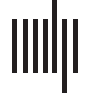RESEARCH ARTICLE
Supramodal Sentence Processing in the Human
Brain: fMRI Evidence for the Influence of Syntactic
Complexity in More Than 200 Participants
Julia Uddén1,2,3,4
, Annika Hultén1,2
Karin Harbusch5, Antal van den Bosch2
, Jan-Mathijs Schoffelen2, Nietzsche Lam1,2,
, Gerard Kempen1
, Karl Magnus Petersson1,2,
and Peter Hagoort1,2
1Max Planck Institute for Psycholinguistics, Nijmegen, the Netherlands
2Donders Institute for Brain, Cognition and Behaviour, Centre for Cognitive Neuroimaging, Radboud University, Nijmegen,
the Netherlands
3Department of Linguistics, Stockholm University, Stockholm, Sweden
4Department of Psychology, Stockholm University, Stockholm, Sweden
5Department of Computer Science, University of Koblenz-Landau, Koblenz, Germany
Keywords: complexity, fMRI, sentence processing, supramodal, unification
ABSTRACT
This study investigated two questions. One is: To what degree is sentence processing beyond
single words independent of the input modality (speech vs. reading)? The second question is:
Which parts of the network recruited by both modalities is sensitive to syntactic complexity?
These questions were investigated by having more than 200 participants read or listen
to well-formed sentences or series of unconnected words. A largely left-hemisphere
frontotemporoparietal network was found to be supramodal in nature, i.e., independent of
input modality. In addition, the left inferior frontal gyrus (LIFG) and the left posterior middle
temporal gyrus (LpMTG) were most clearly associated with left-branching complexity. The left
anterior temporal lobe showed the greatest sensitivity to sentences that differed in right-
branching complexity. Moreover, activity in LIFG and LpMTG increased from sentence onset
to end, in parallel with an increase of the left-branching complexity. While LIFG, bilateral
anterior temporal lobe, posterior MTG, and left inferior parietal lobe all contribute to the
supramodal unification processes, the results suggest that these regions differ in their
respective contributions to syntactic complexity related processing. The consequences of
these findings for neurobiological models of language processing are discussed.
INTRODUCTION
In order to extract meaning from the orthographic patterns or from the speech sounds, multiple
processing steps are involved. One important step is to retrieve relevant word information from
long-term memory (the mental lexicon; Altmann, 1998). This information includes the mor-
phological makeup of words, their syntactic features, and lexical aspects of their meaning.
But this is not enough. In many cases a simple concatenation of individual word meanings
will not result in a correct interpretation (Jackendoff, 2002). The reason is that in language,
words that belong together often do not go together (Lashley, 1951). This is what linguists refer
to as non-adjacent dependencies between the lexical elements that make up an utterance.
How to combine word information retrieved from memory into representations of sentence-
level meaning that are constructed on the fly is what we refer to as unification (Hagoort, 2005,
a n o p e n a c c e s s
j o u r n a l
Citation: Uddén, J., Hultén, A.,
Schoffelen, J.-M., Lam, N., Harbusch,
K., van den Bosch, A., Kempen, G.,
Petersson, K. M., & Hagoort, P. (2022).
Supramodal sentence processing in
the human brain: fMRI evidence for
the influence of syntactic complexity
in more than 200 participants.
Neurobiology of Language, 3(4),
575–598. https://doi.org/10.1162/nol_a
_00076
DOI:
https://doi.org/10.1162/nol_a_00076
Supporting Information:
https://doi.org/10.1162/nol_a_00076
Received: 3 March 2021
Accepted: 13 June 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Authors:
Julia Uddén
julia.udden@psychology.su.se
Peter Hagoort
peter.hagoort@mpi.nl
Handling Editor:
Stephen M. Wilson
Copyright: © 2022
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
Unification:
The process of combining word
information, retrieved from memory,
into representations of sentence-level
meaning that are constructed on
the fly.
Unification complexity or
Left-branching complexity
(of a sentence):
The number of non-adjacent
elements that have to be kept online,
at the point in the sentence where
most non-adjacent elements are
simultaneously kept online (i.e., the
maximum used stack depth).
Supramodal process:
A process that is independent of
input modality (e.g., independent of
visual or auditory input modality).
2013; Hagoort & Indefrey, 2014; Vosse & Kempen, 2000, 2009). The number of non-adjacent
elements that have to be kept online determines unification complexity. In this large functional mag-
netic resonance imaging (fMRI) study on sentence processing (N = 204), we address two outstanding
questions. (i) To what extent is the network subserving unification operations independent of the
modality of input (spoken and written)? This was investigated by confronting half of the participants
with the materials in spoken format and half with the same materials in written format. (ii) Which
nodes in the language network are modulated by variation in syntactic complexity? For each sen-
tence presented to the participant, we calculated a measure of complexity, which allowed us to
identify the areas that were most sensitive to complexity variations.
Modality-Independence of Unification
It is generally assumed that at least some aspects of structure-building processes in the spoken
and written modalities are subserved by similar modality-independent operations (Favier &
Huettig, 2021). For instance, in the processing model of structure building called the unifica-
tion framework (Vosse & Kempen, 2000, 2009), the attachment of each new lexical item to the
incrementally constructed syntactic representation of the sentence is identical for both the
visual and the auditory language input, but this has not been explicitly tested. In the memory,
unification, and control framework (Hagoort, 2005, 2013), the word information mainly stored
in the temporal lobe includes specifications of syntax, morphology, and information about
word meaning (Joshi & Schabes, 1997; Vosse & Kempen, 2000). The process of unifying these
lexical information types with the sentence and discourse context is constrained by lexical
features in a process assumed to be supramodal. It has been suggested that after a forward
sweep from sensory cortex to the left temporal and parietal lobe, top-down signals from the
left inferior frontal gyrus (LIFG) re-enter the posterior regions in cycles of reactivation (Baggio &
Hagoort, 2011), establishing a unification network with the involvement of at least two left
hemisphere regions (left temporal/parietal cortex and LIFG, where the LIFG is the higher level
node in the network). Visual and auditory language processing streams are hypothesized to
converge on this supramodal unification network during comprehension, potentially starting
in posterior areas (for a detailed account, see Wilson et al., 2018). The unification network thus
includes both a frontal and a temporoparietal node, but the LIFG is thought to be crucial for the
higher-level unification processes whereas the mental lexicon (or memory component) is
thought to recruit especially the temporoparietal node.
Sentence Complexity
The second aspect that we addressed is related to sentence complexity. Processing complexity
in language processing is often due to the fact that words that belong together do not always go
together; that is, they do not appear in adjacent positions. In a recent study (Futrell et al.,
2015), it was found that there is an almost universal tendency (based on an analysis of 37
languages) to dis-prefer sentences in which structurally related words are far apart, presumably
as a result of the extra processing costs associated with non-adjacency. Nevertheless, non-
adjacency is a common phenomenon in language processing and a hallmark of human lan-
guages. It occurs not only in sentences with a left-branching structure, but also in sentences
with a right-branching structure. There is evidence that on the whole, left-branching structures
are harder to process than right-branching structures. An increased cost of maintenance and
structure building for left-branching sentence aspects compared to right-branching sentence
aspects was first suggested by Fodor and colleagues (Fodor et al., 1974). This claim of an
added processing load for left-branching structures has been supported by evidence from both
production and comprehension studies (Cheung & Kemper, 1992; Kemper, 1986, 1987;
Norman et al., 1992).
Neurobiology of Language
576
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
Snijders et al. (2009) identified the LIFG and the middle temporal gyrus (MTG) as core uni-
fication regions in a study that contrasted sentence processing with the processing of word
lists. In the current study, we used a similar paradigm but we extended it with an explicit
manipulation and measure of sentence complexity.
The neuroimaging literature on sentence complexity is substantial (among many others, see
Caplan et al., 1998; Cooke et al., 2002; Makuuchi et al., 2009; Meltzer et al., 2010; Peelle
et al., 2004; Santi & Grodzinsky, 2010; Vogelzang et al., 2020), but most often the studies have
been restricted to (a) comparing two conditions of complex vs. simpler sentences, and (b) mea-
suring one sensory modality only (but see Braze et al., 2011; Constable et al., 2004; Michael
et al., 2001; Shankweiler et al., 2008). When comparing two conditions of sentence complex-
ity, most studies have realized this manipulation by comparing object-relative to subject-
relative sentences, the former ones known to be more complex than the latter ones (e.g., Braze
et al., 2011; Constable et al., 2004; Cooke et al., 2002; Meltzer et al., 2010; Michael et al.,
2001; Peelle et al., 2004). In addition, some studies (e.g., Caplan et al., 1998; Makuuchi et al.,
2009; Santi & Grodzinsky, 2010; Stromswold et al., 1996) compared more complex center
embedded sentences to simpler right-branching sentences. In our study, a complexity measure
is instead used to calculate the processing complexity for each individual sentence.
Our complexity measure is motivated by two observations. (a) First is the finding that sen-
tences with a left-branching structure (see Figure 1) are particularly hard to process (Cheung &
Kemper, 1992; Kemper, 1986, 1987; Norman et al., 1992). We thus separated left-branching
from right-branching complexity. (b) The second motivating observation is that there is a high
processing cost related to building sentence structure with multiple simultaneous non-local
dependencies. This is found for both natural (Makuuchi et al., 2009) and artificial grammars
(Bahlmann et al., 2008; de Vries et al., 2012; Makuuchi et al., 2009; Uddén & Bahlmann,
2012; Uddén et al., 2012). Based on these two observations, our measure quantifies the
amount of simultaneous left-branching non-local dependencies in a sentence (see Figure 1).
It is of central importance that the sentence complexity is related to the incrementality of pars-
ing from left to right.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Left-branching dependencies point leftward from head to dependent. The left-branching
Figure 1.
processing complexity measure is calculated per sentence, as the maximum simultaneous non-
resolved left-branching verbal dependencies (i.e., maximum number of dependents not yet assigned
to a verb during the incremental parse). For the example sentence, this number equals 3 (reached
after retrieval of the word gekocht). One of the open dependencies (from papegaai) is resolved after
reading gekocht and other two at had, since the participle ( gekocht) needs to be bound to an aux-
iliary (had ), and the auxiliary needs to fill its subject argument with an antecedent that has the right
number marking (i.e., Henk, singular). Included below the sentence are: a literal and non-literal
translation into English, together with the corresponding word list with the translation. Labels on
the arcs: Det: determiner; Mod: modifier; Subj: subject; Iobj: indirect object; Dobj: direct object;
CCmp: complement clause.
Neurobiology of Language
577
Syntactic complexity in supramodal sentence processing
To illustrate the importance of left to right processing, one could think of a stack (i.e., a first-
in-last-out memory architecture) storing words that cannot yet be unified with the rest of the
sentence structure. Each time a left-branching dependency is open, an element is pushed onto
the stack, and only when it is closed is the element popped. The left-branching complexity
measure thus corresponds to the stack depth used for a sentence. However, we do not want
to make a strong assumption that there is a buffer (in the form of a stack or otherwise). If no
buffering of words occurs during incremental processing, the complexity measure still singles
out the sentences that have a high unification load for the following reason: Multiple simul-
taneously open dependencies lead to more options for the dependent to be unified with a
head, when the head arrives. The number of simultaneously open dependencies is thus a com-
plexity measure probing relevant syntactic processes independent of the existence of a stack or
other buffers.
