Retrosplenial Cortical Connectivity with
Frontal Basal Ganglia Networks
Megan E. Monko and Sarah R. Heilbronner
Abstract
■ Previous studies of the retrosplenial cortex (RSC) have focused
on its role in navigation and memory, consistent with its well-
established medial temporal connections, but recent evidence
also suggests a role for this region in reward and decision making.
Because function is determined largely by anatomical connec-
tions, and to better understand the anatomy of RSC, we used
tract-tracing methods to examine the anatomical connectivity be-
tween the rat RSC and frontostriatal networks (canonical reward
and decision-making circuits). We find that, among frontal cortical
regions, RSC bidirectionally connects most strongly with the ante-
rior cingulate cortex, but also with an area of the central–medial
orbito-frontal cortex. RSC projects to the dorsomedial striatum,
and its terminal fields are virtually encompassed by the frontal-
striatal projection zone, suggestive of functional convergence
through the basal ganglia. This overlap is driven by anterior cingu-
late cortex, prelimbic cortex, and orbito-frontal cortex, all of
which contribute to goal-directed decision making, suggesting
that the RSC is involved in similar processes. ■
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
0
9
6
1
9
1
3
6
6
8
/
/
j
o
c
n
_
a
_
0
1
6
9
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
The retrosplenial cortex (RSC) in rodents occupies a large
territory in the posterior medial part of the cerebral cortex.
Its strong connections with medial temporal lobe areas are
well-established (Sugar, Witter, van Strien, & Cappaert,
2011; van Groen & Wyss, 1990, 1992, 2003). The promi-
nence of these connections has motivated theorizing on
RSC function, which has, not surprisingly, focused on
spatial navigation and memory (Kaboodvand, Bäckman,
Nyberg, & Salami, 2018; Auger & Maguire, 2013; Auger,
Mullally, & Maguire, 2012; Keene & Bucci, 2008, 2009;
Vann, Aggleton, & Maguire, 2009; Harker & Whishaw,
2004; Maviel, Durkin, Menzaghi, & Bontempi, 2004;
Parron & Save, 2004; Cho & Sharp, 2001; Maguire, 2001;
Cooper & Mizumori, 1999; Valenstein et al., 1987). For ex-
ample, lesions in the RSC lead to deficits in performance on
a water maze test (Sutherland, Wishaw, & Kolb, 1988) and
to impairments in allocentric spatial memory (Pothuizen,
Aggleton, & Vann, 2008; Vann & Aggleton, 2002, 2004).
However, RSC likely has a broader and more integrative
function than is currently appreciated. In particular, there
is compelling evidence that RSC may play a crucial role in
reward-guided decision making (Hattori, Danskin, Babic,
Mlynaryk, & Komiyama, 2019; Powell et al., 2017; Vedder,
Miller, Harrison, & Smith, 2017; Nelson, Hindley, Haddon,
Vann, & Aggleton, 2014; Tabuchi et al., 2005). For exam-
ple, neural population activity in the RSC encodes a persis-
tent value, with flexible reward history encoding (Hattori
et al., 2019). In addition, the RSC plays a central albeit
poorly delineated role in the default mode network (DMN),
University of Minnesota, Twin Cities
© 2021 Massachusetts Institute of Technology
a network of functionally connected brain regions associated
with high activity at rest, self-projection, episodic memory,
and reward monitoring (Raichle et al., 2001; Shulman et al.,
1997), which has been found in humans (Raichle et al.,
2001), nonhuman primates (Mantini et al., 2011), and rodents
(Lu et al., 2012; Upadhyay et al., 2011). Understanding the
role of RSC is particularly important because it is likely
to become a major research focus in the coming years: It
reaches the dorsal surface of the brain and, therefore, is
readily optically accessible to studies of neural activity using
calcium imaging. Future work aimed at fully understanding
the functions of RSC must take into account its roles in re-
ward, decision making, and DMN processes. However, our
ability to do so is limited by a lack of integrative knowledge
of the anatomical connections of the RSC.
Most studies of reward-guided decision making implicate
frontal cortico-striatal circuits (Fitoussi et al., 2018; Gremel
et al., 2016; Sleezer, Castagno, & Hayden, 2016; Burton,
Nakamura, & Roesch, 2015; Rothwell et al., 2015; Haber &
Behrens, 2014; Yin, Knowlton, & Balleine, 2006; Balleine,
Delgado, & Hikosaka, 2007; O’Doherty et al., 2004). The
functions of these regions and projections are very well-
studied using diverse tools, including lesions, optogenetics,
electrophysiology, and pharmacology. Distinct striatal zones
and their unique afferents have been implicated in different
components of reward-guided decision making. Broadly
speaking, there is consensus that the ventral striatum, dor-
somedial striatum, and dorsolateral striatum, along with their
frontal cortical afferents, have different roles in reward-
guided decision making (Li et al., 2016; O’Hare et al., 2016;
Kimchi & Laubach, 2009; Yin, Ostlund, Knowlton, & Balleine,
2005; Voorn, Vanderschuren, Groenewegen, Robbins, &
Journal of Cognitive Neuroscience 33:6, pp. 1096–1105
https://doi.org/10.1162/jocn_a_01699
Pennartz, 2004; Yin & Knowlton, 2004; Hassani, Cromwell,
& Schultz, 2001). One major hypothesis is that the dorso-
lateral striatum is involved with habits, the dorsomedial
striatum is involved in goal-directed decision making, and
the ventral striatum is involved in establishing stimulus-
value associations.
One way to understand the function of the RSC in
reward-guided decision making is to analyze its anatomical
connections with this canonical, frontal cortico-striatal
decision-making circuitry. Furthermore, the major subre-
gions of the DMN are the RSC and portions of the frontal
cortex, meaning the same cortical circuitry gives us insight
into both reward-guided decision making and DMN func-
tion. Previous studies have demonstrated that there are
significant projections between the RSC and the frontal
cortex and striatum in rats (Shibata & Naito, 2008;
Shibata, Kondo, & Naito, 2004; van Groen & Wyss, 1990,
1992, 2003). However, the projections to the striatum in
particular have not been analyzed in detail, especially in
light of our current understanding of how different zones
of the striatum contribute to reward-guided decision
making. Thus, in this paper, we used anatomical tract-
tracing methods in rats to analyze the connections of the
RSC with the striatum and frontal cortex.
METHODS
Overview
Connectivity of the RSC with the striatum and frontal
cortical subregions was analyzed using anatomical tract-
tracing in rats. Following surgery and perfusion, immuno-
histochemistry was performed and brain slices were
mounted on slides. Labeling in the striatum (anterograde)
and frontal cortex (anterograde and retrograde) were
charted using light microscopy. Cases were registered to
a standard brain for comparison and visualization.
Surgery and Tissue Preparation
All procedures were approved by the Institutional Animal
Care and Use Committee at the University of Minnesota.
Six adult male and five adult female Sprague Dawley rats
(weight = 230–500 g, Charles River) were used for these
studies.
At the start of surgery, animals were anesthetized using a
combination of ketamine (40–90 mg/kg, intraperitoneal
[IP]) and xylazine (5–10 mg/kg, IP). In addition, a nonsteroi-
dal anti-inflammatory drug (Carprofen, 5 mg/kg, IP or subcu-
taneous [SQ]) and an antibiotic (Baytril, 2.5 mg/kg, SQ) were
administered. Saline injections were given periodically to
maintain hydration. Animals were placed in a stereotaxic
frame on top of a heat source. Temperature and foot with-
drawal reflex were monitored throughout to ensure stable
anesthesia. Lidocaine was administered at the incision site.
The skull was exposed, and then burr holes were made to
expose the surface of the brain. The bidirectional tract-tracer
fluororuby (FR, 40–50 nl, 10% in 0.1-M phosphate buffer, pH
7.4, Invitrogen) was injected over 10 min using a 0.5-μL
Hamilton syringe. After each injection, the syringe re-
mained in situ for 10–20 min. The syringe was removed,
and the incision was closed with sutures. Atipamezole
(0.1–1.0 mg/kg, IP) was used to reverse the ketamine/
xylazine combination. Postoperatively, Carprofen (5 mg/kg,
SQ) was delivered every 24 hr for 72 hr total. Targets were
chosen using Paxinos and Watson (2014).
