REVIEW ARTICLE
The Role of the Right Hemisphere in Processing
Phonetic Variability Between Talkers
a n o p e n a c c e s s
j o u r n a l
Keywords: speech perception, talker identity, vocal identity, phonetic variability, right hemisphere,
functional neuroimaging
Sahil Luthra
Psychological Sciences, University of Connecticut, Storrs, CT, USA
ABSTRACT
Neurobiological models of speech perception posit that both left and right posterior temporal brain
regions are involved in the early auditory analysis of speech sounds. However, frank deficits in
speech perception are not readily observed in individuals with right hemisphere damage. Instead,
damage to the right hemisphere is often associated with impairments in vocal identity processing.
Herein lies an apparent paradox: The mapping between acoustics and speech sound categories
can vary substantially across talkers, so why might right hemisphere damage selectively impair
vocal identity processing without obvious effects on speech perception? In this review, I attempt to
clarify the role of the right hemisphere in speech perception through a careful consideration of its
role in processing vocal identity. I review evidence showing that right posterior superior temporal,
right anterior superior temporal, and right inferior / middle frontal regions all play distinct roles
in vocal identity processing. In considering the implications of these findings for neurobiological
accounts of speech perception, I argue that the recruitment of right posterior superior temporal
cortex during speech perception may specifically reflect the process of conditioning phonetic
identity on talker information. I suggest that the relative lack of involvement of other right
hemisphere regions in speech perception may be because speech perception does not necessarily
place a high burden on talker processing systems, and I argue that the extant literature hints at
potential subclinical impairments in the speech perception abilities of individuals with right
hemisphere damage.
INTRODUCTION
A rich neuroscientific literature has established the importance of the brain’s left hemisphere for
processing language. Early patient data demonstrated that damage to left superior temporal
(Wernicke, 1874) and left inferior frontal (Broca, 1861) brain regions can lead to a loss of language
abilities (i.e., aphasia), and recent studies also support a critical role for left hemisphere structures
in the process of speech perception specifically. In particular, a wealth of neuroimaging evidence
suggests that left superior temporal regions are important for imposing category structure on acous-
tically similar speech sounds (Desai et al., 2008; Liebenthal et al., 2010; Luthra, Guediche, et al.,
2019; Mesgarani et al., 2014; Myers, 2007; Yi et al., 2019) and that left inferior frontal regions play
a key role in differentiating between similar speech sound categories (Lee et al., 2012; Myers,
2007; Myers, Blumstein, et al., 2009; Rogers & Davis, 2018; Xie & Myers, 2018).
Relatively less is known about the extent to which the right hemisphere plays a role in speech
perception, which may largely be a result of the fact that damage to the right hemisphere does not
Citation: Luthra, S. (2021). The role of
the right hemisphere in processing
phonetic variability between talkers.
Neurobiology of Language, 2(1),
138–151. https://doi.org/10.1162
/nol_a_00028
DOI:
https://doi.org/10.1162/nol_a_00028
Supporting Information:
https://doi.org/10.1162/nol_a_00028
Received: 17 April 2020
Accepted: 13 November 2020
Competing Interests: The author has
declared that no competing interests
exist.
Corresponding Author:
Sahil Luthra
sahil.luthra@uconn.edu
Handling Editor:
Jonathan Peelle
Copyright: © 2021 Massachusetts
Institute of Technology. Published
under a Creative Commons Attribution
4.0 International (CC BY 4.0) license.
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
typically result in an aphasia (Blumstein & Myers, 2014; Turkeltaub & Branch Coslett, 2010).
Instead, research on the right hemisphere’s role in language processing has largely focused on
its high-level role in processing pragmatic information (Siegal et al., 1996) such as emotional
prosody (Heilman et al., 1984), metaphorical language (Schmidt et al., 2007), and other forms
of nonliteral language, including humor and sarcasm (Mitchell & Crow, 2005). While prominent
neurobiological models (e.g., the Dual Stream Model; Hickok & Poeppel, 2000, 2004, 2007)
have proposed at least some degree of right hemisphere involvement in processing phonetic
information, the precise function of the right hemisphere in speech perception is relatively under-
specified, especially compared to the more detailed characterization of the left hemisphere.
Notably, however, the right hemisphere has been heavily implicated in vocal identity
processing—that is, in processing perceptual information about a voice in order to identify
who is talking (Maguinness et al., 2018; Perrodin et al., 2015). Neuropsychological studies have
linked right hemisphere strokes to deficits in identifying people by voice (Luzzi et al., 2018;
Roswandowitz et al., 2018; Van Lancker & Canter, 1982; Van Lancker & Kreiman, 1987), though
strikingly, patients with right hemisphere damage do not typically show frank deficits in speech
perception. It is puzzling that these patients show deficits in vocal identity processing but not in
speech perception, since talker processing and phonetic processing are known to be closely tied;
the mapping between acoustic information and phonetic information can vary considerably
across talkers, and theoretical accounts of speech perception argue that to perceive the speech
signal accurately, listeners condition phonetic identity on talker information (Johnson, 2008;
Joos, 1948; Kleinschmidt, 2019; Kleinschmidt & Jaeger, 2015). Given that phonetic processing
is tightly linked to talker information, I suggest that by considering the role of the right hemisphere
in processing nonlinguistic information about vocal identity, we might better understand the
role of the right hemisphere in speech perception.
Note that in this review, I use the term “talker processing” largely to refer to the processing of
voice information in support of processing speech, consistent with the use of the term “talker” in
the speech perception literature. In contrast, I use “vocal identity processing” to refer to the pro-
cessing of voice information to determine who is talking. These two processes are assumed to be
theoretically distinct but to rely on some shared cognitive and neural architecture (Maguinness
et al., 2018).
The structure of this review is as follows. After briefly discussing the interdependence between
phonetic processing and talker processing, I review the existing literature on the role of the right
hemisphere in vocal identity processing, paying careful attention to the contributions of different
brain regions. I then consider current perspectives on the role of the right hemisphere in speech
perception before closing with the hypothesis that the right hemisphere (and the right superior
posterior temporal cortex in particular) may play an important role in allowing listeners to
condition phonetic identity on talker information during speech perception.
How Is Phonetic Processing Linked to Talker Processing?
Individual talkers can differ substantially in how they produce their speech sounds, with talkers
varying both In their use of rapid temporal cues such as voice-onset time (VOT; Allen et al., 2003)
and in their use of spectral cues that indicate phoneme identity (Peterson & Barney, 1952). A vast
literature indicates that listeners are highly sensitive to these talker-specific differences in pho-
netic variation and that they adjust the mapping between acoustic information and phonetic cat-
egories accordingly (e.g., Allen & Miller, 2004; Clayards et al., 2008; Kraljic & Samuel, 2005;
Norris et al., 2003; Theodore & Monto, 2019). More generally, theoretical accounts of speech
perception posit that listeners maintain distinct sets of beliefs about how different talkers produce
Neurobiology of Language
139
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
their speech sounds (Kleinschmidt & Jaeger, 2015), meaning that phonetic processing is intrin-
sically linked to talker information.
The interdependence between phonetic processing and talker processing is further highlighted
by studies showing that phonetic processing is facilitated when listeners are familiar with a
particular talker (a talker familiarity effect) and by studies showing that talker processing is facil-
itated when listeners are familiar with the phonetic inventory of a particular language (a language
familiarity effect). With regard to the former, several studies have found that talker familiarity
leads to perceptual gains when processing speech in noise (Kreitewolf, Mathias, & von
Kriegstein, 2017; Nygaard & Pisoni, 1998; Souza et al., 2013), and that talker familiarity makes
it easier to selectively attend to one talker while ignoring another (Holmes et al., 2018; Holmes &
Johnsrude, 2020; Johnsrude et al., 2013; Newman & Evers, 2007). With regard to the language
familiarity effect, a number of studies have demonstrated that talker identification is facilitated
when listeners hear speech in their native language (in which they are familiar with the phonetic
category structure) compared to when they hear speech in a foreign language (in which they are
not; Goggin et al., 1991; Perrachione & Wong, 2007). Talker familiarity effects can be understood
by considering that when listeners receive practice with a particular talker, the acoustic dimen-
sions that are relevant for processing that talker’s voice acquire distinctiveness; if the same dimen-
sions are relevant for both talker processing and phonetic processing, then experience with a
talker should incur performance benefits for phonetic processing (Nygaard & Pisoni, 1998).
Similarly, language familiarity effects can be understood by recognizing that when listeners are
familiar with the phonetic inventory of a particular language, the key acoustic-phonetic dimen-
sions for that language likewise acquire distinctiveness—and if the same dimensions are relevant
for talker processing, then experience with phonetic processing should yield benefits for talker
processing. Taken together, such findings indicate that speech perception and talker processing
are highly interrelated processes.
