RESEARCH ARTICLE
Phonological and Semantic Specialization in
9- to 10-Year-Old Children During
Auditory Word Processing
Jin Wang1,2
, Brianna L. Yamasaki1,3
, and James R. Booth1
1Department of Psychology and Human Development, Vanderbilt University, Nashville, TN
2Harvard Graduate School of Education, Harvard University, Cambridge, MA
3Department of Psychology, Emory University, Atlanta, GA
Keywords: phonology, semantics, language, specialization, development, children
ABSTRACT
One of the core features of brain maturation is functional specialization. Previous research
has found that 7- to 8-year-old children start to specialize in both the temporal and frontal
lobes. However, as children continue to develop their phonological and semantic skills rapidly
until approximately 10 years old, it remained unclear whether any changes in specialization
later in childhood would be detected. Thus, the goal of the current study was to examine
phonological and semantic specialization in 9- to 10-year-old children during auditory word
processing. Sixty-one children were included in the analysis. They were asked to perform a
sound judgment task and a meaning judgment task, each with both hard and easy conditions
to examine parametric effects. Consistent with previous results from 7- to 8-year-old children,
direct task comparisons revealed language specialization in both the temporal and frontal
lobes in 9- to 10-year-old children. Specifically, the left dorsal inferior frontal gyrus showed
greater activation for the sound than the meaning task whereas the left middle temporal gyrus
showed greater activation for the meaning than the sound task. Interestingly, in contrast to the
previously reported finding that 7- to 8-year-old children primarily engage a general control
region during the harder condition for both tasks, we showed that 9- to 10-year-old children
recruited language-specific regions to process the more difficult task conditions. Specifically,
the left superior temporal gyrus showed greater activation for the phonological parametric
manipulation whereas the left ventral inferior frontal gyrus showed greater activation for the
semantic parametric manipulation.
INTRODUCTION
Effective language comprehension and production skills are crucial for success in daily life.
According to the interactive specialization theory (Johnson, 2011), developing complex
cognitive skills, such as those associated with language use, involves a prolonged process
of neural specialization. That is, it is hypothesized that children move through a process during
which cortical regions become more functionally tuned, or more responsive to their preferred
task or contexts than other tasks or contexts, with a region’s “preferred task” referring to a task
(or tasks) that requires a process that a cortical region is selectively sensitive to in the mature
brain. While there has only been limited support for this theory within the domain of language,
there has been evidence for this process of neural specialization within other cognitive
a n o p e n a c c e s s
j o u r n a l
Citation: Wang, J., Yamasaki, B. L., &
Booth, J. R. (2023). Phonological and
semantic specialization in 9- to 10-year-
old children during auditory word
processing. Neurobiology of Language,
4(2), 297–317. https://doi.org/10.1162
/nol_a_00099
DOI:
https://doi.org/10.1162/nol_a_00099
Supporting Information:
https://doi.org/10.1162/nol_a_00099
Received: 20 April 2022
Accepted: 29 December 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Jin Wang
jin_wang@gse.harvard.edu
Handling Editor:
Steven Small
Copyright: © 2023
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
Specialization:
A process where brain regions
become more responsive to their
preferred task than other tasks.
Semantic:
Refers to the meaning structures in
language.
Direct task comparison:
The contrast of brain activation
between similar tasks that differ
primarily in a process of interest.
Phonology:
Refers to the sound structures in
language.
Double dissociation:
Occurs when brain region A
activates more for Task 1 than Task 2,
whereas region B activates more for
Task 2 than Task 1.
Neurobiology of Language
domains such as face processing (e.g., Aylward et al., 2005; Gathers et al., 2004), social
cognition (e.g., Carter & Pelphrey, 2006), reading (e.g., Schlaggar & McCandliss, 2007), and
cognitive control (e.g., Durston et al., 2006). Furthermore, it has been demonstrated that
children with developmental disorders affecting these cognitive processes tend to present with
atypical patterns of brain specialization (Johnson, 2011). Understanding the development of
language-related neural specialization in typically developing children will not only add to
this growing body of research providing empirical support for the interactive specialization
theory but will also inform our understanding of the maturational trajectory of the neural
systems that support language and will provide a foundation on which to better understand
what may be different in children with developmental language disorders.
Successful language use involves the coordination of many different cognitive systems, two
of which are the phonological, or sound processing, and semantic, or meaning processing,
systems. Decades of theoretical, and supporting empirical work, has identified a unique net-
work of regions that support each of these component skills. More specifically, the left superior
temporal gyrus (STG), supramarginal gyrus (SMG), inferior parietal lobule (IPL), and posterior
dorsal inferior frontal gyrus (dIFG) have been shown to support phonological processing.
Whereas, the left middle temporal gyrus (MTG), angular gyrus (AG), anterior fusiform gyrus
(FG), and anterior ventral inferior frontal gyrus (vIFG) have been associated with semantic
processing (e.g., Binder et al., 2009; Friederici & Gierhan, 2013; Hickok & Poeppel, 2007).
Evidence supporting phonological and semantic specialization within these regions has pri-
marily come from work with adults using direct task comparisons, a statistical approach which
cancels out irrelevant processes that are shared between two similar tasks (i.e., phonological
and semantic) and thus identifies regions sensitive purely to each individual task (i.e., phonol-
ogical or semantic processing; see Hodgson et al., 2021, for a recent meta-analysis). Although
neurocognitive theories of language development have suggested that children exhibit pho-
nological and semantic sensitivity in the brain as early as the first two years of life (see review
in Skeide & Friederici, 2016), the evidence for phonological and semantic specialization in
children using the direct task comparison approach is scarce and inconsistent.
Of the few previous studies which have examined phonological and semantic specializa-
tion using direct task comparisons in developing children, findings are mixed. Mathur et al.
(2020) studied 5- to 7-year-old children but did not find differences in brain activation
between a visual rhyming and a visual meaning task. Landi et al. (2010) recruited children
who were 9 to 19 years old and asked them to perform a cross-modal (auditory and visual)
categorical meaning judgment task and a visual rhyme judgment task. They found evidence
of a single dissociation, that is, children engaged the left STG and AG more in the meaning
task than the rhyming task but there were no regions that showed greater engagement during
the rhyming over the meaning task. Similarly, Liu et al. (2012) recruited children aged 11 to
13 years old and compared their brain activation during a visual rhyming and a visual meaning
association task. They too found evidence only for semantic-specific regions, in particular, that
the left vIFG and MTG showed greater activation for the meaning than the rhyming task, but
there was no evidence for phonology-specific regions.
One critical characteristic of the previously described studies is that they all used visual
word stimuli, which may have introduced confounds related to the visual decoding process.
Using an alternative, auditory stimuli approach, Weiss et al. (2018) and Wang, Yamasaki, et al.
(2021) aimed to investigate language-related neural specialization without the potential
confound associated with visual stimuli. Both studies showed support for specialization via
a double dissociation. Weiss et al. (2018) found that 5- to 6-year-old children showed greater
activation in the left STG during an auditory sound judgment compared to an auditory
298
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
meaning judgment task and greater activation in the left MTG during an auditory meaning
judgment task compared to an auditory sound judgment task. In a slightly older sample, Wang,
Yamasaki, et al. (2021) found that 7- to 8-year-old children showed greater activation in the left
dIFG during the sound versus meaning judgment task and greater activation in both the left
vIFG and MTG during the meaning versus sound judgment task. The progression of language
specialization from the temporal lobe to the frontal lobe, as suggested by the findings of Weiss
et al. (2018) and Wang, Yamasaki, et al. (2021), is consistent with the neurocognitive theory of
language development proposed by Skeide and Friederici (2016), which argues that language
processes in the temporal lobe develop earlier than those in the frontal lobe.
Taken together, the limited previous research on phonological and semantic specialization
in children is mixed, ranging from no to strong support for language-related specialization.
These mixed findings may be driven by methodological differences (e.g., visual vs. auditory
tasks) or different age groups selected by different studies. Regardless, it is clear that more work
is needed to better understand whether or not, in line with the interactive specialization theory,
children show evidence for early neural specialization within the language network. As is
reviewed above, previous double-dissociation-based evidence for phonological and semantic
specialization has only been found in younger children, aged 5 to 8 years old, and only when
auditory tasks were used (i.e., Wang, Yamasaki, et al., 2021; Weiss et al., 2018). No double-
dissociation-based evidence has been observed in previous studies examining older children
using visual stimuli (i.e., Landi et al., 2010; Liu et al., 2012). Thus, studying a slightly older
cohort using auditory tasks may help clarify if older children show both phonological and
semantic specialization.
The current study focused on the development of phonological awareness and semantic
association skills, both of which require metalinguistic processing in addition to phonological
and semantic representations. These two language skills have been shown to be closely
associated with children’s reading skills (e.g., Melby-Lervåg et al., 2012; Swart et al., 2017).
According to the developmental theory by Anthony and Francis (2005), phonological aware-
ness progresses from large grain sizes, such as syllabic and rhyme awareness, to small grain
sizes, such as phonemic awareness. The shift to small grain phonemic awareness usually
occurs at 5 to 7 years old, after children learn to read, and becomes stable around 9 to 10
years old (Wagner et al., 1997). Different from phonological processing, which has been
shown to be related to children’s status of dyslexia or risk of reading disability (e.g., Gu &
Bi, 2020; Noordenbos & Serniclaes, 2015; Snowling & Melby-Lervåg, 2016), phonological
awareness places more demand on metalinguistic processing, which is more strongly related
to reading skills, especially for older children or skilled readers (e.g., Wang et al., 2020;
Wang, Pines, et al., 2021). As for the development of semantic associations, the spreading
activation theory by Collins and Loftus (1975) suggests that words with higher frequency of
co-occurrence establish stronger links between concept nodes in semantic retrieval, and thus
should be earlier to develop. Consistent with this, Unger and Fisher (2021) argue that exposure
to co-occurrence regularities is the driving force for semantic knowledge development.
Children’s vocabulary growth rate has been shown to slow down at approximately 10 years
old (Rice & Hoffman, 2015). Thus, as compared to 5- to 8-year-old children, who are
experiencing shifts from child to adult-like phonological and semantic processing, 9- to
10-year-old children are near adult-like, providing an ideal age range to examine the potential
emergence of adult-like specialization within the language network.
