douglas, S., & Willatts, P. (1994). The relationship between
musical ability and literacy skills. Journal of Research in Read-
ing, 17(2), 99–107. https://doi.org/10.1111/j.1467-9817.1994
.tb00057.x
Doust, C., Fontanillas, P., Eising, E., Gordon, S. D., Wang, Z.,
Alagöz, G., Molz, B., 23andMe Research Team, Quantitative
Trait Working Group of the GenLang Consortium, Pourcain, B.
S., Francks, C., Marioni, R. E., Zhao, J., Paracchini, S., Talcott,
J. B., Monaco, A. P., Stein, J. F., Gruen, J. R., Olson, R. K., …
Luciano, M. (2022). Discovery of 42 genome-wide significant loci
associated with dyslexia. Nature Genetics, 54(11), 1621–1629.
https://doi.org/10.1038/s41588-022-01192-y, PubMed: 36266505
Neurobiology of Language
656
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Drayna, D., Manichaikul, A., de Lange, M., Snieder, H., & Spector,
T. (2001). Genetic correlates of musical pitch recognition in
humans. Science, 291(5510), 1969–1972. https://doi.org/10
.1126/science.291.5510.1969, PubMed: 11239158
Eising, E., Mirza-Schreiber, N., de Zeeuw, E. L., Wang, C. A.,
Truong, D. T., Allegrini, A. G., Yang Shapland, C., Zhu, G., Wigg,
K. G., Gerritse, M. L., Molz, B., Alagöz, G., Gialluisi, A.,
Abbondanza, F., Rimfeld, K., van Donkelaar, M., Liao, Z., Jansen,
P. R., Andlauer, T. F. M., … Fisher, S. E. (2022). Genome-wide
analyses of individual differences in quantitatively assessed
reading- and language-related skills in up to 34,000 people.
Proceedings of the National Academy of Sciences, 119(35),
e2202764119. https://doi.org/10.1073/pnas.2202764119,
PubMed: 35998220
Evans, P. D., Mueller, K. L., Gamazon, E. R., Cox, N. J., & Tomblin,
J. B. (2015). A genome-wide sib-pair scan for quantitative lan-
guage traits reveals linkage to chromosomes 10 and 13. Genes,
Brain, and Behavior, 14(5), 387–397. https://doi.org/10.1111/gbb
.12223, PubMed: 25997078
Fava, E., Hull, R., Baumbauer, K., & Bortfeld, H. (2014). Hemody-
namic responses to speech and music in preverbal infants. Child
Neuropsychology, 20(4), 430–448. https://doi.org/10.1080
/09297049.2013.803524, PubMed: 23777481
Fedorenko, E., Patel, A., Casasanto, D., Winawer, J., & Gibson, E.
(2009). Structural integration in language and music: Evidence
for a shared system. Memory and Cognition, 37(1), 1–9. https://
doi.org/10.3758/MC.37.1.1, PubMed: 19103970
Fedorenko, E., & Varley, R. (2016). Language and thought are not
the same thing: Evidence from neuroimaging and neurological
patients. Annals of the New York Academy of Sciences,
1369(1), 132–153. https://doi.org/10.1111/nyas.13046,
PubMed: 27096882
Fernald, A., Taeschner, T., Dunn, J., Papousek, M., de Boysson-
Bardies, B., & Fukui, I. (1989). A cross-language study of pro-
sodic modifications in mothers’ and fathers’ speech to preverbal
infants. Journal of Child Language, 16(3), 477–501. https://doi
.org/10.1017/S0305000900010679, PubMed: 2808569
Fisher, S. E., & Scharff, C. (2009). FOXP2 as a molecular window
into speech and language. Trends in Genetics, 25(4), 166–177.
https://doi.org/10.1016/j.tig.2009.03.002, PubMed: 19304338
Fisher, S. E., & Vernes, S. C. (2015). Genetics and the language sci-
ences. Annual Review of Linguistics, 1, 289–310. https://doi.org
/10.1146/annurev-linguist-030514-125024
Fitch, W. T. (2017). Empirical approaches to the study of language
evolution. Psychonomic Bulletin and Review, 24(1), 3–33.
https://doi.org/10.3758/s13423-017-1236-5 , PubMed:
28150125
Fiveash, A., Bedoin, N., Gordon, R. L., & Tillmann, B. (2021). Pro-
cessing rhythm in speech and music: Shared mechanisms and
implications for developmental speech and language disorders.
Neuropsychology, 35(8), 771–791. https://doi.org/10.1037
/neu0000766, PubMed: 34435803
Flagge, A. G., Estis, J. M., & Moore, R. E. (2016). Short-term mem-
ory performance in 7- and 8-year-old children: The relationship
between phonological and pitch processing. Journal of Speech,
Language, and Hearing Research, 59(5), 1208–1217. https://doi
.org/10.1044/2016_JSLHR-H-14-0317, PubMed: 27682025
Flaugnacco, E., Lopez, L., Terribili, C., Zoia, S., Buda, S., Tilli, S.,
Monasta, L., Montico, M., Sila, A., Ronfani, L., & Schön, D.
(2014). Rhythm perception and production predict reading
abilities in developmental dyslexia. Frontiers in Human Neuro-
science, 8, Article 392. https://doi.org/10.3389/fnhum.2014
.00392, PubMed: 24926248
Foncubierta, J. M., Machancoses, F. H., Buyse, K., & Fonseca-Mora,
M. C. (2020). The acoustic dimension of reading: Does musical
aptitude affect silent reading fluency? Frontiers in Neuroscience,
14, Article 399. https://doi.org/10.3389/fnins.2020.00399,
PubMed: 32410955
Forgeard, M., Schlaug, G., Norton, A., Rosam, C., Iyengar, U., &
Winner, E. (2008). The relation between music and phonological
processing in normal-reading children and children with dys-
lexia. Music Perception, 25(4), 383–390. https://doi.org/10
.1525/mp.2008.25.4.383
Fotidzis, T. S., Moon, H., Steele, J. R., & Magne, C. L. (2018). Cross-
modal priming effect of rhythm on visual word recognition and
its relationships to music aptitude and reading achievement.
Brain Sciences, 8(12), Article 210. https://doi.org/10.3390
/brainsci8120210, PubMed: 30501073
François, C., Teixidó, M., Takerkart, S., Agut, T., Bosch, L., &
Rodriguez-Fornells, A. (2017). Enhanced neonatal brain
responses to sung streams predict vocabulary outcomes by age
18 months. Scientific Reports, 7(1), Article 12451. https://doi
.org/10.1038/s41598-017-12798-2, PubMed: 28963569
Friedman, N. P., Miyake, A., Young, S. E., DeFries, J. C., Corley,
R. P., & Hewitt, J. K. (2008). Individual differences in executive
functions are almost entirely genetic in origin. Journal of Experi-
mental Psychology: General, 137(2), 201–225. https://doi.org/10
.1037/0096-3445.137.2.201, PubMed: 18473654
Fujiki, M., Brinton, B., Morgan, M., & Hart, C. H. (1999).
Withdrawn and sociable behavior of children with language
impairment. Language, Speech, and Hearing Services in Schools,
30(2), 183–195. https://doi.org/10.1044/0161-1461.3002.183,
PubMed: 27764280
Gamazon, E. R., Wheeler, H. E., Shah, K. P., Mozaffari, S. V.,
Aquino-Michaels, K., Carroll, R. J., Eyler, A. E., Denny, J. C., GTEx
Consortium, Nicolae, D. L., Cox, N. J., & Im, H. K. (2015). A
gene-based association method for mapping traits using reference
transcriptome data. Nature Genetics, 47(9), 1091–1098. https://
doi.org/10.1038/ng.3367, PubMed: 26258848
Gamazon, E. R., Zwinderman, A. H., Cox, N. J., Denys, D., &
Derks, E. M. (2019). Multi-tissue transcriptome analyses identify
genetic mechanisms underlying neuropsychiatric traits. Nature
Genetics, 51(6), 933–940. https://doi.org/10.1038/s41588-019
-0409-8, PubMed: 31086352
George, E. M., & Coch, D. (2011). Music training and working
memory: An ERP study. Neuropsychologia, 49(5), 1083–1094.
https://doi.org/10.1016/j.neuropsychologia.2011.02.001,
PubMed: 21315092
Gialluisi, A., Andlauer, T. F. M., Mirza-Schreiber, N., Moll, K.,
Becker, J., Hoffmann, P., Ludwig, K. U., Czamara, D., Pourcain,
B. S., Honbolygó, F., Tóth, D., Csépe, V., Huguet, G., Chaix, Y.,
Iannuzzi, S., Demonet, J. F., Morris, A. P., Hulslander, J., Willcutt,
E. G., … Schulte-Körne, G. (2021). Genome-wide association
study reveals new insights into the heritability and genetic corre-
lates of developmental dyslexia. Molecular Psychiatry, 26(7),
3004–3017. https://doi.org/10.1038/s41380-020-00898-x,
PubMed: 33057169
Goldman, R., & Fristoe, M. (2000). Goldman-Fristoe test of articu-
lation (2nd ed.). American Guidance Service. https://doi.org/10
.1037/t15098-000
González-Trujillo, M. C., Defior, S., & Gutiérrez-Palma, N. (2014).