Animals were euthanized 10–14 days following injection
with an IP injection of a lethal dose of commercial euthanasia
solution (Euthasol, 0.22 mL/kg), and transcardially perfused
with saline followed by 4% paraformaldehyde. Brains were
extracted, postfixed overnight, and cryoprotected in increas-
ing gradients of sucrose (10%, 20%, and 30%). Serial sections
of 50 μm were cut on a freezing microtome into cryoprotec-
tant solution. Serial sections of 50 μm were cut on a freezing
microtome. One in six sections was processed free-floating
for immunocytochemistry to visualize the tracer. Tissue was
incubated in primary anti-FR (1:6000; Invitrogen) in 10%
normal goat serum and 0.3% Triton X-100 (Sigma-Aldrich)
in PO4 for 4 nights at 4°C. After rinsing, the tissue was in-
cubated in biotinylated secondary antibody followed by in-
cubation with the avidin-biotin complex solution (Vectastain
ABC kit, Vector Laboratories). Immunoreactivity was then
visualized using standard 3,30-Diaminobenzidine (DAB) pro-
cedures. Staining was intensified by incubating the tissue
for 5–15 sec in a solution of 0.05% tetrahydrochloride,
0.025% cobalt chloride, 0.02% nickel ammonium sulfate,
and 0.01% H2O2. Sections were mounted by hand onto
gel-coated slides, dehydrated, defatted in xylene, and
coverslipped with Permount.
Microscopy and Analysis
Microscopy was performed using a Zeiss AxioImager M2
microscope. Three representative cases were chosen for
in-depth analysis and 3-D rendering on the basis of their
outstanding transport and lack of contamination. Using
darkfield light microscopy, brain sections and structures,
the injection site, and cortical and striatal terminal fields
were outlined under a 2.0Å~, 4.0Å~, or 10× objective with
Neurolucida software (MBF Bioscience). Terminal fields
were considered dense when they could be visualized at a
low objective (2.0×; criterion as previously utilized in
Heilbronner, Meyer, Choi, & Haber, 2018; Heilbronner &
Haber, 2014; Haynes & Haber, 2013; Mailly, Aliane,
Groenewegen, Haber, & Deniau, 2013; Haber, Kim,
Mailly, & Calzavara, 2006). We distinguished between likely
terminal fields (thin, labeled fibers containing boutons)
and passing fibers (thick fibers without clear boutons). In
addition, using brightfield light microscopy, retrogradely
labeled cells (all within the target region) were counted
using StereoInvestigator software (MBF Bioscience) under
a 20× objective. This stereology software was utilized to
ensure that every part of a region received equal attention
and to avoid biases associated with dark staining and cell
Monko and Heilbronner
1097
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
0
9
6
1
9
1
3
6
6
8
/
/
j
o
c
n
_
a
_
0
1
6
9
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
clusters. We analyzed one out of every six sections, and density
was defined by total number of cells normalized by brain re-
gion size, with size defined by the sum of the areas of the brain
regions across the one in six sections, rather than by volume.
For each case, stacks of coronal sections were created
from chartings and scanned slides. These stacks were im-
ported into IMOD software (Boulder Laboratory for 3-D
Electron Microscopy; Kremer, Mastronarde, & McIntosh,
1996) and combined, and thus, a 3-D reconstruction that
contained the injection sites, terminal fields, and labeled
cells was created for each case separately. To merge mul-
tiple cases together, individual cases were registered in
IMOD to standard rat MRI images that had been converted
to IMOD files ( Wisner, Odintsov, Brozoski, & Brozoski,
2016). Registrations were manually checked and adjusted
according to cortical and striatal landmarks.
We wanted to compare the RSC-striatal projection zone to
previously established frontal-striatal projection zones.
Projection zones for areas CG (anterior cingulate cortex),
PL (prelimbic cortex), MO (medial orbito-frontal cortex),
VOLO (ventral and lateral orbito-frontal cortex), and IL (in-
fralimbic cortex) were drawn from Heilbronner, Rodriguez-
Romaguera, Quirk, Groenewegen, and Haber (2016). In that
paper, we combined prior maps of cortico-striatal projection
fields, with a focus on the projections at anterior–posterior
distance from bregma (AP) +1.1, where the frontal terminal
fields appeared to be maximally segregated. The database
consisted of four cases from IL, 10 from PL, five from CG,
three from MO, and nine from VOLO. To this database,
we added projections from area M2 (three cases) as de-
picted in Reep and Corwin (1999). Overlaps in terminal field
areas at three coronal A-P positions were calculated by con-
verting the pixels of the models in IMOD to a polygon and
finding the area of the intersect using custom MATLAB
(The MathWorks) code.
RESULTS
The cases included in these analyses had the bidirectional
tracer FR injected into the RSC. Resulting injection sites
were confined to the RSC and, together, covered most of
its rostral-caudal length (collectively the cores of the injec-
tion sites ranged from −4.8 to −8.8 AP). We first analyzed
the projections from the RSC to the striatum. Like most cor-
tical areas, the RSC projects to both ipsilateral and contralat-
eral striatum with predominance of the former (McGeorge
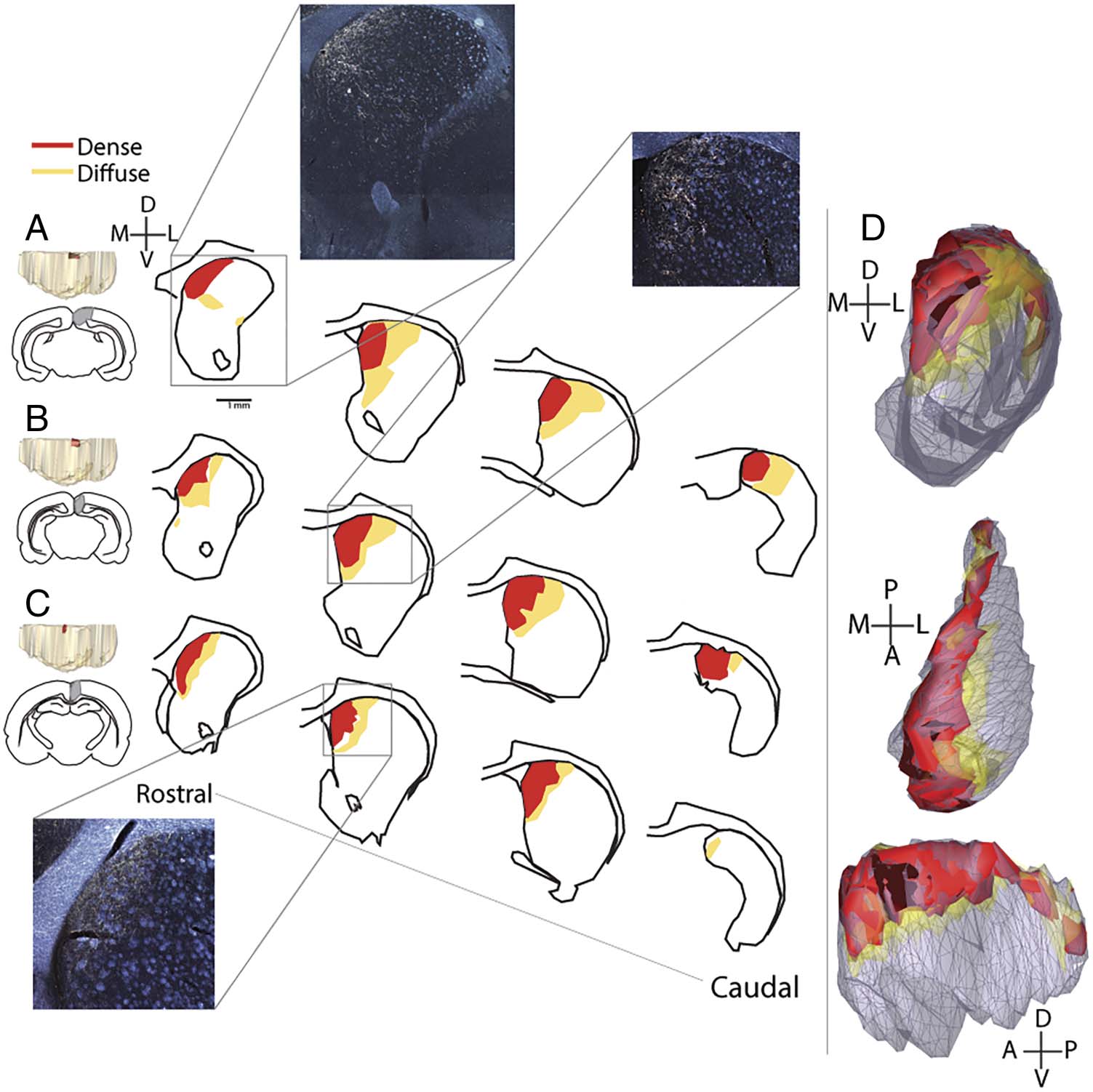
Figure 1. RSC-striatal terminal
fields. Images at far left show
3-D models and outlines for
the three injection sites (A–C).