Using the same experimental design and analytical approach as in Weiss et al. (2018) and
Wang, Yamasaki, et al. (2021), the goal of the current study was to examine phonological and
semantic specialization in 9- to 10-year-old children during auditory word processing. Based
Neurobiology of Language
299
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
on cognitive models of language processing (e.g., Binder et al., 2009; Friederici & Gierhan,
2013; Hickok & Poeppel, 2007; Skeide & Friederici, 2016) and previous findings of neural
specialization in children (e.g., Landi et al., 2010; Liu et al., 2012; Mathur et al., 2020; Wang,
Yamasaki, et al., 2021; Weiss et al., 2018), we expected that 9- to 10-year-old children would
show phonological and semantic specialization in both the frontal and temporal lobe using
direct task comparisons. Specifically, we hypothesized that the auditory meaning judgment
task would elicit greater activation than the auditory sound judgment task in semantically
related regions such as the left IFG, MTG, AG, and/or FG, whereas the auditory sound
judgment task would elicit greater activation than the auditory meaning judgment task in
phonologically related regions such as the left IFG, STG, and/or SMG. Within the left IFG,
it was predicted that the peak of activation for the sound greater than meaning task contrast
would be more dorsal than the peak of activation for the meaning greater than sound task
contrast.
In addition to direct task comparisons, a hard and an easy condition within each task (onset
vs. rhyme and low vs. high association within the sound judgment and meaning judgment
tasks, respectively) was designed. The onset condition is predicted to be more difficult than
the rhyme condition because the former requires the recognition of smaller grain sizes at the
phoneme level which are acoustically less salient and develop later (Anthony & Francis,
2005). The low association condition is predicted to be more difficult than the high association
condition because the former has a lower frequency of co-occurrence (Unger & Fisher, 2021).
We contrasted the hard and easy conditions to examine whether the specialized regions for
each task were also sensitive to within-task differences in difficulty levels. A previous meta-
analysis on adults (Hodgson et al., 2021) showed that during more difficult language tasks both
a domain-general region (i.e., the opercular part of the left IFG), which is active regardless of
task, and language-specific regions (i.e., the orbitalis and triangular part of the left IFG), which
are only responsive to certain language tasks, were engaged. Similarly, Wang, Yamasaki, et al.
(2021) showed in 7- to 8-year-old children that the opercular part of the left IFG was more
active for the hard than easy condition regardless of task, whereas the left STG was uniquely
activated more for the onset than rhyme condition in the sound judgment task. Therefore, we
hypothesized that 9- to 10-year-old children would also show both domain-general and
language-specific regions engaged during the parametric manipulations. Specifically, consis-
tent with the predictions for the direct task comparisons, we expected that regions such as the
left IFG, STG, and/or SMG would show stronger activation for the onset than the rhyme con-
dition during the auditory sound judgment task, whereas regions such as the left IFG, MTG,
AG, and/or FG would show stronger activation for the low than the high association condition
during the auditory meaning judgment task. It was predicted that the peak of activation in the
left IFG for the low greater than high association contrast would be more ventral than the peak
in the left IFG for the onset greater than rhyme contrast. In addition, we hypothesized that a
domain general control region (i.e., the opercular part of the left IFG) would show stronger
activation for the onset and low association conditions during the auditory sound and meaning
judgment tasks, respectively. All of the above hypotheses and the analytic approach outlined
below were preregistered at https://osf.io/5p3es/.
MATERIALS AND METHOD
Participants
Data for this study were pulled from a shared data set on OpenNeuro.org (see data descriptor
by Wang et al., 2022; https://openneuro.org/datasets/ds003604). The specific subjects and
Domain-general:
Brain regions, associated with
cognitive-control mechanisms, that
are recruited across domains,
particularly when tasks are more
difficult.
Language-specific:
Brain regions that are recruited only
during language tasks, and not
others.
Neurobiology of Language
300
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
runs used in the current study as well as the code used to analyze the data were shared on
GitHub (https://github.com/wangjinvandy/PhonSem_Specialization_9_10). The Institutional
Review Board of the University of Texas, Austin, approved all the experimental procedures.
Consent was collected from participants’ parents or guardians and assent was collected from
children before participation in our study.
Parents or guardians were asked to complete an exclusionary survey and a developmental
history questionnaire. All participants enrolled had normal hearing and normal/corrected-to-
normal vision, and had no learning, neurological, or psychiatric disorders. Children were
asked to complete several screening tests, which included five handedness questions, in which
the child had to pretend to write, erase, pick-up, open, and throw something, and the Diag-
nostic Evaluation of Language Variation (DELV) Part 1 Language Variation Status (Seymour
et al., 2003). Children also completed standardized tests to assess their language skills and
nonverbal IQ. General language skill was measured using the Clinical Evaluation of Language
Fundamentals—Fifth Edition (CELF-5; Wiig et al., 2013). Phonological skill was assessed
using the Comprehensive Test of Phonological Processing (CTOPP-2; Wagner et al., 2013).
Nonverbal IQ was measured using the Kaufman Brief Intelligence Test—Second Edition
(KBIT-2; Kaufman & Kaufman, 2004).
One hundred and one 9- to 10-year-old children participated in the sound and meaning
judgment functional magnetic resonance imaging (fMRI) tasks, and those who met the follow-
ing seven criteria were included in the analysis in the current study: (1) complete data obtained
for both runs of the sound and meaning tasks (16 excluded); (2) right-handed, defined as com-
pleting at least three out of the five handedness tasks with their right hand (0 excluded); (3) a
mainstream American English speaker as categorized by the Part I Language Variation Status
subtest on the DELV (0 excluded; this criterion was used because dialects may affect children’s
perception of phonology in spoken words); (4) a standardized IQ score of 80 or higher on the
KBIT-2 (6 excluded); (5) typical language abilities, as indexed by a standardized Core Lan-
guage Scale score of 80 or higher on the CELF-5 (0 excluded); (6) no excessive movement
during the fMRI tasks (5 excluded, see Data Analysis for criteria); and (7) good fMRI task
accuracy (13 excluded, see Experimental Procedure for criteria). In addition, the language test
(i.e., CELF-5), which is more interactive, was always the first standardized test administered.
In the end, 61 children (37 females, 24 males, mean age = 9.20, SD = 0.19, range = 8.96 to
9.87 yr old) were included in the final sample for this study. (Note, we re-analyzed the data by
adding 4 participants who had low IQ scores. We added 4 participants, rather than 6, because
one participant was left-handed, and one had low in-scanner accuracy. We found that all
findings remained the same except that the cluster in the left STG (k = 32) was no longer
significant for the contrast of Onset > Rhyme after family-wise error, or FWE, correction.)
Among the participants, 33 children were included in a previous study examining phonolo-
gical and semantic specialization in 7- to 8-year-old children (Wang, Yamasaki, et al., 2021)
and nine children were included in a previous study on phonological and semantic special-
ization in 5- to 6-year-old children (Weiss et al., 2018).
Experimental Procedure
The sound judgment task
The sound judgment task taps into children’s phonological processing skill for spoken words.
In this task, participants heard a one-syllable word pair presented sequentially through ear-
phones. Children were asked to judge whether the word pair shares any of the same sounds.
Real words were used in the sound judgment task instead of the more traditional use of
Neurobiology of Language
301
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
pseudowords, which might induce semantic processing in addition to phonological process-
ing. However, we designed the task in this way because the aim was to use a direct task com-
parison approach to isolate the regions that were specifically recruited for phonological or
semantic processing, so the type of word had to be kept constant between the sound and
meaning judgment tasks. The sound judgment task included three different experimental con-
ditions: rhyme, onset, and unrelated (see Table 1 for examples, and see a full list of stimuli in
the Supporting Information). Children were expected to press the “yes” button for both the
onset and rhyme conditions and the “no” button for the unrelated condition. In addition to
the three experimental conditions, the task also included a perceptual control condition in
which participants heard two sequentially presented frequency-modulated sounds (i.e.,
“shh-shh”) and were only asked to press the “yes” button. Participants completed two runs
of the task with 12 trials per condition per run for a total of 24 trials for each of the four con-
ditions. The task included a total of 96 trials divided into two separate 48-trial runs. Each audi-
tory word had a duration ranging from 439 to 706 ms. The second word was presented
approximately 1,000 ms after the onset of the first word. Overall, within each trial, the stimuli
duration (i.e., the two words with a brief pause in between) ranged from 1,490 to 1,865 ms and
was followed by a jittered response interval ranging from 1,500 to 2,736 ms. A blue circle
appeared simultaneously with the auditory presentation of the stimuli to help maintain atten-
tion on the task. The blue circle changed to yellow, to provide a 1,000 ms warning for partic-
ipants to respond if they had not already done so, before moving on to the next trial. The total
trial duration ranged from 3,000 to 4,530 ms. Each run lasted approximately 3 min.
The auditory word conditions were designed according to the following standards. For the
onset condition, the word pairs shared the same initial phoneme (corresponding to one letter
at the beginning of their written form). For the rhyme condition, the word pairs shared the
same final vowel and phoneme/cluster (corresponding to two to three letters at the end of their
written form). For the unrelated condition, there were no shared phonemes at any locations in
a word pair although one letter of the written form may be shared on some occasions (e.g.,
land–face). All words were monosyllabic, and all word pairs had no semantic association
based on the University of South Florida Free Association Norms (Nelson et al., 2004). There
were no significant differences between conditions in word length, number of phonemes,
written word frequency, orthographic neighbors, phonological neighbors, semantic neighbors,
Task
Sound task
Meaning task
Table 1.
Experimental conditions in the sound and meaning judgment tasks
Condition
Onset
Rhyme
Unrelated
Perceptual
Low
High
Unrelated
Perceptual
Response
Yes
Brief explanation
Two words share the first sound
Yes
No
Yes
Yes
Yes
No
Yes
Two words share the final sound
Two words do not share sounds
Frequency modulated noise
Two words are weakly associated in meaning
Two words are strongly associated in meaning
Two words are not related in meaning
Frequency modulated noise
Example
Coat–Cup
Wide–Ride
Zip–Cone
Shh–Shh
Dish–Plate
Dog–Cat
Map–Hut
Shh–Shh
Note. In the sound judgment task, children were asked: “Do the two words share any of the same sounds?” In the meaning judgment task, children were asked:
“Do the two words go together?”