The role of nonspeech rhythm in Spanish word reading. Journal
of Research in Reading, 37(3), 316–330. https://doi.org/10.1111/j
.1467-9817.2012.01529.x
Gooch, D., Thompson, P., Nash, H. M., Snowling, M. J., & Hulme,
C. (2016). The development of executive function and language
Neurobiology of Language
657
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
skills in the early school years. The Journal of Child Psychology
and Psychiatry, 57(2), 180–187. https://doi.org/10.1111/jcpp
.12458, PubMed: 26307609
Gordon, R. L., Fehd, H. M., & McCandliss, B. D. (2015). Does
music training enhance literacy skills? A meta-analysis. Frontiers
in Psychology, 6, Article 1777. https://doi.org/10.3389/fpsyg
.2015.01777, PubMed: 26648880
Gordon, R. L., Jacobs, M. S., Schuele, C. M. & McAuley, J. D.
(2015). Perspectives on the rhythm-grammar link and its implica-
tions for typical and atypical language development. Annals of
the New York Academy of Sciences, 1337, 16–25. https://doi
.org/10.1111/nyas.12683, PubMed: 25773612
Gordon, R. L., & Magne, C. L. (2017). Music and the brain: Music
and cognitive abilities. In The Routledge companion to music
cognition (pp. 49–61). Routledge. https://doi.org/10.4324
/9781315194738-5
Gordon, R. L., Ravignani, A., Hyland Bruno, J., Robinson, C. M.,
Scartozzi, A., Embalabala, R., Niarchou, M., 23andMe Research
Team, Cox, N. J., & Creanza, N. (2021). Linking the genomic
signatures of human beat synchronization and learned song
in birds. Philosophical Transactions of the Royal Society B,
376(1835), Article 20200329. https://doi.org/10.1098/rstb.2020
.0329, PubMed: 34420388
Gordon, R. L., Shivers, C. M., Wieland, E. A., Kotz, S. A., Yoder,
P. J., & Devin McAuley, J. (2015). Musical rhythm discrimination
explains individual differences in grammar skills in children.
Developmental Science, 18(4), 635–644. https://doi.org/10
.1111/desc.12230, PubMed: 25195623
Goswami, U. (2011). A temporal sampling framework for develop-
mental dyslexia. Trends in Cognitive Sciences, 15(1), 3–10.
https://doi.org/10.1016/j.tics.2010.10.001, PubMed: 21093350
Gottesman, I. I., & Gould, T. D. (2003). The endophenotype con-
cept in psychiatry: Etymology and strategic intentions. American
Journal of Psychiatry, 160(4), 636–645. https://doi.org/10.1176
/appi.ajp.160.4.636, PubMed: 12668349
Gottesman, I. I., & Shields, J. (1972). Schizophrenia and genetics: A
twin study vantage point. Academic Press.
Graham, S. A., & Fisher, S. E. (2013). Decoding the genetics of speech
and language. Current Opinion in Neurobiology, 23(1), 43–51.
https://doi.org/10.1016/j.conb.2012.11.006, PubMed: 23228431
Grau-Sánchez, J., Münte, T. F., Altenmüller, E., Duarte, E., &
Rodríguez-Fornells, A. (2020). Potential benefits of music playing
in stroke upper limb motor rehabilitation. Neuroscience & Bio-
behavioral Reviews, 112, 585–599. https://doi.org/10.1016/j
.neubiorev.2020.02.027, PubMed: 32092314
Grotzinger, A. D., Rhemtulla, M., de Vlaming, R., Ritchie, S. J.,
Mallard, T. T., Hill, W. D., Ip, H. F., Marioni, R. E., McIntosh,
A. M., Deary, I. J., Koellinger, P. D., Harden, K. P., Nivard,
M. G., & Tucker-Drob, E. M. (2019). Genomic structural equa-
tion modelling provides insights into the multivariate genetic
architecture of complex traits. Nature Human Behaviour, 3(5),
513–525. https://doi.org/10.1038/s41562-019-0566-x, PubMed:
30962613
Grube, M., Kumar, S., Cooper, F. E., Turton, S., & Griffiths, T. D.
(2012). Auditory sequence analysis and phonological skill.
Proceedings of the Royal Society B: Biological Sciences,
279(1746), 4496–4504. https://doi.org/10.1098/rspb.2012
.1817, PubMed: 22951739
Gustavson, D. E., Friedman, N. P., Fontanillas, P., Elson, S. L.,
Palmer, A. A., & Sanchez-Roige, S. (2020). The latent genetic
structure of impulsivity and its relation to internalizing psychopa-
thology. Psychological Science, 31(8), 1025–1035. https://doi.org
/10.1177/0956797620938160, PubMed: 32716714
Gustavson, D. E., Friedman, N. P., Stallings, M. C., Reynolds, C. A.,
Coon, H., Corley, R. P., Hewitt, J. K., & Gordon, R. L. (2021).
Musical instrument engagement in adolescence predicts verbal
ability 4 years later: A twin and adoption study. Developmental
Psychology, 57(11), 1943–1957. https://doi.org/10.1037
/dev0001245, PubMed: 34914455
Gustavson, D. E., Panizzon, M. S., Franz, C. E., Reynolds, C. A.,
Corley, R. P., Hewitt, J. K., Lyons, M. J., Kremen, W. S., &
Friedman, N. P. (2019). Integrating verbal fluency with executive
functions: Evidence from twin studies in adolescence and middle
age. Journal of Experimental Psychology: General, 148(12),
2104–2119. https://doi.org/10.1037/xge0000589, PubMed:
30896200
Hambrick, D. Z., & Tucker-Drob, E. M. (2014). The genetics of
music accomplishment: Evidence for gene–environment
correlation and interaction. Psychonomic Bulletin and Review,
22(1), 112–120. https://doi.org/10.3758/s13423-014-0671-9,
PubMed: 24957535
Hart, S. A., Little, C., & van Bergen, E. (2021). Nurture might be
nature: Cautionary tales and proposed solutions. NPJ Science
of Learning, 6(1), Article 2. https://doi.org/10.1038/s41539-020
-00079-z, PubMed: 33420086
Hausen, M., Torppa, R., Salmela, V. R., Vainio, M., & Särkämö, T.
(2013). Music and speech prosody: A common rhythm. Frontiers
in Psychology, 4, Article 566. https://doi.org/10.3389/fpsyg.2013
.00566, PubMed: 24032022
Hayiou-Thomas, M. E. (2008). Genetic and environmental influ-
ences on early speech, language and literacy development.