How Does the Right Hemisphere Support Vocal Identity Processing?
A focus on the right hemisphere regions involved in talker processing could inform neurobiolog-
ical accounts of phonetic processing, at least to the extent that the same right hemisphere regions
are recruited for both processes. The association between the right hemisphere and vocal identity
processing dates back at least to early clinical studies by Van Lancker and colleagues, who
demonstrated that right-hemisphere stroke patients were more likely than left-hemisphere patients
to show impairments in identifying the voices of celebrities when performing a forced-choice task
(Van Lancker & Canter, 1982; Van Lancker & Kreiman, 1987). Since then, neuroimaging studies
have clarified the role of different right hemisphere regions in vocal identity processing (see
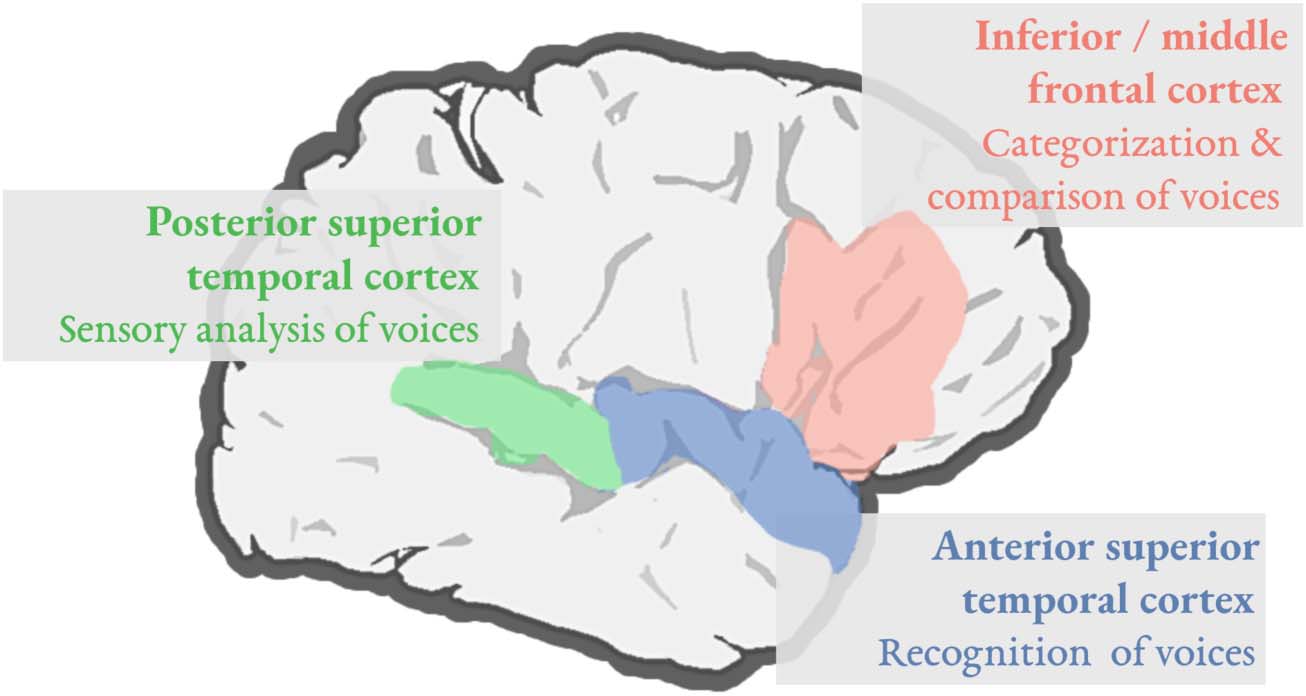
Maguinness et al., 2018, for a recent review). As illustrated in Figure 1, these studies have
revealed that vocal identity processing is largely supported by a set of temporal regions, with
posterior temporal regions (shaded green in Figure 1) playing an important role in the early
sensory analysis of vocal information, and anterior temporal regions (shaded blue) being impor-
tant for vocal identity recognition. While not always recruited in vocal identity processing, right
frontal brain regions (shaded pink) have been implicated in tasks that require listeners to make
comparisons between voices, especially when comparing a vocal sample to a target voice.
Temporal lobe contributions to vocal identity processing
Neuroimaging evidence suggests that there is a posterior–anterior gradient in superior temporal
lobe responses to vocal information, with right posterior temporal regions being thought to play
a larger role in the general sensory processing of voice information (Andics, McQueen,
Petersson, et al., 2010; Belin, Zatorre, Lafaille, et al., 2000; Schall et al., 2014; von Kriegstein
Neurobiology of Language
140
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
Figure 1. Vocal identity processing is supported by a right-lateralized system involving the poste-
rior superior temporal cortex (green), the anterior superior temporal cortex (blue), and the inferior/
middle frontal cortex (pink). The right superior temporal cortex has been implicated in mapping
vocal acoustic information to a person’s identity, with posterior regions underlying the early sensory
analysis of voices and more anterior regions supporting vocal identity recognition. Left temporal
regions (not shown) may contribute to vocal identity processing, with their involvement potentially
depending on the familiarity of the voice being processed. Right inferior and middle frontal regions
play a role during the categorization of vocal stimuli into task-relevant categories, as well as when
listeners must compare a target voice to a vocal sample in working memory, respectively.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
& Giraud, 2004) and right anterior temporal regions being implicated in mapping from vocal
information to a specific identity (Andics, McQueen, Petersson, et al., 2010; Belin, 2006;
Belin, Fecteau, & Bédard, 2004; Imaizumi et al., 1997; Nakamura et al., 2001; von Kriegstein
& Giraud, 2004). Support for the involvement of posterior superior temporal cortex in vocal
identity processing comes from a wide range of studies, including a seminal fMRI study in which
Belin, Zatorre, Lafaille, et al. (2000) examined cortical responses when subjects passively
listened to human vocal stimuli (both speech sounds and nonspeech vocalizations like laughter)
as well as to several types of control stimuli (such as animal sounds, bells, and speech-shaped
white noise). Vocal stimuli elicited robust activation in the superior temporal sulcus (STS) bilat-
erally, but activation on the right was greater both in magnitude and in area than activation on
the left. Notably, the response in the right STS was not specific to speech, as activation in the
right posterior STS did not differ between speech and nonspeech human vocalizations. Belin,
Zatorre, Lafaille, et al. further observed that band-pass filtering the stimuli led to a reduction of
STS activation, and this reduction of activation was associated with worsened behavioral per-
formance in a perceptual judgment task conducted outside the scanner (e.g., deciding whether
the sounds were vocal or nonvocal). Such results indicate that the right STS is involved in dif-
ferentiating between vocal and nonvocal auditory information but do not indicate whether it is
necessary for such discrimination. Evidence for the latter comes from a study by Bestelmeyer
et al. (2011). In that study, the authors first performed a functional localizer to identify the spe-
cific parts of right temporal cortex that were recruited when participants passively listened to
voices compared to nonvocal auditory stimuli. Subsequent transcranial magnetic stimulation
(TMS) to these regions impaired participants’ ability to discriminate between vocal and non-
vocal sounds. Taken together, these findings suggest a critical role for the right posterior STS
in processing the acoustic detail of human voices.
Neurobiology of Language
141
The role of the right hemisphere in speech perception
By contrast, more anterior regions in the right superior temporal cortex seem to be important
when listeners need to map these acoustic details to a specific identity. Belin and Zatorre (2003)
used fMRI to measure the habituation of neural regions in response to a train of stimuli presented
over a short interval. The researchers found that the right anterior STS habituated (i.e., its activity
diminished) when listeners encountered a stream of phonologically distinct syllables that were all
spoken by the same talker. By contrast, this region did not habituate when listeners encountered a
stream of phonologically identical syllables spoken by different talkers. In other words, this
region’s response depended on who was producing the speech but not on what the content of
the speech was. Convergent evidence comes from Formisano et al. (2008), who collected fMRI
data while participants passively listened to different vowels spoken by different talkers. The
authors then trained a machine learning algorithm to classify stimuli on the basis of talker identity
(ignoring vowel identity) and found that the most discriminative voxels were located in right
anterior STS. More recently, Luzzi et al. (2018) reported a case study of a patient who had suffered
a stroke that affected his right anterior STS but did not affect posterior temporal regions; while the
patient was unimpaired in his ability to indicate whether two voices were the same or different, he
was no longer able to recognize his favorite singers on the basis of their voices alone. Overall,
these findings suggest a role for right anterior temporal regions in recognizing vocal identity, as
opposed to low-level processing of voice information.
Consistent with this view, a number of other studies have found that right anterior temporal
regions are recruited when listeners must match vocal details to a known vocal identity. In an
fMRI study by von Kriegstein, Eger, et al. (2003), for instance, greater right anterior STS activation
was observed when listeners attended to vocal information compared to linguistic information.
