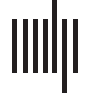RESEARCH ARTICLE
Event-Related Potential Correlates of Learning
to Produce Novel Foreign Phonemes
Henry Railo1,2
, Anni Varjonen2, Minna Lehtonen1,2,3, and Pilleriin Sikka1,2,4,5
1Department of Psychology and Speech-Language Pathology, University of Turku, Turku, Finland
2Turku Brain and Mind Centre, University of Turku, Turku, Finland
3Center for Multilingualism in Society across the Lifespan, Department of Linguistics and Scandinavian Studies,
University of Oslo, Oslo, Norway
4Department of Cognitive Neuroscience and Philosophy, School of Bioscience, University of Skövde, Skövde, Sweden
5Department of Psychology, Stanford University, Stanford, California, USA
Keywords: speaking induced suppression, event-related potential (ERP), phoneme learning
ABSTRACT
Learning to pronounce a foreign phoneme requires an individual to acquire a motor program
that enables the reproduction of the new acoustic target sound. This process is largely based
on the use of auditory feedback to detect pronunciation errors to adjust vocalization. While
early auditory evoked neural activity underlies automatic detection and adaptation to
vocalization errors, little is known about the neural correlates of acquiring novel speech
targets. To investigate the neural processes that mediate the learning of foreign phoneme
pronunciation, we recorded event-related potentials when participants (N = 19) pronounced
native or foreign phonemes. Behavioral results indicated that the participants’ pronunciation of
the foreign phoneme improved during the experiment. Early auditory responses (N1 and P2
waves, approximately 85–290 ms after the sound onset) revealed no differences between
foreign and native phonemes. In contrast, the amplitude of the frontocentrally distributed late
slow wave (LSW, 320–440 ms) was modulated by the pronunciation of the foreign phonemes,
and the effect changed during the experiment, paralleling the improvement in pronunciation.
These results suggest that the LSW may reflect higher-order monitoring processes that signal
successful pronunciation and help learn novel phonemes.
INTRODUCTION
When learning to pronounce foreign phonemes, the individual needs to evaluate how well the
sound they produced matches the target phoneme and, in case of a mismatch, attempt to
improve their phonation. Such learning may be based on fast and automatic unconscious
speech control processes, but it may also depend on later processes that are associated with
the conscious evaluation of phonation. However, relatively little is known about the neural
mechanisms that underlie the learning of foreign phoneme production. In this study, by com-
paring electroencephalographic (EEG) activity evoked by self-produced and passively heard
foreign and native phonemes, we investigate which auditory evoked potentials correlate with
learning to pronounce foreign phonemes.
Previous research on the neural processing of foreign phonemes has focused on examining
changes in auditory evoked activity elicited by passively heard sounds. Studies employing the
oddball paradigm have shown that foreign phonemes elicit weaker mismatch negativity
a n o p e n a c c e s s
j o u r n a l
Citation: Railo, H., Varjonen, A.,
Lehtonen, M., & Sikka, P. (2022). Event-
related potential correlates of learning
to produce novel foreign phonemes.
Neurobiology of Language, 3(4),
599–614. https://doi.org/10.1162
/nol_a_00080
DOI:
https://doi.org/10.1162/nol_a_00080
Received: 18 January 2022
Accepted: 31 August 2022
Competing interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Henry Railo
hmrail@utu.fi
Handling Editor:
Patrick Wong
Copyright: © 2022
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
Phoneme:
A distinct sound unit that can
distinguish one word from another in
a particular language.
Electroencephalography (EEG):
A method for recording brain-related
and other electrical activity from the
scalp.
Event-related potential (ERP):
EEG response produced by a specific
event, such as a sound.
Late slow wave (LSW):
A specific ERP wave with a relatively
late timing (>300 ms after stimulus
onset).
Speaking-induced suppression (SIS):
Attenuation of neural responses to
self-vocalized sounds relative to
passively heard sounds.
responses than native phonemes (Díaz et al., 2008; Näätänen et al., 1997; Peltola et al., 2003),
indicating a poorer ability to discriminate foreign phonemes. Training improves the discrimination
of phonemes, and this is reflected in the mismatch response (Tamminen et al., 2015; Tremblay
et al., 1998). Phoneme discrimination training also increases the amplitude of early (N1/P2)
auditory event-related potentials (ERPs), which has been interpreted as training-induced neuro-
plastic changes in the auditory cortex (Alain et al., 2007; Reinke et al., 2003; Saloranta et al.,
2020). Additionally, studies have shown that the amplitude of a late slow wave (LSW; 300–
500 ms) to passively heard phonemes correlates with phonemic learning (Alain et al., 2007;
Reinke et al., 2003; Saloranta et al., 2020), but the functional role of this correlate is unclear.
Learning to discriminate the acoustic features of passively heard foreign phonemes is
clearly important for learning to produce the phoneme. However, to accurately vocalize novel
phonemes one needs to also learn the motor commands that enable reproduction of the
desired acoustic features. The process of learning to produce novel phonemes involves track-
ing the auditory feedback of one’s own vocalizations to adjust future pronunciation. Studies in
which speech is artificially altered (e.g., by shifting pitch) indicate that individuals can adjust
their speech based on auditory feedback in just 100–150 ms (Hain et al., 2000; Houde & Jordan,
1998). This type of rapid feedback control relies on “efferent copies” of motor commands that
the motor system relays to the auditory cortex to compare how well the produced speech
matches the targeted motor commands (Hickok et al., 2011; Houde & Nagarajan, 2011;
Tourville & Guenther, 2011). Because of this interplay between motor and sensory systems,
the neural processing of passively heard and self-produced sounds differs significantly (Curio
et al., 2000; Houde et al., 2002). Therefore, to understand how individuals learn novel phone-
mic targets, it is crucial to examine electrophysiological responses to self-produced sounds.
The electrophysiological correlate of the feedback control of self-produced speech is the
suppression of auditory evoked activity. Auditory ERPs in response to self-produced sounds
are suppressed in the N1 (100–200 ms) and the P2 (200–300 ms) time windows, as compared
to when the same sounds are passively heard (Behroozmand et al., 2011; Curio et al., 2000;
Heinks-Maldonado et al., 2005; Houde et al., 2002; Railo et al., 2020). This phenomenon is
known as the speaking-induced suppression (SIS). Using magnetoencephalography, Niziolek
et al. (2013) observed that the amplitude of SIS (i.e., amplitude difference between self-
produced and passively heard sounds) tracked variation in phonation: Auditory responses to
phonemes that deviated from prototypical phonemes produced a decreased SIS, and the size
of SIS predicted corrections to vocalization. Similarly, when speech is artificially altered, SIS
decreases, suggesting that the brain detects a mismatch between feedforward motor com-
mands and heard speech (Behroozmand et al., 2009; Behroozmand & Larson, 2011; Chang
et al., 2013).
In addition to enabling online adjustments to pronunciation, such a feedback control mech-
anism may contribute to the process of learning to correctly pronounce foreign phonemes: the
individual learns to better produce the desired sound based on errors in previous vocaliza-
tions. A similar mechanism is assumed to contribute to phonemic learning in children during
speech acquisition. After learning the target sound, the motor commands used to produce the
sound are fine-tuned based on feedback control (Tourville & Guenther, 2011). However, we
are not aware of any studies that have examined whether SIS amplitude tracks the process of
learning to produce a novel foreign phoneme. Many studies have investigated sensorimotor
adaptation of speech to altered auditory feedback, but such learning effects are relatively
short-lived and represent changes to motor programs the individual already knows (e.g.,
Houde & Jordan, 1998; Lametti et al., 2018). In contrast, when learning a novel phoneme,
the individual needs to learn completely novel sensorimotor speech targets.
Neurobiology of Language
600
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
Online adjustments to speech are typically considered rapid low-level processes, but the
processes of learning to pronounce foreign phonemes may also depend on higher-order pro-
cesses that take more time to unfold. SIS typically refers to effects observed in the N1 and P2
time windows, but the comparison of actively spoken and passively heard phonemes some-
times also reveals differences in later time windows where a LSW (300–500 ms) is often
observed in speech tasks. Differences in this late time window are often neglected, likely
because they are too late to reflect automatic feedback speech control, but also because there
is no theoretical framework within which to interpret these findings. Whereas the classical
SIS time windows (N1, P2) likely corresponds to the automatic preconscious evaluation of
self-produced speech, the later time windows may correspond to attentive evaluation of
pronunciation. Because the attentive evaluation of self-produced speech is likely to contribute
to phonemic learning, and because earlier studies suggest that LSW may correlate with pho-
neme discrimination learning (Alain et al., 2007; Reinke et al., 2003; Saloranta et al., 2020), it
is important to investigate not only the earlier time windows (N1 and P2) but also the LSW.