The left-branching complexity was of greatest interest in our neuroimaging study since we
predicted that it would be associated with the greatest processing difficulties. Our left-
branching complexity measure quantifies the maximum simultaneous number of dependents
not yet assigned to the head of a verb phrase. Unification happens between the head of a
phrase and its arguments. For instance, in a noun phrase the determiner and the adjective need
to be unified with the noun as the head of the noun phrase (e.g., The nice lady; see Figure 1).
The longer the distance between the heads and their arguments, and the more dependents and
arguments there are to be maintained, the larger the processing demands (see Figure 1). In this
case, verb phrases are especially relevant, since in general the verb is the nucleus of the prop-
osition that is expressed in the sentence. For instance, upon reading the verb kick, a noun or
the name of a person is expected to fill the argument slot for the agent of the action specified
by the verb (e.g., the man). In addition, another noun is expected to take the argument slot for
the undergoer of the action (e.g., the ball, as in the man kicks the ball ). We expected process-
ing to be more demanding if the arguments precede the head since the head is a stronger
predictor for following arguments than the arguments are predictors for a following head.
Therefore, we only count dependencies with a verbal head when we calculate the processing
complexity. Although there are multiple ways to determine syntactic complexity, we note that
in many cases these measures are highly correlated. (See Supplementary Results in the Sup-
porting Information, which can be found at https://doi.org/10.1162/nol_a_00076.) Therefore,
we surmise that our results are generalizable beyond the specifics of our choice for a measure
of syntactic complexity.
To increase the sensitivity for unification associated with sentence complexity, we made
sure that there was enough variance in the sentence structures. This was achieved by intro-
ducing relative clauses in half of our sentence material, while the other half had mixed sen-
tence structures without relative clauses. Left-branching complexity was higher on average for
sentences with relative clauses (see the Materials and Methods section). The analysis was,
however, not a standard analysis comparing two complexity conditions, but a parametric anal-
ysis probing for the sentence complexity effect across all sentences in the experiment.
In addition to localizing a supramodal structure-building network in the brain, we also
wanted to characterize the temporal dynamics of this network over the course of the sentence.
While temporal dynamics are often studied using oscillatory modulations in electrocorticog-
raphy (ECoG) or magnetoencephalography (MEG; Fedorenko et al., 2016; Lam et al., 2016),
our large-scale fMRI study is a complement to this literature. In addition, testing only sentence
average activations would decrease the sensitivity for potentially subtle modality-dependent
effects of structure building. For this purpose, we divided the sentences into four time bins of
equal length and formed linear contrasts to test increases and decreases in activity over the
Neurobiology of Language
578
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
course of a sentence. For this analysis, we focused on predefined regions of interest (ROIs)
found to be involved in structural unification (Snijders et al., 2009). Based on a recent
meta-analysis of fMRI-studies of both syntactic and semantic sentence-level unification, the
LIFG and the left posterior middle temporal gyrus (LpMTG) were chosen as primary ROIs
(Hagoort & Indefrey, 2014; Tyler et al., 2010, 2013). The role of the left anterior temporal lobe
(LaTL) in structure-building processes during sentence processing is less clear. So far, there is
inconsistent evidence for the role of the aTL in syntactic processing. Some studies (e.g., Brennan
et al., 2012; Rogalsky et al., 2008) report syntactic activations in the LaTL. However, many
other studies have failed to find such activations. Moreover, other studies in patients with
semantic dementia have found mainly semantic impairments (e.g., Chen et al., 2017; Lambon
Ralph et al., 2012). Hence, the role of the aTL in syntactic processing is not yet fully substan-
tiated or clear (cf. Bhattasali et al., 2019; Pallier et al., 2011). To investigate this further, we
therefore also included a third ROI, based on coordinates from the study by Brennan et al.
(2012), in a region in the LaTL.
MATERIALS AND METHODS
Participants
A total of 242 participants volunteered to participate in a larger study—the MOUS study
(Mother of all Unification Studies; Schoffelen et al., 2019)—in which all participants took part
in an fMRI and a MEG session. Of these, 38 participants were excluded (see next paragraph).
The resulting 204 native Dutch speakers had a mean age of 22 years (range: 18–33 years). Half
of the participants read sentences and word lists (visual group, 102 participants, 51 men), and
the other half listened to auditory versions of the same materials (auditory group, 102 partic-
ipants, 51 men). The study was approved by the local ethics committee (CMO; Committee on
Research Involving Human Subjects in the Arnhem-Nijmegen region) and followed guidelines
of the Helsinki declaration. All data of the MOUS study are available as open access with a full
specification of the materials and design features, and shareable in the brain imaging data
structure (BIDS) format (Schoffelen et al., 2019).
All participants were right-handed as assessed by the Bever (Bever et al., 1989) handedness
questionnaire (including familial handedness), had normal or corrected-to-normal vision, and
reported no history of neurological, developmental, or language deficits. We screened for
medication use and excluded anyone on prescription medication. The instructions further con-
tained a statement that no medication, alcohol, or drugs should be used on the day of the
measurement. A total of 38 participants were excluded: (a) 20 because of technical problems
(the most common were problems with data transfer, scanner hardware errors, presentation
software problems resulting in faulty triggers, or absence of comprehension questions); (b)
4 because of poor data quality due to excessive blinking in the MEG session (affecting the
amount of remaining MEG trials after artifact rejection), or more than 3.5 mm movement; (c)
6 because of study interruption; (d) 4 because participants did not fulfill the inclusion criteria
(full list: 18–35 years, right-handed, self-reported Dutch monolingual language background,
normal or corrected-to-normal vision, no self-reported history of neurological, developmental,
or language deficits, MRI-compatibility (not pregnant, no claustrophobia, no incompatible
devices, no incompatible tattoos, no incompatible self-reported operation history or metal in
body)); and (e) 4 because of the participant’s poor compliance with the task as measured with
the comprehension question performance (removal of these four outliers in task performance
meant that all included participants had more than 59% correct, mean 80% correct +/− stan-
dard deviation of 9%; see further details in the Comprehension questions section).
Neurobiology of Language
579
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
Language Stimuli
The stimuli consisted of 360 sentences and their 360 word list counterparts. Sentences were
constructed to vary in complexity. One way to make complexity vary is by introducing a rel-
ative clause (Gibson, 1998). Half of the sentences contained a relative clause; the other half of
the sentences were without a relative clause. Table 1 presents examples of the materials. The
sentences varied between 9 and 15 words in length.
For each sentence, a corresponding word list was created by scrambling the words in the
sentence so that three or more consecutive words did not form a coherent fragment. Since the
sentence and word list conditions contain the same words, the comparison of sentences with
word lists allowed us to probe the sentence-level unification process, while controlling for the
retrieval of lexical items.
An important aspect of the syntactic complexity of the sentences can be formalized in terms
of their dependency structure. The automatic FROG-parser (https://antalvandenbosch
.ruhosting.nl/; https://languagemachines.github.io/frog/) was used to create a dependency tree
for every sentence (see further details in the More on dependency trees section). The resulting
trees were manually checked and corrected.
We calculated the complexity of each sentence on the basis of the dependency structure,
separately for left- and right-branching sentence aspects. For the left-branching processing
complexity measure, at each word we calculated the number of dependents that had not
yet been attached to a verbal head (i.e., the head had not yet been encountered). The maxi-
mum number over the entire sentence was the left-branching complexity for that sentence. In
other words, the left-branching complexity is the maximum number of simultaneously open
left-branching dependencies (see Figure 1). Similarly, for right-branching complexity, at each
word in each sentence, we calculated the number of dependents that would still connect to
verbal heads that had been presented up to that word. The crucial difference between left- and
right-branching constructions is the order of heads and arguments. In left-branching depen-
dencies, the arguments precede the head. In right-branching sentences, the heads are followed
by the arguments further downstream in the sentence. The calculation of the right-branching
complexity measure is thus symmetric with respect to the left-branching complexity measure.
More on dependency trees: Heads, dependents, and the complexity measure
Like the more familiar phrase-structure trees, dependency trees encode syntactic aspects of
word groups (phrases), but the aspects they emphasize differ. Phrase-structure trees specify
the subphrases that a phrase is composed of, for instance, a verb phrase consisting of a verb
followed by a noun phrase. Phrases typically include one word, called the Head, that is more
important than the other subphrases, called dependents. In the example in Figure 1, the verb
Table 1.
Exemplar sentence and word list in Dutch and the literal English translation.
Sentence
Het aardige vrouwtje gaf Henk die een kleurige papegaai gekocht
Word list
zak een kleurige aardige een had die vrouwtje papegaai gaf het
had een zak pitjes.
gekocht pitjes
The nice lady gave Henk, who had bought a colorful parrot, a bag of
bag a colorful nice a had who lady parrot gave the bought
seeds.
seeds
Dit zijn geen regionale problemen zoals die op de Antillen.
zoals geen die Antillen problemen regionale zijn de dit op
These are not regional problems such as those on the Antilles.
such as not those Antilles problems regional are the these on
Neurobiology of Language
580
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
functions as the head because it imposes properties on the noun phrase rather than vice versa:
It assigns accusative case to the noun phrase. Phrase-structure and dependency trees are sim-
ilar because properties encoded by the former usually can be derived from the latter, and
vice versa. We have opted for dependency trees for a practical reason—the availability of a
high-quality dependency parser for Dutch.
Relative clauses are often ambiguous between a restrictive and a non-restrictive/appositive
semantic interpretation. However, we wish to note that these differing interpretations are not
associated with differing syntactic structures (see the analysis in Arnold, 2004, p. 39ff, which
applies to Dutch as well). This means that the parse tree computed by the participants does not
depend on the interpretation they choose. The complexity measure is thus not affected by this
ambiguity. We estimate that in at most five sentences in the materials, the relative clauses
needed a restrictive interpretation in order to make sense. In the written sentences, we did
not use commas to separate the relative clauses from their antecedents, thus leaving the inter-
pretation to the preference of the participants. (In Dutch spelling, such commas are optional,
like in English; but unlike in German, where they are obligatory, irrespective of interpretation.
In the spoken sentences, the sentences were pronounced with a prosodic pattern that was
neutral between the interpretations.) In order to calculate the right-branching sentence com-
plexity measure, we took the maximum of the word-by-word right-branching complexity as
the right-branching complexity of that sentence. Thus, in other words, the right-branching
complexity measure is the maximum number of simultaneously open right-branching
dependencies.
For both left- and right-branching complexity calculations, we focused on verbal heads
since they head clauses. Clausal structure is more encompassing than the more local struc-
ture of phrases headed by other parts of speech (e.g., noun phrases, prepositional phrases,
adjectival phrases). To verify this for our own material, we compared the left-branching com-
plexity between sentences, with and without relative clauses, for verbal and non-verbal
heads. For verbal heads, left-branching complexity was higher for sentences with relative
clauses (mean = 3.0 +/− 0.7 standard deviations) than sentences without (mean = 1.4 +/−
0.9 standard deviations). For non-verbal heads, there was no significant difference between
sentences with relative clauses (mean = 1.9 +/− 0.5 standard deviations) and those without
(mean = 2.1 +/− 0.5 standard deviations). It is assumed that clausal structure exerts the main
influence on cognitive complexity. Note that although our complexity measures are
designed to index processing complexity, for the sake of brevity we will refer to the outcome
of calculations using these measures as left/right-branching complexity.
Task and Procedure
Within a measurement session, the stimuli were presented in a mini-block design, and alter-
nated between a sentence block (5 sentences) and a word list block (5 word lists), for a total of
24 blocks. The type of starting block (sentences or word lists) was counterbalanced across
subjects.