Similar results were observed in an MEG study by Schall et al. (2014), in which greater right
anterior STS activity was observed when subjects had to match a sample of speech to a name
compared to when they had to indicate whether a probe word had been present in the speech
stream. Moreover, the authors observed a strong correlation between the degree of right anterior
STS activity and subjects’ behavioral accuracy on this talker judgment task, suggesting that the
variability in the activity of the right anterior STS might underlie individual differences in voice
recognition. One way to conceptualize these results is to note that in both the study by von
Kriegstein, Eger, et al. (2003) and the study by Schall et al. (2014), listeners were required to com-
pare the incoming auditory signal to their internal representation of a particular vocal identity. As
such, the findings indicate that the right anterior STS may play an important role in matching
complex auditory objects to a stored vocal representation.
The suggestion that right anterior temporal regions are important for identifying a person on the
basis of their voice is particularly striking given studies indicating that the right anterior temporal
cortex is vital for person recognition more broadly (Gainotti, 2007). Individuals with damage to
the right anterior temporal lobe may show selective impairments in identifying people on the
basis of their faces (Damasio, 1990; Gainotti et al., 2003; Tranel et al., 1997) or voices
(Gainotti et al., 2003) alone. As such, right temporal regions are thought to be critically involved
in integrating perceptual information with conceptual person-specific knowledge (Gainotti,
2007). Consistent with this view, Ross et al. (2010) demonstrated that transcranial direct current
stimulation of the right anterior temporal lobe modulated the likelihood that individuals would
recover from a tip-of-the-tongue state when naming celebrities from their photographs. However,
no such effect of stimulation was observed when subjects were shown photographs of famous
places. Such findings point to a critical role of right anterior temporal regions in representing se-
mantic knowledge about person identity specifically. As such, the involvement of right anterior
temporal regions in vocal identity recognition may reflect access to multimodal information
related to person identity (Maguinness et al., 2018; Perrodin et al., 2015).
Neurobiology of Language
142
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
While vocal identity processing is supported predominantly by right hemisphere regions,
there has been some evidence for left hemisphere involvement in this process. In a study by
von Kriegstein and Giraud (2004), for instance, listeners heard speech from talkers who were
personally known to them, as well as speech from relatively unfamiliar talkers, to whom listeners’
previous exposure was limited to a few audio clips presented during a familiarization phase.
Participants heard several sentences spoken by both the familiar and unfamiliar talkers; on each
trial, they had to make a judgment either about the verbal content or about the vocal identity.
Making judgments about vocal identity elicited robust activation of both the right posterior
and right anterior STS, consistent with the characterization of the right posterior STS being
involved in sensory processing of vocal identity and the right anterior STS being involved in vocal
identity recognition. The researchers then examined whether functional connectivity with these
right temporal regions differed as a function of whether the talkers were personally known to the
participants. When participants listened to familiar talkers, there was robust connectivity among
different subregions of the right superior temporal lobe. By contrast, when participants heard
unfamiliar talkers, there was robust connectivity between the right posterior temporal lobe and
the left posterior temporal lobe, suggesting that talker familiarity may modulate the involvement
of left hemisphere regions in vocal identity processing. Other studies have supported the notion
that the involvement of left temporal cortex in vocal identity processing may differ as a function of
talker familiarity (Roswandowitz et al., 2018), and additional work suggests that language famil-
iarity may similarly modulate the involvement of left hemisphere regions in vocal identity pro-
cessing (Perrachione et al., 2009). Nonetheless, at least one study of stroke patients found that
while individuals with right hemisphere damage were impaired in recognizing familiar voices,
the performance of patients with left hemisphere damage was comparable to that of healthy con-
trols (Lang et al., 2009); that is, there was no evidence for a left hemisphere role in processing
familiar voices. Though additional work is needed to clarify the precise contributions of left
temporal cortex, extant data suggest that left posterior temporal regions may play at least some
role in vocal identity processing. Nevertheless, the role of the left hemisphere in processing
vocal identity information is clearly limited, especially in contrast to the well-established role
of the right hemisphere.
Frontal lobe contributions to vocal identity processing
In addition to a role for the right temporal lobe, some studies have posited a role for right frontal
regions in vocal identity recognition, particularly during tasks that require listeners to categorize
voices (Andics, McQueen, & Petersson, 2013; Jones et al., 2015; Zäske et al., 2017) or that require
listeners to compare a voice sample to a referent in working memory (Stevens, 2004). Some
evidence for the former comes from a study by Andics, McQueen, and Petersson (2013), who
presented listeners with a vocal morph continuum where stimuli consisted of two different voices
blended in different proportions. Training was used to establish a category boundary between the
two voices, and participants then completed an fMRI session in which they had to categorize steps
along the morph continuum. Subsequently, a second set of training sessions was administered to
establish a new category boundary, after which participants completed a second fMRI session. The
authors found that the activation of the right inferior frontal cortex depended on the proximity of a
stimulus to the category boundary established during training (regardless of the precise acoustic
details). These findings were interpreted as evidence that the right inferior frontal cortex supports
the categorization of vocal stimuli into vocal identity categories, with the harder-to-categorize
near-boundary stimuli eliciting more activation in right inferior frontal cortex. Consistent with
this finding, Jones et al. (2015) observed that stroke patients who had damage to right frontal
cortex were impaired in their categorization of talker gender when presented with stimuli from
Neurobiology of Language
143
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
male–female continua; critically, the right STS was intact in these patients, suggesting that these
results were not attributable to impairments in early sensory processing. Thus, the right inferior
frontal cortex appears to play a critical role in allowing listeners to evaluate voices with respect
to known vocal categories, whether these categories are task-relevant (e.g., ones established
through training) or socio-indexically derived (i.e., categories based on talker-relevant social cues,
such as gender or sexual orientation; Johnson, 2008; Munson, 2007).
The right frontal cortex has also been implicated in tasks that require listeners to compare
one vocal sample to a second sample held in working memory. In an fMRI study, Stevens (2004)
had participants listen to a series of stimuli while performing a two-back working memory task.
On some blocks, they had to indicate whether the talker producing the current stimulus was the
same as the talker who had produced the stimulus two items previously, and on other blocks,
they had to indicate whether the same word had been produced two items previously. Subjects
showed greater activation in the right middle frontal gyrus when performing the talker two-back
task and greater activation in left inferior frontal gyrus when performing the word two-back task.
Such a finding suggests a role for right frontal regions when subjects have to make explicit
comparisons about vocal identity across stimuli.
Strikingly, the role of right frontal brain areas in vocal identity recognition seems to parallel a
similar role for left frontal regions in phonological processing during speech perception. Just as the
right inferior frontal cortex is strongly recruited when listeners hear stimuli near a vocal category
boundary, the left inferior frontal cortex has been shown to be robustly activated by stimuli near a
phonetic category boundary (Myers, 2007). Similarly, right frontal regions are recruited when
demands on vocal working memory are high, just as left frontal regions are recruited when
demands on phonological processing are high (Burton et al., 2000). More generally, the extant
literature suggests that vocal identity processing is supported by a right-lateralized neural system,
whereas speech perception is supported by an analogous left-lateralized system. To the extent that
phonetic processing is influenced by talker information (as described in How Is Phonetic
Processing Linked to Talker Processing?), it is worth considering how the right hemisphere
may interact with the left to support speech perception; I turn to this question next.
How Might the Right Hemisphere Support Speech Perception?
Though the leftward lateralization of language processing represents a core feature of current
neurobiological models of speech perception (Binder, Frost, et al., 1997; Binder, Swanson,
et al., 1996; Geschwind, 1970; Hickok & Poeppel, 2000, 2004, 2007; Rauschecker & Scott,
2009), there is nevertheless some evidence that the right hemisphere—and right temporal cortex
in particular—does play a role in speech perception. At least one study (Boatman et al., 1998)
demonstrated intact syllable discrimination in a patient whose left hemisphere was sedated
through a sodium amobarbital injection (Wada & Rasmussen, 1960), and functional neuroim-
aging studies of speech perception routinely implicate right temporal structures in speech per-
ception (Belin, Zatorre, Hoge, et al., 1999; Blumstein et al., 2005; Davis et al., 2011; Giraud
et al., 2004; Turkeltaub & Branch Coslett, 2010; Zatorre et al., 1996). More recently, a study by
Kennedy-Higgins et al. (2020) found that listeners’ ability to repeat speech presented against
background noise was impaired when they received TMS above either the left or right superior
temporal gyrus (STG), but not when stimulation was performed at a control site. Collectively,
such findings suggest a nonnegligible role for the right hemisphere in speech perception.