Here, we recorded auditory ERPs when Finnish participants reproduced a native /ö/ phoneme
or a foreign Estonian /õ/ phoneme (Speak condition) after hearing the target spoken by a native
speaker. In a control condition, the participants listened to a recording of the phoneme they had
just spoken (Listen condition). Comparison of the Speak and Listen conditions corresponds to the
SIS, and it also allows us to test whether possible associations between pronunciation and ERP
amplitudes reflect active vocalization or the passive processing of acoustic information. Assum-
ing that participants are not able to produce the foreign phoneme as accurately as the native
phoneme, we expected to observe a smaller suppression of N1 amplitudes to self-produced for-
eign phonemes, as compared to passively heard phonemes (i.e., reduced SIS, indicating that the
auditory system detects a mismatch between produced and attempted sounds). Furthermore,
assuming that participants learn to better pronounce the foreign phoneme during the experi-
ment, we analyzed the trial-by-trial changes in ERP amplitudes in three time windows (N1,
P2, and LSW) to test whether changes in ERP amplitudes track improvements in pronunciation.
MATERIALS AND METHODS
Participants
Twenty-one participants (students at the University of Turku) volunteered for this study. All
participants were Finnish, with normal hearing and with no diagnosed learning disabilities
or neurological disorders. All participants were monolingual and reported no previous expe-
rience in learning Estonian. Two participants were excluded from statistical analyses due to
excessive noise in EEG. Thus, the final sample included 19 participants (range 18–35 years;
17 females, 2 males). The study was conducted according to the principles of the Declaration
of Helsinki. All participants provided informed consent to participate in the study. The exper-
iment was approved by the Ethics Committee for Human Sciences at the University of Turku.
Stimuli and Procedure
An overview of a single experimental trial is presented in Figure 1A. Participants heard a
recording of the Estonian phoneme /õ/, or the Finnish phoneme /ö/, in a random order. We
call this the Cue stimulus condition. The phoneme /õ/ is not part of the Finnish phonological
system and was therefore unfamiliar to the subjects. Acoustically, it resembles the Finnish pho-
neme /ö/ (e.g., Näätänen et al., 1997). After hearing the Cue phoneme, participants attempted
to repeat it as well as possible (Speak condition). After repeating the sound, they heard a play-
back of their produced phoneme (Listen condition). The stimuli were separated by
Neurobiology of Language
601
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
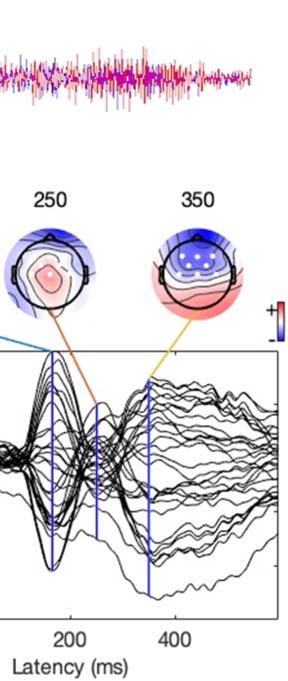
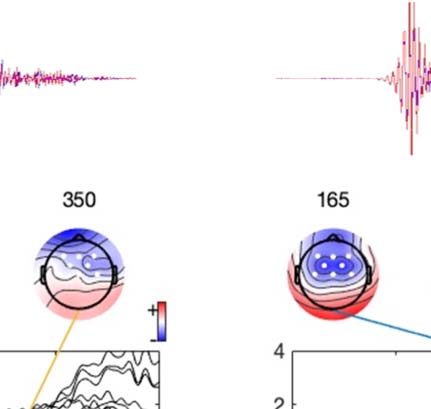
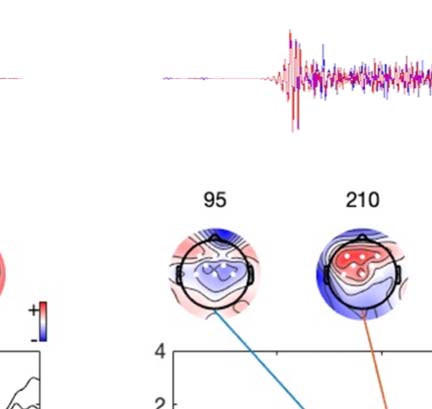
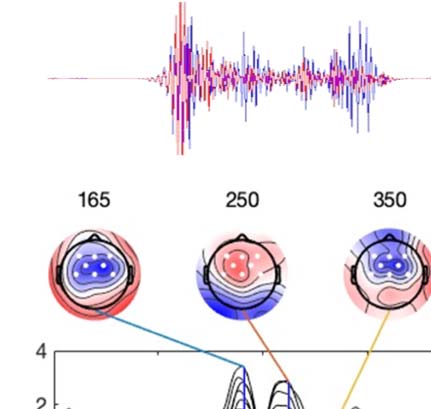
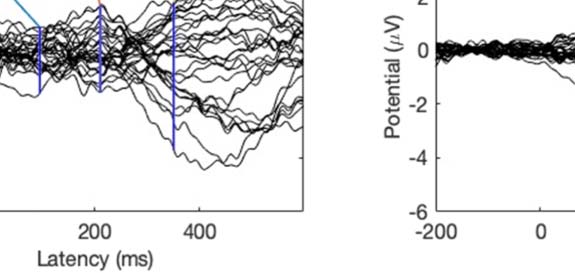
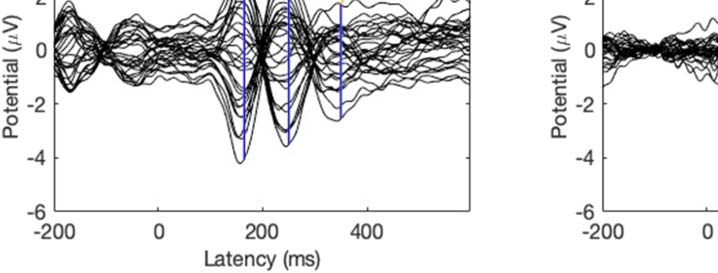
Experimental procedure and ERPs. (A) An overview of a single experimental trial. (B) Auditory signal recorded with the microphone.
Figure 1.
Average across all participants; blue line = foreign phoneme, red = native phoneme. (C) Butterfly ERP plots and scalp maps at three latencies.
Scalp color map ranges from −4 (blue) to 4 μVolt (red). White electrode markers indicate the cluster of electrodes on which the statistical
analysis was performed.
approximately a 2–3 s interval. Intertrial interval was about 4 s. This process was repeated 50
times in a block, and the experiment consisted of five blocks (250 repetitions in total during the
experiment). There was a 2–5 min break between blocks.
Cue stimuli (/ö/ and /õ/) were recorded by the same native Estonian (female) as both pho-
nemes are part of the Estonian language. The two phonemes were approximately the same
amplitude, pitch, and duration (500 ms). To examine how Finnish individuals hear the stimuli,
we asked 21 participants (age range 20–50, 13 females; different participants than those who
took part in the main study reported in this article) to listen to the stimuli, to identify the
phoneme, and to rate how confident (scale 1–4) they were in their response. Twenty out of
the 21 participants correctly identified the native phoneme as /ö/ with high confidence (M =
3.33, SD = 0.86). None of the participants correctly identified the Estonian /õ/ phoneme: Four-
teen participants identified it as Finnish /ö/ (confidence M = 3.14, SD = 0.77), two participants
identified the phoneme as Finnish /y/ (M = 2.0, SD = 0), two as the Russian ы (M = 3.50, 0.70),
two as the Swedish å (M = 4, SD = 0), and one participant believed the phoneme corre-
sponded to the pronunciation of the German ü (confidence = 4). In sum, this result shows, that
Finnish individuals recognize the native phoneme, but misidentify the Estonian phoneme.
The Cue and Listen stimuli were played to the participants from two TEAC LS-X8 speakers
placed about 1 meter to left and right of the participant. Participants’ pronunciations were
recorded using a Trust GXT 242 Lance microphone and saved in wave file format.
EEG Recording
EEG was recorded with 32 passive electrodes (EASYCAP, 2022) placed according to the 10–10
electrode system (Nuwer et al., 1998). Surface electromyograms were measured with two elec-
trodes above and below the lips, and below and to the side of the right eye. Reference elec-
trode was placed on the nose. Ground electrode was placed on the forehead. EEG was
recorded with a NeurOne Tesla amplifier using Version 1.4.1.64 software (Bittium, 2022).
Neurobiology of Language
602
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
Sampling rate was 500 Hz. In addition, the auditory stimuli (Cue, Speak, and Listen conditions)
were recorded as EEG signals using a microphone. This allowed us to accurately mark the
onset times of auditory stimuli on the EEG.
EEG Preprocessing
EEG was processed using EEGLAB Version 14.1.1 software (Delorme & Makeig, 2004) in
MathWorks’s Matlab 2014b. First, we high-pass filtered the microphone signals recorded with
EEG at 100 Hz (to remove noise but keep the sound signal and its transient onset) and used it
to add markers of the onsets of the stimuli on the continuous EEG data. The onset of auditory
stimuli was determined automatically as follows. The microphone signal had to remain above
a specified amplitude threshold for 10 consecutive samples, and then a marker was added to
the sample where the threshold was first crossed. As shown in Figure 1B, this procedure
yielded accurate estimates of phoneme onset times in all three experimental conditions. After
this, the microphone channels were removed from the data.