Neurobiology of Language
302
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
or number of morphemes (Rhyme vs. Onset: ps > 0.123; Rhyme or Onset vs. Unrelated: ps >
0.123; linguistic characteristics were obtained from the English Lexicon Project by Balota
et al., 2007). There were also no significant differences between conditions in phoneme prob-
abilities obtained from a phonotactic probability calculator (Rhyme vs. Onset: ps > 0.302;
Rhyme or Onset vs. Unrelated: ps > 0.203; Vitevitch & Luce, 2004).
The meaning judgment task
The meaning judgment task examines children’s semantic processing skill for spoken words. In
this task, participants heard a one- or two-syllable word pair presented sequentially through
earphones. They were asked to determine whether the word pair goes together semantically.
The task included three different experimental conditions: low association, high association,
and unrelated (see Table 1 for examples, and see a full list of stimuli in the Supporting
Information). Children were expected to press the “yes” button for both the low and high asso-
ciation conditions and the “no” button for the unrelated condition. In addition to the three
experimental conditions, the task included a perceptual control condition in which partici-
pants heard two sequentially presented frequency-modulated sounds (i.e., “shh-shh”) and
were only asked to press the “yes” button. Participants completed two runs of the task with
12 trials per condition per run for a total of 24 trials for each of the four conditions. The task
included a total of 96 trials divided into two separate 48-trial runs. Each auditory word had a
duration ranging from 500 to 700 ms. The second word was presented approximately 1,000 ms
after the onset of the first word. Overall, within each trial, the stimuli duration (i.e., the two
words with a brief pause in between) ranged from 1,500 to 1,865 ms and was followed by a
jittered response interval ranging from 1,800 to 2,701 ms. A blue circle appeared simulta-
neously with the auditory presentation of the stimuli to help maintain attention on the task.
The blue circle changed to yellow, to provide a 1,000 ms warning for participants to respond
if they had not already done so, before moving on to the next trial. The total trial duration
ranged from 3,300 to 4,565 ms. Each run lasted approximately 3 min.
The auditory word conditions were designed according to the following standards. Words,
with relatively high frequencies, were selected from the database of the University of South
Florida Free Association Norms (Nelson et al., 2004). The low and high association semantic
relationships were determined using the forward cue-to-target strength (FSG) values reported
from the norm. The low association condition was defined as word pairs having a weak
semantic association with FSG values between 0.14 and 0.39 (mean = 0.27, SD = 0.07).
The high association condition was defined as word pairs having a strong semantic association
with FSG values between 0.40 and 0.85 (mean = 0.64, SD = 0.13). The unrelated condition
was defined as word pairs that had no FSG values. The FSG values reflect the proportion of
subjects in the group who produce a particular target in the presence of the cue word. The
norming is not based on children, and therefore, is a limitation of the design in this study.
However, as can be seen below, the behavioral data from this study showed that children
responded more poorly to the low association word pairs, which suggests that the intended
manipulation of semantic association strength was successful in our sample of young children
(see statistics in Results). There were no significant differences in association strength between
the two runs of the meaning judgment task ( ps > 0.425). There were also no significant
differences between conditions in word length, number of phonemes, number of syllables,
written word frequency, orthographic neighbors, phonological neighbors, semantic neighbors,
or number of morphemes (High vs. Low: ps > 0.167; High or Low vs. Unrelated: ps >
0.068; linguistic characteristics were obtained from the English Lexicon Project by Balota
et al., 2007).
Neurobiology of Language
303
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
Participants who scored within an acceptable accuracy range and demonstrated no
response bias on the fMRI tasks were included in the final analysis. Specifically, to be
included, children had to score greater than or equal to 50% on the perceptual and
rhyme/high conditions (to ensure that children were engaged in and capable of performing
the tasks), and children had to have an accuracy difference between the rhyme/high condition
(requiring a “yes” response) and the unrelated condition (requiring a “no” response) of lower
than 40% (to ensure that there was no apparent response bias during the tasks). The average
reaction time (RT) for each condition was based on correct trials only and was calculated from
the onset of the second word for the three experimental word conditions, and the onset of
the trial for the perceptual control condition. Reaction times, which were less than or greater
than 3 standard deviations from the mean of all correct trials within a run, or were less than
250 ms, were excluded.
Data Acquisition
Participants lay in the scanner with a response button box placed in their right hand. To keep
participants focused on the task, visual stimuli were projected onto a screen, viewed via a
mirror attached to the head coil. Participants wore earphones to hear the auditory stimuli,
and two pads placed in between the earphones and the head coil were used to reduce move-
ment and attenuate scanner noise.
Images were acquired using a 3.0 T Skyra Siemens scanner with a 64-channel head coil.
The blood oxygen level dependent (BOLD) signal was measured using a susceptibility
weighted single-shot echo planar imaging (EPI) method. Functional images were acquired with
multiband EPI. The following parameters were used: TR = 1,250 ms, TE = 30 ms, flip angle =
80°, matrix size = 128 × 128, FOV = 256 mm2, slice thickness = 2 mm without gaps, number
of slices = 56, multiband acceleration factor = 4, voxel size = 2 × 2 × 2 mm. A high resolution
T1-weighted MPRAGE scan was acquired with the following scan parameters: TR = 1,900 ms,
TE = 2.34 ms, matrix size = 256 × 256, FOV = 256 mm2, slice thickness = 1 mm, number of
slices = 192.
Data Analysis
Statistical Parametric Mapping 12 (SPM12; Ashburner et al., 2021) was used to analyze the
MRI data. First, all functional images were realigned to their mean functional image across
runs. The anatomical image was segmented and warped to a pediatric tissue probability
map template to get the transformation field. An anatomical brain mask was created by com-
bining the segmented products (i.e., grey, white, and cerebrospinal fluid) and then applied to
its original anatomical image to produce a skull-stripped anatomical image. All functional
images, including the mean functional image, were then co-registered to the skull-stripped
anatomical image. All functional images were then normalized to a pediatric template by
applying the transformation field to them and re-sampled with a voxel size of 2 × 2 ×
2 mm. The pediatric tissue probability map template was created using CerebroMatic (Wilke
et al., 2017), a tool that makes SPM12 compatible pediatric templates with user-defined age,
sex, and magnetic field parameters. The unified segmentation parameters estimated from
1,919 participants (Wilke et al., 2017; parameters downloaded from Universitätsklinikum
Tübingen, 2022) were used. We defined our parameters as a magnetic field strength of
3.0 T, age range from 9 to 11 years old with one-month intervals, and sex as two females
and two males at each age interval to obtain our age-appropriate pediatric template. After
Neurobiology of Language
304
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
normalization, smoothing was applied to all the functional images with a 6 mm isotropic
Gaussian kernel.
To reduce movement effects on the brain signal, Art-Repair (Mazaika et al., 2009) was
used to identify outlier volumes, which were defined as those with volume-to-volume head
movement exceeding 1.5 mm in any direction, head movements greater than 5 mm in any
direction from the mean functional image across runs, or deviations of more than 4% from
the mean global signal intensity. These outlier volumes were then repaired using interpolation
based on the nearest non-outlier volumes. Participants included in our study had no more
than 10% of the volumes and no more than six consecutive volumes repaired within each
run. The movement criteria were based on those used in previous studies using the same
experimental design (e.g., Wang, Yamasaki, et al., 2021; Weiss et al., 2018). Generally speak-
ing, these criteria are more stringent than those used by other research with younger cohorts
(e.g., 5- to 7-year-old children by Mathur et al., 2020; 8-year-old children by Girard et al.,
2022) and less stringent than those used by other research with older children (e.g., 6- to
19-year-old children by Baker et al., 2020). Six motion parameters estimated during realign-
ment were entered during first level modeling as regressors of no interest, and the repaired
volumes were deweighted.
Statistical analyses at the first level were calculated using an event-related design. A high
pass filter with a cutoff of 128 s and an SPM default mask threshold of 0.5 were applied. All
experimental trials were included as individual events for analysis and modeled using a
canonical hemodynamic response function (HRF). All four conditions in each task run (i.e.,
onset, rhyme, unrelated and perceptual in the sound judgment task, and low, high, unrelated
and perceptual in the meaning judgment task) were taken as regressors of interest and entered
into the general linear model. We compared the related conditions (i.e., Onset + Rhyme) with
the perceptual condition during the sound judgment task to obtain the brain activation map for
phonological processing within each participant. We compared the related conditions (i.e.,
Low + High) with the perceptual condition during the meaning judgment task to obtain the
brain activation map for semantic processing within each participant. To examine neural
specialization within each participant, we compared brain activation between the two tasks
(the sound task (Related > Perceptual) > the meaning task (Related > Perceptual), or the
meaning task (Related > Perceptual) > the sound task (Related > Perceptual)]. To examine
the parametric modulation effect within each participant, we contrasted the two Related
conditions within each task (Onset > Rhyme in the sound task, and Low > High in the
meaning task).
In the second-level analyses, task comparison contrast maps from each individual (i.e., the
sound task > the meaning task, or the meaning task > the sound task) were entered into a one-
sample t test to generate a brain specialization map at the group level for either phonological
or semantic processing. Contrast maps for the parametric modulations from each individual
(i.e., Onset > Rhyme or Low > High) were also entered into a one sample t test to generate
a parametric modulation map at the group level for either phonological or semantic process-
ing. We used the SPM12 small volume FWE correction to determine the significance of a clus-
ter within our functional language mask. The functional language mask reflects the union of
activation for the Related > Perceptual in the sound and meaning judgment tasks within a
literature-based anatomical mask. The literature-based anatomical mask included the left
IFG, STG, MTG, SMG, AG, IPL, and FG (Binder et al., 2009; Friederici & Gierhan, 2013;
Hickok & Poeppel, 2007) and is consistent with the mask used by Weiss et al. (2018) and
Wang, Yamasaki, et al. (2021) with younger children. Results at the whole brain level were
also calculated using the SPM12 FWE correction.
Neurobiology of Language
305
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
In addition, exploratory analyses were performed to detect whether the strength of phonol-
ogical and semantic specialization was associated with children’s language skill. Specifically,
we carried out two voxel-wise regression analyses to examine the correlation between brain
activation for the contrast Sound (Related > Perceptual) > Meaning (Related > Perceptual) and
raw scores on the Phoneme Isolation subtest of the CTOPP-2 (Wagner et al., 2013) and the
correlation between brain activation for the contrast Meaning (Related > Perceptual) > Sound
(Related > Perceptual) and raw scores on the Word Classes subtest of the CELF-5 (Wiig et al.,
2013).
