REVIEW
The road ahead in clinical
network neuroscience
Linda Douw1,∗
, Edwin van Dellen2,3,∗
, Alida A. Gouw4,5, Alessandra Griffa6, Willem de Haan4,5,
Martijn van den Heuvel6,7, Arjan Hillebrand4, Piet Van Mieghem8, Ida A. Nissen4,
Willem M. Otte9,10, Yael D. Reijmer11, Menno M. Schoonheim1, Mario Senden12,13,
Elisabeth C. W. van Straaten4, Betty M. Tijms5,
Prejaas Tewarie4, and Cornelis J. Stam4
1Department of Anatomy and Neuroscience, Amsterdam Neuroscience, Vrije Universiteit Amsterdam, Amsterdam UMC,
Amsterdam, The Netherlands
2Department of Psychiatry, Brain Center, University Medical Center Utrecht, Utrecht, The Netherlands
3Melbourne Neuropsychiatry Centre, University of Melbourne and Melbourne Health, Melbourne, Australia
4Department of Neurology, Clinical Neurophysiology and MEG Center, Amsterdam Neuroscience, Vrije Universiteit
Amsterdam, Amsterdam UMC, Amsterdam, The Netherlands
5Alzheimer Center Amsterdam, Department of Neurology, Amsterdam Neuroscience, Vrije Universiteit Amsterdam,
Amsterdam UMC, Amsterdam, The Netherlands
6Connectome Lab, Department of Neuroscience, section Complex Trait Genetics, Center for Neurogenomics and Cognitive
Research, Amsterdam Neuroscience, Vrije Universiteit Amsterdam, Amsterdam UMC, Amsterdam, The Netherlands
7Department of Clinical Genetics, Amsterdam Neuroscience, Vrije Universiteit Amsterdam, Amsterdam UMC,
Amsterdam, The Netherlands
8Faculty of Electrical Engineering, Mathematics and Computer Science, Delft University of Technology, Delft, The Netherlands
9Biomedical MR Imaging and Spectroscopy Group, Center for Image Sciences, University Medical Center Utrecht and Utrecht
University, Utrecht, The Netherlands
10Department of Pediatric Neurology, Brain Center, University Medical Center Utrecht,
Utrecht, The Netherlands
11Department of Neurology, Brain Center, University Medical Center Utrecht, Utrecht, the Netherlands
12Department of Cognitive Neuroscience, Faculty of Psychology and Neuroscience, Maastricht University, Maastricht,
The Netherlands
13Maastricht Brain Imaging Centre, Faculty of Psychology and Neuroscience, Maastricht University, Maastricht,
The Netherlands
Authors contributed equally.
∗
Keywords: Connectome, Graph analysis, Neuroimaging, Neurophysiology, Computational modeling,
Network neuroscience, Clinical application
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
ABSTRACT
Clinical network neuroscience, the study of brain network topology in neurological and
psychiatric diseases, has become a mainstay field within clinical neuroscience. Being a
multidisciplinary group of clinical network neuroscience experts based in The Netherlands,
we often discuss the current state of the art and possible avenues for future investigations.
These discussions revolve around questions like “How do dynamic processes alter the
underlying structural network?” and “Can we use network neuroscience for disease
classification?” This opinion paper is an incomplete overview of these discussions and
expands on ten questions that may potentially advance the field. By no means intended
as a review of the current state of the field, it is instead meant as a conversation starter
and source of inspiration to others.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
What are the large-scale network principles governing neuronal communication, cognition,
and ultimately human behavior? In history, there have been two main views on the neural
correlates of behavior. The first stipulates that the brain consists of separate parts or compo-
nents, each responsible for a particular function. Proponents of this localizationist view were
a n o p e n a c c e s s
j o u r n a l
Citation: Douw, L., van Dellen, E.,
Gouw, A. A., Griffa, A., de Haan, W.,
van den Heuvel, M., . . . Stam, C. J.
(2019). The road ahead in clinical
network neuroscience. Network
Neuroscience, 3(4), 969–993.
https://doi.org/10.1162/netn_a_00103
DOI:
https://doi.org/10.1162/netn_a_00103
Received: 24 January 2019
Accepted: 23 July 2019
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Linda Douw
l.douw@amsterdamumc.nl
Edwin van Dellen
E.vanDellen-2@umcutrecht.nl
Handling Editor:
Alex Fornito
Copyright: © 2019
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
The road ahead in clinical network neuroscience
for instance Franz Joseph Gall and Paul Broca. The opposing view has centered on the unitary,
integrative nature of the brain, assuming it impossible to attribute particular functions to either
structure or function of particular brain regions. This view was supported by for instance Jean
Pierre Flourens and Karl Lashley. The very existence of the journal in which this manuscript
is published indicates a new era in neuroscience: After centuries of exploration of these op-
posing views separately, network neuroscience offers a mathematical framework and model
of the brain that combines global integration and local specialization in both structural and
functional networks. This framework thereby may be exclusively able to combine the best of
both worlds in a quantitative and theory-governed manner.
The seminal work of Watts and Strogatz on the structural neuronal organization of the nema-
tode Caenorhabditis elegans together with the introduction of power-law and scale-free graphs
by Barabasi and Albert (Barabasi & Albert, 1999; Watts & Strogatz, 1998) have founded this
new research field, and the number of published works in the field of network neuroscience
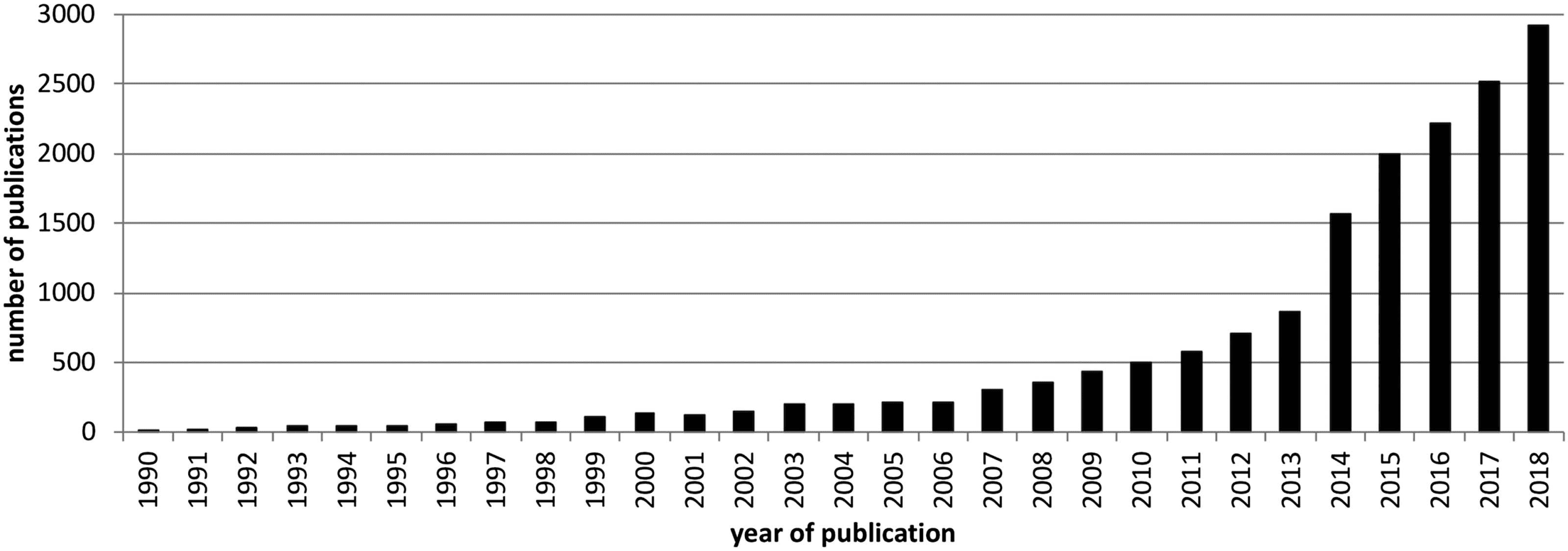
has been on a steep growth curve ever since (see Figure 1). Recent volumes and reviews have
drawn up the current state of the art from multiple perspectives: There are ample overviews of
what is currently known about network neuroscience in the context of methodology and net-
work organization (Fornito, Zalesky, & Bullmore, 2016; Sporns, 2010), computational model-
ing (Bassett, Zurn, & Gold, 2018), and clinical studies (Crossley et al., 2014; Fornito, Bullmore,
& Zalesky, 2017; O’Neill et al., 2018; Stam, 2014). However, there is still a need to tackle new
challenges and develop novel approaches for future clinical brain network studies.
In this paper, we aim to identify key questions in clinical network neuroscience in the style
initiated by Hilbert (the famous mathematical agenda; Hilbert, 1902) and more recently used
by Stephan, Bach et al. (2016) and Stephan, Binder et al. (2016). It is the product of discus-
sions of the informal group of Netherlands Network Neuroscientists (N3), a multidisciplinary
collection of scientists who have been working on network neuroscience since its inception.
This paper and the questions we address here are by no means meant to be complete or rep-
resentative and are not intended as a review of the field. Rather, the work is a collection of
views on the current state of the field and the most promising and interesting research ques-
tions for the near future (see Table 1 for an overview of questions addressed). It naturally falls
Figure 1. Number of publications on network neuroscience per year between 1990 and 2018
Network Neuroscience
970
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
Table 1. Overview of key questions in this piece
Questions
Part I: Theoretical, methodological, and conceptual challenges
How can we overcome methodological hurdles towards reliable and reproducible applications
of clinical network neuroscience?
How do dynamic processes alter the underlying structural network?
What is the role of time-varying dynamics in structure-function coupling?
How can combined computational modeling and experimental work be used in clinical applications?
What are the implications of directionality in the macroscopic brain network?
Part II: Clinical challenges
How can we increase our understanding of disease through network trajectories?
What is needed to use brain network characteristics as biomarkers?
Can we use network neuroscience for disease classification?
Can we systematically bridge the gap between brain network interventions in silico and in vivo?
Are evolution and dissolution driving factors in disease connectomics?
into two parts: a theoretical section on structural and (dynamic) functional network topology,
and a section on future applications of network neuroscience to clinical practice. Our hope
is that this paper will spark ideas and facilitate discussions and may be used for inspiration by
network neuroscientists, clinicians, and other interested readers.
PART I: THEORETICAL, METHODOLOGICAL, AND CONCEPTUAL CHALLENGES
How can we overcome methodological hurdles towards reliable and reproducible applications of clinical
network neuroscience?
An initial major hurdle for clinical network neuroscience is reliability of measures used
(Colclough et al., 2016; Jin, Seol, Kim, & Chung, 2011; Noble et al., 2017; Sinke et al.,
2018) and reproducibility within subjects (Deuker et al., 2009; Garcés, Martín-Buro, & Maestú,
2016; Hallquist & Hillary, 2019; Telesford et al., 2010). Current methodological challenges in
neuroimaging and neurophysiology studies are found on multiple levels, including general
neuroimaging-related methodological and statistical issues (Eklund, Nichols, & Knutsson, 2016;
Poldrack et al., 2017), as well as (homogenization of) preprocessing pipelines tailored to ac-
curate estimation of both structural and functional connections (Esteban et al., 2019; Kale,
Zalesky, Gollo, & Sporns, 2018; Maier-Hein et al., 2017). All these methodological choices
currently lack a gold standard, and are rightfully tailored to individual research questions, re-
lating to which some pitfalls are more important than others. Overall, however, the multitude
of processing choices diminishes comparability between studies, as different studies almost
never use the exact same methodology at all levels, ranging from scanning or recording to
structural or functional network reconstruction. As a result, network neuroscience studies are
generally weakly reproducible and have limited reliability, while rapid methodological innova-
tions in the field may question the validity of studies performed only a few years ago. Network
neuroscientists need to continuously inform themselves about the current state of the art and
incorporate advances in overarching methodologies.
Apart from these general methodological challenges, network neuroscience has several
methodological challenges that are specific to the field. Generally, longer recording times
for functional connectivity studies based on resting-state functional MRI (rsfMRI), electroen-
cephalography (EEG), and magnetoencephalography (MEG) may increase reliability and
Network Neuroscience
971
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
reproducibility of findings (Birn et al., 2013; Fraschini et al., 2016; Liuzzi et al., 2017), but
are possibly limitedly available in practice, particularly in patient populations. Additionally,
network neuroscience generally makes use of predefined node definitions that have been crit-
ically evaluated in recent years, for example concerning the validity of standard brain atlases
(Power, Schlaggar, Lessov-Schlaggar, & Petersen, 2013; Zalesky, Fornito, & Bullmore, 2010).
Edge definitions also have inherent issues, such as the optimal measure to quantify structural
and functional connectivity per modality (Bonilha et al., 2015; Colclough et al., 2016) and the
issue of thresholding approaches, which impacts the estimated topology of the reconstructed
functional (van Wijk, Stam, & Daffertshofer, 2010) or structural network (Maier-Hein et al.,
2017). Thresholding techniques may also induce more noise to the network, further compli-
cating their use in clinical practice (Zalesky et al., 2016).
Whereas much effort and literature are currently directed at developing better data-driven,
computational methods to improve analysis pipelines, theory-inspired approaches may also
aid in increasing reliability and reproducibility of network neuroscience. With respect to thresh-
olding for instance, more data-driven approaches include efficiency cost optimization, propor-
tional thresholding, and probabilistic thresholding (De Vico Fallani, Latora, & Chaves, 2017;
van den Heuvel et al., 2017; Vasa, Bullmore, & Patel, 2018). A more network-based theoretical
strategy towards thresholding is the minimum spanning tree. The idea of this approach is that
analysis of a core subgraph or “backbone” may overcome some of the issues of reproducibility
and within-subject variability, particularly in heterogeneous populations (Stam et al., 2014;
van Dellen et al., 2018), while recapitulating the most important aspects of the underlying sys-
tem (Tewarie et al., 2015). Keeping these analysis- and preprocessing-related caveats in mind,
we now proceed with more conceptual perspectives in the remainder of this review.
How do dynamic processes alter the underlying structural network?
The brain is a highly plastic organ, which constantly adapts to varying input from the environ-
ment and from its own internal perturbations. Many, often complex, molecular mechanisms are
involved in homeostatic synaptic plasticity (Pozo & Goda, 2010). In addition, structural plas-
ticity plays a crucial role in maintaining the overall organization of efficient brain networks
(Butz & van Ooyen, 2013; Fauth & Tetzlaff, 2016; Yin & Yuan, 2015). Up to now (dynamic)
functional connections are usually seen as an effect of the underlying structural connectivity.
However, functional communication may also impact the structural connections between any
set of brain regions. Studies using techniques such as transcranial direct current stimulation
(tDCS) or transcranial magnetic stimulation (TMS), which intercede on the functional commu-
nication between neurons noninvasively, have reported on alterations in structural connectivity
following functional stimulation (Lindenberg, Nachtigall, Meinzer, Sieg, & Flöel, 2013; Zheng
& Schlaug, 2015). This type of functional modulatory influence is also likely to play an im-
portant role in the plasticity of brain networks during development, learning, and response
to disease. In addition, understanding and predicting the effect of various treatment interven-
tions on multimodal brain networks could benefit from a better understanding of the brain as
an “adaptive network,” where the structural network changes as a result of activity or other
dynamic processes (de Haan, van Straaten, Gouw, & Stam, 2017).