The coordinates of the spread
of the injection sites in the
anterior/posterior direction are
A. −4.8 to −8.6 AP, B. −4.8
to −8.8 AP, and C. −5.0 to
−6.0 AP. Drawings show coronal
striatal sections with terminal
fields from rostral to caudal
striatum. Photomicrographs
show coronal darkfield images
of ipsilateral striatal terminal
fields. White arrows point
to dense terminal fields. (D)
Collective model of the striatal
projection zone of the RSC
from coronal (top), horizontal
(middle), and sagittal (bottom)
views, all cases combined. Scale
bar = 1 mm.
1098
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
0
9
6
1
9
1
3
6
6
8
/
/
j
o
c
n
_
a
_
0
1
6
9
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
& Faull, 1989). Therefore, we analyzed ipsilateral striatal pro-
jections, but there were similarly situated, less dense, contra-
lateral terminal fields as well. Cases were extremely similar in
their striatal projections and, thus, are described here to-
gether. Fibers could be visualized exiting the injection site
and traveling ventrally and rostrally toward the dorsal stria-
tum through the cingulum bundle and the white matter di-
rectly adjacent to it. These fibers formed longitudinal bands
of terminal fields that extended for nearly the entire rostral-
caudal distance of the ipsilateral caudoputamen. At the most
caudal levels of the caudoputamen, terminal fields were
present only dorsally. Rostrally, where the caudoputamen
widens in the medial-lateral dimension, terminal fields were
restricted to the dorsomedial component. They frequently
reached the dorsomedial boundary between the caudo-
putamen and the lateral ventricle (Figure 1). In all cases,
both dense and diffuse projections could be observed.
The dense projections were the most consistently dorso-
medial, whereas the lighter projections extended further
ventrally and laterally. However, even diffuse projections
did not extend into the nucleus accumbens.
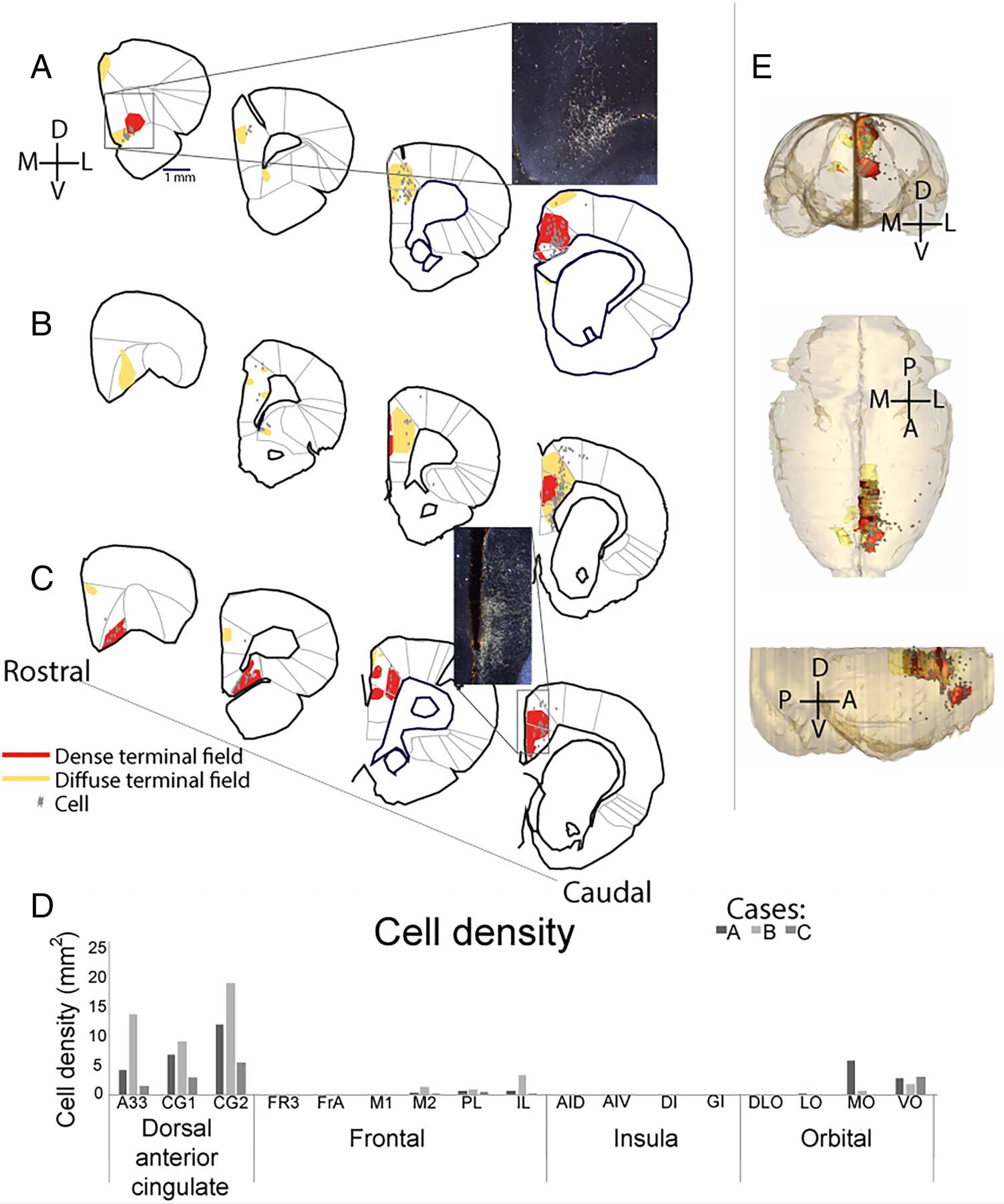
Next, we examined connections between RSC and differ-
ent regions within the frontal cortex (Figure 2). Although
there were some quantitative differences between the cases
(Figure 2D), qualitatively, we observed consistent bidirec-
tional connections primarily with the dorsal anterior cingu-
late areas (CG1, CG2, and A33) and a specific location
within the central-medial orbito-frontal cortex that sits at
the border between ventral orbital and MO. Much weaker,
but present, connections were observed with IL, PL, M2,
and M1. Virtually no connectivity was observed with lateral
orbito-frontal cortex and frontal insular regions. Thus,
although there is a consistent patch of connectivity in the
central-medial orbito-frontal cortex, the medial frontal
Figure 2. RSC-frontal
projections. Terminal fields and
labeled cells following injections
into the RSC, (A–C) Drawings of
frontal terminal fields (red =
dense terminal fields, yellow =
diffuse terminal fields; gray
dots = cells; all shown for each
injection site). (Each case is a
different injection site, matched
to Figure 1.) (D) Cell density
from each case in frontal
regions. Results are shown for
each case (A–C, as above). (E)
Collective model of the frontal
projections of the RSC from
coronal (top), horizontal
(middle), and sagittal (bottom)
views, all cases combined.