However, while left and right temporal structures are both routinely recruited for speech
perception, they do not respond equally to acoustic information. In particular, left temporal
regions seem to respond preferentially to rapid changes in the auditory signal, whereas right
Neurobiology of Language
144
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
temporal regions appear to have a general preference for processing low-frequency modulations
in the acoustic signal (Belin, McAdams, et al., 1998; Robin et al., 1990; Schwartz & Tallal, 1980;
Scott et al., 2000). On the basis of these and other findings, Poeppel (2003) proposed the
asymmetric sampling in time (AST) hypothesis. Under this view, the left hemisphere samples
the speech signal at a relatively fast rate (40 Hz) and as such is well-suited for processing rapidly
changing acoustic information (fluctuations on the order of approximately 25 ms); as such, left
temporal processing is thought to be reflected in neuronal oscillations that occur in the gamma
frequency band. By contrast, the right hemisphere has a slower rate of temporal integration (5 Hz),
allowing it to process signal fluctuations that occur on the order of approximately 200 ms; right
temporal activity is thought to be reflected in theta-band neuronal oscillations. Notably, the right
hemisphere preference for low-frequency modulations has been observed both with speech
(Abrams et al., 2008) and nonspeech stimuli (Boemio et al., 2005; Zatorre & Belin, 2001), sug-
gesting that asymmetric sampling is a core property of temporal cortex rather than being specific
to speech perception. Key to the AST hypothesis is the premise that the processing preferences of
the two hemispheres depend on the physical properties of the auditory signal.
The AST can readily explain an association between the right hemisphere and processing the
prosody of speech, for instance, as prosodic cues are conveyed over a relatively large temporal
window (Poeppel, 2003). However, rightward lateralization is not always observed for prosodic
processing, with the precise lateralization depending on a number of factors, including the
control task used (Kreitewolf, Friederici, & von Kriegstein, 2014). Moreover, a number of studies
have demonstrated left hemisphere involvement in prosodic processing when such information
conveys linguistic information, whether lexical (Gandour, Tong, et al., 2004; Gandour, Wong,
et al., 2002) or syntactic (van der Burght et al., 2019). In one such study, van der Burght et al.
observed robust activation of the left inferior frontal gyrus when prosodic information in a
speech sample determined syntactic structure but not when prosody was not needed for
resolving the sentence’s syntax. These results are consistent with the view that while hemi-
spheric asymmetries in processing auditory information may be partly attributable to the phys-
ical acoustic properties of the signal, the extent to which each hemisphere is involved may
also largely depend on the functional use of the signal (Van Lancker, 1980).
The functional view predicts that right hemisphere involvement in speech perception is not
limited simply to instances when listeners integrate auditory information over a long temporal
window—rather, the involvement of the right hemisphere in speech perception may specifically
reflect the process of conditioning phonetic processing on talker information (Kreitewolf,
Gaudrain, & von Kriegstein, 2014b; Luthra, Correia, et al., 2020; Myers & Mesite, 2014; Myers
& Theodore, 2017; von Kriegstein, Smith, et al., 2010). Some evidence for this hypothesis comes
from a study by von Kriegstein, Smith, et al. (2010), in which listeners heard stimulus trains that
varied in syllable identity, amplitude, and/or vocal tract length (an acoustic parameter that differs
across talkers). Listeners performed either a one-back speech task (in which they had to indicate if
the current stimulus matched the preceding stimulus in syllable identity) or a control task (either a
one-back talker task or a one-back amplitude task). The authors observed that the left posterior
STG was sensitive to vocal tract length (i.e., to acoustic information associated with talker identity).
Moreover, von Kriegstein, Smith, et al. (2010) found that during the speech task, the functional
connections between the left posterior STG and its right hemisphere analogue differed as a func-
tion of vocal tract length. The authors interpreted their findings as evidence that when listeners
process talker-specific information in support of speech recognition, both the left and right tem-
poral cortex are recruited.
Additional support for this perspective comes from a study by Myers and Theodore (2017), in
which listeners were exposed to two talkers who differed in their productions of the sound /k/.
Neurobiology of Language
145
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
Specifically, the talkers differed in whether they produced /k/ with a relatively short or long
VOT ( an acoustic-phonetic cue that distinguishes the voiceless sound / k/ from its voiced
counterpart, /g/); notably, processing VOT requires integrating over a relatively short temporal
window. After being familiarized with these two talkers, listeners completed an MRI scan during
which they performed phonetic categorization on the words “cane” and “gain”; critically, during
this phonetic categorization task, listeners heard both talker-typical and talker-atypical variants of
the word “cane.” Myers and Theodore found that the functional activation of the right STG de-
pended on whether the “cane” variant heard was typical or atypical of that talker. Such a result is
consistent with the functional view of hemispheric asymmetries, which holds that despite being
a short-duration cue, VOT would be processed by the right hemisphere if it was informative of
talker identity. Additionally, the authors observed that the more typical the acoustic-phonetic
variant was of a talker, the more tightly coupled the activity between the right STG and left tem-
poral cortex. Taken together, these findings support the perspective that the right temporal cortex
may support a listener’s ability to adapt to the idiosyncratic ways that different talkers produce
their speech sounds; this may be achieved through the activity of the right temporal cortex itself
or through interactions between the right temporal cortex and left temporal regions associated
with phonetic processing.
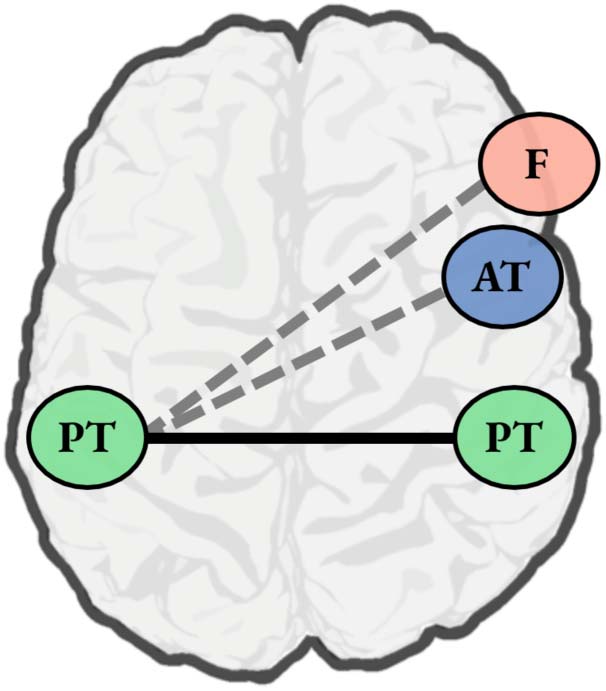
While there are documented functional connections between left posterior temporal regions
involved in phonetic processing and right posterior temporal regions involved in the early
analysis of vocal detail, there does not appear to be a strong role for functional connections be-
tween left posterior temporal regions and other right hemisphere regions associated with vocal
identity processing (Figure 2). In considering why this might be, it is worth noting that these other
regions are primarily associated with explicitly mapping vocal information to a known identity (in
the case of right anterior temporal areas) or are recruited only when listeners are tasked with
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2.
Speech perception involves interactions between left posterior temporal regions impli-
cated in phonetic processing and right posterior temporal regions associated with the perceptual
analysis of vocal information. These interactions may specifically reflect the process of conditioning
phonetic identity on talker information. In this figure, posterior temporal regions are depicted by
green circles with the label “PT,” and the established functional connection between them is indi-
cated via a solid black line. However, the literature on speech perception does not suggest a strong
role for other regions involved in vocal identity processing—namely, right anterior temporal cortex
(blue circle labeled “AT”) and right inferior/middle frontal cortex (pink circle labeled “F”). It may be
the case that these other right hemisphere regions only interact with left posterior temporal cortex
(dashed gray lines) when demands on talker processing are high.
Neurobiology of Language
146
The role of the right hemisphere in speech perception
categorizing or comparing between vocal samples (in the case of right frontal regions). That is,
these regions are only recruited when demands on vocal identity processing are high.
In ecological instances of speech perception, however, listeners may not need to make ex-
plicit judgments about talker identity; indeed, listeners can typically leverage myriad sources
of context to identify a talker’s intended phoneme, be they syntactic (Fox & Blumstein, 2016),
semantic (Borsky et al., 1998), lexical (Ganong, 1980), or visual (Frost et al., 1988; McGurk &
MacDonald, 1976). As such, the involvement of right anterior temporal and right frontal re-
gions in phonetic processing may be limited to situations where the demands on the talker
identification system is high, such that talker identity uniquely determines the mapping be-
tween acoustics and phonemes. I suggest that future studies assess this hypothesis directly,
investigating both the activation of these right hemisphere regions and their functional connec-
tions to left temporal regions involved in phonetic processing.