We rejected channels containing artifacts using EEGLAB’s pop_rejchan function based on
kurtosis, spectrum, and probability measures. Then, data were 1 Hz high-pass filtered (Klug &
Gramann, 2020) using pop_eegfiltnew function, and 50 Hz line noise was reduced using the
ZapLine plugin (de Cheveigné, 2020). We used artifact substance reconstruction to clean con-
tinuous EEG using a cutoff parameter at 20 (Chang et al., 2020). We then average-referenced
the data and ran independent component analysis (ICA) using the extended infomax algo-
rithm. After the ICA, we used the DIPFIT plug-in (SCCN, 2019) for localizing equivalent dipole
locations of the independent components. The rejection threshold was set at 100 (no dipoles
were rejected) and two dipoles constrain in symmetry. We used IClabel to automatically cat-
egorize components into brain-based and various non–brain-based categories (Pion-Tonachini
et al., 2019). Components with residual variance <15%, and >70% probability that the com-
ponent is brain based, were not considered artifactual (i.e., other components were removed).
After this, the removed channels were interpolated.
Next, the data were low-pass filtered at 40 Hz and cut into segments starting 200 ms before
stimulus onset and ending 600 ms after stimulus onset. Artifactual trials were removed using the
pop_jointprob function (local and global thresholds = 3). The average number of trials per par-
ticipant in the Finnish Cue condition was 97 (median = 99.5, SD = 9.1) and in the Estonian Cue
condition 109 (median = 112, SD = 10.1). The average number of trials in the Finnish Speak
condition was 75 (median = 78, SD = 21.0) and in the Estonian Speak condition 88 (median =
99.5, SD = 28.3). In the Finnish Listen condition, the average trial number was 85 (median = 87,
SD = 14.4) and in the Estonian Listen condition 95 (median = 98.5, SD = 16.4).
Statistical Analyses
We used mixed-effects linear regression analysis to test if Condition (Speak vs. Listen) and
Phoneme (Native /ö/ vs. Foreign /õ/) factors influenced ERPs at N1, P2, and LSW time windows
in single-trial data. The benefit of mixed-effects models is that analysis can be performed on
single-trial data while taking into account variation between participants. The analyses were
performed on afrontal-central electrode cluster (average of amplitudes across electrodes F3,
Fz, F4, FC1, FC2, C3, Cz, and C4, shown in Figure 1C). The analysis was run in Matlab
2014b. The Listen condition and Native phoneme were set as reference categories in the regres-
sion models (i.e., intercept is the Native phoneme in the Listen condition). In addition, trial
number was included in the model as a continuous regressor, because we were interested in
examining if ERP amplitudes changed as the experiment progressed (possibly due to learning).
Neurobiology of Language
603
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
Each condition had its own running trial number (e.g., trial number 10 in Speak/Foreign indi-
cated trial 10 in this specific condition). The trial number regressor was z scored (i.e., intercept of
the model represents responses around experiment midpoint). The model included all the three
predictors and their interactions as fixed-effects regressors (i.e., Condition × Phoneme × Trial
number). The model included the intercept and Speak/Listen conditions as participant-wise ran-
dom effects because more complex random effect structures (based on Akaike and Bayesian
information criteria) led to inferior models. Separate models were fitted for each ERP component
(N1, P2, and LSW, as described below), and the models were pruned by removing outliers.
To examine participants’ ability to pronounce the foreign phoneme, recordings of the par-
ticipants’ pronunciation were rated by two native Estonians (one of the raters was co-author
P. S.). During the rating procedure, the recordings were presented in random order, one partic-
ipant at a time. Participants’ pronunciation was rated on a scale from 1 to 4. Rating 1 denoted
vocalizations that did not resemble /õ/ at all (e.g., the participant vocalized the phoneme /ö/).
Rating 2 denoted poor vocalizations that had a little resemblance to /õ/, but the produced pho-
neme still clearly deviated from the target. Rating 3 denoted good vocalizations that clearly
resembled /õ/ without capturing the sound perfectly. Rating 4 corresponded to an excellent
pronunciation of /õ/ (i.e., the pronunciation resembled a native speaker’s pronunciation of the
phoneme). After examining interrater correlations (Figure 2C), the two ratings were averaged for
statistical analyses. Pronunciation of native phonemes was not rated because, being part of
Finnish phonetics, we assumed that participants have no difficulties in pronouncing these.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
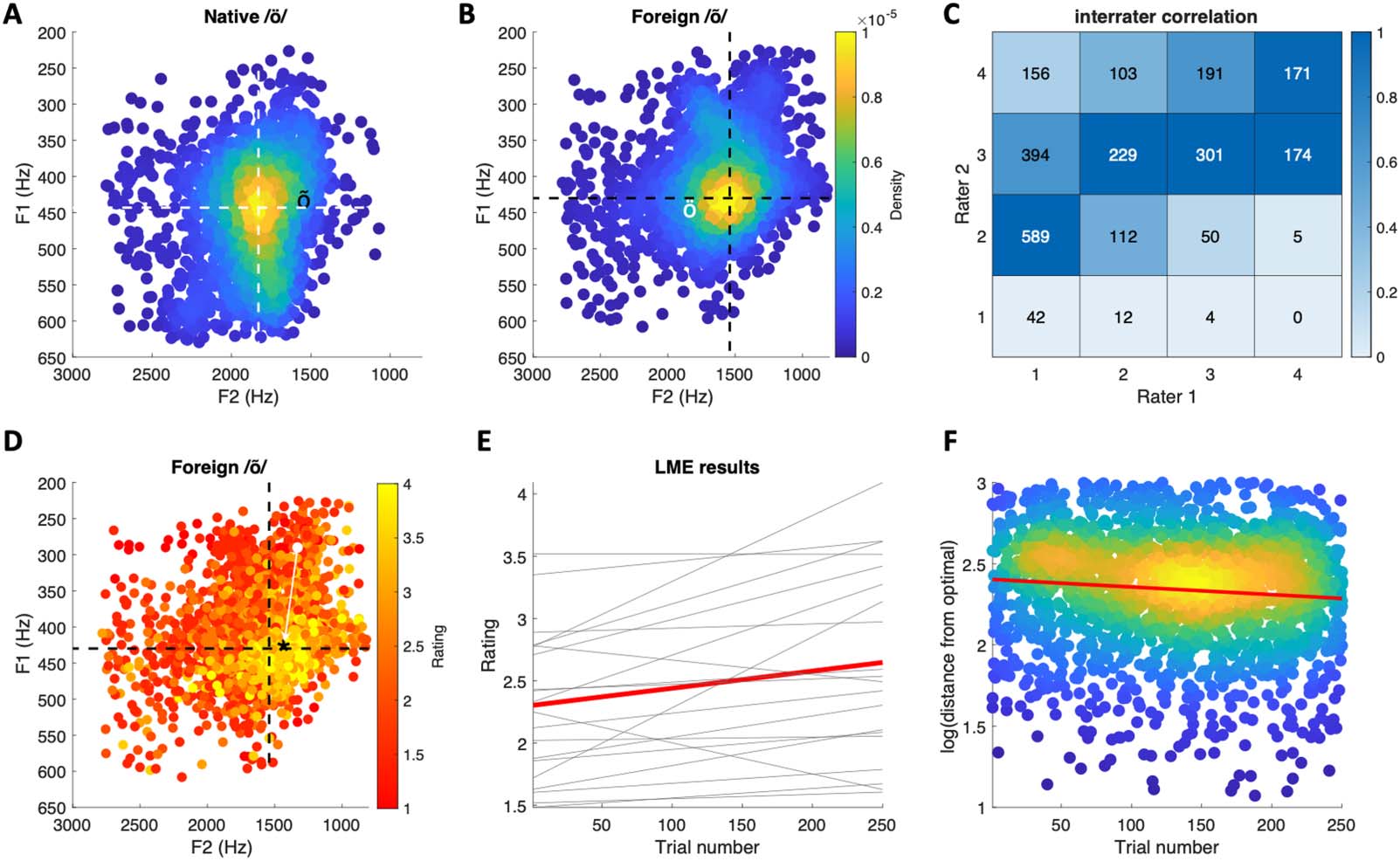
Figure 2. Behavioral results. (A) Formant plot of single-trial estimates of native phoneme /ö/. The white dashed lines indicate the center of mass
of the formant distribution. The black ‘õ’ symbol indicates the center of mass of the foreign phoneme. (B) Formant plot of single-trial estimates of
foreign phoneme /õ/. The black dashed lines indicate the center of mass of the formant distribution. The white ‘ö’ symbol indicates the center of
mass of the native phoneme. Color code indicates the density of data points. (C) Interrater correlation in phoneme ratings. Color indicates
proportion of ratings (scaled per column). (D) Phoneme plot of foreign /õ/ pronunciation color coded by rating. The ‘*’ symbol indicates the
center of mass of vocalizations with the highest rating, and the arrow indicates the distance of a single observation from this point. (E) Results of
the linear mixed-effects model showing that ratings increased as the function of trials. The red line is the group average, and light lines indicate
different participants’ results. (F) As trial number increases, acoustic features of the foreign phoneme approach the target. The red line depicts
the mixed-effects regression results (group average). Scatter density plots were made using the kernal smoothing function by Nils (2021).
Neurobiology of Language
604
EEG correlates of foreign phoneme learning
Formant:
Broad peak in the frequency
spectrum of a speech sound.
The aim of the ratings was to obtain a numerical index reflecting the external validity of
pronunciation. To verify that the ratings reflect acoustic parameters of the target phoneme,
we examined how well the frequency of the two first formants of participants’ vocalizations
predicted the (averaged) ratings. The first and second formats of the participants’ single-trial
vocalizations were estimated from the wave files using linear predictive coding in Matlab.