Scale bar = 1 mm.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
0
9
6
1
9
1
3
6
6
8
/
/
j
o
c
n
_
a
_
0
1
6
9
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Monko and Heilbronner
1099
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
0
9
6
1
9
1
3
6
6
8
/
/
j
o
c
n
_
a
_
0
1
6
9
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
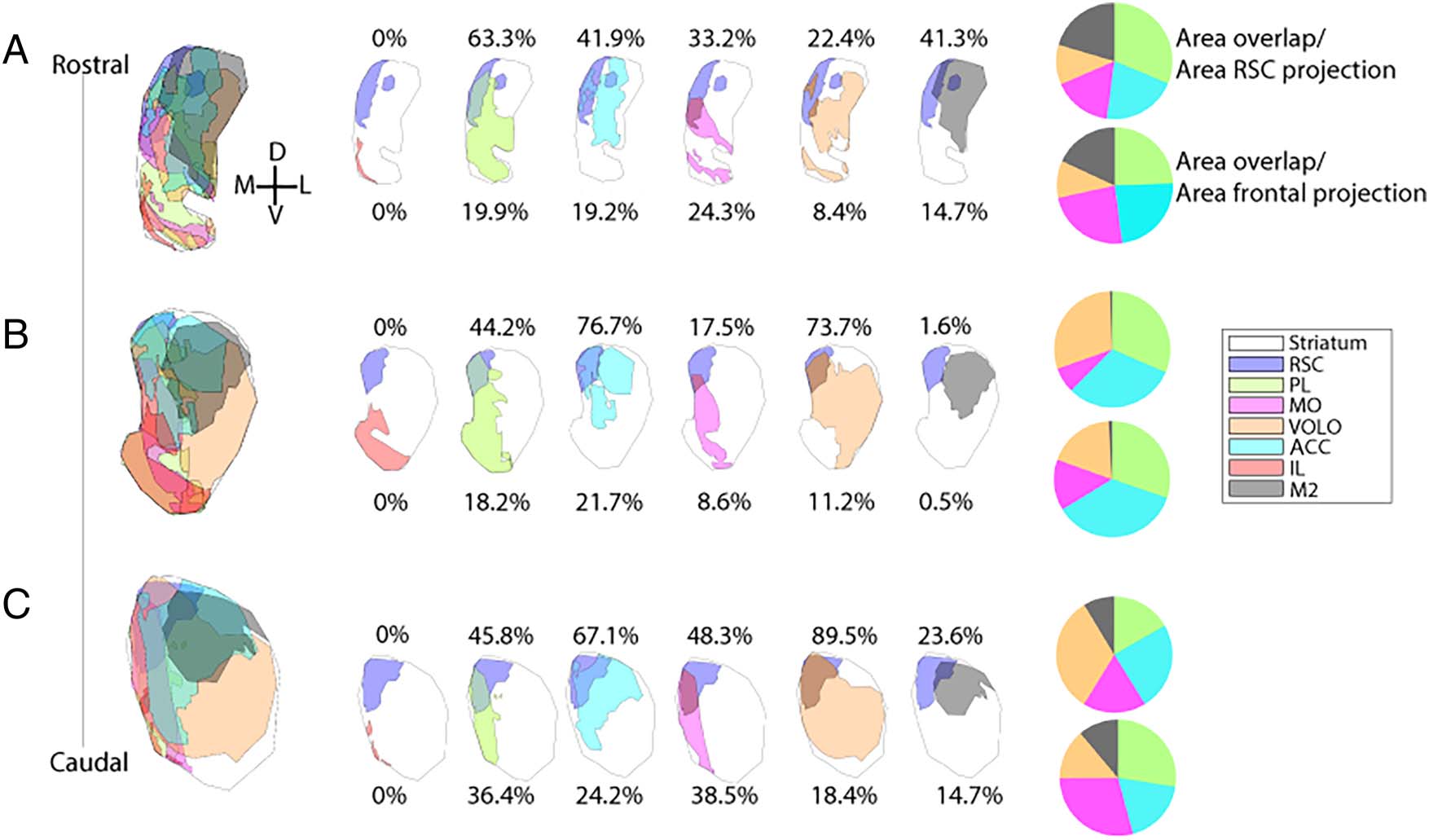
Figure 3. Cortico-striatal overlap. Analyses of overlap of the RSC projection zone in the striatum with select frontal area projection zones. Drawings
depict the RSC projection zone (purple) at AP +2.3 (A), AP +1.1 (B), and AP −0.1 (C) compared to IL (red), PL (yellow-green), CG (light blue), MO
(purple), VOLO (orange), and M2 (black) projection zones. Numbers and pie charts show the area of the RSC-striatal projection zone encapsulated
within the frontal area’s projection zone, divided by the area of the RSC projection (top) or the area of the frontal projection (bottom). AP =
anterior–posterior distance from bregma; IL = infralimbic cortex; PL = prelimbic cortex; CG = anterior cingulate cortex; MO = medial orbital cortex;
VOLO = ventral and lateral orbital cortex.
cortex has, overall, much stronger connections with the RSC
than the orbito-frontal cortex does. In addition, in both the
medial and orbito-frontal cortices, RSC connectivity is with
the central portions, rather than the ventral/dorsal or
medial/lateral edges.
We then analyzed the overlap between the striatal termi-
nal fields from the RSC and the striatal projection zones of
various frontal regions, as previously defined (Heilbronner
et al., 2016; Reep & Corwin, 1999; Figure 3). At all A-P levels
examined, the RSC does not occupy striatal territory iso-
lated from frontal projections. Instead, large portions of
the RSC projection zone overlap with terminal fields from
frontal cortical regions (at A-P +2.3, > 90%; at A-P +1.1 >
95%; at A-P −0.1, > 98%). At A-P +1.1, the database of frontal
terminal fields is greatest, and the projection zones from
these regions are maximally segregated, so we will consider
this topography in detail here (although note other levels
diagrammed in Figure 3). The greatest percentage of over-
lap is with central–medial prefrontal regions (prelimbic and
anterior cingulate cortices) and central and lateral orbito-
frontal regions. Smaller zones of overlap are observed be-
tween the RSC projection zones and those from M2 and
MO. At this level and all levels, there is no overlap between
the RSC projection zone and the infralimbic projection
zone. Finally, although the RSC-striatal projection is not
isolated from frontal-striatal projections, no frontal-striatal
projection zone is as uniquely confined to the dorsome-
dial caudoputamen as the RSC terminal fields are.
DISCUSSION
Here, we examined in detail the projections of the RSC to
frontal cortico-basal ganglia circuits. We found that the
RSC projects exclusively to the dorsomedial striatum,
which also receives dense projections from dorsal anterior
cingulate, ventral and lateral orbital areas, and prelim-
bic cortex. The RSC has bidirectional connections with
dorsal anterior cingulate regions, central-medial orbito-
frontal cortex, and, to a lesser extent, PL and IL. These re-
sults are in full agreement with prior anatomical work
in both rats and mice (Hintiryan et al., 2016; Hunnicutt
et al., 2016; Shibata & Naito, 2008; Shibata et al., 2004;
Cheatwood, Reep, & Corwin, 2003; Reep, Cheatwood, &
Corwin, 2003; van Groen & Wyss, 1992, 2003); here, our
goal was to synthesize frontal and striatal projections
from a functional perspective.
While RSC has historically been known for its role in
navigation and memory functions likely shared with the
medial temporal lobe, it also seems to play an important
role in reward-guided decision making, a process that is
most commonly associated with frontal cortico-basal ganglia
regions and circuits. The dorsomedial striatum and its frontal
cortical afferents are known to be involved with goal-
directed decision making rather than stimulus value or
procedural learning. For example, inhibition of orbito-
frontal-dorsomedial striatal neurons caused mice to be un-
able to shift to goal-directed action control (Gremel et al.,
1100
Journal of Cognitive Neuroscience
Volume 33, Number 6
2016). The placement of RSC within this circuitry calls for
similar manipulation studies focused on testing the role
of RSC-striatal neurons in goal-directed behavior.
The RSC has mainly been studied in the context of spatial
navigation and memory due in part to its strong anatomical
connectivity with medial temporal lobe structures (Mitchell,
Czajkowski, Zhang, Jeffery, & Nelson, 2018; Vann et al.,
2009). The cortico-basal ganglia loops of the medial tempo-
ral lobe are both distinct and overlapping with those of the
RSC. The hippocampus, for example, projects mainly to the
medial nucleus accumbens, positioning it quite differently
than the RSC projection (Groenewegen, Vermeulen-Van
der Zee, te Kortschot, & Witter, 1987). However, very ros-
trally in the caudoputamen, the hippocampus projects just
laterally to the lateral ventricle, similar to the rostral RSC-
striatal projection zone. Similarly, entorhinal and peririhinal
cortices primarily project to the nucleus accumbens, but
also dorsal and medial borders of the caudoputamen, over-
lapping with the RSC projection zone (McIntyre, Kelly, &
Staines, 1996; McGeorge & Faull, 1989). Taken together,
the bulk of the medial temporal lobe structures project
mainly to the nucleus accumbens, yet these structures do
also project to the dorsomedial caudoputamen in such a
way as to provide some overlap with the RSC projection
zone there, although this overlap is not consistent across
rostral versus caudal caudoputamen. Similarly, with the
exception of perirhinal cortex, medial temporal lobe
frontal projections are more strongly connected with ven-
tral prefrontal cortex, rather than dorsal (Insausti, Herrero,
& Witter, 1997; Verwer, Meijer, Van Uum, & Witter, 1997;
McIntyre et al., 1996; Jay, Glowinski, & Thierry, 1989).
RSC-frontal projections mostly parallel the striatal projec-
tion zone overlap. Cortico-basal ganglia loops have been
described as both parallel (Miyachi, 2009; Alexander,
DeLong, & Strick, 1986) and integrative (Groenewegen,
Voorn, & Scheel-Krüger, 2016; Draganski et al., 2008;
Haber et al., 2006). Furthermore, tract-tracing experiments
have found that connected cortical areas overlap in their
striatal terminal fields (Selemon & Goldman-Rakic, 1985),
creating a network with some unifying function (Selemon
& Goldman-Rakic, 1988). This concept has been extended
to human resting-state functional connectivity, in which
distributed, correlated activity defines such functional net-
works as the DMN, the dorsal attention network, and the
frontoparietal network (Choi, Yeo, & Buckner, 2012; Yeo
et al., 2011). Here, we find that, in rats, projections between
the RSC and the frontal lobe roughly, but do not precisely,
match the overlap in terminal fields in the striatum. CG, for
example, projects to the dorsomedial and dorsocentral
striatum, overlapping considerably with the RSC striatal-
projection zone, and also has strong connections with the
RSC. The IL, by contrast, does not overlap with the RSC-
striatal projection zone and has only very weak connections
with the RSC. However, the PL has only weak connections
with the RSC but projects strongly to the dorsomedial stria-
tum. One possibility is that this connection serves an integra-
tive function for decision-making processes at the level of the
basal ganglia. This integrative function might relate to the
RSC’s function in navigation, providing information about
location to the areas that are involved in decision making.