Journal of Communication Disorders, 41(5), 397–408. https://
doi.org/10.1016/j.jcomdis.2008.03.002, PubMed: 18538338
Heard, M., & Lee, Y. S. (2020). Shared neural resources of rhythm
and syntax: An ALE meta-analysis. Neuropsychologia, 137,
Article 107284. https://doi.org/10.1016/j.neuropsychologia
.2019.107284, PubMed: 31783081
Heath, A. C., Kessler, R. C., Neale, M. C., Hewitt, J. K., Eaves, L. J.,
& Kendler, K. S. (1993). Testing hypotheses about direction of
causation using cross-sectional family data. Behavior Genetics,
23(1), 29–50. https://doi.org/10.1007/ BF01067552, PubMed:
8476389
Hedge, C., Powell, G., & Sumner, P. (2018). The reliability paradox:
Why robust cognitive tasks do not produce reliable individual
differences. Behavior Research Methods, 50(3), 1166–1186.
https://doi.org/10.3758/s13428-017-0935-1, PubMed:
28726177
Höhle, B., Pauen, S., Hesse, V., & Weissenborn, J. (2014). Discrimi-
nation of rhythmic pattern at 4 months and language performance
at 5 years: A longitudinal analysis of data from German-learning
children. Language Learning, 64(s2), 141–164. https://doi.org/10
.1111/lang.12075
Holliman, A. J., Wood, C., & Sheehy, K. (2010). The contribution of
sensitivity to speech rhythm and non-speech rhythm to early
reading development. Educational Psychology, 30(3), 247–267.
https://doi.org/10.1080/01443410903560922
Honing, H. (2018). On the biological basis of musicality. Annals of
the New York Academy of Sciences, 1423(1), 51–56. https://doi
.org/10.1111/nyas.13638, PubMed: 29542134
Huang, W.-T., Nan, Y., Dong, Q., & Liu, C. (2015). Just-noticeable
difference of tone pitch contour change for Mandarin congenital
amusics. The Journal of the Acoustical Society of America,
138(1), EL99–EL104. https://doi.org/10.1121/1.4923268,
PubMed: 26233070
Huss, M., Verney, J. P., Fosker, T., Mead, N., & Goswami, U. (2011).
Music, rhythm, rise time perception and developmental dyslexia:
Neurobiology of Language
658
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Perception of musical meter predicts reading and phonology.
Cortex, 47(6), 674–689. https://doi.org/10.1016/j.cortex.2010
.07.010, PubMed: 20843509
Hutchins, S. (2018). Early childhood music training and associated
improvements in music and language abilities. Music Perception,
35(5), 579–593. https://doi.org/10.1525/mp.2018.35.5.579
Jackendoff, R. (2009). Parallels and nonparallels between language
and music. Music Perception, 26(3), 195–204. https://doi.org/10
.1525/mp.2009.26.3.195
James, C. E., Altenmüller, E., Kliegel, M., Krüger, T. H. C., Van De
Ville, D., Worschech, F., Abdili, L., Scholz, D. S., Jünemann, K.,
Hering, A., Grouiller, F., Sinke, C., & Marie, D. (2020). Train the
brain with music (TBM): Brain plasticity and cognitive benefits
induced by musical training in elderly people in Germany and
Switzerland, a study protocol for an RCT comparing musical
instrumental practice to sensitization to music. BMC Geriatrics,
20(1), Article 418. https://doi.org/10.1186/s12877-020-01761-y,
PubMed: 33087078
Jantzen, M. G., Large, E. W., & Magne, C. (2016). Editorial: Overlap
of neural systems for processing language and music. Frontiers in
Psychology, 7, Article 876. https://doi.org/10.3389/fpsyg.2016
.00876, PubMed: 27378976
Kaiser, J. (2016, February 25). NIH’s 1-million-volunteer precision
medicine study announces first pilot projects. Science Insider.
https://www.science.org/content/article/nih-s-1-million-volunteer
-precision-medicine-study-announces-first-pilot-projects. https://
doi.org/10.1126/science.aaf4108
Kalashnikova, M., Goswami, U., & Burnham, D. (2019). Sensitivity
to amplitude envelope rise time in infancy and vocabulary devel-
opment at 3 years: A significant relationship. Developmental Sci-
ence, 22(6), Article e12836. https://doi.org/10.1111/desc.12836,
PubMed: 31004544
Kapa, L. L., & Erikson, J. A. (2020). The relationship between word
learning and executive function in preschoolers with and without
developmental language disorder. Journal of Speech, Language,
and Hearing Research, 63(7), 2293–2307. https://doi.org/10
.1044/2020_JSLHR-19-00342, PubMed: 32546080
Kasdan, A. V., Burgess, A. N., Pizzagalli, F., Scartozzi, A., Chern,
A., Kotz, S. A., Wilson, S. M., & Gordon, R. L. (2022). Identifying
a brain network for musical rhythm: A functional neuroimaging
meta-analysis and systematic review. Neuroscience & Biobehav-
ioral Reviews, 136, Article 104588. https://doi.org/10.1016/j
.neubiorev.2022.104588, PubMed: 35259422
Kasdan, A. V., Gordon, R. L., & Lense, M. D. (2022). Neurophysi-
ological correlates of dynamic beat tracking in individuals with
Williams syndrome. Biological Psychiatry: Cognitive Neurosci-
ence and Neuroimaging, 7(11), 1183–1191. https://doi.org/10
.1016/j.bpsc.2020.10.003, PubMed: 33419711
Kong, X. Z., Tzourio-Mazoyer, N., Joliot, M., Fedorenko, E., Liu, J.,
Fisher, S. E., & Francks, C. (2020). Gene expression correlates of
the cortical network underlying sentence processing. Neurobiol-
ogy of Language, 1(1), 77–103. https://doi.org/10.1162/nol_a
_00004
Kotler, J., Mehr, S. A., Egner, A., Haig, D., & Krasnow, M. M. (2019).
Response to vocal music in Angelman syndrome contrasts with
Prader-Willi syndrome. Evolution and Human Behavior, 40(5),
420–426. https://doi.org/10.1016/j.evolhumbehav.2019.05.003,
PubMed: 32655274
Kotov, R., Waszczuk, M. A., Krueger, R. F., Forbes, M. K., Watson,
D., Clark, L. A., Achenbach, T. M., Althoff, R. R., Ivanova, M. Y.,
Michael Bagby, R., Brown, T. A., Carpenter, W. T., Caspi, A.,
Moffitt, T. E., Eaton, N. R., Forbush, K. T., Goldberg, D., Hasin,
D., Hyman, S. E., … Zimmerman, M. (2017). The hierarchical
taxonomy of psychopathology (HiTOP): A dimensional alterna-
tive to traditional nosologies. Journal of Abnormal Psychology,
126(4), 454–477. https://doi.org/10.1037/abn0000258,
PubMed: 28333488
Kotz, S. A., Ravignani, A., & Fitch, W. T. (2018). The evolution
of rhythm processing. Trends in Cognitive Sciences, 22(10),
896–910. https://doi.org/10.1016/j.tics.2018.08.002, PubMed:
30266149
Kotz, S. A., & Schwartze, M. (2010). Cortical speech processing
unplugged: A timely subcortico-cortical framework. Trends in
Cognitive Sciences, 14(9), 392–399. https://doi.org/10.1016/j
.tics.2010.06.005, PubMed: 20655802
Kovas, Y., Voronin, I., Kaydalov, A., Malykh, S. B., Dale, P. S., &
Plomin, R. (2013). Literacy and numeracy are more heritable than
intelligence in primary school. Psychological Science, 24(10),
2048–2056. https://doi.org/10.1177/0956797613486982,
PubMed: 24002885
Krapohl, E., Patel, H., Newhouse, S., Curtis, C. J., von Stumm, S.,
Dale, P. S., Zabaneh, D., Breen, G., O’Reilly, P. F., & Plomin, R.
(2018). Multi-polygenic score approach to trait prediction.
Molecular Psychiatry, 23(5), 1368–1374. https://doi.org/10
.1038/mp.2017.163, PubMed: 28785111
Kraus, N., & Chandrasekaran, B. (2010). Music training for the
development of auditory skills. Nature Reviews Neuroscience,
11(8), 599–605. https://doi.org/10.1038/nrn2882, PubMed:
20648064
Kunert, R., Willems, R. M., Casasanto, D., Patel, A. D., & Hagoort,
P. (2015). Music and language syntax interact in Broca’s area: An
fMRI study. PLOS ONE, 10(11), Article e0141069. https://doi.org
/10.1371/journal.pone.0141069, PubMed: 26536026
Ladányi, E., Persici, V., Fiveash, A., Tillmann, B., & Gordon, R. L.