RESULTS
Behavioral Results
Figure 2A and 2B show phoneme plots of participants’ native and foreign phoneme pronun-
ciations, respectively. Consistent with previous reports, pronunciation of the two phonemes
differ especially in F2 (e.g., Asu & Teras, 2009; Näätänen et al., 1997). The ratings of the
two native Estonian speakers moderately correlated with each other (rho = 0.54; linearly
weighed Cohen’s kappa = 0.19; Figure 2C) but suggested that the participants had, in general,
difficulties in pronouncing the foreign phoneme (mean rating = 2.46, SD = 0.81). To verify that
the ratings captured systematic variation in the acoustics of the foreign phoneme, we used a
linear mixed-effects model to examine how well the F1 and F2 frequencies predicted the
ratings. The model included F1 and F2 frequencies and their interaction as fixed-effects pre-
dictors, and the same regressors and the intercept were included as by-participant random
effects variables. The results indicated that F1 frequency (t = 1.97, p = 0.04), F2 frequency
(t = 4.06, p < 0.001), and their interaction (t = 2.83, p = 0.004) predicted roughly 67%
(adjusted R2) of variation in the ratings.
An additional linear mixed-effects model indicated that, on average, ratings of participants’
pronunciation of the foreign phoneme improved as a function of trials (β = 0.0014, t = 2.87,
p = 0.004; the model included random, by-participant variation in the intercept and trial
number regressor). This result is visualized in Figure 2E, which shows the model results for
individual participants (thin gray lines) in addition to the group average (red line). Finally, to
verify that the increase in ratings as a function of trials reflected true changes in the acoustic
features of pronunciation, we calculated the Euclidean distance of single-trial foreign pho-
neme vocalizations from the “optimal” F1 and F2 formant combination (see white arrow in
Figure 2D for visualization). The Linear mixed-effects model with trial number as a predictor
indicated that formants approached “optimal” as a function of trial (t = −2.60; random-effect
structure included by-participant intercept and trial number).
ERP Results
Butterfly plots of the ERPs in different experimental conditions are shown in Figure 1C. In both
the Cue and Listen conditions, prominent N1 (negative peak at 160 ms) and P2 (positive peak
at 250 ms) waves were observed. In addition, the LSW was observed around 350–500 ms after
stimulus onset. As expected, the amplitude of the ERPs was suppressed in the Speak condition
relative to the Listen condition, indicating SIS. In addition, the peak latency of the N1 and P2
waves were earlier and the scalp topography somewhat different in the Speak condition (as
compared to the Listen and Cue conditions).
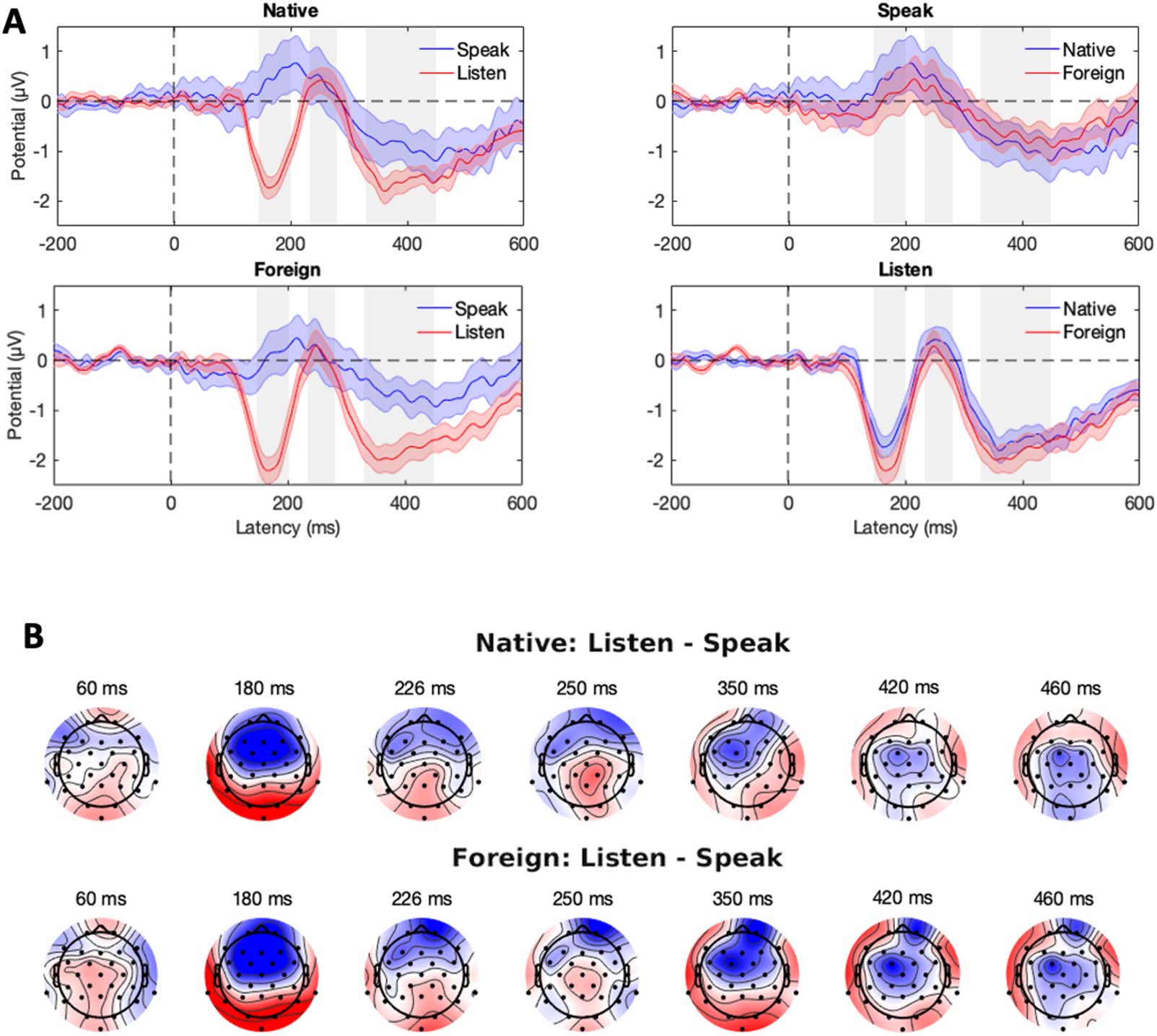
The critical comparison between the Listen and Speak conditions is visualized in Figure 3.
Scalp distributions of the difference between the Listen and Speak conditions—reflecting
SIS—are shown in Figure 3B, separately for the Native and Foreign phoneme conditions.
We statistically analyzed how the experimental manipulations influenced the N1 (150–200 ms),
P2 (230–290 ms), and LSW (320–440 ms) amplitudes. Based on the scalp maps of prominent
Neurobiology of Language
605
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Speaking-induced suppression (SIS). (A) Grand-average ERPs on the frontal-central electrode cluster (F3, Fz, F4, FC1, FC2, C3, Cz,
Figure 3.
and C4). The shaded area is the standard error of the mean. The gray rectangles indicate the N1, P2, and LSW time windows, respectively. (B)
Scalp maps show the SIS separately for the native and foreign phoneme conditions. Color bar ranges from −2 (blue) to 2 μV (red).
waves (see Figure 1C), statistical analyses were performed on the frontal-central electrode
cluster (F3, Fz, F4, FC1, FC2, C3, Cz, C4).
N1 Amplitudes
The results of the linear mixed-effects regression analyses on N1 amplitudes are presented in
Table 1. The intercept represents the average N1 amplitude in the Listen/Native condition. The
effect of Trial shows that the amplitude does not change statistically significantly (p = 0.17) as
a function of trial. The main effect of the Speak condition indicates that, on average, ampli-
tudes were 1.94 μV more positive in the Speak condition than in the Listen condition (p <
0.001), reflecting the SIS. This effect did not change statistically significantly during the
experiment (Trial:Speak interaction, p = 0.67). N1 amplitudes to foreign phonemes were,
on average, amplified by 0.38 μV (p < 0.001). The lack of Speak:Foreign interaction (p =
0.63) indicates that the SIS did not differ between the Foreign and Native conditions. Finally,
the lack of a three-way Trial:Speak:Foreign interaction indicates that the SIS in the Foreign
condition was not modulated by trial number (p = 0.88). The results of the model did not
change markedly if the model was pruned by removing the (largely redundant) Trial regressor.
Neurobiology of Language
606
EEG correlates of foreign phoneme learning
Table 1.
Results of the mixed-effects regression on N1 amplitudes (df = 6157)
Name
Intercept
Trial
Speak
Foreign
Trial:Speak
Trial:Foreign
Speak:Foreign
Trial:Speak:Foreign
Estimate
−1.54
0.12
1.94
−0.38
0.06
−0.19
0.09
−0.03
SE
0.19
0.09
0.45
0.12
0.13
0.12
0.18
0.18
t
−7.98
1.37
4.30
−3.21
0.43
−1.62
0.49
−0.15
p
<0.001
0.17
<0.001
<0.001
0.67
0.11
0.63
0.88
Lower
95% CI
−1.92
Upper
95% CI
−1.16
−0.05
1.06
−0.62
−0.20
−0.43
−0.26
−0.37
0.29
2.83
−0.15
0.31
0.04
0.43
0.32
Because the latency of the N1 wave peaked earlier in the Speak than in the Listen condi-
tion, we repeated the analysis with the N1 amplitudes between 84–104 ms in the Speak con-
dition. The overall pattern of results was similar to that reported in Table 1.