RESULTS
Table 2 shows the mean, standard deviation, and range of the accuracies and RTs for each
condition during the sound and the meaning judgment fMRI tasks. All children included in
the study had above chance level (i.e., 50%) overall performance for each task (meansound =
88.7%, t (60) = 45.83, p < 0.001; meanmeaning = 90.8%, t (60) = 48.94, p < 0.001). In addition,
the hard condition showed significantly lower accuracy and longer RTs than the easy condition
in both the sound (t (60)ACC = 9.587, p < 0.001, t (60)RT = −3.086, p = 0.003) and the meaning
(t (60)ACC = 3.448, p = 0.001, t (60)RT = −6.087, p < 0.001) judgment tasks, supporting the
validity of the parametric manipulation in the current study.
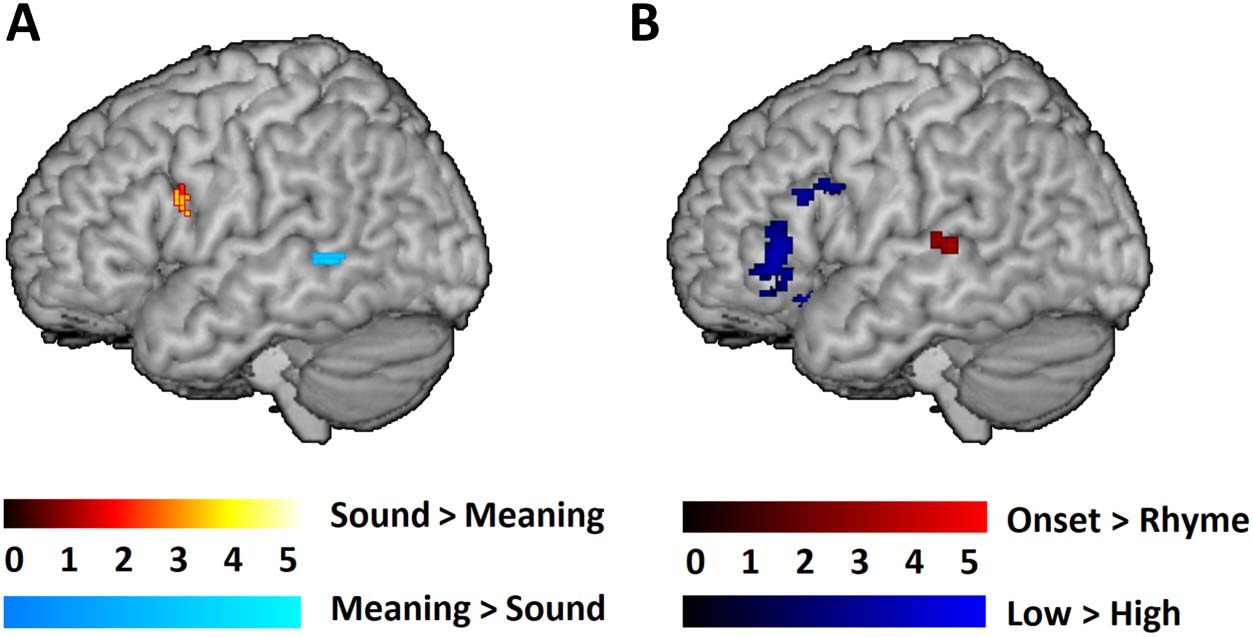
The preregistered (https://osf.io/5p3es/) univariate voxel-wise results within the language
mask for the direct task comparisons and parametric manipulations are shown in Table 3
and Figure 1. The direct comparisons between the sound and meaning judgment tasks (see
Figure 1A) revealed significantly greater activation for the sound judgment task in the left oper-
cular part of the IFG and significantly greater activation for the meaning judgment task in the
left MTG. (Brain specialization maps after controlling covariates of no interest including task
accuracy, nonverbal IQ, and core language skill are displayed in Table S1 and Figure S1 in the
Supporting Information. The main findings remained the same except that the opercular part of
the left IFG was no longer significant for the contrast of Low > High in the meaning task.) In
terms of the parametric manipulations, the comparison between the onset and rhyme condi-
tions within the sound judgment task showed significant clusters in the left STG/STS (superior
temporal sulcus; see Figure 1B). The comparison between the low and high association con-
ditions within the meaning judgment task showed significant clusters in both the
triangular/orbitalis and opercular part of the IFG (see Figure 1B). As is shown in Figure 1,
Table 2.
Behavioral performance during the sound and the meaning judgment tasks
Accuracy (%)
Reaction time (ms)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Tasks
Sound task
Conditions
Onset
Mean (SD)
75.8 (15.1)
Range
29.1–92.4
Mean (SD)
1,240 (179)
Rhyme
91.7 (8.5)
Unrelated
89.3 (8.6)
Perceptual
97.8 (3.1)
Meaning task
Low
86.7 (11.7)
High
90.7 (9.6)
Unrelated
88.2 (10.3)
Perceptual
97.4 (4.0)
66.7–100
62.5–100
87.5–100
54.2–100
58.3–100
62.5–100
83.3–100
Neurobiology of Language
Range
924–1,651
933–1,565
1,191 (160)
1,296 (190)
1,002–1,749
1,225 (451)
1,244 (153)
1,158 (164)
1,325 (162)
1,263 (487)
516–2,267
832–1,663
799–1,559
934–1,736
566–2,478
306
Phonological and semantic specialization in 9- to 10-year-olds
Table 3. Voxel-wise analysis significant results within the combined functional and literature-
based anatomical mask
Brain regions
Sound task (Related > Perceptual) > Meaning task (Related > Perceptual)
Brodmann
area
Peak coordinate
(MNI)
Number
of voxels
T value
Opercular left IFG
44
−42 2 24
103
5.06
Meaning task (Related > Perceptual) > Sound task (Related > Perceptual)
Left MTG
21
−58 −46 0
Onset > Rhyme in the sound task
Left STG/STS
22
−66 −32 6
Low > High in the meaning task
Triangular/orbitalis
45/47
left IFG
Opercular left IFG
44
−54 32 2
−50 12 28
37
39
288
71
4.28
3.69
4.40
4.09
Note. IFG = inferior frontal gyrus, MTG = middle temporal gyrus, STG = superior temporal gyrus, STS = superior
temporal sulcus, MNI = Montreal Neurological Institute.
the significant clusters found in the parametric manipulations did not overlap with the signif-
icant clusters found in the direct task comparisons. Finally, the regression analyses for the
preregistered exploratory analyses did not reveal any significant clusters between
specialization-related brain activation and phonological awareness or semantic association
skill as measured by the standardized tests.
The non-preregistered univariate voxel-wise results at the whole brain level for each task,
the direct task comparisons, and the parametric manipulations are displayed in Table 4 and
Figure 2. We focused our summary of results on the regions of interest specified in the
literature-based anatomical mask, including their homologues in the right hemisphere. We
Figure 1. The univariate voxel-wise results within the combined functional and literature-based
anatomical mask. (A) Task comparisons: Sound (Related > Perceptual) > Meaning (Related > Per-
ceptual) in hot colors; Meaning (Related > Perceptual) > Sound (Related > Perceptual) in cold
colors. (B) Parametric manipulations: Onset > Rhyme within the sound task in red; Low > High
within the meaning task in blue. All clusters were significant at a voxel-wise p < 0.001 uncorrected,
and a cluster-wise p < 0.05 family-wise-error corrected, using the SPM12 small volume correction.
Neurobiology of Language
307
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
Table 4.