(2020). Is atypical rhythm a risk factor for developmental speech
and language disorders? WIREs Cognitive Science, 11(5), Article
e1528. https://doi.org/10.1002/wcs.1528, PubMed: 32244259
Lagrois, M. É., Palmer, C., & Peretz, I. (2019). Poor synchronization
to musical beat generalizes to speech. Brain Sciences, 9(7),
Article 157. https://doi.org/10.3390/brainsci9070157, PubMed:
31277417
Lê, M., Quémart, P., Potocki, A., Gimenes, M., Chesnet, D., &
Lambert, E. (2020). Rhythm in the blood: The influence of
rhythm skills on literacy development in third graders. Journal
of Experimental Child Psychology, 198, Article 104880. https://
doi.org/10.1016/j.jecp.2020.104880, PubMed: 32622068
Lee, P. H., Anttila, V., Won, H., Feng, Y. C. A., Rosenthal, J., Zhu, Z.,
Tucker-Drob, E. M., Nivard, M. G., Grotzinger, A. D., Posthuma,
D., Wang, M. M. J., Yu, D., Stahl, E. A., Walters, R. K., Anney,
R. J. L., Duncan, L. E., Ge, T., Adolfsson, R., Banaschewski, T.,
… Smoller, J. W. (2019). Genomic relationships, novel loci, and
pleiotropic mechanisms across eight psychiatric disorders. Cell,
179(7), 1469–1482. https://doi.org/10.1016/j.cell.2019.11.020,
PubMed: 31835028
Lee, Y. S., Ahn, S., Holt, R. F., & Schellenberg, E. G. (2020). Rhythm
and syntax processing in school-age children. Developmental
Psychology, 56(9), 1632–1641. https://doi.org/10.1037
/dev0000969, PubMed: 32700950
Leggieri, M., Thaut, M. H., Fornazzari, L., Schweizer, T. A., Barfett,
J., Munoz, D. G., & Fischer, C. E. (2019). Music intervention
approaches for Alzheimer’s disease: A review of the literature.
Frontiers in Neuroscience, 13, Article 132. https://doi.org/10
.3389/fnins.2019.00132, PubMed: 30930728
Lense, M. D., Ladányi, E., Rabinowitch, T. C., Trainor, L., &
Gordon, R. (2021). Rhythm and timing as vulnerabilities in neu-
rodevelopmental disorders. Philosophical Transactions of the
Neurobiology of Language
659
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Royal Society B, 376(1835), Article 20200327. https://doi.org/10
.1098/rstb.2020.0327, PubMed: 34420385
Leong, V., Kalashnikova, M., Burnham, D., & Goswami, U.
(2017). The temporal modulation structure of infant-directed
speech. Open Mind, 1(2), 78–90. https://doi.org/10.1162
/OPMI_a_00008
Leppänen, P. H. T., Hämäläinen, J. A., Salminen, H. K., Eklund,
K. M., Guttorm, T. K., Lohvansuu, K., Puolakanaho, A., &
Lyytinen, H. (2010). Newborn brain event-related potentials
revealing atypical processing of sound frequency and the
subsequent association with later literacy skills in children with
familial dyslexia. Cortex, 46(10), 1362–1376. https://doi.org/10
.1016/j.cortex.2010.06.003, PubMed: 20656284
Lewis, B. A., Freebairn, L., Tag, J., Benchek, P., Morris, N. J.,
Iyengar, S. K., Gerry Taylor, H., & Stein, C. M. (2018). Heritability
and longitudinal outcomes of spelling skills in individuals with
histories of early speech and language disorders. Learning and
Individual Differences, 65, 1–11. https://doi.org/10.1016/j.lindif
.2018.05.001, PubMed: 30555216
Loui, P., Kroog, K., Zuk, J., Winner, E., & Schlaug, G. (2011). Relat-
ing pitch awareness to phonemic awareness in children: Implica-
tions for tone-deafness and dyslexia. Frontiers in Psychology, 2,
Article 111. https://doi.org/10.3389/fpsyg.2011.00111, PubMed:
21687467
Lundetræ, K., & Thomson, J. M. (2018). Rhythm production at
school entry as a predictor of poor reading and spelling at the
end of first grade. Reading and Writing, 31(1), 215–237. https://
doi.org/10.1007/s11145-017-9782-9, PubMed: 29367807
Magne, C., Jordan, D. K., & Gordon, R. L. (2016). Speech rhythm
sensitivity and musical aptitude: ERPs and individual differences.
Brain and Language, 153–154, 13–19. https://doi.org/10.1016/j
.bandl.2016.01.001, PubMed: 26828758
Magne, C., Schön, D., & Besson, M. (2006). Musician children
detect pitch violations in both music and language better than
nonmusician children: Behavioral and electrophysiological
approaches. Journal of Cognitive Neuroscience, 18(2), 199–211.
https://doi.org/10.1162/jocn.2006.18.2.199, PubMed: 16494681
Mankel, K., Barber, J., & Bidelman, G. M. (2020). Auditory categor-
ical processing for speech is modulated by inherent musical
listening skills. NeuroReport, 31(2), 162–166. https://doi.org/10
.1097/ WNR.0000000000001369, PubMed: 31834142
Mankel, K., & Bidelman, G. M. (2018). Inherent auditory skills
rather than formal music training shape the neural encoding of
speech. Proceedings of the National Academy of Sciences of
the United States of America, 115(51), 13129–13134. https://
doi.org/10.1073/pnas.1811793115, PubMed: 30509989
Marie, C., Magne, C., & Besson, M. (2011). Musicians and the
metric structure of words. Journal of Cognitive Neuroscience,
23(2), 294–305. https://doi.org/10.1162/jocn.2010.21413,
PubMed: 20044890
Martin, A., Schurz, M., Kronbichler, M., & Richlan, F. (2015). Read-
ing in the brain of children and adults: A meta-analysis of 40
functional magnetic resonance imaging studies. Human Brain
Mapping, 36(5), 1963–1981. https://doi.org/10.1002/ hbm
.22749, PubMed: 25628041
Mehr, S., Krasnow, M., Bryant, G., & Hagen, E. (2021). Origins of
music in credible signaling. Behavioral and Brain Sciences, 44,
Article E60. https://doi.org/10.1017/S0140525X20000345,
PubMed: 32843107
Mehr, S. A., Kotler, J., Howard, R. M., Haig, D., & Krasnow, M. M.
(2017). Genomic imprinting is implicated in the psychology of
music. Psychological Science, 28(10), 1455–1467. https://doi
.org/10.1177/0956797617711456, PubMed: 28857689
Mekki, Y., Guillemot, V., Lemaître, H., Carrión-Castillo, A., Forkel,
S., Frouin, V., & Philippe, C. (2022). The genetic architecture of
language functional connectivity. NeuroImage, 249, Article
118795. https://doi.org/10.1016/j.neuroimage.2021.118795,
PubMed: 34929384
Merchant, H., Grahn, J., Trainor, L., Rohrmeier, M., & Fitch, W. T.
(2015). Finding the beat: A neural perspective across humans
and non-human primates. Philosophical Transactions of the Royal
Society B: Biological Sciences, 370(1664), Article 20140093.
https://doi.org/10.1098/rstb.2014.0093, PubMed: 25646516
Merrett, D. L., Peretz, I., & Wilson, S. J. (2013). Moderating vari-
ables of music training induced neuroplasticity: A review and
discussion. Frontiers in Psychology, 4, Article 606. https://doi
.org/10.3389/fpsyg.2013.00606, PubMed: 24058353
Miller, J. A., Ding, S. L., Sunkin, S. M., Smith, K. A., Ng, L., Szafer,
A., Ebbert, A., Riley, Z. L., Royall, J. J., Aiona, K., Arnold, J. M.,
Bennet, C., Bertagnolli, D., Brouner, K., Butler, S., Caldejon, S.,
Carey, A., Cuhaciyan, C., Dalley, R. A., … Lein, E. S. (2014).
Transcriptional landscape of the prenatal human brain. Nature,
508(7495), 199–206. https://doi.org/10.1038/nature13185,
PubMed: 24695229
Milovanov, R., & Tervaniemi, M. (2011). The interplay between
musical and linguistic aptitudes: A review. Frontiers in Psychology,
2, Article 321. https://doi.org/10.3389/fpsyg.2011.00321,
PubMed: 22125541
Mithen, S. (Ed.). (2005). Creativity in human evolution and prehis-
tory. Routledge. https://doi.org/10.4324/9780203978627
Molino, J. (2000). Toward an evolutionary theory of music and lan-
guage. In N. L. Wallin, B. Merker, & S. Brown (Eds.), The origins
of music (pp. 165–176). MIT Press.