Comprehension questions
In order to check for compliance, 10% of the trials were followed by a yes/no question about
the content of the just-presented sentence/word list. Half of the questions on the sentences
addressed the content of the sentence (e.g., Did grandma give a cookie to the girl? ) whereas
the other half, and all of the questions on the word lists, addressed one of the main content
words (e.g., Was the word ‘grandma’ mentioned? ). Half of the questions on complex relative
Neurobiology of Language
581
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
clause sentences concerned the content from the relative clause, to make sure the participants
comprehended all parts of the sentence. Subjects answered the question by pressing a button
for “Yes”/“No” with their left index and middle finger, respectively. Based on the answers to the
catch trial comprehension questions, subjects were defined as outliers (i.e., subjects with a
negative distance >1.5 times the interquartile range, from the mean) and were excluded from
further analysis, as not compliant with the task. This procedure resulted in a threshold of
59% percent correct, which all 204 included subjects passed (mean 80% correct +/− standard
deviation of 9%).
Stimulus presentation
For visual stimulus presentation, sentences/word lists were presented word-by-word with a
mean duration of 351 ms for each word (minimum of 300 ms and maximum of 1,400 ms,
depending on word length, etc., see further final paragraph of this section). Corresponding
visual sentences and word lists had the same total duration. The median duration of whole
sentences/word lists was 8.3 s (range 6.2–12 s). Auditory sentences had a median duration
of 4.2 s (range 2.8–6.0 s), and were spoken at a natural pace. The matching word list words
were also read at a natural pace, with a brief pause between words, averaging 7.7 s (range 5.5–
11.1 s) per word list.
Each participant was presented with 60 sentences and 60 word lists, either entirely in the
visual or the auditory modality. The full stimulus material consisted of 180 sentences and 180
word lists, divided into three subsets such that each participant saw 2/3 of the stimuli set in the
MEG session (120 trials of each condition) and 1/3 in the fMRI session (60 trials). Across par-
ticipants, each subset was presented as many times in MEG as in fMRI.
Each participant was presented with each stimulus once, in either the sentence or the word
list condition, but not in both. The presentation of the sentence and word list versions of the
items was counterbalanced across subjects. The auditory versions of the stimuli were recorded
by a female native Dutch speaker. The word lists were pronounced with a neutral (i.e., flat)
prosody and a clear pause (at least 100 ms for the auditory word lists and around 300 ms for
the visual word lists) between each word.
To indicate sentence and word list blocks, the start of each block began with a 1,500 ms
block type indication, zinnen ‘sentences’ or woorden ‘words’. Only in sentences did the first
word begin with a capital letter, while the last word ended with a full stop. The intertrial inter-
val was jittered between 3,200–4,200 ms. During this period, an empty screen was presented,
followed by a fixation cross.
The visual stimuli were presented with a LCD projector (vertical refresh rate of 60 Hz) sit-
uated outside the fMRI scanning room, and projected via mirrors onto the screen inside the
measurement room. All stimuli were presented in a black monospaced font on a gray back-
ground at the center of the screen within a visual angle of 4 degrees using Presentation soft-
ware ( Version 16.0, Neurobehavioral Systems, Inc.; https://www.neurobs.com/). For auditory
stimulus presentation, sounds were presented via plastic tubes and ear pieces to both ears.
Before the experiment, the hearing threshold was determined individually, and the stimuli
were then presented at an intensity of 50 dB above the hearing threshold, with the obtained
volume individually pretested on top of the echo planar imaging (EPI) sequence noise, to verify
that all stimuli were clearly audible.
Each word was separated by an empty screen for 300 ms before the onset of the next word.
The presentation time of each word was weighted by the number of letters in the word, pro-
viding a natural reading experience (c.f. Nieuwland & Van Berkum, 2006). In this way reading
Neurobiology of Language
582
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
of the words was also matched to some extent to the total duration of the audio version of the
stimuli. For any given sentence (or word list) the variable presentation duration of a single
word was a function of the following quantities: (i) the total duration of the audio version of
the sentence/word list (audiodur), (ii) the number of words in the sentence (nwords), (iii) the
number of letters per word (nletters), and (iv) the total number of letters in the sentence (sumn-
letters). Specifically, the duration (in ms) of a single word was defined as: (nletters/sumnletters) *
(audiodur + 2000 − 150 * nwords). It is reasonable to include both number of words and
number of letters in this formula for adaptive rapid serial visual presentation, as previous
research found decreased task load ratings for adaptive compared to fixed rapid serial visual
presentation (i.e., with fixed presentation time for each word), using both parameters (Ö quist &
Goldstein, 2003). The minimum duration of short words was set to 300 ms irrespective of the
relative weighting described by the formula. While there are limitations to this method, as it
leaves out multiple factors known to affect reading time (e.g., word frequency), it is an ade-
quate approximation suitable for languages with fairly regular grapheme–phoneme correspon-
dences such as Dutch (as opposed to, e.g., English). Note that this formula does not apply to
the auditory sentences, which were spoken at a natural pace.
MRI Data Acquisition
The data were acquired with a SIEMENS Trio 3T scanner using a 32-channel head coil. We
used a whole head T2*-weighted echo-planar blood oxygenation level-dependent (EPI-BOLD)
sequence. The single-echo 2D ascending slice acquisition sequence (partial brain coverage)
had the following specifications: Volume TR = 2.00 s, TE = 35 ms, 90 degree flip-angle,
29 oblique slices, slice-matrix size = 64 × 64, slice thickness = 3.0 mm, slice gap 0.5 mm,
FOV = 224 mm, voxel size (3.5 × 3.5 × 3.0 mm3 during acquisition; interpolated to 2 × 2 ×
2 mm3 in SPM). The number of volumes acquired per participant was around 650 for the
auditory group and around 850 volumes for the visual group. The exact number of volumes
varies slightly (e.g., 10 volumes) per participant due to different response times to the com-
prehension questions.
For the structural MR image volume, a high-resolution T1-weighted magnetization-
prepared rapid gradient-echo pulse sequence was used (MP-RAGE; TE = 3.03 ms, 8 degree
flip-angle, 1 slab, slice-matrix size = 256 × 256, slice thickness = 1 mm, FOV = 256 mm, isotropic
voxel-size = 1.0 × 1.0 × 1.0 mm3).
Preprocessing
The primary imaging data were checked for quality, including checks of subject movement
and signal drop out. The data were preprocessed with statistical parametric mapping software
(SPM8; Welcome Trust Centre for Neuroimaging, London, UK; www.fil.ion.ucl.ac.uk/spm),
which was also used for statistical analysis at the first- and second-level. Coordinates are
reported in Montreal Neurological Institute (MNI) space throughout.
The first three EPI-BOLD volumes were removed to ensure T1-equilibrium. The remaining
volumes were (1) realigned to correct for individual subject movement and (2) corrected for
differences in slice-acquisition time.
The mean EPI-BOLD volumes were co-registered to the structural image (i.e., the structural
image was the reference and the mean EPI-BOLD volume was the source image), and this
transformation was then applied to all EPI-BOLD volumes. Structural images were spatially
normalized to the structural image (T1) template provided by SPM8, using affine regulariza-
tion. The transformation matrices generated by the normalization algorithm were applied to
Neurobiology of Language
583
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
the corresponding functional EPI-BOLD volumes. All structural and functional images were
spatially filtered with an isotropic 3D spatial Gaussian filter kernel (full width at half maximum;
FWHM = 10 mm).
Statistical Analysis
The fMRI data were analyzed statistically, using the general linear model framework and sta-
tistical parametric mapping (Friston et al., 2007) in a two-step mixed-effects summary-statistics
procedure.
First-level models
The single-subject fixed effect analyses included a temporal high-pass filter (cycle cut-off at
128 s) to account for various low-frequency effects. All linear models included the six realign-
ment parameters from the movement correction. All models included two regressors modeling
the sentence and word list conditions from the onset of the stimulus throughout the duration of
its presentation and the following three regressors: the intertrial interval, the interblock interval,
as well as the comprehension questions. This minimal model (a part of all models) was used to
probe the language/sentence processing network (compared to a low-level baseline, more spe-
cifically the interblock interval) as a localization of language relevant regions. Intertrial and
interblock intervals were modeled separately as we wanted to use the interblock intervals
as a low-level baseline potentially less affected by lingering processing of the stimulus after
its presentation. Comprehension questions were modeled from the onset of the question until
after the reply (hence including the reply). Additional models (including additional regressors)
as well as contrasts are described in more detail below.
Left- vs. right-branching complexity. Regressors containing two sets (corresponding to sentences
and word lists, respectively) of five parametric sentence/word list modulators, included: (1)
number of letters; (2) number of syllables; (3) number of words; all of them referred to as con-
trol regressors; (4) the right-branching complexity; and (5) the left-branching complexity. The
timing of these modulators does not differ from the timing of the sentences/word lists they
modulate (i.e., from the onset of the stimulus throughout the duration of its presentation).
All of the regressors were used both for the auditory and visual analyses. These parametric
modulators were entered as user-specified regressors after convolution with the canonical
hemodynamic response function (HRF). To control for lexical level features, we associated
each scrambled word list version of a sentence with the left- and right-complexity measure
of that sentence and contrasted complexity for sentences and word lists (using the contrasts
[0 0 0 0 0 0 0 0 1 0 0 0 0 −1 0]; [0 0 0 0 0 0 0 0 0 1 0 0 0 0 −1]; where the first five columns are
described in the First-level models section, the next five columns are the modulators of sen-
tences, and the last five are modulators of word lists). To verify the expectation that complexity
should affect sentences and not unstructured word lists, we also created contrasts probing the
sentences and word lists separately, against the implicit baseline (using the contrasts [1 0 0 −1];
[0 1 0 −1]). We also formed the reverse contrasts (word lists > sentences, i.e., negative effects of
complexity) to those described in this paragraph.
In order to analyze activation changes
Left-branching processing complexity: Temporal dynamics.
over sentences, we performed two analyses: One was a contrast analysis with four time bins
and one a finite impulse response (FIR) analysis with seven time points. Note that there is no
exact correspondence between words or other structures and time bins/time points. We used
the complexity measures calculated on the whole sentence level in these analyses.
Neurobiology of Language
584
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
The first analysis divided each sentence/word list presentation into four equally long time
bins. For the time-bin analysis, parametric modulators were specified using pmod (parametric
modulation) functionality implemented in SPM8. The order in which regressors are entered
matters for this analysis, since each pmod regressor is orthogonalized with respect to the so
called unmodulated regressors, as well as earlier pmod regressors (Mumford et al., 2015). Con-
trol regressors (i.e., numbers of words, letters, and syllables) were thus entered before the
regressors encoding the complexity measures, and the right-branching complexity measure
was entered before the left-branching complexity measure, when assessing the effects of
left-branching complexity and, conversely, when assessing the effects of right-branching
complexity.
We focused on the two ROIs that were most clearly and selectively responsive to left-
branching complexity: the LIFG and LpMTG. To assess increases and decreases over the
course of the sentences, we formed first-level linear contrasts across the complexity regressors
corresponding to each time bin (e.g., [−2 −1 1 2]).
In the second analysis, time courses were extracted from the LIFG and LpMTG ROIs using
the FIR model as implemented in MarsBar (Brett et al., 2002). The FIR model does not assume
any particular shape of the hemodynamic response, but estimates the signal at each time bin
(henceforth called time windows) separated by one TR (in our case 2 s), up to 24 s after stim-
ulus onset, adjusting for effects of stimuli overlapping in time. As the naturally spoken sen-
tences created shorter auditory presentation times, we used the visual sample in this analysis.