Voxel-wise analysis significant results at the whole brain level
Brain regions
Sound task (Related > Perceptual)
Brodmann area
Peak coordinate (MNI)
Number of voxels
T value
Left STG
Right STG
Left fusiform
Right insula
Left putamen
Left thalamus
Left supplementary motor area
Right lingual gyrus
Left calcarine
Right precentral gyrus
Right MTG
Meaning task (Related > Perceptual)
Right MTG/STG
Left STG/MTG/IFG
Left inferior temporal gyrus
Right para-hippocampus
22
22
37
13
–
–
6
19
17
6
21
21/22
22/21
20
–
−62 8 0
62 −6 −4
−42 −44 −16
32 20 2
−16 8 6
−10 −16 6
−6 6 60
16 −48 4
−16 −50 8
54 −4 46
56 −74 2
66 −4 −2
−64 −10 2
−38 −14 −28
22 −6 −22
Sound task (Related > Perceptual) > Meaning task (Related > Perceptual)
Left precentral gyrus
Right precentral gyrus
Left middle occipital lobe
6
6
19
−58 6 24
52 2 36
−42 −90 10
Meaning task (Related > Perceptual) > Sound task (Related > Perceptual)
Left MTG
Onset > Rhyme in the sound task
Left postcentral gyrus
Left insula
Left caudate
Left precentral gyrus
Low > High in the meaning task
Left orbitalis/triangular IFG
Left supplementary motor area
Neurobiology of Language
21
4
13
–
3
47/45
8
−62 −46 −4
−34 −10 30
−22 26 16
−16 −2 26
−40 −18 62
−32 22 −4
−6 20 50
7725
2691
1648
255
1654
110
431
136
131
118
132
2659
5369
1338
172
953
307
108
144
115
141
142
502
389
131
17.04
14.19
9.16
6.79
6.73
6.51
5.64
5.15
5.06
5.03
4.77
14.79
14.08
8.07
5.16
5.86
4.57
4.03
5.20
4.99
4.74
4.74
4.10
4.74
4.13
308
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
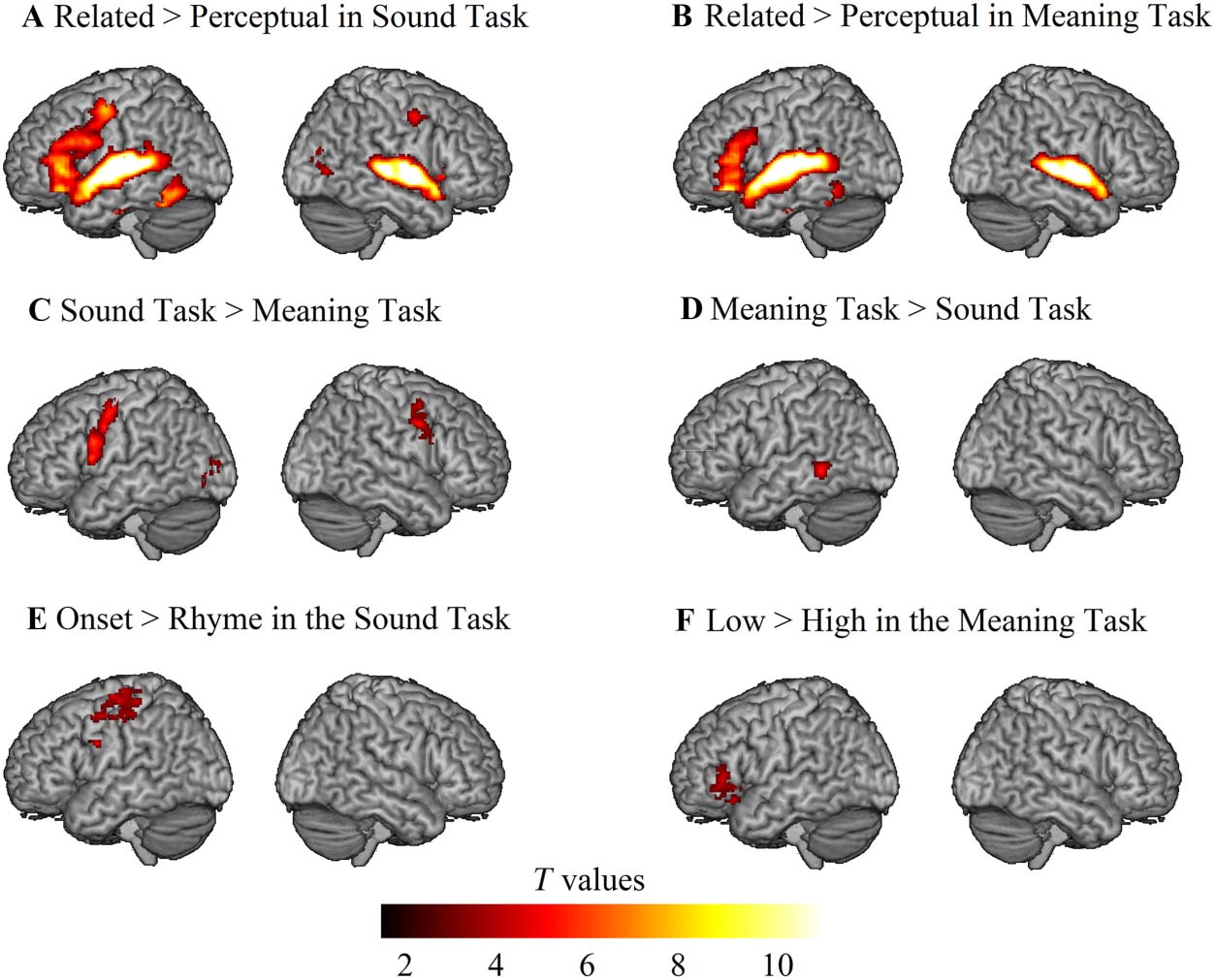
Figure 2. The univariate voxel-wise results at the whole-brain level. (A) Related > Perceptual in the
Sound Task; (B) Related > Perceptual in the Meaning Task; (C) Sound Task (Related > Perceptual) >
Meaning Task (Related > Perceptual); (D) Meaning Task (Related > Perceptual) > Sound Task
(Related > Perceptual); (E) Parametric manipulation in the Sound Task (Onset > Rhyme); (F) Para-
metric manipulation in the Meaning Task (Low > High). All clusters were significant at a voxel-wise
p < 0.001 uncorrected, and cluster-wise p < 0.05 family-wise-error corrected in SPM12 at the whole
brain level.
found that the left IFG, bilateral STG/MTG, and bilateral FG were commonly activated across
the sound (Figure 2A) and meaning (Figure 2B) judgment tasks. Direct task comparisons did
not reveal clusters in the literature-based anatomical areas that showed greater activation for
the sound task (Figure 2C), whereas the meaning task induced greater activation in the left
MTG (Figure 2D). As for the parametric manipulation effects, no clusters in the literature-based
anatomical areas showed greater activation for the onset than the rhyme condition within the
sound judgment task (Figure 2E), whereas the orbitalis/triangular part of the left IFG exhibited
greater activation for the low than the high association condition within the meaning judgment
task (Figure 2F). In terms of the regression analysis, we did not find any significant clusters
showing a correlation between brain activation for specialization and language skills as mea-
sured by standardized testing at the whole brain level.
DISCUSSION
This study examined phonological and semantic specialization in 9- to 10-year-old children
using the same experimental design and analytical approach as used in previous studies with
younger children aged 5 to 6 years old (Weiss et al., 2018) and 7 to 8 years old (Wang,
Yamasaki, et al., 2021). Similar to the findings with 7- to 8-year-old children, the current study
showed phonological and semantic specialization in both the frontal and temporal lobes. Spe-
cifically, the left dIFG showed greater activation for the sound than the meaning judgment task,
whereas the left MTG showed greater activation for the meaning than the sound judgment
task. In terms of the parametric manipulations, we found that the left STG showed greater acti-
vation for the onset than the rhyme condition during the sound judgment task, whereas both
Neurobiology of Language
309
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
the left vIFG and the opercular part of the left IFG showed greater activation for the low than
high association condition during the meaning judgment task. Overall, this pattern of results
suggests that, despite a few differences, phonological and semantic specialization in 9- to
10-year-old children remains similar to that observed in 7- to 8-year-old children.
The current study found that the left dIFG showed greater activation for the sound than the
meaning task in 9- to 10-year-old children, showing support for phonological specialization in
the frontal lobe. This finding is consistent with a previous study on 7- to 8-year-old children
(Wang, Yamasaki, et al., 2021), which also showed phonological specialization in the frontal
but not the temporal lobes. Phonological specialization in the temporal lobe was only
observed in a study with even younger children aged 5 to 6 years old (Weiss et al., 2018),
in which the left STG showed greater activation for the sound than the meaning judgment task.
It has been hypothesized that the left STG is associated with the quality of phonological rep-
resentation, whereas the left dIFG is associated with phonological access and manipulation
(e.g., Boets et al., 2013; Myers et al., 2009). Together with the previous studies on younger
children (Wang, Yamasaki, et al., 2021; Weiss et al., 2018), the current study suggests a devel-
opmental trajectory of phonological specialization from the temporal to the frontal lobe.
Previous research has also shown that younger children vary in the representational quality
of phonological forms in STG as a function of skill (e.g., Wang et al., 2020), whereas older
children vary in the efficiency of phonological access in IFG as a function of skill (Wang, Pines,
et al., 2021). It is likely that although both representation and access are needed to perform
phonological awareness tasks, young children rely more on the quality of phonological rep-
resentation in the left STG, whereas children older than 7 to 8 years old consistently rely more
on the efficiency of phonological access and manipulation in the left dIFG.
However, unlike the left dIFG observed in the previous study on 7- to 8-year-old children,
which was localized to the triangular part of the left IFG (Wang, Yamasaki, et al., 2021), the left
dIFG observed in the current study on 9- to 10-year-old children was in the opercular part of
the left IFG. Previous research suggests that the opercular part of the left IFG functions as a
domain-general control region and is engaged in difficult tasks (e.g., Hodgson et al., 2021).
Consistent with this interpretation, the overall accuracy for the sound task was significantly
lower than that for the meaning task (85.6% for the sound task, 88.5% for the meaning task,
t = −2.905, p = 0.005), suggesting that the sound task was more difficult than the meaning task.
However, when task accuracy was controlled for, the opercular part of the left IFG was still
significantly more active for the sound than the meaning task, suggesting its role specifically in
phonologically related control processes. In line with this argument, previous research has
shown that the opercular part of the left IFG is a core region for phonological processing,
whereas the triangular part of the left IFG is involved in semantic processing in adults (e.g.,
Costafreda et al., 2006; Katzev et al., 2013; Poldrack et al., 1999). Thus, the progression of
phonological specialization from the triangular to opercular part of the left IFG likely suggests
that phonological access and manipulation in 9- to 10-year-old children, as compared to 7- to
8-year-old children, is more adult-like and, potentially, more effective. In support of this argu-
ment, we observed that the mean accuracies for the onset and rhyme conditions (i.e., onset:
76%, rhyme: 92%) in the current study were higher than those found for 7- to 8-year-old chil-
dren (i.e., onset: 70%, rhyme: 88%; Wang, Yamasaki, et al., 2021). However, because the cur-
rent study is cross-sectional, future studies with longitudinal designs are needed to examine
the developmental trajectory that is suggested by the combination of results from the current
study and previous work.
As for semantic specialization, we found that the left MTG showed greater activation for the
meaning than the sound judgment task. Previous research has already shown that the left MTG
Neurobiology of Language
310
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
is a semantic processing region and is reliably engaged in the analysis of semantic relatedness
(e.g., Binder et al., 2009; Enge et al., 2021). Skeide et al. (2014) also found that the temporal
lobe shows gradual specialization for semantic processing and away from syntactic processing
from 7 to 8 years old. Thus, the finding of semantic specialization in the left MTG in the current
study is not surprising. This finding is also consistent with the study by Liu et al. (2012) with 11-
to 13-year-old children using visual tasks and previous studies using the same experimental
design and analytical approach with 7- to 8-year-old children (Wang, Yamasaki, et al., 2021)
and 5- to 6-year-old children (Weiss et al., 2018), in which children also showed greater acti-
vation for the meaning than the sound judgment task in the left MTG, at a similar location.
However, unlike the previous study on 7- to 8-year-old children (Wang, Yamasaki, et al.,
2021), the current study did not find that the left vIFG showed greater activation for the mean-
ing than the sound judgment task in 9- to 10-year-old children. The lack of a significant finding
in the left vIFG may be a power issue. When the statistical threshold was lowered from voxel-
wise p < 0.001 to voxel-wise p < 0.005, we found a cluster showing greater activation for the
meaning than the sound judgment task in the orbitalis/triangular part of the left IFG (peak MNI
−46 34 −14, k = 17), a similar location to that found previously with 7- to 8-year-old children.