Furthermore, the observation that naturalistic speech perception does not necessarily place a
strong burden on talker processing systems may hint at why frank deficits in speech perception
are not observed in individuals with right hemisphere damage. I suggest that the impact of right
hemisphere damage (and damage to right posterior temporal cortex in particular) may only be
observable in tasks that specifically require listeners to condition phonetic identity on talker
information. Future work testing this hypothesis in right hemisphere patients will therefore be
important in elucidating a potential subclinical impairment.
DISCUSSION
The acoustic signal simultaneously conveys linguistic information about speech sounds as well as
nonlinguistic information about vocal identity, and in general, the process of speech perception
is not independent from processing talker information (Mullennix & Pisoni, 1990). In this review, I
have attempted to clarify the nature of right hemisphere involvement in speech perception by
focusing on its role in vocal identity processing. As depicted in Figure 1, vocal identity processing
entails the contributions of right posterior temporal cortex, right anterior temporal cortex, and
right inferior/middle frontal cortex. Based on the functional view of hemispheric contributions
to processing auditory information (Van Lancker, 1980), I presented evidence that the recruitment
of right posterior temporal regions during speech perception may reflect the process of condition-
ing phonetic identity on talker information. I noted that right anterior temporal and right frontal
regions are not strongly implicated during speech perception (Figure 2), and I suggested that the
limited involvement of these regions may reflect the fact that in ecological speech perception,
demands on talker processing are relatively low. In closing, I suggest that our understanding of
the role of the right hemisphere in speech perception may be improved by focusing specifically
on conditions where demands on talker processing are high (e.g., when a listener must appeal to
talker information in order to know how to map the speech signal onto phonetic categories).
Future work of this sort may also elucidate potential subclinical impairments in speech percep-
tion in individuals who have sustained damage to the right hemisphere.
ACKNOWLEDGMENTS
I am thankful to Emily Myers, Jim Magnuson, Rachel Theodore, Gerry Altmann, Eiling Yee,
Jonathan Peelle, and three anonymous reviewers for their feedback on previous versions of this
manuscript. This work was supported by an NSF Graduate Research Fellowship awarded to the
author. The publication of this work was supported by the program in Science of Learning & Art of
Communication at the University of Connecticut, which is supported by the National Science
Foundation under Grant DGE-1747486.
Neurobiology of Language
147
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
FUNDING INFORMATION
Sahil Luthra, National Science Foundation (http://dx.doi.org/10.13039/501100008982), Award
ID: Graduate Research Fellowship. James S. Magnuson, National Science Foundation (http://dx
.doi.org/10.13039/501100008982), Award ID: NRT 1747486.
REFERENCES
Abrams, D. A., Nicol, T., Zecker, S., & Kraus, N. (2008). Right-
hemisphere auditory cortex is dominant for coding syllable patterns
in speech. The Journal of Neuroscience, 28(15), 3958–3965. DOI:
https://doi.org/10.1523/ JNEUROSCI.0187-08.2008, PMID:
18400895, PMCID: PMC2713056
Allen, J. S., & Miller, J. L. (2004). Listener sensitivity to individual talker
differences in voice-onset-time. The Journal of the Acoustical
Society of America, 115(6), 3171–3183. DOI: https://doi.org/10
.1121/1.1701898, PMID: 15237841
Allen, J. S., Miller, J. L., & DeSteno, D. (2003). Individual talker differ-
ences in voice-onset-time. The Journal of the Acoustical Society of
America, 113(1), 544–552. DOI: https://doi.org/10.1121/1
.1528172, PMID: 12558290
Andics, A., McQueen, J. M., & Petersson, K. M. (2013). Mean-based
neural coding of voices. NeuroImage, 79, 351–360. DOI: https://
doi.org/10.1016/j.neuroimage.2013.05.002, PMID: 23664949
Andics, A., McQueen, J. M., Petersson, K. M., Gál, V., Rudas, G., &
Vidnyánszky, Z. (2010). Neural mechanisms for voice recogni-
tion. NeuroImage, 52(4), 1528–1540. DOI: https://doi.org/10
.1016/j.neuroimage.2010.05.048, PMID: 20553895
Belin, P. (2006). Voice processing in human and non-human primates.
Philosophical Transactions of the Royal Society B: Biological
Sciences, 361(1476), 2091–2107. DOI: https://doi.org/10.1098
/rstb.2006.1933, PMID: 17118926, PMCID: PMC1764839
Belin, P., Fecteau, S., & Bédard, C. (2004). Thinking the voice: Neural
correlates of voice perception. Trends in Cognitive Sciences, 8(3),
129–135. DOI: https://doi.org/10.1016/j.tics.2004.01.008, PMID:
15301753
Belin, P., McAdams, S., Smith, B., Savel, S., Thivard, L., Samson, S.,
& Samson, Y. (1998). The functional anatomy of sound intensity
discrimination. Journal of Neuroscience, 18(16), 6388–6394.
DOI: https://doi.org/10.1523/ JNEUROSCI.18-16-06388.1998,
PMID: 9698330, PMCID: PMC6793181
Belin, P., & Zatorre, R. J. (2003). Adaptation to speaker’s voice in
right anterior temporal lobe. Neuroreport, 14(16), 2105–2109.
DOI: https://doi.org/10.1097/00001756-200311140-00019,
PMID: 14600506
Belin, P., Zatorre, R. J., Hoge, R., Evans, A. C., & Pike, B. (1999).
Event-related fMRI of the auditory cortex. NeuroImage, 10(4),
417–429. DOI: https://doi.org/10.1006/nimg.1999.0480, PMID:
10493900
Belin, P., Zatorre, R. J., Lafaille, P., Ahad, P., & Pike, B. (2000).
Voice-selective areas in human auditory cortex. Nature, 403
(6767), 309–312. DOI: https://doi.org/10.1038/35002078,
PMID: 10659849
Bestelmeyer, P. E. G., Belin, P., & Grosbras, M.-H. (2011). Right
temporal TMS impairs voice detection. Current Biology, 21(20),
R838–R839. DOI: https://doi.org/10.1016/j.cub.2011.08.046,
PMID: 22032183
Binder, J. R., Frost, J. A., Hammeke, T. A., Cox, R. W., Rao, S. M., &
Prieto, T. (1997). Human brain language areas identified by func-
tional magnetic resonance imaging. The Journal of Neuroscience,
17(1), 353–362. DOI: https://doi.org/10.1523/JNEUROSCI.17-01
-00353.1997, PMID: 8987760, PMCID: PMC6793702
Binder, J. R., Swanson, S. J., Hammeke, T. A., Morris, G. L.,
Mueller, W. M., Fischer, M., Benbadis, S., Frost, J. A., Rao, S. M.,
& Haughton, V. M. (1996). Determination of language dominance
using functional MRI. Neurology, 46, 978–984. DOI: https://doi
.org/10.1212/ WNL.46.4.978, PMID: 8780076
Blumstein, S. E., & Myers, E. B. (2014). Neural systems underlying
speech perception. In K. N. Ochsner & S. Kosslyn (Eds.), The
Oxford Handbook of Cognitive Neuroscience, Volume 1
(pp. 507–523). Oxford University Press. DOI: https://doi.org
/10.1093/oxfordhb/9780199988693.013.0025
Blumstein, S. E., Myers, E. B., & Rissman, J. (2005). The perception
of voice onset time: An fMRI investigation of phonetic category
structure. Journal of Cognitive Neuroscience, 17(9), 1353–1366.
DOI: https://doi.org/10.1162/0898929054985473, PMID:
16197689
Boatman, D., Hart, J., Lesser, R. P., Honeycutt, N., Anderson, N. B.,
Miglioretti, D., & Gordon, B. (1998). Right hemisphere speech
perception revealed by amobarbital injection and electrical inter-
ference. Neurology, 51(2), 458–464. DOI: https://doi.org/10
.1212/ WNL.51.2.458, PMID: 9710019
Boemio, A., Fromm, S., Braun, A., & Poeppel, D. (2005).
Hierarchical and asymmetric temporal sensitivity in human auditory
cortices. Nature Neuroscience, 8(3), 389–395. DOI: https://doi.org
/10.1038/nn1409, PMID: 15723061
Borsky, S., Tuller, B., & Shapiro, L. P. (1998). “How to milk a coat:”
The effects of semantic and acoustic information on phoneme
categorization. The Journal of the Acoustical Society of America,
103(5), 2670–2676. DOI: https://doi.org/10.1121/1.422787,
PMID: 9604360
Broca, P. (1861). Remarques sur le siège de la faculté du langage
articulé, suivies d’une observation d’aphémie (perte de la parole).
Bulletin et Memoires de La Société Anatomique de Paris, 6,
330–357.