Network science considers a network as consisting of two essential ingredients: (a) a struc-
ture (or graph, topology) that can be associated with “hardware” and (b) a function (or process,
related to “software”). This duality between structure and function distinguishes network sci-
ence from graph theory, which mainly studies the organization of a fixed graph, and systems
Network Neuroscience
972
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
theory that focuses on dynamic processes. A better understanding of plasticity of brain net-
works, having both structural and functional components, can be obtained by developing ap-
propriate computational models of adaptive networks, where the function interacts with the
structure and vice versa. While adaptive network models are challenging to define because of
the interaction of two types of dynamics (i.e., of the nodes and the links) on different timescales,
a number of studies have shown the feasibility of this approach. A particularly popular class
of dynamic processes on networks, called the “local rule–global emergent behavior” (LrGe)
class, describes the collective action of the local rules executed at each node that give rise to
complex, emergent global behavior (Van Mieghem & van de Bovenkamp, 2015). Some exam-
ples of the LrGe class are epidemic models such as susceptible-infected-susceptible (SIS) and
susceptible-infected-recovered (SIR) models and general reaction-diffusion processes (Pastor-
Satorras, Castellano, Van Mieghem, & Vespignani, 2015). Adaptive LrGe models may also be
a powerful class of model for use in network neuroscience.
So far, more biologically informed models have been used to infer global dynamics from
local properties. Indeed, a model of coupled neural masses using a combination of func-
tional (synaptic) and structural plasticity could explain the emergence of modularity and time-
dependent recovery from lesions (Stam, Hillebrand, Wang, & Van Mieghem, 2010). A model
of homeostatic structural plasticity was introduced by Butz and colleagues, and was shown to
be effective in explaining recovery from brain lesions (Butz, van Ooyen, & Worgotter, 2009).
It has also been shown that including some form of synaptic plasticity may improve the fit
of models of fMRI BOLD signals or oscillatory MEG signals to empirical data (Hellyer, Jachs,
Clopath, & Leech, 2016; Rocha, Koçillari, Suweis, Corbetta, & Maritan, 2018). Papadopoulos
and colleagues showed that plasticity in a Kuramoto model of coupled oscillators resulted
in complex topology of the structural and functional networks (Papadopoulos, Kim, Kurths, &
Bassett, 2017). In a computational model of Alzheimer’s disease, inclusion of activity-dependent
loss of synaptic strength could explain various features of the evolution of the disease pro-
cess and the possible functional response to different types of treatment (de Haan, Mott, van
Straaten, Scheltens, & Stam, 2012; de Haan et al., 2017).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
Typically, computational models of plasticity in brain networks involve systems of coupled
harmonic oscillators or neural mass models. Such systems require nonlinear stochastic differ-
ential equations and are difficult to analyze mathematically. However, dynamic processes that
change the underlying graph have also been studied with more simplistic models. The sim-
plest and most tractable model of the above-described LrGe class is the SIS epidemic spread
on a graph, in which nodes can be in two states, namely infected or healthy. The local rule is
twofold: If a node is infected, (a) it can infect its direct healthy neighbors with infection rate
β, and (b) it can heal with rate δ. Both the infection and the healing or recovery process are
independent. These simple rules lead to complex emergent, global network dynamics charac-
terized by two regimes after a sufficiently long time: either an endemic regime develops, in
which the epidemic is still active throughout the network, or a healthy regime exists, where the
epidemic has disappeared from the network. The two regimes are separated by a sharp phase
transition at a precise ratio β/δ close to the inverse of the spectral radius of the adjacency matrix
of the graph. Nearly all LrGe models feature such high-level description.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
As such, an epidemic process on a graph with activity-dependent rewiring that changes
the structural network can give rise to very rich behavior, while still allowing to obtain
analytical understanding of the model (Gross, Dommar D’lima, & Blasius, 2006). One lim-
itation of this rewiring SIS model used by Gross and colleagues was the fact that the num-
ber of links was kept constant. More recently, Guo and colleagues introduced the adaptive
Network Neuroscience
973
The road ahead in clinical network neuroscience
susceptible-infected-susceptible (ASIS) model on graphs (Guo, Trajanovski, van de Bovenkamp,
Wang, & Van Mieghem, 2013). The ASIS model does not have the limitation of a fixed density,
produces rich behavior such as a phase transition and modularity, yet still allows some analyt-
ical computation. As a next step, a modification of the ASIS model was introduced to capture
the phenomenon of (structural) Hebbian learning (Trajanovski, Guo, & Van Mieghem, 2015).
A major challenge for future studies of adaptive brain networks is its integration with more
biologically and neurophysiologically realistic models. Ideally, such models of adaptive brain
networks could be fitted to the data of individual patients, and serve as the starting point for
diagnosis, prediction of treatment effects, and prognosis (Bansal, Nakuci, & Muldoon, 2018).
What is the role of time-varying dynamics in structure-function coupling?
Structural brain network cores—sets of brain regions and connections occupying a central
topological position in the network—are believed to play a key role in facilitating commu-
nication and integration of information in the network, thus shaping functional connectivity
patterns and underpinning cognitive abilities (Avena-Koenigsberger, Misic, & Sporns, 2017;
van den Heuvel & Sporns, 2013; Zamora-López, Chen, Deco, Kringelbach, & Zhou, 2016). The
topology of the structural network and the presence of cores constrain the landscape of func-
tional dynamics taking place on the network (Boccaletti, Latora, Moreno, Chavez, & Hwang,
2006). Other properties of the structural network that shape the functional network include
the Euclidean distance between regions (Alexander-Bloch et al., 2013), the structural degrees
of functionally connected regions (Stam et al., 2016; Tewarie et al., 2014), and detours along
the shortest paths in the structural network (Goñi et al., 2014). Computational models that
simulate neuronal interactions on anatomical networks can reproduce the emergence of func-
tional networks (Deco, Jirsa, & McIntosh, 2011) and dynamic characteristics of neuroimaging
recordings (Hansen, Battaglia, Spiegler, Deco, & Jirsa, 2015).
Although several studies have aimed to formalize a mapping between structural and func-
tional networks, the relationship between the structural and functional dimensions in the brain
is only partially understood. While there is no one-to-one correspondence between struc-
tural and functional connectivity in the brain, static functional networks can be understood in
terms of a weighted sum of direct and indirect paths on the underlying structural networks
(Bettinardi et al., 2017; Mehta-Pandejee, Robinson, Henderson, Aquino, & Sarkar, 2017; Meier
et al., 2016; Robinson, Henderson, Matar, Riley, & Gray, 2009). However, the temporal, time-
varying dimension of functional connectivity is a key aspect of communication models and
a basic expression of mental processes (Avena-Koenigsberger et al., 2017; Chang & Glover,
2010; Hutchison et al., 2013; O’Neill et al., 2018). This dimension is ignored in most studies
that investigate the structure-function relationship in the brain, mainly because of difficulties in
measuring and quantifying dynamics (Griffa et al., 2017; Lurie et al., 2018; Vidaurre et al.,
2018). We argue that an empirical understanding of the relation between structural network
topology and dynamics of functional connectivity is a fundamental prerequisite for the devel-
opment (or refinement) of models of brain functioning.
Recent work has begun to explore how time-evolving functional networks during the rest-
ing state emerge from a single structural network (Amico et al., 2017; Cabral, Kringelbach, &
Deco, 2017; Tewarie et al., 2018). Moreover, task states have been shown to affect directed
or effective connectivity, as well as local dynamics exhibited by cortical hub regions (Senden,
Reuter, van den Heuvel, Goebel, & Deco, 2017; Senden et al., 2018). For instance, a transi-
tion from unstructured to oscillatory behavior exhibited by cortical rich-club regions (i.e., by
regions that occupy a central position in the structural network) demarcates task states from
Network Neuroscience
974
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
rest (Senden et al., 2017). The observed timescale closely matches those that characterize the
transition between structured and unstructured functional connectivity configurations as mea-
sured in other studies (Glomb, Ponce-Alvarez, Gilson, Ritter, & Deco, 2018), as well as the
evolution of performance on diverse tasks (Gilden, Thornton, & Mallon, 1995). This suggests a
complex interplay between structural network organization, local dynamics of both core and
periphery, functional network formation, and task performance.
Dynamics may vary over multiple timescales. For example, disease and task states occur
in the context of a dynamically (and periodically) varying vigilance state driven by the circa-
dian rhythm. Only a handful of studies have investigated the effect of the circadian rhythm
on functional coupling during rest in healthy populations (Blautzik et al., 2013; Hodkinson
et al., 2014; Koenis et al., 2013). Research on how the circadian rhythm affects the dynamics
of brain communication in clinical populations or during task performance appears to be en-
tirely lacking. Given that the circadian rhythm affects attention, learning, and decision-making
(cf. Schmidt, Collette, Cajochen, & Peigneux, 2007), the circadian rhythm likely constitutes
an important factor that affects brain dynamics and functional network formation in a manner
that is also dependent on the underlying structural network. Unraveling the structure-function
interplay calls for an integrative approach involving multimodal neuroimaging and computa-
tional modeling. A tangible step forward would be to report both structure and function in the
setting of within-scan and circadian variation.
How can combined computational modeling and experimental work be used in clinical applications?
Application of network analysis to human brain data may be useful, but often remains phe-
nomenological in its assessment of brain functioning. Computational modeling helps in for-
malizing the assumed relationships between structure and function that we seem to observe.
In order to identify the mechanisms that underlie the organization of (structural and functional)
brain networks and that ultimately lead to behavior, a theoretical framework is needed, based
on computational neuroscience (Bassett et al., 2018). However, unlike in physics, the feedback
loop between theory and experiment is almost nonexistent in computational network neuro-
science. To facilitate modeling studies in network neuroscience, the three steps of denotation,
demonstration, and interpretation of models in science are viewed as essential by Hughes
(1997). These three steps involve establishing a model that captures relevant aspects of the tar-
get system (denotation), studying the model analytically or numerically to demonstrate effects
within the model (demonstration), and relating these effects back to the target system as well
as validating the model by comparing (simulated) model behavior with empirical observations
of the target system (interpretation).
A multitude of simulation models are available that are based on different assumptions, pa-
rameters (and corresponding parameter spaces), and dynamics, making the first step of deno-
tation already difficult (Prinz, Bucher, & Marder, 2004). Different models can uphold different
oscillatory and network behavior (Abeysuriya et al., 2018; Ashwin, Coombes, & Nicks, 2016;
Daffertshofer, Ton, Pietras, Kringelbach, & Deco, 2018): some models may support partial
synchronization, whereas others are able to support scale-free synchronization (Daffertshofer
et al., 2018). It is thus important to be aware of the question one aims to address (i.e., which as-
pects of the target system one is interested in) and to choose the model appropriately. Different
groups have, for instance, gained important insights from nonbiological or conceptual models
(Miši´c et al., 2014; Stam et al., 2016), with the argument that the finer details of a model are
irrelevant as long as the model is able to simulate realistic behavior on a systems level. For ex-
ample, dynamic models that belong to the same type or class of models behave similarly near a
Network Neuroscience
975
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
Multistability:
Coexistence of multiple stable
attractors in the state space of the
system or network.
Metastability:
Nonlinear dynamics definition:
The dwelling tendency of trajectories
in phase space without convergence
to an attractor (or remnants of
attractors). (This definition is used in
the context of this manuscript.)
Network science definition:
Quasi-stable state of the epidemics
in an SIS model beyond the epidemic
threshold. (This definition is
mentioned here only to avoid
confusion;it is not used in the
context of the current manuscript.)
Attractor:
Subspace in phase space to which a
trajectory converges for certain initial
conditions.
Limit cycle:
An isolated periodic oscillator, which
is a solution for a dynamical system
that repeats itself in time.
Chaos:
A periodic long-term behavior in a
deterministic system that exhibits
sensitive dependence on initial
conditions.
Fixed point:
A trajectory of a dynamical system
that does not change in time. It is
also often called an equilibrium or
steadystate.
phase transition (e.g., synchronization and epidemics act similarly around the phase transition).
However, these nonbiological models usually do not support multi- or metastability, which
is likely the dynamic regime in which the brain operates (Cabral et al., 2014; Kringelbach,
McIntosh, Ritter, Jirsa, & Deco, 2015). Temporal fluctuations in functional networks are charac-
terized by formation and dissolution of several subnetworks. Switching between subnetworks
can be achieved in systems that support multi- or metastability, and therefore these dynamic
regimes could be responsible for the emergence of time-varying fluctuations in empirical con-
nectivity (Cabral et al., 2014; Kringelbach et al., 2015; Tewarie et al., 2018). Simplistic models
such as epidemic spreading models/diffusion models usually do not support metastability or
multistability and are therefore limited in their ability to explain transitions between network
states observed in empirical time-varying data. Even when a computational model fits em-
pirical data, the results and potential insight gained from the model will largely depend on
the richness of its supporting dynamics (e.g., limit cycle, chaotic behavior, fixed points), and
thus conclusions can be highly biased by a limited repertoire of dynamic regimes. Common
behavior and generalizations across models have to some extent been studied in the context
of healthy brain networks (Messé, Rudrauf, Benali, & Marrelec, 2014), and may also guide
correct interpretation.
The second step, demonstration, is illustrated by Hughes through the following example:
“The wave theory of light represents light as a wave motion. It invites us to think about op-
tical phenomena in terms of the propagation of waves, and so to anticipate and explain the
behavior of light” (Hughes, 1997, p. S331). In this example, a mathematical model of light as
wave propagation could be physically demonstrated as wave motion in a water tank. A clear
feedback loop between modeled experiments and empirical data would arguably yield a lot of
improvement in the relevance of modeling studies for network neuroscience. Clinical network
science offers an opportunity, because the impact of brain disease can be studied over time
and compared with model-based prediction. Proven examples are models that predict altered
functional dynamics (e.g., Deco et al., 2011; Honey & Sporns, 2008) as a result of structural
brain damage, where the modeled dynamics resemble empirical findings (Butz & van Ooyen,
2013; de Haan et al., 2012; Rubinov, McIntosh, Valenzuela, & Breakspear, 2009; van Dellen
et al., 2013). The comparison to empirical data can then be used to further refine the under-
lying model. This is a slow iterative process, which will eventually lead to more insight into
mechanisms of neuronal oscillations and connectivity, and disease-related changes thereof.
The last step, interpretation, is to translate these inferences to the target system. In order to
be truly synergetic, the observations in our models or algorithms should lead to predictions on
the versatility, power, and energy consumption of the human brain, and on the fundamentals
that shape cognition and behavior.
What are the implications of directionality in the macroscopic brain network?