The human RSC is part of the larger structure of the
posteromedial cortex, which is the main hub of the DMN.
This network, which is more active during resting states than
cognitive tasks (Greicius, Krasnow, Reiss, & Menon, 2003),
also includes frontal and temporal regions. Mirroring the
RSC-striatal anatomical connections in rats, the DMN is func-
tionally connected with the medial caudate in humans (Choi
et al., 2012). The DMN in rodents, which includes the RSC
(Lu et al., 2012; Upadhyay et al., 2011), the central-medial
orbito-frontal cortex, and the dorsomedial frontal cortex,
seems to closely match the anatomical connectivity pattern
shown here. A new theory of the role of the DMN posits that,
in monkeys, much of the DMN participates in cognitive shift-
ing (Arsenault, Caspari, Vandenberghe, & Vanduffel, 2018).
Therefore, one possibility is that the RSC is involved in at-
tentional shifting in rodents as a function of its striatal ter-
ritory. As a hub of the DMN, RSC in humans is known to be
impaired in various psychiatric and neurological disorders
(Doucet et al., 2020; He et al., 2018; Satyshur, Layden,
Gowins, Buchanan, & Gollan, 2018; Martino et al., 2016;
Wu et al., 2016; Baker et al., 2014; Cowdrey, Filippini, Park,
Smith, & McCabe, 2014; Hafkemeijer, van der Grond, &
Rombouts, 2012; Anderson et al., 2011; Bluhm et al.,
2009). Understanding the biology of the DMN will require
a detailed, translational explication of the functional roles
of the RSC, not just in navigation and memory, but also
in decision making.
Acknowledgments
We thank Tanya Casta, Adriana Cushnie, Mark Grier, and Anish
Sethi for assistance with data collection. This work was supported
by the National Institute of Mental Health (R01118257), the
MnDrive Brain Conditions Initiative, and the Brain & Behavior
Research Foundation.
Reprint requests should be sent to Sarah Heilbronner, Depart-
ment of Neuroscience, University of Minnesota, 2-164 Jackson
Hall, 321 Church St SE, Minneapolis, MN 5545, or via e-mail:
heilb028@umn.edu.
Author Contributions
Megan E. Monko: Conceptualization; Data curation; Formal
analysis; Investigation; Project administration; Visualization;
Writing—Original draft; Writing—Review & editing. Sarah R.
Heilbronner: Conceptualization; Formal analysis; Funding
acquisition; Investigation; Methodology; Project administra-
tion; Supervision; Visualization; Writing—Original draft;
Writing—Review & editing.
Funding Information
National Institute of Mental Health (http://dx.doi.org/10
.13039/100000025), grant number: R01118257. Brain and
Monko and Heilbronner
1101
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
0
9
6
1
9
1
3
6
6
8
/
/
j
o
c
n
_
a
_
0
1
6
9
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Behavior Research Foundation (http://dx.doi.org/10
.13039/100000874), grant number: NARSAD Young
Investigator Award. MnDrive Brain Conditions Initiative.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed a
persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/W = .108, and W/W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/W = .102, and W/W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
REFERENCES
Alexander, G. E., DeLong, M. R., & Strick, P. L. (1986). Parallel
organization of functionally segregated circuits linking basal
ganglia and cortex. Annual Review of Neuroscience, 9,
357–381. DOI: https://doi.org/10.1146/annurev.ne.09.030186
.002041, PMID: 3085570
Anderson, J. S., Druzgal, T. J., Lopez-Larson, M., Jeong, E.-K.,
Desai, K., & Yurgelun-Todd, D. (2011). Network anticorrelations,
global regression, and phase-shifted soft tissue correction.
Human Brain Mapping, 32, 919–934. DOI: https://doi.org/10
.1002/hbm.21079, PMID: 20533557, PMCID: PMC3220164
Arsenault, J. T., Caspari, N., Vandenberghe, R., & Vanduffel, W.
(2018). Attention shifts recruit the monkey default mode
network. Journal of Neuroscience, 38, 1202–1217. DOI:
https://doi.org/10.1523/JNEUROSCI.1111-17.2017, PMID:
29263238, PMCID: PMC6596261
Auger, S. D., & Maguire, E. A. (2013). Assessing the mechanism
of response in the retrosplenial cortex of good and poor
navigators. Cortex, 49, 2904–2913. DOI: https://doi.org
/10.1016/j.cortex.2013.08.002, PMID: 24012136, PMCID:
PMC3878422
Auger, S. D., Mullally, S. L., & Maguire, E. A. (2012). Retrosplenial
cortex codes for permanent landmarks. PLoS One, 7, e43620.
DOI: https://doi.org/10.1371/journal.pone.0043620, PMID:
22912894, PMCID: PMC3422332
Baker, J. T., Holmes, A. J., Masters, G. A., Yeo, B. T. T., Krienen,
F., Buckner, R. L., et al. (2014). Disruption of cortical
association networks in schizophrenia and psychotic bipolar
disorder. JAMA Psychiatry, 71, 109–118. DOI: https://doi
.org/10.1001/jamapsychiatry.2013.3469, PMID: 24306091,
PMCID: PMC4435541
Balleine, B. W., Delgado, M. R., & Hikosaka, O. (2007). The
role of the dorsal striatum in reward and decision-making.
Journal of Neuroscience, 27, 8161–8165. DOI: https://doi
.org/10.1523/JNEUROSCI.1554-07.2007, PMID: 17670959,
PMCID: PMC6673072
Bluhm, R., Williamson, P., Lanius, R., Théberge, J., Densmore,
M., Bartha, R., et al. (2009). Resting state default-mode
network connectivity in early depression using a seed region-
of-interest analysis: Decreased connectivity with caudate
nucleus. Psychiatry and Clinical Neurosciences, 63, 754–761.
DOI: https://doi.org/10.1111/j.1440-1819.2009.02030.x,
PMID: 20021629
Burton, A. C., Nakamura, K., & Roesch, M. R. (2015). From
ventral-medial to dorsal-lateral striatum: Neural correlates of
reward-guided decision-making. Neurobiology of Learning
and Memory, 117, 51–59. DOI: https://doi.org/10.1016/j.nlm
.2014.05.003, PMID: 24858182, PMCID: PMC4240773
Cheatwood, J. L., Reep, R. L., & Corwin, J. V. (2003). The
associative striatum: Cortical and thalamic projections to the
dorsocentral striatum in rats. Brain Research, 968, 1–14.
DOI: https://doi.org/10.1016/S0006-8993(02)04212-9, PMID:
12644259
Cho, J., & Sharp, P. E. (2001). Head direction, place, and
movement correlates for cells in the rat retrosplenial cortex.
Behavioral Neuroscience, 115, 3–25. DOI: https://doi.org
/10.1037/0735-7044.115.1.3, PMID: 11256450
Choi, E. Y., Yeo, B. T. T., & Buckner, R. L. (2012). The
organization of the human striatum estimated by intrinsic
functional connectivity. Journal of Neurophysiology, 108,
2242–2263. DOI: https://doi.org/10.1152/jn.00270.2012,
PMID: 22832566, PMCID: PMC3545026
Cooper, B. G., & Mizumori, S. J. (1999). Retrosplenial cortex
inactivation selectively impairs navigation in darkness.
NeuroReport, 10, 625–630. DOI: https://doi.org/10.1097
/00001756-199902250-00033, PMID: 10208601
Cowdrey, F. A., Filippini, N., Park, R. J., Smith, S. M., & McCabe, C.
(2014). Increased resting state functional connectivity in
the default mode network in recovered anorexia nervosa.
Human Brain Mapping, 35, 483–491. DOI: https://doi
.org/10.1002/hbm.22202, PMID: 23033154, PMCID:
PMC6869597
Doucet, G. E., Janiri, D., Howard, R., O’Brien, M., Andrews-
Hanna, J. R., & Frangou, S. (2020). Transdiagnostic and
disease-specific abnormalities in the default-mode network
hubs in psychiatric disorders: A meta-analysis of resting-state
functional imaging studies. European Psychiatry, 63, e57.