P2 Amplitudes
The results of the analyses regarding the P2 time window are shown in Table 2. Here, the only
marginally statistically significant effect is the reduced P2 amplitude in the Foreign condition
(p = 0.05). The results did not markedly change when the model was pruned by removing the
Trial regressor.
LSW Amplitudes
The results regarding the LSW time window are shown in Table 3. As indicated by the main
effect of Speak condition, the estimated SIS was 0.67 μV in the Native condition (p = 0.05). In
the Foreign condition, the LSW amplitude for passively heard phonemes was slightly larger
(0.26 μV) when compared to the Native condition (p = 0.04). The SIS was 0.43 μV larger in
Table 2.
Results of the mixed-effects regression on P2 amplitudes (df = 6143)
Name
Intercept
Trial
Speak
Foreign
Trial:Speak
Trial:Foreign
Speak:Foreign
Trial:Speak:Foreign
Estimate
0.21
0.01
−0.09
−0.23
0.20
−0.21
0.12
0.11
SE
0.27
0.09
0.46
0.12
0.13
0.12
0.18
0.18
t
0.79
0.11
−0.20
−1.94
1.52
−1.76
0.67
0.63
p
0.43
0.92
0.84
0.05
0.13
0.08
0.50
0.53
Lower
95% CI
−0.32
Upper
95% CI
0.74
−0.16
−1.00
−0.46
−0.06
−0.44
−0.23
−0.23
0.18
0.82
0.00
0.45
0.02
0.46
0.45
607
Neurobiology of Language
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
Table 3.
Results of the mixed-effects regression on LSW amplitudes (df = 6123)
Name
Intercept
Trial
Speak
Foreign
Trial:Speak
Trial:Foreign
Speak:Foreign
Trial:Speak:Foreign
Estimate
−1.83
−0.01
0.67
−0.26
0.00
−0.22
0.43
0.49
SE
0.26
0.09
0.34
0.12
0.13
0.12
0.18
0.18
t
−7.01
−0.07
2.00
−2.08
−0.03
−1.80
2.35
2.68
p
<0.001
Lower
95% CI
−2.34
Upper
95% CI
−1.32
0.95
0.05
0.04
0.97
0.07
0.02
0.01
−0.18
0.01
−0.50
−0.27
−0.46
0.07
0.13
0.17
1.33
−0.02
0.26
0.02
0.78
0.84
the Foreign condition than in the Native condition (Speak:Foreign, p = 0.02). Finally, the Trial:
Speak:Foreign interaction (p = 0.01) suggests that the difference in the SIS between Foreign
and Native conditions changed throughout the experiment. The Trial:Speak:Foreign interac-
tion coefficient indicates that, as trial number increased one z unit (roughly 30 trials), the
amplitude of the Speak:Foreign interaction increased by 0.49 μV.
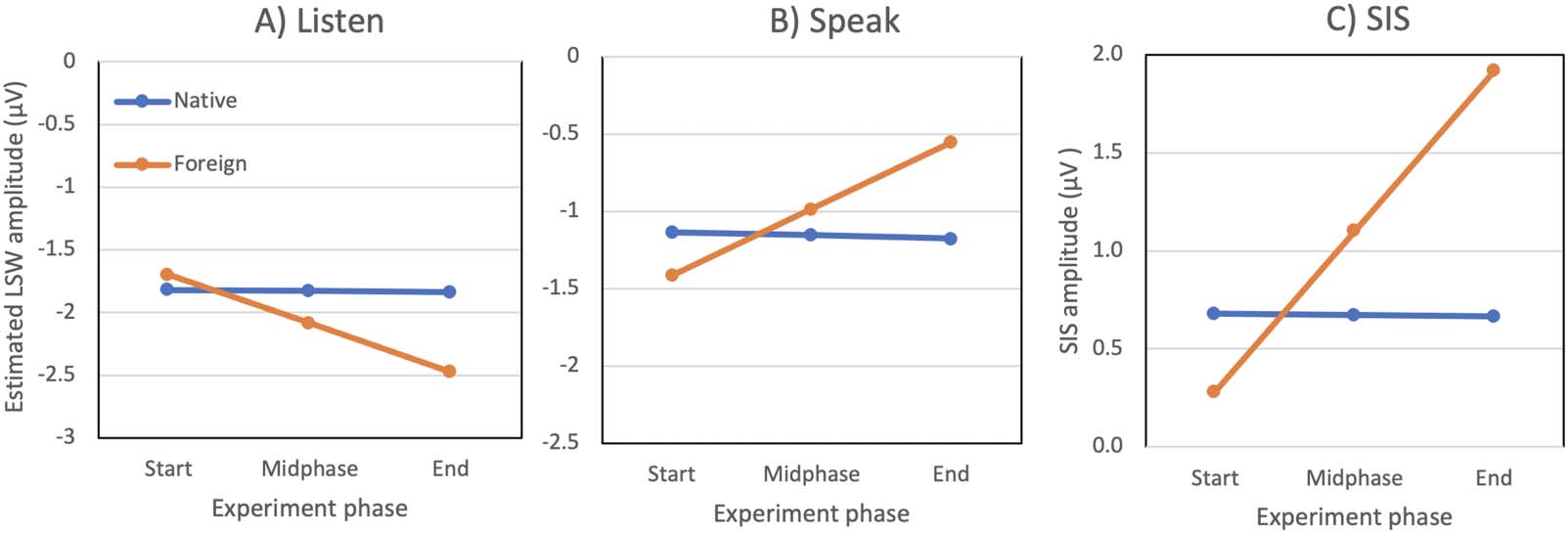
Figure 4 visualizes the modeled LWS amplitude and the resulting SIS at three different phases
of the experiment. For native phonemes (blue lines), the LSW amplitude, and consequently the
SIS, stayed approximately constant throughout the experiment. In contrast, the LSW evoked by
the foreign phonemes (orange lines) changed across the trials: Whereas in the Listen condition
the LSW amplitudes became more negative, in the Speak condition they became more positive.
As a result, the SIS increased throughout the experiment (from 0.28 μV to 1.91 μV).
Correlation Between Pronunciation Ratings and ERPs
The results of the analyses performed so far indicate that foreign phonemes modulate LSW
amplitudes and that this effect changes as a function of trials. To test if these changes
Figure 4. Estimated late slow wave (LSW) amplitudes. (A) Listen condition, (B) Speak condition, and (C) corresponding speaking-induced
suppression (SIS), during different phases of the experiment. The blue line represents the Native phoneme condition and the orange line the
Foreign phoneme condition. The SIS evoked by foreign phonemes increased throughout the experiment, whereas it stayed constant for native
phonemes. The experiment phase refers to the Trial regressor in Table 3 (i.e., −1.7, 0, and 1.7 z-units at the Start, Midphase, and End of the
experiment, respectively).