The study of structural and functional brain networks has mainly been based on link existence,
leading to undirected networks. Undirected networks can be represented by symmetric ma-
trices (such as the adjacency and Laplacian matrix; see Van Mieghem, 2011). In many cases,
the process on the network steers items (e.g., information, packets, traffic, flows) over links
in a certain direction and thus defines a direction of link (Van Mieghem, 2018). Often, di-
rected links are represented by an asymmetric adjacency matrix. Unidirectionality is a useful
simplification to allow progress in the field: analysis of directed graphs is considerably more
complicated than analysis of undirected graphs.
Network Neuroscience
976
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
The nodes or vertices of the structural brain network correspond to cortical and/or subcorti-
cal regions, and the links or edges between them represent bidirectional physical connections
(axonal projections) between regions. A functional connection exists if there is a statistical
dependency between regional activity patterns, which is thought to relate to the amount (or
probability) of communication taking place between brain areas. Although this bidirectionality
probably holds true for certain brain areas that are structurally connected, other functional con-
nections are likely unidirectional (Harriger, van den Heuvel, & Sporns, 2012; Honey, Kotter,
Breakspear, & Sporns, 2007; Kale et al., 2018; Scannell, Burns, Hilgetag, O’Neil, & Young,
1999; White, Southgate, Thomson, & Brenner, 1986). On a microscopic level, the structural
connections between neurons are considered largely unidirectional, as are functional con-
nections, since action potentials from sending neurons set up postsynaptic potentials in the
receiving neurons. Although this in itself does not mean that the interactions between neu-
ronal circuits, that is, macroscopic brain regions, are also necessarily unidirectional, there is
experimental evidence that such interactions exist at the macroscopic level as well. Indeed,
different brain regions may serve as preferred senders or receivers of information, giving rise
to (long-range) feed-forward and feed-back interactions, as well as global patterns of directed
functional connectivity (Bastos et al., 2015; Hillebrand et al., 2016). The directionality of func-
tional information transfer may depend on structural network characteristics such as node de-
gree (Moon, Lee, Blain-Moraes, & Mashour, 2015), suggesting that processes such as brain
development may dynamically alter directionality patterns.
However, it has proven difficult to develop methods to characterize directionality in the
human brain. Structural directionality at the axonal level cannot be accurately studied in hu-
mans in vivo. Functionally, different computational approaches estimate directionality, such
as dynamic causal modeling, phase transfer entropy, and Granger causality. Not only have the
assumptions of these methods themselves been challenged (Seth, Barrett, & Barnett, 2015),
it has also been questioned whether they are applicable in certain modalities. In particular,
fMRI may not be suitable to estimate directionality because of the low temporal resolution of
the signals measured (Smith, Pillai, Chen, & Horwitz, 2012; Wen, Rangarajan, & Ding, 2013).
Gilson and colleagues show theoretically and numerically that the use of covariances with
both zero and nonzero time shifts is the key to infer directed connectivity in fMRI data (Gilson,
Moreno-Bote, Ponce-Alvarez, Ritter, & Deco, 2016). Furthermore, accurate estimation of the
underlying directed functional network connectivity requires that the time shift for covariances
matches the time constant of the dynamical system, which currently is beyond what we can
measure in fMRI.
The nonproven assumption of these methods is that the timing of phases or extent of statisti-
cal prediction embedded in the measured functional signals reflects direction in the underlying
neuronal interactions. Moreover, the recorded time series are often noisy, the number of data
points is limited, and differences in, for example, the signal-to-noise ratios of two time series
may lead to erroneous conclusions about directional coupling (Bastos & Schoffelen, 2015). It is
not yet clear at what timescales directed interactions occur at the macroscopic level. Estimat-
ing brief directed interactions is beyond the capabilities of current macroscopic approaches
because of the limited number of data points, yet using longer time-windows might mask the
true underlying directionality. Similarly, averaging over trials to boost sensitivity might mask
intertrial variability in the preferred direction of interactions. Furthermore, while bivariate cou-
pling estimates may be affected by (hidden) common sources, multivariate measures are de-
pendent on the choice of model order, or require a large number of data points (Bastos &
Schoffelen, 2015). Testing against appropriate surrogate data may help to avoid false positives
Network Neuroscience
977
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
(Paluš & Vejmelka, 2007), but many measures may not be sensitive to true functional interac-
tions because the data cannot be sampled sufficiently within the required time frame.
We therefore need to develop more sensitive and reliable estimators of functional direction-
ality. Until such measures have been developed, should we refrain from including directional-
ity in our estimates of network topology? Directionality may be much more than an additional
source of insight into the brain network: viewing macroscopic networks as bidirectional may
even lead to erroneous reconstructions of their topology (Kale et al., 2018). The work by Kale
and colleagues also suggests that as long as directionality is strictly controlled in terms of false
positives, structural directionality should not be ignored, even if the directionality estimator is
not perfect: The specificity of directional connections has a larger effect on estimated network
topology than their sensitivity. However, it remains an open question whether these conclu-
sions also hold for functional brain networks.
Additionally, perturbed functional directionality patterns have been described in neuro-
logical and psychiatric disorders such as Alzheimer’s disease (AD), Parkinson’s disease (PD),
and delirium (Boon, Hillebrand, Olde Dubbelink, Stam, & Berendse, 2017; Engels et al., 2017;
Numan et al., 2017; van Dellen et al., 2014; van Wijk, Cagnan, Litvak, Kühn, & Friston, 2018),
and relate to seizure spread in patients with epilepsy (Wilke, van Drongelen, Kohrman, & He,
2010). The topology of the directed functional network may relate to nodal (i.e., brain regional)
vulnerability to pathology: Recent fundamental work suggests that topologically different parts
of directed networks have distinct sensitivities to the removal (and addition) of regions and con-
nections (Goltsev, Timár, & Mendes, 2017). Moreover, a stereotypical spreading of pathology
in progressive disorders has been observed (e.g., Braak et al., 2003). Misfolded disease-related
proteins may transfer transsynaptically from neuron to neuron (Brettschneider, Del Tredici,
Lee, & Trojanowski, 2015; Wang et al., 2017), suggesting that disease progression might be
predicted on the basis of the topology of the directed functional network. The implications of
directional brain networks will therefore likely factor into our understanding of neurological
and psychiatric disease.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
PART II: CLINICAL CHALLENGES
How can we increase our understanding of neurological and psychiatric disease through
network trajectories?
The application of network theory to neuroimaging and neurophysiology has uncovered asso-
ciations between cognitive (dys)functioning and brain network behavior. However, these are
mainly cross-sectional findings: Most network neuroscientific studies either demonstrate group
differences in the setting of neurological or psychiatric disease, or report on associations be-
tween network topology and cognitive functioning at the group level. Indeed, interindividual
differences in network organization are much larger than intraindividual variations (Gratton
et al., 2018). However, although functional brain network characteristics can be used for per-
sonal identification (Demuru et al., 2017; Finn et al., 2015) and are related to normal cogni-
tive functioning, these characteristics may be unrelated to the cognitive deterioration seen in
neurodegenerative disease. Cognition and both structural and functional brain network topol-
ogy may dynamically change concurrently or separately. The very essence of neurological and
psychiatric disease from a network neuroscience viewpoint, namely that they induce changing
network topology that is in itself dynamic and often progressive, might therefore be obscured
in strictly controlled cross-sectional case-control studies. Viewing brain network topology, be
it structural or functional, at a single time point as the sole signature of any specific disease
may then yield incorrect and ambiguous results (Tijms et al., 2013). Adaptive network models,
Network Neuroscience
978
The road ahead in clinical network neuroscience
described in Section I, may incorporate the time-varying interplay between structural and func-
tional connectivity and networks in brain disease. Experimentally, the collection of large co-
horts of cross-sectional data (e.g., the Human Connectome Project) should be complemented
by efforts and funding to collect high-quality longitudinal datasets of particular patient pop-
ulations. A longitudinal approach not only is crucial for insight into disease trajectories, but
also opens the gates towards network neuroscience–based prognostic biomarkers.
The essence here is that simply including more subjects is not always better than facilitat-
ing dense longitudinal measurements in fewer participants, particularly when studying elusive
symptoms like cognition and behavior. In healthy subjects, several (small and predominantly
functional) longitudinal datasets with high temporal sampling are available (e.g., the MyCon-
nectome project, Poldrack et al., 2015, and the Midnight Scan Club, Gordon et al., 2017)
and have yielded important insights into the individuality of brain network signatures (Braga
& Buckner, 2017). However, this type of focused longitudinal research has rarely been per-
formed in clinical populations, even though it has been recognized that declining cognition
and psychiatric disease may not be linear and may depend on multiple interacting factors,
including structural and functional connectivity (Jones et al., 2016; Schoonheim, Meijer, &
Geurts, 2015; Stam, 2014; van Os, Guloksuz, Vijn, Hafkenscheid, & Delespaul, 2019). Study
designs are needed that allow assessment of where any patient with a neurological or psychi-
atric disease is on a hypothesized group-level trajectory of brain network change.
As a feasible way forward, we propose to complement studies using single time points and
large samples in future case-control studies of network topology with longitudinal studies that
are tailored to the patient population at hand. Such studies should start off with a double base-
line at two consecutive moments, with the interval determined using information about the
disease at hand: Highly progressive diseases, such as high-grade glioma, should be sampled
at shorter intervals than more stable syndromes, such as mild cognitive impairment as a pro-
dromal phase of Alzheimer’s disease. In order to establish multimodal network trajectories, we
do not only consider network topology at the absolute first time point, but also assess whether
multimodal network topology is in the process of altering in a meaningful way between the
first and second baseline within each patient. Collecting data at the same interval in matched,
healthy controls should allow for adequate correction for variation between measurements
due to measurement error, artifacts, modality-specific noise, and so on. The remaining effects
may be interpreted as disease-specific dynamics, reflecting possible inflection points of the in-
dividual patient on the hypothesized group-level trajectory. This double baseline would then
ideally be followed by longitudinal sampling of the entire disease course, allowing for further
characterization of multimodal network changes, possibly going hand in hand with functional
decline. Ultimately, this type of “trajectory” would allow us to better understand (cognitive)
symptoms in disease, but more importantly may offer new avenues for prediction and treatment
of decline.
What is needed to use brain network characteristics as biomarkers?
Damage to the structural and functional connectome plays an important role in a variety of
brain disorders. A major goal of disease connectomics is to map commonalities and differences
across disorders, but also within patients, to facilitate diagnosis and track disease progression.
Expanding study designs from examining single disorders to sets of disorders may reveal general
principles of brain vulnerability, and specific and shared multimodal connectome pathology
across diseases (Cauda et al., 2018; Crossley et al., 2014). However, there are yet few signs
that structural and/or functional brain network analysis may in fact change clinical practice
Network Neuroscience
979
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
(Castellanos, Di Martino, Craddock, Mehta, & Milham, 2013; Fornito, Zalesky, & Breakspear,
2013). For example, whereas both visual and spectral analysis of EEG and MEG are informative
for the diagnosis and prognosis of certain dementia types (Engels et al., 2016; Gouw & Stam,
2016; Gouw et al., 2017; Olde Dubbelink et al., 2014), functional connectivity and network
characteristics seem to add relatively limited clinical value to standard clinical brain imaging
(Dauwan et al., 2016; Nissen et al., 2018).
The first clinical application of brain network analysis would be the development of a
biomarker: “A characteristic that is objectively measured and evaluated as an indicator of nor-
mal biological processes, pathogenic processes, or pharmacologic responses to a therapeutic
intervention” (Biomarkers Definitions Working Group, 2001, p. 91). Structural and functional
brain network characteristics need to be sufficiently reliable, reproducible, sensitive, and
specific to serve as biomarkers. An increasing number of studies report the sensitivity of
multimodal brain network characteristics to differentiate between patients and controls (Ding
et al., 2013; Vecchio et al., 2018). An important and easily implemented step towards ade-
quate investigation of brain network measures as biomarkers is to consistently report reliability,
reproducibility, sensitivity, and specificity of any measure in classifying patients and controls,
instead of only reporting statistically significant group differences. Somewhat more difficult,
but possibly influential in this context, is creating a large database with all (multimodal) data
from both healthy subjects and different patient populations in order to crosslink different
investigations.
Another direction is to use brain stimulation techniques as controlled network perturba-
tions, in order to understand the dynamic adaptation of brain networks in healthy and disease
states and thus contribute to clinical translation. For instance, TMS can be used to either infer
reversible “virtual lesions” in healthy controls (Klomjai, Katz, & Lackmy-Vallée, 2015) or ame-
liorate brain dysfunction, such as treatment of major depression disorder (Perera et al., 2016).
Recent work has determined clinically useful localized targets, based on predominantly func-
tional network topology, that elicit consistent, predictable behavioral responses and treatment
effects (Fox et al., 2014; Fox, Liu, & Pascual-Leone, 2013; Weigand et al., 2018). Moreover, it
is becoming clear that the functional network topological profile of the target region is predic-
tive of individual patients’ clinical outcome, for instance in patients with Parkinson’s disease
(Koirala et al., 2018) or depression (Downar et al., 2014). The success of translation of net-
work neuroscientific approaches to clinical practice may in this context be facilitated by the
controlled perturbation achieved by neurostimulation, which is more easily understood and
tested than for instance disease progression or systemic treatments.
Can we use network neuroscience for disease classification?
Network neuroscience may help to classify brain diseases. Current neurological and espe-
cially psychiatric disease classifications are often based on symptom co-occurrence, much
more than strict knowledge of pathophysiology. Previously separate disease entities may ac-
tually share important pathology, like alpha-synuclein deposition across neurodegenerative
diseases (Zhang, Nie, & Chen, 2018). Instead of investigating the structural and functional
network correlates of the current classification of disease, we argue that comparison of multi-
modal network patterns within and across classes may bring new insight into (shared) clinical
trajectories and potential intervention strategies.
Sets of structural and/or functional network characteristics may be combined to classify pa-
tient groups using machine-learning algorithms, such as random forest algorithms (van Diessen,
Network Neuroscience
980
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
Otte, Braun, Stam, & Jansen, 2013). These algorithms have the advantage that they provide in-
dications of how much each network feature contributes to the classification result, allowing
better insight into specific versus general brain disease mechanisms. For example, approx-
imately 50% of variance in different dimensions of psychopathological symptoms that are
notoriously difficult to quantify, such as psychosis, mood, and fear, may be explained by only
small sets of functional brain network characteristics (Xia et al., 2018).
