RESEARCH ARTICLE
Activation in Right Dorsolateral Prefrontal Cortex
Underlies Stuttering Anticipation
Eric S. Jackson1
, Swethasri Dravida2
Vincent Gracco3,4
, Xian Zhang2
, and Joy Hirsch2,5,6
, J. Adam Noah2
,
a n o p e n a c c e s s
j o u r n a l
Citation: Jackson, E. S., Dravida, S.,
Zhang, X., Noah, J. A., Gracco, V., &
Hirsch, J. (2022). Activation in right
dorsolateral prefrontal cortex underlies
stuttering anticipation. Neurobiology
of Language, 3(3), 469–494. https://doi
.org/10.1162/nol_a_00073
DOI:
https://doi.org/10.1162/nol_a_00073
Supporting Information:
https://doi.org/10.1162/nol_a_00073
Received: 2 August 2021
Accepted: 16 May 2022
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Eric S. Jackson
ej34@nyu.edu
Handling Editor:
Kate E. Watkins
Copyright: © 2022
Massachusetts Institute of Technology
Published under a Creative Commons
Attribution 4.0 International
(CC BY 4.0) license
The MIT Press
1Department of Communicative Sciences and Disorders, New York University, New York, USA
2Department of Psychiatry, Yale School of Medicine, New Haven, CT, USA
3Haskins Laboratories, New Haven, CT, USA
4McGill University, Montreal, Canada
5Department of Neuroscience, Department of Comparative Medicine, Yale School of Medicine, New Haven, CT, USA
6Department of Medical Physics and Biomedical Engineering, University College London, London, UK
Keywords: stuttering, anticipation, disfluency, fNIRS, frontoparietal network, error-likelihood
monitoring, action-stopping
ABSTRACT
People who stutter learn to anticipate many of their overt stuttering events. Despite the critical
role of anticipation, particularly how responses to anticipation shape stuttering behaviors,
the neural bases associated with anticipation are unknown. We used a novel approach to
identify anticipated and unanticipated words, which were produced by 22 adult stutterers in
a delayed-response task while hemodynamic activity was measured using functional near
infrared spectroscopy (fNIRS). Twenty-two control participants were included such that each
individualized set of anticipated and unanticipated words was produced by one stutterer and
one control participant. We conducted an analysis on the right dorsolateral prefrontal cortex
(R-DLPFC) based on converging lines of evidence from the stuttering and cognitive control
literatures. We also assessed connectivity between the R-DLPFC and right supramarginal gyrus
(R-SMG), two key nodes of the frontoparietal network (FPN), to assess the role of cognitive
control, and particularly error-likelihood monitoring, in stuttering anticipation. All analyses
focused on the five-second anticipation phase preceding the go signal to produce speech.
The results indicate that anticipated words are associated with elevated activation in the
R-DLPFC, and that compared to non-stutterers, stutterers exhibit greater activity in the
R-DLPFC, irrespective of anticipation. Further, anticipated words are associated with reduced
connectivity between the R-DLPFC and R-SMG. These findings highlight the potential roles of
the R-DLPFC and the greater FPN as a neural substrate of stuttering anticipation. The results
also support previous accounts of error-likelihood monitoring and action-stopping in stuttering
anticipation. Overall, this work offers numerous directions for future research with clinical
implications for targeted neuromodulation.
INTRODUCTION
Stuttering is a complex neurodevelopmental communication disorder that often negatively
impacts social, emotional, and professional opportunities for more than 50 million adults
worldwide. The disorder manifests itself to listeners as intermittent interruptions in speech pro-
duction including part-syllable repetitions and audible and inaudible prolongations of sounds.
However, these behaviors do not always accompany stuttering events because most, if not all,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
stutterers develop the remarkable ability to anticipate stuttering. As a result, they can alter their
speech plan prior to execution by, for example, avoiding, stalling, or using a speaking strategy
(e.g., pull-out, easy onset; Jackson et al., 2015, 2019). This potential discrepancy between the
internal experience of the speaker (i.e., sensing upcoming speech breakdown) and how stut-
tering manifests itself to the listener (i.e., as something out of the ordinary) has important clin-
ical implications because how individuals respond to anticipation shapes their communicative
experiences. Behavioral and qualitative investigations of anticipation, especially in recent
years, have improved our understanding of the phenomenon, but it is critical to augment this
evidence with a neural account of stuttering anticipation, especially given the covert nature of
anticipation. The purpose of this study was to initiate a brain-based understanding of stuttering
anticipation by linking neural activation to self-reported anticipation and subsequent stuttering
behaviors.
The Anticipation of Stuttering
Stuttering anticipation refers to the sense or prescience that upcoming speech will be stuttered,
should the speaker execute their speech plan as originally intended without alterations
(Jackson et al., 2015; Wingate, 1975). Anticipation occurs on a temporal continuum from a
longer-term or looming sense of impending stuttering to a shorter-term immediate sense of
upcoming stuttering. For example, a speaker may anticipate a word months in advance of
saying it (a student knowing at the beginning of the semester that they have to say a certain
word in a presentation at the end of the semester); minutes or seconds before (when they are
about to introduce themselves); or immediately before executing speech. Anticipation is
driven by error-likelihood monitoring whereby the speaker learns associations between
“errors” (i.e., stuttered utterances) and listener reactions or other environmental consequences,
thereby learning to predict the occurrences of these errors (Arenas, 2012, 2017; Garcia-Barrera
& Davidow, 2015). While adult stutterers, as a group, predict stuttering with high accuracy in
experimental settings (greater than 90% accuracy; Knott et al., 1937; Milisen, 1938; Van Riper,
1936), there is a range in which speakers report anticipating stuttering, from, “sometimes” to
“always” (Jackson et al., 2015). Anticipation is a relatively stable feature such that anticipated
or feared words are stuttered in experiments even three months after they are identified by
participants (Mersov et al., 2018). Arguably most important in the speaker’s experience is
how they learn or choose to respond to anticipation, whether by avoiding, approaching, or
implementing physical speaking strategies that prevent stuttering from coming to the surface
(Jackson et al., 2015, 2019). In this way, responding to anticipation is mediated by cognitive
control.
Cognitive Control
Cognitive control refers to the ability to orchestrate brain functions to complete a given task or
reach a certain goal (Miller, 2000). Cognitive control encompasses planning, initiating, and
inhibiting actions or tasks, and being flexible and vigilant to tasks in response to environmental
demands (Niendam et al., 2012). All of these processes are involved in responding to stuttering
anticipation. For example, when a stutterer knows that they are going to stutter, they must
initiate (or choose not to initiate) a response which may include avoidance or using a speaking
strategy (Jackson et al., 2015, 2019); they may inhibit responses due to fear of negative reac-
tions from the listener; they must be flexible with the challenge at hand (i.e., not being able to
say what they want to say when they want to say it); and they must remain vigilant to their goal
(i.e., producing speech). While numerous studies have examined cognitive control in children
Neurobiology of Language
470
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
and adults who stutter (for review, see Anderson & Ofoe, 2019), no studies have assessed the
relationship between cognitive control and anticipation directly.
A Potential Neural Substrate of Stuttering Anticipation
While anticipation is pervasive in the stuttering experience and contributes significantly to the
negative impact on quality of life for stutterers (Jackson et al., 2015; Tichenor & Yaruss, 2019),
the neural underpinnings of anticipation and related cognitive control processes are unknown.
Neurofunctional investigations of stuttering have instead focused on the speech motor net-
work, revealing atypical activation in left perisylvian and motor areas along the arcuate and
superior longitudinal fasciculus, and their homologous regions in the right hemisphere, as well
as atypical activity in basal ganglia and cerebellum (Braun et al., 1997; S.-E. Chang et al.,
2011; De Nil et al., 2000; Fox et al., 2000; Kell et al., 2009; Neef et al., 2016; Neumann
et al., 2004; Preibisch et al., 2003; Toyomura et al., 2018). Some of these studies reported
significant findings outside of the speech motor network in areas related to cognitive control,
even though these areas were not the focus of those investigations. For example, it is widely
known that the right dorsolateral prefrontal cortex (R-DLPFC) plays a critical role in cognitive
control processes (Koechlin et al., 2003; MacDonald et al., 2000; Miller, 2000; Ridderinkhof
et al., 2004). Stutterers exhibit elevated activation in the R-DLPFC (Kell et al., 2009; Lu et al.,
2009; Neef et al., 2018), and treatment temporarily reduces activity in R-DLPFC while reduc-
ing stuttering symptoms (De Nil et al., 2004; Neumann et al., 2005). Kell et al. (2009) also
found that stutterers who reported recovering from stuttering without treatment did not show
elevated activation in R-DLPFC, suggesting that these patterns reflect compensatory efforts not
learned in therapy (e.g., avoiding, stalling, or using other self-learned speaking strategies).
However, because anticipation and the R-DLPFC were not the focus of these studies, the
potential relevance of the R-DLPFC to stuttering anticipation, and to stuttering more broadly,
is unknown.
We focus primarily on the R-DLPFC in this study, but other regions and networks are likely
recruited during anticipation and responding to anticipation. Modern imaging and computa-
tional approaches, including task-induced and task-free connectivity analysis, have identified
multiple non-overlapping and distributed brain networks that underlie cognitive control,
including the frontoparietal network (FPN), salience network, cingulo-opercular network,
and dorsal and ventral attention networks (D’Esposito & Postle, 2002; Menon & D’Esposito,
2021; Niendam et al., 2012). The FPN, which includes the R-DLPFC and right supramarginal
gyrus (R-SMG) (Menon & D’Esposito, 2021), is particularly relevant because it co-activates
with multiple other networks (e.g., salience network, cingulo-opercular network) to carry
out the diverse processes associated with cognitive control (Marek & Dosenbach, 2018).
The FPN initiates and flexibly modulates interactions between the salience and cingulo-
opercular networks (Marek & Dosenbach, 2018), which both include the anterior cingulate
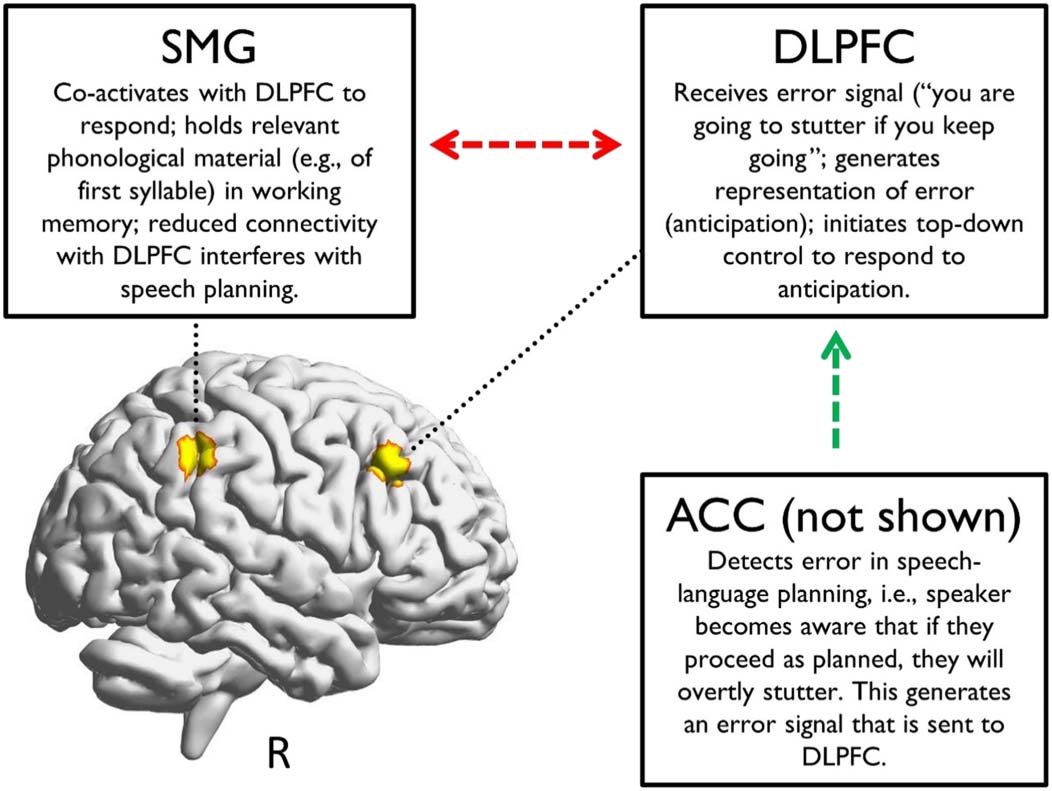
cortex (ACC). ACC underlies error-likelihood monitoring (Brown & Braver, 2005): DLPFC
co-activates with ACC, whereby ACC underlies the detection of errors in response to unin-
tended outcomes and generates error signals, and DLPFC holds task-relevant information in
working memory and initiates subsequent actions (Alexander & Brown, 2015; Holroyd &
Yeung, 2012). It is reasonable to propose that the ACC underlies stuttering anticipation—the
recognition of the breakdown or “glitch” in speech-language planning—and reasonable to
predict that the R-DLPFC underlies initiating a response to this breakdown. Given the strong
bidirectional connections within the FPN (Goldman-Rakic, 1988; Menon & D’Esposito, 2021;
Mesulam, 1998), particularly between R-DLPFC and R-SMG, it is also reasonable to hypoth-
esize that anticipation destabilizes these connections, resulting in altered connectivity.