Burton, M. W., Small, S. L., & Blumstein, S. E. (2000). The role of
segmentation in phonological processing: An fMRI investigation.
Journal of Cognitive Neuroscience, 12(4), 679–690. DOI: https://
doi.org/10.1162/089892900562309, PMID: 10936919
Clayards, M., Tanenhaus, M. K., Aslin, R. N., & Jacobs, R. A.
(2008). Perception of speech reflects optimal use of probabilistic
speech cues. Cognition, 108(3), 804–809. DOI: https://doi.org
/10.1016/j.cognition.2008.04.004, PMID: 18582855, PMCID:
PMC2582186
Damasio, A. (1990). Face agnosia and the neural substrates of
memory. Annual Review of Neuroscience, 13(1), 89–109. DOI:
https://doi.org/10.1146/annurev.ne.13.030190.000513, PMID:
2183687
Davis, M. H., Ford, M. A., Kherif, F., & Johnsrude, I. S. (2011). Does
semantic context benefit speech understanding through “top–down”
processes? Evidence from time-resolved sparse fMRI. Journal of
Cognitive Neuroscience, 23(12), 3914–3932. DOI: https://doi.org
/10.1162/jocn_a_00084, PMID: 21745006
Desai, R., Liebenthal, E., Waldron, E., & Binder, J. R. (2008). Left
posterior temporal regions are sensitive to auditory categorization.
Journal of Cognitive Neuroscience, 20(7), 1174–1188. DOI:
Neurobiology of Language
148
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
https://doi.org/10.1162/jocn.2008.20081, PMID: 18284339,
PMCID: PMC3350814
Formisano, E., De Martino, F., Bonte, M., & Goebel, R. (2008).
“Who” is saying “what”? Brain-based decoding of human voice
and speech. Science, 322(5903), 970–973. DOI: https://doi.org/10
.1126/science.1164318, PMID: 18988858
Fox, N. P., & Blumstein, S. E. (2016). Top-down effects of syntactic
sentential context on phonetic processing. Journal of Experimental
Psychology: Human Perception and Performance, 42(5), 730–741.
DOI: https://doi.org/10.1037/a0039965, PMID: 26689310
Frost, R., Repp, B. H., & Katz, L. (1988). Can speech perception be
influenced by simultaneous presentation of print? Journal of
Memory and Language, 27(6), 741–755. DOI: https://doi.org/10
.1016/0749-596X(88)90018-6
Gainotti, G. (2007). Different patterns of famous people recognition
disorders in patients with right and left anterior temporal lesions:
A systematic review. Neuropsychologia, 45(8), 1591–1607. DOI:
https://doi.org/10.1016/j.neuropsychologia.2006.12.013, PMID:
17275042
Gainotti, G., Barbier, A., & Marra, C. (2003). Slowly progressive
defect in recognition of familiar people in a patient with right
anterior temporal atrophy. Brain, 126(4), 792–803. DOI: https://
doi.org/10.1093/brain/awg092, PMID: 12615639
Gandour, J., Tong, Y., Wong, D., Talavage, T., Dzemidzic, M., Xu, Y.,
Li, X., & Lowe, M. (2004). Hemispheric roles in the perception of
speech prosody. NeuroImage, 23(1), 344–357. DOI: https://doi
.org/10.1016/j.neuroimage.2004.06.004, PMID: 15325382
Gandour, J., Wong, D., Lowe, M., Dzemidzic, M., Satthamnuwong,
N., Tong, Y., & Li, X. (2002). A cross-linguistic fMRI study of
spectral and temporal cues underlying phonological processing.
Journal of Cognitive Neuroscience, 14(7), 1076–1087. DOI: https://
doi.org/10.1162/089892902320474526, PMID: 12419130
Ganong, W. F. (1980). Phonetic categorization in auditory word per-
ception. Journal of Experimental Psychology: Human Perception
and Performance, 6(1), 110–125. DOI: https://doi.org/10.1037
/0096-1523.6.1.110
Geschwind, N. (1970). The organization of language and the brain.
Science, 170(3961), 940–944. DOI: https://doi.org/10.1126/science
.170.3961.940, PMID: 5475022
Giraud, A. L., Kell, C., Thierfelder, C., Sterzer, P., Russ, M. O.,
Preibisch, C., & Kleinschmidt, A. (2004). Contributions of sen-
sory input, auditory search and verbal comprehension to corti-
cal activity during speech processing. Cerebral Cortex, 14(3),
247–255. DOI: https://doi.org/10.1093/cercor/bhg124, PMID:
14754865
Goggin, J. P., Thompson, C. P., Strube, G., & Simental, L. R. (1991).
The role of language familiarity in voice identification. Memory
& Cognition, 19(5), 448–458. DOI: https://doi.org/10.3758
/BF03199567, PMID: 1956306
Heilman, K. M., Bowers, D., Speedie, L., & Branch Coslett, H.
(1984). Comprehension of affective and nonaffective prosody.
Neurology, 34(7), 917–921. DOI: https://doi.org/10.1212/ WNL
.34.7.917, PMID: 6539867
Hickok, G., & Poeppel, D. (2000). Towards a functional neuroanatomy
of speech perception. Trends in Cognitive Sciences, 4(4), 131–138.
DOI: https://doi.org/10.1016/S1364-6613(00)01463-7
Hickok, G., & Poeppel, D. (2004). Dorsal and ventral streams: A
framework for understanding aspects of the functional anatomy
of language. Cognition, 92(1–2), 67–99. DOI: https://doi.org/10
.1016/j.cognition.2003.10.011, PMID: 15037127
Hickok, G., & Poeppel, D. (2007). The cortical organization of
speech processing. Nature Reviews Neuroscience, 8(5), 393–402.
DOI: https://doi.org/10.1038/nrn2113, PMID: 17431404
Holmes, E., Domingo, Y., & Johnsrude, I. S. (2018). Familiar voices
are more intelligible, even if they are not recognized as familiar.
Psychological Science, 29(10), 1575–1583. DOI: https://doi.org
/10.1177/0956797618779083, PMID: 30096018
Holmes, E., & Johnsrude, I. S. (2020). Speech spoken by familiar
people is more resistant to interference by linguistically similar
speech. Journal of Experimental Psychology: Learning, Memory
and Cognition, 46(8), 1465–1476. DOI: https://doi.org/10
.1037/xlm0000823, PMID: 32105143
Imaizumi, S., Mori, K., Kiritani, S., Kawashima, R., Sugiura, M.,
Fukuda, H., Itoh, K., Kato, T., Nakamura, A., Hatano, K.,
Kojima, S., & Nakamura, K. (1997). Vocal identification of
speaker and emotion activates differerent brain regions.
NeuroReport, 8(12), 2809–2812. DOI: https://doi.org/10.1097
/00001756-199708180-00031, PMID: 9295122
Johnson, K. A. (2008). Speaker normalization in speech perception.
In D. B. Pisoni & R. E. Remez (Eds.), The handbook of speech
perception (pp. 363–389). Blackwell Publishing. DOI: https://
doi.org/10.1002/9780470757024.ch15
Johnsrude, I. S., Mackey, A., Hakyemez, H., Alexander, E., Trang,
H. P., & Carlyon, R. P. (2013). Swinging at a cocktail party: Voice
familiarity aids speech perception in the presence of a competing
voice. Psychological Science, 24(10), 1995–2004. DOI: https://
doi.org/10.1177/0956797613482467, PMID: 23985575
Jones, A. B., Farrall, A. J., Belin, P., & Pernet, C. R. (2015).
Hemispheric association and dissociation of voice and speech
information processing in stroke. Cortex, 71, 232–239. DOI:
https://doi.org/10.1016/j.cortex.2015.07.004, PMID: 26247409
Joos, M. (1948). Acoustic phonetics. Language, 24(2), 5–136. DOI:
https://doi.org/10.2307/522229
Kennedy-Higgins, D., Devlin, J. T., Nuttall, H. E., & Adank, P.
(2020). The causal role of left and right superior temporal gyri
in speech perception in noise: A transcranial magnetic stimula-
tion study. Journal of Cognitive Neuroscience, 32(6), 1092–1103.
DOI: https://doi.org/10.1162/jocn_a_01521, PMID: 31933438
Kleinschmidt, D. F. (2019). Structure in talker variability: How much
is there and how much can it help? Language, Cognition and
Neuroscience, 34(1), 43–68. DOI: https://doi.org/10.1080
/23273798.2018.1500698, PMID: 30619905, PMCID:
PMC6320234
Kleinschmidt, D. F., & Jaeger, T. F. (2015). Robust speech percep-
tion: Recognize the familiar, generalize to the similar, and adapt
to the novel. Psychological Review, 122(2), 148–203. DOI:
https://doi.org/10.1037/a0038695, PMID: 25844873, PMCID:
PMC4744792
Kraljic, T., & Samuel, A. G. (2005). Perceptual learning for speech: Is
there a return to normal? Cognitive Psychology, 51(2), 141–178.