Integration of network neuroscience in projects such as the Research Domain Criteria (RDoC)
may bring further insight into the specificity of structural and functional network alterations in
relation to symptom and behavioral domains, treatment response, and prognosis (Insel et al.,
2010). The lack of clinical yield when it comes to network neuroscience may lie in the mis-
match between rigid diagnostic constructs based on groups of signs and symptoms, and a de
facto overlap (or continuum) across diagnostic classes, which share etiological factors, poly-
genic risks, and comorbidities (Crocq, 2018). Moreover, diagnoses such as schizophrenia or
bipolar disorders group patients together that are highly heterogeneous in terms of clinical
manifestation, treatment response, and, possibly, pathophysiological mechanisms. The RDoC
represents a paradigm shift in this sense, proposing to map individuals in terms of functional do-
mains (e.g., cognitive systems) and related constructs (e.g., working memory) on a full range of
variations independent from classic diagnostic boundaries, and spanning the normal (healthy)
to abnormal (pathological) spectrum (Cuthbert & Insel, 2013; Insel et al., 2010). Network neu-
roscience can bring a fundamental contribution to the RDoC paradigm, by defining robust
measurements (such as connectivity and topological measures) across levels of analysis, and
by integrating those levels of analysis into a unified object of investigation, for example with
multilayer network models (Braun et al., 2018). On the one hand, this line of research can
identify dimensional and neurobiological constructs that cut across diagnostic classes. For ex-
ample, patterns of functional segregation in the brain network identify psychopathological di-
mensions across psychiatric groups (Xia et al., 2018). On the other hand, the synergy between
RDoC and network neuroscience may help in identifying valid phenotypes within diagnostic
classes. For example, measures of functional connectivity within brain subnetworks identify
subtypes of depression (Drysdale et al., 2017), which could define optimal targets for treat-
ment selection (Furman & Trivedi, 2019) and personalized intervention and prognosis (Cocchi
& Zalesky, 2018). Clustering algorithms for (multilayer) patient networks, where edges repre-
sent multidimensional similarity between individuals, could also deliver meaningful subtypes
within the RDoC framework (Pai & Bader, 2018; Stefanik et al., 2018).
Finally, psychiatric symptoms may be understood from multiple levels of explanation,
based on, for example, cultural, social, brain imaging, and molecular information (Hugdahl
& Sommer, 2018). Multilayer network approaches may help to come to a conceptual integra-
tion of these levels of information (Gosak et al., 2018). This framework may also include novel
approaches such as network analysis of symptoms as well as digital tracing of interaction
patterns, termed “digital phenotyping” (Lydon-Staley, Barnett, Satterthwaite, & Bassett, 2018),
which quantifies interactions with digital devices, providing novel signatures of psychopathology.
Can we systematically bridge the gap between brain network interventions in silico and in vivo?
Neurological and psychiatric diseases are dynamic in various dimensions, as their symptoma-
tology and multimodal brain network topology change over time. Any intervention in these
diseases is therefore aimed at somehow halting or reversing the pathological multimodal net-
work change that has occurred or is imminent. There are many ways to modify brain structure
and function (e.g., surgery, medication, [non]invasive stimulation, cognitive therapy), but at
Network Neuroscience
981
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
present, interventions are usually performed without considering brain connectivity, network
embedding, or plasticity (see Section I). This has various potential disadvantages: (a) Interven-
tions may not be as efficient as they could be (Horn et al., 2017; Sale, Mattingley, Zalesky,
& Cocchi, 2015); (b) Unexpected adverse effects may occur because of the complex network
(re)organization of the human brain, for instance because of time-dependent effects (Cocchi
et al., 2016); (c) Clinical trial and error to find optimal treatment is slow, burdensome, and ex-
pensive; and (d) The reasons why a certain intervention does or does not work remain unclear
(Sale et al., 2015).
Computational network neuroscience has been used to model the impact of different
types of pathology and disease stages on the structure and functioning of brain networks (Proix,
Bartolomei, Guye, & Jirsa, 2017). An appealing next step is simulation of treatment effects
before putting them in practice (Hughes’ “demonstration”): a kind of “virtual trial” (de Haan
et al., 2017). An example of this approach has been reported for Alzheimer’s disease: In a
computational model that coupled 78 neural mass models according to human, DTI-based
topology, damage was applied to the network based on the local levels of neuronal activity
(“activity-dependent degeneration”). Simultaneously, virtual interventions that applied differ-
ent levels of enhancement or suppression of neuronal excitability were compared for their
ability to maintain normal network topology over time (de Haan et al., 2017). This led to spe-
cific predictions for the optimal stimulation protocol, which can be verified in vivo.
In order to cross the barrier towards use in clinical practice, we need to find a method
to match theoretical and practical treatment strategies. In nonhuman experiments, combina-
tions of neuronal stimulation and accurate network consequence acquisition and analysis pro-
duce fascinating new insights into the organizational principles of the brain (Gollo, Roberts,
& Cocchi, 2017; Wang, Hutchings, & Kaiser, 2015). Feasible analogues in humans could
be transcranial direct current stimulation (tDCS), transcranial alternating current stimulation
(tACS), transcranial focused ultrasound (tFUS), or the previously described TMS: stimulation
techniques that alter neuronal excitability, and thereby influence functional network proper-
ties (Sale et al., 2015). Since tDCS can be performed during MEG recording, we have a way of
simultaneously altering and recording large-scale functional networks. The challenge is then
to find reliable ways to produce (and reproduce) desired network manipulations, and turn
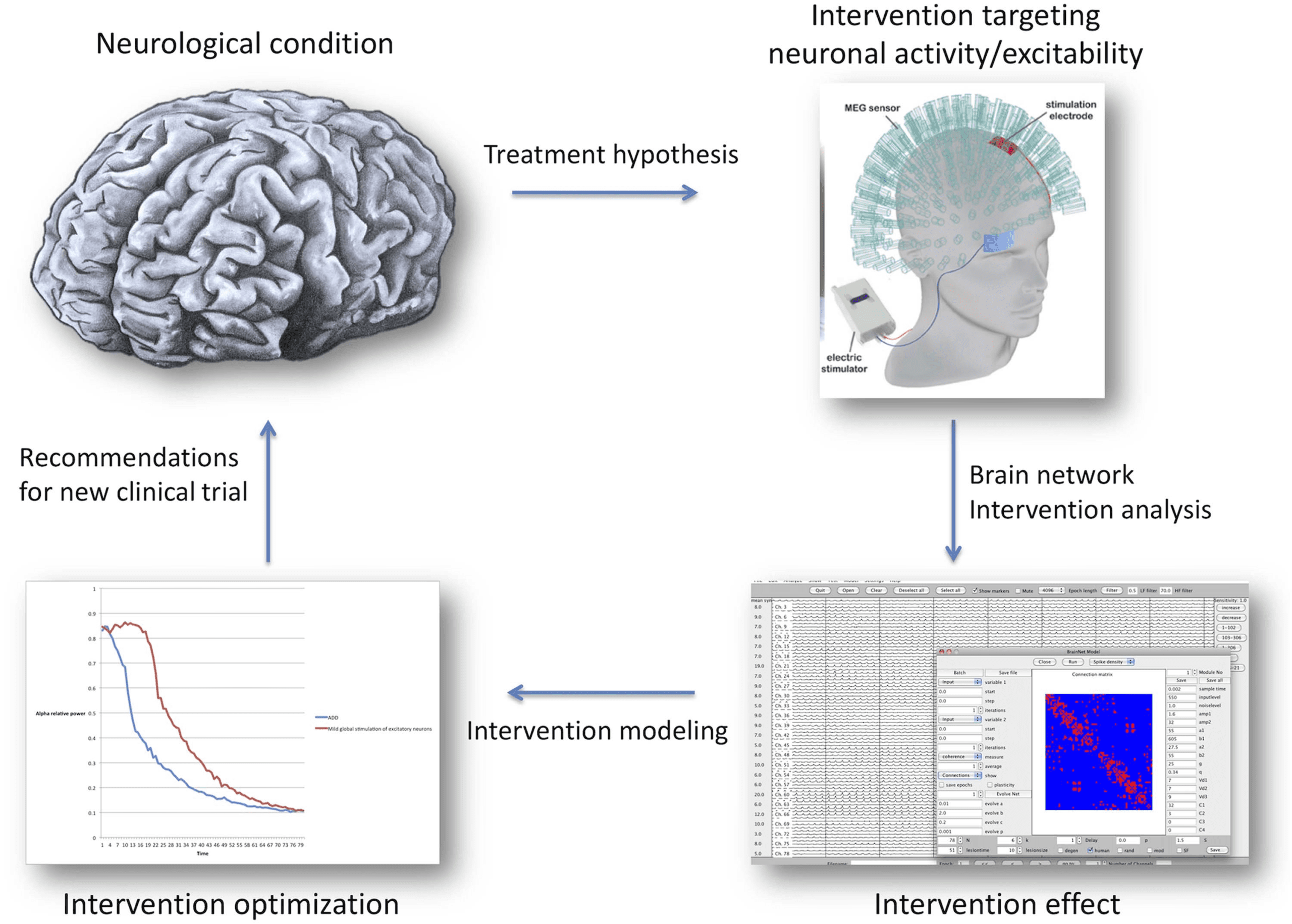
successful virtual interventions into real ones (see Figure 2).
Are evolution and dissolution driving factors in disease connectomics?
An evolutionary approach with network neuroscience may aid in understanding the biology of
disease: Comparing the topological organization of the human structural and functional brain
network to that of other species may enhance our understanding of evolutionary processes that
have shaped human brain wiring (Buckner & Krienen, 2013). Comparative connectomics may
further our understanding of human brain network topology and its implications for function-
ing in both health and disease (Rilling & van den Heuvel, 2018; van den Heuvel, Bullmore, &
Sporns, 2016). A hypothesis that has been posited for human-specific diseases is that “the evo-
lution of brain wiring in support of complex brain function in humans may have increased the
vulnerability to brain dysfunction in disease” (van den Heuvel et al., 2018, p. 2). Schizophre-
nia for example has been suggested to relate to increased evolutionary pressure on structural
brain connectivity in the human brain (Burns, 2004; van den Heuvel et al., 2018), arguably
increasing the vulnerability of the human brain to various brain disorders (Gollo et al., 2018).
Network Neuroscience
982
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
t
.
Figure 2. Using in silico and in vivo experiments to advance interventions. A tentative flowchart for the integration of network interven-
tion modeling and clinical studies. Systematic intervention modeling produces predictions for clinical experiments, for example, by altering
neuronal excitability and testing large-scale network consequences. Vice versa, observed treatment effects can be used to validate and im-
prove model predictions. This mutually reinforcing approach can improve and speed up treatment development, keeping patient burden at a
minimum while providing more insight into treatment success or failure.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
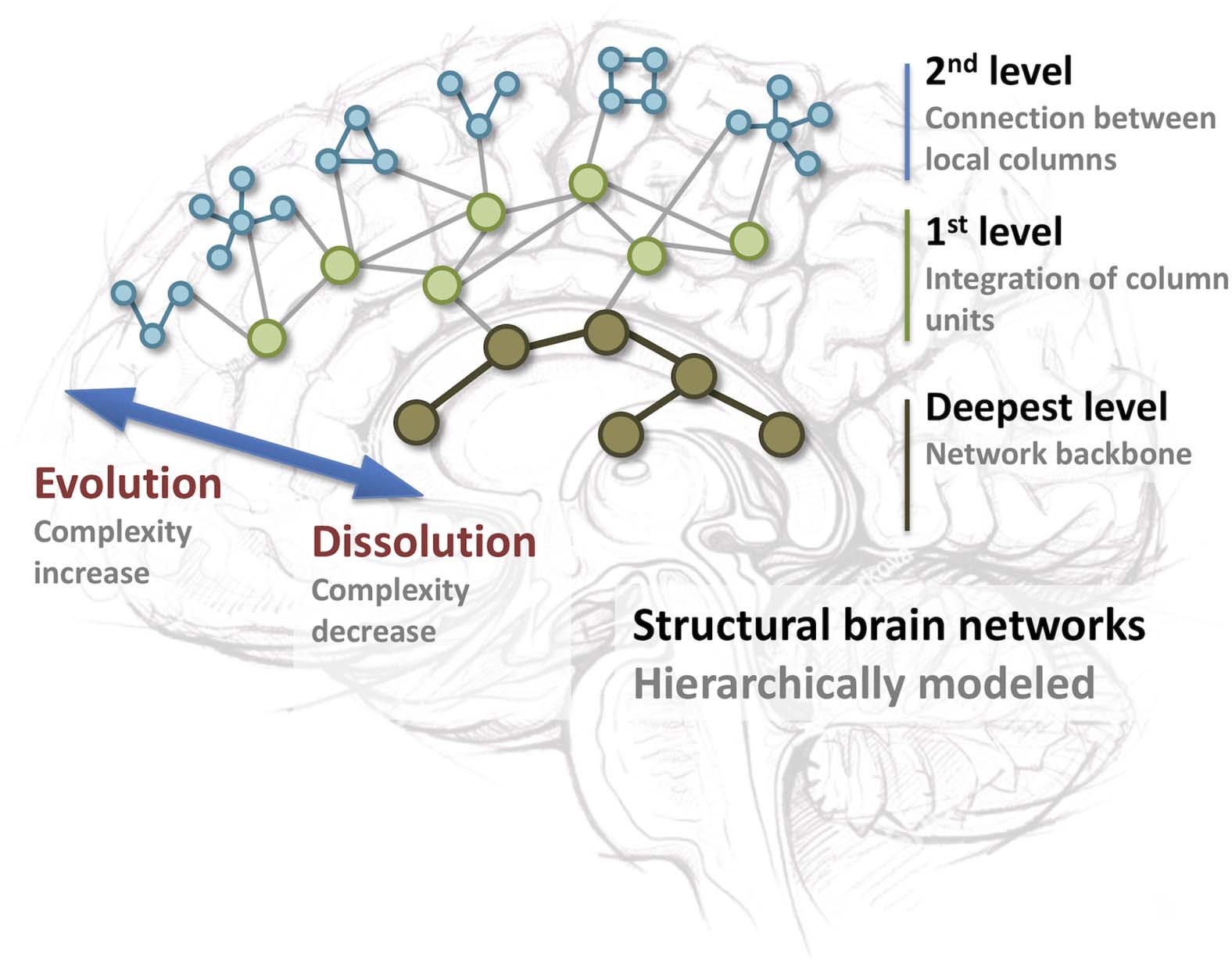
Using a pathological developmental view, loss of organization of the brain network may
be investigated using mapping of dissolution as opposed to evolution. This framework maps
end-stage network configurations, irrespective of timescales, in terms of evolution versus dis-
solution. An obvious example of a relevant disease population to do so is dementia, in which
different pathological entities lead to a final common pathway of network dissolution. Another
example is epilepsy, wherein the concept of a singular focus has shifted to a more widespread
interconnected epileptogenic network (Englot, Konrad, & Morgan, 2016; Kramer & Cash, 2012;
van Diessen, Diederen, Braun, Jansen & Stam, 2013). Seizures may recur several years after
initially successful epilepsy surgery, possibly because of aberrant plasticity and newly grown
connections due to ongoing epileptic activity from (microscopic) parts of the network. John
Hughlings Jackson (1835–1911), the founder of contemporary epileptology, already viewed
epilepsy as a network disease (Jackson, 1884). He saw the brain as a hierarchically ordered
construction of connected brain centers, in which the complexity of connections, due to an
Network Neuroscience
983
The road ahead in clinical network neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3.