Since our average visual stimulus duration was 8.3 s and our average intertrial interval was
3.7 s, we analyzed the first seven time windows, covering 0–14 s post stimulus onset. We
analyzed the differences in percent signal change between high (left-branching complexity
>=3) and low (left-branching complexity <3) complexity, by introducing two sets of regressors
(high/low) for sentences and word lists in the FIR model. The differences in percent signal
change for high–low complexity were tested with a paired t test at all time points. We cor-
rected for multiple comparisons over seven time points using Bonferroni correction, resulting
in a corrected alpha level of 0.05/7 = 0.007. In a follow-up analysis, we tested whether the
complexity effects came in different time windows for LpMTG, compared to LIFG. We com-
pared the difference scores of the percent signal change for high complexity sentences – low
complexity sentences, for LpMTG-LIFG, using a paired t test.
Second-level statistics and visualization
The second level comparison used a two-sample unequal-variance t test. Visual and auditory
data were modeled separately and then compared using an additional model. Five such
models were created: One of each is reported in the first three sections of the Results and
two models (where one was a FIR model) in the fourth section of the Results. (See the
supplement for additional results.) Conjunctions were tested against the conjunction null
(Nichols et al., 2005). As we used the conservative version of conjunction analysis, we are
only observing voxels/regions where the null hypothesis is rejected in both modalities.
Throughout the article, statistics are reported with the following criterion for significance: pres-
ence of family-wise error (FWE) corrected voxels or clusters at pFWE < 0.05, both in the whole
brain analysis and in the ROI analysis, using a threshold of p < 0.005 uncorrected. In other
words, while significant clusters and peak voxels often coincide, we report clusters even if
there were no significant peak voxels in them, and we report a peak voxel even if it would
be outside of a cluster. In tables, all clusters significant at pFWE < 0.05 on the whole brain level
are reported including locations of voxels belonging to a cluster. Location and statistics of sig-
nificant peak voxels (within or outside significant clusters) are also reported.
Neurobiology of Language
585
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
ROI analysis
In the ROI analysis, we used 10 mm spherical ROIs around meta-analytically established acti-
vation hotspots of semantic and syntactic unification, reported in Talairach space (Hagoort &
Indefrey, 2014). The two top coordinates for semantic and syntactic unification were close to
each other, one pair in the LIFG and another in the LpMTG. We used the mean coordinate
between the syntactic and semantic peak for our ROI analysis. This resulted in the (Talairach
space) coordinates [x y z] = [−45 20 10] for LIFG and [−50 −45 5] for LpMTG, which we then
transformed to MNI space using the Talairach Daemon (https://www.talairach.org/).
In addition, a LaTL ROI was investigated in the left- vs. right-branching complexity. This was
again a 10 mm spherical ROI around the peak coordinate reported for the building of sentence
structure in a related study (Brennan et al., 2012). These were [x y z] = [−50 11 −16] in MNI
space.
Anatomical inference
All local maxima are reported as MNI coordinates. Assignment of clusters and voxels to gyri,
sulci, and Brodmann areas (for LIFG) were done using the Anatomy toolbox (Version 21)
implemented in SPM (Eickhoff et al., 2005).
RESULTS
Sentence Processing vs. Low-Level Baseline
We here present the second-level analysis of the per-subject generated first-level contrast.
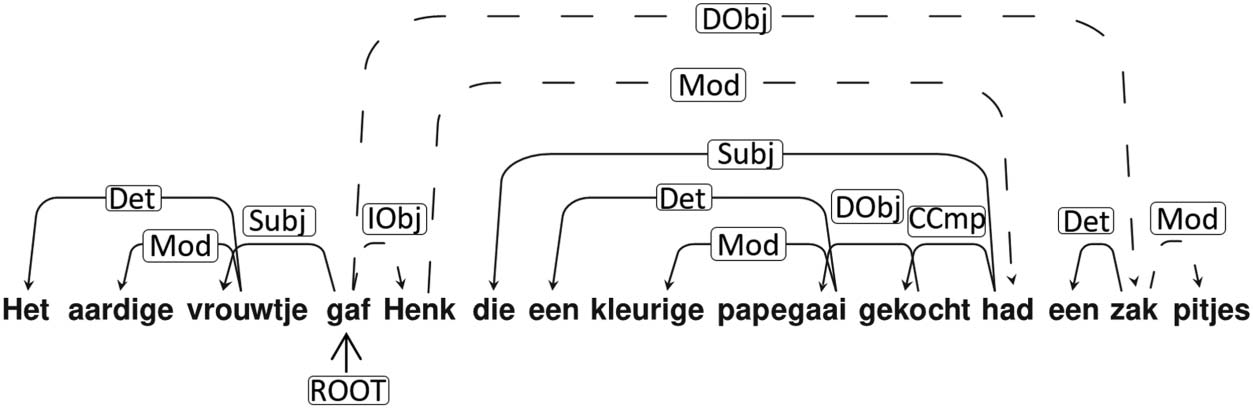
We verified that our paradigm activated the classical perisylvian language network by
comparing activation during sentence processing with a low-level baseline (i.e., fixation/rest
interblock interval). A significant cluster spanning left anterior to posterior superior temporal
cortices (middle and superior temporal gyrus), left inferior frontal gyrus (BA 44/45), the left
inferior parietal lobe (LIPL), and left fusiform gyrus was observed (see Figure 2A). Two addi-
tional clusters (right superior temporal and right fusiform gyrus) as well as significant voxels in
left orbitofrontal cortex and in left precentral and left superior frontal gyrus were also observed.
Left-Branching Complexity
To investigate supramodal unification (Hagoort, 2005; Vosse & Kempen, 2000), we created
conjunctions over visual and auditory contrasts of parametrically varied sentence complexity.
We controlled for lexical features correlated with sentence complexity by subtracting corre-
sponding word list parameters in the following way. Each scrambled word list version of a
sentence was associated with the left- and right-branching complexity measure of that sen-
tence. We analyzed the parametric effects of left-branching complexity for sentences com-
pared to the corresponding effects for word lists (henceforth referred to as sentences > word
lists contrasts).
There was a significant interaction between the parametric effect of left-branching com-
plexity in sentences compared to word lists throughout the left perisylvian language network
(Figure 2B and Table 2). In this comparison, a significant cluster was observed in posterior
middle/superior temporal cortices, anterior superior temporal gyrus (anterior temporal lobe),
left inferior frontal gyrus (BA 44 and 45), and left inferior parietal cortex. There were no sig-
nificant effects in the reverse comparison, word lists > sentences. To consolidate the findings in
the sentences > word lists contrast, we verified that complexity had no effect on word lists by
Neurobiology of Language
586
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
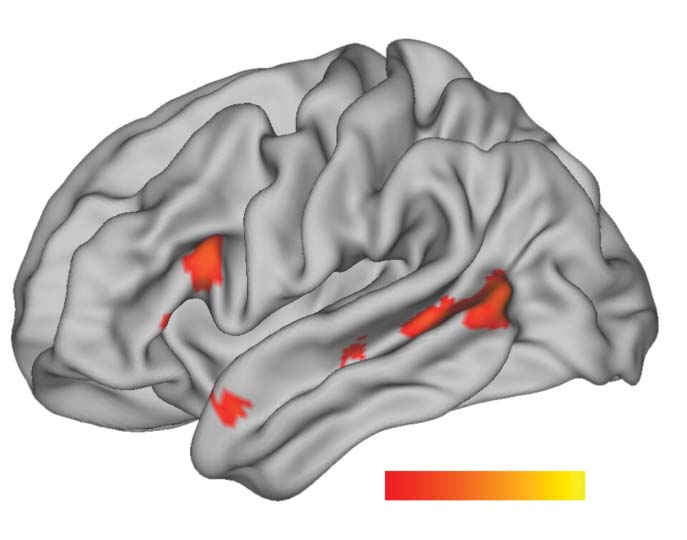
Figure 2. Results images were created based on the conjunction of data of the visual (N = 102)
and the auditory (N = 102) presentation of the materials, using a threshold of p < 0.005 uncorrected,
for the contrasts with significant results at the cluster or voxel level, using a criterion of pFWE < 0.05.
(A) Effect of sentences > low-level baseline. (B) Parametric effect of left-branching complexity in
sentences > word lists (word list complexity is the complexity of the corresponding sentence). Peaks
in a significant temporal cluster were observed in LpMTG and LaTL. An LIFG ROI contained a sig-
nificant cluster. (C) Parametric effect of left-branching complexity in sentences (i.e., without com-
paring word lists). Peaks in a large significant frontotemporal cluster, were observed in LpMTG,
LaTL, and LIFG. LpMTG: left posterior middle temporal gyrus; LaTL: left anterior temporal lobe;
LIFG: left inferior frontal gyrus.
investigating the effect of left-branching processing load in sentences and word lists separately.
There were no significant effects of complexity in the word list condition. In contrast, we
observed a positive effect of left-branching complexity for sentences in the left perisylvian lan-
guage network (Figure 2C and Table 2). There was no significant activity in the reverse
comparison.
Neurobiology of Language
587
Syntactic complexity in supramodal sentence processing
Table 2.
Supramodal (parametric) effect of left-branching complexity.
Cluster size
Cluster pFWE
MNI coordinates
y
x
z
Voxel pFWE
Voxel t201
Region
Left-branching*
Sentences > words
LpMTG/LaTL/LIFG/LIPL
2,678
0.001
LpMTG/STS
LaTL + LIFG (BA 44,45), LIPL
−52
−56
−30
2
−4
−16
0.003
0.042
5.13
4.47
Left-branching
Sentences
LpMTG/LaTL/LIFG/LIPL
4,225
<0.001
LpMTG/STS
LaTL
LIFG (BA45)
LIFG (BA44)
LIPL I
LIPL II
LIPL III
Right-branching
Sentences
−50
−54
−48
−56
−50
−54
−52
−36
8
28
18
−54
−56
−54
−2
−18
0
14
16
18
12
<0.001
<0.001
0.001
0.003
0.011
0.013
0.019
7.67
6.40
5.19
5.15
4.82
4.77
4.68
ROI LaTL, 10 mm, SVC
37
0.048
Note. No significant activations (at either ROI level) for the opposite contrasts, i.e., Left-branching, Words > Sentences; negative effect of Left-branching, Sen-
tences. LpMTG: left posterior middle temporal gyrus; LaTL: left anterior temporal lobe; LIFG: left inferior frontal gyrus; LIPL: left inferior parietal lobe; STS:
superior temporal sulcus; ROI: region of interest; SVC: small volume correction; FWE: family-wise error; MNI: Montreal Neurological Institute.