DOI: https://doi.org/10.1016/j.cogpsych.2005.05.001, PMID:
16095588
Kreitewolf, J., Friederici, A. D., & von Kriegstein, K. (2014).
Hemispheric lateralization of linguistic prosody recognition in
comparison to speech and speaker recognition. NeuroImage,
102(P2), 332–344. DOI: https://doi.org/10.1016/j.neuroimage
.2014.07.038, PMID: 25087482
Kreitewolf, J., Gaudrain, E., & von Kriegstein, K. (2014). A neural
mechanism for recognizing speech spoken by different speakers.
NeuroImage, 91, 375–385. DOI: https://doi.org/10.1016/j
.neuroimage.2014.01.005, PMID: 24434677
Kreitewolf, J., Mathias, S. R., & von Kriegstein, K. (2017). Implicit
talker training improves comprehension of auditory speech in
noise. Frontiers in Psychology, 8(SEP), 1–8. DOI: https://doi
.org/10.3389/fpsyg.2017.01584, PMID: 28959226, PMCID:
PMC5603660
Neurobiology of Language
149
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
Lang, C. J. G., Kneidl, O., Hielscher-Fastabend, M., & Heckmann, J. G.
(2009). Voice recognition in aphasic and non-aphasic stroke
patients. Journal of Neurology, 256(8), 1303–1306. DOI: https://
doi.org/10.1007/s00415-009-5118-2, PMID: 19353219
Lee, Y.-S., Turkeltaub, P., Granger, R., & Raizada, R. D. S. (2012).
Categorical speech processing in Broca’s area: An fMRI study using
multivariate pattern-based analysis. Journal of Neuroscience,
32(11), 3942–3948. DOI: https://doi.org/10.1523/ JNEUROSCI
.3814-11.2012, PMID: 22423114, PMCID: PMC6703443
Liebenthal, E., Desai, R., Ellingson, M. M., Ramachandran, B.,
Desai, A., & Binder, J. R. (2010). Specialization along the left
superior temporal sulcus for auditory categorization. Cerebral
Cortex, 20(12), 2958–2970. DOI: https://doi.org/10.1093/cercor
/bhq045, PMID: 20382643, PMCID: PMC2978244
Luthra, S., Correia, J. M., Kleinschmidt, D. F., Mesite, L. M., &
Myers, E. B. (2020). Lexical information guides retuning of neural
patterns in perceptual learning for speech. Journal of Cognitive
Neuroscience, 32(10), 2001–2012. DOI: https://doi.org/10
.1162/jocn_a_01612, PMID: 32662731
Luthra, S., Guediche, S., Blumstein, S. E., & Myers, E. B. (2019).
Neural substrates of subphonemic variation and lexical competition
in spoken word recognition. Language, Cognition and Neuroscience,
34(2), 141–169. DOI: https://doi.org/10.1080/23273798
.2018.1531140, PMID: 31106225, PMCID: PMC6516505
Luzzi, S., Coccia, M., Polonara, G., Reverberi, C., Ceravolo, G.,
Silvestrini, M., Fringuelli, F., Baldinelli, S., Provinciali, L., &
Gainotti, G. (2018). Selective associative phonagnosia after right
anterior temporal stroke. Neuropsychologia, 116, 154–161. DOI:
https://doi.org/10.1016/j.neuropsychologia.2017.05.016, PMID:
28506806
Maguinness, C., Roswandowitz, C., & von Kriegstein, K. (2018).
Understanding the mechanisms of familiar voice-identity recognition
in the human brain. Neuropsychologia, 116, 179–193. DOI: https://
doi.org/10.1016/j.neuropsychologia.2018.03.039, PMID: 29614253
McGurk, H., & MacDonald, J. (1976). Hearing lips and seeing voices.
Nature, 264, 746–748. DOI: https://doi.org/10.1038/264746a0,
PMID: 1012311
Mesgarani, N., Cheung, C., Johnson, K. A., & Chang, E. F. (2014).
Phonetic feature encoding in human superior temporal gyrus.
Science, 343(6174), 1006–1011. DOI: https://doi.org/10.1126
/science.1245994, PMID: 24482117, PMCID: PMC4350233
Mitchell, R. L. C., & Crow, T. J. (2005). Right hemisphere language
functions and schizophrenia: The forgotten hemisphere? Brain,
128(5), 963–978. DOI: https://doi.org/10.1093/ brain/awh466,
PMID: 15743870
Mullennix, J. W., & Pisoni, D. B. (1990). Stimulus variability and
processing dependencies in speech perception. Perception &
Psychophysics, 47(4), 379–390. DOI: https://doi.org/10.3758
/BF03210878, PMID: 2345691, PMCID: PMC3512111
Munson, B. (2007). The acoustic correlates of perceived masculinity,
perceived femininity, and perceived sexual orientation. Language
and Speech, 50(1), 125–142. DOI: https://doi.org/10.1177
/00238309070500010601, PMID: 17518106
Myers, E. B. (2007). Dissociable effects of phonetic competition
and category typicality in a phonetic categorization task: An
fMRI investigation. Neuropsychologia, 45(7), 1463–1473. DOI:
https://doi.org/10.1016/j.neuropsychologia.2006.11.005, PMID:
17178420, PMCID: PMC1876725
Myers, E. B., Blumstein, S. E., Walsh, E., & Eliassen, J. (2009).
Inferior frontal regions underlie the perception of phonetic category
invariance. Psychological Science, 20(7), 895–903. DOI: https://
doi.org/10.1111/j.1467-9280.2009.02380.x, PMID: 19515116,
PMCID: PMC2851201
Myers, E. B., & Mesite, L. M. (2014). Neural systems underlying
perceptual adjustment to non-standard speech tokens. Journal of
Memory and Language, 76, 80–93. DOI: https://doi.org/10.1016
/j.jml.2014.06.007, PMID: 25092949, PMCID: PMC4118215
Myers, E. B., & Theodore, R. M. (2017). Voice-sensitive brain networks
encode talker-specific phonetic detail. Brain and Language, 165,
33–44. DOI: https://doi.org/10.1016/j.bandl.2016.11.001, PMID:
27898342, PMCID: PMC5237402
Nakamura, K., Kawashima, R., Sugiura, M., Kato, T., Nakamura, A.,
Hatano, K., Nagumo, S., Kubota, K., Fukuda, H., Ito, K., &
Kojima, S. (2001). Neural substrates for recognition of familiar
voices: A PET study. Neuropsychologia, 39(10), 1047–1054.
DOI: https://doi.org/10.1016/S0028-3932(01)00037-9
Newman, R. S., & Evers, S. (2007). The effect of talker familiarity on
stream segregation. Journal of Phonetics, 35(1), 85–103. DOI:
https://doi.org/10.1016/j.wocn.2005.10.004
Norris, D., McQueen, J. M., & Cutler, A. (2003). Perceptual learning
in speech. Cognitive Psychology, 47(2), 204–238. DOI: https://doi
.org/10.1016/S0010-0285(03)00006-9
Nygaard, L. C., & Pisoni, D. B. (1998). Talker-specific learning in
speech perception. Perception and Psychophysics, 60(3), 355–376.
DOI: https://doi.org/10.3758/BF03206860, PMID: 9599989
Perrachione, T. K., Pierrehumbert, J. B., & Wong, P. C. M. (2009).
Differential neural contributions to native- and foreign-language
talker identification. Journal of Experimental Psychology: Human
Perception and Performance, 35(6), 1950–1960. DOI: https://doi
.org/10.1037/a0015869, PMID: 19968445, PMCID: PMC2792570
Perrachione, T. K., & Wong, P. C. M. (2007). Learning to recognize
speakers of a non-native language: Implications for the functional
organization of human auditory cortex. Neuropsychologia, 45(8),
1899–1910. DOI: https://doi.org/10.1016/j.neuropsychologia
.2006.11.015, PMID: 17258240
Perrodin, C., Kayser, C., Abel, T. J., Logothetis, N. K., & Petkov, C. I.
(2015). Who is that? Brain networks and mechanisms for identi-
fying individuals. Trends in Cognitive Sciences, 19(12), 783–796.