Neurobiology of Language
608
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
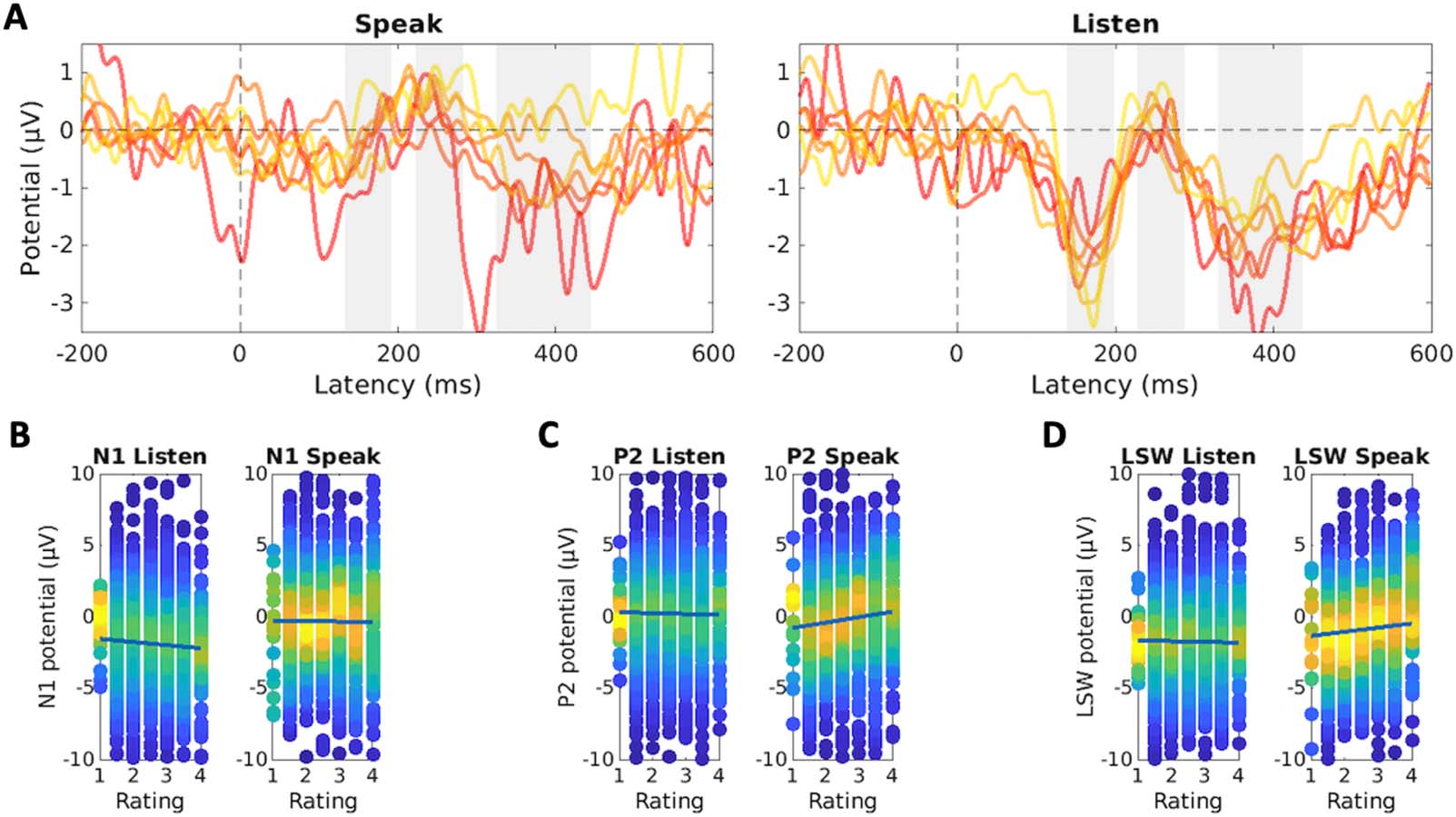
Figure 5. Event-related potentials (ERPs) in the Foreign condition as a function of pronunciation rating. (A) Grand-average ERPs for different
rating categories in the Speak and Listen conditions (frontal-central electrode cluster). The gray rectangles indicate the N1, P2, and LSW time
windows. Yellow lines represent the highest ratings, red lines the lowest (see Figure 2D). (B) N1 amplitude as a function of rating, separately for
Speak and Foreign conditions. (C) P2 amplitude as a function of rating, separately for Speak and Foreign conditions. (D) LSW amplitude as a
function of rating, separately for Speak and Foreign conditions. In panels B–D the dots indicate single-trial ERP amplitudes, and the color
represents the density of the observations. The lines in panels B–D display the results of the linear mixed-effects regression model.
correlate with improvements in the pronunciation of the foreign phoneme, we performed
mixed-effects regression analyses to examine if differences in N1, P2, and LSW amplitudes
were predicted by the ratings of foreign phoneme pronunciation accuracy (Rating). The model
also included the Speak factor (Speak vs. Listen condition), and the interaction between Speak
and Rating. The grand-average ERPs are presented in Figure 5A. In the N1 time window
(Figure 5B), the main effect of Rating (p = 0.08) and Speak:Rating interaction (p = 0.27) indi-
cated that N1 amplitudes were not modulated by the accuracy of pronunciation. When the
N1 amplitude in the Speak condition was calculated based on an earlier time window (84–
140 ms), the main effect of Rating was statistically significant (p = 0.03; Speak:Rating inter-
action p = 0.27), suggesting that higher ratings were associated with enhanced N1 amplitudes
in both Speak and Listen conditions. P2 amplitudes (Figure 5C) did not change as a function
of trial in the Listen condition (main effect of Rating, p = 0.63), but P2 was enhanced for
phonemes with higher ratings (Speak:Rating, p = 0.014). As shown in Figure 5D, a similar
pattern was observed in the LSW time window (main effect of Rating: p = 0.68; Speak:Rating
interaction: p = 0.048).
Analysis of Cue Sounds
We analyzed the ERPs produced by the Cue stimuli using linear mixed-effects regression
models. The analysis included a factor indicating Condition (Native vs. Foreign phoneme),
running (z scored) trial number, and their interaction. This analysis tests to what extent differ-
ences in the Listen and Speak conditions can be observed for stimuli not pronounced by the
participants. In the N1 time window, foreign phonemes produced stronger ERPs (β = −0.25, t =
−1.92, p = 0.054), and this effect increased through the experiment (β = −0.20, t = −2.02, p =
0.042). Models for P2 and LSW amplitudes did not reveal any statistically significant effects.
Neurobiology of Language
609
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
DISCUSSION
When learning to pronounce a novel foreign phoneme, the individual needs to acquire novel
speech targets based on auditory feedback and by trial and error. We examined whether a
well-known marker of auditory feedback control—SIS of auditory evoked activity—is modu-
lated when participants pronounce foreign vs. native phonemes. Specifically, we observed that
activity in a relatively late time window (LSW, about 300 ms after vocalization onset) was
modulated by the pronunciation of foreign phonemes. This effect is interesting because only
LSW amplitudes evoked by self-produced foreign phonemes (but not passively heard foreign
phonemes) changed over the course of the experiment, and this effect correlated positively
with pronunciation accuracy. When participants vocalized native phonemes, the LSW
remained constant throughout the experiment. Because the change in the LSW response
parallels improvements in pronunciation, and the effect was specific to pronouncing foreign
phonemes, we suggest this result reflects neural processes that mediate the acquisition of
novel phonemes.
In line with a large body of research (Behroozmand et al., 2011; Behroozmand & Larson,
2011; Curio et al., 2000; Heinks-Maldonado et al., 2005; Houde et al., 2002; Knolle et al.,
2019; Niziolek et al., 2013), we observed a strong SIS. The question arises as to why we did
not observe differences between Foreign and Native conditions in SIS in the classical N1 and
P2 time windows. Behavioral results indicated that participants had difficulties in pronouncing
the foreign /õ/. This suggests that participants did not yet have a clearly defined acoustic or
motor target for the foreign phoneme, and their phonation was likely largely based on native
phonemes (consistent with this, the formants of the foreign phoneme were biased toward the
native /ö/ phoneme). A difference in SIS is not expected if the pronunciation of foreign
phoneme relies on similar motor programs as that of native phonemes. The lack of differences
in SIS between foreign and native phonemes should not therefore be taken as evidence that the
mismatch between desired and produced speech does not modulate SIS. Many previous
studies have shown that when vocalization does not match the target, SIS is reduced in
amplitude (Behroozmand et al., 2009; Behroozmand & Larson, 2011; Chang et al., 2013;
Niziolek et al., 2013). Moreover, the present findings showed that better accuracy in pro-
nouncing foreign phonemes correlated positively with P2 amplitudes in the Speak condition.
This indicates that better foreign pronunciation was associated with reduced SIS in the P2
time window.
The fact that we did not find any differences in SIS in the early time windows suggests that
learning to better pronounce the foreign phoneme was not strongly associated with low-level
auditory feedback control mechanisms. Instead, the change in pronunciation accuracy corre-
lated with LSW amplitudes. When the participants passively listened to the foreign phoneme,
the LSW amplitude became more negative across the experiment. This result parallels the
findings reported by Alain et al. (2007) and Reinke et al. (2003), who observed that phoneme
discrimination training was associated with a modulated LSW. But when the participants in the
present study actively pronounced the foreign phoneme, the LSW amplitude became more
positive across trials. Consequently, the SIS evoked by foreign phonemes increased as a func-
tion of trials. Control analyses of the Cue sounds (/ö/ and /õ/ phonemes spoken by a native
speaker) indicated that the difference in the LSW amplitudes cannot be attributed to acoustic
differences between the /ö/ and /õ/ phonemes.
What cognitive or behavioral processes does the change in the LSW amplitude to self-
produced foreign phonemes reflect? First, because the pronunciation of foreign phonemes
improved across the experiment, the LSW amplitude does not appear to reflect fatigue or
Neurobiology of Language
610
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
inattentiveness. Moreover, if the change in LSW reflects fatigue, LSW should be negatively
associated with pronunciation accuracy, which is the opposite of what we observed. Second,
it could be suggested that the LSW evoked by self-produced phonemes is an error signal, indi-
cating a mismatch between the produced and attempted vocalization. However, if LSW
reflects an error signal, it should correlate negatively with pronunciation accuracy. Instead,
in the present study, enhanced amplitude of SIS in the LSW time window indicated improved
pronunciation. These findings suggest that the LSW may reflect processing of successful
vocalizations, which enables the participant to notice and adjust future vocalizations.
The positive amplitude shift in LSW coincides with the timing and topography of the P3a
wave, which reflects attentional capture of salient or motivationally relevant stimuli (Knolle
et al., 2019; Polich, 2007). The P3a wave is elicited by sensory stimuli that require a behav-
ioral response form the participant (e.g., participant needs to classify a stimulus and respond
using a button press; Pitts et al., 2012; Scheerer & Jones, 2018), which likely explains why a
P3a wave was not observed in the present study. Although we did not observe a P3a wave per
se, the neurocognitive basis of the positive shift in the LSW amplitude may be similar to the
mechanisms producing the P3a.