Neurobiology of Language
471
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
While it was not possible in the current design to disentangle anticipation and responding
to anticipation, the reader should be aware of these somewhat distinct but overlapping pro-
cesses. We conceive of anticipation as an event—the point in time at which the speaker
becomes aware that should they proceed as planned, they will overtly stutter—and responding
to anticipation as the constellation of processes, involving the cognitive control system, that
underlies how a speaker chooses to proceed in light of the knowledge that they are likely
about to stutter. In the current design and given the hemodynamic lag associated with fNIRS,
it was not possible to disentangle anticipation and responding to anticipation, and thus,
the current study examined the processes involved in anticipation generally. Techniques
with better temporal resolution than functional near-infrared spectroscopy (fNIRS; i.e.,
magnetoencephalography/electroencephalography (MEG/EEG)) are more suited to disentan-
gle these two processes.
The current study examined the relationship between R-DLPFC and stuttering anticipation,
which may clarify the significance of previous and seemingly incidental findings of elevated
activation in R-DLPFC in stutterers (e.g., Kell et al., 2009; Neef et al., 2018). In a first visit, we
used a clinical interview to determine individual-specific anticipated and unanticipated words
(Jackson et al., 2020). In a second visit, which occurred between three and 10 days after the
first visit, we used fNIRS to measure cortical activation immediately prior to participants pro-
ducing the anticipated and unanticipated words in a delayed-response task. We focused on
superficial cortical structures in part due to imaging depth restrictions associated with fNIRS.
While we were not able to measure activation from deeper structures such as ACC, fNIRS
offered several advantages compared to other techniques (e.g., functional magnetic resonance
imaging (fMRI), EEG, MEG), including (1) robustness to speech movement artifact, and (2)
allowing participants to produce speech while they sat upright and across from a communi-
cative partner, which increased the likelihood of anticipation. fNIRS has also been validated as
a tool to measure DLPFC activation associated with anticipation (Vassena et al., 2019). A
matched control group was included to test whether stutterers recruit R-DLPFC differently than
non-stutterers. Each anticipated and unanticipated word list was produced by a stutterer and a
control speaker. We conducted a region of interest (ROI) analysis of R-DLPFC and hypothe-
sized that (1) anticipation would be associated with greater activation in R-DLPFC, reflecting
cognitive control processes associated with responding to anticipation, and (2) stutterers
would exhibit greater activation than control speakers in R-DLPFC during this same time
period. We also assessed functional connectivity to test whether anticipation was associated
with reduced intrinsic connectivity within the FPN, specifically between R-DLPFC and R-SMG.
MATERIALS AND METHODS
This study was approved by the Institutional Review Boards at New York University and Yale
University. Consent was obtained for all participants in accordance with the Declaration
of Helsinki.
Participants
Twenty-seven adult stutterers were recruited through the first author’s clinical network, mass
emails distributed by Friends: The National Association of Young People Who Stutter and the
National Stuttering Association, and by word of mouth. After the fNIRS screening (see below),
22 stutterers (9 female; mean age = 31.9, SD = 9.1; three left-handed) and 22 control speakers
(10 female; mean = 27.4, SD = 8.0; three left-handed) participated in the study. Control par-
ticipants were recruited after the stuttering participants so that they could be matched for age,
Neurobiology of Language
472
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
gender, and stimuli (see below). Male-to-female ratio was lower than what is typically
observed in the stuttering literature (59% vs. ∼75–80%). All participants were between the
ages of 18 and 50, reported that American English was their primary language (multilingual
was acceptable as long as English was learned during early childhood [younger than 6 years of
age]), and reported negative histories of neurological, speech-language, psychological, learn-
ing, and hearing impairment. Participant characteristics, including age, gender, treatment
history, and extent score from the Stuttering Anticipation Scale (SAS; Jackson et al., 2018)
are included in Table 1.
ID
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
Table 1.
Participant data
Age
29
Gender
F
Treatment history
on and off in elementary school
F
M
M
F
F
M
M
M
M
F
F
F
F
M
M
M
M
F
M
M
F
7 years
2 weeks (intensive program)
6 years
3 years
roughly 2 years
8 years
2.5 years
9 months
2 years
off and on for years
12–15 years
6+ years
none
8 years
10 years, on and off
approx. 10 years
about 7 years
about 6 months
about 7 years (on and off )
none
3 months
35
23
34
26
48
21
23
29
34
37
39
23
47
42
18
30
29
39
25
22
50
32
9
M
SD
%TS
43%
36%
0%
0%
87%
80%
27%
23%
45%
98%
23%
76%
93%
30%
15%
70%
51%
25%
55%
21%
55%
15%
40%
30%
SAS
50
100
76
79
95
87
90
77
80
80
75
86
89
98
90
75
76
67
76
70
77
99
81
12
Note. Treatment history descriptions written as reported by participants. TS = Trials Stuttered; SAS = Stuttering
Anticipation Scale (extent score out of 100, 0 = never, 100 = always). ID = participant; M = mean; SD = standard
deviation.
Neurobiology of Language
473
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
First visit (stutterers only)
The first visit comprised (1) the stuttering assessment, (2) the clinical interview to determine
participant-specific stimuli, and (3) the screening for fNIRS. Only the stuttering group partici-
pated in two visits; the fNIRS screening for the control group took place on the same day as
fNIRS testing.
Diagnostic assessment
The stuttering group was identical to that in Jackson et al. (2020). That study validated the
clinical interview used here, and the only overlapping data between that study and the current
study are the speech classification data (stuttered/ambiguous/fluent, interrater reliability). Stut-
tering was diagnosed by the first author, an American Speech-Language-Hearing Association
certified speech-language pathologist (SLP) with more than 10 years of expertise in stuttering
intervention.
All stuttering participants (1) self-reported as a person who stutters; and (2) exhibited three
or more stuttering-like disfluencies (Yairi & Ambrose, 1992) with temporally aligned physical
concomitants (e.g., eye blinking, head movements) during a 5–10 min conversation. Partici-
pants also completed the SAS (Jackson et al., 2018), which provided self-report ratings of the
extent of anticipation based on a 0–100 (never–always) visual analog scale (“How much do
you anticipate stuttering?”).
Clinical interview
The clinical interview was described previously in Jackson et al. (2020), and will be described
briefly here. Interviews were conducted at New York University and Yale University, and for
two participants remotely. The purpose of the interview was to identify 10 words that the
participants anticipated they would stutter (hence: anticipated words) and 10 words that
they did not anticipate they would stutter (hence: unanticipated words), resulting in individual
lists for each participant. This approach extends previous methods for identifying
anticipated/unanticipated words (Bowers et al., 2012; Mersov et al., 2018; Wymbs et al.,
2013), by including clinical inference (e.g., asking participants whether stuttered
words/sounds not immediately identified by participants as anticipated or “feared”
words/sounds should be included as anticipated words), as well as using counseling tech-
niques to create an environment in which participants were comfortable to identify/reveal
feared words. The words were used to create the stimuli for fNIRS testing, which consisted
of short questions or sentence completions that would require the participant to produce
the words. Anticipated and unanticipated words were matched for length (number of sylla-
bles). Each stutterer was matched with a control participant who produced the same set of
words.
fNIRS screening
The goal of the fNIRS screening was to determine whether a reliable hemodynamic signal
could be acquired from each participant. This is because factors such as bone density and
skull thickness weaken fNIRS signals (Krall & Dawson-Hughes, 1993; Okada & Delpy,
2003). Screening fNIRS participants limits acquisition of invalid data (Zhang et al., 2017).
The screening consisted of a finger tapping task during which participants were required to
tap their fingers in an alternating pattern for 15 s then rest for 15 s, for a total of 3 min (right
hand). Typically, this task elicits a robust response in left motor/premotor areas. Here, a
response was determined to be reliable if there was sufficient separation between
Neurobiology of Language
474
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
oxyhemoglobin (HbO) and deoxyhemoglobin (HbR) signals in any channel in left
motor/premotor areas based on visual inspection of the event-triggered average. Importantly,
potential participants were excluded before the study began. That is, no participants were
excluded from the study after their signals were determined to be reliable. Twenty-seven stut-
terers were screened, and 22 participated in the study; twenty-eight controls were screened,
and 22 participated in the study.
Second visit
The second visit (for the stuttering group) occurred between 3 and 10 days after the first visit,
and included fNIRS testing in the Brain Function Laboratory at Yale. Participant-specific stimuli
were created between the first and second visits based on the anticipated and unanticipated
word lists established during the clinical interview (first visit). Each word list was used to create
the stimuli for one stuttering and one control speaker. Stimuli included simple questions or
sentence completions that required one-word responses (e.g., “You can fly in an ______” ➔
airplane. “What month comes after June?” ➔ July). All stimulus questions were approximately
between 1 and 3 s. Participants were exposed to the stimuli before the experiment to minimize
the potential impact of language retrieval and formulation processes (i.e., they became familiar
with the questions and answers prior to the experiment).
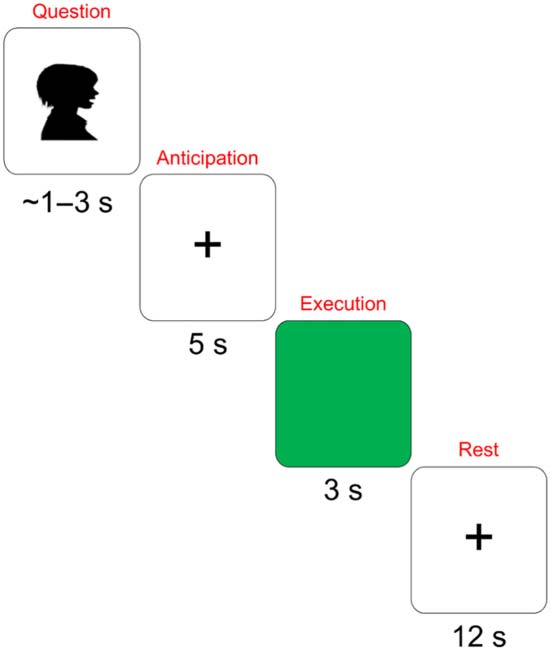
Figure 1 depicts the task timeline. The question was presented auditorily while participants
viewed a cross on the monitor. Verbal responses were delayed by 5 s. Participants responded
when the screen turned green. Participants were asked to look straight ahead and to try to
remain still. The paradigm included interactive and alone conditions, but the condition con-
trast was not the focus of the current study. The two conditions were pooled in all analyses.
During the interactive condition, participants responded to questions asked by the examiner,
who was seated directly across from the participant and in full view. Participants were
instructed to look at the examiner while responding (e.g., “make sure you’re looking at me
when you respond”). The experimenter was given a cue on a separate monitor (unseen by
the participant) just before the go signal was presented, allowing him to look at the participants
when they responded. During the alone condition, participants responded to prerecorded
stimuli, the same questions as asked by the examiner as described above, while alone in
the testing room. Questions during the interactive condition were matched, to the best of
Figure 1. Task timeline. The question/sentence completion was presented auditorily. The antici-
pation period was 5 s; participants looked at a fixation cross during this time. The green screen
signaled participants to produce the word. Participants again looked at the cross during the rest
period.
Neurobiology of Language
475
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
the examiner’s ability, to the prerecorded questions in terms of duration and prosody. The
paradigm included a total of 80 trials: 4 interactive runs, 4 alone runs; each run included
10 words. Each word was produced 4 times.
Data Acquisition
Behavioral
Speech data were acquired via acoustic and video recordings. Acoustic signals were recorded
using a head-mounted microphone with a pop-screen filter set at the same fixed distance for
each participant. Video was captured using a Logitech c920 HD 1080p video camera
mounted on the participant’s monitor.
Neural
Data collection methods have been described previously (e.g., Hirsch et al., 2018, 2021), and
are also described here. Hemodynamic signals were acquired using an 80-fiber continuous-
wave fNIRS system (Shimadzu LABNIRS, Kyoto, Japan) with a temporal resolution of 27 ms.
Forty emitters and forty detectors were arranged in a 134-channel layout covering bilateral
frontal, temporal, parietal, and occipital lobes. Depending on the size of the participant’s
head, caps with optode distances of either 2.75 cm or 3 cm were used. Three wavelengths
of light (780, 805, and 830 nm) were delivered by each LABNIRS emitter. Absorption was
converted to concentration changes for deoxyhemoglobin (HbR) and oxyhemoglobin (HbO)
using the Beer-Lambert Law (Matcher et al., 1995). After the experiment, anatomical locations
of optodes were determined based on standard head landmarks (inion, nasion, top center [Cz],
and left and right tragi) using a Patriot 3D Digitizer (Polhemus, Colchester, VT) and linear
transform techniques (Eggebrecht et al., 2012; Ferradal et al., 2014; Okamoto & Dan,
2005). Montreal Neurological Institute (MNI) coordinates for the channels were obtained
using NIRS-SPM (Ye et al., 2009) in MATLAB (Mathworks, Natick, MA), and corresponding
anatomical locations were determined for each channel. See Table S1 for group median coor-
dinates, atlas-based probabilities, and anatomical regions for each channel; see Figure S1 for a
visual representation. (Supporting Information can be found at https://doi.org/10.1162/nol_a
_00073.) Channels were clustered into anatomical regions based on shared anatomy. The
average number of channels per region was 2.69 ± 1.40.