Because we did not conduct an a priori power analysis it is difficult to determine if the lack of
the finding is due to a lack of power or reflects no effect, and more research is needed to
examine the replicability of this weak finding. In contrast to the observed frontal specialization
for phonological processing, the lack of observed semantic specialization in the frontal lobe in
9- to 10-year-old children may have also been driven by the fact that the meaning judgment
task was easier than the sound judgment task, as the frontal lobe is known to be engaged par-
ticularly when tasks are more difficult or demanding (e.g., Burton et al., 2000; Chiou et al.,
2018; Katzev et al., 2013; Okada et al., 2018; Xie & Myers, 2018). Overall, the similarity in the
pattern of findings for semantic specialization across multiple studies (e.g., Liu et al., 2012;
Wang, Yamasaki, et al., 2021; Weiss et al., 2018) likely suggests that the semantic system
remains stable over middle childhood. This argument is supported by a previous meta-analysis
on the semantic system in developing children ages 4 to 15 years old (Enge et al., 2021), in
which they found little evidence for age-related changes across childhood and high overlap
with the adult semantic system.
In terms of the parametric manipulations, we found that the left STG showed greater acti-
vation for the onset than the rhyme condition during the sound judgment task. This finding is
consistent with previous studies with 7- to 8-year-old and 5- to 6-year-old children (Wang,
Yamasaki, et al., 2021; Weiss et al., 2018) showing that the left STG was more engaged for
the onset than the rhyme condition during the sound judgment task. The left STG has been
consistently shown to be associated with phonological representation (Boets et al., 2013;
Mesgarani et al., 2014) and was found engaged more for onset than rhyme processing in children
with higher phonological skill (e.g., Wang, Joanisse, & Booth, 2021). Thus, it is unsurprising
that the onset condition, which requires more precise phonological representations, engaged
the left STG more than the rhyme condition during the sound judgment task. However, some-
what surprisingly, we did not observe any significant clusters in the left IFG in 9- to 10-year-old
children for the phonological parametric manipulation. This is in contrast to the previously
reported finding that in 7- to 8-year-old children the left opercular part of the left IFG was more
active for the onset than the rhyme condition. The engagement of the opercular part of the left
IFG found in 7- to 8-year-old children was hypothesized to be reflective of a general cognitive
control process because both the sound and the meaning tasks elicited parametric effects in
this area and direct task comparisons revealed no activation difference. The lack of a signif-
icant parametric effect in the opercular part of the left IFG in the current study might suggest
Neurobiology of Language
311
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
that 9- to 10-year-old children rely on other mechanisms to deal with the more difficult onset
judgment. At the whole brain level, the left precentral gyrus was consistently activated during
the sound judgment task and was more activated for the onset than the rhyme condition
(although at a more dorsal location). This area is close to the Exner’s area, a region associated
with handwriting, bridging orthographic and motion systems (Roux et al., 2009). Although
children’s phonemic awareness could appear as early as 4 to 5 years old (Anthony & Francis,
2005), learning to read connects written and spoken languages and thus sculpts the nature of
phonological processing in the brain (e.g., Wang et al., 2020; Wang, Pines, et al., 2021).
Therefore, 9- to 10-year-old children in the current study may have employed a strategy of
spelling out the spoken words to help perform the more fine-grained phonological awareness
task (i.e., the onset judgment). However, given that we did not have a spelling measurement to
confirm the function of this area, more studies are needed to examine this speculation.
For the semantic parametric manipulation analysis, we found that both the left vIFG and the
left opercular part of the left IFG showed greater activation for the low compared to the high
association condition during the meaning judgment task. Previous literature on adults and chil-
dren (e.g., Poldrack et al., 1999; Wang, Yamasaki, et al., 2021) has already shown that the left
vIFG is a region specialized for semantic processing, and that, in adults, it is more engaged
when semantic associations are atypical or more demanding (e.g., Chiou et al., 2018; Katzev
et al., 2013). In the current study, children performed significantly worse in the low than the
high association condition (t (60) = 3.45, p = 0.001), suggesting that the low association con-
dition was more difficult. This difference in difficulty between conditions may be driving the
engagement of the opercular part of the left IFG, as it is a general cognitive control region often
recruited to promote a non-automatic but appropriate response (e.g., Novick et al., 2010). In
comparison with the previous finding that 7- to 8-year-old children only recruited the opercu-
lar part of the left IFG for the low association condition (Wang, Yamasaki, et al., 2021), the
additional finding of a parametric effect in a semantically specialized region (i.e., the left vIFG)
in the current study suggests that 9- to 10-year-old children start to engage more task-specific
regions to solve more difficult semantic problems.
It is interesting to note that when looking at the parametric effects for the sound and meaning
judgment tasks together, the patterns of activation do not overlap. This may suggest that 9- to
10-year-old children rely more on task-specific regions to deal with more difficult language tasks,
which is different from the previous findings with 7- to 8-year-old children (Wang, Yamasaki,
et al., 2021), where the parametric effects were largely overlapping in the opercular part of
the left IFG. To examine if the parametric effects observed in each task were task-specific or
domain-general, additional analyses comparing the parametric effects across tasks were con-
ducted. In 7- to 8-year-old children, Wang, Yamasaki, et al. (2021) showed no task differences,
suggesting that those children tended to use the same general control mechanisms to tackle
more difficult language problems. However, using the same analytical approach, it was found,
in the current study, that the left STG (peak MNI −60 −18 12, k = 132) was significantly more
activated for the sound (Onset > Rhyme) than the meaning (Low > High) parametric manipu-
lation. In addition, a small nonsignificant cluster in the left vIFG (peak MNI −58 30 6, k = 2)
was found to be activated more for the meaning (Low > High) than the sound (Onset > Rhyme)
parametric manipulation. When the voxel-wise threshold was lowered from p < 0.001 to p <
0.005, we found a relatively big cluster at the same location (peak MNI −58 30 6, k = 37) in
the left vIFG. The weaker parametric effect in the meaning judgment task as compared to the
sound judgment task parallels the accuracy difference between the hard and easy conditions
in the meaning judgment task (i.e., 4%, Low: 86.7%, High: 90.7%) versus that of the sound
judgment task (i.e., 15.9%, Onset: 75.8%, Rhyme: 91.7%). Overall, these additional analyses
Neurobiology of Language
312
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
help to confirm that unlike 7- to 8-year-old children who tend to rely on a domain-general
control system, 9- to 10-year-old children use more language-specific regions to deal with
difficult language tasks.
The implication of a developmental progression from reliance on domain-general to
domain-specific processing in the current study is consistent with a domain-relative framework
for cognitive development (e.g., Karmiloff-Smith, 2015; Sloutsky, 2010), which argues that the
infant brain comes equipped with biases that are relevant to, but not initially specific to, pro-
cessing different kinds of input. Therefore, domain-general learning mechanisms serve as a
beginning state and domain-specific mechanisms gradually appear via neural competition
and stimulation from the environment. Although this theory has not yet been supported by
language specialization studies, this argument has consistently been supported by previous
studies on word learning during infancy using computational and behavioral approaches.
For example, Mayor and Plunkett (2010) developed a neurocomputational model and showed
that domain-specific word learning constraints can emerge out of domain-general, associative
learning principles when confronted with a structured environment. Consistent with this argu-
ment, Namy (2012) reviewed previous studies and found that word learning started out gen-
eral and became largely a domain-specific ability over the course of the second year. Similarly,
Samuelson and McMurray (2017) argued that domain-general processes played an important
role in initial word learning. Although word learning likely draws on different processes than
those employed in the semantic association and phonological awareness tasks in the current
study, our findings suggest that the underlying neural development during language processing
from domain-general to domain-specific is similar. Karmiloff-Smith (1998) argues that under-
standing development itself is the key to understanding the complexity of developmental
language disorders, because deviance in this developmental trajectory is likely to result in a
cascade of dynamic subtle deficits across domains rather than a single, static domain-specific
one. Our finding of a shift from relying on domain-general to domain-specific mechanisms in
9- to 10-year-old children during more difficult language processing provides critical develop-
mental information. That is, typical children continue to develop domain-specificity during
word processing until 9 to 10 years old, before which domain-general ability might play a
larger role in word processing, especially during more difficult language tasks.
As for the proposed exploratory analyses, we did not find any significant clusters showing a
correlation between brain specialization and language skills, which is consistent with previous
finding with 7- to 8-year-old children (Wang, Yamasaki, et al., 2021). According to the mem-
ory, unification, and control model by Hagoort (2016), regions in the temporal cortex subserve
knowledge representations that have been laid down in memory during acquisition. In con-
trast, frontal regions, which are structurally and functionally connected to temporal regions,
support memory retrieval, decomposition, and unification operations. The lack of a brain-
behavioral correlation may be because specialization in the frontal lobe provided children
beyond 7 to 8 years old with more flexibility in their division of labor in terms of engaging
the representation and control systems during specific language tasks. According to the
distributions-to-associations model by Shrager and Siegler (1998), children can either retrieve
an answer using their stored knowledge representation or adaptively utilize back-up strategies
when facing a problem (Siegler, 1988). Unlike 5- to 6-year-old children, who rely primarily on
knowledge representation stored in the temporal lobe (Weiss et al., 2018), children older than
7 to 8 years old might have access to more back-up strategies by engaging their frontal lobes.
The larger individual differences in the choice of strategies could lead to more difficulty finding
brain-behavior correlations in older children than in younger children. In addition, behavioral
performance on standardized tests is a byproduct of multiple cognitive processes, including
Neurobiology of Language
313
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
attention and working memory. However, brain specialization, as operationalized in the cur-
rent study as a double dissociation between tasks, reflects language-specific processes. Direct
task comparisons of brain activity between two similar tasks within a language mask allowed
us to cancel out irrelevant processes that were shared by the two tasks (e.g., attention, working
memory) and extract brain activity that is specific to language processing. This mismatch of the
underlying processes measured by behavioral performance and brain activity reduces the like-
lihood of finding brain-behavior correlations. Although we do not know how skill is related to
language specialization observed in 9- to 10-year-old children, as compared to younger chil-
dren (Wang, Yamasaki, et al., 2021; Weiss et al., 2018), older children clearly showed
increased specialization, which likely means that children’s brains become increasingly mod-
ularized into regions with unique computational principles. The interactive specialization
theory (Johnson, 2011) hypothesizes that brain interaction drives specialization. Thus, we
think that this increased specialization should be related to interactions across brain regions,
but this needs to be systematically investigated with future studies.