DOI: https://doi.org/10.1016/j.tics.2015.09.002, PMID:
26454482, PMCID: PMC4673906
Peterson, G. E., & Barney, H. L. (1952). Control methods used in a study
of the vowels. The Journal of the Acoustical Society of America,
24(2), 175–184. DOI: https://doi.org/10.1121/1.1906875
Poeppel, D. (2003). The analysis of speech in different temporal inte-
gration windows: Cerebral lateralization as “asymmetric sampling
in time.” Speech Communication, 41(1), 245–255. DOI: https://
doi.org/10.1016/S0167-6393(02)00107-3
Rauschecker, J. P., & Scott, S. K. (2009). Maps and streams in the
auditory cortex: Nonhuman primates illuminate human speech
processing. Nature Neuroscience, 12(6), 718–724. DOI: https://
d o i . o r g / 1 0 . 1 0 3 8 / n n . 2 3 3 1 , P M I D : 1 9 4 7 1 2 7 1 , P M C I D :
PMC2846110
Robin, D. A., Tranel, D., & Damasio, H. (1990). Auditory percep-
tion of temporal and spectral events in patients with focal left and
right cerebral lesions. Brain and Language, 39(4), 539–555. DOI:
https://doi.org/10.1016/0093-934X(90)90161-9
Rogers, J. C., & Davis, M. H. (2018). Inferior frontal cortex contri-
butions to the recognition of spoken words and their constituent
speech sounds. Journal of Cognitive Neuroscience, 29(5), 919–936.
DOI: https://doi.org/10.1162/jocn_a_01096, PMID: 28129061,
PMCID: PMC6635126
Ross, L. A., McCoy, D., Wolk, D. A., Branch Coslett, H., & Olson, I. R.
(2010). Improved proper name recall by electrical stimulation of the
anterior temporal lobes. Neuropsychologia, 48(12), 3671–3674.
DOI: https://doi.org/10.1016/j.neuropsychologia.2010.07.024,
PMID: 20659489
Neurobiology of Language
150
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
The role of the right hemisphere in speech perception
Roswandowitz, C., Kappes, C., Obrig, H., & Von Kriegstein, K.
(2018). Obligatory and facultative brain regions for voice-identity
recognition. Brain, 141(1), 234–247. DOI: https://doi.org/10
.1093/brain/awx313, PMID: 29228111, PMCID: PMC5837691
Schall, S., Kiebel, S. J., Maess, B., & von Kriegstein, K. (2014). Voice
identity recognition: Functional division of the right STS and its
behavioral relevance. Journal of Cognitive Neuroscience, 27(2),
280–291. DOI: https://doi.org/10.1162/jocn_a_00707, PMID:
25170793
Schmidt, G. L., DeBuse, C. J., & Seger, C. A. (2007). Right hemisphere
metaphor processing? Characterizing the lateralization of seman-
tic processes. Brain and Language, 100(2), 127–141. DOI: https://
doi.org/10.1016/j.bandl.2005.03.002, PMID: 17292739
Schwartz, J., & Tallal, P. (1980). Rate of acoustic change may
underlie hemispheric specalization for speech perception.
Science, 207(4437), 1380–1381. DOI: https://doi.org/10.1126
/science.7355297, PMID: 7355297
Scott, S. K., Blank, C. C., Rosen, S., & Wise, R. J. S. (2000). Identifica-
tion of a pathway for intelligible speech in the left temporal lobe.
Brain, 123(12), 2400–2406. DOI: https://doi.org/10.1093/brain
/123.12.2400, PMID: 11099443, PMCID: PMC5630088
Siegal, M., Carrington, J., & Radel, M. (1996). Theory of mind and
pragmatic understanding following right hemisphere damage.
Brain and Language, 53(1), 40–50. DOI: https://doi.org/10.1006
/brln.1996.0035, PMID: 8722898
Souza, P., Gehani, N., Wright, R., & McCloy, D. (2013). The advan-
tage of knowing the talker. Journal of the American Academy of
Audiology, 24(8), 689–700. DOI: https://doi.org/10.3766/jaaa
.24.8.6, PMID: 24131605, PMCID: PMC3801269
Stevens, A. A. (2004). Dissociating the cortical basis of memory for
voices, words and tones. Cognitive Brain Research, 18(2), 162–171.
DOI: https://doi.org/10.1016/j.cogbrainres.2003.10.008, PMID:
14736575
Theodore, R. M., & Monto, N. R. (2019). Distributional learning for
speech reflects cumulative exposure to a talker’s phonetic distri-
butions. Psychonomic Bulletin and Review, 26(3), 985–992.
DOI: https://doi.org/10.3758/s13423-018-1551-5, PMID:
30604404, PMCID: PMC6559869
Tranel, D., Damasio, H., & Damasio, A. R. (1997). A neural basis
for the retrieval of conceptual knowledge. Neuropsychologia,
35(10), 1319–1327. DOI: https://doi.org/10.1016/S0028-3932(97)
00085-7
Turkeltaub, P. E., & Branch Coslett, H. (2010). Localization of sub-
lexical speech perception components. Brain and Language, 114(1),
1–15. DOI: https://doi.org/10.1016/j.bandl.2010.03.008, PMID:
20413149, PMCID: PMC2914564
van der Burght, C. L., Goucha, T., Friederici, A. D., Kreitewolf, J., &
Hartwigsen, G. (2019). Intonation guides sentence processing in
the left inferior frontal gyrus. Cortex, 117, 122–134. DOI: https://
doi.org/10.1016/j.cortex.2019.02.011, PMID: 30974320
Van Lancker, D. R. (1980). Cerebral lateralization of pitch cues in
the linguistic signal. Papers in Linguistics, 13(2), 201–277. DOI:
https://doi.org/10.1080/08351818009370498
Van Lancker, D. R., & Canter, G. J. (1982). Impairment of voice and
face recognition in patients with hemispheric damage. Brain and
Cognition, 1(2), 185–195. DOI: https://doi.org/10.1016/0278
-2626(82)90016-1
Van Lancker, D. R., & Kreiman, J. (1987). Voice discrimination
and recognition are separate abilities. Neuropsychologia, 25(5),
829–834. DOI: https://doi.org/10.1016/0028-3932(87)90120-5
von Kriegstein, K., Eger, E., Kleinschmidt, A., & Giraud, A. L. (2003).
Modulation of neural responses to speech by directing attention to
voices or verbal content. Cognitive Brain Research, 17(1), 48–55.
DOI: https://doi.org/10.1016/S0926-6410(03)00079-X
von Kriegstein, K., & Giraud, A. L. (2004). Distinct functional sub-
strates along the right superior temporal sulcus for the processing
of voices. NeuroImage, 22(2), 948–955. DOI: https://doi.org/10
.1016/j.neuroimage.2004.02.020, PMID: 15193626
von Kriegstein, K., Smith, D. R., Patterson, R. D., Kiebel, S. J., &
Griffiths, T. D. (2010). How the human brain recognizes speech
in the context of changing speakers. Journal of Neuroscience, 30(2),
629–638. DOI: https://doi.org/10.1523/ JNEUROSCI.2742
-09.2010, PMID: 20071527, PMCID: PMC2824128
Wada, J., & Rasmussen, T. (1960). Intracarotid injection of sodium
amytal for the lateralization of cerebral speech dominance:
Experimental and clinical observations. Journal of Neurosurgery,
17(2), 266–282. DOI: https://doi.org/10.3171/jns.1960.17.2.0266
Wernicke, C. (1874). Der aphasische Symptomencomplex: Eine
psychologische Studie auf anatomischer Basis. Cohn.
Xie, X., & Myers, E. B. (2018). Left inferior frontal gyrus sensitivity to
phonetic competition in receptive language processing: A com-
parison of clear and conversational speech. Journal of Cognitive
Neuroscience, 30(3), 267–280. DOI: https://doi.org/10.1162/jocn
_a_01208, PMID: 29160743
Yi, H. G., Leonard, M. K., & Chang, E. F. (2019). The encoding of
speech sounds in the superior temporal gyrus. Neuron, 102(6),
1096–1110. DOI: https://doi.org/10.1016/j.neuron.2019.04.023,
PMID: 31220442, PMCID: PMC6602075
Zäske, R., Awwad Shiekh Hasan, B., & Belin, P. (2017). It doesn’t
matter what you say: fMRI correlates of voice learning and recog-
nition independent of speech content. Cortex, 94, 100–112. DOI:
https://doi.org/10.1016/j.cortex.2017.06.005, PMID: 28738288,
PMCID: PMC5576914
Zatorre, R. J., & Belin, P. (2001). Spectral and temporal processing in
human auditory cortex. Cerebral Cortex, 11(10), 946–953. DOI:
https://doi.org/10.1093/cercor/11.10.946, PMID: 11549617
Zatorre, R. J., Meyer, E., Gjedde, A., & Evans, A. C. (1996). PET
studies of phonetic processing of speech: Review, replication,
and reanalysis. Cerebral Cortex, 6(1), 21–30. DOI: https://doi.org
/10.1093/cercor/6.1.21, PMID: 8670635
Neurobiology of Language
151
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
2
1
1
3
8
1
9
0
8
8
7
5
n
o
_
a
_
0
0
0
2
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3