Schematic representation of evolution versus dissolution as a theoretical framework to-
wards understanding epilepsy. Structural brain networks are hierarchically ordered from 100 million
cortical minicolumns and 1–2 million cortical columns to distributed column groups and deeper
underlying subcortical network structures. Because of the number and interconnectivity of columns,
complexity is highest at the cortical column connectivity level and decreases at lower topological
levels, namely at the levels of integration of columns and the deeper network backbone. Com-
plexity is defined here as the relative independence of subnetworks and their properties from the
integrative network properties at large. An important hallmark of many network disorders is a shift in
the network complexity, which is characterized by evolutionary processes of rewiring that increase
complexity and network dissolution that decreases complexity.
exponential growth in network nodes and additional layering, increases when we move from
deeper regions toward the neocortex. Hughlings Jackson then labeled this increase in com-
plexity as evolution and the decrease as dissolution (see Figure 3). According to his theory,
brain disorders originated not from malfunctioning of one or two specific brain centers, but
mainly from deterioration of connections between multiple centers. Epilepsy was then charac-
terized by dissolution of brain networks, so a reversed evolution with a reduction in connection
complexity. With this theory he introduced an alternative framework that differed from the, at
that time, dominant, binary all-or-nothing concept and preoccupation with regional pathology.
Although it has been abandoned for decades, network analysis of longitudinal neuroimaging
and physiological datasets may be able to assess the value of understanding evolution and
dissolution of networks in the clinical setting of epilepsy.
Statistical models that allow the mapping of evolution-dissolution in (disease) networks are
the exponential random graph model (Sinke, Dijkhuizen, Caimo, Stam, & Otte, 2016) or topo-
logical motifs morphospace mapping (Morgan, Achard, Termenon, Bullmore, & Vértes, 2018).
Both techniques decompose the network in easy to understand building blocks. In addition,
Network Neuroscience
984
The road ahead in clinical network neuroscience
the morphospace maps the networks into a common space, allowing the quantification of net-
work relationships over time and the driving forces behind network evolution or dissolution.
This approach may also help to assess whether already changed network topologies, for ex-
ample because of years of suffering from medication-refractory epilepsy, remain vulnerable to
disease despite initial treatment.
CONCLUSION
We have identified a set of questions that may guide future research. Two main themes deserve further
attention: (a) the methodological challenges of brain network analysis and (b) the translation of
findings to gain understanding of neurological and psychiatric diseases and ultimately improve
clinical practice. We have delineated a number of routes forward with respect to these themes,
which are without doubt also being explored concurrent to the writing of this review piece.
It is worth mentioning in this context that progress might not occur through experimentation
within a settled paradigm or explanatory framework, or according to a tradition of investiga-
tion, which Thomas Kuhn termed “normal science” in his seminal work on scientific revolu-
tions (Kuhn, 1962). In (clinical) network neuroscience, such tradition may relate to step-by-step
exclusion of hurdles standing in the way of progress, instead of fundamentally challenging the
way we investigate brain networks. We therefore invite the field of network neuroscience to not
only address obvious hurdles towards full fruition of clinical network neuroscience, but also remain
open to unexpected thoughts and ideas. Or, to speak with Paul Feyerabend: Anything goes.
AUTHOR CONTRIBUTIONS
All authors contributed in the conceptualization and writing of the manuscript. Linda Douw
and Edwin van Dellen reviewed and edited contributions of the other authors, merged these
contributions into an original draft, and revised the manuscript based on peer-review. Linda
Douw: Conceptualization; Writing – original draft; Writing – review & editing. Edwin van
Dellen: Conceptualization; Writing – original draft; Writing – review & editing. Alida A. Gouw:
Conceptualization; Writing – original draft. Alessandra Griffa: Conceptualization; Writing –
original draft. Willem de Haan: Conceptualization; Writing – original draft. Martijn van den
Heuvel: Conceptualization; Writing – original draft. Arjan Hillebrand: Conceptualization;
Writing – original draft. Piet Van Mieghem: Conceptualization; Writing – original draft. Ida
A. Nissen: Conceptualization; Writing – original draft. Willem M. Otte: Conceptualization;
Writing – original draft. Yael D. Reijmer: Conceptualization; Writing – original draft. Menno M.
Schoonheim: Conceptualization; Writing – original draft. Mario Senden: Conceptualization;
Writing – original draft. Elisabeth C.W. van Straaten: Conceptualization; Writing – original
draft. Betty M. Tijms: Conceptualization; Writing – original draft. Prejaas Tewarie: Concep-
tualization; Writing – original draft. Cornelis J. Stam: Conceptualization; Writing – original
draft.
FUNDING INFORMATION
Linda Douw, Netherlands Organization for Scientific Research (NWO) Vidi grant, Award ID:
198.015. Edwin van Dellen, The Netherlands Organization for Health Research and Devel-
opment (ZonMW) GGZ fellowship, Award ID: 60-63600-98-711. Edwin van Dellen, UMC
Utrecht Clinical Research Talent Fellowship. Award ID: NA Alessandra Griffa, Swiss National
Science Foundation, Award ID: SNSF #P2ELP3_172087. Martijn van den Heuvel, ALW open,
Award ID: ALWOP.179; Martijn van den Heuvel, NWO Vidi grant, Award ID: 452-16-015.
Martijn van den Heuvel, MQ Fellowship. Award ID: NA Ida A. Nissen, Netherlands Epilepsy
Network Neuroscience
985
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
Fund, Award ID: 95105006. Willem M. Otte, NWO VENI grant, Award ID: 016.168.038.
Menno M. Schoonheim, Dutch MS Research Foundation, Award ID: 13-820; Mario Senden,
EU Horizon 2020 program, Q2 Award ID: 737691-HBP SGA2. Betty M. Tijms, ZonMW, Award
ID: 73305056.
REFERENCES
Abeysuriya, R. G., Hadida, J., Sotiropoulos, S. N., Jbabdi, S., Becker,
R., Hunt, B. A. E., . . . Woolrich, M. W.
(2018). A biophysical
model of dynamic balancing of excitation and inhibition in fast
oscillatory large-scale networks. PLoS Computational Biology,
14(2), e1006007. https://doi.org/10.1371/journal.pcbi.1006007
Alexander-Bloch, A. F., Vértes, P. E., Stidd, R., Lalonde, F., Clasen,
L., Rapoport, J., . . . Gogtay, N. (2013). The anatomical distance of
functional connections predicts brain network topology in health
and schizophrenia. Cerebral Cortex, 23(1), 127–138. https://doi.
org/10.1093/cercor/bhr388
Amico, E., Bodart, O., Rosanova, M., Gosseries, O., Heine, L.,
Van Mierlo, P., . . . Laureys, S. (2017). Tracking dynamic in-
teractions between structural and functional connectivity: A
TMS/EEG-dMRI study. Brain Connectivity, 7(2), 84–97. https://
doi.org/10.1089/brain.2016.0462
Ashwin, P., Coombes, S., & Nicks, R. (2016). Mathematical frame-
works for oscillatory network dynamics in neuroscience. Journal
of Mathematical Neuroscience, 6(1), 2. https://doi.org/10.1186/
s13408-015-0033-6
Avena-Koenigsberger, A., Misic, B., & Sporns, O. (2017). Commu-
nication dynamics in complex brain networks. Nature Reviews
Neuroscience, 19(1), 17–33. https://doi.org/10.1038/nrn.2017.
149
Bansal, K., Nakuci, J., & Muldoon, S. F. (2018). Personalized brain
network models for assessing structure-function relationships.
Current Opinion in Neurobiology, 52, 42–47. https://doi.org/10.
1016/j.conb.2018.04.014
Barabasi, A. L., & Albert, R. (1999). Emergence of scaling in random
networks. Science, 286(5439), 509–512.
Bassett, D. S., Zurn, P., & Gold, J. I. (2018). On the nature and use of
models in network neuroscience. Nature Reviews Neuroscience,
1. https://doi.org/10.1038/s41583-018-0038-8
Bastos, A. M., & Schoffelen, J.-M. (2015). A tutorial review of func-
tional connectivity analysis methods and their interpretational
pitfalls. Frontiers in Systems Neuroscience, 9, 175. https://doi.
org/10.3389/fnsys.2015.00175
Bastos, André Moraes, Vezoli, J., Bosman, C. A., Schoffelen, J.-M.,
Oostenveld, R., Dowdall, J. R., . . . Fries, P. (2015). Visual areas
exert feedforward and feedback influences through distinct fre-
quency channels. Neuron, 85(2), 390—401. https://doi.org/10.
1016/j.neuron.2014.12.018
Bettinardi, R. G., Deco, G., Karlaftis, V. M., Van Hartevelt, T. J.,
Fernandes, H. M., Kourtzi, Z., . . . Zamora-López, G. (2017). How
structure sculpts function: Unveiling the contribution of anatom-
ical connectivity to the brain’s spontaneous correlation structure.
Chaos: An Interdisciplinary Journal of Nonlinear Science, 27(4),
047409. https://doi.org/10.1063/1.4980099
Biomarkers Definitions Working Group. (2001). Biomarkers and
surrogate endpoints: Preferred definitions and conceptual frame-
work. Clinical Pharmacology and Therapeutics, 69(3), 89–95.
https://doi.org/10.1067/mcp.2001.113989
Birn, R. M., Molloy, E. K., Patriat, R., Parker, T., Meier, T. B., Kirk,
G. R., . . . Prabhakaran, V. (2013). The effect of scan length on
the reliability of resting-state fMRI connectivity estimates. Neuro-
Image, 83, 550–558. https://doi.org/10.1016/j.neuroimage.2013.
05.099
Blautzik, J., Vetter, C., Peres, I., Gutyrchik, E., Keeser, D., Berman,
A., . . . Meindl, T. (2013). Classifying fMRI-derived resting-state
connectivity patterns according to their daily rhythmicity. Neuro-
Image, 71, 298–306. https://doi.org/10.1016/j.neuroimage.2012.
08.010
Boccaletti, S., Latora, V., Moreno, Y., Chavez, M., & Hwang, D. U.
(2006). Complex networks: Structure and dynamics. Physics Reports,
424, 175–308.
Bonilha, L., Gleichgerrcht, E., Fridriksson, J., Rorden, C., Breedlove,
J. L., Nesland, T., . . . Focke, N. K.
(2015). Reproducibility
of the structural brain connectome derived from diffusion tensor
imaging. PLoS ONE, 10(8), e0135247. https://doi.org/10.1371/
journal.pone.0135247
Boon, L. I., Hillebrand, A., Olde Dubbelink, K. T. E., Stam, C. J.,
& Berendse, H. W. (2017). Changes in resting-state directed
connectivity in cortico-subcortical networks correlate with cog-
nitive function in Parkinson’s disease. Clinical Neurophysiol-
ogy, 128(7), 1319–1326. https://doi.org/10.1016/j.clinph.2017.
04.024
Braak, H., Del Tredici, K., Rüb, U., de Vos, R. A. I., Jansen Steur,
E. N. H., & Braak, E. (2003). Staging of brain pathology related
to sporadic Parkinson’s disease. Neurobiology of Aging, 24(2),
197–211.
Braga, R. M., & Buckner, R. L. (2017). Parallel interdigitated dis-
tributed networks within the individual estimated by intrinsic
functional connectivity. Neuron, 95(2), 457–471.e5. https://doi.
org/10.1016/j.neuron.2017.06.038
Braun, U., Schaefer, A., Betzel, R. F., Tost, H., Meyer-Lindenberg, A.,
& Bassett, D. S. (2018). From maps to multi-dimensional network
mechanisms of mental disorders. Neuron, 97(1), 14–31. https://
doi.org/10.1016/j.neuron.2017.11.007
Brettschneider, J., Del Tredici, K., Lee, V. M.-Y., & Trojanowski, J. Q.
(2015). Spreading of pathology in neurodegenerative diseases: A
focus on human studies. Nature Reviews Neuroscience, 16(2),
109–120. https://doi.org/10.1038/nrn3887
Buckner, R. L., & Krienen, F. M. (2013). The evolution of distributed
association networks in the human brain. Trends in Cognitive
Sciences, 17(12), 648–665. https://doi.org/10.1016/j.tics.2013.
09.017
Burns, J. K. (2004). An evolutionary theory of schizophrenia: Corti-
cal connectivity, metarepresentation, and the social brain. Behav-
ioral and Brain Sciences, 27(6), 855–885; discussion 831–855.
Network Neuroscience
986
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
Butz, M., & van Ooyen, A. (2013). A simple rule for dendritic spine
and axonal bouton formation can account for cortical reorgani-
zation after focal retinal lesions. PLoS Computational Biology,
9(10). https://doi.org/10.1371/journal.pcbi.1003259
Butz, M., van Ooyen, A., & Worgotter, F. (2009). A model for corti-
cal rewiring following deafferentation and focal stroke. Frontiers
in Computational Neuroscience, 3, 10.
Cabral, J., Kringelbach, M. L., & Deco, G. (2017). Functional con-
nectivity dynamically evolves on multiple time-scales over a static
structural connectome: Models and mechanisms. NeuroImage,
160, 84–96. https://doi.org/10.1016/J.NEUROIMAGE.2017.03.
045
Cabral, J., Luckhoo, H., Woolrich, M., Joensson, M., Mohseni, H.,
Baker, A., . . . Deco, G. (2014). Exploring mechanisms of spon-
taneous functional connectivity in MEG: How delayed network
interactions lead to structured amplitude envelopes of band-pass
filtered oscillations. NeuroImage, 90, 423–435. https://doi.org/
10.1016/j.neuroimage.2013.11.047
Castellanos, F. X., Di Martino, A., Craddock, R. C., Mehta, A. D., &
Milham, M. P. (2013). Clinical applications of the functional con-
nectome. NeuroImage, 80, 527–540. https://doi.org/10.1016/j.
neuroimage.2013.04.083
Cauda, F., Nani, A., Manuello, J., Premi, E., Palermo, S., Tatu, K.,
. . . Costa, T. (2018). Brain structural alterations are distributed
following functional, anatomic and genetic connectivity. Brain.
https://doi.org/10.1093/brain/awy252
Chang, C., & Glover, G. H.
(2010). Time-frequency dynamics
of resting-state brain connectivity measured with fMRI. Neuro-
Image, 50(1), 81–98. https://doi.org/10.1016/j.neuroimage.2009.
12.011
Cocchi, L., Sale, M. V., Gollo, L. L., Bell, P. T., Nguyen, V. T.,
Zalesky, A., . . . Mattingley, J. B. (2016). A hierarchy of timescales
explains distinct effects of local inhibition of primary visual cor-
tex and frontal eye fields. eLife, 5. https://doi.org/10.7554/eLife.
15252
Cocchi, L., & Zalesky, A. (2018). Personalized transcranial magnetic
stimulation in psychiatry. Biological Psychiatry: Cognitive Neu-
roscience and Neuroimaging, 3(9), 731–741. https://doi.org/10.
1016/j.bpsc.2018.01.008
Colclough, G. L., Woolrich, M. W., Tewarie, P. K., Brookes, M. J.,
Quinn, A. J., & Smith, S. M. (2016). How reliable are MEG
resting-state connectivity metrics? NeuroImage, 138, 284–293.
https://doi.org/10.1016/j.neuroimage.2016.05.070
Crocq, M. A. (2018). Can psychopathology and neuroscience co-
exist in psychiatric classifications? Dialogues in Clinical Neuro-
science, 20(3), 155–160.