−54
8
−20
0.024
3.13
Across all contrasts, the number of modality-specific aspects were minimal. There were no
significant effects for visual > auditory or auditory > visual contrasts for left-branching com-
plexity in the sentences > word lists comparison. There were no significant activations for
auditory > visual for the left-branching complexity measure for sentences. There were, how-
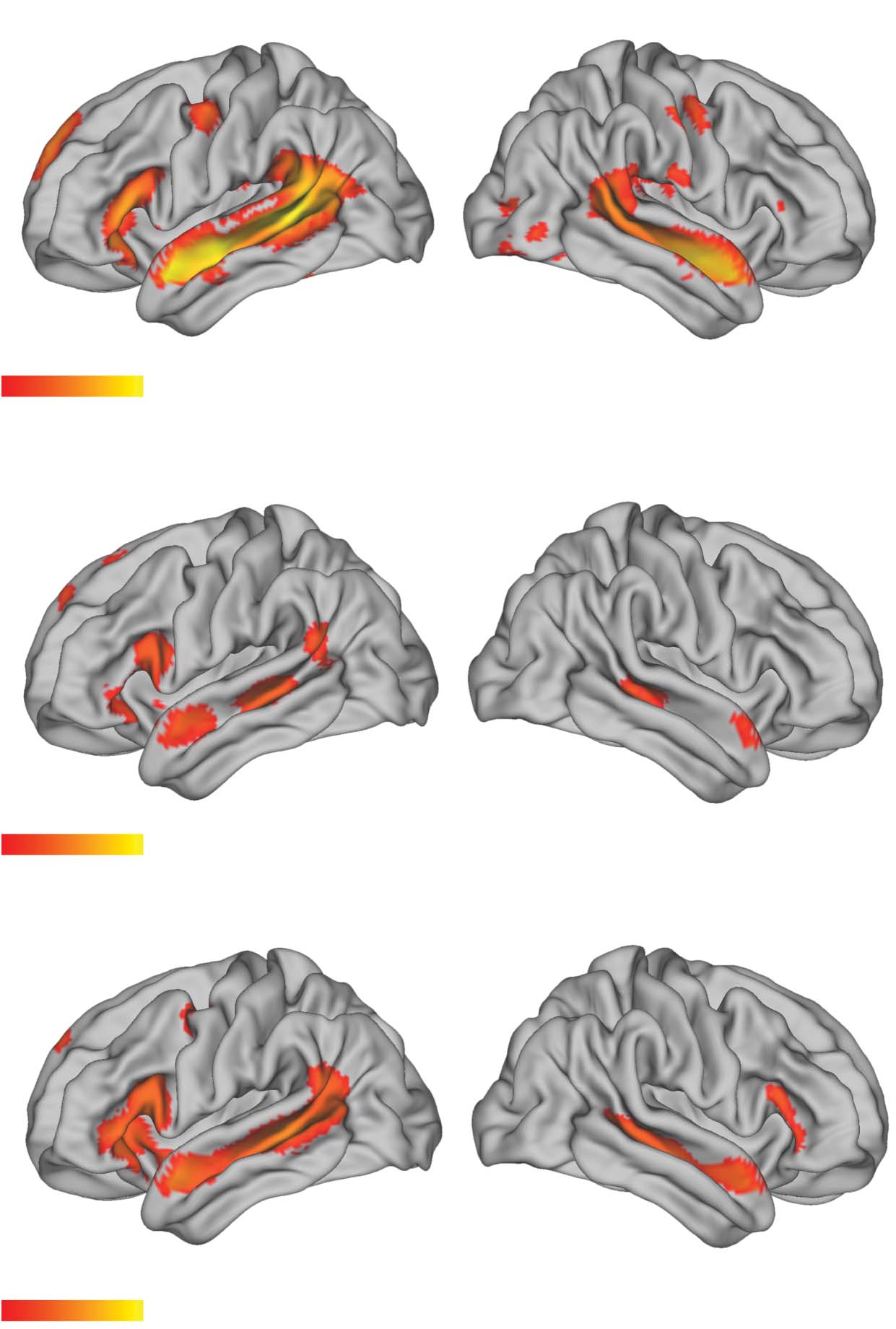
ever, visual > auditory effects for the left-branching complexity measure for sentences (Figure 3,
Table 3). There were clusters observed in the bilateral superior temporal gyrus (STG) / Heschl’s
gyrus and left subiculum. In a follow-up analysis, we tested whether the observed effects in this
visual > auditory contrast was due to an effect of left-branching complexity in the
opposite/negative direction (i.e., a higher BOLD response for lower complexity) for the audi-
tory group. This was the case (see Table 3).
Neurobiology of Language
588
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
Figure 3. Visual > auditory effect for the left-branching complexity measure, for sentences. There
was generally an absence of modality-specific effects across almost all contrasts reported, with this
contrast as one of the exceptions. In a follow-up analysis, we tested whether the observed effects in
this visual > auditory contrast was due to an effect of left-branching complexity in the
opposite/negative direction (i.e., higher BOLD response for lower complexity) for the auditory
group. This was the case (see Table 3).
Effects of Right-Branching Complexity
For the corresponding comparisons for right-branching complexity to those reported in the
Language Stimuli section, no significant effects were found for sentences > word lists nor
for word lists > sentences. For the separate sentence contrast, there were no significant voxels
or clusters. Our ROI analysis included three ROIs (LIFG, LpMTG, and LaTL). The LaTL ROI
contained a significant cluster for an increase in right-branching complexity. No significant
effects were observed for word lists nor for the corresponding comparisons in the reverse
direction.
There were no significant effects for visual > auditory or auditory > visual contrasts for the
right-branching complexity measure for sentences. There were no significant activations for
visual > auditory for right-branching complexity in the sentences > word lists comparison.
Left-Branching Complexity: Temporal Dynamics
Unification processes operate incrementally during word-by-word presentation of sentences.
The unification load will thus vary over time, in particular for complex sentences. To describe
the temporal dynamics of sentence unification, we divided each sentence and word list into
four time bins of equal length. We focused on the left-branching complexity measure, since it
showed the most prominent effects. For the ROI analysis, we used the two ROIs that were most
clearly active specifically to left-branching complexity: the LIFG and LpMTG. The right-
branching complexity parameter and control parameters (number of words, number of sylla-
bles, and number of letters) were entered into the model to ensure that we were studying the
contribution of left-branching complexity to dynamic changes over time. We created linear
contrasts probing monotonic increases and decreases in the parametric effect of the complex-
ity measures across the four time bins. We expected temporal dynamics related to sentence
unification to be stronger in sentences compared to word lists.
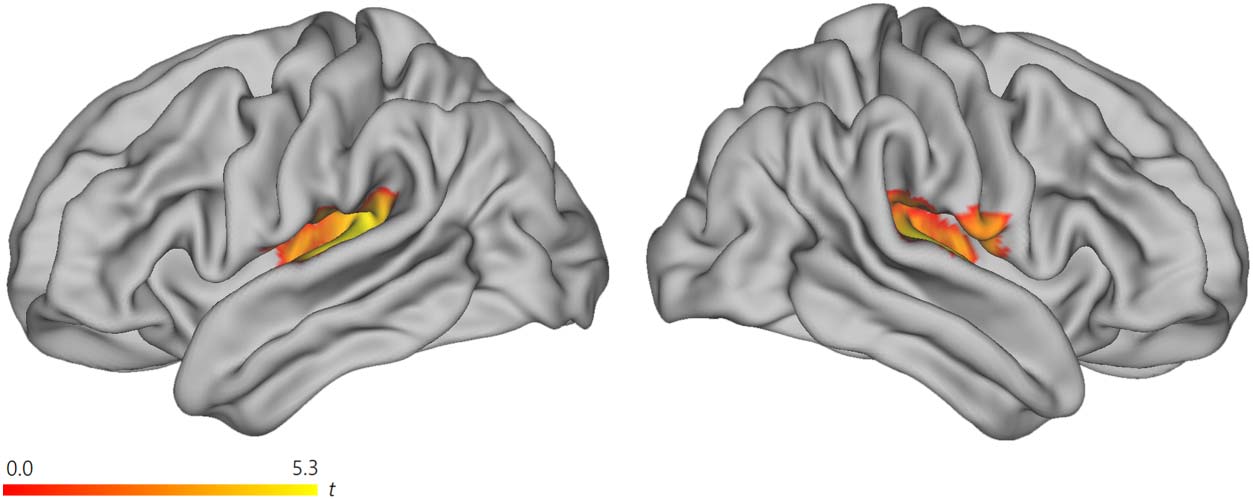
As shown in Figure 4A and Table 4, we observed a monotonic increase related to sentence
unification in the LpMTG ROI. When analyzing sentences and word lists separately, a mono-
tonic increase over sentences was observed in both the LIFG and LpMTG ROIs. No decrease
was observed. There were no effects of modality (visual > auditory or auditory > visual) for the
contrasts reported in this section.
Neurobiology of Language
589
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
Region
Left-branching
Sentences
Visual > auditory
LSTG/Heschl’s gyrus
Table 3. Modality specific effects left-branching complexity.
Cluster size
Cluster pFWE
MNI coordinates
y
x
z
Voxel pFWE
Voxel t201
1,575
0.011
RSTG/Heschl’s gyrus
1,764
0.009
Left subiculum
1,221
0.031
Right-branching
Sentences > word lists
Auditory > visual
48
44
−16
−24
−38
30
8
10
12
<0.001
0.001
5.74
5.44
<0.001
5.73
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
ROI LIFG 10 mm, SVC
36
0.048
−52
20
10
0.003
2.96
Left-branching
Negative direction
Sentences
Auditory
LSTG
1,988
0.004
RSTG
1,741
0.007
Left fusiform gyrus
−38
−48
50
−32
−30
−20
−16
−38
12
8
8
−10
<0.001
<0.001
<0.001
0.009
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
6.16
5.81
6.10
4.87
Note. No significant activations for visual > auditory or auditory > visual in: Left, Sentences > word lists; Right, Sentences. No significant activations for
auditory > visual in: Left, Sentences. No significant activations for visual > auditory in: Right, Sentences > word lists. LSTG: left superior temporal gyrus; RSTG:
right superior temporal gyrus; LIFG: left inferior frontal gyrus; ROI: region of interest; LIPL: left inferior parietal lobe; SVC: small volume correction; FWE: family-
wise error; MNI: Montreal Neurological Institute.
To follow up on this finding, we extracted percent signal change time courses (using a FIR
model) from the LIFG and LpMTG ROIs in the reading group (Figure 4B). The visual sample
has longer presentation times compared to the naturally spoken sentences. Thus, more robust
estimates of changes in the BOLD response over the sentence are expected for this part of the
sample. As the finite impulse response model allows the extraction of time courses from ROIs
Neurobiology of Language
590
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
.