Moreno, S. (2009). Can music influence language and cognition?
Contemporary Music Review, 28(3), 329–345. https://doi.org/10
.1080/07494460903404410
Moreno, S., Bialystok, E., Barac, R., Schellenberg, E. G., Cepeda,
N. J., & Chau, T. (2011). Short-term music training enhances verbal
intelligence and executive function. Psychological Science, 22(11),
1425–1433. https://doi.org/10.1177/0956797611416999,
PubMed: 21969312
Moreno-Morales, C., Calero, R., Moreno-Morales, P., & Pintado, C.
(2020). Music therapy in the treatment of dementia: A systematic
review and meta-analysis. Frontiers in Medicine, 7, Article 160.
https://doi.org/10.3389/fmed.2020.00160, PubMed: 32509790
Moritz, C., Yampolsky, S., Papadelis, G., Thomson, J., & Wolf, M.
(2013). Links between early rhythm skills, musical training, and
phonological awareness. Reading and Writing, 26(5), 739–769.
https://doi.org/10.1007/s11145-012-9389-0
Morley, I. (2013). The prehistory of music: Human evolution,
archaeology, and the origins of musicality. Oxford University
Press. https://doi.org/10.1093/acprof:osobl/9780199234080.001
.0001
Morrill, T. H., Devin McAuley, J., Dilley, L. C., & Hambrick, D. Z.
(2015). Individual differences in the perception of melodic
contours and pitch-accent timing in speech: Support for
domain-generality of pitch processing. Journal of Experimental
Psychology: General, 144(4), 730–736. https://doi.org/10.1037
/xge0000081, PubMed: 26214165
Mosing, M. A., Madison, G., Pedersen, N. L., Kuja-Halkola, R., &
Ullén, F. (2014). Practice does not make perfect: No causal effect
of music practice on music ability. Psychological Science, 25(9),
1795–1803. https://doi.org/10.1177/0956797614541990,
PubMed: 25079217
Mosing, M. A., Verweij, K. J. H., Madison, G., & Ullén, F. (2016).
The genetic architecture of correlations between perceptual
Neurobiology of Language
660
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
timing, motor timing, and intelligence. Intelligence, 57, 33–40.
https://doi.org/10.1016/j.intell.2016.04.002
Münte, T., Altenmüller, E., & Jäncke, L. (2002). The musician’s brain
as a model of neuroplasticity. Nature Reviews Neuroscience, 3(6),
473–478. https://doi.org/10.1038/nrn843, PubMed: 12042882
Nair, P. S., Raijas, P., Ahvenainen, M., Philips, A. K., Ukkola-Vuoti,
L., & Järvelä, I. (2021). Music-listening regulates human micro-
RNA expression. Epigenetics, 16(5), 554–566. https://doi.org/10
.1080/15592294.2020.1809853, PubMed: 32867562
Nan, Y., Sun, Y., & Peretz, I. (2010). Congenital amusia in speakers
of a tone language: Association with lexical tone agnosia. Brain,
133(9), 2635–2642. https://doi.org/10.1093/ brain/awq178,
PubMed: 20685803
National Center for Education Statistics. (2019). Data point:
Adult literacy in the United States. Institute of Education
Sciences, U.S. Department of Education. https://nces.ed.gov
/pubs2019/2019179/index.asp
Newcomer, P., & Hammill, D. (1997). Test of oral language devel-
opment: Primary (3rd ed.). Pro-Ed.
Niarchou, M., Gustavson, D. E., Sathirapongsasuti, J. F., Anglada-
Tort, M., Eising, E., Bell, E., McArthur, E., Straub, P., 23andMe
Research Team, McAuley, J. D., Capra, J. A., Ullén, F., Creanza,
N., Mosing, M. A., Hinds, D. A., Davis, L. K., Jacoby, N., &
Gordon, R. L. (2022). Genome-wide association study of musical
beat synchronization demonstrates high polygenicity. Nature
Human Behaviour, 6, 1292–1309. https://doi.org/10.1038
/s41562-022-01359-x, PubMed: 35710621
Nicholls, H. L., John, C. R., Watson, D. S., Munroe, P. B., Barnes,
M. R., & Cabrera, C. P. (2020). Reaching the end-game for
GWAS: Machine learning approaches for the prioritization of
complex disease loci. Frontiers in Genetics, 11, Article 350.
https://doi.org/10.3389/fgene.2020.00350, PubMed: 32351543
Nitin, R., Gustavson, D. E., Aaron, A., Boorom, O. A., Bush, C. T.,
Wiens, N., Vaughan, C., Persici, V., Blain, S. D., Hambrick, D. Z.,
Camarata, S. C., McAuley, J. D., Gordon, R. L. (in press). Explor-
ing individual differences in musical rhythm and grammar skills
in school-aged children with typically developing language.
Scientific Reports.
Norman-Haignere, S. V., Feather, J., Boebinger, D., Brunner, P.,
Ritaccio, A., McDermott, J. H., Schalk, G., & Kanwisher, N.
(2022). A neural population selective for song in human auditory
cortex. Current Biology, 32(7), 1470–1484. https://doi.org/10
.1016/j.cub.2022.01.069, PubMed: 35196507
Overy, K. (2003). Dyslexia and music: From timing deficits to musi-
cal intervention. Annals of the New York Academy of Sciences,
999, 497–505. https://doi.org/10.1196/annals.1284.060,
PubMed: 14681173
Ozernov-Palchik, O., Wolf, M., & Patel, A. D. (2018). Relationships
between early literacy and nonlinguistic rhythmic processes in
kindergarteners. Journal of Experimental Child Psychology, 167,
354–368. https://doi.org/10.1016/j.jecp.2017.11.009, PubMed:
29227852
Patel, A. D. (2003). Rhythm in language and music: Parallels and
differences. Annals of the New York Academy of Sciences, 999,
140–143. https://doi.org/10.1196/annals.1284.015, PubMed:
14681127
Patel, A. D. (2008). Music as a transformative technology of the
mind. Cambridge University Press.
Patel, A. D. (2012). Language, music, and the brain: A resource-
sharing framework. In P. Rebuschat, M. Rohrmeier, I. Cross, &
J. A. Hawkins (Eds.), Language and music as cognitive systems
(pp. 204–223). Oxford University Press. https://doi.org/10.1093
/acprof:oso/9780199553426.003.0022
Patel, A. D. (2014). Can nonlinguistic musical training change the
way the brain processes speech? The expanded OPERA hypoth-
esis. Hearing Research, 308, 98–108. https://doi.org/10.1016/j
.heares.2013.08.011, PubMed: 24055761
Patel, A. D. (2021). Vocal learning as a preadaptation for the
evolution of human beat perception and synchronization. Philo-
sophical Transactions of the Royal Society B, 376(1835), Article
20200326. https://doi.org/10.1098/rstb.2020.0326, PubMed:
34420384
Perani, D. (2012). Functional and structural connectivity for lan-
guage and music processing at birth. Rendiconti Lincei, 23(3),
305–314. https://doi.org/10.1007/s12210-012-0168-2
Peretz, I., & Vuvan, D. T. (2017). Prevalence of congenital amusia.
European Journal of Human Genetics, 25(5), 625–630. https://doi
.org/10.1038/ejhg.2017.15, PubMed: 28224991
Peretz, I., Vuvan, D., Lagrois, M.-É., & Armony, J. L. (2015). Neural
overlap in processing music and speech. Philosophical Transac-
tions of the Royal Society B: Biological Sciences, 370(1664),
Article 20140090. https://doi.org/10.1098/rstb.2014.0090,
PubMed: 25646513
Perrachione, T. K., Fedorenko, E. G., Vinke, L., Gibson, E., & Dilley,
L. C. (2013). Evidence for shared cognitive processing of pitch in
music and language. PLOS ONE, 8(8), Article e73372. https://doi
.org/10.1371/journal.pone.0073372, PubMed: 23977386
Persici, V., Blain, S. D., Iversen, J. R., Key, A. P., Kotz, S. A.,
McAuley, J. D., & Gordon, R. L. (2022). Individual differences
in neural markers of beat processing predict spoken grammar
skills in six-year-old children. PsyArXiv. https://doi.org/10
.31234/osf.io/9ebvc
Peynircioğlu, Z. F., Durgunoglu, A. Y., & Úney-Küsefog(cid:1)lu, B. (2002).