Data Processing
Behavioral
Errors, which comprised non-productions or incorrect productions due to participants forget-
ting answers or producing erroneous speech, were not included in any analyses (Table 2).
Reaction time was calculated as the time between the go signal (i.e., the green screen) and
speech onset as defined by the first articulatory movement or accessory behavior (Table 3).
Articulatory onset was marked as the first articulatory movement based on visual inspection
using Davinci Resolve (Black Magic Design, Australia), which allowed for frame-by-frame
scanning (29.97 frames per second) of the recordings of participants’ faces. Interrater reliability
between the first author and a SLP blind to the study yielded a Cohen’s weighted kappa of 0.89
(p < 0.05), indicating strong agreement (McHugh, 2012). We used articulatory onset because
(1) inaudible sound prolongations (blocks) typically included observable movement such as
posturing; and (2) it appeared that for some participants, video and audio were not synchro-
nized due to technical error. Although determining neural correlates of stuttered speech was
not the primary goal of this study, we include a comparison of stuttered and fluent speech for
Neurobiology of Language
476
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
completeness. A three-point rating system was used to classify stuttering response type: 0 indi-
cated unambiguous fluency; 1 indicated ambiguity (unclear whether stuttered or fluent); and 2
indicated unambiguous stuttering (Jackson et al., 2020).
Neural
Data processing was similar to that previously reported (Hirsch et al., 2017, 2018; Zhang et al.,
2017). Baseline drift was removed using a NIRS-SPM detrending procedure. Global compo-
nents were removed using a principal component analysis spatial filter (Zhang et al., 2016),
which is comparable to using short-source channels (Noah et al., 2021). Channels were
rejected if the root mean square (RMS ) of the raw signal was 10 times greater than the group
mean RMS, which resulted in a mean of 1.48 of 134 (1.1%) channels per participant rejected.
Little to no motion artifact was observed in the data, based on visual inspection, likely because
participants were instructed to remain still during the task. This is typical for compliant adult
participants (Noah et al., 2021). Further, there was no speech movement observed during the
anticipation phase. HbR and HbO signals were acquired. The fNIRS data sets for each subject
were reshaped into 3D volume images for the general linear model (GLM) analysis using
SPM8 ( Wellcome Trust, London, UK). Beta value coordinates were converted to standard
MNI space using NIRS-SPM (Ye et al., 2009). Contrast images were rendered on a standardized
MNI brain template using a p-value threshold of 0.05 and cluster size threshold of 50 voxels,
for visual representation. Anatomical locations of peak voxel activity were identified using the
Brodmann area Talairach atlas (Lancaster et al., 2000).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Statistical Analysis
Behavioral
To examine reaction time and stuttering response type (stuttered/ambiguous/fluent), linear
mixed effects models were fit using the lme4 (Bates et al., 2014) and lmerTest packages in
R (Kuznetsova et al., 2017; R Core Team, 2014). The MuMIn package (Barton, 2020) was
used to calculate estimated R2 for model fit. For reaction time, the model included word type
(anticipated/unanticipated), word length (number of syllables), trial, and stuttering response
type as fixed factors, and participant as a random factor to account for expected variation
due to individual differences. Wilcoxon rank sum tests were also used to assess reaction
time differences for stuttered, ambiguous, and fluent speech. For stuttering response type,
the model included word type, word length, and trial as fixed factors, and participant as a
random factor.
Neural
All fNIRS analyses followed standard voxel-wise GLM techniques (Friston et al., 1994, 1995)
adapted for fNIRS (e.g., Descorbeth et al., 2020; Hirsch et al., 2021). Analyses targeted the
anticipation phase, i.e., the 5 s time window between the end of the question and the go
cue (see Figure 1). All included voxels were within 2 cm from the cortical surface. The primary
ROI analyses focused on the R-DLPFC; secondary ROI analyses examined activation in the
right inferior frontal gyrus (R-IFG) and right pre-supplementary motor area (R-preSMA). Con-
nectivity between the R-DLPFC and R-SMG was also assessed.
ROI analyses. The mask for R-DLPFC was created by generating a 10 mm sphere using the
MarsBar toolbox (Brett et al., 2002) and xyz coordinates [50 26 38] from Kell et al. (2009).
Five ROI analyses of R-DLPFC were conducted.
Neurobiology of Language
477
Stuttering and anticipation
(1) Anticipated vs. Unanticipated: Activation associated with anticipated vs. unanticipated
words was compared by using SPM8 to convolve a 5 s block regressor during the anticipation
phase, with a standard hemodynamic response function (HRF) that was fitted to the data. The
first-level analysis yielded two beta values for each participant in each run (i.e., 10 trials per
run), for anticipated and unanticipated words (five anticipated, five unanticipated). The results
of the second-level contrast of anticipated vs. unanticipated words were projected onto a
standardized MNI template image using SPM (p < 0.05 and cluster size of >50 voxels, uncor-
rected). Note that the “whole-brain” image was used only to test the ROIs; however, whole-
brain results for this analysis, and all analyses below, are included as supplementary material.
Significance for the ROI analysis was tested using a one-tailed t test in SPM (p < 0.05) by
determining overlap between the second-level image (anticipated vs. unanticipated) and the
mask. Two additional analyses were conducted to test whether the R-DLPFC ROI was signif-
icantly activated during the anticipation phase for anticipated and unanticipated words
(separately). The second-level results, which compared activation for both anticipated and
unanticipated words to rest, were projected onto the same MNI template, and the ROI anal-
yses were carried out as described above.
(2) Stuttered vs. Fluent: To compare activation related to response type within stutterers
(stuttered, ambiguous, fluent), the regressor was modulated in height (0 = fluent, 1 = ambigu-
ous, 2 = stuttered ) in the first-level analysis. The assumption was that stuttered speech yielded
more (or less) activity than ambiguous responses, which yielded more (or less) activity than
fluent responses, effectively providing a “contrast” of stuttered and fluent speech. Beta values
were compared to zero using a one-tailed t test in SPM. The ROI analysis was conducted as
described above.
(3) Stutterers vs. Controls: The ROI group level comparison was similar to the anticipated vs.
unanticipated contrast described above, except for the second-level contrast. The 5 s block
regressor was convolved with the HRF, irrespective of whether the word was anticipated.
Results of the second-level contrast for stutterers vs. controls were projected onto a standard-
ized MNI template image, and the ROI analysis was carried out as described above.
(4) Task vs. Rest (controls only): We also conducted an analysis of the controls only, during
the same 5 s window. The purpose of this analysis was to determine whether the R-DLPFC ROI
was activated prior to speech execution, which could indicate whether R-DLPFC is involved in
speech motor planning in unimpaired speakers. The second-level results, which compared
activation of the controls to rest, were projected onto the same MNI template, and the ROI
analysis was carried out as described above.
(5) Interactive vs. Alone: Finally, we compared activation between the interactive and alone
conditions to justify our decision to exclude “condition” as a factor from all of the models.
Confirmation of the null hypothesis would suggest that condition is not contributing to the
anticipated and unanticipated responses.
We completed two additional ROI analyses. First, we used two control ROIs to confirm
that our findings in R-DLPFC were specific to stuttering anticipation, and not, for example,
general to stuttering or due to systemic artifact. These included L-DLPFC, the homologue of
the R-DLPFC ROI, and right precentral gyrus (R-preCG) from Belyk et al. (2017), the most
recent activation-likelihood meta-analysis of state stuttering (i.e., stuttered vs. fluent). ROIs
were 10 mm spheres with centroids [−50 26 38] for L-DLPFC and [54 −14 34] for R-preCG.
Second, we tested two key superficial cortical nodes of the action-stopping network (i.e.,
R-IFG and R-preSMA), as it has been proposed that action-stopping is associated with stut-
tering anticipation (Arenas, 2017; Hannah & Aron, 2021; Neef et al., 2018). Centroid
Neurobiology of Language
478
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
coordinates were obtained from recent meta-analyses of stuttering: [46 23 −5] for R-IFG,
included as a “state” finding in Belyk et al. (2015); and [15 13 59] for R-preSMA, included
as a “trait” finding in Budde et al. (2014). Trait findings were used for R-preSMA because
state findings were not reported in either meta-analysis. The coordinates were used to create
10 mm ROI spheres.
Functional connectivity. Psychophysiological Interaction (PPI) analysis (Friston et al., 1994,
2003) was used to examine functional connectivity between R-DLPFC and R-SMG. PPI com-
putations were performed on residual components of the modeled task (anticipated vs. unan-
ticipated), after the GLM was effectively removed. Significant correlations are thought to reflect
dynamic neural coupling, though not necessarily related to the task. PPI analysis was con-
ducted using the gPPI toolbox (McLaren et al., 2012) with SPM8. The PPI analysis can be
described by the following equations:
Yk ¼ H xað
Þ;
(cid:4)
(cid:1)
Yi ¼ H xa * gp
(cid:3)
(cid:5)
(cid:4)
* βi þ H gp
(cid:1)
(cid:5)
(cid:3)
* βp þ Yk * βk þ ei;
(1)
(2)
in which the HRF is represented by H, and H(x) is the convolution of signal X with kernel H.
The demeaned time course is represented by gp where 1 is task time and −1 is rest. βi is the
PPI beta value, whereas βp is the beta value for the task, βk is the beta value of the time course
of the seed, and Yk represents the fNIRS data collected at the seed region. Here, k is the func-
tionally defined cluster for the seed region based on the GLM. xa represents the estimated
neural activity for the seed region; the residual error is represented by ei. R-DLPFC had
ROI coordinates [50 26 38], and R-SMG coordinates were determined using the atlas from
NIRS-SPM (Rorden & Brett, 2000) and creating a 10 mm sphere that was projected onto the
brain surface. Each area was used both as a seed and a target, resulting in two comparisons in
total. Two-tailed t tests were used to compare residual activity for anticipated versus
unanticipated words, and the Holm-Bonferroni method was used to correct for familywise
error rates.
RESULTS
Behavioral
Two linear mixed effects models (not included in Jackson et al., 2020) were fit to examine
reaction time and response type. Mean reaction time across all stuttering participants was
348.27 ms (SD = 132.34 ms). See Table 2. Reaction time was significantly impacted by word
type (anticipated/unanticipated), such that anticipated words had longer reaction times (^β =
11.45, t = 2.13, p < 0.05). Reaction time was also impacted by trial (^β = −0.34, t = −2.91, p <
0.01), such that reaction time decreased as the experiment progressed. Reaction time was not
impacted by word length (syllables) (^β = −0.07, t = −0.73, p > 0.05) or stuttering response type
(^β = −5.45, t = −1.54, p > 0.05), indicating that reaction time for stuttered trials was not sig-
nificantly longer than that for fluent trials. R2 for the reaction time model was 0.24. In addition,
post hoc Wilcoxon rank sum tests did not reveal differences in reaction time between stuttered
and ambiguous trials ( W = 203, p > 1.10), ambiguous and fluent trials ( W = 190, p > 0.10), or
stuttered and fluent trials ( W = 152, p > 0.10). Interrater reliability for reaction time between
the first author and a SLP blind to the study yielded a Cohen’s weighted kappa of 0.79 (p <
0.05), indicating moderate to strong agreement (McHugh, 2012).
Neurobiology of Language
479
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
Table 2.
Reaction time means and standard deviations, as determined by first movement
ID
1
Fluent
352.14
SD
103.71
Ambiguous
286.03
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
–
362.68
115.64
398.99
126.61
311.76
120.47
–
–
386.14
90.07
341.00
187.38
310.22
72.77
370.68
109.21
–
–
302.33
75.50
333.70
100.11
360.40
148.11
480.53
147.94
327.57
115.33
290.32
281.56
99.55
87.84
421.30
132.68
515.72
122.92
323.34
76.61
304.64
160.21
362.01
125.17
358.90
–
307.75
300.33
467.18
–
517.24
406.51
296.29
433.81
–
361.51
305.89
300.33
524.39
322.58
340.85
302.18
446.17
517.24
260.29
133.48
310.76
352.49
SD
69.08
101.19
168.84
47.19
–
50.61
132.49
60.60
129.24
–
114.57
64.95
75.91
195.26
70.79
68.24
84.48
208.43
306.75
27.92
47.19
147.09
–
Stuttered
297.39
368.22
SD
108.48
97.08
–
–
–
458.53
272.23
300.33
324.43
282.50
367.07
262.28
303.49
560.34
294.77
282.73
330.44
361.51
489.97
288.55
285.78
367.07
336.10
–
–
–
109.47
87.77
62.43
104.50
81.94
144.78
94.69
69.47
154.65
111.89
69.12
140.16
134.58
134.20
110.64
97.98
95.85
–
Note. Data not obtained in empty cells due to technical complications. ID = participant; SD = standard
deviation.