/
l
Syntactic complexity in supramodal sentence processing
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
(A) In red, left-branching complexity: increase across four time bins for sentences. Results images were created based on the con-
Figure 4.
junction of the data for visual and auditory presentations of the materials, using a threshold of p < 0.005 uncorrected, for the contrasts with
significant results at the cluster or voxel level, using a criterion of pFWE < 0.05. In the LpMTG ROI, there was a significantly (using SVC) steeper
increase for sentences > word lists. (B) To illustrate the observed dynamics in (A) further, with time courses using a FIR model, we focused on
the written sentences and word lists which have longer presentation times for sentences (compared to the naturally spoken sentences) and thus
provides more robust estimates of changes in the BOLD response over the sentence. Complexity difference scores (high–low) of % signal
change from LIFG and LpMTG ROIs, in sentences and word lists are plotted over seven time windows. Note that we are thus contrasting
sentences against sentences and word lists against word lists (not sentences against word lists). Red shaded areas represent +/− 1 standard error
of the mean around each data point for sentences and likewise for word lists, in green shade. The % signal change is relative to the mean
activity of this ROI in the whole session; thus, note that the first time point (0–2 s) can be active since a few words were already presented
during this time. The complexity effect within sentences was significant at 8–10 s in LpMTG (t101 = 3.28, pBonf = 0.01) and at 10–12 s in LIFG
(t101 = 2.82, pBonf = 0.04). Both of these survive Bonferroni correction for testing seven time points. The seventh time point (12–14 s) contains
the intertrial interval, on average. All four mean time courses returned to the session mean at this time point. (C) The complexity effect for
sentences occurred in an earlier time window for LpMTG, compared to LIFG. There was a significant interaction between the sentence com-
plexity effect in LpMTG vs. LIFG, at 8–10 s vs. 10–12 s (t101 = 2.39, p = 0.02). The data in (C) are the same as in the corresponding time
windows in (B). LpMTG: left posterior middle temporal gyrus; LIFG: left inferior frontal gyrus; FWE: family-wise error; ROI: region of interest;
SVC: small volume correction; FIR: finite impulse response; BOLD: blood oxygen level-dependent.
and does not assume any particular shape of the hemodynamic response, this was an appro-
priate follow-up to investigate any potential differences in time courses between the ROIs and
to characterize the time courses in greater detail. We analyzed the differences in percent (%)
signal change between high (left-branching complexity >=3, 168 sentences) and low (left-
branching complexity <3, 192 sentences) complexity as a function of time. The complexity
effect (high–low) for sentences was significant at 8–10 s in the LpMTG ROI (t101 = 3.28, pBonf =
0.01) and at 10–12 s in the LIFG ROI (t101 = 2.82, pBonf = 0.04). We Bonferroni-corrected for testing
seven time points. The complexity effect for sentences thus occurred in an earlier time window in
LpMTG than in LIFG. There was a significant difference between the sentence complexity effect
in LpMTG–LIFG, at 8–10 s vs. 10–12 s (t101 = 2.39, p = 0.02, see Figure 4C). The seventh
time point (12–14 s) contains the intertrial interval.
Neurobiology of Language
591
Syntactic complexity in supramodal sentence processing
Table 4.
Supramodal (parametric) effect of left-branching complexity, increase across 4 time bins.
Cluster size
Cluster pFWE
MNI coordinates
y
x
z
Voxel pFWE
Voxel t201
Region
Left-branching
Sentences > words
ROI LpMTG 10 mm, SVC
74
0.034
−46
−42
0
0.025
3.29
Left-branching
Sentences
ROI LpMTG 10 mm, SVC
242
0.029
no significant peak voxels
ROI LIFG 10 mm, SVC
−50
20
16
0.028
3.25
Note. No significant activations (at either ROI level) for: any of the corresponding decreases; Right, Sentences > words; Right, Sentences. Furthermore, there
were no effects of modality (visual > auditory or auditory > visual). ROI: region of interest; LpMTG: left posterior middle temporal gyrus; SVC: small volume
correction; LIFG: left inferior frontal gyrus; FWE: family-wise error; MNI: Montreal Neurological Institute.
DISCUSSION
Our study provides support for the hypothesis that unification should be understood as a
supramodal process, common to sentences presented as speech or text. The current results
show that the supramodal network subserving unification includes the LIFG, the bilateral
MTG (both anterior and posterior parts), as well as LIPL. We observed an effect of left-
branching complexity in the perisylvian language regions, irrespective of modality. Moreover,
we observed an absence of modality specific effects related to unification processes (i.e., no
effects when comparing visual > auditory or auditory > visual samples). This finding provides
further support for unification as a process that is not modality specific. Our results thus sup-
port a view where the visual and auditory input streams converge in a supramodal unification
network, most clearly in LIFG and LpMTG.
Previous studies comparing complex vs. simple sentences have implicated the inferior fron-
tal and temporal heteromodal association cortices as supramodal regions associated with sen-
tence structure manipulations (Braze et al., 2011; Constable et al., 2004; Michael et al., 2001;
Shankweiler et al., 2008). Our large sample size and the use of parametrical variation of pro-
cessing complexity increase both sensitivity and validity compared to these previous studies.
Our results support the view that LIFG and the bilateral MTG (both anterior and posterior parts)
are key nodes of the supramodal unification network. Moreover, our results suggest that addi-
tional regions (e.g., LIPL; see further discussion below) could be considered a part of this net-
work (Binder et al., 2009). These regions correspond well to the overlap of written and spoken
comprehension studies on sentences reported in a recent meta-analysis study (Walenski et al.,
2019) and in a study using contrasts of speech vs. backwards speech and written sentences vs.
scrambled written sentences (Wilson et al., 2018). The results are largely in agreement with the
findings reported by Friederici and colleagues (for an overview of these findings, see Friederici,
2017), who also found complexity effects in both LIFG and posterior temporal cortex. In addi-
tion, Friederici (2017) refers to activations in anterior STG as related to local structure building,
which presumably refers to the structure building at the phrasal level that others (Snijders
et al., 2009) have found to be related to LpMTG.
Neurobiology of Language
592
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
The effects of left-branching complexity show increased activation levels in mainly left peri-
sylvian language regions for more syntactically complex sentences. Moreover, the results show
that the effect of left-branching processing complexity in LIFG and LpMTG increased toward
and even beyond the end of the sentence. The corresponding changes were nonsignificant
for LIPL, and similar sentence progression effects were not observed for the right-branching
structures. Thus, in summary, left-branching complexity effects are stronger and show different
dynamics over the sentence compared to right-branching complexity effects. This highlights
the fact that the neuronal underpinnings of sentence processing operations cannot be fully
appreciated without understanding how a complexity measure indexes the sentence processing
operations. Another good example of this approach can be found in a recent ECoG study on
visual sentence processing (Nelson et al., 2017; see also Fedorenko et al., 2016). By using many
processing models to predict neural activity over the sentence, they found evidence for a struc-
ture building process in LIFG and posterior superior temporal cortices. It is possible that a sub-
stantial part of the variance in the neural dynamics explained by our left-branching complexity
measure is shared with their word-by-word increases in high gamma for every word in a
constituent. Our results complement their results in five ways: (1) by showing a functional asym-
metry in neural processing of dependencies that go from the head to the dependent (i.e., left-
branching) compared to the other way around; (2) by showing the spatial specificity of these
effects with fMRI; (3) by a 15 fold increase in the number of participants to ensure robust effects;
(4) by increasing the variability of the sentence structures used, since Nelson et al. (2017)
restricted their sentence materials to only a single sentence structure; and finally (5) by extending
these results from the visual modality only to the auditory modality as well, showing that the
effects of left-branching complexity are likely to be supramodal. The absence of modality spe-
cific effects for the temporal dynamics analyses supports this conclusion. While it is easier to
compare the increased response over the sentence to the oscillatory results discussed above,
our results may also be compared with the results of Pallier et al. (2011). They found that fMRI
responses got increasingly slower for larger constituent sizes in the more posterior LIFG pars
triangularis, but not in the more anterior LIFG pars orbitalis (matching the posterior location of
our LIFG finding; see Figure 4A), as well as for similar locations to ours in anterior and posterior
superior temporal sulcus (pSTS). In addition, Regev et al. (2013) report highly similar time courses
for spoken and written narratives in LIFG and pSTS (and additionally in precuneus and angular
gyrus, regions that might be attributed to processing of their narrative stimuli). Below we interpret
the results by focusing on the operations targeted by our left-branching complexity measure, using
the effects of right-branching complexity for contrast and comparison.
A crucial aspect of language processing is the binding of the head of a verb phrase to its
arguments. At least two factors determine the processing complexity of unifying a verb phrase.
One is the position–distance between the head and its arguments in a sentence. The second is
the order of heads and arguments. There are several possible reasons for this, such as an addi-
tional processing cost of storing unbounded arguments, as well as an asymmetry in predictive
power (the head is a stronger prior for arguments than vice versa). In our study we computed a
complexity measure for each individual sentence in the experiment, based on a formal char-
acterization of the processing consequences of simultaneous unbounded arguments. This
complexity measure modulated activity in LIFG and LpMTG in a parametric way, strongly sug-
gesting that these regions together play a crucial role in the unification of lexical elements (i.e.,
words) into an overall structural sentence configuration.
When studying the activation dynamics across the sentence in the different nodes of the
supramodal unification network, we observed a particularly interesting pattern of results in
LIFG and LpMTG. In these regions, activity increased toward the end of the sentence for
Neurobiology of Language
593
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
left-branching structures. No such effect was observed in LIPL (see Figure 4). We calculated the
maximum point of simultaneous unification of unbounded arguments (usually referred to as
dependents for other phrases, e.g., noun phrase, than verb phrases), which occurred on aver-
age around halfway into our sentences. Furthermore, the left-branching complexity measure
indexes sentences with many simultaneous non-attached constituents, a cost which increases
with each non-attached constituent presented, until a culmination point around halfway into
the sentences. Since the priors for unification are relatively low in left-branching dependencies
compared to right-branching dependencies, increased activation is expected toward the later
parts of the sentence, given the increased number of unbounded arguments. In addition to this
online processing complexity, new elements to the sentence structure are incrementally added
and maintained. Hence the overall complexity of the output of online parsing operations is
highest at the end of the sentence. This seems to be especially reflected in the IFG activation.
An additional novel finding is that the complexity effect had an earlier time course in LpMTG
than in LIFG. Both areas contribute to syntactic processing (Hagoort & Indefrey, 2014; Matchin
& Wood, 2020). The earlier complexity effect in LpMTG might also include the consequences
of the online processing complexity, next to the full sentence complexity that seems to dom-
inate LIFG. Another possibility is that there are physiological delays, for example, in the met-
abolic consequences of neural activity that we observe through the hemodynamic response,
that differ across brain regions. This is something to keep in mind when interpreting the result
of the FIR analysis, which does not assume any particular shape of the HRF. Further explora-
tion using additional modalities might be warranted: We note that recent meta-analytic results
on written, spoken, and signed language suggested posterior LIFG in particular as an amodal
hub (see Trettenbrein et al., 2021).
Right-branching complexity produced stronger activation than left-branching complexity in
LaTL. This could be due to the stronger expectations about upcoming lexical-semantic infor-
mation in the former case. This view on LaTL contribution to structure building fits well with
the current views on the anterior temporal lobe function within linguistic and conceptual pro-
cessing more generally (Patterson et al., 2007). There is, furthermore, evidence that conceptual
combinations are processed in the anterior temporal lobe (e.g., Baron & Osherson, 2011). In
line with our view, Pallier et al. (2011) only observed structure building (as indexed by an
increasing response to increasing constituent size) in LaTL for materials with real words,
and not in a Jabberwocky condition, where only posterior temporal and inferior frontal regions
were observed (see also Goucha & Friederici, 2015).
The measure of simultaneous non-local dependencies that we used correlates with the
presence of non-local dependencies as well as their total length. Our results thus show the
neural underpinnings of the ubiquitous difficulty observed with processing non-adjacent
dependencies (Gibson, 1998; Grodner & Gibson, 2005). In addition, our results reflect the
increased difficulty of unifying complex left- compared to right-branching sentence aspects,
at least in the context of a language like Dutch, which features considerable proportions of
both. Our results suggest that simultaneity (or overlap) of multiple unresolved non-adjacent
dependencies, rather than linear distance of non-adjacent dependencies, is a major factor con-
tributing to the difficulty of processing non-adjacent dependencies (see additional analyses
supporting this conclusion in the supplementary material). This highlights the importance of
understanding which sentence processing operation(s) a complexity measure taps into. Previ-
ous literature indeed shows neurocognitive segregation of processing operations, drawing on
computational models quantified as different (complexity) measures calculated on words (e.g.,
Bozic et al., 2010), sentences (e.g., Lopopolo et al., 2021), or narratives (e.g., Bhattasali et al.,
2019; Willems et al., 2016).