Phonological awareness and musical aptitude. Journal of
Research in Reading, 25(1), 68–80. https://doi.org/10.1111
/1467-9817.00159
Picciotti, P. M., Bussu, F., Calò, L., Gallus, R., Scarano, E., Di Cintio,
G., Cassarà, F., & D’Alatri, L. (2018). Correlation between musi-
cal aptitude and learning foreign languages: An epidemiological
study in secondary school Italian students. Acta Otorhinolaryn-
gologica Italica, 38(1), 51–55. https://doi.org/10.14639/0392
-100X-1103, PubMed: 29756615
Politimou, N., Dalla Bella, S., Farrugia, N., & Franco, F. (2019).
Born to speak and sing: Musical predictors of language develop-
ment in pre-schoolers. Frontiers in Psychology, 10, Article 948.
https://doi.org/10.3389/fpsyg.2019.00948, PubMed: 31231260
Pruett, D. G., Shaw, D. M., Chen, H. H., Petty, L. E., Polikowsky,
H. G., Kraft, S. J., Jones, R. M., & Below, J. E. (2021). Identifying
developmental stuttering and associated comorbidities in elec-
tronic health records and creating a phenome risk classifier. Jour-
nal of Fluency Disorders, 68, Article 105847. https://doi.org/10
.1016/j.jfludis.2021.105847, PubMed: 33894541
Psaty, B. M., O’Donnell, C. J., Gudnason, V., Lunetta, K. L., Folsom,
A. R., Rotter, J. I., Uitterlinden, A. G., Harris, T. B., Witteman,
J. C. M., & Boerwinkle, E. (2009). Cohorts for heart and aging
research in genomic epidemiology (CHARGE) consortium:
Design of prospective meta-analyses of genome-wide association
studies from 5 cohorts. Circulation: Cardiovascular Genetics,
2(1), 73–80. https://doi.org/10.1161/CIRCGENETICS.108
.829747, PubMed: 20031568
Purcell, S. (2002). Variance components models for gene-
environment interaction in twin analysis. Twin Research, 5(6),
554–571. https://doi.org/10.1375/136905202762342026,
PubMed: 12573187
Rice, M. L., Zubrick, S. R., Taylor, C. L., Hoffman, L., & Gayán, J.
(2018). Longitudinal study of language and speech of twins at 4
Neurobiology of Language
661
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
and 6 years: Twinning effects decrease, zygosity effects disap-
pear, and heritability increases. Journal of Speech, Language,
and Hearing Research, 61(1), 79–93. https://doi.org/10.1044
/2017_JSLHR-L-16-0366, PubMed: 29255901
Ríos-López, P., Molinaro, N., & Lallier, M. (2019). Tapping to a beat
in synchrony predicts brain print sensitivity in pre-readers. Brain
and Language, 199, Article 104693. https://doi.org/10.1016/j
.bandl.2019.104693, PubMed: 31539633
Rocha, S., Attaheri, A., Choisdealbha, A.N., Brusini, P., Flanagan,
S., Mead, N., Boutris, P., Gibbon, S., Olawole-Scott, H., Ahmed,
H., & Goswami, U. (2020, July 6–9). Baby Rhythm: Early infant
audio-motor rhythmic synchronisation predicts later language
outcomes. Poster presented at the Virtual International Congress
on Infant Studies.
Roncaglia-Denissen, M. P., Roor, D. A., Chen, A., & Sadakata, M.
(2016). The enhanced musical rhythmic perception in second
language learners. Frontiers in Human Neuroscience, 10, Article
288. https://doi.org/10.3389/fnhum.2016.00288, PubMed:
27375469
Sallat, S., & Jentschke, S. (2015). Music perception influences
language acquisition: Melodic and rhythmic-melodic perception
in children with specific language impairment. Behavioural
Neurology, 2015, Article 606470. https://doi.org/10.1155/2015
/606470, PubMed: 26508812
Sammler, D., & Elmer, S. (2020). Advances in the neurocognition of
music and language. Brain Sciences, 10(8), Article 509. https://
doi.org/10.3390/brainsci10080509, PubMed: 32748810
Samuelsson, S., Olson, R., Wadsworth, S., Corley, R., DeFries, J. C.,
Willcutt, E., Hulslander, J., & Byrne, B. (2007). Genetic and envi-
ronmental influences on prereading skills and early reading and
spelling development in the United States, Australia, and Scandi-
navia. Reading and Writing, 20(1–2), 51–75. https://doi.org/10
.1007/s11145-006-9018-x
Sanchez-Alonso, S., & Aslin, R. N. (2020). Predictive modeling of
neurobehavioral state and trait variation across development.
Developmental Cognitive Neuroscience, 45, Article 100855.
https://doi.org/10.1016/j.dcn.2020.100855, PubMed: 32942148
Sanchez-Alonso, S., & Aslin, R. N. (2022). Towards a model of
language neurobiology in early development. Brain and
Language, 224, Article 105047. https://doi.org/10.1016/j.bandl
.2021.105047, PubMed: 34894429
Sauer-Zavala, S., Gutner, C. A., Farchione, T. J., Boettcher, H. T.,
Bullis, J. R., & Barlow, D. H. (2017). Current definitions of
“transdiagnostic” in treatment development: A search for consen-
sus. Behavior Therapy, 48(1), 128–138. https://doi.org/10.1016/j
.beth.2016.09.004, PubMed: 28077216
Savage, J. E., Jansen, P. R., Stringer, S., Watanabe, K., Bryois, J., de
Leeuw, C. A., Nagel, M., Awasthi, S., Barr, P. B., Coleman, J. R. I.,
Grasby, K. L., Hammerschlag, A. R., Kaminski, J. A., Karlsson, R.,
Krapohl, E., Lam, M., Nygaard, M., Reynolds, C. A., Trampush,
J. W., … Posthuma, D. (2018). Genome-wide association
meta-analysis in 269,867 individuals identifies new genetic
and functional links to intelligence. Nature Genetics, 50(7),
912–919. https://doi.org/10.1038/s41588-018-0152-6, PubMed:
29942086
Schellenberg, E. G. (2015). Music training and speech perception:
A gene-environment interaction. Annals of the New York Acad-
emy of Sciences, 1337(1), 170–177. https://doi.org/10.1111/nyas
.12627, PubMed: 25773632
Schellenberg, E. G. (2020). Correlation = causation? Music training,
psychology, and neuroscience. Psychology of Aesthetics,
Creativity, and the Arts, 14(4), 475–480. https://doi.org/10.1037
/aca0000263
Schlaug, G., Forgeard, M., Zhu, L., Norton, A., Norton, A., &
Winner, E. (2009). Training-induced neuroplasticity in young
children. Annals of the New York Academy of Sciences, 1169(1),
205–208. https://doi.org/10.1111/j.1749-6632.2009.04842.x,
PubMed: 19673782
Schön, D., & Tillmann, B. (2015). Short- and long-term rhythmic
interventions: Perspectives for language rehabilitation. Annals
of the New York Academy of Sciences, 1337, 32–39. https://doi
.org/10.1111/nyas.12635, PubMed: 25773614
Seesjärvi, E., Särkämö, T., Vuoksimaa, E., Tervaniemi, M., Peretz, I.,
& Kaprio, J. (2016). The nature and nurture of melody: A twin
study of musical pitch and rhythm perception. Behavior Genet-
ics, 46(4), 506–515. https://doi.org/10.1007/s10519-015-9774-y,
PubMed: 26650514
Seither-Preisler, A., Parncutt, R., & Schneider, P. (2014). Size and
synchronization of auditory cortex promotes musical, literacy,
and attentional skills in children. Journal of Neuroscience,
34(33), 10937–10949. https://doi.org/10.1523/ JNEUROSCI
.5315-13.2014, PubMed: 25122894
Selzam, S., Coleman, J. R. I., Caspi, A., Moffitt, T. E., & Plomin, R.
(2018). A polygenic p factor for major psychiatric disorders.