DOI: https://doi.org/10.1192/j.eurpsy.2020.57, PMID:
32466812, PMCID: PMC7355168
Draganski, B., Kherif, F., Klöppel, S., Cook, P. A., Alexander, D. C.,
Parker, G. J. M., et al. (2008). Evidence for segregated and
integrative connectivity patterns in the human basal ganglia.
Journal of Neuroscience, 28, 7143–7152. DOI: https://doi
.org/10.1523/JNEUROSCI.1486-08.2008, PMID: 18614684,
PMCID: PMC6670486
Fitoussi, A., Renault, P., Le Moine, C., Coutureau, E., Cador, M.,
& Dellu-Hagedorn, F. (2018). Inter-individual differences
in decision-making, flexible and goal-directed behaviors:
Novel insights within the prefronto-striatal networks. Brain
Structure & Function, 223, 897–912. DOI: https://doi.org
/10.1007/s00429-017-1530-z, PMID: 29026986
Greicius, M. D., Krasnow, B., Reiss, A. L., & Menon, V. (2003).
Functional connectivity in the resting brain: A network
analysis of the default mode hypothesis. Proceedings of the
National Academy of Sciences, U.S.A., 100, 253–258. DOI:
https://doi.org/10.1073/pnas.0135058100, PMID: 12506194,
PMCID: PMC140943
Gremel, C. M., Chancey, J. H., Atwood, B. K., Luo, G., Neve, R.,
Ramakrishnan, C., et al. (2016). Endocannabinoid modulation
of orbitostriatal circuits gates habit formation. Neuron, 90,
1312–1324. DOI: https://doi.org/10.1016/j.neuron.2016.04.043,
PMID: 27238866, PMCID: PMC4911264
Groenewegen, H. J., Vermeulen-Van der Zee, E., te Kortschot, A.,
& Witter, M. P. (1987). Organization of the projections from
the subiculum to the ventral striatum in the rat. A study using
anterograde transport of Phaseolus vulgaris leucoagglutinin.
Neuroscience, 23, 103–120. DOI: https://doi.org/10.1016
/0306-4522(87)90275-2, PMID: 3683859
1102
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
0
9
6
1
9
1
3
6
6
8
/
/
j
o
c
n
_
a
_
0
1
6
9
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Groenewegen, H. J., Voorn, P., & Scheel-Krüger, J. (2016).
Limbic-basal ganglia circuits parallel and integrative aspects.
In J.-J. Soghomonian (Ed.), The basal ganglia: Innovations
in cognitive neuroscience (pp. 11–45). Cham: Springer.
DOI: https://doi.org/10.1007/978-3-319-42743-0_2
Haber, S. N., & Behrens, T. E. J. (2014). The neural network
underlying incentive-based learning: Implications for interpreting
circuit disruptions in psychiatric disorders. Neuron, 83,
1019–1039. DOI: https://doi.org/10.1016/j.neuron.2014
.08.031, PMID: 25189208, PMCID: PMC4255982
Haber, S. N., Kim, K.-S., Mailly, P., & Calzavara, R. (2006).
Reward-related cortical inputs define a large striatal region in
primates that interface with associative cortical connections,
providing a substrate for incentive-based learning. Journal of
Neuroscience, 26, 8368–8376. DOI: https://doi.org/10.1523
/JNEUROSCI.0271-06.2006, PMID: 16899732, PMCID:
PMC6673798
Hafkemeijer, A., van der Grond, J., & Rombouts, S. A. R. B.
(2012). Imaging the default mode network in aging and
dementia. Biochimica et Biophysica Acta, 1822, 431–441.
DOI: https://doi.org/10.1016/j.bbadis.2011.07.008, PMID:
21807094
Harker, K. T., & Whishaw, I. Q. (2004). Impaired place navigation
in place and matching-to-place swimming pool tasks follows
both retrosplenial cortex lesions and cingulum bundle lesions
in rats. Hippocampus, 14, 224–231. DOI: https://doi.org/10
.1002/hipo.10159, PMID: 15098727
Hassani, O. K., Cromwell, H. C., & Schultz, W. (2001). Influence
of expectation of different rewards on behavior-related
neuronal activity in the striatum. Journal of Neurophysiology,
85, 2477–2489. DOI: https://doi.org/10.1152/jn.2001.85.6.2477,
PMID: 11387394
Hattori, R., Danskin, B., Babic, Z., Mlynaryk, N., & Komiyama, T.
(2019). Area-specificity and plasticity of history-dependent
value coding during learning. Cell, 177, 1858–1872. DOI:
https://doi.org/10.1016/j.cell.2019.04.027, PMID: 31080067,
PMCID: PMC6663310
Haynes, W. I. A., & Haber, S. N. (2013). The organization
of prefrontal-subthalamic inputs in primates provides an
anatomical substrate for both functional specificity and
integration: Implications for basal ganglia models and deep
brain stimulation. Journal of Neuroscience, 33, 4804–4814.
DOI: https://doi.org/10.1523/JNEUROSCI.4674-12.2013,
PMID: 23486951, PMCID: PMC3755746
He, C., Chen, Y., Jian, T., Chen, H., Guo, X., Wang, J., et al.
(2018). Dynamic functional connectivity analysis reveals
decreased variability of the default-mode network in
developing autistic brain. Autism Research, 11, 1479–1493.
DOI: https://doi.org/10.1002/aur.2020, PMID: 30270547
Heilbronner, S. R., & Haber, S. N. (2014). Frontal cortical
and subcortical projections provide a basis for segmenting
the cingulum bundle: Implications for neuroimaging
and psychiatric disorders. Journal of Neuroscience, 34,
10041–10054. DOI: https://doi.org/10.1523/JNEUROSCI
.5459-13.2014, PMID: 25057206, PMCID: PMC4107396
Heilbronner, S. R., Meyer, M. A. A., Choi, E. Y., & Haber, S. N.
(2018). How do cortico-striatal projections impact on
downstream pallidal circuitry? Brain Structure & Function,
223, 2809–2821. DOI: https://doi.org/10.1007/s00429-018
-1662-9, PMID: 29654360
Heilbronner, S. R., Rodriguez-Romaguera, J., Quirk, G. J.,
Groenewegen, H. J., & Haber, S. N. (2016). Circuit-based
corticostriatal homologies between rat and primate.
Biological Psychiatry, 80, 509–521. DOI: https://doi.org
/10.1016/j.biopsych.2016.05.012, PMID: 27450032, PMCID:
PMC5438202
Hintiryan, H., Foster, N. N., Bowman, I., Bay, M., Song, M. Y.,
Gou, L., et al. (2016). The mouse cortico-striatal projectome.
Nature Neuroscience, 19, 1100–1114. DOI: https://doi.org
/10.1038/nn.4332, PMID: 27322419, PMCID: PMC5564682
Hunnicutt, B. J., Jongbloets, B. C., Birdsong, W. T., Gertz, K. J.,
Zhong, H., & Mao, T. (2016). A comprehensive excitatory
input map of the striatum reveals novel functional organization.
eLife, 5, e19103. DOI: https://doi.org/10.7554/eLife.19103,
PMID: 27892854, PMCID: PMC5207773
Insausti, R., Herrero, M. T., & Witter, M. P. (1997). Entorhinal
cortex of the rat: Cytoarchitectonic subdivisions and the
origin and distribution of cortical efferents. Hippocampus,
7, 146–183. DOI: https://doi.org/10.1002/(SICI)1098-1063
(1997)7:2<146::AID-HIPO4>3.0.CO;2-L, PMID: 9136047
Jay, T. M., Glowinski, J., & Thierry, A. M. (1989). Selectivity of
the hippocampal projection to the prelimbic area of the
prefrontal cortex in the rat. Brain Research, 505, 337–340.
DOI: https://doi.org/10.1016/0006-8993(89)91464-9
Kaboodvand, N., Bäckman, L., Nyberg, L., & Salami, A. (2018).
The retrosplenial cortex: A memory gateway between the
cortical default mode network and the medial temporal
lobe. Human Brain Mapping, 39, 2020–2034. DOI: https://
doi.org/10.1002/hbm.23983, PMID: 29363256, PMCID:
PMC6866613
Keene, C. S., & Bucci, D. J. (2008). Neurotoxic lesions of
retrosplenial cortex disrupt signaled and unsignaled
contextual fear conditioning. Behavioral Neuroscience,
122, 1070–1077. DOI: https://doi.org/10.1037/a0012895,
PMID: 18823164
Keene, C. S., & Bucci, D. J. (2009). Damage to the retrosplenial
cortex produces specific impairments in spatial working
memory. Neurobiology of Learning and Memory, 91,
408–414. DOI: https://doi.org/10.1016/j.nlm.2008.10.009,
PMID: 19026755
Kimchi, E. Y., & Laubach, M. (2009). Dynamic encoding
of action selection by the medial striatum. Journal of
Neuroscience, 29, 3148–3159. DOI: https://doi.org/10.1523
/JNEUROSCI.5206-08.2009, PMID: 19279252, PMCID:
PMC3415331
Kremer, J. R., Mastronarde, D. N., & McIntosh, J. R. (1996).