Crossley, N. A., Mechelli, A., Scott, J., Carletti, F., Fox, P. T.,
McGuire, P., & Bullmore, E. T. (2014). The hubs of the human
connectome are generally implicated in the anatomy of brain dis-
orders. Brain, 137(Pt. 8), 2382–2395. https://doi.org/10.1093/
brain/awu132
Cuthbert, B. N., & Insel, T. R. (2013). Toward the future of psychi-
atric diagnosis: The seven pillars of RDoc. BMC Medicine, 11(1),
126. https://doi.org/10.1186/1741-7015-11-126
Daffertshofer, A., Ton, R., Pietras, B., Kringelbach, M. L., & Deco, G.
(2018). Scale-freeness or partial synchronization in neural mass
phase oscillator networks: Pick one of two? NeuroImage. https://
doi.org/10.1016/J.NEUROIMAGE.2018.03.070
Dauwan, M., van der Zande, J., van Dellen, E., Sommer, I., Schel-
tens, P., Lemstra, A., & Stam, C.
(2016). Random forest to dif-
ferentiate dementia with Lewy bodies from Alzheimer’s disease.
Alzheimer’s and Dementia: Diagnosis. Assessment and Disease
Monitoring, 4, 99–106.
de Haan, W., Mott, K., van Straaten, E. C. W., Scheltens, P., & Stam,
C. J. (2012). Activity dependent degeneration explains hub vul-
nerability in Alzheimer’s disease. PLoS Computational Biology,
8(8), e1002582. https://doi.org/10.1371/journal.pcbi.1002582
de Haan, W., van der Flier, W. M., Koene, T., Smits, L. L., Scheltens,
P., & Stam, C. J. (2012). Disrupted modular brain dynamics re-
flect cognitive dysfunction in Alzheimers disease. NeuroImage,
59(4), 3085–3093. https://doi.org/10.1016/j.neuroimage.2011.
11.055
de Haan, W., van Straaten, E. C. W., Gouw, A. A., & Stam,
C. J. (2017). Altering neuronal excitability to preserve network
connectivity in a computational model of Alzheimer’s disease.
PLoS Computational Biology, 13(9), e1005707. https://doi.org/
10.1371/journal.pcbi.1005707
De Vico Fallani, F., Latora, V., & Chaves, M.
(2017). A topologi-
cal criterion for filtering information in complex brain networks.
PLoS Computational Biology, 13(1), e1005305.
Deco, G., Jirsa, V. K., & McIntosh, A. R.
(2011). Emerging con-
cepts for the dynamical organization of resting-state activity in
the brain. Nature Reviews Neuroscience, 12(1), 43–56.
Demuru, M., Gouw, A. A., Hillebrand, A., Stam, C. J., van Dijk,
B. W., Scheltens, P., . . . Visser, P. J. (2017). Functional and effec-
tive whole brain connectivity using magnetoencephalography to
identify monozygotic twin pairs. Scientific Reports, 7(1), 9685.
Deuker, L., Bullmore, E. T., Smith, M., Christensen, S., Nathan, P. J.,
Rockstroh, B., & Bassett, D. S. (2009). Reproducibility of graph
metrics of human brain functional networks. NeuroImage, 47(4),
1460–1468. https://doi.org/10.1016/j.neuroimage.2009.05.035
Ding, J. R., An, D., Liao, W., Li, J., Wu, G.-R., Xu, Q., . . . Chen, H.
(2013). Altered functional and structural connectivity networks
in psychogenic non-epileptic seizures. PLoS ONE, 8(5), e63850.
https://doi.org/10.1371/journal.pone.0063850
Downar, J., Geraci, J., Salomons, T. V., Dunlop, K., Wheeler, S.,
McAndrews, M. P., . . . Giacobbe, P. (2014). Anhedonia and
reward-circuit connectivity distinguish nonresponders from respond-
ers to dorsomedial prefrontal repetitive transcranial magnetic
stimulation in major depression. Biological Psychiatry, 76(3),
176–185. https://doi.org/10.1016/j.biopsych.2013.10.026
Drysdale, A. T., Grosenick, L., Downar, J., Dunlop, K., Mansouri,
F., Meng, Y., . . . Liston, C. (2017). Resting-state connectiv-
ity biomarkers define neurophysiological subtypes of depres-
sion. Nature Medicine, 23(1), 28–38. https://doi.org/10.1038/
nm.4246
Eklund, A., Nichols, T. E., & Knutsson, H. (2016). Cluster failure:
Why fMRI inferences for spatial extent have inflated false-positive
rates. Proceedings of the National Academy of Sciences, 113(28),
7900–7905. https://doi.org/10.1073/pnas.1602413113
Engels, M. M. A., Hillebrand, A., van der Flier, W. M., Stam, C. J.,
Scheltens, P., & van Straaten, E. C. W.
(2016). Slowing of hip-
pocampal activity correlates with cognitive decline in early onset
Alzheimer’s disease. An MEG study with virtual electrodes.
Network Neuroscience
987
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
Frontiers in Human Neuroscience, 10, 238. https://doi.org/10.3389/
fnhum.2016.00238
Engels, M. M. A., Yu, M., Stam, C. J., Gouw, A. A., van der Flier,
(2017). Directional
W. M., Scheltens, P., . . . Hillebrand, A.
information flow in patients with Alzheimer’s disease. A source-
space resting-state MEG study. NeuroImage: Clinical, 15, 673–
681. https://doi.org/10.1016/j.nicl.2017.06.025
Englot, D. J., Konrad, P. E., & Morgan, V. L. (2016). Regional
and global connectivity disturbances in focal epilepsy, re-
lated neurocognitive sequelae, and potential mechanistic under-
pinnings. Epilepsia, 57(10), 1546–1557. https://doi.org/10.1111/
epi.13510
Esteban, O., Markiewicz, C. J., Blair, R. W., Moodie, C. A., Isik, A. I.,
Erramuzpe, A., . . . Gorgolewski, K. J.
fMRIPrep: A ro-
bust preprocessing pipeline for functional MRI. Nature Methods,
16(1), 111–116. https://doi.org/10.1038/s41592-018-0235-4
Fauth, M., & Tetzlaff, C. (2016). Opposing effects of neuronal ac-
tivity on structural plasticity. Frontiers in Neuroanatomy, 10, 75.
https://doi.org/10.3389/fnana.2016.00075
(2019).
Finn, E. S., Shen, X., Scheinost, D., Rosenberg, M. D., Huang, J.,
Chun, M. M., . . . Constable, R. T. (2015). Functional connectome
Identifying individuals using patterns of brain
fingerprinting:
connectivity. Nature Neuroscience, 18(11), 1664–1671. https://
doi.org/10.1038/nn.4135
Fornito, A., Bullmore, E. T., & Zalesky, A. (2017). Opportunities
and challenges for psychiatry in the connectomic era. Biological
Psychiatry: Cognitive Neuroscience and Neuroimaging, 2(1),
9–19. https://doi.org/10.1016/j.bpsc.2016.08.003
Fornito, A., Zalesky, A., & Breakspear, M. (2013). Graph analysis of
the human connectome: Promise, progress, and pitfalls. NeuroI-
mage, 80, 426–444. https://doi.org/10.1016/j.neuroimage.2013.
04.087
Fornito, A., Zalesky, A., & Bullmore, E. T. (2016). Fundamentals of
brain network analysis Amsterdam. Netherlands: Academic Press.
Fox, M. D., Buckner, R. L., Liu, H., Chakravarty, M. M., Lozano,
A. M., & Pascual-Leone, A. (2014). Resting-state networks link
invasive and noninvasive brain stimulation across diverse psy-
chiatric and neurological diseases. Proceedings of the National
Academy of Sciences, 111(41), E4367–E4375. https://doi.org/10.
1073/pnas.1405003111
Fox, M. D., Liu, H., & Pascual-Leone, A. (2013). Identification
of reproducible individualized targets for treatment of depres-
sion with TMS based on intrinsic connectivity. NeuroImage,
66, 151–160. https://doi.org/10.1016/J.NEUROIMAGE.2012.10.
082
Fraschini, M., Demuru, M., Crobe, A., Marrosu, F., Stam, C. J.,
& Hillebrand, A. (2016). The effect of epoch length on esti-
mated EEG functional connectivity and brain network organisa-
tion. Journal of Neural Engineering, 13(3), 036015. https://doi.
org/10.1088/1741-2560/13/3/036015
Furman, J. L., & Trivedi, M. H. (2019). Biomarker-based treatment
selection: A precision medicine approach for depression. Neuro-
biology of Depression, 331–340. https://doi.org/10.1016/B978-
0-12-813333-0.00029-9
Garcés, P., Martín-Buro, M. C., & Maestú, F. (2016). Quantifying
the test-retest reliability of magnetoencephalography resting-
state functional connectivity. Brain Connectivity, 6(6), 448–460.
https://doi.org/10.1089/brain.2015.0416
Gilden, D. L., Thornton, T., & Mallon, M. W. (1995). 1/f noise in
human cognition. Science, 267(5205), 1837–1839.
Gilson, M., Moreno-Bote, R., Ponce-Alvarez, A., Ritter, P., & Deco,
G. (2016). Estimation of directed effective connectivity from fMRI
functional connectivity hints at asymmetries of cortical connec-
tome. PLoS Computational Biology, 12(3), e1004762. https://doi.
org/10.1371/journal.pcbi.1004762
Glomb, K., Ponce-Alvarez, A., Gilson, M., Ritter, P., & Deco, G. (2018).
Stereotypical modulations in dynamic functional connectivity ex-
plained by changes in BOLD variance. NeuroImage, 171, 40–54.
https://doi.org/10.1016/J.NEUROIMAGE.2017.12.074
Gollo, L. L., Roberts, J. A., & Cocchi, L. (2017). Mapping how lo-
cal perturbations influence systems-level brain dynamics. Neuro-
Image, 160, 97–112. https://doi.org/10.1016/j.neuroimage.2017.
01.057
Gollo, L. L., Roberts, J. A., Cropley, V. L., Di Biase, M. A., Pantelis,
C., Zalesky, A., & Breakspear, M. (2018). Fragility and volatility of
structural hubs in the human connectome. Nature Neuroscience,
21(8), 1107–1116. https://doi.org/10.1038/s41593-018-0188-z
Goltsev, A. V., Timár, G., & Mendes, J. F. F. (2017). Sensitivity of
directed networks to the addition and pruning of edges and
vertices. Physical Review E, 96(2–1), 022317. https://doi.org/10.
1103/PhysRevE.96.022317
Goñi, J., van den Heuvel, M. P., Avena-Koenigsberger, A., Velez de
Mendizabal, N., Betzel, R. F., Griffa, A., . . . Sporns, O. (2014).
Resting-brain functional connectivity predicted by analytic mea-
sures of network communication. Proceedings of the National
Academy of Sciences, 111(2), 833–838. https://doi.org/10.1073/
pnas.1315529111
Gordon, E. M., Laumann, T. O., Gilmore, A. W., Newbold, D. J.,
Greene, D. J., Berg, J. J., . . . Dosenbach, N. U. F. (2017). Precision
functional mapping of individual human brains. Neuron, 95(4),
791–807.e7. https://doi.org/10.1016/j.neuron.2017.07.011
Gosak, M., Markoviˇc, R., Dolenšek, J., Slak Rupnik, M., Marhl, M.,
(2018). Network science of biological
Stožer, A., & Perc, M.
systems at different scales: A review. Physics of Life Reviews, 24,
118–135. https://doi.org/10.1016/J.PLREV.2017.11.003
Gouw, A. A., & Stam, C. J. (2016). Electroencephalography in the
differential diagnosis of dementia. Epileptologie, 33, 173–182.
Gouw, A. A., Alsema, A. M., Tijms, B. M., Borta, A., Scheltens, P.,
Stam, C. J., & van der Flier, W. M. (2017). EEG spectral analysis as
a putative early prognostic biomarker in nondemented, amyloid
positive subjects. Neurobiology of Aging, 57, 133–142. https://
doi.org/10.1016/j.neurobiolaging.2017.05.017
Gratton, C., Laumann, T. O., Nielsen, A. N., Greene, D. J., Gordon,
E. M., Gilmore, A. W., . . . Petersen, S. E.
(2018). Functional
brain networks are dominated by stable group and individual fac-
tors, not cognitive or daily variation. Neuron, 98(2), 439–452.e5.
https://doi.org/10.1016/j.neuron.2018.03.035
Griffa, A., Ricaud, B., Benzi, K., Bresson, X., Daducci, A.,
Vandergheynst, P., . . . Hagmann, P. (2017). Transient networks
of spatio-temporal connectivity map communication pathways
in brain functional systems. NeuroImage, 155, 490–502. https://
doi.org/10.1016/j.neuroimage.2017.04.015
Gross, T., Dommar D’lima, C. J., & Blasius, B. (2006). Epidemic
dynamics on an adaptive network. Physical Review Letters, 96.
https://doi.org/10.1103/PhysRevLett.96.208701
Network Neuroscience
988
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
Guo, D., Trajanovski, S., van de Bovenkamp, R., Wang, H., &
Van Mieghem, P.
(2013). Epidemic threshold and topologi-
cal structure of susceptible-infectious-susceptible epidemics in
adaptive networks. Physical Review E, Statistical, Nonlinear,
and Soft Matter Physics, 88(4), 042802. https://doi.org/10.1103/
PhysRevE.88.042802
Hallquist, M. N., & Hillary, F. G. (2019). Graph theory approaches
to functional network organization in brain disorders: A critique
for a brave new small-world. Network Neuroscience, 3(1), 1–26.
https://doi.org/10.1162/netn_a_00054
Hansen, E. C. A., Battaglia, D., Spiegler, A., Deco, G., & Jirsa, V. K.
(2015). Functional connectivity dynamics: Modeling the switch-
ing behavior of the resting state. NeuroImage, 105, 525–535.
https://doi.org/10.1016/J.NEUROIMAGE.2014.11.001
Harriger, L., van den Heuvel, M. P., & Sporns, O. (2012). Rich
club organization of macaque cerebral cortex and its role in net-
work communication. PLoS ONE, 7(9), e46497. https://doi.org/
10.1371/journal.pone.0046497
Hellyer, P. J., Jachs, B., Clopath, C., & Leech, R. (2016). Local inhib-
itory plasticity tunes macroscopic brain dynamics and allows the
emergence of functional brain networks. NeuroImage, 124(Pt. A),
85–95. https://doi.org/10.1016/j.neuroimage.2015.08.069
Hilbert, D. (1902). Mathematical problems. Bulletin of the American
Mathematical Society, 8(10), 437–480. https://doi.org/10.1090/
S0002-9904-1902-00923-3
Hillebrand, A., Tewarie, P., van Dellen, E., Yu, M., Carbo, E. W. S.,
Douw, L., . . . Stam, C. J. (2016). Direction of information flow
in large-scale resting-state networks is frequency-dependent.
Proceedings of the National Academy of Sciences, 113(14),
3867–3872. https://doi.org/10.1073/pnas.1515657113
Hodkinson, D. J., O’Daly, O., Zunszain, P. A., Pariante, C. M.,
Lazurenko, V., Zelaya, F. O., . . . Williams, S. C. R. (2014). Circa-
dian and homeostatic modulation of functional connectivity and
regional cerebral blood flow in humans under normal entrained
Journal of Cerebral Blood Flow and Metabolism,
conditions.