Table 3 shows the amount of stuttered, ambiguous, and fluent trials for each participant.
For all trials, 43.6% were stuttered (2), 43.3% were fluent (0), and 13.1% were ambiguous (1),
or not unambiguously stuttered or fluent (reported in Jackson et al., 2019; see Table 3). Inter-
rater reliability between the first author and an SLP with 8 years of experience (blind to the
study) yielded a Cohen’s weighted kappa of 0.85 (p < 0.05), indicating high agreement. The
remaining response type data were not reported in Jackson et al. (2020). 53.9% of anticipated
words were unambiguously stuttered and 33.4% of unanticipated words were unambiguously
stuttered. 35.4% of anticipated words were unambiguously fluent whereas 51.2% of unantic-
ipated words were unambiguously fluent. In addition, 10.7% of anticipated words were
ambiguous and 15.41% of unanticipated words were ambiguous. It is important to note that
Neurobiology of Language
480
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
Table 3.
Stuttering response type by participant, including percentages
ID Trials
1
80
Errors
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
80
80
80
60
70
70
80
80
80
80
80
80
80
80
80
80
80
80
80
80
60
4
4
3
1
3
1
1
0
2
0
1
0
1
9
0
0
1
4
0
5
2
Unambiguously
fluent (0)
38
38
70
75
2
7
38
27
37
0
50
7
5
47
49
10
18
30
33
58
33
33
736
%
49%
53%
93%
98%
4%
10%
49%
35%
46%
3%
63%
9%
6%
60%
73%
13%
23%
39%
43%
73%
41%
43%
Ambiguous
(1)
7
9
6
2
5
6
12
34
7
0
12
12
1
8
10
14
21
29
2
5
2
16
222
%
9%
11%
8%
3%
6%
8%
15%
43%
9%
0%
15%
15%
1%
10%
13%
18%
26%
36%
3%
6%
4%
21%
% across data set
43.30%
13.10%
Table continued on next page.
Unambiguously
stuttered (2)
34
29
0
0
52
54
19
18
36
78
18
60
74
24
12
56
41
20
41
17
40
9
%
43%
36%
0%
0%
87%
80%
27%
23%
45%
98%
23%
76%
93%
30%
15%
70%
51%
25%
55%
21%
55%
15%
Anticipated
words that were
unambiguously
stuttered
26
25
0
0
29
30
17
11
24
40
7
30
38
20
11
34
24
11
33
10
24
6
%
65%
63%
0%
0%
97%
89%
49%
28%
60%
100%
18%
78%
95%
50%
28%
85%
60%
28%
90%
25%
68%
20%
742
43.60%
458
53.88%
words characterized as ambiguous using the Jackson et al. (2020) approach would most likely
have been categorized as fluent with a standard binary stuttered/fluent distinction that is most
commonly applied clinically. There was more stuttering for anticipated than unanticipated
words, as expected (^β = −0.36, t = −9.76, p < 0.001). There was also more stuttering for
Neurobiology of Language
481
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
Table 3.
continued from previous page.
ID
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
Unanticipated
words that were
unambiguously
stuttered
8
4
0
0
23
24
2
7
12
38
11
30
36
4
1
22
17
9
8
7
16
3
284
33.41%
%
20%
10%
0%
0%
77%
71%
6%
18%
30%
95%
28%
75%
90%
10%
3%
55%
43%
23%
20%
18%
43%
10%
Anticipated
words that were
unambiguously
fluent
13
10
38
38
0
1
11
13
13
0
27
0
2
16
20
2
8
15
3
28
13
19
301
35.41%
%
33%
30%
98%
98%
3%
3%
34%
35%
33%
0%
68%
0%
5%
40%
58%
5%
20%
38%
10%
70%
33%
63%
Note. ID = participant.
Unanticipated
words that were
unambiguously
fluent
25
28
32
37
2
6
27
14
24
0
23
7
3
31
29
8
10
15
30
30
20
14
435
Anticipated
words that
were
ambiguous %
3%
1
8%
3%
3%
0%
9%
17%
38%
8%
0%
15%
23%
0%
10%
15%
10%
20%
35%
0%
5%
0%
17%
3
1
1
0
3
6
15
3
0
6
9
0
4
6
4
8
14
0
2
0
5
91
%
65%
75%
88%
98%
7%
20%
77%
35%
60%
5%
58%
18%
8%
80%
88%
20%
25%
40%
75%
75%
50%
50%
Unanticipated
words that
were
ambiguous
6
6
5
1
5
3
6
19
4
0
6
3
1
4
4
10
13
15
2
3
2
11
131
%
15%
15%
13%
3%
17%
9%
17%
48%
10%
0%
15%
8%
3%
10%
10%
25%
33%
38%
5%
8%
8%
40%
51.18%
10.71%
15.41%
longer than shorter words (^β = 0.11, t = 5.84, p < 0.001) and for earlier than later trials (^β =
−0.004, t = −4.42, p < 0.001), reflecting a reduction in stuttering over the course of the exper-
iment. R2 for the stuttering response type model was 0.38. Pearson’s correlation test indicated
that stuttering rate, expressed as the percentage of trials stuttered for each participant, and
Neurobiology of Language
482
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
extent of anticipation for each participant (i.e., SAS extent score) were not related (t = 0.47,
p > 0.10).
Neural
ROI analyses
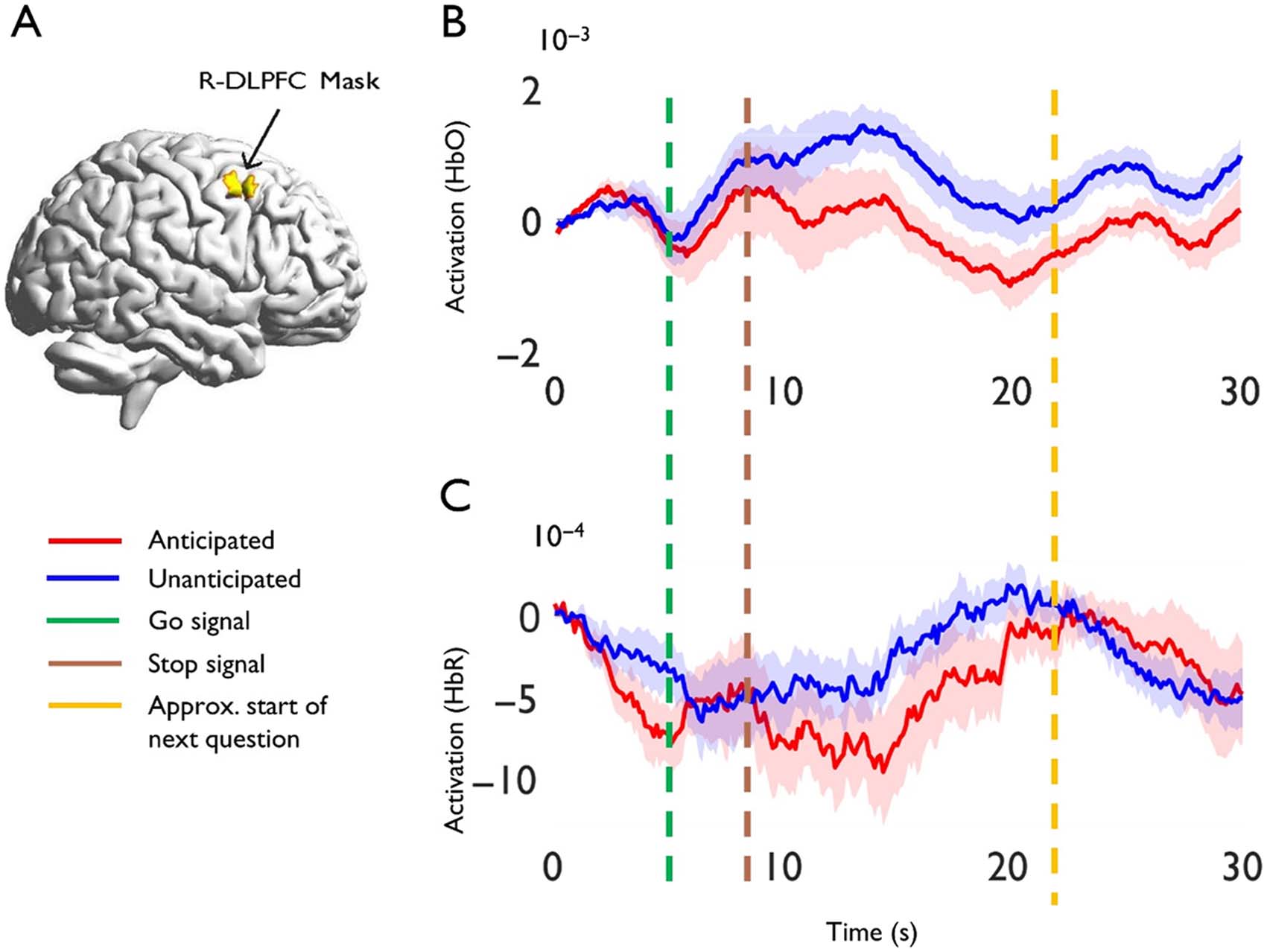
(1) Anticipated vs. Unanticipated: The primary contrast compared activation in the R-DLPFC
mask associated with anticipated vs. unanticipated words. Anticipated words were associated
with greater activation than unanticipated words (p = 0.0217) based on HbR, but not HbO
(p > 0.05). Figure 2 includes the ROI illustration (A) and event-related averages for anticipated
vs. unanticipated words, for HbO (B) and HbR (C), respectively. The HbR difference between
event-related averages at 5 s (i.e., the end of the anticipation phase) for anticipated vs. unan-
ticipated words approached significance (t = 1.67, p = 0.0567), though for HbO, signals were
similar (t = 0.57, p > 0.05). Note that the remaining analyses in this section focus on HbR due
to the null HbO results, as well as evidence suggesting that HbR (1) is more strongly correlated
to the blood oxygen level dependent signal (Boas et al., 2004, 2014; Ferrari & Quaresima,
2012); (2) has greater spatial specificity than HbO (Noah et al., 2021; Zhang et al., 2016);
(3) has been validated for speech-language tasks (Zhang et al., 2017); and (4) is less sensitive
to systemic effects (e.g., heart rate, breathing, bloodflow) than HbO (Franceschini et al., 2003;
Kirilina et al., 2012; Santosa et al., 2019; Scholkmann et al., 2013; Tachtsidis & Scholkmann,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Region of interest analyses. (A) ROI mask for right dorsolateral prefrontal cortex
(R-DLPFC) [58 26 38]. A 10 mm sphere was created using the MarsBar toolbox (Brett et al.,
2002) and the xjView toolbox (https://www.alivelearn.net/xjview). The image was then projected
onto the template using BrainNet Viewer (Xia et al., 2013). (B–C) Event-related averages of activity
for the stuttering group within the R-DLPFC ROI for anticipated vs. unanticipated words, for
Oxyhemoglobin (HbO) and DeOxyhemoglobin (HbR), respectively. 0 seconds is the beginning
of the anticipation phase, and 30 s is shown because the hemodynamic response could feasibly
last this long. HbR signals prior to sign reversal such that decreasing values reflect increases in acti-
vation strength. Shading represents standard error of the mean.
Neurobiology of Language
483
Stuttering and anticipation
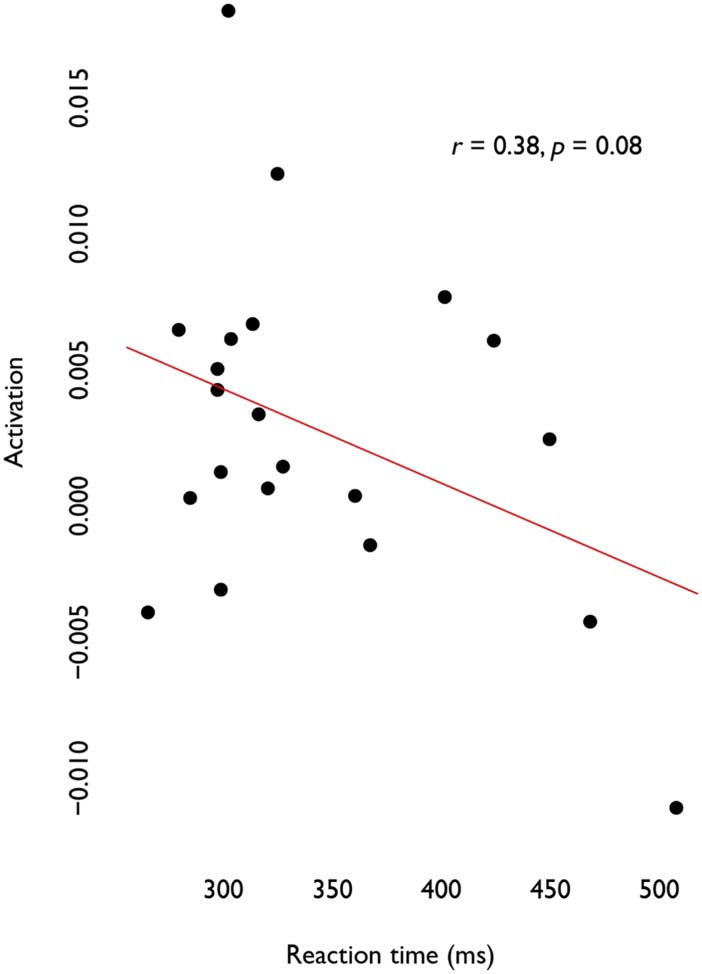
Figure 3. Correlation between activation and reaction time (ms) in R-DLPFC (approached signif-
icance). Deoxyhemoglobin (HbR) signals prior to sign reversal such that decreasing values reflect
increases in activation strength.