Consciously noticing successful vocalizations is likely a key part of learning to pronounce
foreign phonemes because the process requires identifying new acoustic speech targets; it is
difficult to improve one’s vocalization of foreign phonemes unless one consciously knows
what to aim for. Noticing successful pronunciations could also be interpreted as motivation-
ally relevant reinforcement signals that drive learning. Like the LSW effect we observed, cor-
rectly performed actions and feedback after a successful performance are associated with a
positive amplitude shift in frontal-central locations (called the “reward positivity”), often inter-
preted as a correlate of reinforcement signals (Carlson et al., 2011; Glazer et al., 2018;
Holroyd & Coles, 2002; Hoy et al., 2021; Ullsperger et al., 2014). Consistent with this, indi-
viduals learn better from feedback following successful trials (rather than errors; Arbel et al.,
2013; Chiviacowsky & Wulf, 2007). Although, in the present study, the participants were not
provided direct feedback on the success of their pronunciation, the playback of their own
pronunciation likely functioned as type of feedback signal (Parrell, 2021). Reinforcement
learning likely plays a role in learning to produce foreign phonemes but the topic remains little
studied (Parrell, 2021).
Based on the similarities to P3a and reward positivity, we suggest that the positive shift in
the LSW amplitude to self-produced foreign phonemes may reflect motivational, saliency, or
reinforcement signals. These enable the individual to notice successful pronunciations, which
then translates to improved pronunciation during the experiment. By successful pronunciation
we do not simply refer to the absence of vocalization errors, but to pronunciation that finds its
target surprisingly well. These successful pronunciations could be “planned” (i.e., in the sense
that this was what the individual attempted to do), but they could also be due to pronunciation
errors that, by coincidence, match the target sound better than intended.
Assuming that the sources that contribute to P3a also contribute to LSW, the positive shift in
the LSW amplitude could also reflect participants’ conscious confidence in their performance.
Frömer et al. (2021) showed that P3a amplitudes correlated with participants’ confidence on
the success of their actions. The authors suggest that confidence approximates the amount of
noise in the efference copy signals (i.e., their precision), which together with the efference
copy and feedback, allow the individuals to make inferences about the success of their action.
Their results showed that participants with more accurately “calibrated” confidence (i.e., more
accurate estimate of noise in efference copies) learned better (Frömer et al., 2021). This
Neurobiology of Language
611
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
suggests that LSW could reflect higher-order monitoring mechanisms: Whereas SIS in the N1
time window may reflect automatic “online” corrections based on the efference copy (Hickok
et al., 2011; Houde & Nagarajan, 2011; Tourville & Guenther, 2011), the LSW may reflect
monitoring processes that integrate multiple sources of information (e.g., auditory feedback
on one’s pronunciation, predictions based on the efference copy, and the individual’s confi-
dence in the accuracy of these predictions).
In conclusion, our results show that the ability to correctly pronounce a novel foreign
phoneme correlates with the amplitude of a late slow ERP (320–440 ms after vocalization).
Activity during this time window was differently modulated when participants pronounced
foreign (as compared to native) phonemes, and the effect changed during the experiment, par-
alleling improvements in pronunciation. We propose that activity in the LSW time window
reflects high-level performance monitoring processes that signal successful pronunciations
and help learn motor commands that produce novel acoustic speech targets. If our interpre-
tation is correct, we expect that the effect in the LSW time window (difference between
Foreign and Native conditions) disappears once the individual has successfully learned to pro-
duce the novel phoneme (i.e., there is little room, or need for further improvement), and the
pronunciation of the foreign phoneme has automatized. A better understanding of the pro-
cesses that mediate the acquisition of novel phonemes may, in the future, help shed light
on various speech-related phenomena, such as the neural processes underlying native lan-
guage acquisition, second language learning, and rehabilitation of speech deficits.
ACKNOWLEDGMENTS
We thank Ita Puusepp for help with the ratings. We thank Teemu Laine for help with the exper-
imental setup and equipment.
FUNDING INFORMATION
Minna Lehtonen, Research Council of Norway, Award ID: 223265. Pilleriin Sikka, Alfred
Österlunds Stiftelse (https://dx.doi.org/10.13039/501100005390). Pilleriin Sikka, Emil Aaltonen
Foundation research grant.
AUTHOR CONTRIBUTIONS
Henry Railo: Conceptualization; Formal analysis; Methodology; Supervision; Writing—
original draft; Writing—review and editing. Anni Varjonen: Formal analysis; Investigation;
Writing—original draft. Minna Lehtonen: Methodology; Writing—review & editing. Pilleriin
Sikka: Methodology; Writing—review & editing.
DATA AVAILABILITY STATEMENT
The data, preprocessing, and analysis scripts are available at https://osf.io/wnd2j/.
REFERENCES
Alain, C., Snyder, J. S., He, Y., & Reinke, K. S. (2007). Changes in
auditory cortex parallel rapid perceptual learning. Cerebral
Cortex, 17(5), 1074–1084. https://doi.org/10.1093/cercor
/bhl018, PubMed: 16754653
Arbel, Y., Goforth, K., & Donchin, E. (2013). The good, the bad, or
the useful? The examination of the relationship between the
feedback-related negativity (FRN ) and long-term learning
outcomes. Journal of Cognitive Neuroscience, 25(8), 1249–1260.
https://doi.org/10.1162/jocn_a_00385, PubMed: 23489147
Asu, E. L., & Teras, P. (2009). Estonian. Journal of the International
Phonetic Association, 39(3), 367–372. https://doi.org/10.1017
/S002510030999017X
Behroozmand, R., Karvelis, L., Liu, H., & Larson, C. R. (2009).
Vocalization-induced enhancement of the auditory cortex
Neurobiology of Language
612
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
responsiveness during voice F0 feedback perturbation. Clinical
Neurophysiology, 120(7), 1303–1312. https://doi.org/10.1016/j
.clinph.2009.04.022, PubMed: 19520602
Behroozmand, R., & Larson, C. R. (2011). Error-dependent modu-
lation of speech-induced auditory suppression for pitch-shifted
voice feedback. BMC Neuroscience, 12, Article 54. https://doi
.org/10.1186/1471-2202-12-54, PubMed: 21645406
Behroozmand, R., Liu, H., & Larson, C. R. (2011). Time-dependent
neural processing of auditory feedback during voice pitch error
detection. Journal of Cognitive Neuroscience, 23(5), 1205–1217.
https://doi.org/10.1162/jocn.2010.21447, PubMed: 20146608
Bittium. (2022). NeurOne Tesla ( Version 1.4.1.64) [EEG amplifier
and software]. https://www.bittium.com/medical/bittium-neurone
Carlson, J. M., Foti, D., Mujica-Parodi, L. R., Harmon-Jones, E., &
Hajcak, G. (2011). Ventral striatal and medial prefrontal BOLD
activation is correlated with reward-related electrocortical
activity: A combined ERP and fMRI study. NeuroImage, 57(4),
1608–1616. https://doi.org/10.1016/j.neuroimage.2011.05.037,
PubMed: 21624476
Chang, C. Y., Hsu, S. H., Pion-Tonachini, L., & Jung, T. P. (2020).
Evaluation of artifact subspace reconstruction for automatic arti-
fact components removal in multi-channel EEG recordings. IEEE
Transactions on Biomedical Engineering, 67(4), 1114–1121.
https://doi.org/10.1109/ TBME.2019.2930186, PubMed:
31329105
Chang, E. F., Niziolek, C. A., Knight, R. T., Nagarajan, S. S., &
Houde, J. F. (2013). Human cortical sensorimotor network under-
lying feedback control of vocal pitch. Proceedings of the
National Academy of Sciences, 110(7), 2653–2658. https://doi
.org/10.1073/pnas.1216827110, PubMed: 23345447
Chiviacowsky, S., & Wulf, G. (2007). Feedback after good trials
enhances learning. Research Quarterly for Exercise and Sport,
78(2), 40–47. https://doi.org/10.1080/02701367.2007
.10599402, PubMed: 17479573
Curio, G., Neuloh, G., Numminen, J., Jousmäki, V., & Hari, R.
(2000). Speaking modifies voice-evoked activity in the human
auditory cortex. Human Brain Mapping, 9, 183–191. https://doi
.org/10.1002/(SICI)1097-0193(200004)9:4<183::AID-HBM1>3
.0.CO;2-Z, PubMed: 10770228
de Cheveigné, A. (2020). ZapLine: A simple and effective method
to remove power line artifacts. NeuroImage, 207, Article 116356.
https://doi.org/10.1016/j.neuroimage.2019.116356, PubMed:
31786167
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source tool-
box for analysis of single-trial EEG dynamics including indepen-
dent component analysis. Journal of Neuroscience Methods,
134(1), 9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009,
PubMed: 15102499
Díaz, B., Baus, C., Escera, C., Costa, A., & Sebastián-Gallés, N.
(2008). Brain potentials to native phoneme discrimination reveal
the origin of individual differences in learning the sounds of a
second language. Proceedings of the National Academy of
Sciences, 105(42), 16083–16088. https://doi.org/10.1073/pnas
.0805022105, PubMed: 18852470
EASYCAP. (2022). EASYCAP [electrodes]. https://www.easycap.de/
Frömer, R., Nassar, M. R., Bruckner, R., Stürmer, B., Sommer, W., &
Yeung, N. (2021). Response-based outcome predictions and
confidence regulate feedback processing and learning. ELife,
10, Article e62825. https://doi.org/10.7554/eLife.62825,
PubMed: 33929323
Glazer, J. E., Kelley, N. J., Pornpattananangkul, N., Mittal, V. A., &
Nusslock, R. (2018). Beyond the FRN: Broadening the time-course
of EEG and ERP components implicated in reward processing.