Neurobiology of Language
594
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
In conclusion, we have characterized the basic spatiotemporal dynamics of a supramodal
unification network consisting of the LIFG, bilateral MTG, and the LIPL. The finding that activ-
ity in this network increases over the sentence is novel (see possibly related oscillatory mod-
ulations in ECoG and MEG, respectively: Fedorenko et al., 2016; Lam et al., 2016). These
results support our interpretation that binding of unbounded arguments increases toward the
end of the sentence, resulting in stronger activations in LIFG and LpMTG. We also show the
neural underpinnings of the ubiquitous difficulty observed with processing overlapping non-
adjacent dependencies is independent of modality. Both listening to speech and reading acti-
vate the same neuronal circuitry once the modality-specific input is mapped onto lexical
items, in the service of the brain’s capacity to make sense beyond the processing of single
words.
ACKNOWLEDGMENTS
We thank Laura Arendsen, Manuela Schuetze, Tineke de Haan, and Charlotte Poulisse for
assisting with stimuli construction, participant recruitment, data collection, and preprocessing.
FUNDING INFORMATION
This work was supported by the Max Planck Society. Julia Uddén, Riksbankens Jubileumsfond
(https://dx.doi.org/10.13039/501100004472). Julia Uddén, Swedish Collegium of Advanced
Studies, the Swedish Brain Foundation (Hjärnfonden postdoc grant, to J.U.). Peter Hagoort,
the Spinoza Prize. Peter Hagoort, the Royal Netherlands Academy Professorship Prize. Peter
Hagoort, the NWO Grant Language in Interaction, grant number 024.001.006.
AUTHOR CONTRIBUTIONS
Julia Uddén: Conceptualization; Data curation; Formal analysis; Investigation; Methodology;
Project administration; Resources; Software; Writing – original draft; Writing – review & edit-
ing. Annika Hultén: Conceptualization; Data curation; Investigation; Methodology; Project
administration; Software. Jan-Mathijs Schoffelen: Resources. Nietzsche Lam: Data curation;
Investigation; Project administration; Software. Karin Harbusch: Resources. Antal van den
Bosch: Resources. Gerard Kempen: Resources. Karl Magnus Petersson: Conceptualization;
Methodology; Resources. Peter Hagoort: Conceptualization; Methodology; Writing – original
draft; Writing – review & editing.
REFERENCES
Altmann, G. T. M. (1998). The ascent of Babel: An exploration of
language, mind, and understanding. Oxford University Press.
https://doi.org/10.1093/acprof:oso/9780198523772.001.0001
Arnold, D. (2004). Non-restrictive relative clauses in construction
based HPSG. In S. Müller (Ed.), Proceedings of the 11th inter-
national conference on head-driven phrase structure grammar
(pp. 27–47). OJS. https://doi.org/10.21248/hpsg.2004.2
Baggio G., & Hagoort P. (2011). The balance between memory and
unification in semantics: A dynamic account of the N400. Lan-
guage and Cognitive Processes, 26(9), 1338–1367. https://doi.org
/10.1080/01690965.2010.542671
Bahlmann, J., Schubotz, R. I., & Friederici, A. D. (2008). Hierarchi-
cal artificial grammar processing engages Broca’s area.
NeuroImage, 42(2), 525–534. https://doi.org/10.1016/j
.neuroimage.2008.04.249, PubMed: 18554927
Baron, S. G., & Osherson, D. (2011). Evidence for conceptual com-
bination in the left anterior temporal lobe. NeuroImage, 55(4),
1847–1852. https://doi.org/10.1016/j.neuroimage.2011.01.066,
PubMed: 21281723
Bever, T. G., Carrithers, C., Cowart, W., & Townsend, D. J. (1989).
Language processing and familial handedness. In A. M. Galaburda
(Ed.), From reading to neurons (pp. 341–357). MIT Press.
Bhattasali, S., Fabre, M., Luh, W.-M., Al Saied, H., Constant, M.,
Pallier, C., Brennan, J. R., Spreng, R. N., & Hale, J. (2019). Loca-
lising memory retrieval and syntactic composition: An fMRI study
of naturalistic language comprehension. Language, Cognition
Neurobiology of Language
595
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
and Neuroscience, 34(4), 491–510. https://doi.org/10.1080
/23273798.2018.1518533
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and
meta-analysis of 120 functional neuroimaging studies. Cerebral
Cortex, 19(12), 2767–2796. https://doi.org/10.1093/cercor
/bhp055, PubMed: 19329570
Bozic, M., Tyler, L. K., Ives, D. T., Randall, B., & Marslen-Wilson,
W. D. (2010). Bihemispheric foundations for human speech
comprehension. Proceedings of the National Academy of Sci-
ences, 107(40), 17439–17444. https://doi.org/10.1073/pnas
.1000531107, PubMed: 20855587
Braze, D., Mencl, W. E., Tabor, W., Pugh, K. R., Constable, R. T.,
Fulbright, R. K., Magnuson, J. S., Van Dyke, J. A., & Shankweiler,
D. P. (2011). Unification of sentence processing via ear and eye:
An fMRI study. Cortex, 47(4), 416–431. https://doi.org/10.1016/j
.cortex.2009.11.005, PubMed: 20117764
Brennan, J., Nir, Y., Hasson, U., Malach, R., Heeger, D. J., & Pylkkanen,
L. (2012). Syntactic structure building in the anterior temporal
lobe during natural story listening. Brain and Language, 120(2),
163–173. https://doi.org/10.1016/j.bandl.2010.04.002,
PubMed: 20472279
Brett, M., Anton, J.-L., Valabregue, R., & Poline, J.-B. (2002, June
2–6). Region of interest analysis using an SPM toolbox [Paper
presentation]. The 8th International Conference on Functional
Mapping of the Human Brain, Sendai, Japan. Available on
CD-ROM in NeuroImage, 16(2), abstract 497.
Caplan, D., Alpert, N., & Waters, G. (1998). Effects of syntactic struc-
ture and propositional number on patterns of regional cerebral
blood flow. Journal of Cognitive Neuroscience, 10(4), 541–552.
https://doi.org/10.1162/089892998562843, PubMed: 9712683
Chen, L., Lambon Ralph, M. A., & Rogers, T. T. (2017). A unified
model of human semantic knowledge and its disorders. Nature
Human Behaviour, 1, Article 0039. https://doi.org/10.1038
/s41562-016-0039, PubMed: 28480333
Cheung, H., & Kemper, S. (1992). Competing complexity metrics and
adults’ production of complex sentences. Applied Psycholinguis-
tics, 13(1), 53–76. https://doi.org/10.1017/S0142716400005427
Constable, R. T., Pugh, K. R., Berroya, E., Mencl, W. E., Westerveld,
M., Ni, W., & Shankweiler, D. (2004). Sentence complexity and
input modality effects in sentence comprehension: An fMRI
study. NeuroImage, 22(1), 11–21. https://doi.org/10.1016/j
.neuroimage.2004.01.001, PubMed: 15109993
Cooke, A., Zurif, E. B., DeVita, C., Alsop, D., Koenig, P., Detre, J.,
Gee, J., Pinãngo, M., Balogh, J., & Grossman, M. (2002). Neural
basis for sentence comprehension: Grammatical and short-term
memory components. Human Brain Mapping, 15(2), 80–94.
https://doi.org/10.1002/hbm.10006, PubMed: 11835600
de Vries, M. H., Petersson, K. M., Geukes, S., Zwitserlood, P., &
Christiansen, M. H. (2012). Processing multiple non-adjacent
dependencies: Evidence from sequence learning. Philosophical
Transcriptions of the Royal Society B, 367(1598), 2065–2076.
https://doi.org/10.1098/rstb.2011.0414, PubMed: 22688641
Eickhoff, S. B., Stephan, K. E., Mohlberg, H., Grefkes, C., Fink,
G. R., Amunts, K., & Zilles, K. (2005). A new SPM toolbox for
combining probabilistic cytoarchitectonic maps and functional
imaging data. NeuroImage, 25(4), 1325–1335. https://doi.org
/10.1016/j.neuroimage.2004.12.034, PubMed: 15850749
Favier, S., & Huettig, F. (2021). Long-term written language expe-
rience affects grammaticality judgements and usage but not pri-
ming of spoken sentences. Quarterly Journal of Experimental
Psychology, 74(8), 1378–1395. https://doi.org/10.1177
/17470218211005228, PubMed: 33719762
Fedorenko, E., Scott, T. L., Brunner, P., Coon, W. G., Pritchett, B.,
Schalk, G., & Kanwisher, N. (2016). Neural correlate of the con-
struction of sentence meaning. Proceedings of the National Acad-
emy of Sciences, 113(41), E6256–E6262. https://doi.org/10.1073
/pnas.1612132113, PubMed: 27671642
Fodor, J. A., Bever, T. G., & Garrett, M. F. (1974). The psychology of
language: An introduction to psycholinguistics and generative
grammar. McGraw-Hill.
Friederici, A. D. (2017). Language in our brain: The origins of a
uniquely human capacity. MIT Press. https://doi.org/10.7551
/mitpress/9780262036924.001.0001
Friston, K. J., Ashburner, J. T., Kiebel, S. J., Nichols, T. E., & Penny,
W. D. (Eds.). (2007). Statistical parametric mapping: The analysis
of functional brain images. Academic Press. https://doi.org/10
.1016/B978-012372560-8/50002-4
Futrell, R., Mahowald, K., & Gibson, E. (2015). Large-scale evidence
of dependency length minimization in 37 languages. Proceedings
of the National Academy of Sciences, 112(33), 10336–10341.
https://doi.org/10.1073/pnas.1502134112, PubMed: 26240370
Gibson, E. (1998). Linguistic complexity: Locality of syntactic
dependencies. Cognition, 68(1), 1–76. https://doi.org/10.1016
/S0010-0277(98)00034-1, PubMed: 9775516
Goucha, T., & Friederici, A. D. (2015). The language skeleton
after dissecting meaning: A functional segregation within Broca’s
area. NeuroImage, 114, 294–302. https://doi.org/10.1016/j
.neuroimage.2015.04.011, PubMed: 25871627
Grodner, D., & Gibson, E. (2005). Consequences of the serial
nature of linguistic input for sentenial complexity. Cognitive
S c i e n c e , 2 9 ( 2 ) , 2 6 1 – 2 9 0 . h t t p s : / / d o i . o r g / 1 0 . 1 2 0 7
/s15516709cog0000_7, PubMed: 21702774
Hagoort, P. (2005). On Broca, brain, and binding: A new frame-
work. Trends in Cognitive Science, 9(9), 416–423. https://doi
.org/10.1016/j.tics.2005.07.004, PubMed: 16054419
Hagoort, P. (2013). MUC (Memory, unification, control) and
beyond. Frontiers in Psychology, 4, Article 416. https://doi.org
/10.3389/fpsyg.2013.00416, PubMed: 23874313
Hagoort, P., & Indefrey, P. (2014). The neurobiology of language
beyond single words. Annual Review of Neuroscience, 37,
347–362. https://doi.org/10.1146/annurev-neuro-071013
-013847, PubMed: 24905595
Jackendoff, R. (2002). Foundations of language: Brain, meaning,
grammar, evolution. Oxford University Press. https://doi.org/10
.1093/acprof:oso/9780198270126.001.0001
Joshi, A. K., & Schabes, Y. (1997). Tree adjoining grammars. In G.