34(9), 1493–1499. https://doi.org/10.1038/jcbfm.2014.109
Honey, C. J., Kotter, R., Breakspear, M., & Sporns, O. (2007). Net-
work structure of cerebral cortex shapes functional connectivity
on multiple time scales. Proceedings of the National Academy
of Sciences, 104(24), 10240–10245.
Honey, C. J., & Sporns, O. (2008). Dynamical consequences of le-
sions in cortical networks. Human Brain Mapping, 29(7), 802–809.
Horn, A., Reich, M., Vorwerk, J., Li, N., Wenzel, G., Fang, Q., . . .
(2017). Connectivity predicts deep brain stimula-
Fox, M. D.
tion outcome in Parkinson disease. Annals of Neurology, 82(1),
67–78. https://doi.org/10.1002/ana.24974
Hugdahl, K., & Sommer, I. E.
(2018). Auditory verbal hallucina-
tions in schizophrenia from a levels of explanation perspective.
Schizophrenia Bulletin, 44(2), 234–241. https://doi.org/10.1093/
schbul/sbx142
Hughes, R. I. G. (1997). Models and representation. Philosophy of
Science, 64, S325–S336. https://doi.org/10.1086/392611
Hutchison, R. M., Womelsdorf, T., Allen, E. A., Bandettini, P. A.,
Calhoun, V. D., Corbetta, M.,
(2013). Dy-
namic functional connectivity: Promise, issues, and interpre-
tations. NeuroImage, 80, 360–378. https://doi.org/10.1016/j.
neuroimage.2013.05.079
. Chang, C.
.
.
Insel, T., Cuthbert, B., Garvey, M., Heinssen, R., Pine, D. S., Quinn,
K., . . . Wang, P. (2010). Research Domain Criteria (RDoc): To-
ward a new classification framework for research on mental dis-
orders. American Journal of Psychiatry, 167(7), 748–751. https://
doi.org/10.1176/appi.ajp.2010.09091379
Jackson, J. H. (1884). The Croonian Lectures on evolution and dis-
solution of the nervous system. British Medical Journal, 1(1215),
703–707.
Jin, S. H., Seol, J., Kim, J. S., & Chung, C. K. (2011). How reliable
are the functional connectivity networks of MEG in resting states?
Journal of Neurophysiology, 106(6), 2888–2895.
Jones, D. T., Knopman, D. S., Gunter,
J.,
Vemuri, P., Boeve, B. F., . . . Jack, C. R. (2016). Cascading network
failure across the Alzheimer’s disease spectrum. Brain, 139(2),
547–562. https://doi.org/10.1093/brain/awv338
J. L., Graff-Radford,
Kale, P., Zalesky, A., Gollo, L. L., & Sporns, O. (2018). Estimating the
impact of structural directionality: How reliable are undirected
connectomes? Network Neuroscience, 2(2), 259–284. https://
doi.org/10.1162/netn_a_00040
Klomjai, W., Katz, R., & Lackmy-Vallée, A. (2015). Basic principles
of transcranial magnetic stimulation (TMS) and repetitive TMS
(rTMS). Annals of Physical and Rehabilitation Medicine, 58(4),
208–213. https://doi.org/10.1016/j.rehab.2015.05.005
Koenis, M. M. G., Romeijn, N., Piantoni, G., Verweij, I., van der
Werf, Y. D., Van Someren, E. J. W., & Stam, C. J. (2013). Does
sleep restore the topology of functional brain networks? Human
Brain Mapping, 34(2), 487–500. https://doi.org/10.1002/hbm.
21455
Koirala, N., Fleischer, V., Glaser, M., Zeuner, K. E., Deuschl, G.,
Volkmann, J., . . . Groppa, S. (2018). Frontal lobe connectiv-
ity and network community characteristics are associated with
the outcome of subthalamic nucleus deep brain stimulation in
patients with Parkinson’s disease. Brain Topography, 31(2),
311–321. https://doi.org/10.1007/s10548-017-0597-4
Kramer, M. A., & Cash, S. S. (2012). Epilepsy as a disorder of cortical
network organization. Neuroscientist, 18(4), 360–372.
Kringelbach, M. L., McIntosh, A. R., Ritter, P., Jirsa, V. K., & Deco,
G. (2015). The rediscovery of slowness: Exploring the timing of
cognition. Trends in Cognitive Sciences, 19(10), 616–628. https://
doi.org/10.1016/j.tics.2015.07.011
Kuhn, T. S. (1962). The structure of scientific revolutions. Chicago,
IL: University of Chicago Press.
Lindenberg, R., Nachtigall, L., Meinzer, M., Sieg, M. M., & Flöel,
(2013). Differential effects of dual and unihemispheric mo-
A.
Journal of Neuroscience,
tor cortex stimulation in older adults.
33(21), 9176–9183. https://doi.org/10.1523/JNEUROSCI.0055-
13.2013
Liuzzi, L., Gascoyne, L. E., Tewarie, P. K., Barratt, E. L., Boto, E., &
Brookes, M. J. (2017). Optimising experimental design for MEG
resting state functional connectivity measurement. NeuroImage,
155, 565–576. https://doi.org/10.1016/j.neuroimage.2016.11.
064
Lurie, D. J., Kessler, D., Bassett, D. S., Betzel, R. F., Breakspear, M.,
Keilholz, S., . . . Calhoun, V. D. (2018). On the nature of resting
fMRI and time-varying functional connectivity. PsyArxiv.
Lydon-Staley, D. M., Barnett, I., Satterthwaite, T. D., & Bassett, D. S.
(2018). Digital phenotyping for psychiatry: Accommodating
Network Neuroscience
989
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
data and theory with network science methodologies. PsyArxiv.
https://doi.org/10.31234/OSF.IO/MYB8C
Maier-Hein, K. H., Neher, P. F., Houde,
J.-C., Côté, M.-A.,
Garyfallidis, E., Zhong, J., . . . Descoteaux, M. (2017). The chal-
lenge of mapping the human connectome based on diffusion
tractography. Nature Communications, 8(1), 1349. https://doi.
org/10.1038/s41467-017-01285-x
(2017).
Mehta-Pandejee, G., Robinson, P. A., Henderson, J. A., Aquino,
Inference of direct and multistep
K. M., & Sarkar, S.
effective connectivities from functional connectivity of the brain
and of relationships to cortical geometry. Journal of Neuroscience
Methods, 283, 42–54. https://doi.org/10.1016/j.jneumeth.2017.
03.014
Meier, J., Tewarie, P., Hillebrand, A., Douw, L., Van Dijk, B. W.,
Stufflebeam, S. M., & Van Mieghem, P. (2016). A mapping be-
tween structural and functional brain networks. Brain Connectiv-
ity, 6(4). https://doi.org/10.1089/brain.2015.0408
Messé, A., Rudrauf, D., Benali, H., & Marrelec, G. (2014). Relat-
ing structure and function in the human brain: Relative contri-
butions of anatomy, stationary dynamics, and nonstationarities.
PLoS Computational Biology, 10(3), e1003530.
Miši´c, B., Sporns, O. & McIntosh, A. R. (2014). Communication ef-
ficiency and congestion of signal traffic in large-scale brain net-
works. PLoS Computational Biology, 10(1), e1003427. https://
doi.org/10.1371/journal.pcbi.1003427
Moon, J.-Y., Lee, U., Blain-Moraes, S., & Mashour, G. A. (2015).
General relationship of global topology, local dynamics, and di-
rectionality in large-scale brain networks. PLoS Computational
Biology, 11(4), e1004225. https://doi.org/10.1371/journal.pcbi.
1004225
Morgan, S. E., Achard, S., Termenon, M., Bullmore, E. T., & Vértes,
P. E. (2018). Low-dimensional morphospace of topological mo-
tifs in human fMRI brain networks. Network Neuroscience,
02(02), 285–302. https://doi.org/10.1162/netn_a_00038
Nissen, I. A., Stam, C. J., van Straaten, E. C. W., Wottschel, V.,
Reijneveld, J. C., Baayen, J. C., . . . Hillebrand, A.
(2018). Lo-
calization of the epileptogenic zone using interictal MEG and
machine learning in a large cohort of drug-resistant epilepsy pa-
tients. Frontiers in Neurology, 9, 647. https://doi.org/10.3389/
fneur.2018.00647
Noble, S., Scheinost, D., Finn, E. S., Shen, X., Papademetris, X.,
McEwen, S. C., . . . Constable, R. T. (2017). Multisite reliability of
MR-based functional connectivity. NeuroImage, 146, 959–970.
https://doi.org/10.1016/J.NEUROIMAGE.2016.10.020
Numan, T., Slooter, A. J. C., van der Kooi, A. W., Hoekman, A. M. L.,
Suyker, W. J. L., Stam, C. J., & van Dellen, E. (2017). Functional
connectivity and network analysis during hypoactive delirium
and recovery from anesthesia. Clinical Neurophysiology, 128(6),
914–924. https://doi.org/10.1016/j.clinph.2017.02.022
O’Neill, G. C., Tewarie, P., Vidaurre, D., Liuzzi, L., Woolrich,
M. W., & Brookes, M.
large-scale
electrophysiological networks: A technical review. NeuroIm-
age, 180(Pt. B), 559–576. https://doi.org/10.1016/j.neuroimage.
2017.10.003
(2018). Dynamics of
J.
Olde Dubbelink, K. T. E., Hillebrand, A., Twisk, J. W. R., Deijen,
J. B., Stoffers, D., Schmand, B. A., . . . Berendse, H. W. (2014).
Predicting dementia in parkinson disease by combining neuro-
physiologic and cognitive markers. Neurology, 82(3), 263–270.
https://doi.org/10.1212/WNL.0000000000000034
Pai, S., & Bader, G. D. (2018). Patient similarity networks for precision
Journal of Molecular Biology, 430(18), 2924–2938.
medicine.
https://doi.org/10.1016/J.JMB.2018.05.037
Paluš, M., & Vejmelka, M. (2007). Directionality of coupling from
bivariate time series: How to avoid false causalities and missed
connections. Physical Review E, 75(5), 056211. https://doi.org/
10.1103/PhysRevE.75.056211
Papadopoulos, L., Kim, J. Z., Kurths, J., & Bassett, D. S. (2017). De-
velopment of structural correlations and synchronization from
adaptive rewiring in networks of Kuramoto oscillators. Chaos,
27(7), 073115. https://doi.org/10.1063/1.4994819
Pastor-Satorras, R., Castellano, C., Van Mieghem, P., & Vespig-
nani, A. (2015). Epidemic processes in complex networks. Re-
views of Modern Physics, 87(3), 925–979. https://doi.org/10.
1103/RevModPhys.87.925
Perera, T., George, M. S., Grammer, G., Janicak, P. G., Pascual-
Leone, A., & Wirecki, T. S. (2016). The Clinical TMS Society con-
sensus review and treatment recommendations for TMS therapy
for major depressive disorder. Brain Stimulation, 9(3), 336–346.
https://doi.org/10.1016/j.brs.2016.03.010
Poldrack, R. A., Baker, C.
J.,
I., Durnez,
Matthews, P. M., Munafò, M. R., . . . Yarkoni, T. (2017). Scanning
the horizon: Towards transparent and reproducible neuroimaging
research. Nature Reviews Neuroscience, 18(2), 115–126. https://
doi.org/10.1038/nrn.2016.167
J., Gorgolewski, K.
Poldrack, R. A., Laumann, T. O., Koyejo, O., Gregory, B., Hover, A.,
Chen, M. Y., . . . Mumford, J. A. (2015). Long-term neural and
physiological phenotyping of a single human. Nature Communi-
cations, 6, 8885. https://doi.org/10.1038/ncomms9885
Power, J. D., Schlaggar, B. L., Lessov-Schlaggar, C. N., & Petersen,
S. E. (2013). Evidence for hubs in human functional brain net-
works. Neuron, 79(4), 798–813. https://doi.org/10.1016/j.neuron.
2013.07.035
Pozo, K., & Goda, Y. (2010). Unraveling mechanisms of homeo-
static synaptic plasticity. Neuron, 66(3), 337–351. https://doi.
org/10.1016/j.neuron.2010.04.028
Prinz, A. A., Bucher, D., & Marder, E. (2004). Similar network ac-
tivity from disparate circuit parameters. Nature Neuroscience,
7(12), 1345–1352. https://doi.org/10.1038/nn1352
Proix, T., Bartolomei, F., Guye, M., & Jirsa, V. K. (2017). Individual
brain structure and modelling predict seizure propagation. Brain,
140(3), 641–654. https://doi.org/10.1093/brain/awx004
Rilling, J. K., & van den Heuvel, M. P. (2018). Comparative primate
connectomics. Brain, Behavior and Evolution, 91(3), 170–179.
https://doi.org/10.1159/000488886
Robinson, P., Henderson, J., Matar, E., Riley, P., & Gray, R. (2009).
Dynamical reconnection and stability constraints on cortical net-
work architecture. Physical Review Letters, 103(10), 1–4. https://
doi.org/10.1103/PhysRevLett.103.108104
Rocha, R. P., Koçillari, L., Suweis, S., Corbetta, M., & Maritan, A.
(2018). Homeostatic plasticity and emergence of functional net-
works in a whole-brain model at criticality. Scientific Reports,
8(1), 15682. https://doi.org/10.1038/s41598-018-33923-9
Rubinov, M., McIntosh, A. R., Valenzuela, M. J., & Breakspear, M.
(2009). Simulation of neuronal death and network recovery in
Network Neuroscience
990
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
a computational model of distributed cortical activity. American
Journal of Geriatric Psychiatry, 17(3), 210–217.
Sale, M. V., Mattingley, J. B., Zalesky, A., & Cocchi, L. (2015). Imaging
human brain networks to improve the clinical efficacy of non-
invasive brain stimulation. Neuroscience and Biobehavioral
Reviews, 57, 187–198. https://doi.org/10.1016/j.neubiorev.2015.
09.010
Scannell, J. W., Burns, G. A., Hilgetag, C. C., O’Neil, M. A.,
& Young, M. P.
(1999). The connectional organization of
the cortico-thalamic system of the cat. Cerebral Cortex, 9(3),
277–299.
Schmidt, C., Collette, F., Cajochen, C., & Peigneux, P. (2007). A
time to think: Circadian rhythms in human cognition. Cogni-
tive Neuropsychology, 24(7), 755–789. https://doi.org/10.1080/
02643290701754158
Schoonheim, M. M., Meijer, K. A., & Geurts, J. J. G. (2015). Net-
in multiple sclero-
work collapse and cognitive impairment
sis. Frontiers in Neurology, 6, 82. https://doi.org/10.3389/fneur.
2015.00082
Senden, M., Reuter, N., van den Heuvel, M. P., Goebel, R., &
Deco, G. (2017). Cortical rich club regions can organize state-
dependent functional network formation by engaging in oscilla-
tory behavior. NeuroImage, 146, 561–574. https://doi.org/10.