2016; Zhang et al., 2016). Still, Figure S2 and Table S2 include the uncorrected whole-brain
results for both chromophores.
The two within-group analyses that compared activation to rest provided some support that
anticipated words are associated with increased activation (p = 0.02), whereas there was no
change for unanticipated words (p = 0.82). Thus, it appears that prior to execution, antici-
pated words recruited the R-DLPFC ROI, whereas unanticipated words did not. Lastly, the
correlation between reaction time and anticipation approached significance (r = 0.38, p =
0.08), indicating that for anticipated words, reaction time may increase as activation increases
(Figure 3).
(2) Stuttered vs. Fluent: Activation in the R-DLPFC mask, using the same 5 s time window
during the anticipation phase, was also greater for stuttered vs. fluent trials (p = 0.0039). See
Figure S3 and Table S3 for uncorrected whole-brain results for HbR as well as HbO. Given that
anticipation is strongly associated with stuttering (above), it is possible that the previous results
for the anticipated vs. unanticipated contrast may be in part due to atypical planning associ-
ated with stuttering that is not necessarily related to anticipation. It was not possible to
compare activation associated with anticipated/stuttered, anticipated/fluent, unanticipated/
stuttered, and unanticipated/fluent, due to the unbalanced distribution of stuttering within par-
ticipants (see Table 3) and the relatively limited number of trials. Importantly, however, 33.4%
of unanticipated words were unambiguously stuttered, and 35.4% of anticipated words were
unambiguously fluent (35.4%).
(3) Stutterers vs. Controls: Activation in the R-DLPFC mask, during the anticipation phase,
was greater for stutterers compared to control speakers (p = 0.0442), irrespective of anticipa-
tion and stuttering. See Figure S4 and Table S4 for uncorrected whole-brain results for HbR
and HbO.
Neurobiology of Language
484
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
(4) Task vs. Rest (controls only): We did not find evidence that R-DLPFC was significantly
activated during the “anticipation” phase in the control group (p = 0.1764). This may indicate that
the area in the R-DLPFC mask was not recruited for speech planning, which is in line with previous
accounts of speech motor control that do not attribute speech motor function to R-DLPFC.
(5) Interactive vs. Alone: Condition was not a variable of interest in the current study, but it
was important to confirm that condition (interactive vs. alone) did not contribute to R-DLPFC
overactivation. A significant difference between the interactive and alone conditions was not
observed (p = 0.4826).
Significant differences in activation were not observed for anticipated vs. unanticipated
words for the two control ROIs: L-DLPFC (p = 0.99); R-preCG (p = 0.95). The lack of signif-
icant differences in these areas provides evidence that the differences observed in the
R-DLPFC ROI were due to the contrast (anticipated vs. unanticipated) and not to general
differences associated with the stuttering brain or systemic artifact. In addition, significant
activation differences were not observed between anticipated vs. unanticipated words in
R-IFG (p = 0.75) or R-preSMA (p = 0.39).
Functional connectivity
Functional connectivity between the R-DLPFC and R-SMG was assessed using each node as a
seed and target, and Holm-Bonferroni correction was applied for the two comparisons.
Compared to unanticipated words, anticipated words were associated with lower intrinsic
connectivity when R-DLPFC was the seed and R-SMG was the target (t = −2.89, p = 0.01),
and also when R-SMG was the seed and R-DLPFC was the target (t = −2.06, p = 0.05). This
reduction in functional connectivity between the R-DLPFC and R-SMG for anticipated relative
to unanticipated words is taken as evidence of involvement of the FPN in stuttering
anticipation.
DISCUSSION
That stutterers anticipate overt stuttering events is well known, but the neural substrates of
anticipation had not been studied previously. In this study, we used a novel, clinically inspired
approach to identify anticipated and unanticipated words in a relatively large sample of adults
who stutter. The words were produced in a delayed-response paradigm while neural signals
were recorded with fNIRS. We identified R-DLPFC as a neural substrate of stuttering anticipa-
tion. A connectivity analysis was also conducted to explore whether the FPN, specifically the
R-DLPFC and R-SMG, is associated with stuttering anticipation. Results are discussed in the
context of theoretical accounts of stuttering anticipation, error-likelihood monitoring, and
action-stopping. Our findings and potential limitations, as well as possible clinical implica-
tions, are also discussed in the following sections.
Right Dorsolateral Prefrontal Cortex Underlies Stuttering Anticipation
The primary hypothesis was confirmed—anticipated words are associated with greater
pre-execution activation in the R-DLPFC, compared to unanticipated words. This means that
the production of words previously identified by participants as being difficult or likely to be
stuttered, up-regulates activation in this area. It was also shown that stutterers exhibit greater
activation than non-stutterers during this same time period, irrespective of anticipation and
stuttering, and while anticipated words elicited activation in the R-DLPFC ROI in stutterers,
unanticipated words of stutterers, and all words produced by controls, did not elicit activation
in this area. Further, anticipated words were associated with longer reaction times, and there
Neurobiology of Language
485
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
was some indication that as activation in R-DLPFC increased, so did reaction time (for antic-
ipated words only). This extra time may be due to speakers delaying speech onset until the
word can appear fluent to listeners, or “letting the stuttering pass,” which could be a function
of the R-DLPFC. Our results provide some clarification of Kell et al. (2009), which found that
stutterers exhibit greater activation than controls in the R-DLPFC, but that after therapy this is
not the case, suggesting that therapy down-regulates an overactive R-DLPFC. Kell et al. (2009)
also found that stutterers who reportedly “recovered” from stuttering after early childhood
exhibited similar activation compared to controls in the R-DLPFC, suggesting that elevated
activation in the R-DLPFC is a maladaptive response to stuttering. Our results suggest that
the Kell et al. (2009) result was due to stuttering anticipation. It is important to highlight that
we focused on longer- (vs. shorter-) term anticipation in this study, as stimuli were identified
between 3 and 10 days prior to the fNIRS experiment. While it is possible that the neural pro-
cesses for longer- and shorter-term anticipation overlap, many words identified as anticipated
before the experiment were not stuttered during the experiment, indicating that they may not
have been anticipated either, suggesting that there may be differences in the underlying pro-
cesses related to longer- vs. shorter-term anticipation. Future studies can attempt to disentangle
these types of anticipation to provide clarity on the time scales associated with anticipation.
Stuttered speech was also associated with elevated activation in R-DLPFC, calling into
question whether atypical activation in this area was due to anticipation, or aberrant planning
associated with stuttering. Given our novel approach, it was not possible to completely differ-
entiate processes associated with anticipation and aberrant planning because we did not
have enough power to make comparisons between anticipated-stuttered, anticipated-fluent,
unanticipated-stuttered, unanticipated-fluent speech. fNIRS, like fMRI, relies on the slow
hemodynamic response, which requires longer trials. With 80 trials already, each lasting at
least 20 s, the experiment lasted approximately 30 min, which pushed the comfort threshold
for participants. Together with the unpredictability of stuttering, this resulted in too few trials
for the aforementioned analysis. However, there is reason to believe that activation differences
were in fact driven by anticipation. First, a significant portion of anticipated/unanticipated
words were fluent/stuttered, respectively. 33% of anticipated words were unambiguously
fluent, whereas 33% of unanticipated words were unambiguously stuttered. Second, it is likely
that anticipation was present throughout the 5 s anticipation phase, whereas it is unlikely that
speech planning processes would comprise the entire 5 s anticipation phase. Third, we did not
find evidence that non-stutterers activate R-DLPFC during the anticipation phase, suggesting
that R-DLPFC does not play a primary role in speech planning. This is consistent with work
on speech motor control or production models that do not include DLPFC as a part of the
speech motor network (e.g., Forseth et al., 2021; Guenther, 2016; Sörös et al., 2006). Future
work can tease these processes apart by obtaining enough trials to make the necessary statis-
tical comparisons. It should also be noted, however, that due to the dynamic nature of both
anticipation and stuttering, it may not be possible to completely differentiate these processes,
especially because speech execution, or the possibility of it, is required to elicit anticipation
in speakers.
Error-Likelihood Monitoring
Anticipation is likely driven by error-likelihood monitoring, which refers to the ability to predict
errors based on prior experience making those errors. In their original account, Brown and
Braver (2005) showed that ACC learns to predict the likelihood of errors, generating a “warning
signal” to heighten readiness or initiate cognitive control in response to predicted errors.
Arenas (2012, 2017) and Garcia-Barrera and Davidow (2015) extended the error-likelihood
Neurobiology of Language
486
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
Figure 4. A model of stuttering anticipation. Anticipation develops via associative learning. Mem-
ory traces of associations form between overtly stuttered utterances and environmental/listener
reactions. ACC detects upcoming stuttering via error-likelihood monitoring and signals R-DLPFC,
which initiates cognitive control to respond to anticipation, thereby elevating activation in
R-DLPFC. R-DLPFC initiates activation within the broader FPN to support the response. Atypical
connection between R-DLPFC and areas within the FPN, such as SMG, impedes the speaker’s ability
to adaptively respond to anticipation. ACC = anterior cingulate cortex; R-DLPFC = right dorsolateral
prefrontal cortex; FPN = frontoparietal control network; R-SMG = right supramarginal gyrus.
account to stuttering, whereby associative learning is the basis for anticipation. The speaker
learns that some words/sounds are difficult, and when the speaker next says these
words/sounds, they are primed to respond to upcoming stuttering. DLPFC works in concert
with ACC to detect and respond to anticipated errors, such that ACC underlies the detection
of errors in response to unintended outcomes, and subsequently generates error signals,
whereas DLPFC generates representations of these errors including holding task-relevant infor-
mation in working memory, and initiating subsequent actions (Alexander & Brown, 2015;
Botvinick et al., 2001; Donoso et al., 2014; Holroyd & Yeung, 2012; Kolling et al., 2012). Inter-
estingly, while Brown and Braver (2005) primarily focused on ACC, they also found error-
likelihood effects in the R-DLPFC and R-SMG, which were less connected functionally for
anticipated vs. unanticipated words in the current study. Thus, anticipation may destabilize
the FPN, potentially reducing speakers’ control in responding to anticipation. We propose a
model that extends the Arenas and Garcia-Barrera and Davidow accounts to the right FPN
(see Figure 4). The ACC detects the error (i.e., anticipates) and subsequently generates an error
signal that is sent to the R-DLPFC, which coupled with R-SMG, holds this information in working
memory and initiates a response. Ultimately, we could not measure ACC due to imaging depth
restrictions associated with fNIRS. Future work can test this proposal using fMRI.
Action-stopping
A related interpretation involves the action-stopping network. Action-stopping has been stud-
ied primarily in the lab using the stop-signal task, in which participants are required to stop an
initiated response when a signal occurs. Aron and colleagues (Aron, 2011; Wessel & Aron,
2017) proposed a prefrontal-basal ganglia-thalamocortical model of action-stopping in which
prefrontal areas (R-IFG, R-preSMA) receive sensory information and subsequently produce a
Neurobiology of Language
487
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
stop command, which is then transmitted to the subthalamic nucleus. The subthalamic
nucleus then relays this information to the globus pallidus interna of the basal ganglia, which
then inhibits the excitatory drive to motor cortex, i.e., to stop the action (Aron, 2011; Wessel &
Aron, 2017). It has been proposed that this mechanism underlies stuttering by inducing a
global inhibition response (over-suppression) that impedes the execution of successive motor
programs (Arenas, 2017; Aron et al., 2014; Neef et al., 2018). Our results did not support this
hypothesis—we did not find evidence of heightened activity in R-IFG or R-preSMA. This may
be because the global inhibition account attempts to explain stuttering events at a speech
motor control level, whereas anticipation occurs at a cognitive control level.