In summary, the current study showed phonological and semantic specialization in both
the temporal and frontal lobe in 9- to 10-year-old children, similar to that observed in 7- to
8-year-old children. However, different from 7- to 8-year-old children who seem to rely on a
general cognitive control region, 9- to 10-year-old children appear to rely on language-specific
regions to deal with more difficult language tasks. Overall, this study suggests that at 9 to
10 years old, children’s phonological and semantic specialization in the brain continues to
become more adult-like.
FUNDING INFORMATION
James R. Booth, National Institutes of Health (https://dx.doi.org/10.13039/100000002), Award
ID: DC013274.
AUTHOR CONTRIBUTIONS
Jin Wang: Conceptualization; Formal analysis; Writing – original draft; Writing – review &
editing. Brianna L. Yamasaki: Conceptualization; Writing – review & editing. James R. Booth:
Conceptualization; Funding acquisition; Writing – review & editing.
DATA AVAILABILITY STATEMENT
All data were pulled from a published data set on OpenNeuro.org (see detailed description in
Wang et al., 2022; https://openneuro.org/datasets/ds003604). The specific subjects and runs
used in the current study as well as the code used to analyze the data were shared on GitHub
https://github.com/wangjinvandy/PhonSem_Specialization_9_10.
REFERENCES
Anthony, J. L., & Francis, D. J. (2005). Development of phonological
awareness. Current Directions in Psychological Science, 14(5),
255–259. https://doi.org/10.1111/j.0963-7214.2005.00376.x
Ashburner, J., Barnes, G., Chen, C.-C., Daunizeau, J., Flandin, G.,
Friston, K., Gitelman, D., Glauche, V., Henson, R., Hutton, C.,
Jafarian, A., Kiebel, S., Kilner, J., Litvak, V., Mattout, J., Moran,
R., Penny, W., Phillips, C., Razi, A., … Zeidman, P. (2021).
SPM12 manual. Wellcome Centre for Human Neuroimaging.
Aylward, E. H., Park, J. E., Field, K. M., Parsons, A. C., Richards,
T. L., Cramer, S. C., & Meltzoff, A. N. (2005). Brain activation
during face perception: Evidence of a developmental change.
Journal of Cognitive Neuroscience, 17(2), 308–319. https://doi
.org/10.1162/0898929053124884, PubMed: 15811242
Baker, K., Carlson, H. L., Zewdie, E., & Kirton, A. (2020). Develop-
mental remodelling of the motor cortex in hemiparetic children
with perinatal stroke. Pediatric Neurology, 112, 34–43. https://doi
.org/10.1016/j.pediatrneurol.2020.08.004, PubMed: 32911261
Balota, D. A., Yap, M. J., Cortese, M. J., Hutchison, K. A., Kessler, B.,
Loftis, B., Neely, J. H., Nelson, D. J., Simpson, G. B., & Treiman, R.
(2007). The English lexicon project. Behavior Research Methods,
39(3), 445–459. https://doi.org/10.3758/BF03193014, PubMed:
17958156
Neurobiology of Language
314
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and meta-
analysis of 120 functional neuroimaging studies. Cerebral Cortex,
19(12), 2767–2796. https://doi.org/10.1093/cercor/ bhp055,
PubMed: 19329570
Boets, B., de Beeck, H. P. O., Vandermosten, M., Scott, S. K.,
Gillebert, C. R., Mantini, D., Bulthé, J., Sunaert, S., Wouters, J.,
& Ghesquière, P. (2013). Intact but less accessible phonetic
representations in adults with dyslexia. Science, 342(6163),
1251–1254. https://doi.org/10.1126/science.1244333, PubMed:
24311693
Burton, M. W., Small, S. L., & Blumstein, S. E. (2000). The role of
segmentation in phonological processing: An fMRI investigation.
Journal of Cognitive Neuroscience, 12(4), 679–690. https://doi
.org/10.1162/089892900562309, PubMed: 10936919
Carter, E. J., & Pelphrey, K. A. (2006). School-aged children exhibit
domain-specific responses to biological motion. Social Neuro-
s c i e n c e , 1 ( 3 – 4 ) , 3 9 6 – 4 11 . h t t p s : / / d o i . o r g / 1 0 . 1 0 8 0
/17470910601041382, PubMed: 18174911
Chiou, R., Humphreys, G. F., Jung, J., & Lambon Ralph, M. A.
(2018). Controlled semantic cognition relies upon dynamic and
flexible interactions between the executive “semantic control”
and hub-and-spoke “semantic representation” systems. Cortex,
103, 100–116. https://doi.org/10.1016/j.cortex.2018.02.018,
PubMed: 29604611
Collins, A. M., & Loftus, E. F. (1975). A spreading-activation theory
of semantic processing. Psychological Review, 82(6), 407–428.
https://doi.org/10.1037/0033-295X.82.6.407
Costafreda, S. G., Fu, C. H., Lee, L., Everitt, B., Brammer, M. J., &
David, A. S. (2006). A systematic review and quantitative
appraisal of fmri studies of verbal fluency: Role of the left inferior
frontal gyrus. Human Brain Mapping, 27(10), 799–810. https://
doi.org/10.1002/hbm.20221, PubMed: 16511886
Durston, S., Davidson, M. C., Tottenham, N., Galvan, A., Spicer, J.,
Fossella, J. A., & Casey, B. J. (2006). A shift from diffuse to focal
cortical activity with development. Developmental Science, 9(1),
1–8. https://doi.org/10.1111/j.1467-7687.2005.00454.x,
PubMed: 16445387
Enge, A., Rahman, R. A., & Skeide, M. A. (2021). A meta-analysis of
fMRI studies of semantic cognition in children. NeuroImage, 241,
Article 118436. https://doi.org/10.1016/j.neuroimage.2021
.118436, PubMed: 34329724
Friederici, A. D., & Gierhan, S. M. E. (2013). The language network.
Current Opinion in Neurobiology, 23(2), 250–254. https://doi.org
/10.1016/j.conb.2012.10.002, PubMed: 23146876
Gathers, A. D., Bhatt, R., Corbly, C. R., Farley, A. B., & Joseph, J. E.
(2004). Developmental shifts in cortical loci for face and object
recognition. Neuroreport, 15(10), 1549–1553. https://doi.org/10
.1097/01.wnr.0000133299.84901.86, PubMed: 15232281
Girard, C., Bastelica, T., Léone, J., Epinat-Duclos, J., Longo, L., &
Prado, J. (2022). Nurturing the mathematical brain: Home
numeracy practices are associated with children’s neural
responses to Arabic numerals. Psychological Science, 33(2),
196–211. https://doi.org/10.1177/09567976211034498,
PubMed: 35108141
Gu, C., & Bi, H.-Y. (2020). Auditory processing deficit in individuals
with dyslexia: A meta-analysis of mismatch negativity. Neurosci-
ence & Biobehavioral Reviews, 116, 396–405. https://doi.org/10
.1016/j.neubiorev.2020.06.032, PubMed: 32610180
Hagoort, P. (2016). MUC (memory, unification, control): A model
on the neurobiology of language beyond single word processing.
In Neurobiology of language (pp. 339–347). Academic Press.
https://doi.org/10.1016/B978-0-12-407794-2.00028-6
Hickok, G., & Poeppel, D. (2007). The cortical organization of
speech processing. Nature Reviews Neuroscience, 8(5), 393–402.
https://doi.org/10.1038/nrn2113, PubMed: 17431404
Hodgson, V. J., Lambon Ralph, M. A., & Jackson, R. L. (2021).
Multiple dimensions underlying the functional organisation
of the language network. NeuroImage, 241, Article 118444.
https://doi.org/10.1016/j.neuroimage.2021.118444, PubMed:
34343627
Johnson, M. H. (2011). Interactive specialization: A domain-general
framework for human functional brain development? Develop-
mental Cognitive Neuroscience, 1(1), 7–21. https://doi.org/10
.1016/j.dcn.2010.07.003, PubMed: 22436416
Karmiloff-Smith, A. (1998). Development itself is the key to under-
standing developmental disorders. Trends in Cognitive Sciences,
2(10), 389–398. https://doi.org/10.1016/S1364-6613(98)01230
-3, PubMed: 21227254
Karmiloff-Smith, A. (2015). An alternative to domain-general or
domain-specific frameworks for theorizing about human evo-
lution and ontogenesis. AIMS Neuroscience, 2(2), 91–104.
https://doi.org/10.3934/ Neuroscience.2015.2.91, PubMed:
26682283
Katzev, M., Tüscher, O., Hennig, J., Weiller, C., & Kaller, C. P.
(2013). Revisiting the functional specialization of left inferior
frontal gyrus in phonological and semantic fluency: The crucial
role of task demands and individual ability. Journal of Neurosci-
ence, 33(18), 7837–7845. https://doi.org/10.1523/JNEUROSCI
.3147-12.2013, PubMed: 23637175
Kaufman, A. S., & Kaufman, N. L. (2004). Kaufmann Brief Intelli-
gence Test (2nd ed.). Pearson Assessments.
Landi, N., Mencl, W. E., Frost, S. J., Sandak, R., & Pugh, K. R.
(2010). An fMRI study of multimodal semantic and phonological
processing in reading disabled adolescents. Annals of Dyslexia,
60(1), 102–121. https://doi.org/10.1007/s11881-009-0029-6,
PubMed: 20049657
Liu, L., Wang, W., You, W., Li, Y., Awati, N., Zhao, X., Booth, J. R.,
& Peng, D. (2012). Similar alterations in brain function for
phonological and semantic processing to visual characters in
Chinese dyslexia. Neuropsychologia, 50(9), 2224–2232. https://
doi.org/10.1016/j.neuropsychologia.2012.05.026, PubMed:
22698991
Mathur, A., Schultz, D., & Wang, Y. (2020). Neural bases of pho-
nological and semantic processing in early childhood. Brain
Connectivity, 10(5), 212–223. https://doi.org/10.1089/ brain
.2019.0728, PubMed: 32308014
Mayor, J., & Plunkett, K. (2010). A neurocomputational account of
taxonomic responding and fast mapping in early word learning.
Psychological Review, 117(1), 1–31. https://doi.org/10.1037
/a0018130, PubMed: 20063962
Mazaika, P. K., Hoeft, F., Glover, G. H., & Reiss, A. L. (2009).