Translational Psychiatry, 8, Article 205. https://doi.org/10.1038
/s41398-018-0217-4, PubMed: 30279410
Shen, L., & Thompson, P. M. (2020). Brain imaging genomics: Inte-
grated analysis and machine learning. Proceedings of the IEEE,
108(1), 125–162. https://doi.org/10.1109/ JPROC.2019
.2947272, PubMed: 31902950
Slater, J., & Kraus, N. (2016). The role of rhythm in perceiving
speech in noise: A comparison of percussionists, vocalists and
non-musicians. Cognitive Processing, 17, 79–87. https://doi.org
/10.1007/s10339-015-0740-7, PubMed: 26445880
Slater, J., Kraus, N., Woodruff Carr, K., Tierney, A., Azem, A., &
Ashley, R. (2018). Speech-in-noise perception is linked to rhythm
production skills in adult percussionists and non-musicians. Lan-
guage, Cognition and Neuroscience, 33(6), 710–717. https://doi
.org/10.1080/23273798.2017.1411960, PubMed: 31475217
Slater, J., Skoe, E., Strait, D. L., O’Connell, S., Thompson, E., &
Kraus, N. (2015). Music training improves speech-in-noise per-
ception: Longitudinal evidence from a community-based music
program. Behavioural Brain Research, 291, 244–252. https://doi
.org/10.1016/j.bbr.2015.05.026, PubMed: 26005127
Slevc, L. R. (2012). Language and music: Sound, structure, and
meaning. WIREs Cognitive Science, 3(4), 483–492. https://doi
.org/10.1002/wcs.1186, PubMed: 26301531
Slevc, L. R., & Miyake, A. (2006). Individual differences in
second-language proficiency: Does musical ability matter? Psy-
chological Science, 17(8), 675–681. https://doi.org/10.1111/j
.1467-9280.2006.01765.x, PubMed: 16913949
Smit, D. J. A., Andreassen, O. A., Boomsma, D. I., Burwell, S. J.,
Chorlian, D. B., de Geus, E. J. C., Elvsåshagen, T., Gordon,
R. L., Harper, J., Hegerl, U., Hensch, T., Iacono, W. G., Jawinski,
P., Jönsson, E. G., Luykx, J. J., Magne, C. L., Malone, S. M.,
Medland, S. E., Meyers, J. L., … Wright, M. J. (2021). Large-scale
collaboration in ENIGMA-EEG: A perspective on the meta-
analytic approach to link neurological and psychiatric liability
genes to electrophysiological brain activity. Brain and Behavior,
11(8), Article e02188. https://doi.org/10.1002/ brb3.2188,
PubMed: 34291596
Snowling, M. J. (2001). From language to reading and dyslexia.
Dyslexia, 7(1), 37–46. https://doi.org/10.1002/dys.185,
PubMed: 11305230
Sodini, S. M., Kemper, K. E., Wray, N. R., & Trzaskowski, M. (2018).
Comparison of genotypic and phenotypic correlations:
Neurobiology of Language
662
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
Cheverud’s conjecture in humans. Genetics, 209(3), 941–948.
https://doi.org/10.1534/genetics.117.300630, PubMed:
29739817
Stein, C. M., Lu, Q., Elston, R. C., Freebairn, L. A., Hansen, A. J.,
Shriberg, L. D., Taylor, H. G., Lewis, B. A., & Iyengar, S. K.
(2011). Heritability estimation for speech-sound traits with devel-
opmental trajectories. Behavior Genetics, 41(2), 184–191. https://
doi.org/10.1007/s10519-010-9378-5, PubMed: 20623172
Steinbrink, C., Knigge, J., Mannhaupt, G., Sallat, S., & Werkle, A.
(2019). Are temporal and tonal musical skills related to phonol-
ogical awareness and literacy skills? – Evidence from two
cross-sectional studies with children from different age groups.
Frontiers in Psychology, 10, Article 805. https://doi.org/10.3389
/fpsyg.2019.00805, PubMed: 31040806
Strait, D. L., Hornickel, J., & Kraus, N. (2011). Subcortical process-
ing of speech regularities underlies reading and music aptitude in
children. Behavioral and Brain Functions, 7, Article 44. https://
doi.org/10.1186/1744-9081-7-44, PubMed: 22005291
Subramanian, A., Tamayo, P., Mootha, V. K., Mukherjee, S., Ebert,
B. L., Gillette, M. A., Paulovich, A., Pomeroy, S. L., Golub, T. R.,
Lander, E. S., & Mesirov, J. P. (2005). Gene set enrichment
analysis: A knowledge-based approach for interpreting
genome-wide expression profiles. Proceedings of the National
Academy of Sciences, 102(43), 15545–15550. https://doi.org/10
.1073/pnas.0506580102, PubMed: 16199517
Sudlow, C., Gallacher, J., Allen, N., Beral, V., Burton, P., Danesh, J.,
Downey, P., Elliott, P., Green, J., Landray, M., Liu, B., Matthews,
P., Ong, G., Pell, J., Silman, A., Young, A., Sprosen, T., Peakman,
T., & Collins, R. (2015). UK Biobank: An open access resource
for identifying the causes of a wide range of complex diseases
of middle and old age. PLOS Medicine 12(3), Article e1001779.
https://doi.org/10.1371/journal.pmed.1001779, PubMed:
25826379
Sutherland, M. E., Zatorre, R. J., Watkins, K. E., Hervé, P. Y.,
Leonard, G., Pike, B. G., Witton, C., & Paus, T. (2012). Anatomical
correlates of dynamic auditory processing: Relationship to liter-
acy during early adolescence. NeuroImage, 60(2), 1287–1295.
https://doi.org/10.1016/j.neuroimage.2012.01.051, PubMed:
22266413
Swaminathan, S., & Schellenberg, E. G. (2020). Musical ability,
music training, and language ability in childhood. Journal of
Experimental Psychology: Learning, Memory, and Cognition,
46(12), 2340–2348. https://doi.org/10.1037/xlm0000798,
PubMed: 31750723
Swaminathan, S., Schellenberg, E. G., & Venkatesan, K. (2018).
Explaining the association between music training and reading
in adults. Journal of Experimental Psychology: Learning, Memory,
and Cognition, 44(6), 992–999. https://doi.org/10.1037
/xlm0000493, PubMed: 29389190
Thakur, D., Martens, M. A., Smith, D. S., & Roth, E. (2018).
Williams syndrome and music: A systematic integrative review.
Frontiers in Psychology, 9, Article 2203. https://doi.org/10.3389
/fpsyg.2018.02203, PubMed: 30487769
Tichko, P., & Skoe, E. (2018). Musical experience, sensorineural
auditory processing, and reading subskills in adults. Brain Sciences,
8(5), Article 77. https://doi.org/10.3390/brainsci8050077, PubMed:
29702572
Tierney, A., & Kraus, N. (2013). Music training for the development
of reading skills. Progress in Brain Research, 207, 209–241.
https://doi.org/10.1016/B978-0-444-63327-9.00008-4, PubMed:
24309256
Tierney, A., White-Schwoch, T., MacLean, J., & Kraus, N. (2017).
Individual differences in rhythm skills: Links with neural
consistency and linguistic ability. Journal of Cognitive Neurosci-
ence, 29(5), 855–868. https://doi.org/10.1162/jocn_a_01092,
PubMed: 28129066
Tilot, A. K., Khramtsova, E. A., Liang, D., Grasby, K. L., Jahanshad,
N., Painter, J., Colodro-Conde, L., Bralten, J., Hibar, D. P., Lind,
P. A., Liu, S., Brotman, S. M., Thompson, P. M., Medland, S. E.,
Macciardi, F., Stranger, B. E., Davis, L. K., Fisher, S. E., & Stein,
J. L. (2021). The evolutionary history of common genetic variants
influencing human cortical surface area. Cerebral Cortex, 31(4),
1873–1887. https://doi.org/10.1093/cercor/bhaa327, PubMed:
33290510
Tomblin, J. B., & Buckwalter, P. R. (1998). Heritability of poor lan-
guage achievement among twins. Journal of Speech, Language,
and Hearing Research, 41(1), 188–199. https://doi.org/10.1044
/jslhr.4101.188, PubMed: 9493744
Trehub, S. E., & Trainor, L. (1998). Singing to infants: Lullabies and
play songs. In C. Rovee-Collier, L. Litsitt, & H. Hayne (Eds.),
Advances in infancy research (pp. 43–78). Ablex Publishing.