Disentangling reactive and proactive control, two forms of action-stopping, may provide
clarity regarding the distinction between speech motor and cognitive control processes asso-
ciated with stuttering. Reactive control is stimulus-driven and habitual or automatic, whereas
proactive control is prospective and goal-directed (Hannah & Aron, 2021). Reactive control is
supported by the hyperdirect cortico-subthalamic-pallidal-thalamo-cortical pathway, and
includes superficial cortical structures, the R-IFG, and R-preSMA (Jahanshahi et al., 2015). This
pathway is likely associated with the global inhibition that has been proposed to prevent the
execution of successive motor programs (e.g., Arenas, 2017; Aron et al., 2014; Neef et al.,
2018), i.e., to cause or trigger stuttering events. Proactive control, on the other hand, is
regulated by the indirect fronto-striato-pallido-thalamo-cortical pathway, which includes the
R-DLPFC, R-SMG, caudate, and thalamus (A. Chang et al., 2017; Chikazoe et al., 2009;
Jahanshahi et al., 2015; Jahfari et al., 2010; Marek & Dosenbach, 2018; Smittenaar et al.,
2013). Neef et al. (2018) proposed that proactive control may underlie responses to stuttering
anticipation, which is supported by the current findings of overactivation in the R-DLPFC and
reduced connectivity between R-DLPFC and R-SMG. Thus, while the indirect and hyperdirect
pathways share neural circuitry, the hyperdirect pathway may underlie global suppression to
prevent the succession of speech gestures (i.e., the stuttering event), whereas the indirect path-
way seems to be related to how the speaker responds to knowing that the stuttering event is
going to happen. The temporal dynamics associated with fNIRS as well as the slow hemody-
namic response makes fNIRS a sub-optimal tool to test this hypothesis. Future studies could
use fMRI and MEG to tease apart the specific spatial and temporal contributions (respectively)
of both pathways, as well as interactions with the error monitoring system (including the ACC),
to the manifestation of overt stuttering events and the speaker’s anticipation of them.
Clinical Implications
The current work may inform neuromodulation techniques that have recently been applied to
stuttering in clinical trials. Chesters et al. (2018) tested the impact of anodal transcranial direct
current stimulation (tDCS) of left inferior frontal cortex on overt severity in 30 adult stutterers.
They found that the treatment significantly reduced overt stuttering severity at a 1-week follow-up
assessment, and at 6 weeks for reading but not conversational speech (Chesters et al., 2018).
Garnett et al. (2019) tested the impact of anodal tDCS on overt severity in 14 adult stutterers.
They did not find significant effects on overt stuttering severity, though they found that the
atypically strong association between overt severity and right thalamocortical activity was
attenuated, especially in severe stutterers. It may be that the modest effects reported to date
are due to the focus on single areas, specifically in the speech network. Stuttering is not simply
a speech motor control problem. Our data show that the cognitive and sensorimotor processes
that underlie anticipation and subsequent overt stuttering elicit elevated activation in R-DLPFC,
as well as potentially other areas within the FPN. Down-regulating R-DLPFC using tDCS and
concurrently up-regulating, for example, left speech-language areas (premotor cortex/inferior
Neurobiology of Language
488
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
frontal gyrus), which are typically under-active in speakers who stutter, could be the basis for
clinical trials of the effects of tDCS in stuttering therapy.
Behaviorally, the anticipation of stuttering allows stutterers to disguise stuttering, such that
there will be a discrepancy between what listeners hear or see and what speakers experience.
While anticipating stuttering, the speaker may experience anxiety, fear, shame, or other cog-
nitive responses, but the listener may not be privy to this information, creating misunderstand-
ing between the speaker and the listener that could lead to negative listener perceptions of
stuttering. For example, a listener might judge a speaker for looking “nervous” or for not being
intelligent because they do not respond in a timely manner (as perceived by the listener).
Understanding how the brain functions during anticipation could mitigate this discrepancy
through increased understanding of the phenomenon and public awareness. This could also
improve how SLPs work with people who stutter, particularly with respect to helping clients
develop adaptive responses to anticipation. Developing such adaptive responses during
therapy can be challenging, primarily because of the unobservable or “hidden” nature of
anticipation. A brain-based understanding of anticipation may provide an entry point to begin
discussing anticipation with their clients.
Limitations
One limitation of this study is that the hypothesized effect in R-DLPFC—activation greater for
anticipated vs. unanticipated words—was only evident based on the HbR signal, not HbO.
For other motor tasks (e.g., finger tapping), effects are expected to be present for both chromo-
phores. However, it is important to highlight that there is evidence that HbR may be a more reli-
able signal for speech tasks. For example, HbR is more strongly correlated to the blood oxygen
level dependent signal compared to HbO (Boas et al., 2004, 2014; Ferrari & Quaresima, 2012),
has greater spatial specificity than HbO (Noah et al., 2021; Zhang et al., 2016), and has been
validated for speech-language tasks (Zhang et al., 2017). Further, HbR is less sensitive to systemic
effects (e.g., heart rate, breathing, blood flow) than HbO (Franceschini et al., 2003; Kirilina et al.,
2012; Santosa et al., 2019; Scholkmann et al., 2013; Tachtsidis & Scholkmann, 2016; Zhang
et al., 2016), which may be relevant to stuttering because stuttering speakers may experience
elevated anxiety/heart rate prior to speaking tasks. Neither anxiety nor autonomic data were
collected in the current study, but either may have contributed to activation in R-DLPFC. Future
studies could examine the impact of systemic effects on HbO and HbR, particularly for popula-
tions that may exhibit greater systemic effects such as anxiety. This would allow researchers both
to determine whether systemic effects differentially impact HbO/HbR, and also to differentiate
between anticipation and anxiety, to the extent that the latter is even possible.
Another limitation is that the design did not allow us to differentiate between the awareness
of upcoming stuttering and the speaker’s response to the knowledge that upcoming speech
will be stuttered (should they initiate their speech plan as intended). Thus, anticipation and
responding to anticipation are conflated. Future work can address this limitation, for example,
by testing the model of anticipation proposed herein, which postulates that the initial aware-
ness of upcoming speech breakdown may occur in ACC, whereas the response to anticipation
may be supported by R-DLPFC and the broader FPN. A third limitation is that anticipation was
not measured during the actual experiment (e.g., by using a button press), and therefore there
was no indication of the extent to which participants anticipated, if they anticipated at all.
There were several reasons for this. First, we did not want to increase the length of the exper-
iment, which was already nearing the participant comfort threshold. Identifying whether
anticipation occurred could be difficult for some, if not all, participants, which would require
Neurobiology of Language
489
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
additional time. Second, the act of identifying anticipation may alter neural responses. If
identification comes before the trial, the act of identifying itself could alter the neural response.
If identification occurs after the trial, responses may be biased based on whether the trial was
overtly stuttered. One compromise could be to ask participants to identify the extent of antic-
ipation for all words that were previously identified as anticipated just prior to the experiment.
Participants could be presented with stimuli as they would during the actual experiment,
which may help with identifying whether a word would be anticipated.
Conclusion
This study determined that the R-DLPFC is a likely neural substrate of stuttering anticipation, and
also that anticipation may be associated with reduced connectivity within the right hemisphere
FPN. The results support accounts of error-likelihood monitoring and action-stopping and their
association with stuttering events. Future investigations will benefit from adapting the current
paradigm for use with fMRI and MEG to determine the relationships between error-likelihood
monitoring, action-stopping, and stuttering, and whether functional connectivity within the FPN
and related networks (salience, cingulo-opercular) is weaker for anticipated words. It will also
be critical to study children who stutter so that the developmental bases of anticipation can
be determined. Finally, results from this study may inform clinical trials that test the efficacy
of neuromodulation in stuttering therapy, particularly by focusing not only on speech motor
control networks, but also cognitive control and related networks.
ACKNOWLEDGMENTS
The authors thank Drs. Joan Orpella, Adam Buchwald, and Christian A. Kell for reading earlier
versions of this manuscript. Most importantly, the authors thank the participants. The science
of stuttering will not advance without the willing participation of speakers who stutter, and we
are greatly appreciative of their commitment and time.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
FUNDING INFORMATION
Eric S. Jackson, National Institute on Deafness and Other Communication Disorders (https://dx
.doi.org/10.13039/100000055), Award ID: R21DC017821. Swethasri Dravida, National Insti-
tutes of Health (https://dx.doi.org/10.13039/100000002), Award ID: T32GM007205.
Swethasri Dravida, National Institutes of Health (https://dx.doi.org/10.13039/100000002),
Award ID: F30MH116626. Joy Hirsch, National Institute of Mental Health, Award ID:
R01MH111629. Joy Hirsch, National Institute of Mental Health, Award ID: R01MH119430.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
AUTHOR CONTRIBUTIONS
Eric S. Jackson: Conceptualization: Lead; Formal analysis: Equal; Funding acquisition: Lead;
Investigation: Lead; Methodology: Lead; Project administration: Lead; Writing: Lead. Swethasri
Dravida: Conceptualization: Supporting; Formal analysis: Equal; Investigation: Supporting;
Methodology: Supporting; Writing: Supporting. Xian Zhang: Conceptualization: Supporting;
Formal analysis: Equal;
Investigation: Supporting; Methodology: Supporting; Software:
Lead; Writing: Supporting. J. Adam Noah: Conceptualization: Supporting; Formal analysis:
Supporting; Investigation: Supporting; Methodology: Supporting; Writing: Supporting. Vincent
Gracco: Conceptualization: Supporting; Methodology: Supporting; Supervision: Supporting;
Writing: Supporting. Joy Hirsch: Conceptualization: Supporting; Methodology: Supporting;
Supervision: Supporting; Writing: Supporting.
Neurobiology of Language
490
Stuttering and anticipation
REFERENCES
Alexander, W. H., & Brown, J. W. (2015). Hierarchical error repre-
sentation: A computational model of anterior cingulate and
dorsolateral prefrontal cortex. Neural Computation, 27(11),
2354–2410. https://doi.org/10.1162/NECO_a_00779, PubMed:
26378874
Anderson, J. D., & Ofoe, L. C. (2019). The role of executive
function in developmental stuttering. Seminars in Speech and
Language, 40(4), 305–319. https://doi.org/10.1055/s-0039
-1692965, PubMed: 31311055
Arenas, R. M. (2012). The role of anticipation and an adaptive
monitoring system in stuttering: A theoretical and experimental
investigation [Unpublished doctoral dissertation]. University of
Iowa.
Arenas, R. M. (2017). Conceptualizing and investigating the con-
textual variability of stuttering: The speech and monitoring inter-
action (SAMI) framework. Speech, Language and Hearing, 20(1),
15–28. https://doi.org/10.1080/2050571X.2016.1221877
Aron, A. R. (2011). From reactive to proactive and selective control:
Developing a richer model for stopping inappropriate responses.
Biological Psychiatry, 69(12), e55–e68. https://doi.org/10.1016/j
.biopsych.2010.07.024, PubMed: 20932513
Aron, A. R., Robbins, T. W., & Poldrack, R. A. (2014). Inhibition and
the right inferior frontal cortex: One decade on. Trends in Cogni-
tive Sciences, 18(4), 177–185. https://doi.org/10.1016/j.tics.2013
.12.003, PubMed: 24440116
Barton, K. (2020). Mu-MIn: Multi-model inference (R Package Version
0.12. 2/r18. 2009) [Computer software]. https://CRAN.R-project.org/
Bates, D. M., Maechler, M., Bolker, B., & Walker, S. (2014). lme4:
Linear mixed-effects models using ‘Eigen’ and S4 (R package
version 1.1-7) [Computer software]. https://CRAN.R-project.org
/package=lme4
Belyk, M., Kraft, S. J., & Brown, S. (2015). Stuttering as a trait or
state: An ALE meta-analysis of neuroimaging studies. European
Journal of Neuroscience, 41(2), 275–284. https://doi.org/10
.1111/ejn.12765, PubMed: 25350867
Belyk, M., Kraft, S. J., & Brown, S. (2017). Stuttering as a trait or a state
revisited: Motor system involvement in persistent developmental
stuttering. European Journal of Neuroscience, 45(4), 622–624.
https://doi.org/10.1111/ejn.13512, PubMed: 28191730
Boas, D. A., Dale, A. M., & Franceschini, M. A. (2004). Diffuse
optical imaging of brain activation: Approaches to optimizing
image sensitivity, resolution, and accuracy. NeuroImage,
23(S1), S275–S288. https://doi.org/10.1016/j.neuroimage.2004
.07.011, PubMed: 15501097
Boas, D. A., Elwell, C. E., Ferrari, M., & Taga, G. (2014). Twenty
years of functional near-infrared spectroscopy: Introduction for
the special issue. NeuroImage, 85(Pt. 1), 1–5. https://doi.org/10
.1016/j.neuroimage.2013.11.033, PubMed: 24321364
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen,
J. D. (2001). Conflict monitoring and cognitive control. Psycho-
logical Review, 108(3), 624–652. https://doi.org/10.1037/0033
-295X.108.3.624, PubMed: 11488380
Bowers, A., Saltuklaroglu, T., & Kalinowski, J. (2012). Autonomic
arousal in adults who stutter prior to various reading tasks
intended to elicit changes in stuttering frequency. International
Journal of Psychophysiology, 83(1), 45–55. https://doi.org/10
.1016/j.ijpsycho.2011.09.021, PubMed: 22044550
Braun, A. R., Varga, M., Stager, S., Schulz, G., Selbie, S., Maisog,
J. M., Carson, R. E., & Ludlow, C. L. (1997). Altered patterns of
cerebral activity during speech and language production in
developmental stuttering. An H2(15)O positron emission
tomography study. Brain, 120(5), 761–784. https://doi.org/10
.1093/brain/120.5.761, PubMed: 9183248
Brett, M., Anton, J.-L., Valabregue, R., & Poline, J.-B. (2002, June 2–6).