Methods and software for fMRI analysis of clinical subjects. Neu-
roImage, 47(S1), S58. https://doi.org/10.1016/S1053-8119(09)
70238-1
Melby-Lervåg, M., Lyster, S. A. H., & Hulme, C. (2012). Phonolo-
gical skills and their role in learning to read: A meta-analytic
review. Psychological Bulletin, 138(2), 322–352. https://doi.org
/10.1037/a0026744, PubMed: 22250824
Mesgarani, N., Cheung, C., Johnson, K., & Chang, E. F. (2014).
Phonetic feature encoding in human superior temporal gyrus.
Science, 343(6174), 1006–1010. https://doi.org/10.1126
/science.1245994, PubMed: 24482117
Myers, E. B., Blumstein, S. E., Walsh, E., & Eliassen, J. (2009).
Inferior frontal regions underlie the perception of phonetic
category invariance. Psychological Science, 20(7), 895–903.
Neurobiology of Language
315
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
https://doi.org/10.1111/j.1467-9280.2009.02380.x, PubMed:
19515116
Namy, L. L. (2012). Getting specific: Early general mechanisms give
rise to domain-specific expertise in word learning. Language
Learning and Development, 8(1), 47–60. https://doi.org/10
.1080/15475441.2011.617235
Nelson, D. L., McEvoy, C. L., & Schreiber, T. A. (2004). The
University of South Florida free association, rhyme, and word
fragment norms. Behavior Research Methods, Instruments, & Com-
puters, 36(3), 402–407. https://doi.org/10.3758/ BF03195588,
PubMed: 15641430
Noordenbos, M. W., & Serniclaes, W. (2015). The categorical per-
ception deficit in dyslexia: A meta-analysis. Scientific Studies of
Reading, 19(5), 340–359. https://doi.org/10.1080/10888438
.2015.1052455
Novick, J. M., Trueswell, J. C., & Thompson-Schill, S. L. (2010).
Broca’s area and language processing: Evidence for the cognitive
control connection. Language and Linguistics Compass, 4(10),
906–924. https://doi.org/10.1111/j.1749-818X.2010.00244.x
Okada, K., Matchin, W., & Hickok, G. (2018). Phonological feature
repetition suppression in the left inferior frontal gyrus. Journal of
Cognitive Neuroscience, 30(10), 1549–1557. https://doi.org/10
.1162/jocn_a_01287, PubMed: 29877763
Poldrack, R. A., Wagner, A. D., Prull, M. W., Desmond, J. E.,
Glover, G. H., & Gabrieli, J. D. (1999). Functional specialization
for semantic and phonological processing in the left inferior pre-
frontal cortex. NeuroImage, 10(1), 15–35. https://doi.org/10.1006
/nimg.1999.0441, PubMed: 10385578
Rice, M. L., & Hoffman, L. (2015). Predicting vocabulary growth in
children with and without specific language impairment: A lon-
gitudinal study from 2;6 to 21 years of age. Journal of Speech,
Language, and Hearing Research, 58(2), 345–359. https://doi
.org/10.1044/2015_JSLHR-L-14-0150, PubMed: 25611623
Roux, F.-E., Dufor, O., Giussani, C., Wamain, Y., Draper, L.,
Longcamp, M., & Démonet, J.-F. (2009). The graphemic/motor
frontal area Exner’s area revisited. Annals of Neurology, 66(4),
537–545. https://doi.org/10.1002/ana.21804, PubMed:
19847902
Samuelson, L. K., & McMurray, B. (2017). What does it take to
learn a word? Wiley Interdisciplinary Reviews: Cognitive Science,
8(1–2), Article e1421. https://doi.org/10.1002/wcs.1421,
PubMed: 27911490
Schlaggar, B. L., & McCandliss, B. D. (2007). Development of neu-
ral systems for reading. Annual Review of Neuroscience, 30(1),
475–503. https://doi.org/10.1146/annurev.neuro.28.061604
.135645, PubMed: 17600524
Seymour, H. N., Roeper, T. W., & De Villiers, J. (2003). Diagnostic
Evaluation of Language Variation: Screening test. NCS Pearson.
Shrager, J., & Siegler, R. S. (1998). SCADS: A model of children’s
strategy choices and strategy discoveries. Psychological Science,
9(5), 405–410. https://doi.org/10.1111/1467-9280.00076
Siegler, R. S. (1988). Individual differences in strategy choices:
Good students, not-so-good students, and perfectionists. Child
Development, 59(4), 833–851. https://doi.org/10.2307
/1130252, PubMed: 3168623
Skeide, M. A., Brauer, J., & Friederici, A. D. (2014). Syntax gradu-
ally segregates from semantics in the developing brain. Neuro-
Image, 100, 106–111. https://doi.org/10.1016/j.neuroimage
.2014.05.080, PubMed: 24927987
Skeide, M. A., & Friederici, A. D. (2016). The ontogeny of the cor-
tical language network. Nature Reviews Neuroscience, 17(5),
323–332. https://doi.org/10.1038/nrn.2016.23, PubMed:
27040907
Sloutsky, V. M. (2010). Mechanisms of cognitive development:
Domain-general learning or domain-specific constraints? Cogni-
tive Science, 34(7), 1125–1130. https://doi.org/10.1111/j.1551
-6709.2010.01132.x, PubMed: 21564246
Snowling, M. J., & Melby-Lervåg, M. (2016). Oral language deficits
in familial dyslexia: A meta-analysis and review. Psychological
Bulletin, 142(5), 498–545. https://doi.org/10.1037/bul0000037,
PubMed: 26727308
Swart, N. M., Muijselaar, M. M., Steenbeek-Planting, E. G., Droop,
M., de Jong, P. F., & Verhoeven, L. (2017). Differential lexical pre-
dictors of reading comprehension in fourth graders. Reading and
Writing, 30(3), 489–507. https://doi.org/10.1007/s11145-016
-9686-0, PubMed: 28255199
Unger, L., & Fisher, A. V. (2021). The emergence of richly organized
semantic knowledge from simple statistics: A synthetic review.
Developmental Review, 60, Article 100949. https://doi.org/10
.1016/j.dr.2021.100949, PubMed: 33840880
Universitätsklinikum Tübingen. 2022. Klinik fur Kinder- und Jegen-
dmedizin [software and parameters]. https://www.medizin.uni
-tuebingen.de/kinder/en/research/neuroimaging/software
Vitevitch, M. S., & Luce, P. A. (2004). A web-based interface to
calculate phonotactic probability for words and nonwords in
English. Behavior Research Methods, Instruments, & Computers,
36(3), 481–487. https://doi.org/10.3758/BF03195594, PubMed:
15641436
Wagner, R. K., Torgesen, J. K., Rashotte, C. A., Hecht, S. A., Barker,
T. A., Burgess, S. R., Donahue, J., & Garon, T. (1997). Changing
relations between phonological processing abilities and
word-level reading as children develop from beginning to skilled
readers: A 5-year longitudinal study. Developmental Psychology,
33(3), 468–479. https://doi.org/10.1037/0012-1649.33.3.468,
PubMed: 9149925
Wagner, R. K., Torgesen, J. K., Rashotte, C. A., & Pearson, N. A.
(2013). Comprehensive test of phonological processing (2nd ed.).
Pro-Ed. https://doi.org/10.1037/t52630-000
Wang, J., Joanisse, M. F., & Booth, J. R. (2020). Neural representa-
tions of phonology in temporal cortex scaffold longitudinal
reading gains in 5- to 7-year-old children. NeuroImage, 207,
Article 116359. https://doi.org/10.1016/j.neuroimage.2019
.116359, PubMed: 31733372
Wang, J., Joanisse, M. F., & Booth, J. R. (2021). Letter fluency in
7–8-year-old children is related to the anterior, but not poste-
rior, ventral occipito-temporal cortex during an auditory pho-
nological task. Developmental Cognitive Neuroscience, 47,
Article 100898. https://doi.org/10.1016/j.dcn.2020.100898,
PubMed: 33341533
Wang, J., Lytle, M. N., Weiss, Y., Yamasaki, B. L., & Booth, J. R.
(2022). A longitudinal neuroimaging dataset on language pro-
cessing in children ages 5, 7, and 9 years old. Scientific Data,
9(1), Article 4. https://doi.org/10.1038/s41597-021-01106-3,
PubMed: 35013348
Wang, J., Pines, J., Joanisse, M., & Booth, J. R. (2021). Reciprocal
relations between reading skill and the neural basis of phonolo-
gical awareness in 7- to 9-year-old children. NeuroImage, 236,
Article 118083. https://doi.org/10.1016/j.neuroimage.2021
.118083, PubMed: 33878381
Wang, J., Yamasaki, B. L., Weiss, Y., & Booth, J. R. (2021). Both fron-
tal and temporal cortex exhibit phonological and semantic spe-
cialization during spoken language processing in 7- to 8-year-old
children. Human Brain Mapping, 42(11), 3534–3546. https://doi
.org/10.1002/hbm.25450, PubMed: 33951259
Weiss, Y., Cweigenberg, H. G., & Booth, J. R. (2018). Neural spe-
cialization of phonological and semantic processing in young
Neurobiology of Language
316
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Phonological and semantic specialization in 9- to 10-year-olds
children. Human Brain Mapping, 39(11), 4334–4348. https://doi
.org/10.1002/hbm.24274, PubMed: 29956400
Wiig, E. H., Secord, W. A., & Semel, E. (2013). Clinical Evaluation
of Language Fundamentals: CELF-5. Pearson.
Wilke, M., Altaye, M., Holland, S. K., & CMIND Authorship
Consortium. (2017). CerebroMatic: A versatile toolbox for
spline-based MRI template creation. Frontiers in Computational
Neuroscience, 11, Article 5. https://doi.org/10.3389/fncom.2017
.00005, PubMed: 28275348
Xie, X., & Myers, E. (2018). Left inferior frontal gyrus sensitivity to
phonetic competition in receptive language processing: A com-
parison of clear and conversational speech. Journal of Cognitive
Neuroscience, 30(3), 267–280. https://doi.org/10.1162/jocn_a
_01208, PubMed: 29160743
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
2
9
7
2
0
7
9
0
0
9
n
o
_
a
_
0
0
0
9
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
317