Turker, S., Reiterer, S. M., Seither-Preisler, A., & Schneider, P.
(2017). “When music speaks”: Auditory cortex morphology as
a neuroanatomical marker of language aptitude and musicality.
Frontiers in Psychology, 8, Article 2096. https://doi.org/10.3389
/fpsyg.2017.02096, PubMed: 29250017
Ujfalussy, J. (1993). The role of music and song in human commu-
nication. In E. Pásztor, J. Vajda, & F. Loew (Eds.), Language and
speech (pp. 6–8). Springer. https://doi.org/10.1007/978-3-7091
-9239-9_2, PubMed: 8498203
Ullén, F., Mosing, M. A., Holm, L., Eriksson, H., & Madison, G.
(2014). Psychometric properties and heritability of a new online
test for musicality, the Swedish musical discrimination test. Per-
sonality and Individual Differences, 63, 87–93. https://doi.org/10
.1016/j.paid.2014.01.057
van der Sluis, S., Posthuma, D., & Dolan, C. V. (2012). A note
on false positives and power in G × E modelling of twin data.
Behavior Genetics, 42(1), 170–186. https://doi.org/10.1007
/s10519-011-9480-3, PubMed: 21748401
van Eck, N. J. & Waltman, L. (2010). Software Survey: VOSviewer, a
computer program for bibliometric mapping. Scientometrics,
84(2). 523–538. https://doi.org/10.1007/s11192-009-0146-3,
PubMed: 20585380
van Zuijen, T. L., Plakas, A., Maassen, B. A. M., Been, P., Maurits,
N. M., Krikhaar, E., van Driel, J., & van der Leij, A. (2012). Tem-
poral auditory processing at 17 months of age is associated with
preliterate language comprehension and later word reading flu-
ency: An ERP study. Neuroscience Letters, 528(1), 31–35. https://
doi.org/10.1016/j.neulet.2012.08.058, PubMed: 22981882
Vinkhuyzen, A. A. E., van der Sluis, S., Posthuma, D., & Boomsma,
D. I. (2009). The heritability of aptitude and exceptional talent
across different domains in adolescents and young adults.
Behavior Genetics, 39(4), 380–392. https://doi.org/10.1007
/s10519-009-9260-5, PubMed: 19288254
Volkow, N. D., Koob, G. F., Croyle, R. T., Bianchi, D. W., Gordon,
J. A., Koroshetz, W. J., Perez-Stable, E. J., Riley, W. T., Block,
M. H., Conway, K., Deeds, B. G., Dowling, G. J., Grant, S.,
Howlett, K. D., Matochik, J. A., Morgan, G. D., Murray, M. M.,
Noronha, A., Spong, C. Y., … Weiss, S. R. B. (2018). The concep-
tion of the ABCD study: From substance use to a broad NIH
collaboration. Developmental Cognitive Neuroscience, 32, 4–7.
https://doi.org/10.1016/j.dcn.2017.10.002, PubMed: 29051027
Wagner, R. K., Torgesen, J. K., & Rashotte, C. A. (1999). Compre-
hensive test of phonological processing (CTOPP). Pro-Ed.
Walters, C. E., Jr., Nitin, R., Margulis, K., Boorom, O., Gustavson,
D. E., Bush, C. T., Davis, L. K., Below, J. E., Cox, N. J., Camarata,
Neurobiology of Language
663
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Understanding musicality-language links
S. M., & Gordon, R. L. (2020). Automated phenotyping tool for
identifying developmental language disorder cases in health sys-
tems data (APT-DLD): A new research algorithm for deployment
in large-scale electronic health record systems. Journal of Speech,
Language, and Hearing Research, 63(9), 3019–3035. https://doi
.org/10.1044/2020_JSLHR-19-00397, PubMed: 32791019
Watanabe, K., Stringer, S., Frei, O., Umićević Mirkov, M., de
Leeuw, C., Polderman, T. J. C., van der Sluis, S., Andreassen,
O. A., Neale, B. M., & Posthuma, D. (2019). A global overview
of pleiotropy and genetic architecture in complex traits. Nature
Genetics, 51(9), 1339–1348. https://doi.org/10.1038/s41588-019
-0481-0, PubMed: 31427789
Weinert, S. (1992). Deficits in acquiring language structure: The
importance of using prosodic cues. Applied Cognitive Psychol-
ogy, 6(6), 545–571. https://doi.org/10.1002/acp.2350060607
Weissbrod, O., Flint, J., & Rosset, S. (2018). Estimating SNP-based
heritability and genetic correlation in case-control studies
directly and with summary statistics. The American Journal of
Human Genetics, 103(1), 89–99. https://doi.org/10.1016/j.ajhg
.2018.06.002, PubMed: 29979983
Wesseldijk, L. W., Gordon, R. L., Mosing, M. A., & Ullén, F. (2021).
Music and verbal ability—A twin study of genetic and environ-
mental associations. Psychology of Aesthetics, Creativity, and the
Arts. Advance online publication. https://doi.org/10.1037
/aca0000401
Wesseldijk, L. W., Mosing, M. A., & Ullén, F. (2019). Gene-environ-
ment interaction in expertise: The importance of childhood envi-
ronment for musical achievement. Developmental Psychology,
55(7), 1473–1479. https://doi.org/10.1037/dev0000726,
PubMed: 30883154
White, E. J., Hutka, S. A., Williams, L. J., & Moreno, S. (2013).
Learning, neural plasticity and sensitive periods: Implications
for language acquisition, music training and transfer across the
lifespan. Frontiers in Systems Neuroscience, 7, Article 90.
https://doi.org/10.3389/fnsys.2013.00090, PubMed: 24312022
Wong, P., & Cheng, M. W. (2020). On the relationship between
general auditory sensitivity and speech perception: An
examination of pitch and lexical tone perception in 4- to
6-year-old children. Journal of Speech, Language, and Hearing
Research, 63(2), 487–498. https://doi.org/10.1044/2019_JSLHR
-19-00104, PubMed: 32073343
Woodard, K., Pozzan, L., & Trueswell, J. C. (2016). Taking your
own path: Individual differences in executive function and lan-
guage processing skills in child learners. Journal of Experimental
Child Psychology, 141, 187–209. https://doi.org/10.1016/j.jecp
.2015.08.005, PubMed: 26454180
Woodruff Carr, K., White-Schwoch, T., Tierney, A. T., Strait, D. L., &
Kraus, N. (2014). Beat synchronization predicts neural speech
encoding and reading readiness in preschoolers. Proceedings
of the National Academy of Sciences of the United States of
America, 111(40), 14559–14564. https://doi.org/10.1073/pnas
.1406219111, PubMed: 25246562
Yates, K. M., Moore, D. R., Amitay, S., & Barry, J. G. (2019). Sensi-
tivity to melody, rhythm, and beat in supporting speech-in-noise
perception in young adults. Ear and Hearing, 40(2), 358–367.
https://doi.org/10.1097/AUD.0000000000000621, PubMed:
29965864
Yu, M., Xu, M., Li, X., Chen, Z., Song, Y., & Liu, J. (2017). The
shared neural basis of music and language. Neuroscience, 357,
208–219. https://doi.org/10.1016/j.neuroscience.2017.06.003,
PubMed: 28602921
Zhang, J., Meng, Y., Wu, C., & Zhou, D. Q. (2017). Writing system
modulates the association between sensitivity to acoustic cues in
music and reading ability: Evidence from Chinese-English bilin-
gual children. Frontiers in Psychology, 8, Article 1965. https://doi
.org/10.3389/fpsyg.2017.01965, PubMed: 29170647
Zuk, J., Andrade, P. E., Andrade, O. V. C. A., Gardiner, M., & Gaab,
N. (2013). Musical, language, and reading abilities in early Por-
tuguese readers. Frontiers in Psychology, 4, Article 288. https://
doi.org/10.3389/fpsyg.2013.00288, PubMed: 23785339
Zuk, J., & Gaab, N. (2018). Evaluating predisposition and training in
shaping the musician’s brain: The need for a developmental per-
spective. Annals of the New York Academy of Sciences, 1423,
40–50. https://doi.org/10.1111/nyas.13737, PubMed: 29799116
Neurobiology of Language
664
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
6
1
5
2
0
6
2
9
2
2
n
o
_
a
_
0
0
0
7
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3