Region of interest analysis using an SPM toolbox [Paper presenta-
tion]. The 8th International Conference on Functional Mapping of
the Human Brain, Sendai, Japan.
Brown, J. W., & Braver, T. S. (2005). Learned predictions of error
likelihood in the anterior cingulate cortex. Science, 307(5712),
1118–1121. https://doi.org/10.1126/science.1105783, PubMed:
15718473
Budde, K. S., Barron, D. S., & Fox, P. T. (2014). Stuttering, induced
fluency, and natural fluency: A hierarchical series of activation
likelihood estimation meta-analyses. Brain and Language, 139,
99–107. https://doi.org/10.1016/j.bandl.2014.10.002, PubMed:
25463820
Chang, A., Ide, J. S., Li, H.-H., Chen, C.-C., & Li, C.-S. R. (2017).
Proactive control: Neural oscillatory correlates of conflict antic-
ipation and response slowing. eNeuro, 4(3), Article 0061-
17.2017. https://doi.org/10.1523/ ENEURO.0061-17.2017,
PubMed: 28560315
Chang, S.-E., Horwitz, B., Ostuni, J., Reynolds, R., & Ludlow, C. L.
(2011). Evidence of left inferior frontal–premotor structural and
functional connectivity deficits in adults who stutter. Cerebral
Cortex, 21(11), 2507–2518. https://doi.org/10.1093/cercor
/bhr028, PubMed: 21471556
Chesters, J., Möttönen, R., & Watkins, K. E. (2018). Transcranial direct
current stimulation over left inferior frontal cortex improves
speech fluency in adults who stutter. Brain, 141(4), 1161–1171.
https://doi.org/10.1093/brain/awy011, PubMed: 29394325
Chikazoe, J., Jimura, K., Hirose, S., Yamashita, K., Miyashita, Y., &
Konishi, S. (2009). Preparation to inhibit a response comple-
ments response inhibition during performance of a stop-signal
task. Journal of Neuroscience, 29(50), 15870–15877. https://doi
.org/10.1523/JNEUROSCI.3645-09.2009, PubMed: 20016103
De Nil, L. F., Kroll, R. M., Kapur, S., & Houle, S. (2000). A positron
emission tomography study of silent and oral single word reading
in stuttering and nonstuttering adults. Journal of Speech, Lan-
guage and Hearing Research, 43(4), 1038–1053. https://doi.org
/10.1044/jslhr.4304.1038, PubMed: 11386470
De Nil, L. F., Kroll, R. M., Lafaille, S. J., & Houle, S. (2004). A pos-
itron emission tomography study of short- and long-term treat-
ment effects on functional brain activation in adults who stutter.
Journal of Fluency Disorders, 28(4), 357–380. https://doi.org/10
.1016/j.jfludis.2003.07.002, PubMed: 14643070
Descorbeth, O., Zhang, X., Noah, J. A., & Hirsch, J. (2020). Neural
processes for live pro-social dialogue between dyads with socio-
economic disparity. Social Cognitive and Affective Neuroscience,
15(8), 875–887. https://doi.org/10.1093/scan/nsaa120, PubMed:
32879986
D’Esposito, M., & Postle, B. R. (2002). The neural basis of working
memory storage, rehearsal, and control processes: Evidence from
patient and functional magnetic resonance imaging studies. In
L. R. Squire & D. L. Schacter (Eds.), Neuropsychology of memory
(pp. 215–224). Guilford Press.
Donoso, M., Collins, A. G., & Koechlin, E. (2014). Foundations of
human reasoning in the prefrontal cortex. Science, 344(6191),
1481–1486. https://doi.org/10.1126/science.1252254, PubMed:
24876345
Eggebrecht, A. T., White, B. R., Ferradal, S. L., Chen, C., Zhan, Y.,
Snyder, A. Z., Dehghani, H., & Culver, J. P. (2012). A quantitative
spatial comparison of high-density diffuse optical tomography
Neurobiology of Language
491
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
and fMRI cortical mapping. NeuroImage, 61(4), 1120–1128.
https://doi.org/10.1016/j.neuroimage.2012.01.124, PubMed:
22330315
Ferradal, S. L., Eggebrecht, A. T., Hassanpour, M., Snyder, A. Z., &
Culver, J. P. (2014). Atlas-based head modeling and spatial nor-
malization for high-density diffuse optical tomography: In vivo
validation against fMRI. NeuroImage, 85(1), 117–126. https://doi
.org/10.1016/j.neuroimage.2013.03.069, PubMed: 23578579
Ferrari, M., & Quaresima, V. (2012). A brief review on the history of
human functional near-infrared spectroscopy (fNIRS) develop-
ment and fields of application. NeuroImage, 63(2), 921–935.
https://doi.org/10.1016/j.neuroimage.2012.03.049, PubMed:
22510258
Forseth, K. J., Pitkow, X., Fischer-Baum, S., & Tandon, N. (2021).
What the brain does as we speak. BioRxiv. https://doi.org/10
.1101/2021.02.05.429841
Fox, P. T., Ingham, R. J., Ingham, J. C., Zamarripa, F., Xiong, J.-H., &
Lancaster, J. L. (2000). Brain correlates of stuttering and syllable
production: A PET performance-correlation analysis. Brain,
123(10), 1985–2004. https://doi.org/10.1093/ brain/123.10
.1985, PubMed: 11004117
Franceschini, M. A., Fantini, S., Thompson, J. H., Culver, J. P., &
Boas, D. A. (2003). Hemodynamic evoked response of the
sensorimotor cortex measured noninvasively with near-infrared
optical imaging. Psychophysiology, 40(4), 548–560. https://doi
.org/10.1111/1469-8986.00057, PubMed: 14570163
Friston, K. J., Harrison, L., & Penny, W. (2003). Dynamic causal
modelling. NeuroImage, 19(4), 1273–1302. https://doi.org/10
.1016/S1053-8119(03)00202-7, PubMed: 12948688
Friston, K. J., Holmes, A. P., Poline, J. B., Grasby, P. J., Williams,
S. C. R., Frackowiak, R. S., & Turner, R. (1995). Analysis of fMRI
time-series revisited. NeuroImage, 2(1), 45–53. https://doi.org/10
.1006/nimg.1995.1007, PubMed: 9343589
Friston, K. J., Holmes, A. P., Worsley, K. J., Poline, J.-P., Frith, C. D.,
& Frackowiak, R. S. (1994). Statistical parametric maps in func-
tional imaging: A general linear approach. Human Brain Map-
ping, 2(4), 189–210. https://doi.org/10.1002/hbm.460020402
Garcia-Barrera, M. A., & Davidow, J. H. (2015). Anticipation in
stuttering: A theoretical model of the nature of stutter prediction.
Journal of Fluency Disorders, 44, 1–15. https://doi.org/10.1016/j
.jfludis.2015.03.002, PubMed: 25841698
Garnett, E. O., Chow, H. M., Choo, A. L., & Chang, S.-E. (2019).
Stuttering severity modulates effects of non-invasive brain stimu-
lation in adults who stutter. Frontiers in Human Neuroscience,
13, Article 411. https://doi.org/10.3389/fnhum.2019.00411,
PubMed: 31824276
Goldman-Rakic, P. S. (1988). Topography of cognition: Parallel dis-
tributed networks in primate association cortex. Annual Review
of Neuroscience, 11(1), 137–156. https://doi.org/10.1146
/annurev.ne.11.030188.001033, PubMed: 3284439
Guenther, F. H. (2016). Neural control of speech. MIT Press. https://
doi.org/10.7551/mitpress/10471.001.0001
Hannah, R., & Aron, A. R. (2021). Towards real-world generaliz-
ability of a circuit for action-stopping. Nature Reviews Neurosci-
ence, 22(9), 538–552. https://doi.org/10.1038/s41583-021
-00485-1, PubMed: 34326532
Hirsch, J., Noah, J. A., Zhang, X., Dravida, S., & Ono, Y. (2018). A
cross-brain neural mechanism for human-to-human verbal com-
munication. Social Cognitive and Affective Neuroscience, 13(9),
907–920. https://doi.org/10.1093/scan/nsy070, PubMed:
30137601
Hirsch, J., Tiede, M., Zhang, X., Noah, J. A., Salama-Manteau, A., &
Biriotti, M. (2021). Interpersonal agreement and disagreement
during face-to-face dialogue: An fNIRS investigation. Frontiers
in Human Neuroscience, 14, Article 606397. https://doi.org/10
.3389/fnhum.2020.606397, PubMed: 33584223
Hirsch, J., Zhang, X., Noah, J. A., & Ono, Y. (2017). Frontal temporal
and parietal systems synchronize within and across brains during
live eye-to-eye contact. NeuroImage, 157, 314–330. https://doi
.org/10.1016/j.neuroimage.2017.06.018, PubMed: 28619652
Holroyd, C. B., & Yeung, N. (2012). Motivation of extended behav-
iors by anterior cingulate cortex. Trends in Cognitive Sciences,
16(2), 122–128. https://doi.org/10.1016/j.tics.2011.12.008,
PubMed: 22226543
Jackson, E. S., Gerlach, H., Rodgers, N. H., & Zebrowski, P. M.
(2018). My client knows that he’s about to stutter: How can
we address stuttering anticipation during therapy with young
people who stutter? Seminars in Speech and Language, 39,
356–370. https://doi.org/10.1055/s-0038-1667164, PubMed:
30142646
Jackson, E. S., Gracco, V., & Zebrowski, P. M. (2020). Eliciting stut-
tering in laboratory contexts. Journal of Speech, Language, and
Hearing Research, 63(1), 143–150. https://doi.org/10.1044/2019
_JSLHR-S-19-0173, PubMed: 31835000
Jackson, E. S., Rodgers, N. H., & Rodgers, D. B. (2019). An explor-
atory factor analysis of action responses to stuttering anticipation.
Journal of Fluency Disorders, 60, 1–10. https://doi.org/10.1016/j
.jfludis.2019.03.001, PubMed: 30875585
Jackson, E. S., Yaruss, J. S., Quesal, R. W., Terranova, V., & Whalen,
D. H. (2015). Responses of adults who stutter to the anticipation
of stuttering. Journal of Fluency Disorders, 45, 38–51. https://doi
.org/10.1016/j.jfludis.2015.05.002, PubMed: 26065618
Jahanshahi, M., Obeso, I., Rothwell, J. C., & Obeso, J. A. (2015). A
fronto–striato–subthalamic–pallidal network for goal-directed
and habitual inhibition. Nature Reviews Neuroscience, 16(12),
719–732. https://doi.org/10.1038/nrn4038, PubMed: 26530468
Jahfari, S., Stinear, C. M., Claffey, M., Verbruggen, F., & Aron, A. R.
(2010). Responding with restraint: What are the neurocognitive
mechanisms? Journal of Cognitive Neuroscience, 22(7),
1479–1492. https://doi.org/10.1162/jocn.2009.21307, PubMed:
19583473
Kell, C. A., Neumann, K., von Kriegstein, K., Posenenske, C., von
Gudenberg, A. W., Euler, H., & Giraud, A.-L. (2009). How the
brain repairs stuttering. Brain, 132(10), 2747–2760. https://doi
.org/10.1093/brain/awp185, PubMed: 19710179
Kirilina, E., Jelzow, A., Heine, A., Niessing, M., Wabnitz, H., Brühl,
R., Ittermann, B., Jacobs, A. M., & Tachtsidis, I. (2012). The phys-
iological origin of task-evoked systemic artefacts in functional
near infrared spectroscopy. NeuroImage, 61(1), 70–81. https://
doi.org/10.1016/j.neuroimage.2012.02.074, PubMed:
22426347
Knott, J. R., Johnson, W., & Webster, M. J. (1937). Studies in the
psychology of stuttering: II A quantitative evaluation of expecta-
tion of stuttering in relation to the occurrence of stuttering. Jour-
nal of Speech Disorders, 2(1), 20–22. https://doi.org/10.1044/jshd
.0201.20
Koechlin, E., Ody, C., & Kouneiher, F. (2003). The architecture of
cognitive control in the human prefrontal cortex. Science,
302(5648), 1181–1185. https://doi.org/10.1126/science
.1088545, PubMed: 14615530
Kolling, N., Behrens, T. E., Mars, R. B., & Rushworth, M. F.