1016/J.NEUROIMAGE.2016.10.044
Senden, M., Reuter, N., van den Heuvel, M. P., Goebel, R., Deco, G.,
& Gilson, M. (2018). Task-related effective connectivity reveals
that the cortical rich club gates cortex-wide communication.
Human Brain Mapping, 39(3), 1246–1262. https://doi.org/10.1002/
hbm.23913
Seth, A. K., Barrett, A. B., & Barnett, L. (2015). Granger causality anal-
ysis in neuroscience and neuroimaging. Journal of Neuroscience,
35(8), 3293–3297. https://doi.org/10.1523/JNEUROSCI.4399-14.
2015
Sinke, M. R. T., Dijkhuizen, R. M., Caimo, A., Stam, C. J., &
Bayesian exponential random graph
Otte, W. M.
modeling of whole-brain structural networks across lifespan.
NeuroImage, 135, 79–91. https://doi.org/10.1016/j.neuroimage.
2016.04.066
(2016).
Sinke, M. R. T., Otte, W. M., Christiaens, D., Schmitt, O., Leemans,
A., van der Toorn, A., . . . Dijkhuizen, R. M.
(2018). Diffusion
MRI-based cortical connectome reconstruction: Dependency on
tractography procedures and neuroanatomical characteristics.
Brain Structure and Function, 223(5), 2269–2285. https://doi.
org/10.1007/s00429-018-1628-y
Smith, J. F., Pillai, A., Chen, K., & Horwitz, B. (2012). Effective con-
nectivity modeling for fMRI: Six issues and possible solutions us-
ing linear dynamic systems. Frontiers in Systems Neuroscience,
5. https://doi.org/10.3389/fnsys.2011.00104
Sporns, O. (2010). Networks of the brain. Cambridge, MA: MIT Press.
Stam, C. J. (2014). Modern network science of neurological disor-
ders. Nature Reviews Neuroscience, 15(10), 683–695. https://
doi.org/10.1038/nrn3801
Stam, C. J., Hillebrand, A. J., Wang, H., & Van Mieghem, P. (2010).
Emergence of modular structure in a large-scale brain network
with interactions between dynamics and connectivity. Frontiers
in Computational Neuroscience, 4, 133.
Stam, C. J., Tewarie, P., van Dellen, E., van Straaten, E. C. W.,
Hillebrand, A., & Van Mieghem, P. (2014). The trees and the for-
est: Characterization of complex brain networks with minimum
spanning trees. International Journal of Psychophysiology, 92(3),
129–138. https://doi.org/10.1016/j.ijpsycho.2014.04.001
Stam, C. J., van Straaten, E. C. W., van Dellen, E., Tewarie, P.,
Gong, G., Hillebrand, A., . . . Van Mieghem, P. (2016). The
relation between structural and functional connectivity pat-
terns in complex brain networks. International Journal of Psycho-
physiology, 103, 149–160. https://doi.org/10.1016/j.ijpsycho.
2015.02.011
Stefanik, L., Erdman, L., Ameis, S. H., Foussias, G., Mulsant, B. H.,
Behdinan, T., . . . Voineskos, A. N. (2018). Brain-behavior par-
ticipant similarity networks among youth and emerging adults
with schizophrenia spectrum, autism spectrum, or bipolar disor-
der and matched controls. Neuropsychopharmacology, 43(5),
1180–1188. https://doi.org/10.1038/npp.2017.274
Stephan, K. E., Bach, D. R., Fletcher, P. C., Flint, J., Frank, M. J.,
Friston, K. J., . . . Breakspear, M. (2016). Charting the landscape
of priority problems in psychiatry, part 1: Classification and di-
agnosis. The Lancet Psychiatry, 3(1), 77–83. https://doi.org/10.
1016/S2215-0366(15)00361-2
Stephan, K. E., Binder, E. B., Breakspear, M., Dayan, P., Johnstone, E.
C., Meyer-Lindenberg, A., . . ., Friston, K. J. (2016). Charting the
landscape of priority problems in psychiatry, part 2: Pathogene-
sis and aetiology. The Lancet Psychiatry, 3(1), 84–90. https://doi.
org/10.1016/S2215-0366(15)00360-0
Telesford, Q. K., Morgan, A. R., Hayasaka, S., Simpson,
S. L., Barret, W., Kraft, R. A., . . . Laurienti, P. J. (2010). Repro-
ducibility of graph metrics in fMRI networks. Frontiers in Neu-
roinformatics, 4(December), 117. https://doi.org/10.3389/fninf.
2010.00117
Tewarie, P., Hillebrand, A., van Dellen, E., Schoonheim, M. M.,
Barkhof, F., Polman, C. H., . . . Stam, C. J. (2014). Structural
degree predicts functional network connectivity: A multimodal
resting-state fMRI and MEG study. NeuroImage, 97, 296–307.
https://doi.org/10.1016/j.neuroimage.2014.04.038
Tewarie, P., Hunt, B. A. E., O’Neill, G. C., Byrne, A., Aquino,
K., Bauer, M., . . . Brookes, M. J.
(2018). Relationships be-
tween neuronal oscillatory amplitude and dynamic functional
connectivity. Cerebral Cortex. https://doi.org/10.1093/cercor/
bhy136
Tewarie, P., van Dellen, E., Hillebrand, A., & Stam, C. J. (2015). The
minimum spanning tree: An unbiased method for brain network
analysis. NeuroImage, 104, 177–188. https://doi.org/10.1016/j.
neuroimage.2014.10.015
Tijms, B. M., Wink, A. M., de Haan, W., van der Flier, W. M., Stam,
C. J., Scheltens, P., . . . Barkhof, F. (2013). Alzheimer’s disease:
Connecting findings from graph theoretical studies of brain net-
works. Neurobiology of Aging, 34(8), 2023–2036. https://doi.
org/10.1016/j.neurobiolaging.2013.02.020
Trajanovski, S., Guo, D., & Van Mieghem, P. (2015). From epi-
demics to information propagation: Striking differences in struc-
turally similar adaptive network models. Physical Review E, 92(3),
030801. https://doi.org/10.1103/PhysRevE.92.030801
van Dellen, E., Hillebrand, A., Douw, L., Heimans, J. J., Reijneveld, J. C.,
& Stam, C. J. (2013). Local polymorphic delta activity in cortical
Network Neuroscience
991
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
lesions causes global decreases in functional connectivity. NeuroImage,
83. https://doi.org/10.1016/j.neuroimage.2013.06.009
using graph theory. PLoS ONE, 5(10), e13701. https://doi.org/
10.1371/journal.pone.0013701
van Dellen, E., Sommer,
I. E., Bohlken, M. M., Tewarie, P.,
Draaisma, L., Zalesky, A., . . . Stam, C. J. (2018). Minimum span-
ning tree analysis of the human connectome. Human Brain Map-
ping. https://doi.org/10.1002/hbm.24014
van Dellen, E., van der Kooi, A. W., Numan, T., Koek, H. L.,
(2014).
Klijn, F. A. M., Buijsrogge, M. P., . . . Slooter, A. J. C.
Decreased functional connectivity and disturbed directional-
ity of information flow in the electroencephalography of in-
tensive care unit patients with delirium after cardiac surgery.
Anesthesiology, 121(2), 328–335. https://doi.org/10.1097/ALN.
0000000000000329
van den Heuvel, M. P., Bullmore, E. T., & Sporns, O. (2016).
Comparative connectomics. Trends in Cognitive Sciences, 20(5),
345–361. https://doi.org/10.1016/j.tics.2016.03.001
van den Heuvel, M. P., de Lange, S. C., Zalesky, A., Seguin, C.,
Yeo, B. T. T., & Schmidt, R. (2017). Proportional thresholding in
resting-state fMRI functional connectivity networks and conse-
quences for patient-control connectome studies: Issues and rec-
ommendations. NeuroImage, 152, 437–449.
van den Heuvel, M. P., Scholtens, L. H., de Lange, S. C., Pijnenburg,
R., Cahn, W., van Haren, N. E. M., . . . Rilling, J. K. (2018). Evolu-
tionarily developed connections compromised in schizophrenia.
bioRxiv. https://doi.org/10.1101/387506
van den Heuvel, M. P., & Sporns, O. (2013). Network hubs in the
human brain. Trends in Cognitive Sciences, 17(12), 683–696.
https://doi.org/10.1016/j.tics.2013.09.012
van Diessen, E., Diederen, S. J. H., Braun, K. P. J., Jansen, F. E., &
(2013). Functional and structural brain networks in
Stam, C. J.
epilepsy: What have we learned? Epilepsia, 54(11), 1855–1865.
https://doi.org/10.1111/epi.12350
van Diessen, E., Otte, W. M., Braun, K. P. J., Stam, C. J., & Jansen,
F. E. (2013). Improved diagnosis in children with partial epilepsy
using a multivariable prediction model based on EEG network
characteristics. PLoS ONE, 8(4), e59764. https://doi.org/10.1371/
journal.pone.0059764
Van Mieghem, P.
(2011). Graph spectra for complex networks.
New York, NY: Cambridge University Press.
Van Mieghem, P. (2018). Directed graphs and mysterious complex
eigenvalues.
Van Mieghem, P., & van de Bovenkamp, R. (2015). Accuracy cri-
terion for the mean-field approximation in susceptible-infected-
susceptible epidemics on networks. Physical Review E, 91(3).
https://doi.org/10.1103/PhysRevE.91.032812
van Os, J., Guloksuz, S., Vijn, T. W., Hafkenscheid, A., & Delespaul,
P.
(2019). The evidence-based group-level symptom-reduction
model as the organizing principle for mental health care: Time
for change? World Psychiatry, 18(1), 88–96. https://doi.org/10.
1002/wps.20609
van Wijk, B. C. M., Cagnan, H., Litvak, V., Kühn, A. A., & Friston,
K. J. (2018). Generic dynamic causal modelling: An illustrative
application to Parkinson’s disease NeuroImage, 181, 818–830.
https://doi.org/10.1016/j.neuroimage.2018.08.039
van Wijk, B. C. M., Stam, C. J., & Daffertshofer, A. (2010). Com-
paring brain networks of different size and connectivity density
Vasa, F., Bullmore, E. T., & Patel, A. X. (2018). Probabilistic thresh-
olding of functional connectomes: Application to schizophrenia.
NeuroImage, 172, 326–340.
Vecchio, D. F., Miraglia, D. F., Iberite, D. F., Lacidogna, D. G.,
Guglielmi, D. V., Marra, D. C., . . . Rossini, P. P. M. (2018).
Sustainable method for Alzheimer dementia prediction in mild
cognitive impairment: Electroencephalographic connectivity and
graph theory combined with apolipoprotein. E. Annals of Neurol-
ogy, 84(2), 302–314. https://doi.org/10.1002/ana.25289
Vidaurre, D., Hunt, L. T., Quinn, A. J., Hunt, B. A. E., Brookes, M. J.,
(2018). Spontaneous corti-
Nobre, A. C., & Woolrich, M. W.
cal activity transiently organises into frequency specific phase-
coupling networks. Nature Communications, 9(1), 2987. https://
doi.org/10.1038/s41467-018-05316-z
Wang, Y., Balaji, V., Kaniyappan, S., Krüger, L.,
Irsen, S.,
Tepper, K., . . . Mandelkow, E.-M. (2017). The release and
tau via exosomes. Molecular
trans-synaptic transmission of
Neurodegeneration, 12(1), 5. https://doi.org/10.1186/s13024-
016-0143-y
Wang, Y., Hutchings, F., & Kaiser, M. (2015). Computational modeling
of neurostimulation in brain diseases. In Progress in brain re-
search (Vol. 222, pp. 191–228). https://doi.org/10.1016/bs.pbr.
2015.06.012
Watts, D. J., & Strogatz, S. H. (1998). Collective dynamics of “small-
world” networks. Nature, 393(6684), 440–442.
Weigand, A., Horn, A., Caballero, R., Cooke, D., Stern, A. P., Taylor,
S. F., . . . Fox, M. D. (2018). Prospective validation that subgen-
ual connectivity predicts antidepressant efficacy of transcranial
magnetic stimulation sites. Biological Psychiatry, 84(1), 28–37.
https://doi.org/10.1016/j.biopsych.2017.10.028
Wen, X., Rangarajan, G., & Ding, M. (2013). Is Granger causality
a viable technique for analyzing fMRI data? PLoS ONE, 8(7),
e67428. https://doi.org/10.1371/journal.pone.0067428
White, J. G., Southgate, E., Thomson, J. N., & Brenner, S. (1986). The
structure of the nervous system of the nematode Caenorhabditis
elegans. Philosophical Transactions of
the Royal Society of
London B: Biological Sciences, 314(1165), 1–340.
Wilke, C., van Drongelen, W., Kohrman, M., & He, B. (2010). Neo-
cortical seizure foci localization by means of a directed transfer
function method. Epilepsia, 51(4), 564–572. https://doi.org/10.
1111/j.1528-1167.2009.02329.x
Xia, C. H., Ma, Z., Ciric, R., Gu, S., Betzel, R. F., Kaczkurkin, A. N.,
. . . Satterthwaite, T. D. (2018). Linked dimensions of psycho-
pathology and connectivity in functional brain networks. Nature
Communications, 9(1), 3003. https://doi.org/10.1038/s41467-
018-05317-y
Yin, J., & Yuan, Q. (2015). Structural homeostasis in the nervous sys-
tem: A balancing act for wiring plasticity and stability. Frontiers
in Cellular Neuroscience, 8, 439. https://doi.org/10.3389/fncel.
2014.00439
Zalesky, A., Fornito, A., Cocchi, L., Gollo, L. L., van den Heuvel,
M. P., & Breakspear, M. (2016). Connectome sensitivity or speci-
ficity: Which is more important? NeuroImage, 142, 407–420.
https://doi.org/10.1016/j.neuroimage.2016.06.035
Network Neuroscience
992
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The road ahead in clinical network neuroscience
Zalesky, A., Fornito, A., & Bullmore, E. T. (2010). Network-based
statistic: Identifying differences in brain networks. NeuroImage,
53(4), 1197–1207. https://doi.org/10.1016/j.neuroimage.2010.
06.041
Zamora-López, G., Chen, Y., Deco, G., Kringelbach, M. L., & Zhou,
C.
(2016). Functional complexity emerging from anatomical
constraints in the brain: The significance of network modularity
and rich-clubs. Scientific Reports, 6(1), 38424. https://doi.org/10.
1038/srep38424
Zhang, Z., Nie, S., & Chen, L.
(2018). Targeting prion-like pro-
tein spreading in neurodegenerative diseases. Neural Regenera-
tion Research, 13(11), 1875. https://doi.org/10.4103/1673-5374.
239433
Zheng, X., & Schlaug, G. (2015). Structural white matter changes
in descending motor tracts correlate with improvements in mo-
tor impairment after undergoing a treatment course of tDCS
and physical therapy. Frontiers in Human Neuroscience, 9, 229.
https://doi.org/10.3389/fnhum.2015.00229
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
/
3
4
9
6
9
1
8
6
6
8
4
3
n
e
n
_
a
_
0
0
1
0
3
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
993