International Journal of Psychophysiology, 132(Part B), 184–202.
https://doi.org/10.1016/j.ijpsycho.2018.02.002, PubMed:
29454641
Hain, T. C., Burnett, T. A., Kiran, S., Larson, C. R., Singh, S., &
Kenney, M. K. (2000). Instructing subjects to make a voluntary
response reveals the presence of two components to the
audio-vocal reflex. Experimental Brain Research, 130(2), 133–141.
https://doi.org/10.1007/s002219900237, PubMed: 10672466
Heinks-Maldonado, T. H., Mathalon, D. H., Gray, M., & Ford, J. M.
(2005). Fine-tuning of auditory cortex during speech production.
Psychophysiology, 42(2), 180–190. https://doi.org/10.1111/j
.1469-8986.2005.00272.x, PubMed: 15787855
Hickok, G., Houde, J., & Rong, F. (2011). Sensorimotor integration
in speech processing: Computational basis and neural organiza-
tion. Neuron, 69(3), 407–422. https://doi.org/10.1016/j.neuron
.2011.01.019, PubMed: 21315253
Holroyd, C. B., & Coles, M. G. H. (2002). The neural basis of human
error processing: Reinforcement learning, dopamine, and the
error-related negativity. Psychological Review, 109(4), 679–709.
https://doi.org/10.1037/0033-295X.109.4.679, PubMed:
12374324
Houde, J. F., & Jordan, M. I. (1998). Sensorimotor adaptation in
speech production. Science, 279(5354), 1213–1216, https://doi
.org/10.1126/science.279.5354.1213, PubMed: 9469813
Houde, J. F., & Nagarajan, S. S. (2011). Speech production as state
feedback control. Frontiers in Human Neuroscience, 5, Article
82. https://doi.org/10.3389/fnhum.2011.00082, PubMed:
22046152
Houde, J. F., Nagarajan, S. S., Sekihara, K., & Merzenich, M. M.
(2002). Modulation of the auditory cortex during speech: An MEG
study. Journal of Cognitive Neuroscience, 14(8), 1125–1138.
https://doi.org/10.1162/089892902760807140, PubMed:
12495520
Hoy, C. W., Steiner, S. C., & Knight, R. T. (2021). Single-trial
modeling separates multiple overlapping prediction errors during
reward processing in human EEG. Communications Biology,
4(1), Article 910. https://doi.org/10.1038/s42003-021-02426-1,
PubMed: 34302057
Klug, M., & Gramann, K. (2020). Identifying key factors for
improving ICA-based decomposition of EEG data in mobile
and stationary experiments. European Journal of Neuroscience,
54(12), 8406–8420. https://doi.org/10.1111/ejn.14992, PubMed:
33012055
Knolle, F., Schwartze, M., Schröger, E., & Kotz, S. A. (2019). Audi-
tory predictions and prediction errors in response to self-initiated
vowels. Frontiers in Neuroscience, 13, Article 1146. https://doi
.org/10.3389/fnins.2019.01146, PubMed: 31708737
Lametti, D. R., Smith, H. J., Watkins, K. E., & Shiller, D. M. (2018).
Robust sensorimotor learning during variable sentence-level
speech. Current Biology, 28(19), 3106–3113. https://doi.org/10
.1016/j.cub.2018.07.030, PubMed: 30245103
Näätänen, R., Lehtokoski, A., Lennes, M., Cheour, M., Huotilainen,
M., Iivonen, A., Vainio, M., Alku, P., Ilmoniemi, R. J., Luuk, A.,
Allik, J., Sinkkonen, J., & Alho, K. (1997). Language-specific pho-
neme representations revealed by electric and magnetic brain
responses. Nature, 385(6615), 432–434. https://doi.org/10.1038
/385432a0, PubMed: 9009189
Nils. (2021). Scatter plot colored by kernel density estimate
( Version 1.0.0.0) [function]. Matlab Central, Mathworks. https://
se.mathworks.com/matlabcentral/fileexchange/65728-scatter
-plot-colored-by-kernel-density-estimate
Niziolek, C. A., Nagarajan, S. S., & Houde, J. F. (2013). What does
motor efference copy represent?: Evidence from speech production.
Neurobiology of Language
613
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
EEG correlates of foreign phoneme learning
Journal of Neuroscience, 33(41), 16110–16116. https://doi.org/10
.1523/JNEUROSCI.2137-13.2013, PubMed: 24107944
Nuwer, M. R., Comi, G., Emerson, R., Fuglsang-Frederiksen, A.,
Guérit, J.-M., Hinrichs, H., et al. (1998). IFCN standards for
digital recording of clinical EEG. Electroencephalography and
Clinical Neurophysiology, 106(3), 259–261. https://doi.org/10
.1016/S0013-4694(97)00106-5, PubMed: 9743285
Parrell, B. (2021). A potential role for reinforcement learning in speech
production. Journal of Cognitive Neuroscience, 33(8), 1470–1486.
https://doi.org/10.1162/jocn_a_01742, PubMed: 34496380
Peltola, M. S., Kujala, T., Tuomainen, J., Ek, M., Aaltonen, O., &
Näätänen, R. (2003). Native and foreign vowel discrimination
as indexed by the mismatch negativity (MMN) response. Neuro-
science Letters, 352(1), 25–28. https://doi.org/10.1016/j.neulet
.2003.08.013, PubMed: 14615041
Pion-Tonachini, L., Kreutz-Delgado, K., & Makeig, S. (2019).
ICLabel: An automated electroencephalographic independent
component classifier, dataset, and website. NeuroImage, 198,
181–197. https://doi.org/10.1016/j.neuroimage.2019.05.026,
PubMed: 31103785
Pitts, M. A., Martínez, A., & Hillyard, S. A. (2012). Visual processing
of contour patterns under conditions of inattentional blindness.
Journal of Cognitive Neuroscience, 24(2), 287–303. https://doi
.org/10.1162/jocn_a_00111, PubMed: 21812561
Polich, J. (2007). Updating P300: An integrative theory of P3a and
P3b. Clinical Neurophysiology, 118(10), 2128–2148. https://doi
.org/10.1016/j.clinph.2007.04.019, PubMed: 17573239
Railo, H., Nokelainen, N., Savolainen, S., & Kaasinen, V. (2020).
Deficits in monitoring self-produced speech in Parkinson’s
disease. Clinical Neurophysiology, 131(9), 2140–2147. https://
doi.org/10.1016/j.clinph.2020.05.038, PubMed: 32682241
Reinke, K. S., He, Y., Wang, C., & Alain, C. (2003). Perceptual
learning modulates sensory evoked response during vowel
segregation. Cognitive Brain Research, 17(3), 781–791. https://
doi.org/10.1016/S0926-6410(03)00202-7, PubMed: 14561463
Saloranta, A., Alku, P., & Peltola, M. S. (2020). Listen-and-repeat
training improves perception of second language vowel dura-
tion: Evidence from mismatch negativity (MMN ) and N1
responses and behavioral discrimination. International Journal
of Psychophysiology, 147, 72–82. https://doi.org/10.1016/j
.ijpsycho.2019.11.005, PubMed: 31743699
SCCN. (2019). DIPFIT [software plug in]. Swartz Center for Compu-
tational Neuroscience. https://github.com/sccn/dipfit
Scheerer, N. E., & Jones, J. A. (2018). The role of auditory feedback
at vocalization onset and mid-utterance. Frontiers in Psychology,
9, Article 2019. https://doi.org/10.3389/fpsyg.2018.02019,
PubMed: 30459679
Tamminen, H., Peltola, M. S., Kujala, T., & Näätänen, R. (2015).
Phonetic training and non-native speech perception—New
memory traces evolve in just three days as indexed by the mis-
match negativity (MMN) and behavioural measures. Interna-
tional Journal of Psychophysiology, 97(1), 23–29. https://doi.org
/10.1016/j.ijpsycho.2015.04.020, PubMed: 25956191
Tourville, J. A., & Guenther, F. H. (2011). The DIVA model: A neural
theory of speech acquisition and production. Language and
Cognitive Processes, 26(7), 952–981. https://doi.org/10.1080
/01690960903498424, PubMed: 23667281
Tremblay, K., Kraus, N., & McGee, T. (1998). The time course of
auditory perceptual learning: Neurophysiological changes dur-
ing speech-sound training. Neuroreport, 9(16), 3557–3560.
https://doi.org/10.1097/00001756-199811160-00003, PubMed:
9858359
Ullsperger, M., Fischer, A. G., Nigbur, R., & Endrass, T. (2014).
Neural mechanisms and temporal dynamics of performance
monitoring. Trends in Cognitive Sciences, 18(5), 259–267.
https://doi.org/10.1016/j.tics.2014.02.009, PubMed: 24656460
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
9
9
2
0
6
0
8
9
1
n
o
_
a
_
0
0
0
8
0
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
614