(2012). Neural mechanisms of foraging. Science, 336(6077),
95–98. https://doi.org/10.1126/science.1216930, PubMed:
22491854
Krall, E. A., & Dawson-Hughes, B. (1993). Heritable and life-style
determinants of bone mineral density. Journal of Bone and
Neurobiology of Language
492
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
Mineral Research, 8(1), 1–9. https://doi.org/10.1002/jbmr
.5650080102, PubMed: 8427042
Kuznetsova, A., Brockhoff, P. B., & Christensen, R. H. B. (2017).
lmerTest package: Tests in linear mixed effects models. Journal
of Statistical Software, 82(13), 1–26. https://doi.org/10.18637/jss
.v082.i13
Lancaster, J. L., Woldorff, M. G., Parsons, L. M., Liotti, M., Freitas,
C. S., Rainey, L., Kochunov, P. V., Nickerson, D., Mikiten, S. A., &
Fox, P. T. (2000). Automated Talairach atlas labels for functional
brain mapping. Human Brain Mapping, 10(3), 120–131. https://
doi.org/10.1002/1097-0193(200007)10:3<120::AID-HBM30>3
.0.CO;2-8, PubMed: 10912591
Lu, C., Ning, N., Peng, D., Ding, G., Li, K., Yang, Y., & Lin, C.
(2009). The role of large-scale neural interactions for develop-
mental stuttering. Neuroscience, 161(4), 1008–1026. https://
doi.org/10.1016/j.neuroscience.2009.04.020, PubMed:
19364522
MacDonald, A. W., Cohen, J. D., Stenger, V. A., & Carter, C. S.
(2000). Dissociating the role of the dorsolateral prefrontal and
anterior cingulate cortex in cognitive control. Science, 288(5472),
1835–1838. https://doi.org/10.1126/science.288.5472.1835,
PubMed: 10846167
Marek, S., & Dosenbach, N. U. (2018). The frontoparietal network:
Function, electrophysiology, and importance of individual
precision mapping. Dialogues in Clinical Neuroscience, 20(2),
133–140. https://doi.org/10.31887/ DCNS.2018.20.2/smarek,
PubMed: 30250390
Matcher, S. J., Elwell, C. E., Cooper, C. E., Cope, M., & Delpy, D. T.
(1995). Performance comparison of several published tissue
near-infrared spectroscopy algorithms. Analytical Biochemistry,
227(1), 54–68. https://doi.org/10.1006/abio.1995.1252,
PubMed: 7668392
McHugh, M. L. (2012). Interrater reliability: The kappa statistic.
Biochemia Medica, 22(3), 276–282. https://doi.org/10.11613
/BM.2012.031, PubMed: 23092060
McLaren, D. G., Ries, M. L., Xu, G., & Johnson, S. C. (2012). A gen-
eralized form of context-dependent psychophysiological interac-
tions (gPPI): A comparison to standard approaches. NeuroImage,
61(4), 1277–1286. https://doi.org/10.1016/j.neuroimage.2012
.03.068, PubMed: 22484411
Menon, V., & D’Esposito, M. (2021). The role of PFC networks in
cognitive control and executive function. Neuropsychopharma-
cology, 47, 1–14. https://doi.org/10.1038/s41386-021-01152-w,
PubMed: 34408276
Mersov, A., Cheyne, D., Jobst, C., & De Nil, L. (2018). A prelimi-
nary study on the neural oscillatory characteristics of motor
preparation prior to dysfluent and fluent utterances in adults
who stutter. Journal of Fluency Disorders, 55, 145–155. https://
doi.org/10.1016/j.jfludis.2017.05.003, PubMed: 28577876
Mesulam, M. M. (1998). From sensation to cognition. Brain, 121(6),
1013–1052. https://doi.org/10.1093/brain/121.6.1013, PubMed:
9648540
Milisen, R. (1938). Frequency of stuttering with anticipation of
stuttering controlled. Journal of Speech Disorders, 3(4), 207–214.
https://doi.org/10.1044/jshd.0304.207
Miller, E. K. (2000). The prefrontal cortex and cognitive control.
Nature Reviews Neuroscience, 1(1), 59–65. https://doi.org/10
.1038/35036228, PubMed: 11252769
Neef, N. E., Anwander, A., Bütfering, C., Schmidt-Samoa, C.,
Friederici, A. D., Paulus, W., & Sommer, M. (2018). Structural
connectivity of right frontal hyperactive areas scales with stutter-
ing severity. Brain, 141(1), 191–204. https://doi.org/10.1093
/brain/awx316, PubMed: 29228195
Neef, N. E., Bütfering, C., Anwander, A., Friederici, A. D., Paulus,
W., & Sommer, M. (2016). Left posterior-dorsal area 44 couples
with parietal areas to promote speech fluency, while right area
44 activity promotes the stopping of motor responses. Neuro-
Image, 142, 628–644. https://doi.org/10.1016/j.neuroimage
.2016.08.030, PubMed: 27542724
Neumann, K., Euler, H. A., von Gudenberg, A. W., Giraud, A.-L.,
Lanfermann, H., Gall, V., & Preibisch, C. (2004). The nature and
treatment of stuttering as revealed by fMRI: A within- and
between-group comparison. Journal of Fluency Disorders,
28(4), 381–410. https://doi.org/10.1016/j.jfludis.2003.07.003,
PubMed: 14643071
Neumann, K., Preibisch, C., Euler, H. A., von Gudenberg, A. W.,
Lanfermann, H., Gall, V., & Giraud, A.-L. (2005). Cortical plastic-
ity associated with stuttering therapy. Journal of Fluency Disor-
ders, 30(1), 23–39. https://doi.org/10.1016/j.jfludis.2004.12
.002, PubMed: 15769497
Niendam, T. A., Laird, A. R., Ray, K. L., Dean, Y. M., Glahn, D. C.,
& Carter, C. S. (2012). Meta-analytic evidence for a superordi-
nate cognitive control network subserving diverse executive
functions. Cognitive, Affective, & Behavioral Neuroscience,
12(2), 241–268. https://doi.org/10.3758/s13415-011-0083-5,
PubMed: 22282036
Noah, J. A., Zhang, X., Dravida, S., DiCocco, C., Suzuki, T., Aslin,
R. N., Tachtsidis, I., & Hirsch, J. (2021). Comparison of
short-channel separation and spatial domain filtering for removal
of non-neural components in functional near-infrared spectros-
copy signals. Neurophotonics, 8(1), Article 015004. https://doi
.org/10.1117/1.NPh.8.1.015004, PubMed: 33598505
Okada, E., & Delpy, D. T. (2003). Near-infrared light propagation in
an adult head model. II. Effect of superficial tissue thickness on
the sensitivity of the near-infrared spectroscopy signal. Applied
Optics, 42(16), 2915–2921. https://doi.org/10.1364/AO.42
.002915, PubMed: 12790440
Okamoto, M., & Dan, I. (2005). Automated cortical projection of
head-surface locations for transcranial functional brain mapping.
NeuroImage, 26(1), 18–28. https://doi.org/10.1016/j.neuroimage
.2005.01.018, PubMed: 15862201
Preibisch, C., Neumann, K., Raab, P., Euler, H. A., von Gudenberg,
A. W., Lanfermann, H., & Giraud, A.-L. (2003). Evidence for
compensation for stuttering by the right frontal operculum. Neu-
roImage, 20(2), 1356–1364. https://doi.org/10.1016/S1053-8119
(03)00376-8, PubMed: 14568504
R Core Team. (2014). R: A language and environment for statistical
computing [Computer software]. R Foundation for Statistical
Computing. https://www.R-project.org/
Ridderinkhof, K. R., van den Wildenberg, W. P., Segalowitz, S. J., &
Carter, C. S. (2004). Neurocognitive mechanisms of cognitive
control: The role of prefrontal cortex in action selection, response
inhibition, performance monitoring, and reward-based learning.
Brain and Cognition, 56(2), 129–140. https://doi.org/10.1016/j
.bandc.2004.09.016, PubMed: 15518930
Rorden, C., & Brett, M. (2000). Stereotaxic display of brain lesions.
Behavioural Neurology, 12(4), Article 421719. https://doi.org/10
.1155/2000/421719, PubMed: 11568431
Santosa, H., Fishburn, F., Zhai, X., & Huppert, T. J. (2019). Investiga-
tion of the sensitivity-specificity of canonical- and deconvolution-
based linear models in evoked functional near-infrared spectroscopy.
Neurophotonics, 6(2), Article 025009. https://doi.org/10.1117/1
.NPh.6.2.025009, PubMed: 31172019
Scholkmann, F., Gerber, U., Wolf, M., & Wolf, U. (2013). End-tidal
CO2: An important parameter for a correct interpretation in func-
tional brain studies using speech tasks. NeuroImage, 66, 71–79.
Neurobiology of Language
493
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Stuttering and anticipation
https://doi.org/10.1016/j.neuroimage.2012.10.025, PubMed:
23099101
Smittenaar, P., Guitart-Masip, M., Lutti, A., & Dolan, R. J. (2013).
Preparing for selective inhibition within frontostriatal loops. Jour-
nal of Neuroscience, 33(46), 18087–18097. https://doi.org/10
.1523/JNEUROSCI.2167-13.2013, PubMed: 24227719
Sörös, P., Guttman Sokoloff, L., Bose, A., McIntosh, A. R., Graham,
S. J., & Stuss, D. T. (2006). Clustered functional MRI of overt
speech production. NeuroImage, 32(1), 376–387. https://doi.org
/10.1016/j.neuroimage.2006.02.046, PubMed: 16631384
Tachtsidis, I., & Scholkmann, F. (2016). False positives and false
negatives in functional near-infrared spectroscopy: Issues, chal-
lenges, and the way forward. Neurophotonics, 3(3), Article
031405. https://doi.org/10.1117/1.NPh.3.3.031405, PubMed:
27054143
Tichenor, S. E., & Yaruss, J. S. (2019). Repetitive negative thinking,
temperament, and adverse impact in adults who stutter. American
Journal of Speech-Language Pathology, 29(1), 201–215. https://doi
.org/10.1044/2019_AJSLP-19-00077, PubMed: 31846585
Toyomura, A., Fujii, T., Yokosawa, K., & Kuriki, S. (2018). Speech
disfluency-dependent amygdala activity in adults who stutter:
Neuroimaging of interpersonal communication in MRI scanner
environment. Neuroscience, 374, 144–154. https://doi.org/10
.1016/j.neuroscience.2018.01.037, PubMed: 29378280
Van Riper, C. (1936). Study of the thoracic breathing of stutterers
during expectancy and occurrence of stuttering spasm. Journal
of Speech Disorders, 1(3), 61–72. https://doi.org/10.1044/jshd
.0103.61
Vassena, E., Gerrits, R., Demanet, J., Verguts, T., & Siugzdaite, R.
(2019). Anticipation of a mentally effortful task recruits dorsolateral
prefrontal cortex: An fNIRS validation study. Neuropsychologia,
123, 106–115. https://doi.org/10.1016/j.neuropsychologia.2018
.04.033, PubMed: 29705065
Wessel, J. R., & Aron, A. R. (2017). On the globality of motor sup-
pression: Unexpected events and their influence on behavior and
cognition. Neuron, 93(2), 259–280. https://doi.org/10.1016/j
.neuron.2016.12.013, PubMed: 28103476
Wingate, M. E. (1975). Expectancy as basically a short-term pro-
cess. Journal of Speech and Hearing Research, 18(1), 31–42.
https://doi.org/10.1044/jshr.1801.31, PubMed: 1127907
Wymbs, N. F., Ingham, R. J., Ingham, J. C., Paolini, K. E., & Grafton,
S. T. (2013). Individual differences in neural regions functionally
related to real and imagined stuttering. Brain and Language,
124(2), 153–164. https://doi.org/10.1016/j.bandl.2012.11.013,
PubMed: 23333668
Xia, M., Wang, J., & He, Y. (2013). BrainNet Viewer: A network
visualization tool for human brain connectomics. PLOS ONE,
8(7), Article e68910. https://doi.org/10.1371/journal.pone
.0068910, PubMed: 23861951
Yairi, E., & Ambrose, N. (1992). A longitudinal study of stuttering in
children. Journal of Speech, Language, and Hearing Research,
35(4), 755–760. https://doi.org/10.1044/jshr.3504.755,
PubMed: 1405530
Ye, J. C., Tak, S., Jang, K. E., Jung, J., & Jang, J. (2009). NIRS-SPM:
Statistical parametric mapping for near-infrared spectroscopy.
NeuroImage, 44(2), 428–447. https://doi.org/10.1016/j
.neuroimage.2008.08.036, PubMed: 18848897
Zhang, X., Noah, J. A., Dravida, S., & Hirsch, J. (2017). Signal pro-
cessing of functional NIRS data acquired during overt speaking.
Neurophotonics, 4(4), Article 041409. https://doi.org/10.1117/1
.NPh.4.4.041409, PubMed: 28924564
Zhang, X., Noah, J. A., & Hirsch, J. (2016). Separation of the global
and local components in functional near-infrared spectroscopy
signals using principal component spatial filtering. Neuropho-
tonics, 3(1), Article 015004. https://doi.org/10.1117/1.NPh.3.1
.015004, PubMed: 26866047
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
–
p
d
f
/
/
/
/
3
3
4
6
9
2
0
3
9
9
8
7
n
o
_
a
_
0
0
0
7
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
494