Rozenberg & A. Salomaa (Eds.), Handbook of formal languages
(pp. 69–123). Springer-Verlag. https://doi.org/10.1007/978-3-642
-59126-6_2
Kemper, S. (1986). Imitation of complex syntactic constructions by
elderly adults. Applied Psycholinguistics, 7(3), 277–288. https://
doi.org/10.1017/S0142716400007578
Kemper, S. (1987). Life-span changes in syntactic complexity. Jour-
nal of Gerontology, 42(3), 323–328. https://doi.org/10.1093
/geronj/42.3.323, PubMed: 3571869
Lam, N. H. L., Schoffelen, J. M., Uddén, J., Hultén, A., & Hagoort, P.
(2016). Neural activity during sentence processing as reflected in
theta, alpha, beta, and gamma oscillations. NeuroImage, 142,
43–54. https://doi.org/10.1016/j.neuroimage.2016.03.007,
PubMed: 26970187
Lambon Ralph, M. A., Ehsan, S., Baker, G. A., & Rogers, T. T.
(2012). Semantic memory is impaired in patients with unilateral
anterior temporal lobe resection for temporal lobe epilepsy.
Brain, 135(1), 242–258. https://doi.org/10.1093/brain/awr325,
PubMed: 22287382
Neurobiology of Language
596
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
Lashley, K. S. (1951). The problem of serial order in behavior. In L. A.
Jeffress (Ed.), Cerebral mechanisms in behavior (pp. 112–146).
Wiley.
Lopopolo, A., van den Bosch, A., Petersson, K. M., Willems, R. M.
(2021). Distinguishing syntactic operations in the brain: Depen-
dency and phrase-structure parsing. Neurobiology of Language,
2(1), 152–175. https://doi.org/10.1162/nol_a_00029
Makuuchi, M., Bahlmann, J., Anwander, A., & Friederici, A. D.
(2009). Segregating the core computational faculty of human
language from working memory. Proceedings of the National
Academy of Sciences, 106(20), 8362–8367. https://doi.org/10
.1073/pnas.0810928106, PubMed: 19416819
Matchin, W., & Wood, E. (2020). Syntax-sensitive regions of the
posterior inferior frontal gyrus and the posterior temporal lobe
are differentially recruited by production and perception. Cere-
bral Cortex Communications, 1(1), Article tgaa029. https://doi
.org/10.1093/texcom/tgaa029, PubMed: 34296103
Meltzer, J. A., McArdle, J. J., Schafer, R. J., & Braun, A. R. (2010).
Neural aspects of sentence comprehension: Syntactic complexity,
reversibility, and reanalysis. Cerebral Cortex, 20(8), 1853–1864.
https://doi.org/10.1093/cercor/bhp249, PubMed: 19920058
Michael, E. B., Keller, T. A., Carpenter, P. A., & Just, M. A. (2001).
fMRI investigation of sentence comprehension by eye and by ear:
Modality fingerprints on cognitive processes. Human Brain Map-
ping, 13(4), 239–252. https://doi.org/10.1002/ hbm.1036,
PubMed: 11410952
Mumford, J. A., Poline, J.-B., & Poldrack, R. A. (2015). Orthogonal-
ization of regressors in fMRI models. PLOS ONE, 10(4), Article
e0126255. https://doi.org/10.1371/journal.pone.0126255,
PubMed: 25919488
Nelson, M. J., El Karoui, I., Giber, K., Yang, X., Cohen, L., Koopman,
H., Cash, S. S., Naccache, L., Hale, J. T., Pallier, C., & Dehaene, S.
(2017). Neurophysiological dynamics of phrase-structure building
during sentence processing. Proceedings of the National Academy
of Sciences, 114(18), E3669–E3678. https://doi.org/10.1073/pnas
.1701590114, PubMed: 28416691
Nichols, T., Brett, M., Andersson, J., Wager, T., & Poline, J.-B.
(2005). Valid conjunction inference with the minimum statistic.
NeuroImage, 25(3), 653–660. https://doi.org/10.1016/j
.neuroimage.2004.12.005, PubMed: 15808966
Nieuwland, M. S., & Van Berkum, J. J. A. (2006). Individual differ-
ences and contextual bias in pronoun resolution: Evidence from
ERPs. Brain Research, 1118(1), 155–167. https://doi.org/10.1016
/j.brainres.2006.08.022, PubMed: 16956594
Norman, S., Kemper, S., & Kynette, D. (1992). Adults’ reading com-
prehension: Effects of syntactic complexity and working memory.
Journal of Gerontology, 47(4), P258–P265. https://doi.org/10
.1093/geronj/47.4.P258, PubMed: 1624703
Öquist, G., & Goldstein, M. (2003). Towards an improved readabil-
ity on mobile devices: Evaluating adaptive rapid serial visual pre-
sentation. Interacting with Computers, 15(4), 539–558. https://doi
.org/10.1016/S0953-5438(03)00039-0
Pallier, C., Devauchelle, A. D., & Dehaene, S. (2011). Cortical rep-
resentation of the constituent structure of sentences Proceedings
of the National Academy of Sciences, 108(6), 2522–2527. https://
doi.org/10.1073/pnas.1018711108, PubMed: 21224415
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do you
know what you know? The representation of semantic knowl-
edge in the human brain. Nature Reviews Neuroscience, 8,
976–987. https://doi.org/10.1038/nrn2277, PubMed: 18026167
Peelle, J. E., McMillan, C., Moore, P., Grossman, M., & Wingfield,
A. (2004). Dissociable patterns of brain activity during compre-
hension of rapid and syntactically complex speech: Evidence
from fMRI. Brain and Language, 91(13), 315–325. https://doi
.org/10.1016/j.bandl.2004.05.007, PubMed: 15533557
Regev, M., Honey, C. J., Simony, E., & Hasson, U. (2013). Selective
and invariant neural responses to spoken and written narratives.
Journal of Neuroscience, 33(40), 15978–15988. https://doi.org
/10.1523/JNEUROSCI.1580-13.2013, PubMed: 24089502
Rogalsky, C., Matchin, W., & Hickok, G. (2008). Broca’s area, sen-
tence comprehension, and working memory: An fMRI study.
Frontiers in Human Neuroscience, 2, Article 14. https://doi.org
/10.3389/neuro.09.014.2008, PubMed: 18958214
Santi, A., & Grodzinsky, Y. (2010). fMRI adaptation dissociates syn-
tactic complexity dimensions. NeuroImage, 51(4), 1285–1293.
https://doi.org/10.1016/j.neuroimage.2010.03.034, PubMed:
20338244
Schoffelen, J.-M., Oostenveld, R., Lam, N. H. L., Uddén, J., Hultén, A.,
& Hagoort, P. (2019). A 204-subject multimodal neuroimaging
dataset to study language processing. Scientific Data, 6, Article 17.
https://doi.org/10.1038/s41597-019-0020-y, PubMed: 30944338
Shankweiler, D., Mencl, W. E., Braze, D., Tabor, W., Pugh, K. R., &
Fulbright, R. K. (2008). Reading differences and brain: Cortical
integration of speech and print in sentence processing varies with
reader skill. Developmental Neuropsychology, 33(6), 745–775.
https://doi.org/10.1080/87565640802418688, PubMed:
19005913
Snijders, T. M., Vosse, T., Kempen, G., Van Berkum, J. J., Petersson,
K. M., & Hagoort, P. (2009). Retrieval and unification of syntactic
structure in sentence comprehension: An fMRI study using
word-category ambiguity. Cerebral Cortex, 19(7), 1493–1503.
https://doi.org/10.1093/cercor/bhn187, PubMed: 19001084
Stromswold, K., Caplan, D., Alpert, N., & Rauch, S. (1996). Local-
ization of syntactic comprehension by positron emission tomog-
raphy. Brain and Language, 52(3), 452–473. https://doi.org/10
.1006/brln.1996.0024, PubMed: 8653390
Trettenbrein, P. C., Papitto, G., Friederici, A. D., & Zaccarella, E.
(2021). Functional neuroanatomy of language without speech:
An ALE meta-analysis of sign language. Human Brain Mapping,
42(3), 699–712. https://doi.org/10.1002/hbm.25254, PubMed:
33118302
Tyler, L. K., Cheung, T. P., Devereux, B. J., & Clarke, A. (2013). Syn-
tactic computations in the language network: Characterizing
dynamic network properties using representational similarity
analysis. Frontiers in Psychology, 4, Article 271. https://doi.org
/10.3389/fpsyg.2013.00271, PubMed: 23730293
Tyler, L. K., Shafto, M. A., Randall, B., Wright, P., Marslen-Wilson,
W. D., & Stamatakis, E. A. (2010). Preserving syntactic processing
across the adult life span: The modulation of the frontotemporal
language system in the context of age-related atrophy. Cerebral
Cortex, 20(2), 352–364. https://doi.org/10.1093/cercor/bhp105,
PubMed: 19505991
Uddén, J., & Bahlmann J. (2012). A rostro-caudal gradient of
structured sequence processing in the left inferior frontal gyrus.
Philosophical Transactions of the Royal Society B, 367(1598),
2023–2032. https://doi.org/10.1098/rstb.2012.0009, PubMed:
22688637
Uddén, J., Ingvar, M., Hagoort, P., & Petersson, K. M. (2012).
Implicit acquisition of grammars with crossed and nested
non-adjacent dependencies: Investigating the push-down stack
model. Cognitive Science, 36(6), 1078–1101. https://doi.org/10
.1111/j.1551-6709.2012.01235.x, PubMed: 22452530
Vogelzang, M., Thiel, C. M., Rosemann, S., Rieger, J. W., & Ruigendijk,
E. (2020). Neural mechanisms underlying the processing of com-
plex sentences: An fMRI Study. Neurobiology of Language, 1(2),
226–248. https://doi.org/10.1162/nol_a_00011
Neurobiology of Language
597
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Syntactic complexity in supramodal sentence processing
Vosse, T., & Kempen, G. (2000). Syntactic structure assembly in
human parsing: A computational model based on competitive inhi-
bition and a lexicalist grammar. Cognition, 75(2), 105–143. https://
doi.org/10.1016/S0010-0277(00)00063-9, PubMed: 10771275
Vosse, T., & Kempen, G. (2009). The Unification Space imple-
mented as a localist neural net: Predictions and error-tolerance
in a constraint-based parser. Cognitive Neurodynamics, 3(4),
331–346. https://doi.org/10.1007/s11571-009-9094-0, PubMed:
19784798
Walenski, M., Europa, E., Caplan, D., & Thompson, C. K. (2019).
Neural networks for sentence comprehension and production:
An ALE-based meta-analysis of neuroimaging studies. Human
Brain Mapping, 40(8), 2275–2304. https://doi.org/10.1002/hbm
.24523, PubMed: 30689268
Willems, R. M., Frank, S. L., Nijhof, A. D., Hagoort, P., & van den
Bosch, A. (2016). Prediction during natural language compre-
hension. Cerebral Cortex, 26(6), 2506–2516. https://doi.org/10
.1093/cercor/bhv075, PubMed: 25903464
Wilson, S. M., Bautista, A., & McCarron, A. (2018). Convergence of
spoken and written language processing in the superior temporal
sulcus. NeuroImage, 171, 62–74. https://doi.org/10.1016/j
.neuroimage.2017.12.068, PubMed: 29277646
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
4
5
7
5
2
0
4
6
8
1
7
n
o
_
a
_
0
0
0
7
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
598