Computer visualization of three-dimensional image data
using IMOD. Journal of Structural Biology, 116, 71–76. DOI:
https://doi.org/10.1006/jsbi.1996.0013, PMID: 8742726
Li, Y., He, Y., Chen, M., Pu, Z., Chen, L., Li, P., et al. (2016).
Optogenetic activation of adenosine A2A receptor signaling
in the dorsomedial striatopallidal neurons suppresses
goal-directed behavior. Neuropsychopharmacology, 41,
1003–1013. DOI: https://doi.org/10.1038/npp.2015.227,
PMID: 26216520, PMCID: PMC4748425
Lu, H., Zou, Q., Gu, H., Raichle, M. E., Stein, E. A., & Yang, Y.
(2012). Rat brains also have a default mode network.
Proceedings of the National Academy of Sciences, U.S.A.,
109, 3979–3984. DOI: https://doi.org/10.1073/pnas
.1200506109, PMID: 22355129, PMCID: PMC3309754
Maguire, E. A. (2001). The retrosplenial contribution to
human navigation: A review of lesion and neuroimaging
findings. Scandinavian Journal of Psychology, 42, 225–238.
DOI: https://doi.org/10.1111/1467-9450.00233, PMID:
11501737
Mailly, P., Aliane, V., Groenewegen, H. J., Haber, S. N., &
Deniau, J.-M. (2013). The rat prefrontostriatal system
analyzed in 3D: Evidence for multiple interacting functional
units. Journal of Neuroscience, 33, 5718–5727. DOI: https://
doi.org/10.1523/JNEUROSCI.5248-12.2013, PMID: 23536085,
PMCID: PMC6705071
Mantini, D., Gerits, A., Nelissen, K., Durand, J.-B., Joly, O.,
Simone, L., et al. (2011). Default mode of brain function in
monkeys. Journal of Neuroscience, 31, 12954–12962. DOI:
https://doi.org/10.1523/JNEUROSCI.2318-11.2011, PMID:
21900574, PMCID: PMC3686636
Monko and Heilbronner
1103
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
0
9
6
1
9
1
3
6
6
8
/
/
j
o
c
n
_
a
_
0
1
6
9
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Martino, M., Magioncalda, P., Huang, Z., Conio, B., Piaggio, N.,
Duncan, N. W., et al. (2016). Contrasting variability patterns
in the default mode and sensorimotor networks balance in
bipolar depression and mania. Proceedings of National
Academy of Sciences, U.S.A., 113, 4824–4829. DOI: https://
doi.org/10.1073/pnas.1517558113, PMID: 27071087, PMCID:
PMC4855585
Maviel, T., Durkin, T. P., Menzaghi, F., & Bontempi, B. (2004).
Sites of neocortical reorganization critical for remote spatial
memory. Science, 305, 96–99. DOI: https://doi.org/10.1126
/science.1098180, PMID: 15232109
McGeorge, A. J., & Faull, R. L. (1989). The organization of the
projection from the cerebral cortex to the striatum in the rat.
Neuroscience, 29, 503–537. DOI: https://doi.org/10.1016
/0306-4522(89)90128-0, PMID: 2472578
Reep, R. L., & Corwin, J. V. (1999). Topographic organization of
the striatal and thalamic connections of rat medial agranular
cortex. Brain Research, 841, 43–52. DOI: https://doi.org/10
.1016/S0006-8993(99)01779-5, PMID: 10546986
Rothwell, P. E., Hayton, S. J., Sun, G. L., Fuccillo, M. V., Lim, B. K.,
& Malenka, R. C. (2015). Input- and output-specific regulation
of serial order performance by corticostriatal circuits. Neuron,
88, 345–356. DOI: https://doi.org/10.1016/j.neuron.2015
.09.035, PMID: 26494279, PMCID: PMC4618801
Satyshur, M. D., Layden, E. A., Gowins, J. R., Buchanan, A., &
Gollan, J. K. (2018). Functional connectivity of reflective and
brooding rumination in depressed and healthy women.
Cognitive, Affective, & Behavioral Neuroscience, 18,
884–901. DOI: https://doi.org/10.3758/s13415-018-0611-7,
PMID: 29949111
McIntyre, D. C., Kelly, M. E., & Staines, W. A. (1996). Efferent
Selemon, L. D., & Goldman-Rakic, P. S. (1985). Longitudinal
projections of the anterior perirhinal cortex in the rat.
Journal of Comparative Neurology, 369, 302–318. DOI:
https://doi.org/10.1002/(SICI)1096-9861(19960527)
369:2<302::AID-CNE10>3.0.CO;2-J, PMID: 8727002
Mitchell, A. S., Czajkowski, R., Zhang, N., Jeffery, K., & Nelson,
A. J. D. (2018). Retrosplenial cortex and its role in spatial
cognition. Brain and Neuroscience Advances, 2,
2398212818757098. DOI: https://doi.org/10.1177
/2398212818757098, PMID: 30221204, PMCID: PMC6095108
Miyachi, S. (2009). Cortico-basal ganglia circuits—Parallel closed
loops and convergent/divergent connections. Brain and
Nerve, 61, 351–359. DOI: https://doi.org/10.11477/mf
.1416100459, PMID: 19378804
Nelson, A. J. D., Hindley, E. L., Haddon, J. E., Vann, S. D., &
Aggleton, J. P. (2014). A novel role for the rat retrosplenial
cortex in cognitive control. Learning & Memory, 21, 90–97.
DOI: https://doi.org/10.1101/lm.032136.113, PMID: 24434870,
PMCID: PMC3895227
O’Doherty, J., Dayan, P., Schultz, J., Deichmann, R., Friston, K.,
& Dolan, R. J. (2004). Dissociable roles of ventral and dorsal
striatum in instrumental conditioning. Science, 304, 452–454.
DOI: https://doi.org/10.1126/science.1094285, PMID: 15087550
O’Hare, J. K., Ade, K. K., Sukharnikova, T., Van Hooser, S. D.,
Palmeri, M. L., Yin, H. H., et al. (2016). Pathway-specific
striatal substrates for habitual behavior. Neuron, 89, 472–479.
DOI: https://doi.org/10.1016/j.neuron.2015.12.032, PMID:
26804995, PMCID: PMC4887103
Parron, C., & Save, E. (2004). Evidence for entorhinal and
parietal cortices involvement in path integration in the rat.
Experimental Brain Research, 159, 349–359. DOI: https://
doi.org/10.1007/s00221-004-1960-8, PMID: 15526193
Paxinos, G., & Watson, C. (2014). The rat brain in stereotaxic
coordinates (7th ed.). San Diego: Elsevier.
Pothuizen, H. H. J., Aggleton, J. P., & Vann, S. D. (2008). Do rats
with retrosplenial cortex lesions lack direction? European
Journal of Neuroscience, 28, 2486–2498. DOI: https://doi
.org/10.1111/j.1460-9568.2008.06550.x, PMID: 19032585
Powell, A. L., Nelson, A. J. D., Hindley, E., Davies, M., Aggleton,
J. P., & Vann, S. D. (2017). The rat retrosplenial cortex as
a link for frontal functions: A lesion analysis. Behavioural
Brain Research, 335, 88–102. DOI: https://doi.org/10.1016
/j.bbr.2017.08.010, PMID: 28797600, PMCID: PMC5597037
topography and interdigitation of corticostriatal projections
in the rhesus monkey. Journal of Neuroscience, 5, 776–794.
DOI: https://doi.org/10.1523/JNEUROSCI.05-03-00776.1985,
PMID: 2983048, PMCID: PMC6565017
Selemon, L. D., & Goldman-Rakic, P. S. (1988). Common
cortical and subcortical targets of the dorsolateral prefrontal
and posterior parietal cortices in the rhesus monkey:
Evidence for a distributed neural network subserving spatially
guided behavior. Journal of Neuroscience, 8, 4049–4068.
DOI: https://doi.org/10.1523/JNEUROSCI.08-11-04049.1988,
PMID: 2846794, PMCID: PMC6569486
Shibata, H., Kondo, S., & Naito, J. (2004). Organization of
retrosplenial cortical projections to the anterior cingulate,
motor, and prefrontal cortices in the rat. Neuroscience
Research, 49, 1–11. DOI: https://doi.org/10.1016/j.neures
.2004.01.005, PMID: 15099698
Shibata, H., & Naito, J. (2008). Organization of anterior
cingulate and frontal cortical projections to the retrosplenial
cortex in the rat. Journal of Comparative Neurology, 506,
30–45. DOI: https://doi.org/10.1002/cne.21523, PMID:
17990270
Shulman, G. L., Fiez, J. A., Corbetta, M., Buckner, R. L., Miezin,
F. M., Raichle, M. E., et al. (1997). Common blood flow
changes across visual tasks: II. Decreases in cerebral cortex.
Journal of Cognitive Neuroscience, 9, 648–663. DOI: https://
doi.org/10.1162/jocn.1997.9.5.648, PMID: 23965122
Sleezer, B. J., Castagno, M. D., & Hayden, B. Y. (2016). Rule
encoding in orbitofrontal cortex and striatum guides
selection. Journal of Neuroscience, 36, 11223–11237. DOI:
https://doi.org/10.1523/JNEUROSCI.1766-16.2016, PMID:
27807165, PMCID: PMC5148240
Sugar, J., Witter, M. P., van Strien, N. M., & Cappaert, N. L. M.
(2011). The retrosplenial cortex: Intrinsic connectivity and
connections with the (para)hippocampal region in the rat.
An interactive connectome. Frontiers in Neuroinformatics,
5, 7. DOI: https://doi.org/10.3389/fninf.2011.00007, PMID:
21847380, PMCID: PMC3147162
Sutherland, R. J., Wishaw, I. Q., & Kolb, B. (1988). Contributions
of cingulate cortex to two forms of spatial learning and
memory. Journal of Neuroscience, 8, 1863–1872. DOI:
https://doi.org/10.1523/JNEUROSCI.08-06-01863.1988, PMID:
3385478, PMCID: PMC6569344
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J.,
Tabuchi, E., Furusawa, A. A., Hori, E., Umeno, K., Ono, T., &
Gusnard, D. A., & Shulman, G. L. (2001). A default mode of
brain function. Proceedings of the National Academy of
Sciences, U.S.A., 98, 676–682. DOI: https://doi.org/10.1073
/pnas.98.2.676, PMID: 11209064, PMCID: PMC14647
Reep, R. L., Cheatwood, J. L., & Corwin, J. V. (2003). The
associative striatum: Organization of cortical projections to
the dorsocentral striatum in rats. Journal of Comparative
Neurology, 467, 271–292. DOI: https://doi.org/10.1002/cne
.10868, PMID: 14608594
Nishijo, H. (2005). Neural correlates to action and rewards in
the rat posterior cingulate cortex. NeuroReport, 16, 949–953.
DOI: https://doi.org/10.1097/00001756-200506210-00014,
PMID: 15931067
Upadhyay, J., Baker, S. J., Chandran, P., Miller, L., Lee, Y., Marek,
G. J., et al. (2011). Default-mode-like network activation in
awake rodents. PLoS One, 6, e27839. DOI: https://doi.org
/10.1371/journal.pone.0027839, PMID: 22125628, PMCID:
PMC3220684
1104
Journal of Cognitive Neuroscience
Volume 33, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
0
9
6
1
9
1
3
6
6
8
/
/
j
o
c
n
_
a
_
0
1
6
9
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Valenstein, E., Bowers, D., Verfaellie, M., Heilman, K. M., Day,
A., & Watson, R. T. (1987). Retrosplenial amnesia. Brain, 110,
1631–1646. DOI: https://doi.org/10.1093/brain/110.6.1631,
PMID: 3427404
van Groen, T., & Wyss, J. M. (1990). Connections of the
retrosplenial granular a cortex in the rat. Journal of
Comparative Neurology, 300, 593–606. DOI: https://doi
.org/10.1002/cne.903000412, PMID: 2273095
van Groen, T., & Wyss, J. M. (1992). Connections of the
retrosplenial dysgranular cortex in the rat. Journal of
Comparative Neurology, 315, 200–216. DOI: https://doi
.org/10.1002/cne.903150207, PMID: 1545009
van Groen, T., & Wyss, J. M. (2003). Connections of the
retrosplenial granular b cortex in the rat. Journal of
Comparative Neurology, 463, 249–263. DOI: https://doi
.org/10.1002/cne.10757, PMID: 12820159
Vann, S. D., & Aggleton, J. P. (2002). Extensive cytotoxic lesions
of the rat retrosplenial cortex reveal consistent deficits
on tasks that tax allocentric spatial memory. Behavioral
Neuroscience, 116, 85–94. DOI: https://doi.org/10.1037
/0735-7044.116.1.85, PMID: 11895186
Vann, S. D., & Aggleton, J. P. (2004). Testing the importance of
the retrosplenial guidance system: Effects of different sized
retrosplenial cortex lesions on heading direction and spatial
working memory. Behavioural Brain Research, 155, 97–108.
DOI: https://doi.org/10.1016/j.bbr.2004.04.005, PMID:
15325783
Vann, S. D., Aggleton, J. P., & Maguire, E. A. (2009). What does
the retrosplenial cortex do? Nature Reviews Neuroscience,
10, 792–802. DOI: https://doi.org/10.1038/nrn2733, PMID:
19812579
Vedder, L. C., Miller, A. M. P., Harrison, M. B., & Smith, D. M.
(2017). Retrosplenial cortical neurons encode navigational
cues, trajectories and reward locations during goal directed
navigation. Cerebral Cortex, 27, 3713–3723. DOI: https://
doi.org/10.1093/cercor/bhw192, PMID: 27473323, PMCID:
PMC6059095
Verwer, R. W. H., Meijer, R. J., Van Uum, H. F. M., & Witter, M. P.
(1997). Collateral projections from the rat hippocampal
formation to the lateral and medial prefrontal cortex.
Hippocampus, 7, 397–402. DOI: https://doi.org/10.1002
/(SICI)1098-1063(1997)7:4<397::AID-HIPO5>3.0.CO;2-G,
PMID: 9287079
Voorn, P., Vanderschuren, L. J. M. J., Groenewegen, H. J.,
Robbins, T. W., & Pennartz, C. M. A. (2004). Putting a spin
on the dorsal–ventral divide of the striatum. Trends in
Neurosciences, 27, 468–474. DOI: https://doi.org/10.1016
/j.tins.2004.06.006, PMID: 15271494
Wisner, K., Odintsov, B., Brozoski, D., & Brozoski, T. J. (2016).
Ratat1: A digital rat brain stereotaxic atlas derived from
high-resolution MRI images scanned in three dimensions.
Frontiers in Systems Neuroscience, 10, 64. DOI: https://
doi.org/10.3389/fnsys.2016.00064, PMID: 27540358, PMCID:
PMC4973504
Wu, H., Sun, H., Xu, J., Wu, Y., Wang, C., Xiao, J., et al. (2016).
Changed hub and corresponding functional connectivity
of subgenual anterior cingulate cortex in major depressive
disorder. Frontiers in Neuroanatomy, 10, 120. DOI: https://
doi.org/10.3389/fnana.2016.00120, PMID: 28018183, PMCID:
PMC5159433
Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R.,
Lashkari, D., Hollinshead, M., et al. (2011). The organization
of the human cerebral cortex estimated by intrinsic
functional connectivity. Journal of Neurophysiology, 106,
1125–1165. DOI: https://doi.org/10.1152/jn.00338.2011,
PMID: 21653723, PMCID: PMC3174820
Yin, H. H., & Knowlton, B. J. (2004). Contributions of striatal
subregions to place and response learning. Learning &
Memory, 11, 459–463. DOI: https://doi.org/10.1101/lm
.81004, PMID: 15286184, PMCID: PMC498333
Yin, H. H., Knowlton, B. J., & Balleine, B. W. (2006). Inactivation
of dorsolateral striatum enhances sensitivity to changes in the
action-outcome contingency in instrumental conditioning.
Behavioural Brain Research, 166, 189–196. DOI: https://
doi.org/10.1016/j.bbr.2005.07.012, PMID: 16153716
Yin, H. H., Ostlund, S. B., Knowlton, B. J., & Balleine, B. W.
(2005). The role of the dorsomedial striatum in instrumental
conditioning. European Journal of Neuroscience, 22,
513–523. DOI: https://doi.org/10.1111/j.1460-9568.2005
.04218.x, PMID: 16045504
Monko and Heilbronner
1105
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
3
3
6
1
0
9
6
1
9
1
3
6
6
8
/
/
j
o
c
n
_
a
_
0
1
6
9
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3